
Hormonally Induced Changes in Stem and Petiole · Time course changes were observed in petiole and...
7
Plant Physiol. (1973) 52, 554-560 Hormonally Induced Changes in the Stem and Petiole Anatomy W1 and Cellulase Enzyme Patterns in Phaseolus vulgaris L.' Received for publication April 9, 1973 ARTHUR E. LINKINS, LOWELL N. LEWIS, AND RAYMOND L. PALMER Department of Plant Sciences, University of California, Riverside, California 92502 ABSTRACT Time course changes were observed in petiole and stem anatomy and cellulase enzyme patterns in bean (Phaseolus vulgaris L.) explants when 10' or 102 M indoleacetic acid in lanolin paste was applied to acropetal cut surfaces in the presence or absence of ethylene. Auxin (102 M) in the pres- ence of ethylene stimulated rapid ordered cell division and dedifferentiation, with ensuing lateral root formation. Auxin (10-5 M) caused moderate cortical swelling, pit formation in pith parenchyma, and chloroplast development in certain cortical cells. Exogenous ethylene reduced cell division ac- tivity and caused cortical cell swelling and separation. Re- moval of endogenously generated ethylene by mercuric per- chlorate resulted in less ordered cell division patterns and no lateral root formation. Auxin treatments enhanced formation of an active acidic pI cellulase, exogenous ethylene-stimulated formation of an active basic pl cellulase. The absence of basic pl cellulase activity by the removal of endogenously generated ethylene suggests a close dependence of basic pl cellulase activity on ethylene. Based on studies of plants treated with auxin or ethylene, it appears that plant cellulase occurs in different forms (12), de- grades cellulose differentially (17), and exists in different parts of the cell (1, 4). Lewis et al. (12) and Lewis and Varner (13) have associated one form with events related to abscission, while Sheldrake (18-22) has shown cellulase activity associated with xylem, phloem, and laticifer differentiation. These obser- vations suggest that cellulase, or the separate forms of cellu- lase, serve different functions through their specific hydrolytic effects on the cell wall. It is the purpose of this paper to ex- amine the role of the different cellulases in the morphological changes produced by auxin or ethylene treatment in the bean petiole and stem. MATERIALS AND METHODS Plant Materials and Treatments. Phaseolus vulgaris L., cv. Red Kidney, seeds obtained from W. Atlee Burpee Company were germinated and grown under the conditions described by Lewis and Varner (13). Seedlings 10- to 11-days-old were de- bladed, the laminar pulvinus and cotyledons were removed, 1 This work was supported by National Science Foundation Grant GB 17850. and the remaining plant was cut off 4 to 5 cm below the cot- yledonary node. The intact primary axis with the first leaf node petioles were then placed in cups of water and treated as follows: explants had either 10'2 M or 10' M indoleacetic acid in lanolin paste or pure lanolin paste applied to the acropetal cut petiolar surfaces (Fig. 1). In selected experiments the ex- plants were exposed to 15 u1/I ethylene (Matheson Co.) for the final 20 to 24 hr of IAA treatment. At the termination of treatment periods, the lanolin paste was removed from the control explants or explants treated with auxin or ethylene or both, and the 5 mm area below the treated surface was taken for anatomical study or cellulase extraction (Fig. 1). In experiments where ethylene was used as the sole treat- ment, the main stem was severed immediately below the basi- petal end of the nodal pulvinus of the first leaf axis (Fig. 1). Lanolin paste was then applied to the cut surface of the stem before exposure to 5 ,ul/l ethylene in air-tight chambers. This modification was necessary to circumvent the premature cessa- tion of experiments using petiole tissue due to the abscission of the petiole at the nodal axis after 36 to 48 hr treatment. Cellulase Extraction and Purification. Cellulase was ex- tracted by grinding the excised 5-mm segments for 1 min in a VirTis homogenizer with 0.2 M phosphate buffer, pH 6.1, 1.0 M NaCl, and 3% insoluble polyvinylpyrrolidone. The homogenate was passed through a 50 , mesh nylon cloth. The pellet was re- suspended in the above buffer less the polyvinylpyrrolidone, ground for 1 min, and filtered again. The filtered extracts were pooled and centrifuged at 10,000g for 15 min; the resultant supematant solution was centrifuged at 100,OOOg for 1 hr. The final supernatant solution was then dialyzed against two changes of 1 % glycine, pH 6.1, for 2 hr each. The solution was then subjected to isoelectric focusing ac- cording to the procedure of Hagland (6) to separate acidic pl (isoelectric point) and basic pl cellulase. Samples were focused for 18 to 24 hr at 300 v. Determination of Ethylene Generated by Explants. Petioles prepared as described were removed from the explants, treated at their acropetal cut surface with either pure lanolin, or lano- lin with 10' to 10'2 M IAA. The treated petioles were sup- ported by agar in the bottom of a 50-ml flask which was then sealed with a serum cap. Ethylene measurements were deter- mined on 1-ml samples using an Aerograph 600D chromato- graph with a flame ionization detector. Column packing was Poropak Q with 2.54 cm of activated alumina at each end of a 0.32 cm X 71.28 cm stainless steel column (15). Sensitivity of this system was greater than 0.02 nl ethylene/ml of sample. Ethylene Trapping. Endogenously generated ethylene from explants enclosed in an air-tight chamber was trapped using the ethylene trap, mercuric perchlorate, described by Young et al. (25). Histological Techniques. Selected 5-mm segments removed 554 www.plantphysiol.org on June 5, 2020 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.
Transcript of Hormonally Induced Changes in Stem and Petiole · Time course changes were observed in petiole and...

Plant Physiol. (1973) 52, 554-560
Hormonally Induced Changes in the Stem and Petiole AnatomyW1
and Cellulase Enzyme Patterns in Phaseolus vulgaris L.'Received for publication April 9, 1973
ARTHUR E. LINKINS, LOWELL N. LEWIS, AND RAYMOND L. PALMERDepartment of Plant Sciences, University of California, Riverside, California 92502
ABSTRACT
Time course changes were observed in petiole and stemanatomy and cellulase enzyme patterns in bean (Phaseolusvulgaris L.) explants when 10' or 102 M indoleacetic acidin lanolin paste was applied to acropetal cut surfaces in thepresence or absence of ethylene. Auxin (102 M) in the pres-ence of ethylene stimulated rapid ordered cell division anddedifferentiation, with ensuing lateral root formation. Auxin(10-5 M) caused moderate cortical swelling, pit formation inpith parenchyma, and chloroplast development in certaincortical cells. Exogenous ethylene reduced cell division ac-tivity and caused cortical cell swelling and separation. Re-moval of endogenously generated ethylene by mercuric per-chlorate resulted in less ordered cell division patterns and nolateral root formation. Auxin treatments enhanced formationof an active acidic pI cellulase, exogenous ethylene-stimulatedformation of an active basic pl cellulase. The absence ofbasic pl cellulase activity by the removal of endogenouslygenerated ethylene suggests a close dependence of basic plcellulase activity on ethylene.
Based on studies of plants treated with auxin or ethylene, itappears that plant cellulase occurs in different forms (12), de-grades cellulose differentially (17), and exists in different partsof the cell (1, 4). Lewis et al. (12) and Lewis and Varner (13)have associated one form with events related to abscission,while Sheldrake (18-22) has shown cellulase activity associatedwith xylem, phloem, and laticifer differentiation. These obser-vations suggest that cellulase, or the separate forms of cellu-lase, serve different functions through their specific hydrolyticeffects on the cell wall. It is the purpose of this paper to ex-amine the role of the different cellulases in the morphologicalchanges produced by auxin or ethylene treatment in the beanpetiole and stem.
MATERIALS AND METHODS
Plant Materials and Treatments. Phaseolus vulgaris L., cv.Red Kidney, seeds obtained from W. Atlee Burpee Companywere germinated and grown under the conditions described byLewis and Varner (13). Seedlings 10- to 11-days-old were de-bladed, the laminar pulvinus and cotyledons were removed,
1 This work was supported by National Science FoundationGrant GB 17850.
and the remaining plant was cut off 4 to 5 cm below the cot-yledonary node. The intact primary axis with the first leafnode petioles were then placed in cups of water and treated asfollows: explants had either 10'2 M or 10' M indoleacetic acidin lanolin paste or pure lanolin paste applied to the acropetalcut petiolar surfaces (Fig. 1). In selected experiments the ex-plants were exposed to 15 u1/I ethylene (Matheson Co.) for thefinal 20 to 24 hr of IAA treatment. At the termination oftreatment periods, the lanolin paste was removed from thecontrol explants or explants treated with auxin or ethylene orboth, and the 5 mm area below the treated surface was takenfor anatomical study or cellulase extraction (Fig. 1).
In experiments where ethylene was used as the sole treat-ment, the main stem was severed immediately below the basi-petal end of the nodal pulvinus of the first leaf axis (Fig. 1).Lanolin paste was then applied to the cut surface of the stembefore exposure to 5 ,ul/l ethylene in air-tight chambers. Thismodification was necessary to circumvent the premature cessa-tion of experiments using petiole tissue due to the abscissionof the petiole at the nodal axis after 36 to 48 hr treatment.
Cellulase Extraction and Purification. Cellulase was ex-tracted by grinding the excised 5-mm segments for 1 min in aVirTis homogenizer with 0.2 M phosphate buffer, pH 6.1, 1.0 MNaCl, and 3% insoluble polyvinylpyrrolidone. The homogenatewas passed through a 50 , mesh nylon cloth. The pellet was re-suspended in the above buffer less the polyvinylpyrrolidone,ground for 1 min, and filtered again. The filtered extracts werepooled and centrifuged at 10,000g for 15 min; the resultantsupematant solution was centrifuged at 100,OOOg for 1 hr. Thefinal supernatant solution was then dialyzed against twochanges of 1 % glycine, pH 6.1, for 2 hr each.The solution was then subjected to isoelectric focusing ac-
cording to the procedure of Hagland (6) to separate acidic pl(isoelectric point) and basic pl cellulase. Samples were focusedfor 18 to 24 hr at 300 v.
Determination of Ethylene Generated by Explants. Petiolesprepared as described were removed from the explants, treatedat their acropetal cut surface with either pure lanolin, or lano-lin with 10' to 10'2 M IAA. The treated petioles were sup-ported by agar in the bottom of a 50-ml flask which was thensealed with a serum cap. Ethylene measurements were deter-mined on 1-ml samples using an Aerograph 600D chromato-graph with a flame ionization detector. Column packing wasPoropak Q with 2.54 cm of activated alumina at each end ofa 0.32 cm X 71.28 cm stainless steel column (15). Sensitivityof this system was greater than 0.02 nl ethylene/ml of sample.
Ethylene Trapping. Endogenously generated ethylene fromexplants enclosed in an air-tight chamber was trapped usingthe ethylene trap, mercuric perchlorate, described by Younget al. (25).
Histological Techniques. Selected 5-mm segments removed554
www.plantphysiol.orgon June 5, 2020 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

CHANGES IN BEAN ANATOMY AND CELLULASE
Laminar Pulvinus
Zone
5mm Area UIn IAA Treatments
//...- Petiole
PetiolarPulvinus-
5mm Stem Area UsedM S
In 5ppm C2H4 Treatments
ot4' dCotyledon
FIG. 1. Bean explant showing areas treated and removed foranatomical studies or cellulase extraction.
after treatment were immediately placed in Navashin's chromicacid fixative for at least 48 hr in vacuo (9). The fixed tissue wasthen dehydrated in an ethyl alcohol, tertiary butyl alcoholseries and embedded in paraplast (9). Thin sections (6 ,u) werecut and stained with Shelhorn and Drake's sextuple stain (5).Photographs were taken on Kodachrome II, type A-35 mmfilm.
RESULTS
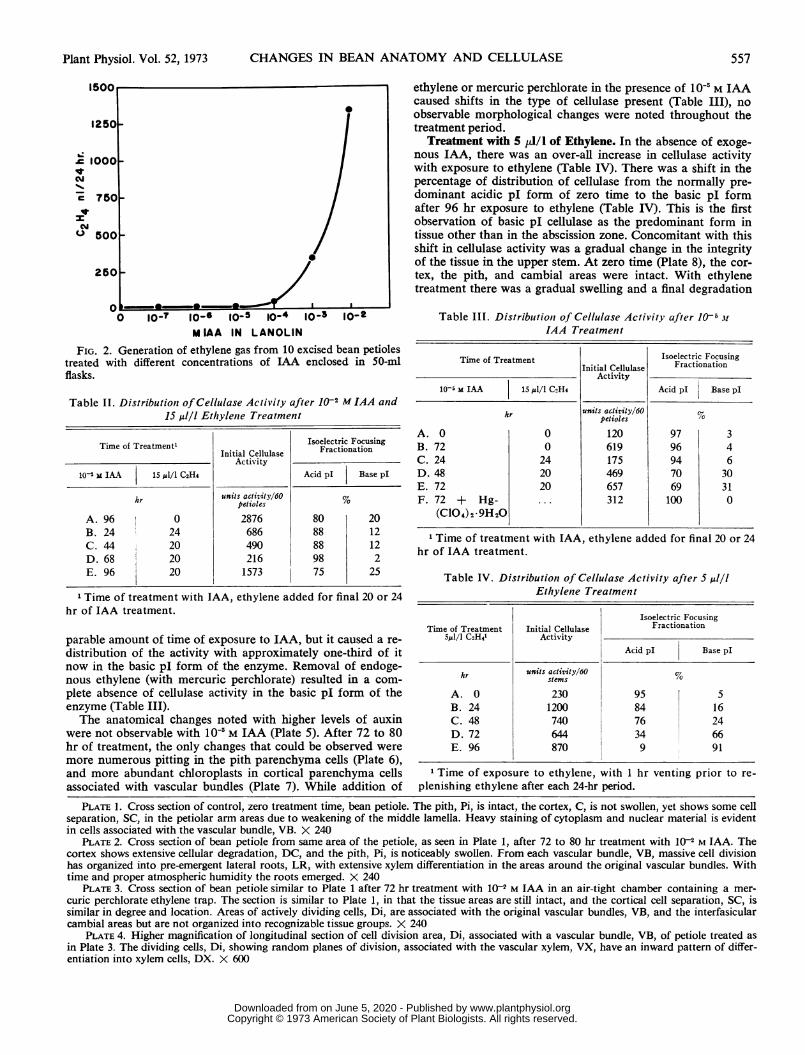
Interaction of 102 M IAA and Ethylene. High auxin levelscaused a large increase in cellulase activity as well as numerousanatomical changes. Cellulase activity in the bean petiole in-creased more than 20 times during a 96-hr exposure to auxin(Table I). During this time root initiation (3, 23), xylem forma-tion (8, 24), and cortical disruption were observed (Plates 1and 2). Xylem formation extended from the areas of differen-tiation in the lateral roots to areas in the pith associated withthe primary vascular bundles. A time course study showed thatcortical and pith tissues began to swell during the first 24 to48 hr; then the swelling ceased and cortical disruption beganas the lateral roots expanded outward.Our studies show that auxin-induced root initiation requires
ethylene. Enclosing 10-' M auxin-treated plants in air-tightchambers caused an increase in atmospheric ethylene concen-trations (Fig. 2), and the changes described in the precedingparagraph were also obtained under these conditions. Whenmercuric perchlorate was placed in the chamber to reduce en-
dogenous ethylene levels to less than 0.02 nl/ml (throughoutthe experiment), no trace of differentiation into lateral rootswas detected. Extensive cell division, however, occurred aroundvascular bundles and in the interfascicular cambium (Plate 3).Plate 4, a higher magnification of a longitudinal section throughthe bundle and cambial area, shows that xylem differentiation
accompanies the extensive cell division even though there is noorganization into lateral roots. There was also evidence of cellseparation in the cortex of the tissues. However, if the ethyl-ene was removed only during the latter 20 to 24 hr of thelonger IAA treatment periods (68-96 hr), the development andemergence of lateral roots was not suppressed. This suggeststhat the role of ethylene in adventitious root initiation is com-pleted during the early part of the IAA treatment when celldivision patterns are established.The removal of endogenously generated ethylene with mer-
curic perchlorate from explants treated with 10'2 M IAA alsohas a striking quantitative and qualitative effect on cellulase inthis tissue (Table I). There was still a large increase in cellulaseactivity compared with nontreated, zero time tissue, but thetotal cellulase activity reached only 1500 units compared toapproximately 2800 units reached in the same time periodwhen endogenously generated ethylene was not removed (Ta-ble I). The qualitative difference in 102 M IAA stimulatedcellulase activity when endogenously generated ethylene is re-moved may be even more significant since this treatment re-sulted in a complete absence of the basic pI form of cellulase(Table I). One hundred per cent of the cellulase activity foundin the tissue treated with auxin in the presence of mercuricperchlorate was found in the acidic pI form of cellulase.
Terminal addition of ethylene to the auxin treatment had nomeasurable effect on petiole anatomy during the first 44 hr ofIAA treatments (treatments B and C, Table II). However, theterminal addition of ethylene to 68 and 96 hr treatments ofIAA caused substantial swelling of the acropetal 5 mm of thepetiole due to the rapid expansion and emergence of the de-veloping lateral roots (treatments D and E, Table II). If theseenclosed plants had no exogenous ethylene added or the endog-enously generated ethylene was removed with mercuric per-chlorate during the last 24 hr of treatment, there was no changein the rate of root emergence compared to enclosed plants withor without exogenous ethylene. This acceleration is probablydue to increased atmospheric humidity or concentration ofvolatile compound(s) other than ethylene.
Addition of ethylene during any of the IAA treatment pe-riods depressed the total measurable cellulase activity (TableII) as noted by Ridge and Osborne (17). However, it did notcause any consistent change in the ratio of acidic pI to basicpl forms of cellulase.
Treatment with 10 MIAA. Treatment of bean petioles with10-' M IAA is the highest auxin concentration that can be ap-plied without stimulating ethylene generation over that ob-served for control petioles (Fig. 2). This level of auxin causeda moderate increase in the activity of the acidic pI form ofcellulase (Table III). Basic pI cellulase still accounted for lessthan 5% of the activity in this treatment. Addition of 15 ,lI/lethylene did not affect the total level of cellulase for a com-
Table I. Distribution of Cellulase Activity afterI 7J2 _f IAA Treatment
Isoelectric FocusingTime of Treatment Initial Cellulase Fractionation
10-2 m IAA ActivityAcid pl Base pI
hr inits activity/60 %
A. 0 120 97 3B. 96 2876 80 20C. 96 + Hg(C104).2 1500 100 0
9H20
555Plant Physiol. Vol. 52, 1973
www.plantphysiol.orgon June 5, 2020 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

LINKINS, LEWIS, AND PALMER
YVB
..
A. 1 ., 4.
PLATE. 1 to 4.
556 Plant Physiol. Vol. 52, 1973
www.plantphysiol.orgon June 5, 2020 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.
CHANGES IN BEAN ANATOMY AND CELLULASE
12501
c 1000
cq'e 750
Nc)o
,,/0
0
II-
5001
2560
n A s -
0 lo-T 10-6 1I sL-4N L -3IN-2
M IAA IN LANOLIN
FIG. 2. Generation of ethylene gas from 10 excised bean petiolestreated with different concentrations of LAA enclosed in 50-mlflasks.
Table II. Distributioni of Cellulase Activity after 1O2 M IAA and15 ,I/l Ethylenie Treatment
Time of Treatment' Isoelectric FocusinInitial Cellulase Fractionation
Activity10-2 M IAA 15 pI/l C2H4 Acid pI Base pI
hr units activity/60 %petiolesA. 96 0 2876 80 20B. 24 24 686 88 12C. 44 20 490 88 12D. 68 20 216 98 2E. 96 20 1573 75 25
1 Time of treatment with IAA, ethylene added for final 20 or 24hr of IAA treatment.
parable amount of time of exposure to IAA, but it caused a re-distribution of the activity with approximately one-third of itnow in the basic pl form of the enzyme. Removal of endoge-nous ethylene (with mercuric perchlorate) resulted in a com-plete absence of cellulase activity in the basic pl form of theenzyme (Table III).The anatomical changes noted with higher levels of auxin
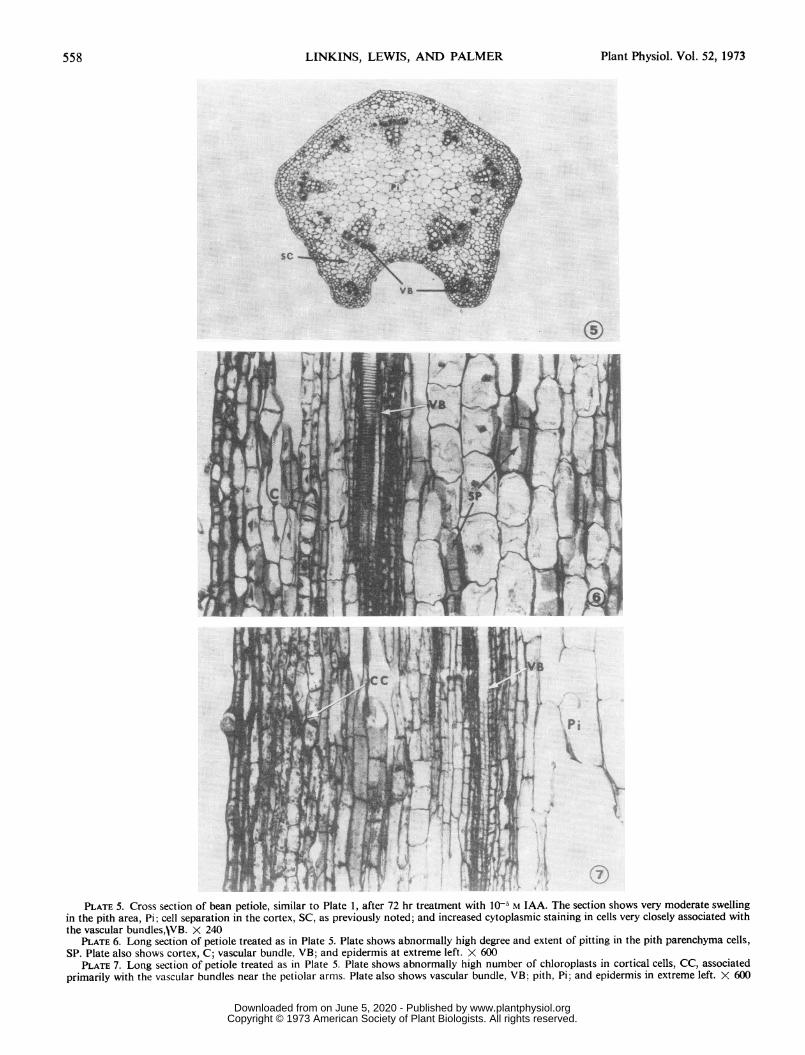
were not observable with 10-" M IAA (Plate 5). After 72 to 80hr of treatment, the only changes that could be observed weremore numerous pitting in the pith parenchyma cells (Plate 6),and more abundant chloroplasts in cortical parenchyma cellsassociated with vascular bundles (Plate 7). While addition of
ethylene or mercuric perchlorate in the presence of 1 0- M IAAcaused shifts in the type of cellulase present (Table III), noobservable morphological changes were noted throughout thetreatment period.
Treatment with 5 Atl/l of Ethylene. In the absence of exoge-nous IAA, there was an over-all increase in cellulase activitywith exposure to ethylene (Table IV). There was a shift in thepercentage of distribution of cellulase from the normally pre-dominant acidic pI form of zero time to the basic pI formafter 96 hr exposure to ethylene (Table IV). This is the firstobservation of basic pl cellulase as the predominant form intissue other than in the abscission zone. Concomitant with thisshift in cellulase activity was a gradual change in the integrityof the tissue in the upper stem. At zero time (Plate 8), the cor-tex, the pith, and cambial areas were intact. With ethylenetreatment there was a gradual swelling and a final degradation
Table III. Distributiont of Cellulase Activity after 105 mIAA Treatment
Time of Treatment Isoelectric FocusingInitial Cellulase Fractionation
Activity10-5 m IAA 15U1/l C2H4 Acid pI Base pI
hr units actirlty/60 %petioles
A. 0 0 120 97 3B. 72 0 619 96 4C. 24 24 175 94 6D. 48 20 469 70 30E. 72 20 657 69 31F. 72 + Hg- ... 312 100 0
(C104) 2-9H20
1 Time of treatment with IAA, ethylene added for final 20 or 24hr of IAA treatment.
Table IV. Distribution of Cellulase Activity after 5 ,ul/lEthylenie Treatment
Isoelectric FocusingTime of Treatment Initial Cellulase Fractionation
Spill C2H41 ActivityAcid pI Base pI
hr units actirity/60 %stems
A. 0 230 95 5B. 24 1200 84 16C. 48 740 76 24D. 72 644 34 66E. 96 870 9 91
1 Time of exposure to ethylene, with 1 hr venting prior to re-plenishing ethylene after each 24-hr period.
PLATE 1. Cross section of control, zero treatment time, bean petiole. The pith, Pi, is intact, the cortex, C, is not swollen, yet shows some cellseparation, SC, in the petiolar arm areas due to weakening of the middle lamella. Heavy staining of cytoplasm and nuclear material is evidentin cells associated with the vascular bundle, VB. X 240
PLATE 2. Cross section of bean petiole from same area of the petiole, as seen in Plate 1, after 72 to 80 hr treatment with 10-2 M IAA. Thecortex shows extensive cellular degradation, DC, and the pith, Pi, is noticeably swollen. From each vascular bundle, VB, massive cell divisionhas organized into pre-emergent lateral roots, LR, with extensive xylem differentiation in the areas around the original vascular bundles. Withtime and proper atmospheric humidity the roots emerged. X 240
PLATE 3. Cross section of bean petiole similar to Plate 1 after 72 hr treatment with 10 2 M IAA in an air-tight chamber containing a mer-curic perchlorate ethylene trap. The section is similar to Plate 1, in that the tissue areas are still intact, and the cortical cell separation, SC, issimilar in degree and location. Areas of actively dividing cells, Di, are associated with the original vascular bundles, VB, and the interfasicularcambial areas but are not organized into recognizable tissue groups. X 240
PLATE 4. Higher magnification of longitudinal section of cell division area, Di, associated with a vascular bundle, VB, of petiole treated asin Plate 3. The dividing cells, Di, showing random planes of division, associated with the vascular xylem, VX, have an inward pattern of differ-entiation into xylem cells, DX. X 600
-I Iv K=--
Plant Physiol. Vol. 52, 1973 557
www.plantphysiol.orgon June 5, 2020 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.
LINKINS, LEWIS, AND PALMER
t~~~~ it.S WiPLATE 5. Cross section of bean petiole, similar to Plate 1, after 72 hr treatment with 10O5 M IAA. The section shows very moderate swelling
in the pith area, Pi; cell separation in the cortex, SC, as previously noted; and increased cytoplasmic staining in cells very closely associated withthe vascular bundles,\VB. X 240
PLATE 6. Long section of petiole treated as in Plate 5. Plate shows abnormally high degree and extent of pitting in the pith parenchyma cells,SP. Plate also shows cortex, C; vascular bundle, VB; and epidermis at extreme left. X 600
PLATE 7. Long section of petiole treated as in Plate 5. Plate shows abnormally high number of chloroplasts in cortical cells, CC, associatedprimarily with the vascular bundles near the petiolar arms. Plate also shows vascular bundle, VB; pith, Pi; and epidermis in extreme left. X 600
Plant Physiol. Vol. 52, 1973558
www.plantphysiol.orgon June 5, 2020 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

CHANGES IN BEAN ANATOMY AND CELLULASE
>N,_,: >- ; {'-'
b~~. w t>ur!ep ' - ". '
PLATE 8. Cross section of bean stem, zero time treatment. Plate shows pith, Pi; cortex, C; vascular bundles, VB; and interfasicular cambium,IC. All tissues are intact. X 240
PLATE 9. Cross section of bean stem, similar to one in Plate 8, after 72 hr of treatment with 5 ,uI/I ethylene gas. There is extensive degradationof the pith, Pi; separation of pith cells from the interfasicular cambium, IC; and moderate swelling of the cortex, C. Cells closely associated withthe cambial cells do not show as extensive cytoplasic staining as noted in control. X 240
of the cortical and pith tissues that was initiated around thecambium cells similar to the changes observed by Radin andLoomis (16) in radish roots. After 72 to 80 hr treatment with5 1il/l ethylene, degradation of pith integrity was extensive(Plate 9). There was no evidence for initiation of cell divisionor lateral root formation as noted in other ethylene treatedplants by Zimmerman and Hitchcock (26).
DISCUSSION
Herbicidal concentrations of IAA are known to induce rootinitiation in stems. Addition of ethylene has been shown to actin a synergistic manner with IAA-induced root initiation instems (7, 11, 14), but that ethylene plays such an integral rolein the initial cellular organization of the root primordia has notbeen previously reported. Although auxin is capable of induc-ing cell division and xylem differentiation in this tissue in theabsence of endogenous ethylene, there appears to be no orderto this cell division and hence no lateral root initiation. On the
other hand, when ethylene was added to the tissue in the ab-sence of exogenous auxin, there appeared to be little or no celldivision and only a breakdown of certain tissues. This wouldindicate that auxin causes cell division, ethylene causes cellbreakdown, and a combination of the two cause lateral rootformation.Accompanying these anatomical changes are some specific
changes in the levels and forms of cellulase present. Bothauxin and ethylene stimulate the over-all level of cellulasepresent. The stimulation due to auxin treatment results pri-marily in an increase in the acidic pl form of the enzyme.When both hormones were present in the tissue at the sametime, both forms of the enzyme accumulated. Since it is ob-vious that the cell walls are undergoing substantial modifica-tion and breakdown in these treatments, it is reasonable to as-sume that cellulase plays an important role in these hormonallyinduced changes. Furthermore, it appears that the acidic pIform of the enzyme is more closely associated with the differ-entiation of the tissue through such cellular changes as cell di-
Plant Physiol. Vol. 52, 1973 559
www.plantphysiol.orgon June 5, 2020 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

LINKINS, LEWIS, AND PALMER
vision and xylem differentiation, while the more basic pI formof the enzyme seems to be more closely associated with cellwall degradation as seen in cortical and pith degradation in thebeanl petiole and in abscission zone breakdown in the bean.This hypothesis is supported by the observations of severalworkers that have shown ethylene treatments cause a mobili-zation of cellulase from cytoplasm into the cell wall. Abelesand Leather (1), Ferrari (4), and unpublished results from thislaboratory have observed this mobilization through infusionexperiments. Jutte and Wardrop (10) showed the generalizeddegradation of plant cell walls by fungal cellulase as occurringby an over-all infusion of the cellulase into the wall eitherthrough pits or over-all surface contact. Thus, the general hy-drolysis of cell walls as observed in bean plants suggests a ma-
jor change in mobilization of plant cellulase either through thesynthesis of new forms or changes in membrane permeabilityto existent forms of cellulase and other structural hydrolases(2).
Ackntowledgment-We thank H. Gardner for his excellent technical assistance.
LITERATURE CITED
1. ABELES, F. G. AN-D G. R. LEATHER. 1971. Abscission: control of cellulasesecretion by ethylene. Planta 97: 87-91.
2. DATKo, A. H. AN-D G. A. 'MACLACHLAN. 1968. In(loleacetic acid and the syn-
thesis of glucanases and pectic enzymes. Plant Physiol. 43: 735742.3. FAN\, D. F. AND G. A. MACLACHLAN. 1966. Control of cellulase activity by
indoleacetic acid. Can. J. Bot. 44: 1025-1034.4. FERRARI, T. E. 1972. Effect of auxin and ethylene on the activity and com-
partmentation of cellulase. Plant Physiol. 49: S2,5. GuRE, E. 1965. The Rational Use of Dyes in Biology and General Staining
AMethods. The Williams & Wilkins Co., Baltimore. p. 312.6. HAGLAND, H. 1967. Isoelectric focusing in natural pH gradients-a technique
of growing importance for fractionation and characterization of proteins.Sci. Tools 14: 17-23.
7. HITCHCOCK, A. E. AND P. W. ZIMMERMAN. 1940. Effects obtained withmixtures of root-inducing and other substances. Contrib. Boyce ThompsonInst. Plant Res. 11: 143-160.
8. JACOBS, W. P. 1954. Acropetial auxin transport and xylem regeneration: A
quantitative study. Aimer. Naturalist 88: 327-337.
Plant Physiol. Vol. 52, 1973
9. JEN-SEN, W. A. 1962. Botanical Histochemistry, Principles and Practice.W. H. Freeman Co., San Francisco. pp. 55-83.
10. JUTTE, S. M. AND A. B. WNARDROP. 1970. Morphological factors relating to
the degradation of wood fibers by cellulase preparations. Acta Bot. Neerl.19: 906-917.
11. KRISHNAMOORTHY, H. N. 1970. Promotion of rooting in mung bean hypocotylcuttings with ethrel, an ethylene releasing compound. Plant Cell Physiol.11: 979-982.
12. LEWIS, L. N., F. T. LEW, P. D. REID, AND J. E. BAR-NES. 1972. Isozymes ofcellulase in the abscission zone of Phaseolus vulgaris. In: D. J. Carr, ed.,Plant Growth Substances, 1970. Springer Vrerlag, Berlin. pp. 234-239.
13. LEWIS, L. N. AND J. E. VARNZER. 1970. Synthesis of cellulase during abscissionof Phaseolus vulgaris leaf explants. Plant Physiol. 46: 194-199.
14. MICHENER, H. D. 1935. Effects of ethylene on plant growth hormone. Science82: 2136-2137.
15. PALMER, R. L., H. Z. HIELD, AND L. N. LEWIS. 1969. Citrus leaf and fruitabscission. In: H. D. Chapman ed., Proceedings First International CitrusSymposium. University of California, Riverside. pp. 1135-1143.
16. RADIN, J. W. AND R. S. LooMis. 1969. Ethylene and carbon dioxide in thegrowth and development of cultured radish roots. Plant Physiol. 44: 1584-1589.
17. RIDGE, I. AND D. J. OSBORN-E. 1969. Cell growth and cellulase: Regulation byethylene and indole-3-acetic acid in shoots of Pisum sativum. Nature 223:318-319.
18. SHELDRAKE, A. R. 1969. Cellulase in latex and its possible significance incell wall differentiation. Planta 89: 82-84.
19. SHELDRAKE, A. R. 1970. Cellulase and cell differentiation in Acer pseudo-plantanus. Planta 95: 167-178.
20. SHELDRAKE, A. R. 1970. Cellulase in latex and its possible significance in
cell differentiation. J. Rubber Inst. 'Malaya 23: 74-75.21. SHELDRAKE, A. R. AND G. J. MOIR. 1970. A cellulase in Hevea latex. Physiol.
Plant. 23: 267-277.22. SHELDRAKE, A. R. AND D. H. NORTHCOTE. 1968. Some constituents of xylem
sap and their possible relationship to xylem differentiation. J. Exp. Bot.19: 681-689.
23. THIMAN-N, K. V. AND F. W. WENT. 1934. On the chemical nature of the root-forming hormone. Proc. Kon. Ned. Akad. Wetensch. Ser. B. 37: 456-459.
24. WETMORE, R. H. 1955. Differentiation of xylem in plants. Science 121: 626-627.
25. YOUNG, R. E., H. K. PRATT, AND J. B. BIALE. 1952. 'Manometric determina-tion of low concentrations of ethylene. Anal. Chem. 24: 551-555.
26. ZIMMERMAN, P. W. A_ND A. E. HITCMCOCK. 1933. Initiation and stimulation ofadventitious roots caused by unsaturated hydrocarbon gases. ContribBoyce Thompson Inst. Plant Res. 5: 351-369.
560
www.plantphysiol.orgon June 5, 2020 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.







![The Barley Uniculme4 Gene Encodes a BLADE-ON- PETIOLE-Like ... · The Barley Uniculme4 Gene Encodes a BLADE-ON-PETIOLE-Like Protein That Controls Tillering and Leaf Patterning1[OPEN]](https://static.fdocuments.us/doc/165x107/5ed141dccd86a73bbf4f388b/the-barley-uniculme4-gene-encodes-a-blade-on-petiole-like-the-barley-uniculme4.jpg)










