![Decreased GABAA receptors and benzodiazepine · PDF fileBenson, Luscher, & Fritschy, 1995a]. ... B3 using the Solver tool of Excel (Microsoft Office XP Professional)toconstruct astandardcurve,whichwas](https://static.fdocuments.us/doc/165x107/5a76874a7f8b9aa3618d49b4/decreased-gabaa-receptors-and-benzodiazepine-benson-luscher-fritschy.jpg)
Heterogeneity of responses to agents that act at the benzodiazepine site on GABAA receptors of...
3
SYNAPSE 11:259-261 (1992) Short Communications Heterogeneity of Responses to Agents That Act at the Benzodiazepine Site on GABA, Receptors of Dorsal Root Ganglion Neurons Freshly Isolated From Adult Rats GEOFFREY WHITE Neurogen Corporation, Branford, Connecticut 06405 KEY WORDS Inhibition, DRG, Patch-clamp, Ion channel GABA exerts an influence on neuronal excitability by activating a C1- current through the GABAAreceptor1 ionophore complex. This complex is a heteromultimeric protein (Boormann et al., 1987; Olsen and Tobin, 1990; Vicini, 1991), whose various subunits are differentially expressed within the brain and possibly within neurons themselves (Juiz et al., 1989; Olsen and Tobin, 1990; Vicini, 1991; Zimprich et al., 1991). Such observations raise the possibility that a topographical pattern of pharmacological and physiological properties of GABA, receptorlionophore complexes exists within the nervous system (Olsen and Tobin, 1990; Vicini, 1991). Functional heterogeneity is well documented in oocytes and transfected cultured mammalian tumor cells with respect to sensitivity to GABA and agents that act at the benzodiazepine receptor (Puia et al., 1991; Sigel et al., 1990). Functional heterogeneity in sensitivity to GABA has recently been demonstrated for neurons iso- lated from adult rats (White, 1992). No electrophysio- logical studies have systematically identified heteroge- neity in responses to benzodiazepines €or intrinsically expressed GABAAreceptor complexes in neurons from adult mammals. This report details heterogeneity in such responses and suggests that individual neurons can express more than one benzodiazepine receptor subtype. Male Sprague-Dawley rats (200-500 g) were anes- thetized with chloral hydrate (400 mg/kg) and decapi- tated. Dorsal root ganglia (DRG) were removed and placed in oxygenated DMEM, 0.4 mglml type I11 trypsin, 0.3 mg/ml type VII collagenase, and 0.1 mg/ml type IV DNase 11. This solution was incubated at 35°C for 45-60 minutes. Individual neurons were dissociated from the ganglia by vigorous shaking. Neurons were devoid of processes as described previously (White et al., 1990). Electrophysiological responses were obtained from 40 neurons using the whole-cell patch-clamp technique 0 1992 WILEY-LISS, INC. with an Axopatch 1D amplifier (Axon Instruments) and pClamp software (Axon Instruments) as described pre- viously (White, 1990). Briefly, subsequent to gaining access to the cell interior, membrane potential was volt- age-clamped to -60 mV. Series resistance (3-6 mn) was compensated by 90%. Current amplitude was <1.5 nA. The external solution contained (mM): 150 NaCl, 5 KC1, 2 CaCl,, 1 MgCl,, 10 HEPES, and 10 glucose; osmolarity was adjusted to 320 using sucrose and pH was adjusted to 7.4 using NaOH. The recording elec- trode solution contained (mM): 130 KC1, 1 CaCl,, 2 MgCl,, 11 EGTA, 10 HEPES, 5 mM MgATP, and 0.5 mM GTP; osmolarity was adjusted to 310 using sucrose and pH was adjusted to 7.4 using KOH. GABA was dissolved in double-distilled/deionized water. Other drugs were dissolved in polyethylene gly- col 400 (PEG) and diluted to a final concentration of 0.002% or less. At this or lower concentrations, PEG exerted no detectable effects on GABA-activated cur- rents. Drugs were applied to neurons using a linear array of 17 200-pm diameter glass tubes placed within 100 pm of the neuron. The delivery rate of the drug- containing solution was -90 plms. Repeated exposure to a given concentration of drug resulted in amplitude differences of less than 5-10%. Concentrations of drugs were usually applied in ascending order. In 5 neurons the initial concentration of GABA was applied after the final drug application until a stable value was reached (3-4 minutes). This value was within 10% of the initial value. In order for the maximal effect of a drug to be ob- served at a given drug concentration, it was necessary to expose the neuron to the drug before applying the drug plus GABA (25 of 25 neurons). Current flow was induced by exposing neurons to 5 pM GABA (Figure 1, Received March 2,1992; accepted in revised form March 25,1992.
-
Upload
geoffrey-white -
Category
Documents
-
view
212 -
download
0
Transcript of Heterogeneity of responses to agents that act at the benzodiazepine site on GABAA receptors of...

SYNAPSE 11:259-261 (1992)
Short Communications
Heterogeneity of Responses to Agents That Act at the Benzodiazepine Site on GABA,
Receptors of Dorsal Root Ganglion Neurons Freshly Isolated From Adult Rats
GEOFFREY WHITE Neurogen Corporation, Branford, Connecticut 06405
KEY WORDS Inhibition, DRG, Patch-clamp, Ion channel
GABA exerts an influence on neuronal excitability by activating a C1- current through the GABAA receptor1 ionophore complex. This complex is a heteromultimeric protein (Boormann et al., 1987; Olsen and Tobin, 1990; Vicini, 1991), whose various subunits are differentially expressed within the brain and possibly within neurons themselves (Juiz et al., 1989; Olsen and Tobin, 1990; Vicini, 1991; Zimprich et al., 1991). Such observations raise the possibility that a topographical pattern of pharmacological and physiological properties of GABA, receptorlionophore complexes exists within the nervous system (Olsen and Tobin, 1990; Vicini, 1991). Functional heterogeneity is well documented in oocytes and transfected cultured mammalian tumor cells with respect to sensitivity to GABA and agents that act at the benzodiazepine receptor (Puia et al., 1991; Sigel et al., 1990). Functional heterogeneity in sensitivity to GABA has recently been demonstrated for neurons iso- lated from adult rats (White, 1992). No electrophysio- logical studies have systematically identified heteroge- neity in responses to benzodiazepines €or intrinsically expressed GABAA receptor complexes in neurons from adult mammals. This report details heterogeneity in such responses and suggests that individual neurons can express more than one benzodiazepine receptor subtype.
Male Sprague-Dawley rats (200-500 g) were anes- thetized with chloral hydrate (400 mg/kg) and decapi- tated. Dorsal root ganglia (DRG) were removed and placed in oxygenated DMEM, 0.4 mglml type I11 trypsin, 0.3 mg/ml type VII collagenase, and 0.1 mg/ml type IV DNase 11. This solution was incubated at 35°C for 45-60 minutes. Individual neurons were dissociated from the ganglia by vigorous shaking. Neurons were devoid of processes as described previously (White et al., 1990).
Electrophysiological responses were obtained from 40 neurons using the whole-cell patch-clamp technique 0 1992 WILEY-LISS, INC.
with an Axopatch 1D amplifier (Axon Instruments) and pClamp software (Axon Instruments) as described pre- viously (White, 1990). Briefly, subsequent to gaining access to the cell interior, membrane potential was volt- age-clamped to -60 mV. Series resistance (3-6 m n ) was compensated by 90%. Current amplitude was <1.5 nA. The external solution contained (mM): 150 NaCl, 5 KC1, 2 CaCl,, 1 MgCl,, 10 HEPES, and 10 glucose; osmolarity was adjusted to 320 using sucrose and pH was adjusted to 7.4 using NaOH. The recording elec- trode solution contained (mM): 130 KC1, 1 CaCl,, 2 MgCl,, 11 EGTA, 10 HEPES, 5 mM MgATP, and 0.5 mM GTP; osmolarity was adjusted to 310 using sucrose and pH was adjusted to 7.4 using KOH.
GABA was dissolved in double-distilled/deionized water. Other drugs were dissolved in polyethylene gly- col 400 (PEG) and diluted to a final concentration of 0.002% or less. At this or lower concentrations, PEG exerted no detectable effects on GABA-activated cur- rents. Drugs were applied to neurons using a linear array of 17 200-pm diameter glass tubes placed within 100 pm of the neuron. The delivery rate of the drug- containing solution was -90 plms. Repeated exposure to a given concentration of drug resulted in amplitude differences of less than 5-10%. Concentrations of drugs were usually applied in ascending order. In 5 neurons the initial concentration of GABA was applied after the final drug application until a stable value was reached (3-4 minutes). This value was within 10% of the initial value.
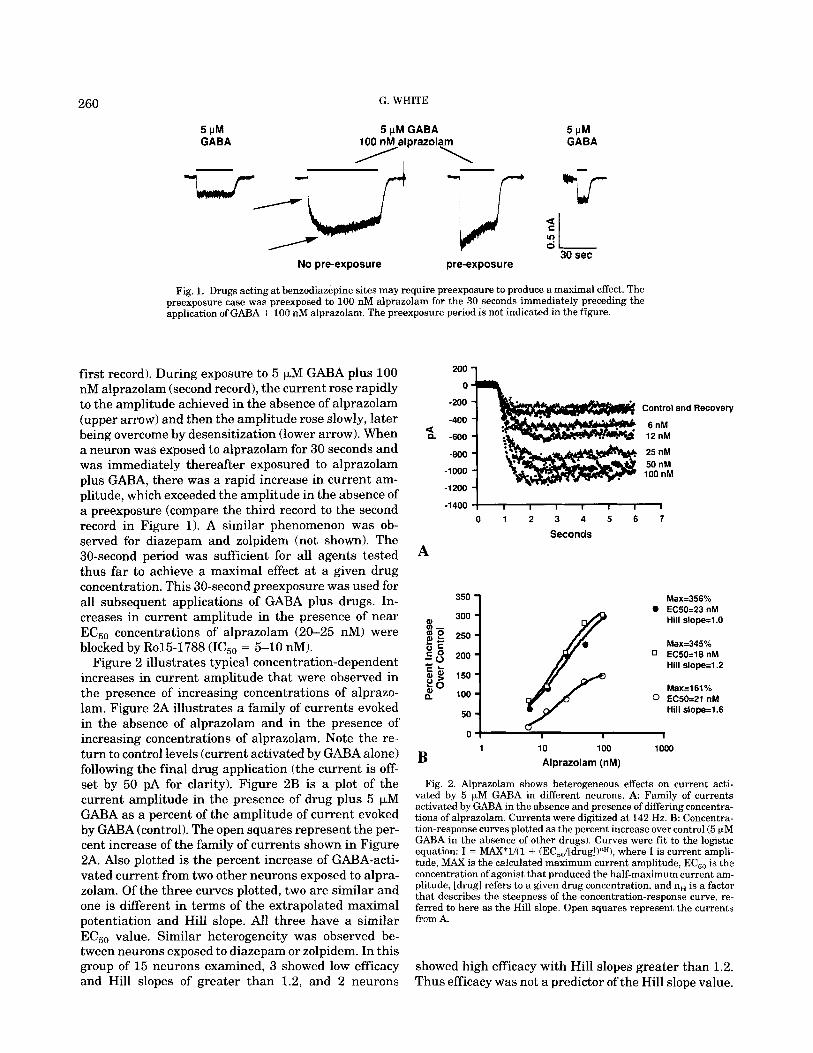
In order for the maximal effect of a drug to be ob- served at a given drug concentration, it was necessary to expose the neuron to the drug before applying the drug plus GABA (25 of 25 neurons). Current flow was induced by exposing neurons to 5 p M GABA (Figure 1,
Received March 2,1992; accepted in revised form March 25,1992.
260
300 - 250 - 2-
c
- L 5 $ 150'
p 100-
P Q 200-
$ 0
50 -
G. WHITE
5 PM GABA
5 pM GABA 100 nM alprazolam
/ , \ 5 PM GABA
- - r v - w -u/u y.' ;I
30 sec /
No pre-exposure pre-exposure
Fig. 1. Drugs acting at benzodiazepine sites may require preexposure to produce a maximal effect. The preexposure case was preexposed to 100 nM alprazolam for the 30 seconds immediately preceding the application of GABA + 100 nM alprazolam. The preexposure period is not indicated in the figure.
first record). During exposure to 5 pM GABA plus 100 nM alprazolam (second record), the current rose rapidly to the amplitude achieved in the absence of alprazolam (upper arrow) and then the amplitude rose slowly, later being overcome by desensitization (lower arrow). When a neuron was exposed to alprazolam for 30 seconds and was immediately thereafter exposured to alprazolam plus GABA, there was a rapid increase in current am- plitude, which exceeded the amplitude in the absence of a preexposure (compare the third record to the second record in Figure 1). A similar phenomenon was ob- served for diazepam and zolpidem (not shown). The 30-second period was sufficient for all agents tested thus far to achieve a maximal effect at a given drug concentration. This 30-second preexposure was used for all subsequent applications of GABA plus drugs. In- creases in current amplitude in the presence of near EC,, concentrations of alprazolam (20-25 nM) were blocked by Ro15-1788 (IC50 = 5-10 nM).
Figure 2 illustrates typical concentration-dependent increases in current amplitude that were observed in the presence of increasing concentrations of alprazo- lam. Figure 2A illustrates a family of currents evoked in the absence of alprazolam and in the presence of increasing concentrations of alprazolam. Note the re- turn to control levels (current activated by GABA alone) following the final drug application (the current is off- set by 50 pA for clarity). Figure 2B is a plot of the current amplitude in the presence of drug plus 5 pM GABA as a percent of the amplitude of current evoked by GABA (control). The open squares represent the per- cent increase of the family of currents shown in Figure 2A. Also plotted is the percent increase of GABA-acti- vated current from two other neurons exposed to alpra- zolam. Of the three curves plotted, two are similar and one is different in terms of the extrapolated maximal potentiation and Hill slope. All three have a similar EC,, value. Similar heterogeneity was observed be- tween neurons exposed to diazepam or zolpidem. In this group of 15 neurons examined, 3 showed low efficacy and Hill slopes of greater than 1.2, and 2 neurons
*0° 0 L Control and Recovery
6 n M 12 nM
25 nM 50 nM 100 nM
-1400 - 0 1 2 3 4 5 6 7
Seconds A
Max=356% 350 1 0 EC50=23nM
Hill slope=l.O
Max=345%
Hill slope=l.2
Max=161%
Hill slope=1.6
0 EC50=18nM
0 EC50=21 nM
o ! I I 1)
1 10 100 1000
B Alprazolam (nM)
Fig. 2. Alprazolam shows heterogeneous effects on current acti- vated by 5 pM GABA in different neurons. A Family of currents activated by GABA in the absence and presence of differing concentra- tions of alprazolam. Currents were digitized at 142 Hz. B: Concentra- tion-response curves plotted as the percent increase over control (5 pM GABA in the absence of other drugs). Curves were fit to the logistic equation: I = MAX*lMl + (EC,Jdrugl)""), where I is current ampli- tude, MAX is the calculated maximum current amplitude, EC,, is the concentration of agonist that produced the half-maximum current am- plitude, [drug] refers to a given drug concentration, and nH is a factor that describes the steepness of the concentration-response curve, re- ferred to here as the Hill slope. Open squares represent the currents from A.
showed high efficacy with Hill slopes greater than 1.2. Thus efficacy was not a predictor of the Hill slope value.

261 MULTIPLE BENZODIAZEPINE SITES IN ADULT NEURONS
Maxd 37% EC50=469 Hill slope=l.41
140 -
m 0 100
2 g 80 - Q u)-
Max=68% EC50=8
60 Hill slope=1.6
8 0 40
20 r=0.9998
&!&
z o
n
- 1 I I I
1 10 100 1000
Zolpidem (nM)
Fig. 3. Plot showing multiple benzodiazepine sites located on a sin- gle neuron. The curve was fit to the sum of two logistic equations (see Fig. 2B legend).
Figure 3 illustrates a plot similar to that of Figure 2B, except that increasing concentrations of zolpidem were applied to the neuron in the presence of 5 p M GABA. The curve is suggestive of two separate binding sites. Curves suggestive of multiple binding sites on a single neuron were observed in 3 of 15 neurons exam- ined. The EC,,s have a difference of 50-fo1d7 which is in agreement with reported differences between type I and type I1 sites for zolpidem in cerebellum and hippo- campal dentate gyrus (Langer et al., 1990).
This report indicates there may be heterogeneity of benzodiazepine receptor-mediated responses in neu- rons from adult rats. Such heterogeneity was predicted from molecular biological experiments in model expres- sion systems (Olsen and Tobin, 1990; Puia et al., 1991; Sigel et al., 1990). The finding that preexposure of neu- rons to drug was necessary to observe the full effects of the drug was unexpected. The requirement of pretreat- ment is probably not an artifact of enzyme treatment because the phenomenon was also observed in fetal hip- pocampal neurons maintained in cell culture for at least 7 days (in preparation). In some instances no po- tentiation was observed unless the neuron was preex- posed to the drug. These observations suggest that GABA can reduce the agonist-like actions of drugs at the benzodiazepine site under the nonequilibrium bind- ing conditions of this study. In fact, the increased rate of fading of GABA-activated currents as reported by Mierlak and Farb (1988) could result from this type of phenomenon. The finding that Hill slopes for agents acting as agonists at the benzodiazepine receptor can
be greater than 1 was unexpected in so far as no reports of this are generally cited or easily found in the litera- ture. The generally reported Hill slope of l could result in some instances from averaging concentration-re- sponse curves from a number of cells. Finally, the present report is consistent with previous evidence that more than one receptor subtype may be expressed by a given neuron (Zimprich et al., 1991). This finding sug- gests that biphasic curves should be observed in model expression systems when the appropriate combinations of GABAA subunits are expressed. If such biphasic curves are not found, then expression systems may not be appropriate for modeling some aspects of the recep- tor inophore complex. The extent to which segregation of receptor subtypes occurs within a single neuron at the synaptic level is currently under investigation.
REFERENCES Boormann, J., Hamill, O.P., and Sakmann, B. (1987) Mechanism of
anion permeation through channels gated by glycine and gamma- aminobutyric acid in mouse cultured spinal neurones. J . Physiol. (Lond.), 385243-286.
Juiz, J.M., Helfert, R.H., Wenthold, R.J., deBlas, A.L., and Altschuler, R.A. (1989) Immunocytochemical localization of the GABA,?/ benzodiazepine receptor in the guinea pig cochlear nucleus: Evl- dence for receptor localization heterogeneity. Brain Res., 504:173- 179.
Langer, S.Z., Arbilla, S., Benavides, J . , and Scatton, B. (1990) Zolpi- dem and alpidem: Two imidazopyridines with selectivity for omega1 and omega3 receptor subtypes. In: GABA and Benzodiazepine Re- ceptor Subtypes. G. Biggio and E. Costa, eds. Raven Press, New York, pp. 61-72.
Mierlak, D. and Farb, D.H. (1988) Modulation of neurotransmitter receptor desensitization: Chlordiazepoxide stimulates fading of the GABA response. J. Neurosci., 8:814-820.
Olsen, R. and Tobin, A.J. (1990) Molecular biology of GABA, recep- tors. FASEB J., 4:146%1480.
Puia, G., Vicini, S., Seeburg, P., and Costa, E. (1991) Influence of recombinant gamma-aminobutyric acid-, receptor subunit composi- tion on the action of allosteric modulators of gamma-aminobutyric acid-gated C1- currents. Mol. Pharmacol., 39:691-696.
Sigel, E., Baur, R., Trube, G., Mohler, H., and Malherbe, P. (1990) The effect of subunit composition of rat brain GABA, receptors on chan- nel function. Neuron, 5:703-711.
Vicini, S. (1991) Pharmacologic significance of the structural heteroge- neity of the GABA, receptor-chloride ion channel complex. Neu- ropsychopharmacology, 4:9-15.
White, G. (1990) GABA, receptor-activated current in dorsal root gan- glion neurons freshly isolated from adult rats. J . Neurophysiol., 6 4 5 7 4 3 .
White, G., Lovinger, D.M., and Weight, F.F. (1990) Ethanol inhibits NMDA-activated current but does not alter GABA-activated current in an isolated adult mammalian neuron. Brain Res., 507:332-336.
White, G., (1992) Heterogeneity in EC,qand nH of GABA, receptors on dorsal root ganglion neurons freshly isolated from adult rats. Brain Res., in press.
Zimprich, F., Zezula, J., Sieghart, W., and Lassman, H. (1991) Immu- nohistochemical localization of the alphal, alpha2 and alpha3 sub- unit of the GABA, receptor in the rat brain. Neurosci. Lett., 127:125-128.


















