
Las moscas de las frutas, Ceratitis capitata y Anastrepha fraterculus ...
Upload
luis-francoCategory
view
213download
1
Insect Biochem., Vol. 9, pp. 31 to 38. 0020-1700/79/0201-0031 $02.00 © Pergamon Press Ltd. 1979. Printed in Great Britain
HISTONE H4 FROM THE FRUIT FLY CERA TITIS CAPITATA. PURIFICATION AND CHARACTERIZATION
Luls FRANCO, ROSA MARIA NIETO-SANDOVAL and JULIAN PERERA
Department of Biochemistry, Faculty of Sciences, Universidad Complutense, Madrid-3, Spain
(Received 22 May 1978)
A~traet--Histone H4 has been isolated, in a pure form, from Ceratit& capitata pharate adults. Its molecular weight is very similar to that of calf thymus H4 histone. The N-terminal residue is blocked and its amino acid composition is similar to that of homologous histones. Four peptides have been isolated from the Ceratitis capitata histone H4, and characterized. Suggestions are made about possible regions in the total sequence which gave rise to them.
Key Word Index: Histones, histone H4, Ceratitis capitata
I N T R O D U C T I O N
SINCE THE discovery o f the repeating structure of chromatin (HEwISH and BURGOYNE, 1973; OLIr~S and OLINS, 1974) a great deal of evidence has accumulated which has led to the conclusion that an octamer formed by two copies o f each H2A, H2B, H3 and H4 histones is wrapped by a well defined length o f D N A (140 base pairs) (NOLL and'KoRNBERG, 1977). The fifth histone, H1, is located outside this nucleosome core and it is not clear how it is associated with the nucleosome.
Al though some recent studies on X-ray diffraction (FiNch et. al., 1977) and neutron scattering (SUAc et aL, 1977) of nucleosome cores have led to a better definition of their shape and size, as well to a more precise location o f the core D N A , the fine details of internal structure in nucleosomes are still obscure.
The precise description o f nucleosome core structure requires an accurate knowledge of the histone regions which are involved in histone-histone or histone--DNA interaction. This knowledge must be, of course, based on that of histone primary structures. Very recently, BrHta et al. (1977) have investigated the sites of interaction between histones H3 and H4, and they have concluded, that the regions important for complex formation are those formed by residues 42-120 in histone H3 and residues 38-102 in histone H4. These studies are of interest because there is an increasing body of evidence that a tetramer formed by these histones may act as a 'kernel ' around which D N A supercoils, in a manner that mimics its native supercoiling, without the intervention o f the other two histones (CAMERIrqI-OTERO et al., 1976).
On the other hand, a contribution to the precise understanding of these relationships among histones in the nucleosome core could be provided by the primary structure o f histones. To determine whether minor differences in the primary structures o f the arginine-rich histories from lower eukaryotes may alter the interaction pattern between them, a study has been carried out in this paper on the purification of the historic H4 from the fruit fly Ceratitis capitata and the isolation of several tryptic peptides from it.
31
M A T E R I A L S AND M E T H O D S
Materials
Sephadex G- 100 (Pharmacia Fine Chemicals AB, Uppsala Sweden), Bio-Gel P-IO Bio-Rad Laboratories, Richmond, Calif., U.S.A.) and Chromobeads Type-P (Technicon Chemicals SA, Belgium) were used in purification and fractionation steps.
Trypsin, twice recrystallized (Worthington Biochemical Co., Freehold, N.J., U.S.A.) and treated with l-chloro-4- phenyl-3-tosylamidobutan-2-one (TPCK), was used for the digestion. Soy-bean trypsin inhibitor was also obtained from Worthington Biochemical Co.
Citraconic anhydride was purchased from Pierce. Rockford, Illinois, U.S.A., and ninhydrin from Sigma Chemical Co., St. Louis, Mo., U.S.A.
Rearing of insects
Culturing of Ceratitis capitata Wied. was carried out as described elsewhere (Muylfio et al., 1970).
Preparation of crude H4
Histone H4 was isolated from Ceratitis capitata pharate adults by extracting crude chromatin with 10~o (w/v) guanidinium chloride in 75~ ethanol, according to the method previously described (FRANCO et aL, 1974). The acetone precipitation of the extract yielded a crude H4 preparation (FRANCO et aL, 1974).
Purification of H4
The final purification of histone H4 was achieved by gel filtration. Crude H4 was dissolved in 0.01 M acetic acid to give a concentration of about 2 mg/ml. After centrifuging (2000 g, 15 min) to remove any insoluble material, the supernatant was loaded onto a column (1.6 cm i.d. × 100 cm) of Sephadex G-100, equil!brated with 0.01 M acetic acid. The elution was carried out with the same solvent. Fractions of 3.6 ml were collected at a flow rate of 30 ml per hr. The protein content of the fractions was spectrophotometrically measured at 230 and 280 nm and the protein-containing fractions were pooled and lyophilized.
Chemical modification
Histone H4 was dissolved in 8M urea to give a concentration of 10 mg/ml, and the pH was adjusted to 8.7 with solid sodium carbonate. Citraconic anhydride was added in 15 gl aliquots with constant stirring and the pH was kept at 8.3-8.8 by adding 5 M sodium hydroxide. On

32 LUIS FRANCO, ROSA MARIA NIETO-SANDOVAL AND JULIAN PERERA
completion of the reaction the mixture was dialyzed against dilute ammonium hydroxide, pH 8.5, and the citraconylated histone was recovered by lyophilization.
Enzymatic digestion
Citraconylated histone H4 was digested with TPCK- treated trypsin (1:80, w/w = trypsin/ histone) in 0.3 M ammonium bicarbonate, pH 8.0, at 37°C for 3 hr. The reaction was then stopped by adding a 1.5-fold excess (w/w) of soy-bean trypsin inhibitor.
Fractionation of the peptide mixtures
Tryptic peptides were submitted to an initial fractionation by gel filtration on a Bio-Gel P-10 (100--200 mesh) column (1.6 cm i.d. x 80 cm), equilibrated with 0.05 M ammonium bicarbonate, pH 8.9. The column was eluted with the same buffer, at 10 ml per hr; 2 ml fractions were collected. The protein content of the eluate was measured as before, and the appropriate fractions were pooled and lyophilized.
These fractions were dissolved in water at pH 4.5 and incubated at 45°C for 6 hr, to reverse the blocking reaction and recover the unmodified peptides. They were then loaded onto 1 cm i.d. x 80 cm columns of sulphonated polystyrene resin (Technicon Chromobeads P-type) previously equilibrated with the first buffer (see below). Peptides were eluted at 40' C, at a flow rate of 0.2 ml pei" rain as follows: 1st 84 ml of 0.2 M pyridine-acetate (pH 3.1); 2nd 84 ml of 2.0 M pyridine-acetate (pH 5.1); 3rd 84 ml of 2.0 M pyridine- acetate (pH 6.5). The elution of peptides was followed by the ninhydrin method of Hms et al. (1956) after alkaline hydrolysis of 0.2 ml aliquots.
Further separation and purification of peptides was achieved, when necessary, by high voltage preparative paper electrophoresis at pH 2.1. The samples were dissolved in 0.02 M HCI and applied to Whatman No. 1 paper. Electrophoresis was carried out in formic acid-acetic acid-water (1:4:45, pH 2.1) at 2000 V during 30 min. Peptides were visualized in two side-vertical strips by using the ninhydrincadmium reagent (HEATHCOTE and HAWORTH, 1969). Peptides were eluted from the central strip by cutting the peptide-containing areas into small pieces and incubating them at 37°C with 0.1 M acetic acid.
Analytical methods
Disc electrophoresis was carried out in 20% polyacrylamide gels according to JOHNS (1967) and urea polyacrylamide gels were run following the procedure of Pa~vtm and CHALKLEY (1969). SDS-polyacrylamide slab gels were run following the method of LAEMMLI (1970).
The N-terminal residue was assayed with dansyl chloride (GRAY, 1972; PEREZ-ARANDA, 1975).
Hydrolysis of protein or peptide samples was carried out at 105°C with 5.7 M HCI containing 0.1% (w/v) phenol in evacuated sealed tubes for 24 hr. Amino acid analyses were performed with a Durrum amino acid analyzer model D-500.
High voltage electrophoresis at pH 2.1 was carried out using formic-acetic acid buffer on Whatman No. 1 paper. Peptides were detected with the ninhydrincadmium reagent (HEATHCOTE and HAWORTH, 1969).
R E S U L T S A N D D I S C U S S I O N
F i g u r e 1A shows the electrophoretic pattern of crude histone H4, prepared as described earlier (FRANCO et al., 1974). It is evident from this figure that histone H4 is contaminated to some extent by a material which did not enter the gel. The final purification of histone H4 was achieved by gel-filtration as described under Materials and Methods. Crude H4 was resolved into two peaks (Fig. 2). Peak I was mainly composed of the high molecular
Table 1. Amino acid composition of histone H4 from Ceratitis capitata, calf thymus and pea
Amino acid C. capitata* Calf thymus¢ Pea++
Aspartic acid 5.4+ 0.2 4.9 4.9 Threonine 6.9 + 0.2 6.9 6.9 Serine 2.2 ___ 0.1 2.0 2.0 Glutamic acid 7.2 + 0.1 5.9 5.9 Proline 0.8 + 0.1 1.0 1.0 Glycine 14.5 + 0.2 16.7 16.7 Alanine 7.4 + 0.2 6.9 6.9 Half-cystine - - - - - - Valine 8.4 + 0.2 8.8 7.8 Methionine 0.7 ___ 0.1 1.0 1.0 Isoleucine 5.5 + 0.2 5.9 6.9 Leucine 7.3 ___ 0.6 7.8 7.8 Tyrosine 3.5 _ 0.2 3.9 3.9 Phenylalanine 2.4 + 0.1 2.0 2.0 Histidine 2.2 + 0.2 2.0 2.0 Lysine 10.0+0.7 10.8 9.8 Arginine 15.5 + 0.9 13.7 14.7 Basic/acidic 2.2 2.5 2.5 Lys/Arg 0.6 0.8 0.7
Compositions are given in mole ~o. * Two histone samples were independently hydrolyzed and
four analysis were performed with each hydrolyzate. This column gives the average of the eight determinations + standard deviation. The values for serine and threonine have been extrapolated to zero-time.
t Based on the amino acid sequence (DELANGE et al., 1969a).
:~ Based on the amino acid sequence (DELANGE et al., 1969b).
weight material, while peak II run on 20% polyacrylamide gels as a single band (Fig. 1) and its mobility was identical to that of calf thymus H4 both in urea gel (Fig. 3) and in SDS gel (Fig. 4). This material will be referred to as pure H4.
The nature of the protein recovered from peak I is still unknown. The possibility o f its being an aggregated form of H4 can be ruled out, because its amino acid composit ion is quite different. This material is now being studied.
The amino acid composit ion of Ceratitis capitata H4 is given in Table 1, which also includes the composit ion of the homologous histones from calf thymus and pea, both calculated from their known amino acid sequences (DELANGE et al., 1969a. b). The high degree of similarity in the composit ion of these three histones is noteworthy. The arginine content of the Ceratitis histoneis closer to the value found in the pea histone than that o f the mammal ian H4. Assuming that the total number of residues in the fly histone is 102, as the similarity in electrophoretic mobility between Ceratitis and thymus histones in SDS- polyacrylamide gels may suggest, it may be concluded that Ceratitis H4 possesses 15 arginyl residues. The pea histone also possesses an extra arginyl residue that substitutes for the lysine 77 in mammal ian sequences. It is not clear whether this substitution also takes place in the Ceratitis histone, because the lysine level, although somewhat lower, may not be significantly different from that in thymus H4. It is also possible that two additional differences in composit ion occur between fly and thymus histones (Table 1). The fly histone has one to two less glycyl residues and perhaps

l lai l lbl l lcl
A B A B C Fig. 1. 20~0 polyacrylamide gel electrophoresis of (A) crude H4; note the presence of a high molecular weight material that does not enter the gel. (B) High molecular weight material removed from H4 by gel- chromatography (fraction I in Fig. 2). (C) Pure H4 (fraction II in Fig. 2). Migration is from top to bottom.
Fig. 3. Urea-polyacrylamide gel electrophoresis of (A) calf thymus arginine-rich histones. (B) Ceratitis capitata pure histone H4. Migration is from top to bottom.
Fig. 4. SDS-polyacrylamide slab gel electrophoresis of(A) calf thymus whole histone. (B) Ceratitis capitata histone H4. (C) Calf thymus histone H4. Migration is from top to bottom.

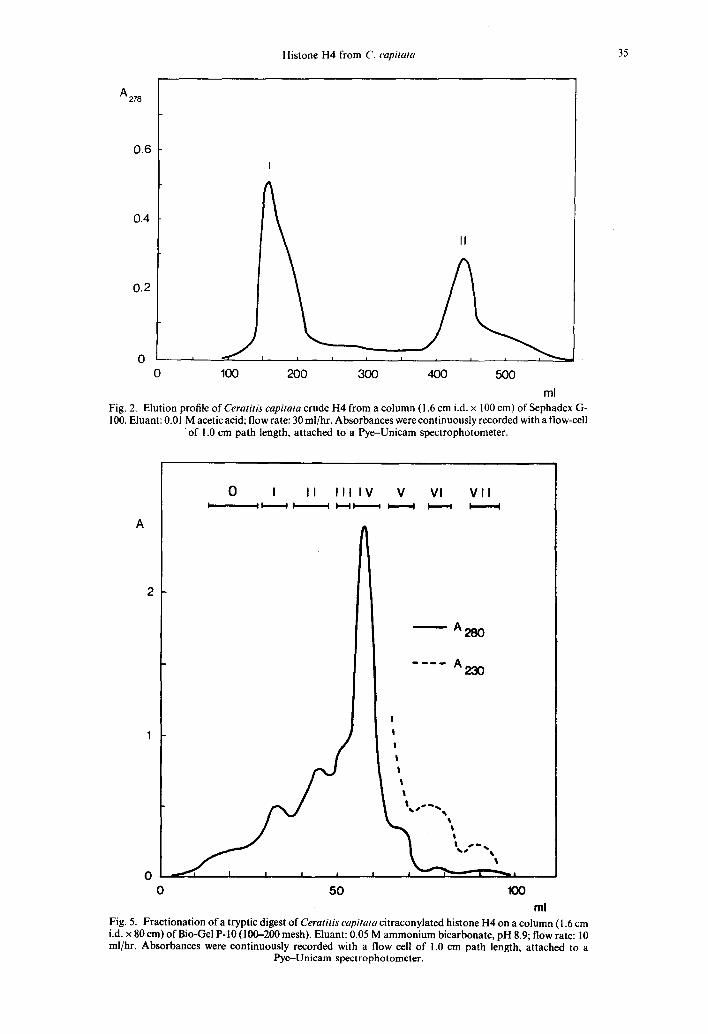
Histone H4 from C. capitata 35
A 278
0.6
0.4
0.2
0 i i
0
I
J I I I I
100 200 300
II
4OO 5OO
ml Fig. 2. Elution profile of Ceratitis capitata crude H4 from a column (1.6 cm i.d. x 100 cm) of Sephadex G- 100. Eluant: 0.01 M acetic acid; flow rate: 30 ml/hr. Absorbances were continuously recorded with a flow-cell
• of 1.0 cm path length, attached to a Pye-Unicam spectrophotometer.
A
2
0 0 50
0 I I I I I I IV V Vl V l l I I I I I I I " ~ I I I I I I I I
A280
230
!
i
!
t t t I
I I I ~ I - " q ~ l
100 ml
Fig. 5. Fractionation o f a tryptic digest of Ceratitis capitata citraconylated histone H4 on a column ( 1.6 cm i.d. x 80 era) o f Bio-Gel P-10 (100-200 mesh). Eluant: 0.05 M a m m o n i u m bicarbonate, pH 8.9; flow rate: 10 ml/hr. Absorbances were continuously recorded with a flow cell of 1.0 cm path length, attached to a
Pye-Unicam spectrophotometer.

36 Luls FRANCO, ROSA MARIA NIETO-SANDOVAL AND JULL~N PERERA
Table 2. Isolation and properties of tryptic peptides from Ceratitis capitata histone H4 (see text)
Bio-gel chromatography IEC HVPE Fraction Elution vol. Fraction Elution vol. Buffer Spots Mobility Peptide
(ml) (ml) .(cm) designation
f m
I 30-38 - - M 170-180 3rd pH 6.5
(- 11 38--48 - -K 125-170 2nd pH 5.1
IV 53--64
V 65-72
t- / - -K 130-140 2nd pH 5.1 - - L 140-145 2nd pH 5.1 - -M 145-160 2nd pH 5.1 - -N 160-170 2nd pH 5.1
- - 110-120 2nd pH 5.1 120-135 2nd pH 5.1
IV(K-N)
V(I-J)
1 6.8 I-M
--1 - -2 --3
- -5
--1
1 5.8 II-K
5,1 IV(K-N)5
7.2 V(l-J)4
one more glutamyl residue. RODRIGUEZ-ALFAGEME et al. (1974) also repor ted an excess of glutamic acid and a deficiency of glycine in Drosophila melanogaster H4 but the differences were not as clear as those repor ted here, and they could not draw any definite conclus ion f rom their results. In fact, our own da ta only allow the above differences to be drawn tentatively, especially tha t of glutamic acid content .
The actual differences between Ceratitis and
m a m m a l i a n H4 his tones can only be unambiguous ly ascertained by sequencing. The appl ica t ion of ei ther manua l or au tomated E d m a n ' s degrada t ion to the intact his tone was not possible in this ins tance because Ceratitis H4 proved to be blocked at the amino terminal end, as revealed by the dansyl chloride method.
Tak ing into account the high percentage oflysyl and arginyl residues, tryptic cleavage after b locking the
Table 3. Amino acid composition of tryptic peptides from the Ceratitis capitata histone H4
I-M II-K Amino acid A B A
Peptide IV-(K-N)5 V-(I-J)4
B A B A B
Asx 7.4 0.7 14.2 2.2 4.1 0.2 4.7 0.2 Thr 10.9 1.1 7.0 1.1 . 2.4 0.1 2.3 0.1 Ser 2.4 0.2 3.3 0.5 6.8 0.3 6.4 0.3 Glx 6.9 0.7 6.9 1.1 6.3 0.3 5.3 0.3 Pro 2.1 0.2 5.9 0.9 . . . . Gly 7.0 0.7 9.2 1.4 10.8 0.5 7.5 0.4 Ala 11.3 1.1 7.9 1.2 18.3 0.8 5.3 0.3 Val 9.8 1.0 3.2 0.5 3.8 0.2 14.0 0.7 Met 3.8 0.4 2.4 0.4 . . . . lie 2.5 0.2 17.5 2.7 3.4 0.1 2.3 0.1 ~ u 6.7 0.7 3.3 0.5 17.4 0.8 15.9 0.8 Tyr 5.4 0.5 2.6 0.4 - - - - 1.6 0.1 Phe 1.7 0.2 1.7 0.3 - - - - 1.6 0.1 His 1.0 0.1 0.9 0 . 1 - - - - 2.2 0.1 Lys 10.9 1.1 7.5 1.2 4.0 0.2 6.1~
0.6 6-N-Me-Lys 6.0 Arg 10.2 1.0 6.4 1.0 22.7 1.0 18.9 1 . 0
Composition* DT2EGA2V2 - DzTEPGA- ALR VLKR MLYK2R 2 IaKR
Columns A give the composition of peptides in moles %. Columns B give the number of residues calculated from Arg = 1. * The one-letter code has been used.

Histone H4 from C. capitata 37
lysyl residues with citraconic anhydride was the method of choice for obtaining peptides of convenient size.
The peptide mixture was initially fractionated by Bio-Gel P- 10 chromatography, as shown in Fig. 5. The additional use of ion-exchange chromatography (IEC) yielded a series of fractions which were analyzed by high voltage paper electrophoresis (HVPE), as summarized in Table 2. Two of these fractions gave single spots (I-M and II-K). The electrophoretic patterns of fractions IV-K to IV-N were complex but similar to some extent. They were pooled and further fractionated by preparative high voltage paper electrophoresis and the peptide corresponding to spot 5 was isolated. In a similar way (Table 2) the peptide V(I-J)4 was obtained. The amino acid composition of the four pure peptides are shown in Table 3.
Assuming a quantitative blocking of lysyl residues and a good yield in tryptic cleavage, the presence of a single arginyl residue in every peptide is to be expected, although the C-terminal peptide would, of course be an exception. For this reason, Table 3 includes the molar composition of the peptides normalized to one arginyl residue.
To allow a preliminary identification of the isolated peptides, parallel experiments were carried out with calf thymus H4 as starting material. In this way, the four peptides whose composition is shown in Table 3 were identified on the following basis: (a) comparison of their compositions with those of the calf thymus H4 peptides, both experimental and calculated from the known mammalian sequence; (b) comparison of their elution behaviour from both Bio-Gel and ion exchange columns; (c) electrophoretic behaviour. Peptides I-M, II-K, IV(K-N)5 and V(I-J)4 are thus thought to proceed respectively from residues 79-95, 24-35, 37-39 and 20-23, in the histone sequence. It is to be noted that a definite identification will only be achieved by obtaining the whole sequence of the peptides, but the extremely conservative nature of
histone H4 may make the above identification reasonable.
Provided that the primary structure of the fly histone is very similar to that of the calf thymus H4, the identified peptides from the Cerat i t i s protein are tentatively located as shown in F!g. 6. It is to be noted that they would come from the more conservative regions of the histone, i.e. those in which no variation has ever been detected. It is thus conceivable that the presumed variations in Cerat i t i s H4 occur in regions other than those identified.
The occurrence of 6-N-methyllysine in peptide V(I-J)4 may be of special interest. Provided that it actually came from the residues 20-23 of the fly histone, it would indicate that this covalent post- synthetic modification takes place at the same residue--lysine 20---as in calf thymus H4.
If the above assumptions are correct, the peptides, whose isolation and purification have been reported here, may come from the N-terminal and C-terminal regions of the histone molecule. They represent about one third of the histone molecule and were identical to the corresponding ones from the calf thymus histone in composition, electrophoretic and chromatographic behaviour, and even in the degree to which the lysine is methylated. This suggests that the variations predicted from the composition of the Cerat i t i s histone occur in the other two thirds of the molecule. It is noteworthy that the variations so far detected in this histone (DELANGE et al., 1969a. b: TYROU et al., 1976) occur in the most hydrophobic region and this might also be the case with the Cerat i t i s histone. Nevertheless, these regions are implicated in the interactions between H4 and H3 (BrHM et al., 1977) which, as previously noted, play a fundamental role in the maintenance of the nucleosome structure, this being well preserved during the biological evolution. However, all these facts are consistent with an amino acid variation in the inner part of the globular region which does not contribute to histone-histone interaction. Further studies on this matter are now in progress.
Ac-SER-GLYLAR~-GLY-LYS-GLY-~LY-LYS-CLY-LEU-GLY-LYS-GLY-~LY -
ALA-LYS-ARG-HIS-ARC.-LYS (Me)-VAL-LEU-ARG-~ $P-A,qN- ILE-CJ,N-C~LY-
< v(i-,~) 4 > <
ILE-THR- LY,q- PRO-ALA- ILE-ARG- ARG-LEU-ALA-ARC~- ARG-GLY-GLY-VAn-
- - II-K ) ~IV (K-N) 5--->
LYS- ARG- ILE- SER-C~LY- LEU- I LE-TYR-GLU-GLU-THR-ARC~-C~LY-VAL- LEU-
I.B. 9 . 1 - - c
LYS-VAL- PHE- LEU-GLU-ASN-VAL- I LE- ARG-ASP- ALA-VAL-THR-TYR-THR-
Ile Cys
GLU- HI S-ALA- LYS- ARG- LYS-THR-VAL-TH~-ALA-MET- ASP-VAL-VAL-TYR-
Arg < I-M
ALA- LEU- LYS- ARG-GLN- GLY- ARG-THR- LEU-TYR-C~LY- DHE-GLY-~LY
>
Fig. 6. Assignment of the purified peptides from the Ceratitis histone H4. The sequence shown in capital letters is that of calf thymus H4 (DELANGE et al.. 1969a). The three variations known up to date are shown under the corresponding residues of the thymus histone: cysteine-73 in Psamechinus miliaris (TYROU et al.,
1976) and isoleucine-60 and arginine-77 in pea (DELANGE et al., 1969b).

38 Luls FRANCO, ROSA MARia NIETO-SANDOVAL AND JULIAN PERERA
Acknowledgements--This work has been supported by a grant from the Comisi6n Asesora de Investigaci6n Cientifica y T6cnica (Spain). We are also very indebted to Prof. A. M. MUNICIO for his invaluable suggestions and helpful criticism, and to Mr. G. GONZALEZ de BUITRAGO for the amino acid analyses.
R E F E R E N C E S
BOHM L., HAYASHI H., CARY P. D., Moss, T., CRANE- ROmNSON C. and BRADBURY E. M. (1977) Sites of histone/histone interaction in the H3.H4 complex. Eur. J. Biochem. 77, 487-493.
CAMERINI-OTERO R. D., SOLLNER-WEBB B. and FELSENFELD G. (1976) The organization of histones and DNA in chromatin: evidence for an arginine-rich histone kernel. ('e//8, 333--347.
DELANGE R. J., FAMBROUGH D. M., SMITH e. L. and BONNER J. (1969a) Call" and pea histone--IV, 11. The complete amino acid sequence of calf thymus histone I V: presence of 6-N-acetyllysine. J. biol. Chem. 244, 319-334.
DELANt3E R. J., FAMBROUGH D. M .. SMITH E. L. and BONNER J. (1969b) Calf and pea histone--IV, 111. Complete amino acid sequence of pea seedling histone IV; comparison with the homologous calf thymus histone. J. biol. Chem. 244, 566%5679.
FINCH J. T., LUTTER L. C., RHODES D., BROWN R. S., RUSHTON B., LEVITT M. and KLUO A. (1977) Structure of nucleosome core particles of chromatin. Nature 269, 29-36.
FRANCO L., MONTERO F.. NAVLET J. M., PERERA J. and RoJO M. C. (1974) Histones from the fruit fly Ceratitis capitata. Isolation and characterization. Eur. J. Biochem. 48, 53-61.
GRAY W. R. (1972) End-group analysis using dansyl chloride. In Methods in Enzymolog.v (Ed. by Hits C. H. W. and TIMASHEEF S. N.) Vol. XXV, pp. 121-138. Academic Press, New York.
HEatncote J. G. and Hawor tn C. (1969) An improved technique for the analysis of amino acids and related compounds on thin layers of cellulose--ll. The
quantitative determination of amino acids in protein hydrolysates J. Chromat. 43, 84-87.
HEWlSH D. R. and BURGOYNE L. A. (1973) Chromatin substructure. The digestion of chromatin DNA at regularly spaced sites by a nuclear deoxiribonuclease. Biochim. biophys. Res. Commun. 52, 504-510.
HIRS C. H. W., MOORE S. and STEIN W. H. (1956) Peptides obtained by tryptic hydrolysis of performic acid oxidiced ribonuclease. J. biol. Chem. 219, 623-642.
JOHNS E. W. (1967) The electrophoresis of histories in polyacrylamide gel and their quantitative determination. Biochem. J. 104, 78-82.
LAEMMU U. K. (1970) Cleavage of.structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685.
MUNICIO A. M., ODRIOZOLA J. M. and Pl~EIrO A. (1970) Biochemistry of the development of the insect Ceratitis capitata. C,mp. Biochem. Physiol. 37, 387-395.
NOLL M. and KORNBERG R. D. (1977)Action of micrococcal nuclease on chromatin and the location of histone HI. J. molec, Biol. 109, 393-404.
OLINS A, L. and Ouys D. E. (1974) Spheroid chromatin units (v bodies). Science 183, 330-332.
PANYtM S. and CHALKLEY R. (1969) High resolution acrylamide gel electrophoresis ofhistones. Archs. Biochem. Biaphl~ 130, 337-346.
PEREZ-ARANDA A. (1975) Estructura primaria de citocromo c de Ceratitis capitata. Ph.D. Thesis. Faculty of Chemistry, University Complutense, Madrid.
RODRIGUEZ-ALFAGEME C., ZWEIDLER A., M AHOWALD A. and COHEN L. H. (1974) Histones of Drosophila embrios. Electrophoretic isolation and structural studies. J. biol. Chem. 249, 3729-3736.
SUAU P., KNEALE G. G., BRADDOCK G. W., BALDWIN" J. P. • and BRADBURY E. M. (1977) A low resolution model for
the chromatin core particle by neutron scattering. Nucl. Acids Res. 4, 3769-3786.
Tvrou D., SAUTIERE P. and BISERTE G. (1976) Covalent structure of the sea urchin bistone H4. FEBS Letts 65, 225-228.





![Targeting the autosomal Ceratitis capitata transformer gene ......tory genes widely conserved in the Tephritidae family (Fig. 1)[6–10]. This taxon includes many other invasive agricultural](https://static.fdocuments.us/doc/165x107/60cedb3f8fcbf6439d79a6ee/targeting-the-autosomal-ceratitis-capitata-transformer-gene-tory-genes-widely.jpg)












