
Hawaii Cooperative Fishery Unit - Directed Research Report Title: …€¦ · Hawaii Cooperative...
56
Hawaii Cooperative Fishery Unit - Directed Research Report Title: The Food and Feeding Habits of the Kumu, Parupeneus porphyreus. By: Mrs. Cherrie A. Mahi Degree: Master of Science (Zoology), Plan B. University of Hawaii Pagination: Total - 54; 23 text, 31 Appendix including 3 Tables and 27 Figures. Abstract The kumu, Parupeneus porphyreus, is one of the most valuable reef fishes in Hawaii. It is a member of the goatfish family (Mullidae) and is endemic to Hawaii. One hundred and ninety-eight specimens, ranging in size from 31 to 306 mm, were collected from four areas around Oahu, Hawaii. Collections were made from July through November, 1968. The anatomy and the contents of the digestive systems were examined The digestive system is short. The stomach is V-shaped with a bulbi- form pylorus. There are 20 pyloric caeca. The instestine has one siphonal loop. Transforming specimens differ in having a shorter, straight intes- tine. They also have sharper pharangeal teeth and longer gill rakers than do the adults. Occurrence of food in the entirA gastrointestinal tract was used in ranking relative fullness. These rankings, compared with time of capture, indicated feeding was nocturnal or crepuscular among larger fish (only very young kumu fed during the day) and that food passed through the ali- mentary canal within 19 hours. Twenty-three food items were identified; 12 of them occurred in >10% of the guts. Crabs were the most important food, followed by other crustaceans (copepods, isopods, other decapods) and other invertebrates. Sand occurred in 57% of the specimens. An ex- tensive comparison of food items vs. fish size demonstrated various rela- tionships. Although crabs were eaten by nearly all fish, only large kumu ate Stenopus and other fish, and only small kumu ate copepods and isopods.
Transcript of Hawaii Cooperative Fishery Unit - Directed Research Report Title: …€¦ · Hawaii Cooperative...

Hawaii Cooperative Fishery Unit - Directed Research Report
Title: The Food and Feeding Habits of the Kumu, Parupeneus porphyreus.
By: Mrs. Cherrie A. Mahi
Degree: Master of Science (Zoology), Plan B. University of Hawaii
Pagination: Total - 54; 23 text, 31 Appendix including 3 Tables and 27 Figures.
Abstract
The kumu, Parupeneus porphyreus, is one of the most valuable reef fishes in Hawaii. It is a member of the goatfish family (Mullidae) and is endemic to Hawaii. One hundred and ninety-eight specimens, ranging in size from 31 to 306 mm, were collected from four areas around Oahu, Hawaii. Collections were made from July through November, 1968. The anatomy and the contents of the digestive systems were examined
The digestive system is short. The stomach is V-shaped with a bulbiform pylorus. There are 20 pyloric caeca. The instestine has one siphonal loop. Transforming specimens differ in having a shorter, straight intestine. They also have sharper pharangeal teeth and longer gill rakers than do the adults.
Occurrence of food in the entirA gastrointestinal tract was used in ranking relative fullness. These rankings, compared with time of capture, indicated feeding was nocturnal or crepuscular among larger fish (only very young kumu fed during the day) and that food passed through the alimentary canal within 19 hours. Twenty-three food items were identified; 12 of them occurred in >10% of the guts. Crabs were the most important food, followed by other crustaceans (copepods, isopods, other decapods) and other invertebrates. Sand occurred in 57% of the specimens. An extensive comparison of food items vs. fish size demonstrated various relationships. Although crabs were eaten by nearly all fish, only large kumu ate Stenopus and other fish, and only small kumu ate copepods and isopods.

THE FOOD AND FEEDING HABITS OF THE KUMU,
PARUPENEUS PORPHYREUS
DIRECTED RESEARCH REPORT
SUBMITTED TO THE GRADUATE SCHOOL OF THE
UNIVERSITY OF HAWAII IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
PLAN B
IN ZOOLOGY
FEBRUARY, 1969
BY
Cherrie Ann Mahi
Committee: John A. Maciolek, Chairman
John E. Randall
Philip Helfrich

rr 'l'HODUCTION
This paper present" a :;1, lJd,V of the stoIlk1.ch content s of kumu
(Parupeneus porphyreus ,Tenkins) from a number of localities, habitat~;,
and derths around Oahu, Hawai.i; as well as a description of the gross
anatomy of the digestive system.
The kwnu is a member of the family JI~ullidae (goatfishes, surrnullets
and red mullets), which .is widely distributed in tropical and subtropical
waters with some species ranging into temperate seas. Nullids are valu-
able food fishes wherever they occur. Because of this, some species J
have been extensively studie:-i, especially in the Hediterranean area. In I
Hawaii, goatfisbcs are captured by trap fishermen and by divers using
spears and surround nets. l ~l.l. rkets pay fishermen an average of $1.35 rJer
lb. for average size (1 - 2 lb) kumu, making this species one of the most
valuable reef fishes in Hawaii.
No author has directly investigated the ecology of goatfishes in
the central Pacific area, dp.spite tbe economic importance of the family.
The other t,{O genera of mullids in Hawaii, Upeneus and Mulloidichthys
have been thoroughly examined ecologically and anatomically in such areas
as Japan and the tied Sea, but the Indopacific genus Parupeneus has not
been studied anywhere. Fortunately broad ecological investigations,
such as that of Hiatt and Strasburg (1960) in the ¥~rshall Islands, have
included members of the genus. The kumu is endemic to Hawaii and thus
has been excluded.
The pu~pose of this study was to gain useful information about the
food and feeding habits of the killnu in order to illuminate some aspects

- ) -
of the hio1ogy of this extrelliely iMlortant reef fish. Hopefully, this
information will some dAy In,ld t,o :m improvement in the fishery.
~JETHODS
Jviyhusband and I (Henry and Cherrie Hethi) collected kUYl!1l f:rClJn four
General areas around Oahu: the North Shore, the Haianae coast, KcmeO'lE:
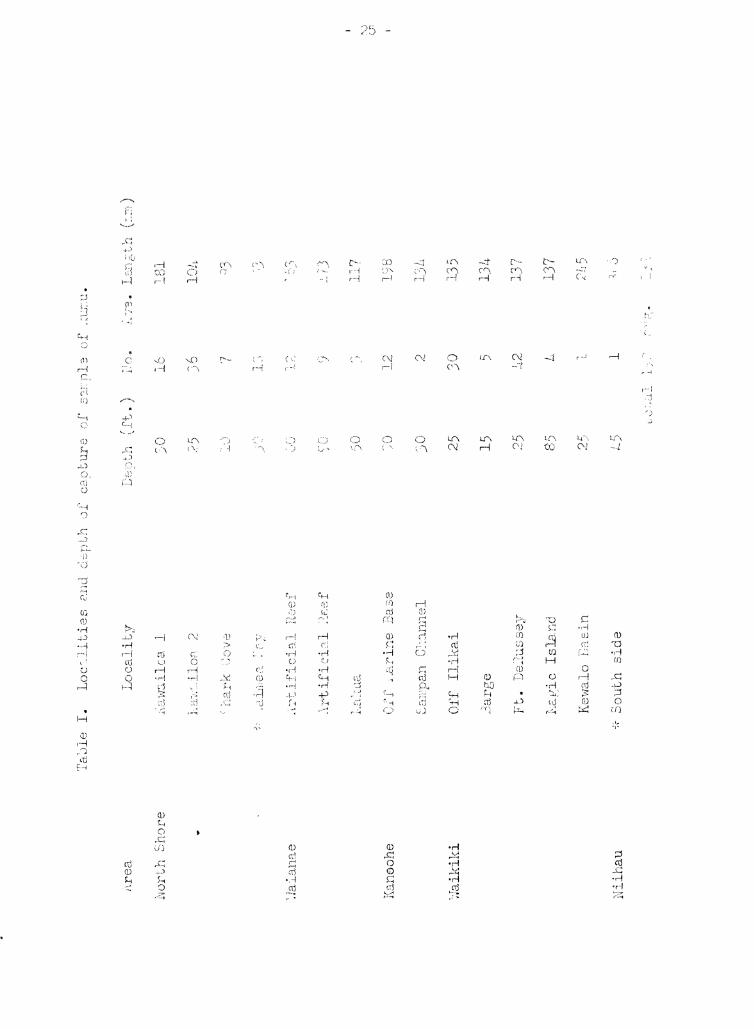
Bay, and Waikiki. Tahle 1 lists the collection localities and depths
of capture in these four areas along with the numbers and averaF~e si7.8s
of specimens from each locality. Depths of collection ranged from 1 to
95 feet, but the majority were about 30 feet. lie collected fish at tines
varying from 6:00 AN to 7:00 PN, but we concentrated on the morning hours.
Our collecti.ons were made between mid July and early November, 1968. In
addition, I borrowed 18 transforming or very small kumu from a collecti'ln
made in June by Mr. GrefE Stanton, and I received the largest specimen
(306 mm) from Hr. Richard Wass who speared it at Niihau in late November.
These borrowed fish are i.ndicated in Table 1. by asterisks.
We used Hawaiian three-prong hand spears to collect the specimens
in all but two cases. We used po~uered rotenone to collect 36 small
specimens at Kawailoa and Mr. Stanton used rotenone to capture the 18
juvenile ftsh that I borrowed.
Immediately after capturing the fish, we measured each one, removed
its digestive tract, and tagged each tract for identification purposes. ,
I preserved all of the digestive systems in 10% formalin with 15 cc of
lonol per gallon added as a color preservative. Three fish were preserved
intact.
I dissected these three average size fish (130 - 140 nun in standard

;,
lengt h) in order to COTllt1i) n; L he p;ross anatomy of the kumu I s dige st i ve
system with that of ~lull.21:d:i~hthys cLuriflamma as described by AI-Huss:d,r;j
(194b) and Upeneus bem:§:si as described by Suyehiro (1942). In addition,
I examined two transformjng newly benthic juvenile kumu (37-41 mrr..) for
comparison with the benthic adults. Henceforth, I shall refer to these
newly arrived kun;u as "transforminp;" and those which have already achie'ved
the appearance of miniature adults as "benthic."
I ranked the degree of fullness of each digestive system from 0 to 3.
If the entire alimentary canal contained food, the rank was 3. If the
stomach was empty but the intestine was full, the rank was 2. If food
remained only in the last fold of the intestine or the rectum, the rank
was 1. If completely empty, the digestive tract was ranked o. I com
pared these ranks with the time of capture to obtain a degree of fullness
for each collection time. I did this by multiplying the percentage of
fish having each rank at anyone time by the rank. I then added the rank
times percent values for each tj~e. Examples of these calculations are
shown in Table II. These total rank times percent values provided a
means for estimating the rate of digestion.
I examined stomach contents separately from intestinal contents so
that I could determine whether there was a reduction in the variety of
food organisms in posterior portions of the gut. I identified the food
organisms as clvsely as their condition pernutted. Useful references
were Edmondson (1962), Barnes (1963), and Hiatt (-unpublished lab manual).
Each food organism was ranked from 1 to 7 according to its relative
volumetric importance in each fish, with 7 being the most important.

I, -
RESULTS
The sample consisted of 19t) specimens ranging in standard length
from 31 to 306 mm. The median 1 eng-th was 130 mm. The mean was 132 mm
with a standard deviation of 53 mm. Figure 1 shows the distribution of
the lengths within the sample. The 198 specimens are divided into 28
size classes, each class havin8 an interval of 10 mm.
Feeding and Digestive Ana.tomy
The anatomy of the digestive system of Parupeneus porphyreus proved
to be very similar to that of Hulloidichthys auriflamma and Upeneus
bensasi. Like Mulloidichthys, there are teeth on the premaxillary and
dentary bones but none of the vomer or palatine bones. Upeneus differs
in having rudimentary vomerine and palatine teeth. The kumu's teeth
are small, conical and-blunt. They are widely spaced in a single row
in each jaw. The pharangeal teeth are fairly well developed. They are
small, pebble-like structures found in dorsal and ventral pads on the
branchial bones (Figure 3). Each pad is sectioned by tough ridges of
flesh.
The gill rRkers are 2/3 the length of the gill filaments (Figure 2).
They are stiff structures with slightly toothed inner edges. There are
28 in allo
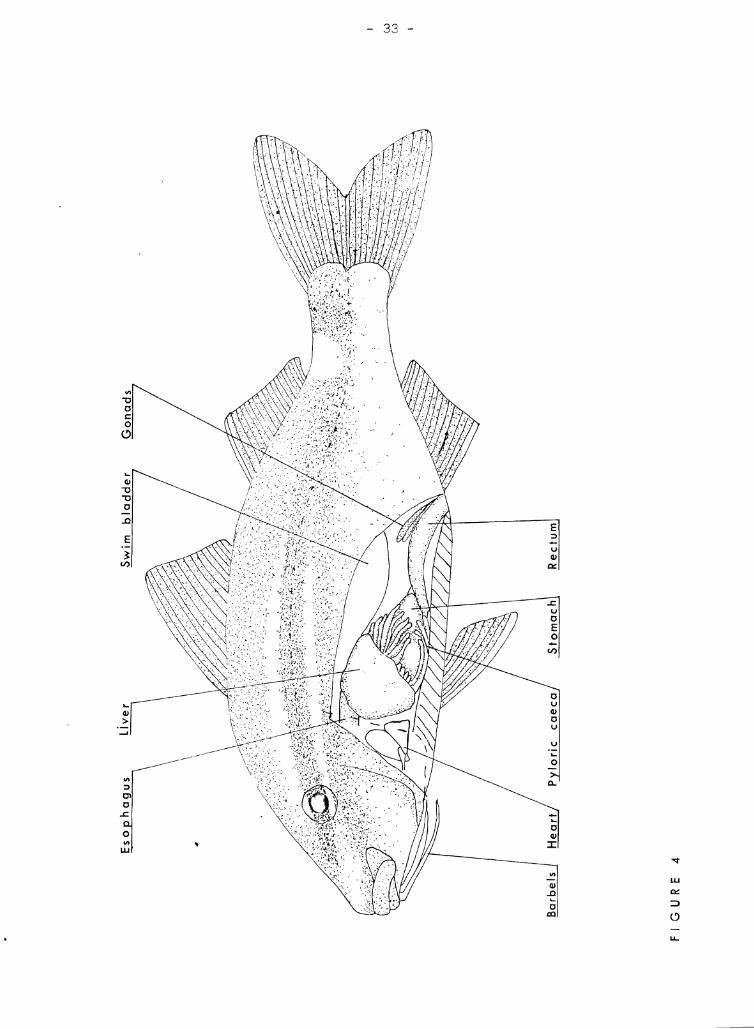
Figure 4 shows the intact digestive system of the kumu. The liver
is fairly large'on the left side, but the right lobe is very small.
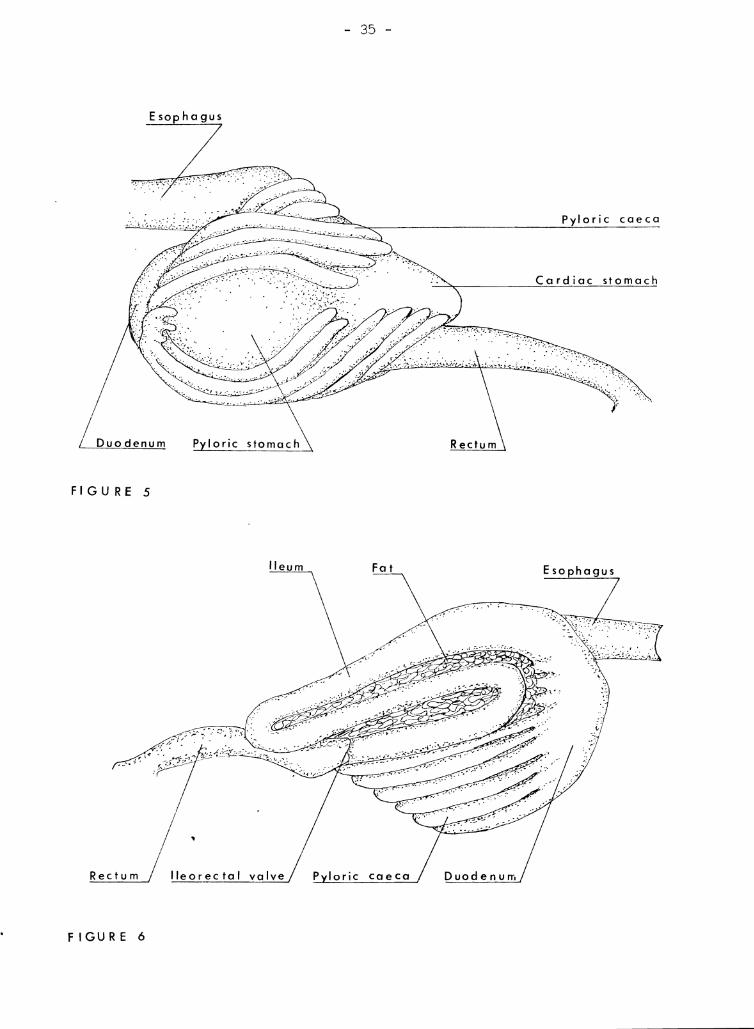
With the liver removed (Figure 5), the finger-like pyloric caeca can
be seen. In some specimens, the pyloric caeca are completely encased in
fat. This fatty tissue is rich in pancreatic cells according to

- 5 -
AI-Husnaini (1946). The l,;rlryri.c ea,f~ca number about 20 and arie>e from a
thickened rJOrtion of the ClU()(lrnU1Tl ,iust heyond the pYloric valve. Ttwy
lie such that in the right view (Figure 6) t.hey hide the stoID!lch. The:}
intestine has one siphonal loop. It arches dorsally across the esophap'u:'o,
runG posteriorl~r to the tip ()f the stomach, then turns sharply anterior
until it reaches the arch of the duodenum where it turns sharnly postcrj.or
again. Like the pyloric caeca, the intestine is frequently encased in
fat. About half--way behreen the duodenum and the tip of the stomach,
the last fold of the ileum enters the rectum. The rectum has a larger
diameter than the inte,3tine and its wall is thicker and darker in color.
In Figure 7, the duodenum hClS been cut and the intestine removed to
expose the large spleen, .."rhich fits into the "VII between the two Darts
of the stomach. After the intefltine, pyloric caeca, and all fat are
removed, the shanG of the stomach can be determined (Figure 8). The
esophagus is short and unmodified. The cardiac portion of the stomach
ends in a blind pouch posteriorly. In MulloiQ.~ this pouch is more
elongate. The f)yloric valve is clearly evident as a sharp constriction
in the bulbiform pylorus before it enters the duodenum.
The division b8tween the esophagus and the stomach can best be
determined by examining the internal structure of the gut (Figure 9) ..
According to AI-Hussaini (1946), the division is where the number of
mucosal ridges 1s SUddenly reduced. There are five mucosal ridges in
the cardiac Dortion of the stomach. These are most pronounced in empty
stomachs, but are always evident. There are only three in the pyloric
portion of the stomach. Between the ridges in the pylorus there is a

- h -
spon,r,:y network of more: lllinlJ~,'~ rLdr~c;:;. According to AI-Hussatni, this
thick that in empty fish, i.t almost obliterates the cavity. The wall
consi~;ts of three layers, Lhe innermost mucosa, the submucosa, and th,cO
mU:'lcularis. A-L H ." (, OJ 6') d t '1 d th .. t 1 "' th ,. - u,;~,a_lnl ,. /q.8 al e, e nlS o. og,y OJ. '. esc la~lers
for ,MulJ.-gidich!:b..Y§., which I assume is similar histologically to ParuP~r:t~21.:2.
The intestine has a few v-leak mucosal ridges in the anterior part of the
duodenum, but the rest of ttw intestine is lined with a spongy net1trork
similar to that of the pylorus. A section of this net1rmrk (small c;Quares)
is enlarged in Figure 9C. Neither I nor AI-Hussaini (1946) could deter-
mine the difference between the duodemlm and the ilelJm. The anterior
1>lall of the duodenum is only slightly thinner than that of the stofficwh,
but it gradually b(,)come,o much thinner until it is abnost membraneous in
the ileum, especially when it is full of food. It is ver:! easy to di~3-
tinguish the rectum hy its thicker vJall and different color. Internally
the muscular ileorectal valve is evident. The spongy network is dark
orange in color and more pronounced than in the intestine.
Transforming kumu which have just entered shallow 'v,rater (Figure 10)
look considerably different from the benthic form. At first glance,
they look more like a.nchovies than goatf'ish. However, the barbels
reveal their true identity as mullids. Even in the tiniest juveniles
I have seen (31 mm), a bar through the eye and a "Thite saddle behind
the soft dorsal fin are present, i1Thich serve to distinguish these as
well as adult kumu from all other Hr..waiian goatfishes.

- '/ .-
The Dharangeal Leeth pad~ [In) ,ohaperi like those of the adults, but
the teeth them"clvcs arc (111: ',': different. Instead of being pebble-]i t~~),
they 11flVe Sll<lrp, recurved points some,,'lt:at resembling cat claws.
The gill raken; (Figure 11) of the transforming fish are relatively
longer than tho~3e of the bent:li.c form. '1'h8;7 are slightly longer than the
gill filaments. At the lower angle of the opercular opening there is "
fringe on the gill cover that meets the poorly developed gill rakers of
that region. The teeth on the inner (xlgcs of the gill rakers a:c-c ';;or,-;
pronounced than in the bf:nth';'c form.
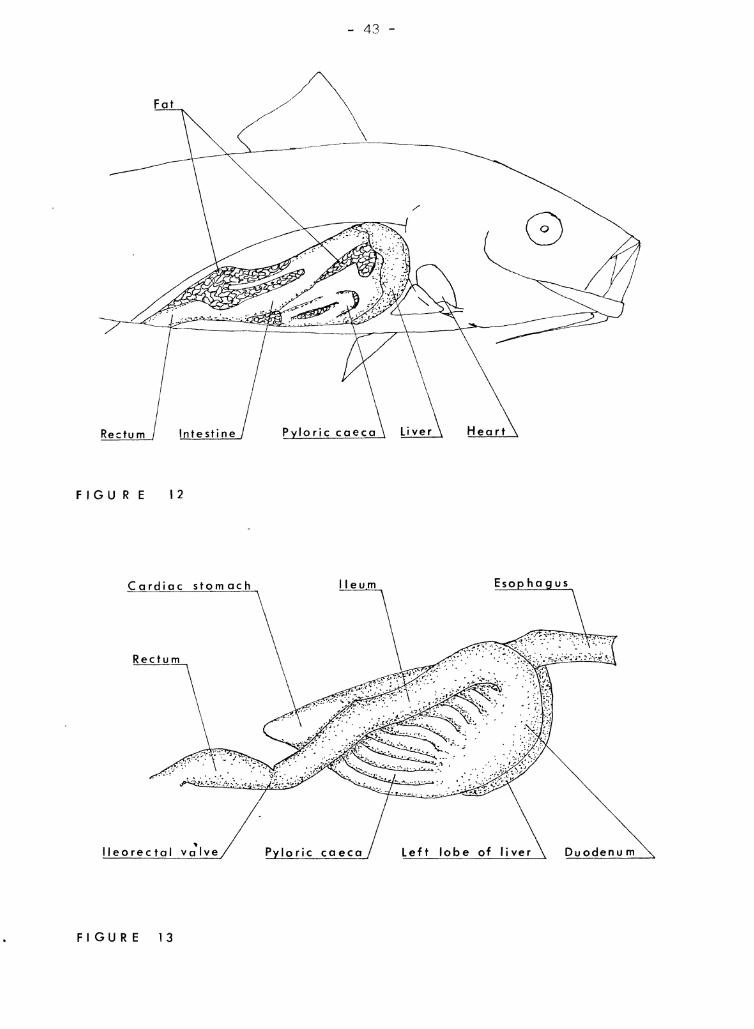
The heart and liver are relatively larr;er in transforming kwnu
(Figure 12). The digestive tr;'Ict is bound into a compact mass by Ir/;s(mterY.
V'men the liver, fat, and IJlet;enteries are removed from the digestive sY:Jtcm
(Figure 13), it resemtiles that of the adult "lith one very c3triking excep-
tion; the inte~3tine is short ,;nei straight \1ithout the siphonal loop of
the A,eiult intestine. Also the pyloric caeca are shorter so that in the
right view, the tip of the cardiac stomach protrudf:3s. In the left view
(Figure 14), the esophagus seems much longer than in benthic fish because
there is no indication of where it ends and the stomach begins. v-Jhen the
pyloric caeca on the left side are removed and the esophagus is lifted
slightly, the 2D1een is revealed. It is sm!'l.ller them that of the adult
and does not fit so tightly into the "V!! of the stomach. The cardiac
portion of the ~tomach is poorly defined and the pylorus is not so bulbous
as that of the bc:nthic form. Internally the digestive system is very
similar to that of the adult.

- ;j -
Food Habits -~-~-'---'-
Ahout one fourth (~:(l 01,;, of' l,)g) of the digestive tracts eX!l.min':,ri
were completely empty. Only those fish crlupht before 9:30 in the morniJI;'!
h:ld food. in their stomach~3. The only exceptions Here fish less than
80 mm long. Fish with ernnty stOTJ1nCDS hut full intestines were most COIlIDun
from 9:30 tL'1til noon. Fish with small ClIrlOunts of food remainin,c; in the
lA.st fold of the intestine and the rectum were f01md for only a short
time between 11:00 AN and 1:30 PJ:v1. The first completely empty fish
appeared at 9=30 AN and the percentage rose steadily until practically
all fish were emptJr by InO Ph. FifSure 16 Sh01'1[S hOrI the degree ot' f1Jl1-
ness (totA.l rank times psrcont) oecreA.ses rapidly "nth time. A point
at 10:00 AJ'vt is excluded because I bOrr01'led the fish from a collection
mA.de earlier in the year and because the fish "Jere very small. Also n
point at 1900 (7:00 PM) was excluded because the collection consisted of
one small specimen. Irregularities occur because small fish tend to
continue oA.ting throughout the day and large fish retain food in their
intestines longer than average size fish.
Twenty-three different kinds of organisms were identified in the
gut contents of the kumu (Tahle III). Crabs were by far the most import-
ant having an average rank of 5.3 out of a :maximum rank of 7.0. The
next most important organisms were isopods and alpheid shrimps each rank-
ing only 2.2. ~ollowing in decreasing order, foraminiferans, gammarids,
Stenopus hispid£§.. and stomatopods ,,,ere also important. The vast majority
of the organisms were crustaceans, but foraminiferans, fish and fish eggs,
mollusks, polychaetes, a tunicate, and even a caterpillar were found.

:)7~(, of the fish contaLncd C',:u. AJthough the quantity of th8 s:md
vClried, it Wrt s £'eDnra ,i':p Ii comn/'l.recJ with the rent of the conLerlt:.
The imnort;:mce or DVffiCr(lUS orRani~m1S varied with the size of th?
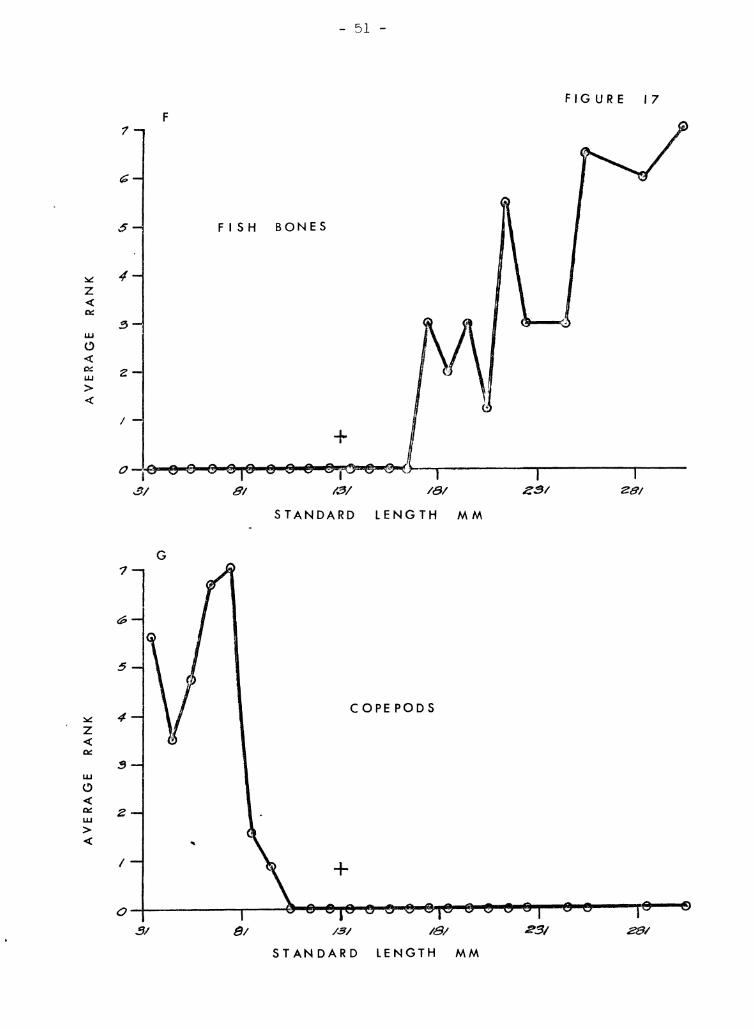
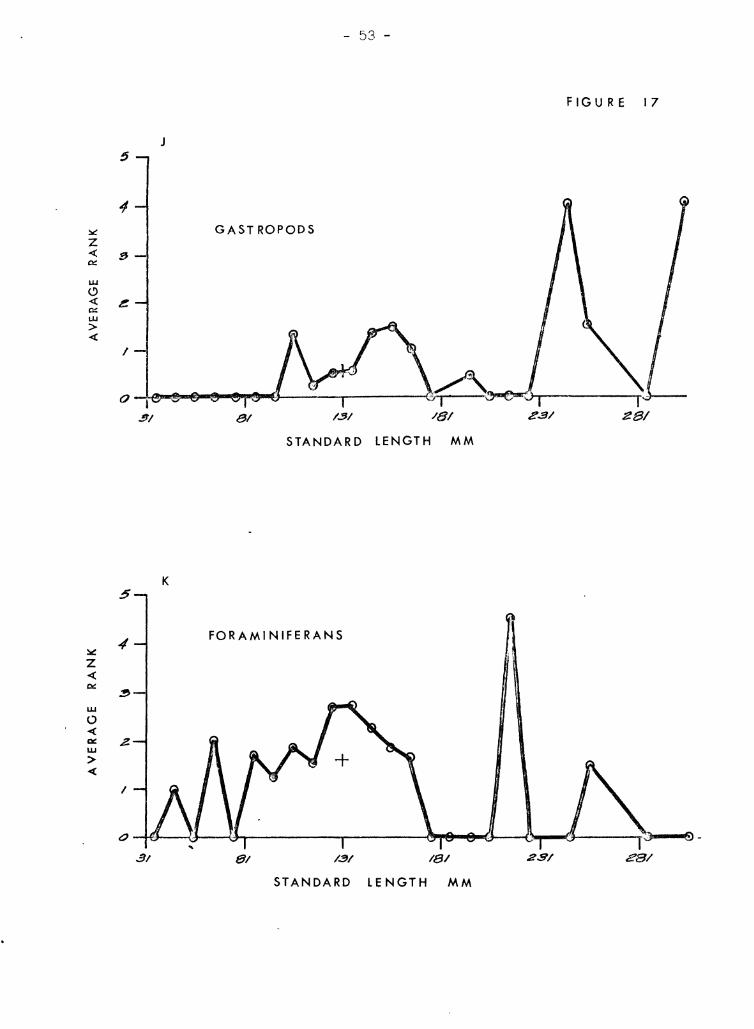
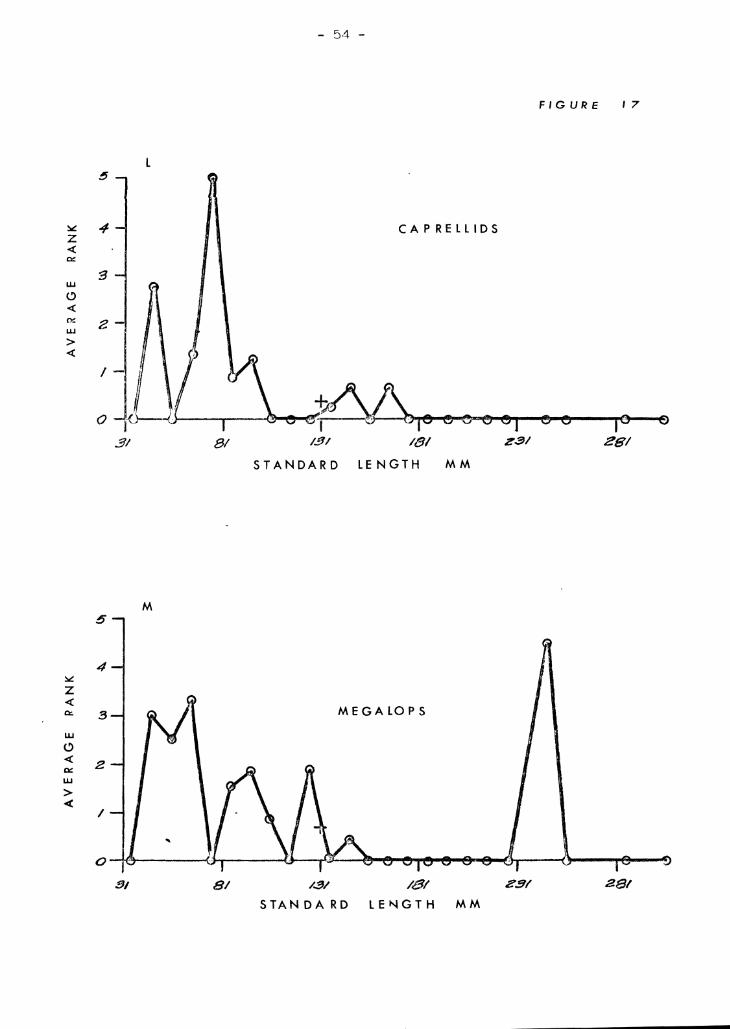
kumu. Figures 17 A-:t.l illustrate tf1GSe re1ationships. Crab ,3 arc venT
inmortant in all but the smalJ.8st kurnu (Figure 1713). The broad :::;i7.8
range help~l account for their overall high rank. 1sopods are import,:::tnt
in small to medium kumu J but, not in the very tiniest nor the larf',er
snecimens (Figure l7A). Alnheid shrimps (Figure 17E) resemble crahs
in that their importance is uniform over a broad size range excluding
very small fish. ~)tom[Jt.oC)ods (Figure l7C) and ostracods (Fir;ure 17]))
have irre{"ular diE;tribut-i0ns l-Ihich demIt seem to be closely related to
size of fish. Fish (Fl pure I'll") and copepods (Figure l7G) are ,just ;~h':
opposite sho,tTinf~ close relationships to size. Copepods ?re found only
in very SDl"'Lll specimen~3, c~nd fish bones are found only in very large
specimens with no overlan of :="ize ranv,8s. Gammarids (Figure 17H) and
StenQ12.1!.~ (Figure 171) are also size related but the differences bet"recn
them are not so extreme so thRt there is overlap in size ranges. Gammarids
are importcmt in small to mp.cliwn fish ,,[hereas SteQ.<?J2.1!:.§. is important in
medium to l,s.rr,e fish. Gastronods and foraminiferans are shown together
(Figurefl l?tY and 17l<:) because they both seem to be related to a third
factor -- S;;Yld. In only a fc, .... cases were they present when sand was
absent. :Host of the gastropods were even smaller than the sand Brains.
One notable exception is a ~~-inch Q:voraea fimbriata which was in the
stomach of a 1,,0 mm fish. Foraminiferans occured mostly in small numbers
except in small fish. Caprellids and megalops crab larvae (Figures I7L

In general, v:hen l(·:;~) Lh.'lll 60 mm in stand,grd lent,-th, kumu eat rnO:;T" '1
, . 1 n' 'I 0' ~ "nd c"f)yoplJ J' c' r From ()O tr) lIe) l'l'l.ril, ih.!'.:: copepous, gaJ!lTn'lr,lC S, ,ICC"" ,);0 c<. .G,.~, .'_ Lea "V .
')arne orfanism~; occur, Lmt .l!:nnocis are more important, and crabs iU'8
bcginninr; to aprenr more frcqurmt ly. From 110 to 180 mm, crabs are
dominant with stomatoDods, 'if;opods, and alpheid shrimps often includ8d.
In kumu greater than lCO mID, fish share domi.nance with crabs. StenonUc3
and alpheids also make Ir'A.;jor (~ontributions.
Food organisms exbibl,Led variations ,.rith locality in addi.tion to
size of fish. Crabs ranked r;reater than 6 in all locriJ_ities except
Km'Jailoa on the North shore v!here they ranlced only 5. One collrc~ction
of large fish from KaHaiJoa Nas dominated by ~1lQ..Tl~ and another
collection of sYJall fiEoh from K'liAmiloa 'VIas dominated by isopods. At
'daikiki, §.!:.2l~2n~us WPtS a ln~ost absent whereas stomatopods and foraminiferans
i"ere disproport.iona tel? imno:rtcmt. At Kaneohe, 9ten~nu~ and fish "Jere
more abundfmt than usual, but this may be because most of the l{aneohe
specimens c::onta:iniriP: food W0re larp;e $
DISCUSSION
The Samnle
Although nly sample was small (198 specimens), it did bear a resem-
blance to the popUlation of kUlnu at the time of collection. At the time ,
I did most of my collecting, the greatest numbers of fish in the en-
viromnent were betl"een 100 and 150 mIll in standard length. These are the
largest classes in Figure L If I had begun collecting earlier, I "JOuld

-- 11 -
have ~n ~he sJaller Gl~e ranc(", .
i vely sn:aller numbers oj' lin'l'er fi~~h. The 31 nm 3pec~L!L'::n DroL',
31 :md hO mm \.re1'8 tranSfOrITiini~ from the nelatzie type. On the othr;r nn,rJe;,
506 rom i~3 by no me;:m s Uv~ Inaximurn SJ_ze for kumu since they reach at
least 600 IllTii.
The sea son of collection is !lot the only factor that bill sed tho
samnle. Spe;lri~lg Ha~3 Lhe nrilJ:ary teclmique for capturing trlG "'1.sh and
this technique i~J inherently selecti ve. Slovier more sluggi sh fish are
most easily SDf)n.red. Perhan~)'.bese fish have fed best or are il~. A1';(\
larGer fish Rr8 [Cenerctlly morA wary, especia,ily in areas sueil (l sv)qikLL.i..
that are frecmer~t,ed ;;08ar Ci.f;hermen. This last fa ctor, as "mIl as
the gener:-ll scarcity of larr;e fish around Oahu, explains the rela ti vely
small size o~' the lal'!'e:t fi.. 8h in the sample.
It didn't t:l.ke me long to find that the later the fish Here caught,
the emptier t,he~r were; t hcrej'ol'e, I began doing most of my collec:,ing
in the ('-drl;! morrdng hours. A niGht collection would have been very
valuable, but we were unable to find the fish even in area s where they
were abundant duri ng the day.
Varley (1~6LI) ,ras -unable to make many generalizations concerning
the relationshin between the ane.tomy of a fi sh IS digestive system and
its diet. One I!eneralization that does apply to goatfishes is
"Carra vores have a shorter inte stine often with fingerlike pyloric

(AI-Husf:;airri., 1%.6 and 19h7)
C2t:';er: the total lcnrLh OJ "the clil~e,3tive system is acout tIle same J.~n h
in having he8 vy muco~'ial ridges :Ln the inte stine. AI-Hussaini (1946) cn)'1-
eluded that these ridr:es Emei t,h e hec1 vy mucous that they f3ecrete prate ct
the lininr; of the diJ.."estiv8 system from abrasion by the large amounts of
sand included in the diet,. According to Handall (personal eomrnun ~caU.()n),
throuGh the sand or silt sI1'J.llo1rring an7-
thinp edible ::mel extrll(l-inl" the exceSf3 [land through the [,;il1s. In th(~
proces5, considerAble 5D.nd if: swallowed. P~~en£~~S. is evidently ifJor8
selecl,ivo in its feeding beci'Lm,e it swaUows only small quantities of
sand. For thi5 reason, heavy filUCO sal ridGes in the intestine are not
ne ce s SClY'Y·"
ene puzzling :l~)pE;Ct of the anatomy of the klllilU concerns the len[';th
of the int,'3c,tineLn tranbformLng kumu cot:lpared 1.nth l)(mthic kwnu. The
intestine is nearly three time~3 lonf~;er in benthic specimen s than in
transforming juveniles. l1andall (1961) reports a similar lengthening of
the intestine in transforming manini (Acanthurus sandvicensi s), but he
relates the chaw:;e to a transition from a zooulankton to herbivorous
diet. Such a drastic alteration in basic diet does not occur in the ,
kumu. Perh2_ps the longer intestine is an adaptation for handlin[,; bulkier
foods such as heavily armored crabs.
An even more puzzling difference it:; in the sharply-pointed pharanp,eal

-- 13 -
teeth o~' thn lJelajtc ,i'.lveni.l.c. ]'.-:rhaps they help to prp.'V(':nt zoo:)lan1:tr;n
from svn.Iil' ir.g hAC I.;: out (,r th(' )I;()utll of the fish while it ::-;wims with it::
mouth aDen catch·L.nr: oUv:r pre;T. The pha:ranf~eal teeth of the l)entJhic r')y-r-,
e.ppcar to be full;7 cnp;'bln of crushinG nrey, but I could find no evirir::nC(7
that the fish actu~l,11y m,E: them for this. Fish caught soon (-tfter fcC)di.r'{~
had inta ct food oren,nisus in their stonachs, inclUding hard-shelled
gnstropoc1n. A questlon thet'f'fore exists as to the fllDction of the nh;',,'-
anf,cal teeth in the adult.
Fc.£dil]p:; H;:1:!2.~~
~:;V results strongJy indicate that only very small kumu fef'd dU~'iw'
the daylir,ht hO'Jr2, CIt least during the fall season. The obvious con
clusion from my res1l1ts is i,:nt mOf,t kUTnu feed at ni.e;ht. HmJever, sevnrM.l
comrnercial fishrcrn,en srd.d Lh'l.t kumu slee"D M.t night. These men liJake Lheir
Ii v irws rJY knolt/inv ",here kurnu ar8 at .'1.11 times and what they are (10in{~.
They move nff tl-e reef to s10cD either in sarga,ssum or on flat simd nc.xtJ
to, hut not under rock,;. ;)ince they don't, feed durinp the day and fish
ermen sac' the,v s1ef-)'O at nif'ht, about ttll that rem:tins is crepu~icular
feeding. Fl1l. J fot0TwtchsLn th e r,ornin C indic~i.te that they certainly feed
&t da"m, <'mel ful J intestines ,1,t dawn seem to indicn.te that they M.lso feed
at dusk. If they begin feedhlg at dusk (say 6:00 PH), digestion must
take a maxiE!1lIJ1 of 19 hours since they are completely empty by 1:00 P}I,
the next day (Figure 16).
I can only Sp8cUl;lte as to ",here kurnu feed. A number of the more
important foud or,f'anisIns such !'is snapping shrimps, stomatopods, fora
minifel'CJ.r1s, ('mel. g,3lTlIj]a,ricis are sano cii-rellers. 1,ilost of the gastropods

-Hi -
in holFJ r: in ro cks.
E':ince they occupy a wide vnri.ct,'" of habittlts rlccording to i~dmonc3~''ln (i.lC?),
and "ince c, 1;' conti' in cel dLffcr(~nt spe cies
Apparent
of the f'i~:;h cont;:liniDf' J'oor
f11rt hccr supnort s this conclu !~i()n.
Numerous authors hcve obE:erved goatfishes prob:inr~ the sand ':lith
their b.qrbels (Devanec)c{\ and Chidarnbaram, 19h9; Herre and }ljonr;:l11nn,
0. ) , .
derived from the first l)c;u;ci,iostef,;q.l ray (Caldwell, 1962). ~;d,o (1937,'1"
b) c3tudiec1 the histolov.v :'lnd function cf the barbels of a ,Jan;mese goat-
fish, Upeneu§ be]]..:?;:.?!. He compared t,he histology of these barbel::) vTith
tbose of nurrlcrCUfc (17,1'",1" fc~nds of fit3hes (~",to 1937c). Of all the diff-
aneous t;--,;cte Luel:::. Fe ckterE':Lned the function of e barbels in feeding
by elirdn2,ting various SFmsp~., find observing the behavior of the operated
a.nd unoperated fish. He concluded that although other senses contribute
to finding :Cood, the b,qrbels are most important, especially in findjng
and recognizing,buried food. He also determined that the barbels are
'to.. t t t" ul" +h . "',,"'ctl"J.e st;""lull" (Sato, 1°,,')'.,7 more senSl,l.ve lJJ rus .~a ory s lm l " an 1,0 ,a . ..J..,ln . _) ,
a, b). Unfortu.rn.tely the papers are rather old and the behavioral
experiment: ~n,re not controlled very ""Jel1. Upene~ ]2.~i char:lcteristi-
C'llly inhabits turbid waters \!lath mud bottoms and very poor visibility.

i.~, nf'i' ' Lvcl;T
f .ish.
The barbel s of the ku; 11 "'-',':. 1', -'
The food ornu1isn,::: in 1',1[e r;ut of each fish it.rere ranked accordin.f, to
OJl c;-!r'~
bodied on~anisrr: such :j S noJ yc~haete 3 nI'ol:H.bl;r don f t last ver;,' lorw il, the
digestive s;.';:.,tclT;. Alth()1](~h this almost certainly introduced some erJ:'C)l',
I donlt think the 81'1'01' is very f,re~:tt, because stomilch contellt,') were no
"lore W''''iAr\Il!'''.il intestirlal cnntentf';. If this factor Nere inlpo::~tnnt, I
should 8X1)CCt a re oid n~ducL~on in v[Tiet;,r from the stomach to the mlcl-
intestine. The bic's ,.,;as f.~re;.jte st for contents that were ranked on the
:~ inee I '\'Iet:; not able to determine where kmnu feed, it is difficult
to relate the food of kumu to the abundance of the food orl3anisms.
l~ccording to ~~dmondson (19tl?), Xanthid crClbs form the Jartjef,t group of
brachyurans in ilcp~Taii. The fact thClt they are also the most imnortanL
food item for tile kumu "Hould seem to indic(1te that kumu tend to eat most
of lvhat is most a.vailablc.
I was unnble to identify an~v of the fish eaten by large kumu. One
set of jnwC) might have belonged to a wrass, but I can't be certrtin.

There ",r:18 .'1bc;olutely no pC':'i_t.iv(, cor nerr!1.tive correlation betv:een the
say that the fiE;}] Here fOf':::cwlal. I wOlJld guess tl1at tiw;V 1trc~re either
fos~)or ial or
fi~3h thnt slcen tmrrieri in the sand.
Fornmin-iferans had a rather higb r!!_nk ~,E3 kumu food, but I doubt
their actual importance in T'lClst of the specimens. Only in VAr-:r sn ial1
[i"h I'fere they found i.n larf';e nmnb-'~r's and in only ti'JO caf3CS Ollt 0:" 1)1)
wert' they present when silnd v!i'if) ab"ent. The same 58ems to be true of
fimbrint,'1 fiL1J;(1 the: st.om~lch. I b r31inve t,r-nl,roram:1niferans and ~ --~-... - -~ --.~,~-
gastroc'O{1S ,""r'; E0'3tl;v incidental and ;lrc; ken alonr with sand durin?
the cant ure of larger orrsanipms.
There .·rere wmy vnrinLion;; frorn locality to locality and from
incH vidual to incU vidu,31. Po::-- exaJJ:D 10, one indi viduCll had eaten nracti-
call;v not>inv but chLtons, I dldn It find chitons in a single other
individu2.l. "hie; af~re(;f: y·lith the findings of v'lirszllbski (1953) for the
red IT.ul1et) 11J11_U'O ix!rbatlls.
The food haLi.ts of the kUInlJ f';enerally compare well "rith those of
other species of mullids studied by other authors. In practically all
cases, crustaceans were dominant, but in no case were crabs as important
as in the kumu., itllith this exception, central Pacific members of the
Strasbnrl';, 1 cencI Randall, 1955). Suyehiro (1942) found that the
Japanese I"Clatfi;ch l.!.I2en~E.f2. PJlli.;~_s..~ feeds on benthic shrimps, polychaetes

- L7 -
in kUlrm.
for l!iullus both in Israel 1tJ?tcrc: -,~~~"-.
irszubski, 19 Lj3) and in the; rJ(;diterrcuv.;!,'1.n, Adriatic and Bl:1C'k Seas
( T l' D"'ka· ,'" 1 ot../. ) IV' ..J...J _ .:> '_. I ( .... , ........ / '--J 'r <I ~ , .• l~lJf; ,"l180 Gat s considerable oU'l:ntitie S of n')ly-
to the lmmu (r;,'1.denat" 19 \ ) .
(Beebe and 'l'ee-VDn, 1 Ci2i; nnd Longley and Hildebrand, 19h1). Like the l<U1iiU,
1\. martiniclW :.~l iY1 ';~ he 8.fternoon ~ - "'~ .. _--_."---------Life Historv
• _",...-__ """ _"-' ____ M> • ..t .. ~
Little is known of the life hi~itor:T of the kumu. Gosline ami Brock
(1960) sta te thf~.t th e yourw fir13t fl,np.;ar in shalloH water in harch and
often found in t1ma "tor!';". ch,;. I>':y observat ions allow me to maJ:e a few
[';uarded C)t,"tcfiicnt;, about th", l:Lfe history. From July to Noveml)t~r when I
ly'aS collecting kmwl, I nC'lcr found any ripe fish. If the yOlli'll': first
apDear in shallow water in harch, the pelagic life must be less than four
months (l;:,tte l\ovember through February). Since I could not find newly
benthic ]-(UFlU Eift~er ,Tlmr~, the hreeding season must be about four months
long also. I TIi\lst qualify these statements because my observations ale
for less Ulan one year and arc limited. Yearly variations are pos~;ilJle.
Usually kumu live at all deaths from one foot to at least 200 feE:t ,but.
larger kumu gencY'nily live in deep8r "i'Tater. Commercial fi shermen say

;, ha hlJllw; h: r' hr: us har; a -",~ • . ---- -, >'- ~- .. ~.
b:,,3i:::-, o:f in1'orm'l tion from
otO.LLtllF, he decided 1,11nL tlie pc1ai;ic "t.p.ge extended from the :mrine:
E',pm·minp: t.ime until th c end of the :,,>c;me surnm.er. Cald1.vell (1962) d(~ C'-
cribes the pelar;ic ,Juvnni18 S of three; He~~tf~rn Atlantic spe ciGs, but,
doo~~ not [~pp.cuJate as to the l~n[·th of larval life. He believe~-3 that
JcetnmOr;JhOfoi.s from tl~(~ .,c type: occur:; after the . ." ,ll1Venl.L e s cmt3r
shaLlow watcT ;md not ('t, cl:~;"lc-:c'Lfic time; aft"r spavminr:.
I knm·r of only om, p()s~;:Lhl.e ob~)ervation of actual spawning in any
p:o,qtLLsh. hond:'l11 (r)8""on;1,1 cOImlUnication) believes he saw one Atlantic
sDecies spav-..rning in mid afternoon. Spa"ming was accomplished by ('to U'O-
\.·TCl.rr! rur:h vTith e.iection nfr:,'1metes1.t the apex of the rush.
Several points vV(lTr:'nt :"urther' stud;T. The time of feeding CAn be
:fixed morc! i1ccur;lLcl;v b~, Clh~)ct'vinF': 1':1.5h at dusk and. at various hour,; of
the nip;hl~. botL extrer,,'~s of Lhe siz.e sen.le need. further sarnplinr", All
of Tt,,:r f'i ~~h werebcntllic ltJhen captured. I w01.l1d like to exar;line gut con-
tents of night livht or traltrl suecimens. Also I would lilce to G3. pture
large (creat.er thnY' ?()O Lim) specirrl':ms early enoui~h in the Dlorning to find
intact fish in Lhr<.r c;tomachs. This tray I could determine the types of
stud;T "IRS not long enough to determine if there are any
seasonal e.:Tectf; on food habits. Suyehiro (1942) reports th3.t Y..Qeneu12,
bens'121 sc~rcnl.y eats during the spa-:ming season. Lipskay.q (1964) fou..'1d
seasonal variation in the relative importance of crustaceans,

Perhap:3 there :1.re :~ inri. 1 ;1 r
I,::.ri;l,tions "rith kllTl'U.
0[' the kumu :is very siJililAr t.!;
fi:'hco>, The p:reatc;c;t di·'i'erence is the presence of n
stonlil.ch i13 of the caee!'.l .?lX., "rith a r,izzard-like pylorus tlnci the inte"tine
hetS one siphoD[l.l loop. ,711'lC':ni1e kmnu differ from adul ts in havinl': lon[ccr
f5.11 ral';:crs J sharply 1. teeth, and a short, "tr::ight
inte st iw'C.
isoDods are also quite imon: i,ant. A ~Tide wl.riety of other crusta e'",3.n8
are cflten in srran.er \11u1.nt itif:f, aC'. l.'T'~ll il s other invert ebra t(; sand sma.ll
.(>' r ~ lS.l. Gmwrn,11;" , very BIl.a.Ii. fi fJh eil~, mOc)tl;'T minute crustaceans f3U ch as
C·' , ir;.-tus beCOJrle iYfnort.an t very ,~ar ly and. are domin.'u-:rt.
by the time +,110 "~i:nu reach IOO mm irl stanci:)rrl length. Isopods are inport.-·
ant in mnL' ;;wJ },:cd tum size fish. LT::'ger crustace:1ns such as alnheicis
,'md stomatoDods are relatively important in medium and large fish. StenoDUS -----
is eaten primarily by large specimens. Only large kurnu eat other fish.
There is consideraple individual and some local variations in food
habits.
At lea"t frml: {July through November, all but the slTallest kUJTlu ~;l.re
crepuscular 0 .. ' no::',sibly nocturnal f8eders.
Digestion probably takes less than 19 hours.


and histolof~Y of the [J,lib;n Lnr;r
trftct of the bottnm-·fe
! 1?1-1
hl-Hllss,:l"..niJ A. H. 1(jh7. The feedinc; h:l,hitc3 ;:,.nd the morpholov,v of ii
alimcn
Invr~rtel!l'a t e Zoolo!~:r, :'avnder s Co., PhiJadr;] obi:, ..
_L ? n.
Haiti, vrith n. Sl]mr;Vl.t"- of the knol·m sP"!cies of marine fish of the
islands of Haiti and (:lnto Domin,o:o. Zoologica. 10: 1-271.
Cadenat, (L '9Sh. l\cl:,"; (] I icht,;.rclor~ie ow~~,t-3.frieaine. VIr. BioloF;ic.
dlAfY'
Caldwell, 1'-1. C. 1962. Develonment and distribution of larval and
juvenile fishes of the family hullidae of the western North Atlantic.
U. S. Fi:::sh and Wildlife Servo, Fish. Bull. 62: 403-457.
Chacko, P. 1. 1'11.9. r.'ood and feeding habits of the fishes o:f' th-:; Gulf
of L::maar. Indian Acad. Sci., Proe. B. 29: 83-97,

:)~.~VZ1neSCll" D. W. (")11.(1 E. C1:jd'~::.LtLr·;:J.(I. 1_9~-~1. The COnL'rJlon food fir3hs~) l)f tIlE;
Goslin,;, {, • j:". liandbook of Hawaiian Fishes.
lTniv. Hawaii Pres:3, Hnw")lulu. 372 D.
Gottlieb, E. 1956. Tht~ n.p:e nnd rro1;rth of the red mullet, £:]1+.1.2:1;'2 ~?:~'Q::~::.'_".~:.
L. in Isr,qel 1.rI,ratc"t'u., 19:-~1-·1955. ;)ca Ii~ish Res. 5.-~tCtt. I;:;rfl~1" f~l.ll_l.
12. ;)Cl
Herre, As • ,~,nd I-I. {~oat fishes or l'~llllidae l'
the PhLLipnines. Phi lir);)ine ,J. eel. 36: S15-l36.
Hiatt, E. H. rtnd D. 'iT.
fish fauna on cora 1 r'c;ci'~3 of the )\~arshall Islands. Ecol. JVJonop:r.
Conmarison of t he food of pullet (1/111.11us Qarh:~.hu~
1',':;C\iterranean, Adriatic, and Black Seas (in Russian,.
English abstract). 1'1"'. SevastoDol skoi Biol. Stat. Akad. fJ;ouk.
UKRRfR. 17: 116-124. from Ref. Zh. Biol. 1965. No. 9II+2.
Longle~T, v'i. l1. hnd S. F. Hildebrand. 19h1. ~3ystematic catalogue of the,
fishee) e)f"' TortuWls, Florida, with observations on color, habits, and
loc:J1 distrirmtion. Carnegie Inst. Wash., Pub. 535. 34: 1-331.

R.'lnd'lJ, J. 1961. ft cor,trilmtion to the biology of the convict
J r: • - ,j •
Sato, J,~. 1,937:1.. ?rc;lirnin.'lry Y'en()~t on the b~1rbels of a J:1DaneSe I.-roat--
(Terilldnck C'!.[1'1 ~jchler:el). Tohoku lT~l~ \I.,
Sci. R~n.) ~~ries L. " L. 11:
Sato, }\;. 1937b. ?urLher si"ldiAs 0'1 the barb,;ls of a Jap;:mese goat-
fish, r:.' I; ('l'ernninck (inc! Sch18f~e1). Tohoku Uiriv.
Sci. Rep_, Snric;s L, Hiol. 11: ?G7-102.
Smith, t). 1.. I), 1 ~f. The S8!1 fishAs of Southern Africa. Central N~J\Vs
) ~ ~ c)()u,th ~\.f':Y'.ic;:\. x-"riii i- ~550 p.
Suyehiro, yr, 1942. 1\ rctUdy on the digestive system and the feeriing
habit~) rl;' ri~;r,. ,TrI.DClJW;if"'; J. 7.001. 10: 1-30].
Varley, I'" .. '-1. 196/+. Some aSD8cts of the nutrition of fish. AClvance.
::~G.L., Londcm. 20: L97-500.
·,rir~3z.ubski, /~. 1953. (In the biology and biotype of the re d mullet,

- >4 -
APPENDIX
Tables 1 through 3
and
Figures 1 through 17m
,G
ill! -, cn ~i l,\ s:-: 1--1
23 :'Y."\ / .. /",\, r- l{\ r'" d) t.:() -.
\, 0\ ('\ (Y', ~ 0-1 ,I r'l " : ,-1 r-1 ,--I r-l I
;:5
r' tj ., 4-1
(') C' ,0 n ~'.- C I, ,,,- (\1 C\J 0 tf\ N -ej r,··l '-1
.\,) ,--, I'i r---; I r-i c, I:::, ;,:i
, __ 1
iCj r~--,
::"-i
'-"
m w\ '~') ') 0 Cl C) L(\ L(\ 4\ l{\ '"
8 t': , ~ ) ~' ',(\ : '\ (\1 ,--j e'l tIJ 0l
+~)
_c) C, \1)
<J ,:-=l 0
'-I
,.f.':
)
r"
e. (',.! C"'_i OJ (j) 'I) in ,--j
en :',' },~, r;j :])
OJ r:'~ ('1 r' ~:>"' U c: '-=1 ·rl '-.. ~) ~:: .-1 ",C) ·-1 (\l ,1) , r-"i r-~ G) ['"1 ·rl ill ,;:l ())
',-1 .:-.~ :> Ci~ ( ~: f~ 0} 'el ,--1 r--i ro-I () ',4 ·r~ .-1 [) ;:1 (f) ·rl
- ~! 0 ~:::j (\i u , (.I : i ,-, r'--~ H ((),
U 0 ,--I 1 (.J .,-i or-f r ' r-I :1) 0 0 0 .,-j .~! ,'G 0 4--1 '+-1 ~'S ~J H oJ 0 ,--j x::
......:l ;--~ : 1 ;.. ~ ,~Cj
• .-j ·ri -) f~~ 1:<0 .C-! "J -~ I" ri"~ +.1 +, ~ eM H 'J-,~ ~ ;';, -, :~3 ,c_~ '~J .::j f ... q ,~j +-j cD -<-,) ,~ G) 0 . 1 ' -.." f·: ... 0J C n'l /., ..... r'':::'" ~ rn
H /;~
Q) ,---l .. '1 \U ~ -~
(1) ~j
('I ,.C U.·,: m <ll .r!
,-' .>:: Iv' ;::J ") "C ;~: 0 :;-~ ~ <ll .:._1 ") (J) ~~ --'-<
'-< ,
" -{ ~~ .C-! .,--l r-i ~) ~~ r.:~ G .r-! ::=- ~::-~
f-:- __ ~ ,-~-t

r-l
;< ..
,'\: ~-·c
~~
Gl ,
)'
d, ~ ~'l~ i
(.--~! "> ~:~ C) C') (,':.~
v. ~-'-I
,1) , . f~ 0 C:j
.4 () :; . l ... : (V'\ '" r---1 ,.,
,'t"1
0 C' '\ ,("-~
\U (1
',' i C) ~--) .~; I :) <t 0
.;i; ,:j -::J ,::.~ i-··j
i ':.-:
(t) r=l ... <.-': <"-.:
.)
r-~ ~ Q .J ,~) ()
,-i I~-i
_t.) :""1 ;:,.;-.
rJ Q) ;-:; ?<
0 'f~ (i) I --+' ..;..J ~-: ,~<; r-i ,:) -r-1 (~ 1
i<...J r-l ,0 0 en ,.)., ~ -1--') I C~ r:. :; ,0 , I " (\) I ",I n}
-!-) I (0 !-) !-"', !
r--1 ;.1 1'- C) 0 (r c~ c) l(,\ U, C)
C-1 LJ
c:i Cl ()
L! i;'.., ~.~) r-j () ~~.-;
<fJ ~-> (.) ·,-1
, rj ~J (",
C ... r"i '-.J f'-i r' 0
r~ ,-I
f,:'; 0
u ~ u-'.
0 H r,~ ~/?
, !---( ":~I 0 0
OJ r-i r O ('3
r::'-l
m u\ 0 0 ~::; -7' (:J 0
""'1 to (\I '0 0 rei r"'1

- )7 -
()r,~:'ni;:;m of " 1. c::; ~- l.ri t,1) orf~t~Lr) i~3r.1 r)vera~C! r[jnk
Cr·~lb~) i .1 I; .1
Isopocl.s '~(I.Cj 2./.
Alnb,id ,'llrimps ! ') f .. ' " /.) ;::, /
ForC'tf:lin i fc~rF1 n s ~rl.l 1.b
Garrm:Pl rid~; :21 .7 1. ')
;~t, er_0...p_tU? Ll i ['P;~9.:11,:l 1..8.2 LD
StoUlClt,opods ;~l .0 1.0
Copepods 14.2 0.8
kep,nlops .til .? O.?
F'ish bones LL5 O. f'1
!
Gastropods 1.7.6 0.6
Ostracods 12.8 0.6
CClprr;llids ? .fl O.h
Fish i:'Wf' f.; 6.1 (9) 0.]
Po1ychaetes 4.1 (6) O.?
Kona crabs 3·14- (5) 0.2
!v~~,rsid s LL. (2) 0.1
Chitons O.? (1) 0.1
Spider crabs 1.4 (2) 0.04
OctODUS 0.7 (1) 0.04
I'unicate 0.7 (1) 0.03
C"ltenil1ar 0.7 (1) 0.03
CUJI1rl.cean 0.7 (1) 0.02
(nurr~bers i.n narcnthp,sis are number~~ of fish containing orgClnisms.)

- 23 -
:[i'i~ure 1
Distribution of 3amn l e of F)g fish ,J,ccordinp: to st,-,nd"",d lenr~th. ~;}1;"tciccl
areas represent lllJI(l.bcr~; of fifh "lith food in stomachs.

1
I 20-1
I I
::c U) I ;:;:= 15_1
u.. o CY w ru :'£ ::::> Z
10-
- 29 -
FIGURE I
! I I
I
\ \
'A.~1---L~\v .I--.-~ J I I j
/.51 /:91 2/1 /:..31 25/ 271 2.91 3//
STANDARD LENGTH MM

- ]0 -
FiJ'ure ?
First n;ill arch frOTi" the 18i't ~i(k of a 140 rrrrn kUIml. Gill rakc;rs DlJr:lber
~,. 1 J<lgure j
Pharanr:e[ll teeth of lL~O mm hmlU. Teeth are pebble-Like. Patches of
teeth are sepnr:t2c1 h:r ridges of tough flesh. ::;p'l.ce b,,,tvJerm un;lcr Dadfl
is concave J and S-:!Clce oet"reen lower pads is convex.

- 21 -
Gill f iJam~nts
Gill arch
FI G U R E 2 '
Upper pharangeal teeth
lower pharangeal teeth
4 t b ---ttill Q r c h
FIG UR E 3

- 32 -
Figure 4
Average size kumu (140 mm) with left side of peritoneal and cardiac
cavities opened to eXDose the gross internal anatom~T. Gonads are in
inacti ve state. The d:1shed line shmvs the position of the operculum.
The body color is generall~r oink, but some small specimens are gray
green and ~)orw::; very large adults are purplish.
33 -
41 "'C "'C 0
..0 E
E ;:)
v GI ~
CI) c.:
:' .:.,~ ..
.c. v 0 E 0
CI)
0 ... V 41 GI > C
V
V ... 0
'" ;:)
Cl 0
.c.
0
'" ~ W
......
'" 41
W
..0 c.: ::> ...
0 <Xl t>
u..

- 94 -
Figure 5
Digesti ve system of a 140 mm kumu vie"l'Ted from the left after removal
from fish. The liver has been removed.
Figure 6
A right view of the digestive system of a 140 nun kumu. The liver has
been removed. This particular fish has relatively small quantities
of fat surrounding the intestine and pyloric caeca.
- 35 -
Pyloric caeca
Cardiac stom~cb
/ I Duodenum Rectum \
FIGURE 5
Ileum Fat
... ~ : . . E'~+7 .:.: ...... ::".! ',' ~-,;\1
Rectum valve P loric caeca Duodenum
FIGURE 6

- 36 -
Figure 7
A right view of the digestive system ,,,,,ith the intestine removed. The
darkened pyloric caeca below the spleen are in a notch which is usually
occunied by the intestine.
Figure 8
A right view of the diw'stivc system with the liver, intestine, Ilyloric
caeca, and spleen removed. The pyloric valve is the constriction between
the pylorus and the duodemun.

Eso ha gu s
.~~:::l;->;:±fi!!f'j
l~d' caeca Fat Duod e n u m
FIGURE 7
Cardiac ~ ___ ~mach E so
p loric sto _--=-" .. moc h Duodenum
FIGURE 8

- 38 -
Figure 9
Internal view of the digestive system showing linings.
A. The stomach of a 11!-0 mm specimen cut open from the left side and
the sides folded back.
H. The intestine and rectum of the SDme snecimen cut open 10ngitudiDcLlly
from the right and the siLles folded back.
C. An enlareed seetLon of the spongy network lining the pylorus and
the inte,,:tine. Small souares in A and !3 represent th c approximate
relative size of the section.

- 39 -
A B
P lorie valve
L~oreelal
c
FIGURE 9

- 40 -
Figure 10
A transforming kumu. Apparently it had just entered shallow water and
become benthic at the time of' capture. It measured 37 mm in standard
length. The pattern is made up of lare;e, widely-spaced melanophores.
Figure 11
The right opercular opening of a 37 nun transforming klll11U with the gill
cover lifted un and folded foreward to expose the gill arches. Gill
rakers are elongate on the first arch but rUdimentary on the other
arches.

- 41 -
FIGURE 10
o erculum
FIGURE II

- 42 -
Figure 12
A 37 rmn transforn'ing kurnu with the right side of the peritoneal and
cardiac cfl .. vities opened to eXDOse the gross intern'3.1 anatomy. The
digestive system is tightly bound by Yflesenteries.
Figure 13
A right view of the digestive system of a tran sforming kumu after removal
from the fish. The right lobe of the liver and the fat and mesenteries
surrounding the digestive system have been removed. Note the short,
straight intestine.
- 43 -
I Intestine I P loric caeca Hea rt
FIGURE 12
~ardia~ stomach
Ileorectal Left lobe of Ii ve\ Quodenu m
FIGURE 13

- 44 -
Figure lL~
A left view of the digestive system of a transforrrdng kumu. The
dashed line shows the Dositioc of the large left. lobe of the liver,
which has been removed.
Figure 15
A left view of the digestive system of a transforming kumu '<lith the
liver and pyloric caecc:. of the left side removed and the esophagus
lifted sli[htl;\\ to expose the spleen. Note the poor development of
the cardiac stomach.

Eso
puodenum
/ /
FIGURE 14
Esophagus
Duodenum
FIGURE 15
- 45 -
Position of left lobe of liver Cardiac stomach
Rectum
P loric c a ec a
/ Cordjac sto mac h
.I
~ectum
Pyloric stomach

- 46 -
Figure 16
The relationship bet1:J"een time of capture and fullness of the dig.;stive
system in kumu. The derree of fullness is the total rank times percent
value for eel eh time (see text ,md Tclble II for explan.1.tion).

- 47 -
FIGURE 16
£4-
2Z0
zOO
180
V)
V)
w !60 z ....... ......I
~ u...
140 LL
0
w IZ() w c.:
<-' UJ
0 I«J
go
~o
4~
20
CJ
5
TI ME OF DAY

- 48 -
Fieures 17 A trLrough H
The relationship between size of kumu and average ranks of the 13 most
important organisms in the diet of kumu. The 8m'l11 cross above 131 mm
on each graph represents the mean rank for the entire sample .
..

- 49 -
5 A
FIGuRE 17
-1 ~
Z 4: !) ISOPODS C"<:
w C> « z .... 0:: W
> « 1-
0-1 ..!II 81 I!!'I 181 C31 ~e/
STANDARD LENGTH MM
B
;5 ~
z « 0::
1-w <.') «
~ 0:: W
> eRA B S «
2
.. /
0
9/ 81 I.!J/ 181
STANDARD LENGTH MM

- 50 -
FIGURE 17 C
~
loc::
Z ..x: Z STOMATOPODS O<!
w () I ..x: 0::
• .1J
> ..x: 0
.31 81 /..91 /13/ e.!J1 E81
STANDARD LENGTH MM
D 3
~ OSTRACODS z « z 0:::
W
() ..x: 1-0:::
W
> ..x:
0 I .31 81 1.:31 181 ~I ZBI
STANDARD LEN GT H MM
E 5
ALPHEIDS
--I loc:: Z ..x: 0::: 3
w () ..x: Z 0::: W
> .. « 1
0
3/ !J/ 13/ .181 2~1 Z!81
STANDARD LENGTH MM
- 51 -
FIGURE 17
F 1
6 F ISH BON ES
~ '" Z « c.:
.;3 w 0 « 0<: Z W
> «
1
+ 0 I IV
..51 81 131 /8/ eSI Z81
STANDARD LENGTH MM
G
(OPEPODS :..:: Z « a.::
.9 w 0 « a.:: :z w > « ..
/
0
.5/ a/ /3/ /8/ 281
STANDARD LENGTH MM

H 1
~ !-
z
) <{ 0::
3 w 0 <{ 0:: t! w > <{
/
0-1 31
1
~
Z <{ 0::
.3 w 0 <{ 0::
2 w > <{
I
0
~I
- 52 -
/3/
STANDARD
STENOPUS
+
tJl I~/
STANDARD
GAMMARIDS
181
LENGTH
181
LENGTH
MM
MM
FIGURE 17
231 281
~/
~
Z 4: ct:
w 0 4: ~
UJ > 4:
w o 4:
!J
I!!
I
0
5
CI: ~ W
> 4:
1
J
I .!II
K
.31
- 53 -
FIGURE 17
GAST ROPODS
~ C:1 I
81 1.51 /81 E3/
STANDARD LENGTH MM
FORAMINIFERANS
81 /3/ 2.5'/ e8/
STANDARD LENGTH MM
w
" < IY
W
> «
!><:
Z -4: C<.
w
" -4: a:: w > <
5
4
.3
Z
I
0
9/
- 54 -
FIGURE 17
l
CAPRElllDS
STANDARD LENGTH MM
M
MEGAlOPS
81 /03/ /8/ Z,5/ 281
STANDARD LENGTH MM


















