Heteroresistance to Meropenem in Carbapenem...
5
JOURNAL OF CLINICAL MICROBIOLOGY, Dec. 2009, p. 4055–4059 Vol. 47, No. 12 0095-1137/09/$12.00 doi:10.1128/JCM.00959-09 Copyright © 2009, American Society for Microbiology. All Rights Reserved. Heteroresistance to Meropenem in Carbapenem-Susceptible Acinetobacter baumannii Alexandros Ikonomidis, 1 * Evangelia Neou, 1 Vassiliki Gogou, 1 Georgia Vrioni, 2 Athanassios Tsakris, 2 and Spyros Pournaras 1 University of Thessaly, Mezourlo, Larissa, Greece, 1 and University of Athens, Athens 11527, Greece 2 Received 13 May 2009/Returned for modification 29 June 2009/Accepted 26 September 2009 The characteristics of carbapenem heteroresistance were studied in 14 apparently carbapenem-susceptible Acinetobacter baumannii isolates. The MICs for carbapenems were determined, and the isolates were genotyped by pulsed-field gel electrophoresis (PFGE) and sequence typing (ST). Population analysis, testing of the stability of the heteroresistant subpopulations, and time-killing assays were performed. The agar dilution MICs of both imipenem and meropenem for the native isolates ranged from 0.25 to 4 mg/liter. The isolates belonged to nine PFGE types and exhibited seven ST allelic profiles. Population analysis revealed subpopu- lations that grew in the presence of imipenem at concentrations of up to 8 mg/liter and meropenem at concentrations of up to 32 mg/liter. The meropenem-heteroresistant subpopulations of 11 isolates exhibited stable resistance with MICs that ranged from 16 to >32 mg/liter; their PFGE profiles were identical to those of the native isolates. Time-kill assays with meropenem revealed less pronounced killing for 10 isolates. These findings indicate that meropenem pressure can produce meropenem-heteroresistant subpopulations that might subsequently select for highly resistant strains. Acinetobacter baumannii is an opportunistic pathogen asso- ciated with severe hospital infections (1, 16), which often need the use of carbapenems as the treatment of last resort. How- ever, reduced susceptibility or resistance to carbapenems is increasingly being observed among A. baumannii clinical iso- lates (2, 5, 25). In a preliminary work, we reported on the growth of distinct colonies within the inhibition halo around carbapenem disks or Etest strips and assessed these colonies as having the pheno- typic manifestation of carbapenem heteroresistance (17). Like- wise, a recent study (12) has described colistin-heteroresistant subpopulations among apparently colistin-susceptible A. bau- mannii clinical isolates, implying an intrinsic potential of the species to overcome drug pressure. These observations suggest that the use of carbapenems or colistin to treat severe multi- drug-resistant A. baumannii infections may lead to the devel- opment of resistance. The present study aimed to evaluate in vitro the incidence of carbapenem heteroresistance, to characterize the traits of the heteroresistant subpopulations, and to assess the efficacies of carbapenems against heteroresistant A. baumannii isolates. The study included carbapenem-susceptible A. baumannii clin- ical isolates that cause infections; such infections are com- monly treated with carbapenems, although it is recognized that the heterogeneity of resistance might affect the outcome. The study also determined how the heterogeneity of resistance might affect the treatment outcome. MATERIALS AND METHODS Study population and susceptibility testing. All single A. baumannii clinical isolates that were recovered in the University Hospital of Larissa (Larissa, Greece), a tertiary-care hospital with 700 beds, over a 6-month period from September 2007 to March 2008 were recorded. The study included all isolates that were initially identified as carbapenem susceptible by use of the Vitek 2 system (bioMe ´rieux, Marcy l’E ´ toile, France) to investigate their putative hetero- resistant phenotype. Imipenem and meropenem MICs were also tested by agar dilution (4). The susceptibilities of the native isolates and their meropenem- heteroresistant subpopulations to other -lactams, aminoglycosides, quinolones, and colistin were determined by disk diffusion. Pseudomonas aeruginosa ATCC 27853 was used as a control. PFGE and ST. The clinical isolates as well as their meropenem-heteroresistant subpopulations were tested by pulsed-field gel electrophoresis (PFGE) of ApaI- digested genomic DNA. The banding patterns were compared by using the criteria proposed by Tenover et al. (23). Sequence typing (ST) was performed by use of a previously described scheme (27). The sequences of both strands were determined at Lark Technologies, Inc. (Takeley, United Kingdom), and were analyzed by using DNAStar software (version 5.07; Lasergene, Madison, WI). STs were assigned by using the scheme that has recently been developed by J. F. Turton and R. Meyers (http://www.hpa-bioinformatics.org.uk/AB/home.php). PCR assays. PCR assays for genes coding for known class B and D carba- penemases (bla IMP , bla VIM , bla SPM , bla SIM, bla GIM , bla OXA-23-like , bla OXA-24-like , bla OXA-58-like ) were performed as described previously (3, 11, 14, 18). Amplifi- cation of the intrinsic bla OXA-51 allele (28) was also done to confirm the pheno- typic identification of the isolates as A. baumannii. Insertion sequences ISAbA1 and ISAbA3 were amplified, and screening for their possible location upstream of the class D carbapenemase genes bla OXA-51 and bla OXA-58 was performed by PCR mapping with primers ISAbA1 forward and OXA-51-like reverse and prim- ers ISAbA3 forward and OXA-58 reverse, respectively (24). The nucleotide sequences of both strands of the PCR products were determined at Lark Tech- nologies, Inc., and sequence analysis was carried out with DNAStar software (version 5.07; Lasergene, Madison, WI). Population analysis and investigation of the stability of the heterogeneous phenotype. Population analyses with imipenem and meropenem were performed by a previously described protocol for P. aeruginosa (19), with some modifica- tions. Briefly, subpopulations were yielded by spreading approximately 10 8 bac- terial CFU (100 l of a starting inoculum containing approximately 10 9 CFU/ml) on Mueller-Hinton agar plates with imipenem or meropenem in serial dilutions at concentrations ranging from 0.5 to 32 mg/liter and incubating the plates for 48 h. The analysis was performed three times for all isolates, and the mean numbers of viable CFU were estimated and plotted on a semilogarithmic graph. * Corresponding author. Mailing address: University Hospital of Larissa, Mezourlo Area 41110, Larissa, Greece. Phone: 0030 2410 682537. Fax: 0030 2410 681570. E-mail: [email protected]. Published ahead of print on 14 October 2009. 4055 on June 30, 2018 by guest http://jcm.asm.org/ Downloaded from
Transcript of Heteroresistance to Meropenem in Carbapenem...

JOURNAL OF CLINICAL MICROBIOLOGY, Dec. 2009, p. 4055–4059 Vol. 47, No. 120095-1137/09/$12.00 doi:10.1128/JCM.00959-09Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Heteroresistance to Meropenem in Carbapenem-SusceptibleAcinetobacter baumannii�
Alexandros Ikonomidis,1* Evangelia Neou,1 Vassiliki Gogou,1 Georgia Vrioni,2Athanassios Tsakris,2 and Spyros Pournaras1
University of Thessaly, Mezourlo, Larissa, Greece,1 and University of Athens, Athens 11527, Greece2
Received 13 May 2009/Returned for modification 29 June 2009/Accepted 26 September 2009
The characteristics of carbapenem heteroresistance were studied in 14 apparently carbapenem-susceptibleAcinetobacter baumannii isolates. The MICs for carbapenems were determined, and the isolates were genotypedby pulsed-field gel electrophoresis (PFGE) and sequence typing (ST). Population analysis, testing of thestability of the heteroresistant subpopulations, and time-killing assays were performed. The agar dilutionMICs of both imipenem and meropenem for the native isolates ranged from 0.25 to 4 mg/liter. The isolatesbelonged to nine PFGE types and exhibited seven ST allelic profiles. Population analysis revealed subpopu-lations that grew in the presence of imipenem at concentrations of up to 8 mg/liter and meropenem atconcentrations of up to 32 mg/liter. The meropenem-heteroresistant subpopulations of 11 isolates exhibitedstable resistance with MICs that ranged from 16 to >32 mg/liter; their PFGE profiles were identical to thoseof the native isolates. Time-kill assays with meropenem revealed less pronounced killing for 10 isolates. Thesefindings indicate that meropenem pressure can produce meropenem-heteroresistant subpopulations thatmight subsequently select for highly resistant strains.
Acinetobacter baumannii is an opportunistic pathogen asso-ciated with severe hospital infections (1, 16), which often needthe use of carbapenems as the treatment of last resort. How-ever, reduced susceptibility or resistance to carbapenems isincreasingly being observed among A. baumannii clinical iso-lates (2, 5, 25).
In a preliminary work, we reported on the growth of distinctcolonies within the inhibition halo around carbapenem disks orEtest strips and assessed these colonies as having the pheno-typic manifestation of carbapenem heteroresistance (17). Like-wise, a recent study (12) has described colistin-heteroresistantsubpopulations among apparently colistin-susceptible A. bau-mannii clinical isolates, implying an intrinsic potential of thespecies to overcome drug pressure. These observations suggestthat the use of carbapenems or colistin to treat severe multi-drug-resistant A. baumannii infections may lead to the devel-opment of resistance.
The present study aimed to evaluate in vitro the incidence ofcarbapenem heteroresistance, to characterize the traits of theheteroresistant subpopulations, and to assess the efficacies ofcarbapenems against heteroresistant A. baumannii isolates.The study included carbapenem-susceptible A. baumannii clin-ical isolates that cause infections; such infections are com-monly treated with carbapenems, although it is recognized thatthe heterogeneity of resistance might affect the outcome. Thestudy also determined how the heterogeneity of resistancemight affect the treatment outcome.
MATERIALS AND METHODS
Study population and susceptibility testing. All single A. baumannii clinicalisolates that were recovered in the University Hospital of Larissa (Larissa,Greece), a tertiary-care hospital with 700 beds, over a 6-month period fromSeptember 2007 to March 2008 were recorded. The study included all isolatesthat were initially identified as carbapenem susceptible by use of the Vitek 2system (bioMerieux, Marcy l’Etoile, France) to investigate their putative hetero-resistant phenotype. Imipenem and meropenem MICs were also tested by agardilution (4). The susceptibilities of the native isolates and their meropenem-heteroresistant subpopulations to other �-lactams, aminoglycosides, quinolones,and colistin were determined by disk diffusion. Pseudomonas aeruginosa ATCC27853 was used as a control.
PFGE and ST. The clinical isolates as well as their meropenem-heteroresistantsubpopulations were tested by pulsed-field gel electrophoresis (PFGE) of ApaI-digested genomic DNA. The banding patterns were compared by using thecriteria proposed by Tenover et al. (23). Sequence typing (ST) was performed byuse of a previously described scheme (27). The sequences of both strands weredetermined at Lark Technologies, Inc. (Takeley, United Kingdom), and wereanalyzed by using DNAStar software (version 5.07; Lasergene, Madison, WI).STs were assigned by using the scheme that has recently been developed by J. F.Turton and R. Meyers (http://www.hpa-bioinformatics.org.uk/AB/home.php).
PCR assays. PCR assays for genes coding for known class B and D carba-penemases (blaIMP, blaVIM, blaSPM, blaSIM, blaGIM, blaOXA-23-like, blaOXA-24-like,blaOXA-58-like) were performed as described previously (3, 11, 14, 18). Amplifi-cation of the intrinsic blaOXA-51 allele (28) was also done to confirm the pheno-typic identification of the isolates as A. baumannii. Insertion sequences ISAbA1and ISAbA3 were amplified, and screening for their possible location upstreamof the class D carbapenemase genes blaOXA-51 and blaOXA-58 was performed byPCR mapping with primers ISAbA1 forward and OXA-51-like reverse and prim-ers ISAbA3 forward and OXA-58 reverse, respectively (24). The nucleotidesequences of both strands of the PCR products were determined at Lark Tech-nologies, Inc., and sequence analysis was carried out with DNAStar software(version 5.07; Lasergene, Madison, WI).
Population analysis and investigation of the stability of the heterogeneousphenotype. Population analyses with imipenem and meropenem were performedby a previously described protocol for P. aeruginosa (19), with some modifica-tions. Briefly, subpopulations were yielded by spreading approximately 108 bac-terial CFU (100 �l of a starting inoculum containing approximately 109 CFU/ml)on Mueller-Hinton agar plates with imipenem or meropenem in serial dilutionsat concentrations ranging from 0.5 to 32 mg/liter and incubating the plates for48 h. The analysis was performed three times for all isolates, and the meannumbers of viable CFU were estimated and plotted on a semilogarithmic graph.
* Corresponding author. Mailing address: University Hospital ofLarissa, Mezourlo Area 41110, Larissa, Greece. Phone: 0030 2410682537. Fax: 0030 2410 681570. E-mail: [email protected].
� Published ahead of print on 14 October 2009.
4055
on June 30, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

P. aeruginosa ATCC 27853 was used as a control for the population analysisexperiments. The frequency of appearance of heteroresistant subpopulations inthe presence of the highest drug concentration was calculated by dividing thenumber of colonies that grew on the antibiotic-containing plate by the colonycounts from the same bacterial inoculum that grew on antibiotic-free plates. Foreach strain, three distinct colonies grown in the presence of the highest drugconcentration were subcultured daily in antibiotic-free medium for 1 week, andthe carbapenem MICs were tested by agar dilution on Mueller-Hinton agarplates to check the stability of the heteroresistant phenotypes.
Meropenem time-killing assays. Killing curve assays with meropenem wereperformed with all isolates, as well as P. aeruginosa ATCC 27853, by previouslydescribed protocols for A. baumannii (12) and gram-positive bacteria (21, 22).Briefly, from a starting bacterial inoculum containing approximately 1.5 � 108
CFU/ml, 100 �l was inoculated into 3 ml of Mueller-Hinton broth (final inocu-
lum, approximately 5 � 106 CFU/ml) containing meropenem at a concentrationof 4 mg/liter. This meropenem concentration was selected, as it was the meansteady-state antibiotic level achieved in sera when the usual doses of meropenemwere administered to healthy volunteers (10). Viable cells were counted byspreading 50-�l aliquots of 10�2 and 10�4 dilutions of the broth cultures inantibiotic-free Muller Hinton agar plates at 1, 3, 6, 9, 12, 24, 36, and 48 h afterantibiotic addition. The analysis was performed three times for all isolates, andthe mean numbers of viable CFU were estimated and plotted on a semilogarith-mic graph.
RESULTS
During the study period, 142 nonrepetitive A. baumanniiisolates were recovered from various wards and clinical sam-ples. Among them, 14 (9.9%) were classified as carbapenemsusceptible by use of the Vitek 2 automated system and werefurther investigated. The isolates were also susceptible to colis-tin and exhibited various profiles of susceptibility to otherantimicrobials used to treat A. baumannii infections (Table 1).PFGE analysis discriminated nine distinct genotypes amongthe 14 study isolates. For each clinical isolate, the native andthe meropenem-heteroresistant populations exhibited identi-cal PFGE profiles. By ST, six isolates belonged to ST1, threebelonged to ST2, and five exhibited various allelic profiles thatwere not assigned a separate ST because they have not yetbeen found among outbreak strains (Table 2). PCR forgenes encoding known carbapenemases (except the intrinsicblaOXA-51-like carbapenemase) was negative. Nucleotide se-quencing showed that the blaOXA-66 and blaOXA-69 alleleswere more common, as anticipated, according to a previousnationwide study (8). No ISAba1 elements were detectedupstream of the gene blaOXA-51-like carbapenemase.
Time-kill studies. In time-kill studies with meropenem, fourisolates (isolates AB5, AB13, AB79, and AB135) were killed ina time-dependent manner. For the remaining isolates, the ini-
TABLE 1. Antimicrobial susceptibility profile of native andmeropenem-heteroresistant colonies
Isolate
Antimicrobial(s) to which isolate was susceptiblea
Native colonies Meropenem-heteroresistant colonies
AB5 SAM -AB13 AMK, PTZ, SAM AMK, PTZ, SAMAB27 PTZ, CIP, SAM PTZ, CIP, SAMAB32 PTZ, SAM PTZ, SAMAB49 SAM SAMAB68 AMK, SAM AMK, SAMAB71 PTZ, SAM PTZ, SAMAB72 SAM SAMAB78 AMK, SAM AMKAB79 AMK AMKAB119 AMK, CIP, PTZ, SAM AMK, CIP, PTZ, SAMAB129AB133 AMK, PTZ, SAM AMKAB135 SAM SAM
a AMK, amikacin; CIP, ciprofloxacin; PTZ, piperacillin-tazobactam; SAM,ampicillin-sulbactam. All clinical isolates were susceptible to imipenem, mero-penem, and colistin.
TABLE 2. Characteristics of the tested isolates
Isolate Date of isolation(day/mo/yr) Ward Specimen blaOXA-51-like
alleleAllelicprofilea
STtypeb
PFGEtype
Agar dilutionMIC (mg/
liter)
Highestconcn ofgrowth in
populationanalyses
(mg/liter)
Agar MIC (mg/liter) ofheterogenous
subpopulations inc:
IPM MEM
IPMd MEMe IPM MEM IPM MEM IPM MEM
AB5 09/10/07 Cardiology Sputum 66 1-1-1 1 I 2 4 4 8 4 2 2 32AB13 28/09/07 Medical Decubitus ulcer 66 1-1-1 1 VI 2 0.5 2 2 2 0.5 4 4AB27 15/09/07 ICUf Bronchial 91 1d-4-1c* NA VII 0.25 1 1 8 0.5 1 0.5 �32AB32 31/10/07 ICU Bronchial 66 7-4-1a* NA V 1 2 1 8 2 1 1 16AB49 26/11/07 Oncology Blood 66 1a-1-1 1 I 4 4 8 16 4 2 1 32AB68 19/12/07 Oncology Sputum 66 1a-1-1 1 I 2 2 8 32 2 8 2 �32AB71 28/11/07 Medical Blood 66 1a-1-1 1 I 0.25 0.25 4 8 1 1 1 32AB72 02/12/07 Orthopedic Pus 94 7-2-7* NA II 1 2 2 8 0.5 1 1 32AB78 07/01/08 Medical Blood 94 7-2-10* NA II 1 2 2 8 2 1 2 32AB79 16/01/08 Medical Decubitus ulcer 69 2-2-2 2 III 0.5 1 2 8 0.5 1 1 8AB119 07/02/08 Neurosurgery Sputum 66 4-2-7* NA VIII 0.25 1 1 8 0.5 1 0.5 8AB129 28/02/08 Neurosurgery Blood 69 2-2-2 2 III 2 1 8 16 1 1 2 �32AB133 27/03/08 Orthopedic Pus 66 1-1-1 1 IX 2 2 4 32 2 1 1 �32AB135 31/03/08 Oncology Sputum 69 2-2-2 2 IV 1 1 4 16 1 1 1 16
a Alleles ompA, csuE, and blaoxa-51-like, respectively.b NA, not assigned, as it did not correspond to outbreak strains. Asterisks, all three alleles correspond to the closest matches.c These MICs were estimated after 1 week of daily subculture in antibiotic-free medium.d IPM, imipenem.e MEM, meropenem.f ICU, intensive care unit.
4056 IKONOMIDIS ET AL. J. CLIN. MICROBIOL.
on June 30, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

tial bactericidal effect was followed by substantial regrowth,which was observed after 9 h to 12 h of incubation for threeisolates (isolates AB27, AB49, and AB68) and after 24 h forseven isolates (isolates AB32, AB71, AB72, AB78, AB119,AB129, and AB133). The survival curves obtained from time-kill studies are shown in Fig. 1.
In vitro characterization of the heteroresistant subpopula-tions. The agar dilution MICs of imipenem for the clinicalisolates ranged from 0.25 to 4 mg/liter. Population analysisassays with imipenem showed that colonies grew in the pres-ence of 8 mg/liter for 3 isolates, whereas the 11 isolates thatremained were within the susceptibility range. However, theheterogeneous growth in the presence of imipenem was notstable; after seven daily subcultures in drug-free medium, theimipenem MICs for the colonies grown in the presence ofthe highest imipenem concentration were similar to thosefor the native isolates (Table 2).
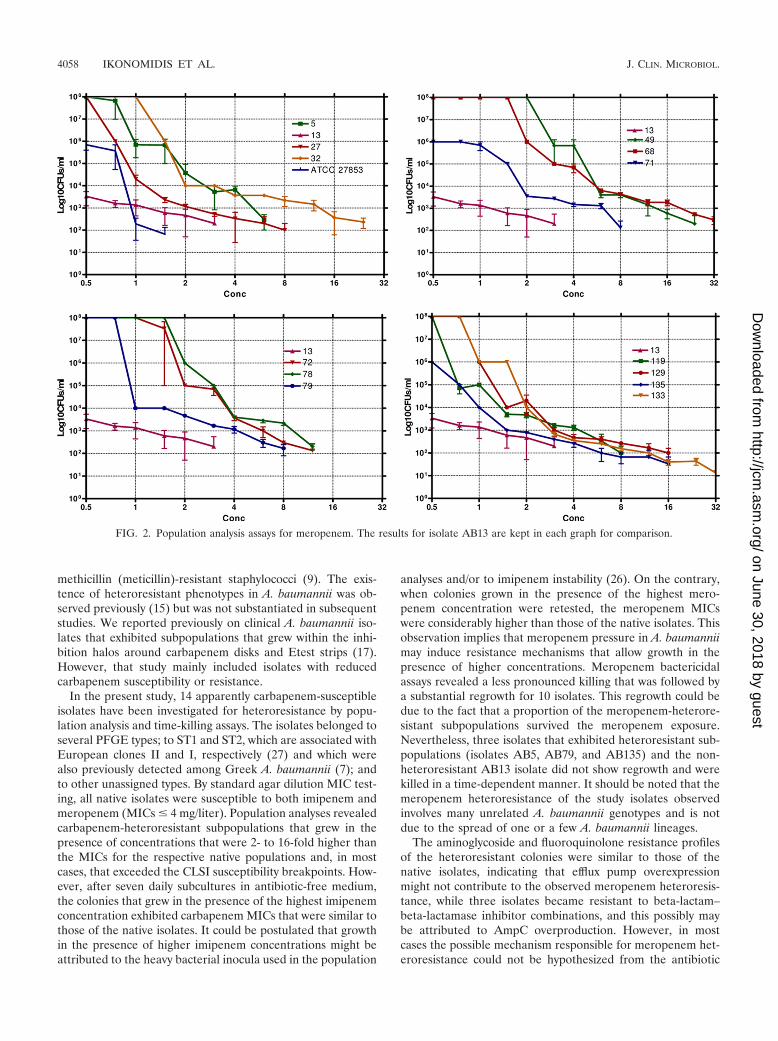
The agar dilution MICs of meropenem for the clinical iso-lates ranged from 0.25 to 4 mg/liter. Population analysis assayswith meropenem revealed colonies that grew in the presence of8 to 32 mg/liter for 13 isolates and in the presence of up to 2mg/liter for isolate AB13 (Table 2; Fig. 2). The heteroresistant
colonies showed stable meropenem resistance when they wereretested by agar dilution after seven daily subcultures in drug-free medium but showed susceptibility to imipenem (Table 2).The frequency of meropenem-heteroresistant subpopulations,as calculated by the population analysis assays, ranged from3 � 10�7 to 5 � 10�5. The antibiotic resistance profiles of mostof the heteroresistant colonies did not exhibit considerabledifferences from the profiles of the native isolates for amino-glycosides, fluoroquinolones, and beta-lactam antibiotics;three isolates, however, acquired resistance to ampicillin-sul-bactam, and one isolate also acquired resistance to piperacil-lin-tazobactam (Table 1).
DISCUSSION
During the last few years, A. baumannii isolates that exhibitcarbapenem resistance have increasingly been isolated andpose substantial therapeutic problems in many regions, includ-ing the United States (6, 20). A further worrisome observationis heteroresistance to carbapenems, which may have implica-tions for the treatment of multiresistant A. baumannii infec-tions. Heteroresistance to meropenem was first detected in
FIG. 1. Time-killing assays for meropenem. The results for isolate AB13 are kept in each graph for comparison.
VOL. 47, 2009 CARBAPENEM-HETERORESISTANT A. BAUMANNII 4057
on June 30, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
methicillin (meticillin)-resistant staphylococci (9). The exis-tence of heteroresistant phenotypes in A. baumannii was ob-served previously (15) but was not substantiated in subsequentstudies. We reported previously on clinical A. baumannii iso-lates that exhibited subpopulations that grew within the inhi-bition halos around carbapenem disks and Etest strips (17).However, that study mainly included isolates with reducedcarbapenem susceptibility or resistance.
In the present study, 14 apparently carbapenem-susceptibleisolates have been investigated for heteroresistance by popu-lation analysis and time-killing assays. The isolates belonged toseveral PFGE types; to ST1 and ST2, which are associated withEuropean clones II and I, respectively (27) and which werealso previously detected among Greek A. baumannii (7); andto other unassigned types. By standard agar dilution MIC test-ing, all native isolates were susceptible to both imipenem andmeropenem (MICs � 4 mg/liter). Population analyses revealedcarbapenem-heteroresistant subpopulations that grew in thepresence of concentrations that were 2- to 16-fold higher thanthe MICs for the respective native populations and, in mostcases, that exceeded the CLSI susceptibility breakpoints. How-ever, after seven daily subcultures in antibiotic-free medium,the colonies that grew in the presence of the highest imipenemconcentration exhibited carbapenem MICs that were similar tothose of the native isolates. It could be postulated that growthin the presence of higher imipenem concentrations might beattributed to the heavy bacterial inocula used in the population
analyses and/or to imipenem instability (26). On the contrary,when colonies grown in the presence of the highest mero-penem concentration were retested, the meropenem MICswere considerably higher than those of the native isolates. Thisobservation implies that meropenem pressure in A. baumanniimay induce resistance mechanisms that allow growth in thepresence of higher concentrations. Meropenem bactericidalassays revealed a less pronounced killing that was followed bya substantial regrowth for 10 isolates. This regrowth could bedue to the fact that a proportion of the meropenem-heterore-sistant subpopulations survived the meropenem exposure.Nevertheless, three isolates that exhibited heteroresistant sub-populations (isolates AB5, AB79, and AB135) and the non-heteroresistant AB13 isolate did not show regrowth and werekilled in a time-dependent manner. It should be noted that themeropenem heteroresistance of the study isolates observedinvolves many unrelated A. baumannii genotypes and is notdue to the spread of one or a few A. baumannii lineages.
The aminoglycoside and fluoroquinolone resistance profilesof the heteroresistant colonies were similar to those of thenative isolates, indicating that efflux pump overexpressionmight not contribute to the observed meropenem heteroresis-tance, while three isolates became resistant to beta-lactam–beta-lactamase inhibitor combinations, and this possibly maybe attributed to AmpC overproduction. However, in mostcases the possible mechanism responsible for meropenem het-eroresistance could not be hypothesized from the antibiotic
FIG. 2. Population analysis assays for meropenem. The results for isolate AB13 are kept in each graph for comparison.
4058 IKONOMIDIS ET AL. J. CLIN. MICROBIOL.
on June 30, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

resistance profiles; the underlying mechanisms are currentlyunder investigation.
The medical records of patients treated with meropenemshowed that six patients were given meropenem for at least 1week and that three of them died from an A. baumannii blood-stream infection while they were receiving meropenem (datanot shown). A relevant publication by Nunez et al. described acase of fatal Acinetobacter meningitis (13). In that case, nineclonally related strains were isolated from cerebrospinal fluid;the first eight strains were carbapenem susceptible, but theninth one was meropenem resistant (MIC � 32 mg/liter) andstill imipenem susceptible. The meropenem-resistant strainwas isolated after two courses of meropenem treatment. Thisobservation is in accordance with our findings, suggesting aresistance mechanism that affects meropenem but not imi-penem and that might result from meropenem pressure.
Conclusion. The observations of the present study suggestthat A. baumannii isolates that are apparently meropenemsusceptible by standard susceptibility testing may contain acertain amount of meropenem-resistant subpopulations, andthose subpopulations could be selected for by the use of sub-optimal therapeutic drug dosages. In this context, the imple-mentation of screening techniques for the identification ofheteroresistant isolates and the elucidation of the underlyingmolecular mechanisms would be of significant importance.
REFERENCES
1. Bergogne-Berezin, E., and K. J. Towner. 1996. Acinetobacter spp. as noso-comial pathogens: microbiological, clinical, and epidemiological features.Clin. Microbiol. Rev. 9:148–165.
2. Bogaerts, P., T. Naas, I. Wybo, et al. 2006. Outbreak of infection by carba-penem-resistant Acinetobacter baumannii producing the carbapenemaseOXA-58 in Belgium. J. Clin. Microbiol. 44:4189–4192.
3. Castanheira, M., M. A. Toleman, R. N. Jones, F. J. Schmidt, and T. R.Walsh. 2004. Molecular characterization of a ß-lactamase gene, blaGIM-1,encoding a new subclass of metallo-ß-lactamase. Antimicrob. Agents Che-mother. 48:4654–4661.
4. Clinical and Laboratory Standards Institute. 2006. Performance standardsfor antimicrobial susceptibility testing, 16th informational supplement.M100-S16. Clinical and Laboratory Standards Institute, Wayne, PA.
5. Coelho, J. M., J. F. Turton, M. E. Kaufmann, J. Glover, N. Woodford, M.Warner, M. F. Palepou, R. Pike, T. L. Pitt, B. C. Patel, and D. M. Livermore.2006. Occurrence of carbapenem-resistant Acinetobacter baumannii clones atmultiple hospitals in London and Southeast England. J. Clin. Microbiol.44:3623–3627.
6. Gaynes, R., J. R. Edwards, and the National Nosocomial Infections Surveil-lance System. 2005. Overview of nosocomial infections caused by gram-negative bacilli. Clin. Infect. Dis. 41:848–854.
7. Giannouli, M., F. Tomasone, A. Agodi, H. Vahaboglu, Z. Daoud, M. Triassi,A. Tsakris, and R. Zarrilli. 2009. Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii strains in intensive care units of multipleMediterranean hospitals. J. Antimicrob. Chemother. 63:828–830.
8. Ikonomidis, A., S. Pournaras, A. Markogiannakis, A. N. Maniatis, E. N.Manolis, and A. Tsakris. 2007. Low genetic diversity of the intrinsic OXA-51-like class D carbapenemases among Acinetobacter baumannii clinical iso-lates in Greece. Epidemiol. Infect. 135:877–881.
9. Kayser, F. H., G. Morenzoni, A. Strassle, and K. Hadorn. 1989. Activity ofmeropenem, against gram-positive bacteria. J. Antimicrob. Chemother.24(Suppl. A):101–112.
10. Lee, C. H., L. H. Su, Y. F. Tang, and J. W. Liu. 2006. Treatment of ESBL-
producing Klebsiella pneumoniae bacteraemia with carbapenems or flo-moxef: a retrospective study and laboratory analysis of the isolates. J. Anti-microb. Chemother. 58:1074–1077.
11. Lee, K., J. H. Yum, D. Yong, H. M. Lee, H. D. Kim, J.-D. Docquier, G. M.Rossolini, and Y. Chong. 2005. Novel acquired metallo-ß-lactamase gene,blaSIM-1, in a class 1 integron from Acinetobacter baumannii clinical isolatesfrom Korea. Antimicrob. Agents Chemother. 49:4485–4491.
12. Li, J., C. R. Rayner, R. L. Nation, R. J. Owen, D. Spelman, K. E. Tan, andL. Liolios. 2006. Heteroresistance to colistin in multidrug-resistant Acineto-bacter baumannii. Antimicrob. Agents Chemother. 50:2946–2950.
13. Nunez, M. L., M. C. Martinez-Toldos, M. Bru, E. Simarro, M. Segovia, andJ. Ruiz. 1998. Appearance of resistance to meropenem during the treatmentof a patient with meningitis by Acinetobacter. Scand. J. Infect. Dis. 30:421–423.
14. Oh, E. J., S. Lee, Y. J. Park, J. J. Park, K. Park, S. I. Kim, M. W. Kang, andB. K. Kim. 2003. Prevalence of metallo-�-lactamase among Pseudomonasaeruginosa and Acinetobacter baumannii in a Korean university hospital andcomparison of screening methods for detecting metallo-�-lactamase. J. Mi-crobiol. Methods 54:411–418.
15. Pascual, A., L. Martinez-Martinez, M. J. Clavijo, M. D. García-Perea, andE. J. Perea. 1997. Comparison of three methods of determining the in-vitrosusceptibilities of Acinetobacter baumannii isolates to imipenem. J. Antimi-crob. Chemother. 40:742–743.
16. Peleg, A. Y., H. Seifert, and D. L. Paterson. 2008. Acinetobacter baumannii:emergence of a successful pathogen. Clin. Microbiol. Rev. 21:538–582.
17. Pournaras, S., A. Ikonomidis, A. Markogiannakis, A. N. Maniatis, and A.Tsakris. 2005. Heteroresistance to carbapenems in Acinetobacter baumannii.J. Antimicrob. Chemother. 55:1055–1056.
18. Pournaras, S., A. Markogiannakis, A. Ikonomidis, L. Kondyli, K. Bethi-mouti, A. N. Maniatis, N. J. Legakis, and A. Tsakris. 2006. Outbreak ofmultiple clones of imipenem-resistant Acinetobacter baumannii isolates ex-pressing OXA-58 carbapenemase in an intensive care unit. J. Antimicrob.Chemother. 57:557–561.
19. Pournaras, S., A. Ikonomidis, A. Markogiannakis, N. Spanakis, A. N. Ma-niatis, and A. Tsakris. 2007. Characterization of clinical isolates of Pseudo-monas aeruginosa heterogeneously resistant to carbapenems. J. Med. Micro-biol. 56:66–70.
20. Quale, J., S. Bratu, D. Landman, and R. Heddurshetti. 2003. Molecularepidemiology and mechanisms of carbapenem resistance in Acinetobacterbaumannii endemic in New York City. Clin. Infect. Dis. 37:214–220.
21. Sakoulas, G., J. Alder, C. Thauvin-Eliopoulos, R. C. Moellering, Jr., andG. M. Eliopoulos. 2006. Induction of daptomycin heterogeneous susceptibil-ity in Staphylococcus aureus by exposure to vancomycin. Antimicrob. AgentsChemother. 50:1581–1585.
22. Swenson, J. M., J. F. Hindler, and J. H. Jorgensen. 2003. Special phenotypicmethods for detecting antimicrobial resistance, p. 1178–1195. In P. R. Mur-ray, E. J. Baron, J. H. Jorgensen, M. A. Pfaller, and R. H. Yolken (ed.),Manual of clinical microbiology, 8th ed. American Society for Microbiology,Washington, DC.
23. Tenover, F. C., R. D. Arbeit, R. V. Goering, P. A. Mickelsen, B. E. Murray,D. H. Persing, and B. Swaminathan. 1995. Interpreting chromosomal DNArestriction patterns produced by pulsed-field gel electrophoresis: criteria forbacterial strain typing. J. Clin. Microbiol. 33:2233–2239.
24. Tsakris, A., A. Ikonomidis, S. Pournaras, N. Spanakis, and A. Markogian-nakis. 2006. Carriage of OXA-58 but not of OXA-51 �-lactamase genecorrelates with carbapenem resistance in Acinetobacter baumannii. J. Anti-microb. Chemother. 58:1097–1099.
25. Tsakris, A., C. Tsioni, S. Pournaras, S. Polyzos, and A. N. Maniatis. 2003.Spread of low-level carbapenem-resistant Acinetobacter baumannii in a ter-tiary care Greek hospital. J. Antimicrob. Chemother. 52:1046–1047.
26. Turgeon, P. L., and C. Desrochers. 1985. Stability of imipenem in Mueller-Hinton agar stored at 4°C. Antimicrob. Agents Chemother. 28:711–712.
27. Turton, J. F., S. N. Gabriel, C. Valderrey, M. E. Kaufmann, and T. L. Pitt.2007. Use of sequence-based typing and multiplex PCR to identify clonallineages of outbreak strains of Acinetobacter baumannii. Clin. Microbiol.Infect. 13:807–815.
28. Turton, J. F., N. Woodford, J. Glover, S. Yarde, M. E. Kaufmann, and T. L.Pitt. 2006. Identification of Acinetobacter baumannii by detection of theblaOXA-51-like carbapenemase gene intrinsic to this species. J. Clin. Micro-biol. 44:2974–2976.
VOL. 47, 2009 CARBAPENEM-HETERORESISTANT A. BAUMANNII 4059
on June 30, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from