
HiLiDe ...jpmorth.dk/wp-content/uploads/2012/01/Gourdon2011CrystGRowDes.… · Published: April 08,...
9
Published: April 08, 2011 r2011 American Chemical Society 2098 dx.doi.org/10.1021/cg101360d | Cryst. Growth Des. 2011, 11, 2098–2106 ARTICLE pubs.acs.org/crystal HiLiDe—Systematic Approach to Membrane Protein Crystallization in Lipid and Detergent Published as part of the Crystal Growth & Design virtual special issue on the 13th International Conference on the Crystallization of Biological Macromolecules (ICCBM13). Pontus Gourdon, †,‡,# Jacob Lauwring Andersen, †,‡ Kim Langmach Hein, †,‡ Maike Bublitz, †,‡ Bjørn Panyella Pedersen, †, ‡ Xiang-Yu Liu, †, ‡, z Laure Yatime, †, ‡ Maria Nyblom, †, ‡ Thorbjørn Terndrup Nielsen, †,|| Claus Olesen, †,^ Jesper Vuust Møller, †,^ Poul Nissen,* ,†,‡ and Jens Preben Morth* ,†,‡,§ † Centre for Membrane Pumps in Cells and Disease—PUMPKIN, Danish National Research Foundation, University of Aarhus, DK-8000 Aarhus C, Denmark ‡ Department of Molecular Biology, Gustav Wieds Vej 10C ) Department of Biotechnology, Chemistry and Environmental Engineering, Aalborg University, Sohngaardsholmsvej 57, 9000 Aalborg, Denmark ^ Institute of Physiology and Biophysics, Ole Worms All e 1185 ’ INTRODUCTION With the aim of obtaining structural information on biomem- branes, high quality membrane protein crystals are required, and methods to obtain these are in constant development. The common and typically preferred method of choice is to maintain a low concentration of detergent above the critical micelle concentration (cmc), to shield the membrane spanning regions during the purification and the crystallization process (reviewed in ref 1). This approach frequently results in “Type II” crystals that are built up through interactions between the hydrophilic surfaces of the molecules (Figure 1a). 2,3 In comparison, “Type I” crystals 2,3 are defined by continuous bilayers of the membrane spanning segments and lipids. These crystals are often described as stacked 2-D crystals. To form actual 2-D crystals, lipid is generally supplemented and the detergent is extracted from the solution to reconstitute the membrane protein into a 2-D crystalline array. 4 In contrast, the 3-D “Type I” crystals are generated in the presence of detergent. Membrane proteins prepared and crystallized directly from a detergent solubilized, native membrane (retaining large amounts of solubilized lipids) typically form such Type I crystals (Table 1). Moreover, the type I crystal lattice is also induced by the cubic-, 5 sponge-, 6 and “bicelle”-phase methods (here denoted lipid-phase methods) 7 (Figure 1c). These latter three techniques make use of specific lipid properties to induce bilayer arrangements, and they have been applied successfully to a subset of membrane proteins. 8 However, they require specific lipid compositions and phase control and are often associated with technical hurdles, such as high viscosity and temperature dependence. In addition, lipid- phase methods are not freely compatible with the vapor-diffusion technique that is dominating the protein crystallization field. Here we present a straightforward and systematic approach to screen for initial crystal conditions and to proceed with rational crystal optimization to obtain well-diffracting “Type I” mem- brane protein crystals. The method centers on a relipidation step with relatively high concentrations of lipid(s) and detergent(s) prior to the crystallization process, and we have therefore abbreviated the approach HILIDE for the high lipid and high detergent concentrations used (Figure 1b). Conditions that pertain to membrane protein samples crystallized directly from solubilized native tissue membranes (Table 1) and the lipid-phase techniques have served as our source of inspiration for the rational approach. Received: October 14, 2010 Revised: April 5, 2011 ABSTRACT: To obtain the initial crystallization hit and the subsequent optimization remain the major bottlenecks in membrane protein structure determination. Here we describe an approach to achieve diffraction quality membrane protein crystals in high concentrations of lipid and detergent (HILIDE). Initial hits are achieved through a controlled variation of the membrane protein/lipid/detergent ratios, and optimized crystals are established by systematic screening with secondary detergents at concentra- tions that are adapted for the method. The approach is generally applicable for native and heterologously expressed membrane proteins, and it is compatible with any crystallization method, such as vapor diffusion and batch crystallization under oil. The method provides a user defined lipidic environment and is in itself not limited to lipids with particular properties, such as the lipid-phase crystallization methods.
-
Upload
nguyenhanh -
Category
Documents
-
view
213 -
download
0
Transcript of HiLiDe ...jpmorth.dk/wp-content/uploads/2012/01/Gourdon2011CrystGRowDes.… · Published: April 08,...

Published: April 08, 2011
r 2011 American Chemical Society 2098 dx.doi.org/10.1021/cg101360d | Cryst. Growth Des. 2011, 11, 2098–2106
ARTICLE
pubs.acs.org/crystal
HiLiDe—Systematic Approach to Membrane Protein Crystallization inLipid and DetergentPublished as part of the Crystal Growth & Design virtual special issue on the 13th International Conference onthe Crystallization of Biological Macromolecules (ICCBM13).
Pontus Gourdon,†,‡,# Jacob Lauwring Andersen,†,‡ Kim Langmach Hein,†,‡ Maike Bublitz,†,‡
Bjørn Panyella Pedersen,†,‡ Xiang-Yu Liu,†,‡,z Laure Yatime,†,‡Maria Nyblom,†,‡ Thorbjørn Terndrup Nielsen,†,||
Claus Olesen,†,^ Jesper Vuust Møller,†,^ Poul Nissen,*,†,‡ and Jens Preben Morth*,†,‡,§
†Centre for Membrane Pumps in Cells and Disease—PUMPKIN, Danish National Research Foundation,University of Aarhus, DK-8000 Aarhus C, Denmark‡Department of Molecular Biology, Gustav Wieds Vej 10C
)Department of Biotechnology, Chemistry and Environmental Engineering, Aalborg University, Sohngaardsholmsvej 57,9000 Aalborg, Denmark^Institute of Physiology and Biophysics, Ole Worms All�e 1185
’ INTRODUCTION
With the aim of obtaining structural information on biomem-branes, high quality membrane protein crystals are required, andmethods to obtain these are in constant development. Thecommon and typically preferred method of choice is to maintaina low concentration of detergent above the critical micelleconcentration (cmc), to shield the membrane spanning regionsduring the purification and the crystallization process (reviewedin ref 1). This approach frequently results in “Type II” crystalsthat are built up through interactions between the hydrophilicsurfaces of the molecules (Figure 1a).2,3 In comparison, “Type I”crystals2,3 are defined by continuous bilayers of the membranespanning segments and lipids. These crystals are often describedas stacked 2-D crystals. To form actual 2-D crystals, lipid isgenerally supplemented and the detergent is extracted from thesolution to reconstitute the membrane protein into a 2-Dcrystalline array.4 In contrast, the 3-D “Type I” crystals aregenerated in the presence of detergent. Membrane proteinsprepared and crystallized directly from a detergent solubilized,native membrane (retaining large amounts of solubilized lipids)typically form such Type I crystals (Table 1). Moreover, the typeI crystal lattice is also induced by the cubic-,5 sponge-,6 and“bicelle”-phase methods (here denoted lipid-phase methods)7
(Figure 1c). These latter three techniques make use of specificlipid properties to induce bilayer arrangements, and they havebeen applied successfully to a subset of membrane proteins.8
However, they require specific lipid compositions and phasecontrol and are often associated with technical hurdles, such ashigh viscosity and temperature dependence. In addition, lipid-phase methods are not freely compatible with the vapor-diffusiontechnique that is dominating the protein crystallization field.
Here we present a straightforward and systematic approach toscreen for initial crystal conditions and to proceed with rationalcrystal optimization to obtain well-diffracting “Type I” mem-brane protein crystals. The method centers on a relipidation stepwith relatively high concentrations of lipid(s) and detergent(s)prior to the crystallization process, and we have thereforeabbreviated the approach HILIDE for the high lipid and highdetergent concentrations used (Figure 1b). Conditions that pertainto membrane protein samples crystallized directly from solubilizednative tissue membranes (Table 1) and the lipid-phase techniqueshave served as our source of inspiration for the rational approach.
Received: October 14, 2010Revised: April 5, 2011
ABSTRACT: To obtain the initial crystallization hit and the subsequent optimizationremain the major bottlenecks in membrane protein structure determination. Here wedescribe an approach to achieve diffraction quality membrane protein crystals in highconcentrations of lipid and detergent (HILIDE). Initial hits are achieved through acontrolled variation of the membrane protein/lipid/detergent ratios, and optimizedcrystals are established by systematic screening with secondary detergents at concentra-tions that are adapted for the method. The approach is generally applicable for native andheterologously expressedmembrane proteins, and it is compatible with any crystallizationmethod, such as vapor diffusion and batch crystallization under oil. The method providesa user defined lipidic environment and is in itself not limited to lipids with particularproperties, such as the lipid-phase crystallization methods.

2099 dx.doi.org/10.1021/cg101360d |Cryst. Growth Des. 2011, 11, 2098–2106
Crystal Growth & Design ARTICLE
We have tested our approach with two new and heterologouslyoverproduced membrane protein targets from our laboratory andusing the generic phospholipid 1,2-dioleoyl-sn-glycero-3-phospha-tidylcholine (DOPC) solubilized by the detergent octaethyleneglycol monododecyl ether (C12E8). We show that crystal optimiza-tion in thepresenceof high amountsof lipid anddetergent canproceedalong similar rationales as with widely used detergent optimizationprocedures. Furthermore, immersion oil is identified as a favorable
sealing agent with our lipidated membrane protein samples. Finally,we conclude that the HILIDEmethod as a rational approach alsoencompasses several other examples in the literature.
’EXPERIMENTAL SECTION
Chemicals. Octaethylene glycol monododecyl ether (C12E8) waspurchased from Nikko Chemicals (Tokyo, Japan) and detergent screenHT from Hampton research (CA, USA). All other detergents werepurchased from Affymetrix (CA, USA). Castor oil, benzyl benzoate, andimmersion oil were purchased from Sigma Aldrich (MO, USA).Mass Spectrometry and NMR Analysis. MS/MS was per-
formed using a Bruker MicroTOFQ mass spectrometer. QuantitativeNMR analyses were conducted in CDCl3 using a BRUKER DRX600spectrometer operating at a field strength of 14.1 T with a 5 mm triple-axis gradient TXI(H/C/N) probe.Membrane Protein Preparation. The full length genes of the
Legionella pneumophila lpg1024 and Listeria monocytogenes lmca1 (genelocus lmo0841) were cloned into the pET-22b(þ) vector (Novagen)and expressed and purified as previously described,12 except thatLpg1024 was engineered (and purified) without an N-terminal tobaccoetch virus protease cleavage (TEV) site and yielded 1 mg per 1 g of wetweight cells. Following metal affinity chromatography on Ni2þ-sephar-ose beads, the eluted proteins were analyzed by SDS-PAGE, andLpg1024 containing fractions were pooled and concentrated to20�25 mg/mL (Vivaspin 50 kDa cut off). Aliquots equivalent to5 mg of concentrated protein were then applied to a Superose 6 size-exclusion column equilibrated in Buffer A (20 mM MOPS-KOH, pH6.8, 80 mMKCl, 20% glycerol, 5 mM BME, 1 mMMgCl2, and 0.28 mMC12E8). The Lpg1024 containing fractions were pooled and concen-trated to 20�25 mg/mL, flash frozen in aliquots of 200 μL in liquidnitrogen, and stored at�80 �C. For final samples, aliquots were dilutedto 10 mg/mL in Buffer A. Naþ,Kþ-ATPase was prepared as from theouter medulla in pig kidney as described previously.13
Re-lipidation. Samples for nine different membrane protein/lipid/detergent concentration ratios were generated for the initial crystal-lization screening. This was achieved by adding 100 μL of concentratedmembrane protein sample (∼10 mg/mL protein, Lpg1024 or LMCA1)to nine glass tubes (9 mm inner diameter, 7 cm high, round-bottom,from AG-Frisinette (Aarhus, Denmark)) pretreated with a thin film ofthe lipid 1,2-dioleoyl-sn-glycero-3-phosphatidylcholine (DOPC). Thefilm was prepared by dispensing DOPC dissolved at 40 mg/mLconcentration in CHCl3, corresponding to 0 (control), 0.15, and 0.3mg ofDOPC into three glass tubes for each lipid concentration, followedby evaporation of the CHCl3 using nitrogen gas (N2) at roomtemperature (preventing the lipid from oxidation). Subsequently, thelipidated protein samples were supplemented with either 0, 0.75, or1.5 mg of (extra) C12E8 from an aqueous 100 mg/mL stock solution, foreach lipid concentration, yielding a total of nine samples (see Table 2).The tubes were stirred at 50 rpm using microstirring bars (5� 2 mm2)for 10�18 h at 4 �C. The insoluble material was subsequently removedby ultracentrifugation at 190.000g for 10 min (using separate table-topcentrifugation tubes), and the transparent supernatants were used instandard hanging-drop crystallization experiments at either 4 or 20 �C.Identification of Crystallization Hit Conditions. The relipi-
dated supernatants were supplemented with 2 mM EGTA, 1 mM AlCl3,and 10 mM NaF (final concentrations) to stabilize the proteins (bothLpg1024 and LMCA1) in the so-called E2-AlF3 conformation(characteristic for P-type ATPases) and then diluted to 5 mg/mL andused for vapor-diffusion hanging drop experiments. Briefly, 1 μL ofprotein solution was mixed with 1 μL of reservoir solution using a pipetand let equilibrate over 450 μL of reservoir solution (siliconized cover-slips from Hampton Research were used throughout). An in-housescreen, with 48 conditions based on PEG2000 MME or PEG6000 and
Figure 1. Membrane protein crystallization with high-lipid, high-detergent concentrations, as compared to traditional techniques.Starting from purified and detergent-solubilized protein, different crys-tallization approaches are devised: (a) traditional crystallization ofdelipidated protein in detergents; (b) crystallization of relipidatedprotein with the HILIDE approach; or (c) lipid cubic phase crystal-lization. In part a, the detergent levels are typicallyminimized to generateinitial Type II crystals which may be optimized using detergent or lipidsscreens. In contrast, large amounts of detergent and lipid (b) or specificbicontinuous lipid phases (c) are supplemented prior to crystallization,yielding Type I crystals. The Type I crystals are improved using similarapproaches as for Type II crystals, albeit with higher concentrations ofdetergent.

2100 dx.doi.org/10.1021/cg101360d |Cryst. Growth Des. 2011, 11, 2098–2106
Crystal Growth & Design ARTICLE
various salts, was employed as reservoir solutions (described in ref 9),and the crystallization experiments were placed at 4 and 18 �C,respectively. All trays (standard 24-well trays from Hampton Research)were sealed with immersion oil (Merck cat. #104699). The experimentswere evaluated by visual inspection under the microscope, and crystalswere tested for diffraction at a synchrotron facility to assess the crystalquality as a function of the lipid-detergent ratio used.Identification of Secondary Detergents. Detergents that
improved the diffraction of crystals were identified using several differentreservoir conditions that provided initial hits (for both Lpg1024 andLMCA1). The 96 detergents screen fromHampton Research, employedat several different concentrations, was used for this purpose. For1 � cmc (in the initial drop), 1 μL of detergent stock solution wasmixed with 4 μL of 1.25 � reservoir solution, and then 1 μL of proteinwas finally mixed with 1 μL of detergent/reservoir combination (nodetergent supplements to the reservoir).Detergent Optimization. The procedure described below was
documented for the pig Naþ,Kþ-ATPase and elaborates on previouslydescribed results.14 Similar procedures were followed for Lpg1024 andLMCA1 using other detergents.12,28,29
Four 25 μL Naþ,Kþ-ATPase samples in Eppendorf tubes (at a ∼10mg/mL protein concentration) were supplemented with 1 μL of C12E8solution to each tube (1, 2, 3, or 4% (w/v) C12E8 in water to tubes 1through 4, respectively). The precipitating solution (14% PEG, 200 mMCholine Chloride, 4% methylpentanediol, 4% glycerol, 4 mMdithiotreitol) was prepared separately and freshly. Finally, six differentsolutions of a secondary detergent (1.0, 2.0, 3.0, 4.0, 5.0, and 6.0% (w/v)β-DDM in water) were prepared. From each of the four protein tubes,six samples were made. The first sample contained 4 μL of the proteinsolution, 4 μL of precipitating solution, and 0.8 μL of 1.0% β-DDM, thesame for the second, except 0.8 μL of 2.0% β-DDM and so forth, andrepeated again for protein samples 2, 3, and 4. Any precipitate thatformed in each of the 8.8μL final samples was removed by centrifugationat 13K rpm for 1 min at room temperature, typically leaving a visiblewhite pellet. The supernatants of each final sample were applied as fouridentical 2 μL drops on coverslips to complete one column of a 24-wellhanging drop crystallization plate. The reservoir solutions would beequivalent to the precipitating solution only with increasing PEGconcentration for each row (14, 15, 16, and 17%) to fine-tune vapordiffusion strength within each column of the trays.
Table 1. Examples of Membrane Proteins Crystallized in High-Lipid and High-Detergent
protein lipid detergent
Type I Crystals from Chromatography Purified Proteins Using the Approach Presented in This Work
10 mg/mL Lpg1024, this work 7.5 mg/mL DOPC 1.5 mg/mL C12E810 mg/mL LMCA1, this work 1 mg/mL DOPC 3.5 mg/mL C12E812 mg/mL SERCA1a9 8 mg/mL DOPC 24 mg/mL C12E8
Examples of Type I Crystals Yielded from Chromatography Purified Proteins Using an Approach Similar to the One Presented in This Work
10 mg/mL rKv1.2-β210 0.1b mg/mL (3:1:1 POPC/POPE/POPG) 3a mg/mL DM
20 mg/mL E.coli Complex I11 2 mg/mL (3:1 DMPC/E. coli PL) 5 mg/mL β-HG
20 mg/mL T. thermophilus Complex I11 2 mg/mL (E. coli PL) 2 mg/mL DDM
Examples of Type I Crystals Yielded Directly from Proteins Solubilized from Native Membranes (As Mimicked by the Approach Presented in This Work)∼9 mg/mL pea LHC-II23 ND, endogenous 10a mg/mL β-NG
∼8 mg/mL bovine rhodopsin24 ND, endogenous ∼18 mg/mL β-NG
20 mg/mL bovine Cyt bc125 4 mg/mL, endogenous 1 mg/mL DMG
10�16 mg/mL rabbit SERCA1a26 ND, endogenous ∼16 mg/mL C12E811 mg/mL pig Naþ,Kþ-ATPase14 ND, endogenous 9 mg/mL C12E82.5 mg/mL shark Naþ,Kþ-ATPase27 2.1 mg/mL, endogenous NA mg/mL C12E8
Examples of Type I Crystals from Purified Proteins Using the Bicelle Technique10 mg/mL hβ2-adrenergic receptor
18 20 mg/mL DMPC 60 mg/mLCHAPSO
12 mg/mL mVDAC119 50 mg/mL DMPC 20 mg/mL CHAPSO
Examples of Non-Type I Crystals from Purified Proteins8 mg/mL spinach LHC-II21 2 mg/mL DGDG 8 mg/mL β-NG
20 mg/mL M. laminosus Cyt b6f20 1 mg/mL DOPC 0.5 mg/mL UDMaConcentration prior to concentration. bUnclear whether it is prior to or after concentration.
Table 2. Membrane Protein/Lipid/Detergent Ratios Generated in the Re-lipidation
0 mg lipid DOPC 0.15 mg lipid DOPC 0.3 mg DOPC
0 mg C12E8 1:0:22a þ 0b (0.13:0:2.8 þ 0)c 1:17:22a þ 0b (0.13:2.1:2.8 þ 0) 1:33:22a þ 0b (0.13:4.3:2.8 þ 0)
0.75 mg C12E8 1:0:22a þ 102b (0.12:0:2.8 þ 12) 1:17:22a þ 102b (0.12:2.0:2.6 þ 12) 1:33:22a þ 102b (0.12:4.0:2.6 þ 12)
1.5 mg C12E8 1:0:22a þ 233b (0.11:0:2.4 þ 26) 1:17:22a þ 233b (0.11:1.9:2.4 þ 26) 1:33:22a þ 233b (0.11:3.7:2.4 þ 26)aRatios ofmembrane protein/DOPC/C12E8 (concentrated). bC12E8 added during the nine relipidations displayed in Figure 2.
cConcentrations (mM)of the components are indicted (in parentheses). One milligram of protein was used in all relipidations. We have assumed that the C12E8 has beenconcentrated ten times during protein concentration (prior to the relipidation) and that the DOPC supplementation has a negligible effect on the finalvolume.

2101 dx.doi.org/10.1021/cg101360d |Cryst. Growth Des. 2011, 11, 2098–2106
Crystal Growth & Design ARTICLE
Lipid Supplementation Technique. A comparison of lipidsupplementation procedures was made to see if DOPC could besupplied to LMCA1 as a cocrystallization experiment rather than by amore elaborate relipidation procedure prior to crystallization. Identicalcrystallization conditions were employed. Prior to the cocrystallizationexperiments, DOPC was solubilized in low concentrations of C12E8 to
obtain saturated solutions with the same membrane protein/lipid/detergent ratios as in the relipidation experiment. The saturated DOPCsolutions (without protein) were obtained by solubilization of 4 mg ofDOPC-film (DOPC dissolved in CHCl3 (40 mg/mL)) in 1 mL ofC12E8-solutions with different concentrations (equivalent to valuesgiven in Table 2) with ultrasonic waves in a water bath (Branson 550)
Figure 2. Flow charts of the high-lipid high-detergent crystallization of a heterologously overproduced membrane protein and of the systematicdetergent optimization approach. (a) The lipid/detergent optimization: (1) The generation of a lipid film using DOPC dissolved in CHCl3. Threedifferent amounts of DOPC are added, resulting in three lipid/membrane protein ratios. (2) Addition of the membrane protein and detergent 1. Threedifferent amounts of detergent 1 are added, resulting in three detergent/membrane protein ratios. Combined with the three lipid/protein ratios, a total ofnine samples with unique lipid/detergent/membrane protein ratios are obtained. (3) Incubation. (4) Removal of aggregates and excess lipid bycentrifugation. (5) A sparse matrix screen was set up for each of the nine unique lipid/detergent to membrane protein ratios. Representative pictures ofthe same condition (green dots) are shown below. (b) Detergent 1 (C12E8) and detergent 2 (β-DDM) optimization for the pig Naþ,Kþ-ATPase.14 Theprecipitating solution added to the drops is identical for all, only with increasing β-DDM concentration along the six columns and increasing PEGconcentration down the rows. Each setup required a detergent optimization window of 0.1�0.4% β-DDM for every new protein batch.14

2102 dx.doi.org/10.1021/cg101360d |Cryst. Growth Des. 2011, 11, 2098–2106
Crystal Growth & Design ARTICLE
for 60 min at 18 �C, and subsequent removal of insoluble materialthrough ultracentrifugation at 190.000g for 10 min (Beckman rotorTLA-100.3, in Microfuge Tube, Polyallomer part # 357448). Onemicroliter of DOPC saturated solution was then mixed with 4 μL1.25 � reservoir solution, and eventually 1 μL of DOPC saturated/reservoir combination was supplemented to 1 μL drops of the proteinsample. Control experiments were performed using the relipidationapproach described above. Crystallization was performed at 18 �C.Immersion Oil for Crystallization. An analysis of the effect of
immersion oil on the HILIDE crystallization methods was conductedusing LMCA1. All experiments were performed using relipidatedLMCA1 samples (see earlier), and identical crystallization conditionswere employed for the different experiments. The wells were sealed witheither Dow Corning Vacuum Grease from Hampton research orimmersion oil from Merck. For the immersion oil supplementedconditions, 0.1, 1, or 10 μL of immersion oil were mixed with 500 μLof solution identical to the reservoir/precipitant solution. Immediatelyprior to the crystallization experiment, the mixture was thoroughlymixed and 1μL extracted and added to a 1 μL protein drop (as describedearlier).
’RESULTS
The two proteins selected for our HILIDE relipidationapproach belong to two different families of the P-type ATPasesuperfamily: The P1-type ATPase Lpg1024
29 from L. pneumophila,and the P2-type ATPase LMCA112,28 from L. monocytogenes.The two targets differ significantly in primary structure, with onlya few conserved sequence motifs distributed over three cyto-plasmic domains and six transmembrane helices in a commontopology. Lpg1024 contains a metal binding domain and twoadditional transmembrane helices at the N-terminus, whileLpg1024 displays four additional transmembrane helices at theC-terminus. The proteins were overproduced in E. coli andsubjected to purification by liquid chromatography in thedetergent octaethylene glycol monododecylether (C12E8).
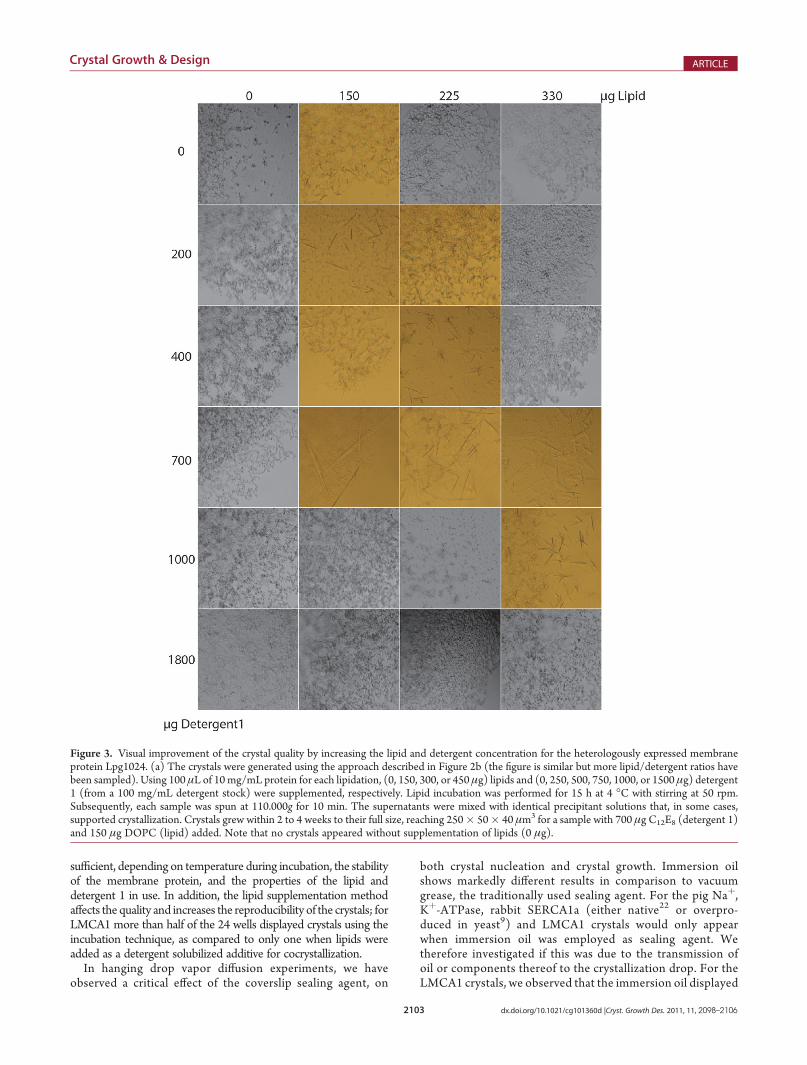
Prior to crystallization, the proteins were relipidated in a two-step procedure. First, a thin film of lipid was generated bydispensing the lipid dissolved in chloroform into glass tubesfollowed by removal of the chloroform by evaporation in a N2-atmosphere tominimize lipid oxidation (Figure 2a, step 1). In thesecond step, the concentrated membrane protein and detergent1 (the detergent used during purification) are added (Figure 2a,step 2) and incubated with stirring for several hours. Due to thehigh lipid content, detergent 1 is, in effect, a limiting factor andadditional detergent needs to be added to allow further lipidsolubilization. By using three different concentrations of the lipidand detergent 1, we end up with nine unique membrane protein/lipid/detergent ratios. These samples were used in standardhanging drop crystallization screens at 4 and 18 �C. Visualinspection of the crystallization drops reveals a clear variationin the crystal morphology and size, depending on the protein/detergent/lipid ratio. Representative pictures from the relipida-tion experiments, crystallized at 20 �C, were taken from eachplate (Figure 2a, step 4, and Figure 3). An apparent optimum ofthe protein/lipid/detergent ratio for each individual target wasidentified and the increased size of the crystals obtained. Im-portantly, and despite an extensive screening for both Lpg1024and LMCA1, we were not able to get diffraction quality crystalswithout our thorough relipidation procedure (Figure 3). Thelipid employed in this work was 1,2-dioleoyl-sn-glycero-3-phos-phatidylcholine (DOPC). We have also successfully growncrystals of these two proteins relipidated in other lipids, including
E. coli lipid extract (Avanti Polar Lipids) as well as soy bean lipids(Sigma), using the identical approach and similar amounts oflipids (data not shown). However, no visual improvements ascompared to DOPCwere observed, and we have used DOPC forour further analysis.
Screening for detergents that modify the protein/lipid/deter-gent micelles has often proven critical in the optimization ofmembrane protein crystals.15 It was, however, unclear if thisprinciple of optimization was compatible with our relipidationprocedure due to the significantly higher levels of lipid anddetergent present during crystallization. The flow chart inFigure 2b outlines the strategy for how we practically approachedthe addition of the second detergent. Importantly, a large numberof detergents needs to be tested and both the first and the second(and possibly a third) detergents must be reassessed in concen-tration ranges during optimization. The range of concentrationsin which we saw the strongest effect we named “the detergentoptimization window”. As an example, we show here the improve-ment of the Type I crystals resulting from addition of β-DDM topig Naþ,Kþ-ATPase solubilized in C12E8 (Figure 2b)14—a similar effect was seen for Lpg1024 and LMCA1.
In the first step, the membrane protein was diluted in a buffercontaining the detergent 1 used in the solubilization or simply abuffer without detergent to reduce the protein concentration(Figure 2b, step 1). In the second step the second detergent(detergent 2—an additive detergent) is added (Figure 2b, step 2).For the Naþ,Kþ-ATPase, the best-diffracting crystals wereobtained with a detergent 2 optimization window of 0.1�0.4%(w/v) β-DDM (Figure 2b, step 4). Probing by a more narrowrange of β-DDM concentrations was unsuccessful due to batch-to-batch variations on the optimal concentration for detergent 2.These levels of detergent 2 (in the range 10�40 times the cmcfor β-DDM) are considerably higher than typically used forcrystal optimization of “Type II” crystals (often reported as 1�3times the cmc). As a consequence, we have adapted the detergentscreen from Hampton Research and employed the detergents atmuch higher concentrations than recommended by the supplier.Using this approach, we successfully identified secondary deter-gents that improved diffraction properties for Lpg1024 with datasets collected at 3.2 Å resolution29 and LMCA1 with data setscollected at 4.3 Å resolution.28 For most of the HILIDE proteincrystals we have generated, including Lpg1024, LMCA1, andrabbit SERCA1a,9 the detergent 2 optima were in the range3�15 times the cmc with similar trends as the one shown inFigure 2b.
The crystal structures of both Lpg1024 and LMCA1 show acrystal lattice arrangement consistent with “Type I” crystalpacking, as is also seen in the case of relipidated rabbit SERCA1aoverproduced in yeast (Figure 4).9 Thus, it appears that theresulting type of packing in the crystals generated within thesuccessful protein/lipid/detergent window is equivalent to thoseobtained by the cubic-,5 sponge-,6 and “bicelle”7 techniques.
The long lipid incubation stage of several hours (also requiredfor the bicelle approach) made us assess the lipid supplementa-tion technique further. However, cocrystallization with DOPClipid dissolved in C12E8 (with the aim to yield equivalent levels asthe relipidation procedure) failed to reproduce the good-qualitycrystals (Figure 5, see the methods section “Lipid Supplementa-tion Technique”). Thus, we recommend the thorough relipida-tion procedure described here as the preferred protocol forcrystallization in high lipid and detergent concentrations. It ishowever possible that a briefer relipidation period would be
2103 dx.doi.org/10.1021/cg101360d |Cryst. Growth Des. 2011, 11, 2098–2106
Crystal Growth & Design ARTICLE
sufficient, depending on temperature during incubation, the stabilityof the membrane protein, and the properties of the lipid anddetergent 1 in use. In addition, the lipid supplementation methodaffects the quality and increases the reproducibility of the crystals; forLMCA1 more than half of the 24 wells displayed crystals using theincubation technique, as compared to only one when lipids wereadded as a detergent solubilized additive for cocrystallization.
In hanging drop vapor diffusion experiments, we haveobserved a critical effect of the coverslip sealing agent, on
both crystal nucleation and crystal growth. Immersion oilshows markedly different results in comparison to vacuumgrease, the traditionally used sealing agent. For the pig Naþ,Kþ-ATPase, rabbit SERCA1a (either native22 or overpro-duced in yeast9) and LMCA1 crystals would only appearwhen immersion oil was employed as sealing agent. Wetherefore investigated if this was due to the transmission ofoil or components thereof to the crystallization drop. For theLMCA1 crystals, we observed that the immersion oil displayed
Figure 3. Visual improvement of the crystal quality by increasing the lipid and detergent concentration for the heterologously expressed membraneprotein Lpg1024. (a) The crystals were generated using the approach described in Figure 2b (the figure is similar but more lipid/detergent ratios havebeen sampled). Using 100 μL of 10 mg/mL protein for each lipidation, (0, 150, 300, or 450 μg) lipids and (0, 250, 500, 750, 1000, or 1500 μg) detergent1 (from a 100 mg/mL detergent stock) were supplemented, respectively. Lipid incubation was performed for 15 h at 4 �C with stirring at 50 rpm.Subsequently, each sample was spun at 110.000g for 10 min. The supernatants were mixed with identical precipitant solutions that, in some cases,supported crystallization. Crystals grew within 2 to 4 weeks to their full size, reaching 250� 50� 40 μm3 for a sample with 700 μg C12E8 (detergent 1)and 150 μg DOPC (lipid) added. Note that no crystals appeared without supplementation of lipids (0 μg).

2104 dx.doi.org/10.1021/cg101360d |Cryst. Growth Des. 2011, 11, 2098–2106
Crystal Growth & Design ARTICLE
a direct effect on crystal nucleation when added directly to thedrop as an additive (Figure 5). The reported components ofthe immersion oil are castor oil and benzyl benzoate at anapproximately 1:3 w/w ratio (Merck cat. #104699). Neithermass spectrometry nor nuclear magnetic resonance (NMR)spectroscopy identified any additional components (data notshown). Both castor oil and benzyl benzoate applied alonefailed to reproduce the effect of immersion oil, while acombination of the two restored the notable effects onnucleation and crystal growth.
’DISCUSSION
Several reports in the literature use high lipid and detergentconcentrations, as we use here, including the high-resolutioncrystal structure of the rat potassium channel rKv1.2-β210 as wellas the multicomponent Complex I from Thermus thermophilusand Escherichia coli with different subunit compositions11 (seealso Table 1). Furthermore, as previously indicated, very similarconditions were present when targets from native tissues werecrystallized (Table 1). Thus, these crystals have been generated
Figure 4. Crystal packing of the LMCA1 and Lpg1024: (a) LMCA1 is represented as ribbons, with transmembrane regions colored green, thecytoplasmic helices in blue, and beta-sheets in red; (b) view rotated 90�. The structure was solved by a molecular replacement structure using diffractiondata extending to 4.5 Å resolution and the SERCA1a structure PDB ID 3FGO.22,28 (c) Lpg1024 is represented as ribbons, with transmembrane regionscolored red and the cytoplasmic domains in green. A single monomer is highlighted by a different shade of color. (d) View rotated 90�.29 The crystalpacking of both LMCA1 and Lpg1024 shows a bilayered “Type I” crystal packing as stimulated by the relipidation approach.
Figure 5. Visual crystal differences between two different lipid supplementation techniques and the use of either immersion oil or vacuum grease.(a) Experience from our laboratory indicates that supplementing lipids using the approach described in this paper (left) is superior to adding similaramounts of detergent-solubilized lipids as an additive (right). Both samples are from the same protein preparation and crystallized with identicalprecipitation agents. (b) Crystals of several membrane proteins, which we have generated in high-lipid and high-detergent concentration, includingrabbit SERCA,22 pig Naþ,Kþ-ATPase,14 and relipidated LMCA1 described in this work, are influenced by the way the crystallization trays are sealed. Forsome, only oil, and not grease, leads to crystal formation. Recent results from our laboratory with LMCA1 indicate that the oil displays a direct effect oncrystal nucleation when added as an additive. The figure demonstrates four situations, with identical membrane protein and precipitant solutions andwith oil or grease used as indicated. We have tested 0.1�10 μL oil per 500 μL reservoir solution (and then transferred 1 μL for 1þ 1 μL crystallizationdrops) with similar results.

2105 dx.doi.org/10.1021/cg101360d |Cryst. Growth Des. 2011, 11, 2098–2106
Crystal Growth & Design ARTICLE
for at least six different types of membrane proteins usingconditions very similar to the ones generated following therelipidation procedure presented here and using a wide set ofcombinations of detergents and lipids. This number is not evenincluding several membrane proteins crystallized by the “bicelle”approach8 at comparable protein/lipid/detergent concentra-tions and ratios (Table 1, exemplified by the human β2-adrener-gic G protein-coupled receptor18 and murine voltage-dependentanion channel19). In addition, the usefulness of a thorough lipid/detergent treatment is supported by nonbilayered crystals thathave emerged from conditions that are similar to the onespresented here; the Cytochrome b6f complex fromMastigocladuslaminosus packed as a Type II crystal and20 light-harvestingcomplex II from spinach ordered in icosahedral liposomes(coined Type III crystals by the authors)21 (Table 1).
Considering the choice of lipid and detergent for a specificmembrane protein, no apparent trend is clear (Table 1). Weassume that the optimal types of lipids and detergents must beestablished for each individual membrane protein based on, e.g.,biochemical assays and screening, but an initial set of testcomponents may otherwise be selected from Table 1. In fact, ageneral usage of other lipid/detergent combinations thanDMPC/CHAPSO originally suggested for the bicelle methodhas already been proposed, but not validated.8
We propose that the simple and systematic relipidationmethodology described here is generally applicable and favors“Type I” membrane protein crystals. Such bilayer arrangementsin the crystal packing are attractive approximations for nativeconditions.
The HILIDE approach should also be considered as analternative to crystallization in pure detergent micelles. In fact,we have rarely observedmembrane proteins of the P-type ATPasefamily to crystallize without a prior relipidation procedure.
Although not tested on an exhaustive number of lipids, ourmethod should in principle work for all physiologically relevantlipids—since only detergent solubilized lipid is required in aprotein-containing supernatant, and vapor diffusion for crystal-lization is used in a visual screening-based approach to identifythe optimal ratios. This is an advantage over the cubic-,5
sponge-,6 and “bicelle”7 techniques, which are specialized toparticular lipid phases. The “optimal” lipid for crystallization islikely independent of both the origin of the recombinant geneand the expression host. If, however, a specific lipid was requiredfor stability or activity of a membrane protein, one would merelyinclude it in the relipidation procedure.
Furthermore, our approach yields nonviscous, nongel phasesolutions at temperatures used for crystallization (4�30 �C).Themethod is also straightforward to implement and compatiblewith ordinary liquid handling tools and robotics, which arebecoming increasingly important in the field. Thus, the describedrelipidation procedure represents a simple technique to incorpo-rate lipids in the sample and obtain samples with favorablecrystallization properties.
The type and the concentration of solubilizing and additivedetergents are of key importance in optimization of membraneprotein crystals. The additive detergents are expected to partitionin lipid-detergent bilayers and they affect crystal packing properties.The screening for additive detergents can follow well-establishedroutines and formulations butmust exploit higher concentrations,typically reaching 3�15 times the cmc or even higher. This is incontrast to 2-D crystallization, where the removal of detergent is adriving force for reconstitution of the lipid bilayers.4,16 In this
context it should be noted that other lipid-detergent micelleinterfering factors, such as salts, polymers, alcohols, and pH,also display strong influences on crystal packing and crystalquality.4,16,17 We therefore recommend that crystal optimizationswith lipid and detergent interfering factors are combined with abroad detergent optimization window, and vice versa.
The effect of immersion oil on nucleation and crystal growth iscompelling, and for some examples in our laboratory, it has beencritical. The effect is seemingly synergetic between castor oil andbenzyl benzoate, and it is likely that the immersion oil stabilizesthe lipid-detergent bilayers, e.g. by incorporation of triglyceridesof the castor oil. The sealing agent is not a typical parameter toinvestigate during crystallization, and the gas permeability of thesealing agent can also cause different results during the crystal-lization experiment. Our result does however show that the directcontact is important for the immersion oil. We did experiencedifficulty with using immersion oil as an actual additive and, thus,recommend to merely try both vacuum grease and immersion oilas sealing agent during crystal screening and optimization.
’CONCLUSIONS
We present a successful and generally applicable method torelipidate membrane proteins for crystallization experiments,dubbed the HILIDE method to indicate the use of high lipidand detergent concentrations. The technique aims at conditionsmimicking solubilized membranes that appears to “re-form” intorepetitive bilayers in the crystal, as stimulated by the crystal-lization conditions. In addition, we describe a systematic ap-proach to improve the crystal packing of these crystals byoptimization of the detergent components of the membraneprotein/lipid/detergent mixture. The technique is well suited forstandard vapor-diffusion experiments and compatible with or-dinary pipetting and robotics tools, and is therefore also ideal forthe generation of samples for automated screening platforms.Webelieve that our results clearly show the importance of detergent/lipid optimization at an early stage in the crystallization process ofmembrane proteins. Keeping the detergent/lipid optimizationon top of the priority list for a membrane protein project willincrease the crystallization probability and thus decrease the totalprotein consumption.
’AUTHOR INFORMATION
Corresponding Author*J.P.M.: phone, þ47 2284 0794; e-mail: j.p.morthncmm.uio.no.P.N.: phone, þ45 89425025; e-mail, [email protected].
Present Addresses§Centre for Molecular Medicine, Nordic EMBL Partnership,University of Oslo, P.O. Box 1137, Blindern, 0318 Oslo, Norway.#Department of Biochemistry and Biophysics, University of Cali-fornia at San Francisco, San Francisco, California 94158, USA.zState Key Laboratory of Protein and Plant Gene Research,College of Life Sciences, Peking University, Beijing, 100871,P.R. China.
’ACKNOWLEDGMENT
This study was supported by postdoctoral fellowships fromthe Swedish Research Council (P.G. and M.N., respectively)and the Carlsberg Foundation (B.P.P. and J.P.M.) and Lund-beckfondet (J.P.M), a Hallas-Møller stipend from the Novo

2106 dx.doi.org/10.1021/cg101360d |Cryst. Growth Des. 2011, 11, 2098–2106
Crystal Growth & Design ARTICLE
Nordisk Foundation (P.N.), and a State Scholarship from theChina Scholarship Council (X.-Y.L). Support was also obtainedby the EU-FP7 program EDICT. We thank Mickael Blaise andPaul A. Tucker for critical reading of the manuscript.
’ABBREVIATIONS
hβ2 adrenergic receptor, human β2-adrenergic G protein-coupled receptor; C12E8, octaethylene glycol monododecylether; CHAPSO, 3-[(3-cholamidopropyl)dimethylammonio]-2-hydroxy-1-propanesulfonate; Cyt b6f, Cytochrome b6fcomplex; Cyt bc1, Cytochrome bc1 complex; DDM, n-dodecyl-β-D-maltoside; DGDG, digalactosyl diacylglycerol; DM, n-decyl-β-D-maltopyranoside;DMG,decanoyl-N-methyl-glucamide;DMPC,dimyristoylphosphatidylcholine;DOPC, 1,2-dioleoyl-sn-glycero-3-phos-phatidylcholine;E. coliPL,E. colipolar lipids extract,MWassumed to be750 g/mol;β-HG, n-heptyl-β-glucoside;LHC-II, light harvestingcomplex II; ND, not determined; β-NG, n-nonyl-β-glucoside;POPC, 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine; POPE,1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine; POPG,1-palmitoyl-2-oleoyl-sn-glycero-3-[phospho-rac-(1-glycerol)];SERCA1a, sarcoplasmic reticulum Ca2þ-ATPase (Oryctolaguscuniculus (Rabbit)); UDM, β-D-undecyl-maltoside; mVDAC1,murine voltage-dependent anion channel.
’REFERENCES
(1) Prive, G. G. Methods 2007, 4, 388–397.(2) Ostermeier, C.; Michel, H. Curr. Opin. Struct. Biol. 1997,
5, 697–701.(3) Michel, H. Trends Biochem. Sci. 1983, 2, 56–59.(4) Hasler, L.; Heymann, J. B.; Engel, A.; Kistler, J.; Walz, T. J. Struct.
Biol. 1998, 2, 162–171.(5) Landau, E. M.; Rosenbusch, J. P. Proc. Natl. Acad. Sci. U. S. A.
1996, 25, 14532–14535.(6) Wohri, A. B.; Johansson, L. C.; Wadsten-Hindrichsen, P.;
Wahlgren, W. Y.; Fischer, G.; Horsefield, R.; Katona, G.; Nyblom, M.;Oberg, F.; Young, G.; Cogdell, R. J.; Fraser, N. J.; Engstrom, S.; Neutze,R. Structure 2008, 7, 1003–1009.(7) Faham, S.; Bowie, J. U. J. Mol. Biol. 2002, 1, 1–6.(8) Johansson, L. C.;Wohri, A. B.; Katona, G.; Engstrom, S.; Neutze,
R. Curr. Opin. Struct. Biol. 2009, 4, 372–378.(9) Jidenko, M.; Nielsen, R. C.; Sorensen, T. L.; Moller, J. V.;
le Maire, M.; Nissen, P.; Jaxel, C. Proc. Natl. Acad. Sci. U. S. A. 2005,33, 11687–11691.(10) Long, S. B.; Campbell, E. B.; Mackinnon, R. Science 2005,
5736, 897–903.(11) Efremov, R. G.; Baradaran, R.; Sazanov, L. A. Nature 2010,
7297, 441–445.(12) Faxen, K.; Andersen, J. L.; Gourdon, P.; Fedosova, N.; Morth,
J. P.; Nissen, P.; Moller, J. V. J. Biol. Chem. 2011, 2, 1609–1617.(13) Jorgensen, P. L. Biochim. Biophys. Acta 1974, 1, 36–52.(14) Morth, J. P.; Pedersen, B. P.; Toustrup-Jensen, M. S.; Sorensen,
T. L.; Petersen, J.; Andersen, J. P.; Vilsen, B.; Nissen, P. Nature 2007,7172, 1043–1049.(15) Seddon, A. M.; Curnow, P.; Booth, P. J. Biochim. Biophys. Acta
2004, 1�2, 105–117.(16) Dolder, M.; Engel, A.; Zulauf, M. FEBS Lett. 1996,
1�2, 203–208.(17) le Maire, M.; Champeil, P.; Moller, J. V. Biochim. Biophys. Acta
2000, 1�2, 86–111.(18) Cherezov, V.; Rosenbaum, D. M.; Hanson, M. A.; Rasmussen,
S. G.; Thian, F. S.; Kobilka, T. S.; Choi, H. J.; Kuhn, P.; Weis, W. I.;Kobilka, B. K.; Stevens, R. C. Science 2007, 5854, 1258–1265.
(19) Ujwal, R.; Cascio, D.; Colletier, J. P.; Faham, S.; Zhang, J.; Toro,L.; Ping, P.; Abramson, J. Proc. Natl. Acad. Sci. U. S. A. 2008,46, 17742–17747.
(20) Zhang, H.; Kurisu, G.; Smith, J. L.; Cramer, W. A. Proc. Natl.Acad. Sci. U. S. A. 2003, 9, 5160–5163.
(21) Liu, Z.; Yan, H.; Wang, K.; Kuang, T.; Zhang, J.; Gui, L.; An, X.;Chang, W. Nature 2004, 6980, 287–292.
(22) Laursen, M.; Bublitz, M.; Moncoq, K.; Olesen, C.; Moeller,J. V.; Young, H. S.; Nissen, P.; Morth, J. P. J. Biol. Chem. 2009,284, 13513–13518.
(23) Standfuss, J.; Terwisscha van Scheltinga, A. C.; Lamborghini,M.; Kuhlbrandt, W. EMBO J. 2005, 5, 919–928.
(24) Okada, T.; Le Trong, I.; Fox, B. A.; Behnke, C. A.; Stenkamp,R. E.; Palczewski, K. J. Struct. Biol. 2000, 1, 73–80.
(25) Xia, D.; Yu, C. A.; Kim,H.; Xia, J. Z.; Kachurin, A.M.; Zhang, L.;Yu, L.; Deisenhofer, J. Science 1997, 5322, 60–66.
(26) Sorensen, T. L.; Olesen, C.; Jensen, A. M.; Moller, J. V.; Nissen,P. J. Biotechnol. 2006, 4, 704–716.
(27) Shinoda, T.; Ogawa, H.; Cornelius, F.; Toyoshima, C. Nature2009, 7245, 446–450.
(28) Andersen, J. L.; Gourdon, P.; Moller, J. V.; Morth, J. P.; Nissen,P. Crystallization and preliminary structural analysis of the Listeriamonocytogenes Ca2þ-ATPase LMCA1. Acta Crystallogr., Sect. F 2011,accepted.
(29) Gourdon, P.; et al. Nature 2011, accepted.
















![Gorilla Wreck Shatter 10.04.2019[2098] - Venom Extracts€¦ · Title: Gorilla Wreck Shatter 10.04.2019[2098].pdf Author: logan Created Date: 10/9/2019 2:17:29 PM](https://static.fdocuments.us/doc/165x107/5f684be189daaa3ad52915a4/gorilla-wreck-shatter-100420192098-venom-extracts-title-gorilla-wreck-shatter.jpg)
