
Gold Nanoparticle Interactions and Impact Upon a Common ...
44
Clemson University TigerPrints All eses eses 12-2009 Gold Nanoparticle Interactions and Impact Upon a Common Biofilm Source: Legionella pneumophila Amber Stojak Clemson University, [email protected] Follow this and additional works at: hps://tigerprints.clemson.edu/all_theses Part of the Biology Commons is esis is brought to you for free and open access by the eses at TigerPrints. It has been accepted for inclusion in All eses by an authorized administrator of TigerPrints. For more information, please contact [email protected]. Recommended Citation Stojak, Amber, "Gold Nanoparticle Interactions and Impact Upon a Common Biofilm Source: Legionella pneumophila " (2009). All eses. 679. hps://tigerprints.clemson.edu/all_theses/679
Transcript of Gold Nanoparticle Interactions and Impact Upon a Common ...

Clemson UniversityTigerPrints
All Theses Theses
12-2009
Gold Nanoparticle Interactions and Impact Upon aCommon Biofilm Source: Legionella pneumophilaAmber StojakClemson University, [email protected]
Follow this and additional works at: https://tigerprints.clemson.edu/all_theses
Part of the Biology Commons
This Thesis is brought to you for free and open access by the Theses at TigerPrints. It has been accepted for inclusion in All Theses by an authorizedadministrator of TigerPrints. For more information, please contact [email protected].
Recommended CitationStojak, Amber, "Gold Nanoparticle Interactions and Impact Upon a Common Biofilm Source: Legionella pneumophila " (2009). AllTheses. 679.https://tigerprints.clemson.edu/all_theses/679

GOLD NANOPARTICLE INTERACTIONS AND IMPACT UPON A COMMON BIOFILM SOURCE: LEGIONELLA PNEUMOPHILA
A Thesis Presented to
the Graduate School of Clemson University
In Partial Fulfillment of the Requirements for the Degree
Master of Sciences Environmental Toxicology
by Amber Reve Stojak
December 2009
Accepted by: Dr. Stephen J. Klaine, Committee Chair
Dr. Tamara L. McNealy Dr. Lisa Bain

ii
ABSTRACT
There exists widespread concern of pathogenic bacteria colonizing and establishing
biofilms in water systems providing a direct exposure route to the public. Legionella are
widely distributed in human-made and natural environments, and colonization lends the
potential to infect human hosts that may lead to the development of Legionnaire’s
Disease. Silver has been used in cooling towers and water filtration systems for removal
of planktonic bacteria and biofilms; however permanent removal of L. pneumophila
biofilms with metal ions is impossible when established biofilms are in hard to access
places such as dead end piping or inside a protozoan host. Bulk metals showed limited
success in the past at the permanent removal of Legionella, but nanometals, unlike their
bulk elemental or molecular form, behave based on their size, shape and surface
chemistries. Some nanometals may provide, perhaps, an interesting avenue for novel
anti-microbial agents. This research investigated interactions between citrate-capped
gold nanoparticles (AuNPs) and L. pneumophila biofilms. Planktonic virulence, growth
kinetics and toxicity were assessed, while biofilms were evaluated for structure, density,
and composition. Transmission electron microscopy (TEM) micrographs indicate that
AuNPs associate with cellular surfaces, the internal cellular compartment and integrate
into the extrapolymeric matrix. Previous research has shown that Au and Ag NPs induce
an antibacterial response in some planktonic bacteria. In contrast, there were no
significant differences of planktonic viability of L. pneumophila treated with 18 nm
AuNPs. However, planktonic assays with 4 nm AuNPs at a concentration of 100 µg/L
indicated a statistically significant decrease in biomass, measured as the optic density

iii
(OD) at 600 nm after 30 hrs of growth. Pigment production, a known virulence factor,
measured after 48 hrs of growth at OD400 for the 4nm AuNPs treatments was also
significantly less than all other treatments. Evaluation of the biofilm using Syto 11 dye
and confocal microscopy indicate an alteration of morphology seen on glass slides. In
treated biofilms, L. pneumophila formed long chain-like assemblages in contrast to a
thick and densely packed biofilm as seen in controls indicating a possible stress response
to the presence of AuNPs. No change in morphology was seen between controls and
polystyrene NP treatments. AuNPs, although not effective in completely inhibiting L.
pneumophila growth and pigment production, exhibit a negative mechanical response
under biofilm forming conditions in response to 18 and 4 nm AuNPs. The results from
this research suggest the potential uses of NPs in destabilization of biofilms. Further
research will attempt to delineate whether the effects from surface chemistries, sizes, or
shapes of the AuNPs are merely mechanical or antimicrobial with L. pneumophila.

iv
ACKNOWLEDGMENTS
I would like to thank my advisor, Dr. Stephen Klaine for teaching me how to
design experiments, think like a toxicologist, and how to become a well rounded scientist.
Also, thanks to my committee members, Dr. Lisa Bain and Dr. Tamara McNealy for their
help and support. I would like to acknowledge the McNealy lab for all their help on
microbiology assays. Dr. Tamara McNealy provided help and effort training me in her
microbiology lab. Dr. McNealy’s Creative Inquiry class was much help; specifically
gratitude goes to Luke Bury and Marie Capelle for their help on methods development
and planktonic assays. Also, all transmission electron and scanning electron images were
taken at Clemson University Electron Microscopy facility and guidance was given from
Dr. Joan Hudson, Dayton Cash, Donald Mulwee and my labmate, Aaron Edgington.
Confocal images were taken at the Clemson Imaging Facility and Nikon imaging facility,
Jordan Hall. Also, acknowledgement goes to my labmates for help with water quality
measurements and test preparations including Sarah Robinson, Brandon Seda, Jeffrey
Gallagher and Aaron Edgington. I would like to acknowledge Clemson University for
funding and support. Also, I could not have completed this work without the
encouragement, motivation, love, and support from my mom, family, friends, and my
very patient boyfriend.

v
TABLE OF CONTENTS
Page
TITLE PAGE............................................................................................................... i
ABSTRACT................................................................................................................ ii
ACKNOWLEDGEMENTS........................................................................................ iv
LIST OF FIGURES.................................................................................................... vi
LIST OF TABLES .................................................................................................... vii
CHAPTER
1. LITERATURE REVIEW............................................................................... 1 Gold nanoparticles: an ancient noble metal with novel uses ........................... 1 Gold nanoparticle interactions with bacteria................................................... 3 The possibility of trophic transfer................................................................... 6 Environmental risk assessment for gold nanoparticles....................................9 References ....................................................................................................11 2. MANUSCRIPT ...........................................................................................12 Discussion and Conclusion ..........................................................................26 References ...................................................................................................31 3. SUPPLEMENTAL MATERIAL ..................................................................33

vi
LIST OF FIGURES
Figure Page (Literature Review) 1. The exponential increase in articles published
on gold and silver nanoparticles.......................................................... 2 2. Transmission electron microscopy micrograph of
E. coli- AuNP exposure...................................................................... 5
3. Transmission electron microscopy of C. fluminea-AuNP interaction ........................................................................................... 8
(Manuscript) 1. Confocal microscopy shows induction of morphology changes
after exposure to AuNPs......................................................................18
2. Bar graph representing both overall biomass and pigment production at all time points ...............................................................20 3. Bar graph representing preliminary results from 4nm AuNP viability assay ..................................................................22 4. Transmission electron micrographs of control, AuNP, and polystyrene exposed L. pneumophila ....................................................................25 (Supplemental Material) 1. Transmission electron micrographs of 18 and 4nm AuNPs and 20nm polystyrene beads before use in experiments ......................................34 2. Transmission electron micrograph of 4nm AuNPs before and after aggregation ....................................................................35 3. Light microscopy image indicating sufficient L. pneumophila biofilm growth after 5 days …………………………………………...36

vii
LIST OF TABLES
Table Page
1. Elemental dispersive x-ray spectroscope % weight reports for Gold in TEM samples ...........................................................................26

1
LITERATURE REVIEW:
Gold nanoparticles: an ancient noble metal with novel uses
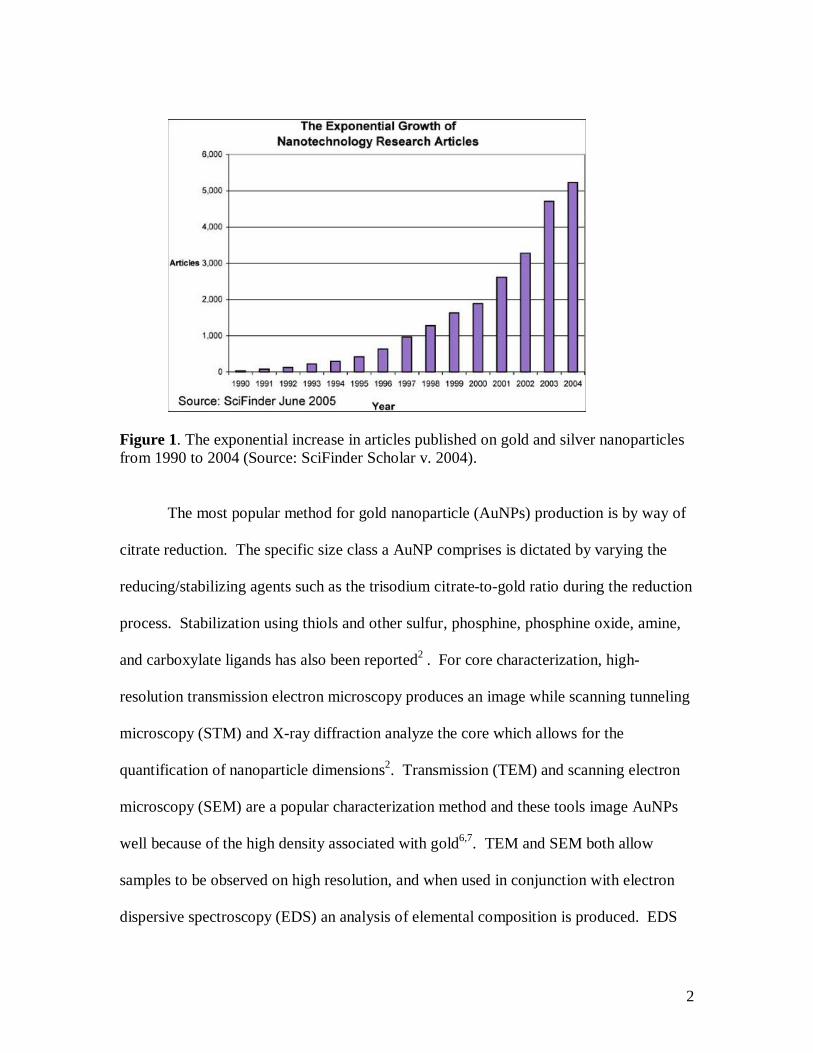
The interest in noble metals, including gold, as nanoparticles has exponentially
increased over the years as reflected in the sheer number of publications on gold and
silver nanoparticles1 (Figure 1). The idea of gold colloids is quite ancient, as it dates
back to the 5th – 4th century B.C. where gold colloids created a ruby red coloration in
glass. The term “colloid” was first used by Graham in 1861, however the use and the
production of nanomaterials boomed in the 20th century. For gold, this is due to its
unique electronic and optical properties2. Gold is used now for inks, films, catalysts,
dyes, drug delivery, and imaging2,3,4,5. Gold nanoparticles have been found to act unlike
their bulk or elemental form. The size, shape, shell or surface chemistry all contributes to
the behavior of a gold nanoparticle. Size alone may control the quantity of nanoparticles
permitted to interact with the organism or material as there is a higher surface area to
volume ratio associated with smaller particles6.
2
Figure 1. The exponential increase in articles published on gold and silver nanoparticles from 1990 to 2004 (Source: SciFinder Scholar v. 2004).
The most popular method for gold nanoparticle (AuNPs) production is by way of
citrate reduction. The specific size class a AuNP comprises is dictated by varying the
reducing/stabilizing agents such as the trisodium citrate-to-gold ratio during the reduction
process. Stabilization using thiols and other sulfur, phosphine, phosphine oxide, amine,
and carboxylate ligands has also been reported2 . For core characterization, high-
resolution transmission electron microscopy produces an image while scanning tunneling
microscopy (STM) and X-ray diffraction analyze the core which allows for the
quantification of nanoparticle dimensions2. Transmission (TEM) and scanning electron
microscopy (SEM) are a popular characterization method and these tools image AuNPs
well because of the high density associated with gold6,7. TEM and SEM both allow
samples to be observed on high resolution, and when used in conjunction with electron
dispersive spectroscopy (EDS) an analysis of elemental composition is produced. EDS

3
utilizes back scattering X-ray when focusing an electron beam on the material in
question6. During simple light microscopy, challenges arise when attempting to depict
the differences between a AuNP scattering light from the background. Scanning near-
field optical microscopy and dark-field microscopy have been utilized in the past for their
imaging capabilities, but have fallen short. The problem with small nanoparticles is their
rapid scattering signal which vanishes and becomes optically undetectable8.
Gold nanoparticle interactions with bacteria
The increase in AuNP research and production calls for examination of their fate
in the environment. In one regard, clinical and industrial systems are interested in the
route of using novel materials such as nanoparticles for the eradication of biofilms that
may be a nuisance when biofouling equipment or a potential health threat when
pathogenic bacteria colonize a human-made system. In another regard, bacteria are a key
player in the ecosystem as they contribute as a food source, to decomposition and cycling
of organic carbon, and other important biogeochemical cycles9 and if toxic to bacteria,
nanomaterials released into the environment may cause a disruption in the ecosystem.
Promising results have been seen in research done using photothermal killing of
bacteria with gold nanoparticles. Photodynamic therapy produces a reactive oxygen
species (singlet of oxygen) that can cause bacterial damage. Gold is considered the
strongest nano-absorber due to the profound plasmon resonance associated with their
conducive electrons8,10. Zharov et al.10 looked at the pathogenic bacteria, Staphylococcus
aureus and used gold particles of sizes 10, 20, and 40 nm that were conjugated with

4
specific antibodies to cause damage to S. aureus cells. They developed a new approach
to produce physical damage to the bacterium when pulsed laser energy was absorbed by
the nanoparticles that had specifically attached to the bacteria. Irradiated nanoparticles
absorbed energy and heat remained at the site causing damage to the bacteria10. Another
study11 took advantage of the photostability of AuNPs and allowed them to absorb near-
infrared radiation. Once near-infrared radiation was absorbed, gold nanoparticles were
capable of transmitting heat. They used antibody-conjugated gold nanorods to bind to P.
aeruginosa. A LIVE/DEAD assay assessed viability of the cells and indicated a
significant decrease in viability for gold nanorod treated cells. TEM micrographs
illustrated large areas of cell membrane disruption either due to nanoparticle explosion
during irradiation, bubble formation and/or thermal disintegration11.
Aside from using photothermal dynamics against bacteria, many researchers are
discovering that gold may provide a level of toxicity of its own without using lasers for
selectively killing bacteria. This result often times questions the use of a surface
chemistry and whether it is the properties associated with the AuNP core itself, or the
surface coating with a purpose of maintaining a stable AuNP suspension. Exposure to
AuNPs has induced a heat shock response in E. coli12, as well as generating a 64%
reduction in viable bacteria13. Zhang et al.13 used a hyperbranched poly(amidoamine)
coated AuNPs and tested their toxicity on a large suite of gram-negative and positive
bacteria. At 2.8 mg/L, AuNPs demonstrated up to a 98% inhibition of bacterial growth.
The cationic charge to this specific surface coating allows for strong ionic interaction
with the bacteria cells and was noted as a possible contributor to the overall inhibition
5
seen in their study13. In comparison, Simon-Deckers et al.5 focused on using only citrate
to stabilize the AuNPs. Past studies have shown that non-functionalized AuNPs do not
interact with the cell wall of the bacteria and offer the explanation as they both have
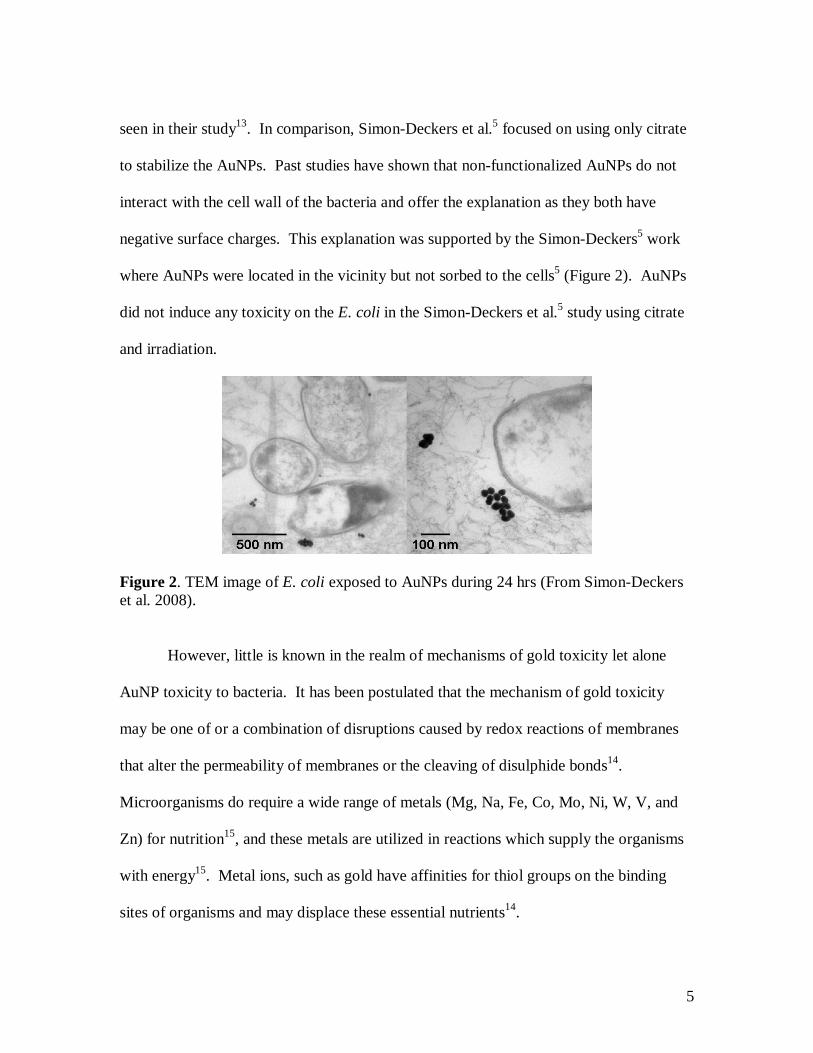
negative surface charges. This explanation was supported by the Simon-Deckers5 work
where AuNPs were located in the vicinity but not sorbed to the cells5 (Figure 2). AuNPs
did not induce any toxicity on the E. coli in the Simon-Deckers et al.5 study using citrate
and irradiation.
Figure 2. TEM image of E. coli exposed to AuNPs during 24 hrs (From Simon-Deckers et al. 2008).
However, little is known in the realm of mechanisms of gold toxicity let alone
AuNP toxicity to bacteria. It has been postulated that the mechanism of gold toxicity
may be one of or a combination of disruptions caused by redox reactions of membranes
that alter the permeability of membranes or the cleaving of disulphide bonds14.
Microorganisms do require a wide range of metals (Mg, Na, Fe, Co, Mo, Ni, W, V, and
Zn) for nutrition15, and these metals are utilized in reactions which supply the organisms
with energy15. Metal ions, such as gold have affinities for thiol groups on the binding
sites of organisms and may displace these essential nutrients14.

6
The possibility of trophic transfer
In contrast to the AuNPs exhibiting a negative toxic affect or not interacting with
the bacterial cell at all, some researchers have demonstrated a possible
compartmentalization and self assembly of AuNPs by bacteria. Bacteria that are capable
of compartmentalizing AuNPs may then serve as a source for trophic transfer of gold in
the environment. Some bacteria and diatoms are capable of producing inorganic
materials, and some bacteria have been shown to synthesize gold nanoparticles. This
“green” approach to producing nanoparticles was examined with an actinomycete
(Rhodococcus sp.) when it synthesized gold nanoparticles of 5-15 nm both on the cell
wall and internally16. There were no reported signs of toxicity during the interaction
between the gold and the actinomycete. A UV-vis spectra recorded the absorbance of
samples before and after the introduction of the AuNPs, and a broad peak at ~ 540nm
was observed, indicating aggregation of gold nanoparticles on or inside the cell. Using x-
ray diffraction to provide Bragg reflections of the gold and the Debye-Scherrer equation
they were able to estimate size of the gold nanoparticles inside a nano-actinomycete
biofilm16. This was not the only study detailing the internalization of gold into an
organism. Nakajima17 found bacteria efficiently removed gold from media and Tsuruta18
found gram-negative bacteria that accumulated gold.
Phytoplankton serves as another possible source in the facilitation of AuNP
transport up a food chain. Their distribution throughout the entire water column and their
relatively large surface to volume ratio are reasons for their exposure to and accumulation

7
of AuNPs4. One study used green algae, Scenedesmus subspicatus, a popular species in
French ecotoxicological tests. Mollusks such as Corbicula fluminea were also studied
and were thought to come into contact with gold nanoparticles based on their nutritional
and respiratory activities4. The analysis of metallothionein (MT) concentrations served
as a measurement of stress caused by the metals in the mollusks. Ten-nanometer AuNPs
were used with a positively charged amine surface chemistry. The S. scendesmus toxicity
test was a direct water-only exposure to AuNPs for 24h. A mortality rate of up to 40%
was reported after 12 hours for the algae (concentration of 1.6 * 105 AuNPs/cell) and the
LD50 occurred at 24 hours. Microscopy showed no AuNPs residing in cells, however
weakening of the cell wall was the expected cause of mortality4. The C. fluminea study
was considered a trophic exposure. The test mollusks were placed in a green algae and
AuNP suspension. AuNPs penetrated epithelia4 (shown by TEM below) however no
structural disturbances were seen. The analysis of the gills and visceral mass showed
elevated levels of metallothionein. Bioconcentration factors of 4,000 and 26,000 were
calculated in gills and visceral mass.

8
Figure 3. TEM of C. fluminea-AuNP interaction (Left: epithelial stomach cell Middle: epithelial branchial cell Right: epithelial cell. Figure from Renault et al. 2008.
Another filter feeder in a water column, Daphnia magna, maintain an integral
position in an aquatic food web. Being filter feeders, there exists a high potential for
contaminant accumulation to occur in this organism. Lovern et al.7 used gold
nanoparticles suspended in trisodium citrate with an average particle size of 17-23 nm to
assess the uptake and release of AuNPs in Daphnia magna. At concentrations of 500 ppb
AuNPs, entry into the D. magna after 6 hours was noted using TEM. Depuration
experiments indicated that a decrease in concentration of the AuNPs occurred over time,
however no bioaccumulation was detected7. Trophic transfer is still possible if D. magna
are exposed to surrounding waters containing AuNPs. D. magna could perhaps be the
first level of a trophic transfer. If an aquatic environment contained AuNPs, D. magna
could then filter feed and ingest the AuNPs and excrete pellets containing the AuNPs that
may then settle onto the benthic life.

9
The potential for bioaccumulation or toxicity of AuNPs has not been examined in
many species representative of every trophic level. Perhaps the least explored for
apparent toxicity or accumulation, and the most studied for the therapeutic effect of gold
is with mammals. When using mice in AuNP research, it is important to understand the
distribution of AuNPs. Available research to date indicates it is the size of the particle
that dictates its fate and ultimate distribution19. Oral administration of AuNPs resulted in
distribution to the kidney, liver, spleen, lungs, and brain with smaller particles
distributing further19. Most therapeutic research either uses AuNPs in a phosphate-saline
buffer injected intravenously or directly into the tumor, or when targeted with an
antibody20. One study by Hainfeld20 et al. examined the treatment of tumors in mice with
AuNPs and used either AuNPs intravenously injected with no x-ray irradiation, x-ray and
AuNPs, or x-ray alone. The AuNPs only treatment did not cause any apparent toxicity
but no therapeutic effect was seen. With x-ray and AuNPs all but one mouse had a tumor
remaining, of which was shrinking, and with x-ray alone the tumors grew to 5 times the
initial size after 1 month20. The lack of apparent toxicity to AuNPs seen in mice is also
the case with humans, as human cells can take up AuNPs with no cytotoxic effects21.
After conducting a literature search, there is a lack of research examining the effects of
AuNPs to mammalian models using the same surface chemistries that have caused known
toxic effects in the previously mentioned species.
Environmental risk assessment for gold nanoparticles
Typically, environmental safety data requires information regarding the fate and
transport of the nanoparticles, the hazards they poses to aquatic, benthic, terrestrial, and

10
atmospheric organisms, along with environmental exposure and risk for different
compartments. The aforementioned studies highlight the important concept of surface
chemistries, and when testing the effects of AuNPs on organisms, particular attention and
thorough assessment should be spent verifying any toxicity associated with the surface
chemistry alone.
For any nanoparticle, size, shape, associated chemistries, and chemistry of the
media may play a large role in dictating effects. The concentration at which AuNPs
become saturated and their kinetics in cellular uptake experiments are influenced by their
physical parameters22. Silver nanoparticles, along with gold, come in a variety of shapes,
and triangular silver nanoparticles have been found to exert more antibacterial action than
other shapes. It is recommended that when designing toxicity testing, all nanoparticle
products and by-products during and resulting from the production process are used as
test materials as well. Finally, once materials are chosen, size characterization,
composition, shape, and purity need to be addressed22. As of now, there is no single
protocol guiding researchers in how to characterize and quantify the fate transport of the
nanoparticles in situ. Fate and transport studies are limited as are values for
concentrations of nanoparticles relevant to a concentration which may be released in the
environment.

11
WORKS CITED
1) Eustis, S.; El-Sayed, M.A. Chem. Soc. Rev. 2006, 35, 209-17. 2) Daniel, M.C.; Astruc, D. Chem. Rev. (Washington, DC, U.S.) 2004, 104, 293-346. 3) Klaine, S.J.; Alvarez, P.J.J.; Batley, G.E.; Fernandes, T.F.; Handy, R.D.; Lyon, D.Y.; Mahendra, S.; McLaughlin, M.J.; and Lead, J.R. Environ. Toxicol. Chem. 2008, 27(9):1825-1851. 4) Renault, S.; Baudrimont, M.; Mesmer-Dudons, N.; Gonzalez, P.; Mornet, S.; Brisson, A. Gold Bulletin. 2008, 41, 116-126. 5) Simon-Deckers, A.; Brun, E.; Gouget, B.; Carrière, M.; Sicard-Roselli, C. Gold Bulletin 2008, 41, 187-194. 6) Weir, E.; Lawlor, A.; Whelan, A.; Regan F. Analyst 2008, 133,835-845. 7) Lovern, S.; Owen, H.; Klaper, R. Nanotoxicology. 2007, 2, 1743-5404. 8) Lindfors, K.; Kalkbrenner, T.; Stoller, P.; Sandoghdar, V. Phys. Rev. Lett. 2004, 93, 037401-1-4. 9) Pusch, M.; Fiebig, D.; Brettar, I.; Eisenmann, H.; Ellis, B.K.; Kaplan, L.A.; Lock, M.A.; Naegeli, M.W.; Traunspurger. Fresh. Biol. 1998, 40, 453-495. 10) Zharov, V.P.; Mercer, K.E.; Galitovskaya, E.N.; Smeltzer, M.S. Biophys. J. 2006, 90, 619-627. 11) Norman, T.J.; Grant, C.D.; Magana.; Zhang J.Z. J. Phys. Chem. 2002, 106, 7005-7012. 12) Hwang E.T.; Lee J.H.; Chae Y.J.; Kim B.C.; Sang B.I.; Gu, M.B. Abstracts, American Institute of Chemical Engineers Meeting, Salt Lake City, UT, USA, November 4–9, 2007, 60e. 13) Zhang, Y.; Peng, H.; Huang, W.; Zhou, Y.; Yan, D. J. Colloid Interface Sci. 2008, 325, 371-376. 14) Reith, F; Lengke, M.F; Falconer, D; Craw, D; and Southam, G. ISME. 2007, 1:567-584. 15) Madigan, M.T.; Martinko, J.M. Brock-Biology of microorganisms, 11th edition. 2006, Prentice Hall: New York, USA. 16) Ahmad, A.; Senapati, S.; Khan, M.I.; Kumar, R.; Ramani, R.; Srinivas, V.; Sastry, M. Nanotechnology. 2003, 14, 824-828. 17) Nakajima, A. WJ Microbiol. Biotech. 2003. 19:369-374. 18) Tsuruta, T. J Gen. Appl. Microbiol. 2004, 50, 221-228. 19) Hillyer, J.F.; Albrecht, R.M. Microsc. Microanal. 1999, 4, 481-490. 20) Hainefeld, J.F.; Slatkin, D.N.; Smilowitz, H.M. Phys. Med. Biol. 2004, 49, 309-315. 21) Connor, E.E.; Mwamuka, J.; Gole, A.; Murphy, C.J.; Wyatt, M.D. Small. 2005, 1, 325-327. 22) Tiede, K.; Hassellov; Breitbarth, E.; Chaudhry, Q.; Boxall, A.B.A. J. of Chromatogr. 2009, 1216, 503-509.

12
Gold Nanoparticle Exposure Alters Legionella
pneumophila Biofilm Formation
Amber R. Stojak, Stephen J. Klaine, and Tamara L. McNealy*
Institute of Environmental Toxicology (CU-ENTOX).
Department of Biological Sciences, Clemson University
509 Westinghouse Road, Pendleton, South Carolina 29670-0709
RECEIVED DATE (to be automatically inserted after your manuscript is accepted
if required according to the journal that you are submitting your paper to)
*To whom correspondence should be addressed: [email protected]
Address: Clemson University, Department of Biological Sciences, 132 Long Hall, Clemson, South Carolina 29634-0314.
ABSTRACT
Outbreaks of Legionella spp. represent a significant risk to human health. Hence,
biocides are used regularly to control populations of pathogenic organisms in many water
systems. Metal salts have shown limited success at permanently removing Legionella
from man-made systems; however, metallic nanoparticles may behave differently from
their ionic forms based on their size, shape and surface chemistries. This research
investigated the interactions between gold nanoparticles and Legionella pneumophila

13
biofilms. Transmission electron microscopy indicated association of both 18 and 4 nm
gold nanospheres with bacterial cells and biofilm. In treated biofilms, L. pneumophila
formed long chain-like assemblages that were unseen in thick, structured control and
polystyrene nanoparticle treated biofilms. This stress response suggests the potential uses
of gold nanoparticles to destabilize biofilms potentially making them more susceptible to
biocides.
KEYWORDS Legionella pneumophila, Biofilms, Gold Nanoparticles
Widespread concern exists for pathogenic bacteria colonization and establishment
of biofilms in water systems due to elevated risk of direct human exposure to disease.
Human-made aquatic environments are home to several pathogenic biofilm-forming
bacteria that populate the elevated temperature, low nutrient environments typically
found in such places1 and Legionella spp. are arguably of greatest concern2. Outbreaks of
Legionella spp. have occurred in water systems which experience periods of non-use
such as in cooling towers or hospital water3,4. This pathogenic bacterium persists in a
variety of complex niches and is often resistant to current biocidal or disinfectant
treatment methods, thus presenting a challenge on the basis for removal in industrial and
medical settings.
Legionella spp. are widely distributed in all waters including, but not limited to,
drinking and cooling systems, plant misters and spa pools. These warm water
environments are conducive to biofilm formation and facilitate the growth and success of
Legionella spp.2. Such biofilms then provide environments that can lead to human

14
exposure and infection resulting in Legionnaire’s Disease (LD)3,5. LD is a form of
pneumonia that can be severe enough to demand hospitalization3 and has a 10-50%
mortality rate if untreated6. Cases of LD increased 70% from 2002 to 2003 in the United
States and recorded numbers of >2000 cases per year from 2003-20052. Water systems
and cooling towers are prominent sources of LD outbreaks due to biofilm colonization
and later dispersal of contaminated aerosols from the showers and towers. In a 2003-
2004 outbreak in Pas-de-Calais, France, 86 cases resulted from contaminated aerosols of
a cooling tower containing amoeba and Legionella spp. These aerosols reached distances
of at least 6 km from the source. Twenty-one percent of the confirmed cases in this
outbreak were fatal7.
Prevention of colonization and therefore dissemination is difficult as biofilms are
resistant to disinfectants, biocides, and antibiotics, grow in hard to reach areas such as
dead end plumbing, or exist in or among complex niches8,9,10. In field trials using
chlorine, quaternary ammonium, pentachlorophenol, alkyl propanediamine, or methylene
bis thiocyanate in cooling towers at concentrations equal to or greater than the effective
dose used in preliminary, laboratory testing, the removal of Legionella pneumophila was
unsuccessful11. The reason for the contradictory results may have been due to the
complex niche L. pneumophila maintain in man-made water systems8. While some
studies have found a reduction in planktonic L. pneumophila growth after exposure to
copper and silver ions, neither metal ions, conventional chlorination, ultraviolet radiation,
nor heating of water in cooling towers and distribution systems have been successful at
the permanent removal of Legionella biofilms3,10. Part of the reason for the failed

15
removal with metal ions is their possible interaction with extrapolymeric matrix (EPS) of
biofilms resulting in reduced metal bioavailability to the bacterial cells12. The anionic
charges from hydroxyl, carboxylate, phosphate, and sulfhydrl functional groups
associated with the EPS may serve as ligands for metal cations. Ligand binding reduces
metal bioavailability and reduces biocidal efficacy 10. A further explanation for the failed
treatment attempts is that Legionella spp. are protected from treatment during their
intracellular invasion and replication in amoeba and can survive for extended periods of
time within amoebal cysts13. Most Legionella spp. in these systems are associated with
biofilms1, and it has been suggested that the biofilms serve as a shelter and nutrient
source for Legionella 9. The buildup of EPS and organic debris associated with this
biofilm prevents or reduces direct contact of biocides with Legionella, thus preventing
the successful removal of Legionella from a system 8.
Although dissolved metals, such as copper and silver, have been unsuccessful at
permanent removal of Legionella biofilms, metal nanoparticles behave differently,
depending on their physical and chemical characteristics14, and therefore may offer novel
mechanisms in treatment options. For example, gold nanoparticles (AuNPs) have been
found to inhibit the growth of Gram-negative and Gram-positive bacteria due to gold
strongly binding to the electron donating groups of the bacteria15. Past studies that
evaluated the anti-microbial effect of AuNPs have stabilized the particles with various
surface chemistries including hyperbranched poly(amidoamine) with terminal
dimethylamine groups ((HPAMAM-N(CH3)2)15 and polyvinyl composite films16. These
surface chemistries also resulted in inhibition of bacterial growth. Aside from causing

16
98% growth inhibition in a large suite of Gram-negative and Gram-positive bacteria at a
concentration of 2.8 mg/L15, AuNPs have been shown to induce a heat shock response in
E. coli17. Metal nanomaterials, if effective at controlling pathogenic biofilms, would be
of use in water cooling towers and recirculating systems. However, despite the fact that
these and other data suggest that nanoparticles can affect bacteria in suspension, virtually
no work has been performed using biofilms18. The goal of this research was to
characterize the effects of gold nanoparticles on L. pneumophila both in biofilm and
planktonic stages.
L. pneumophila (Philadelphia 1) was cultured on buffered charcoal yeast extract
(BCYE, VWR) agar at 37˚C and sampled from the agar plate after 3 days incubation. L.
pneumophila taken from these plates was suspended in buffered yeast extract (BYE)
broth at the start of an assay at experiment-specific optical densities (OD) and dilutions.
Gold nanoparticles were obtained from Dr. Catherine Murphy (University of South
Carolina) in both 18 and 4 nm spherical diameters, 0.22 mM Au, capped with a citrate
coating in a citrate solution. Biofilm assays were performed with both sizes of
nanoparticles to test for a potential size effect. Polystyrene, 20nm fluorescent spheres
coated in carboxylate groups in a 2% azide solution (Fluospheres, Invitrogen) were also
tested to determine if effects were a function of the nanoparticle chemistry. All
nanoparticles were imaged before and after the start of the experiment using transmission
electron microscopy to confirm their size distributions (TEM, Clemson Electron
Microscopy Lab, Hitachi 7600, Supplemental Material Figure 1).

17
Qualitative assessments of biofilms were conducted using Syto 11 (Invitrogen)
cell permeant nucleic acid dye and confocal microscopy. Glass slides were placed
horizontally in Pyrex chambers containing 40 ml of a 0.600 OD600 suspension of L.
pneumophila in BYE broth. Preliminary studies have indicated 5 days were sufficient for
L. pneumophila to establish irreversibly attached biofilms in this system (Supplemental
Material Fig 2). At day 5, BYE broth was removed, slides were rinsed gently by placing
into sterile water and removed and placed horizontally back into position. Forty-
milliliters of moderately hard water (MHW19) produced with 18 mega-ohm water and
reagent grade salts was added (Hardness= 80 mg CaCO3/L alkalinity= 60 mg CaCO3/L
pH=7.7) with 0.7 µg/L AuNPs (1.09 x 104 particles/µL 18nm AuNPs, 1.02 x 106
particles/µL 4nm AuNPs), 1.32 x 109 particles/µL polystyrene beads or MHW alone.
Two days after media exchange, slides were removed, rinsed and methanol fixed for 15
minutes, followed by staining with 5 µM Syto 11 dye in water. Imaging was performed
by confocal microscopy and biofilm morphology and appearance assessed for all
samples.
Controls formed thick, condensed biofilms (Figure 1a). For all assessments for
this study, we considered a biofilm to be fully established when bacterial cells or the EPS
produced by the L. pneumophila remained irreversibly attached to substrates post sterile
water washes. In biofilms treated with 18 nm AuNPs, a morphological change in cells
and biofilms was observed after 2 days. Chain-like assemblages of elongated cells were
noted on all treatment slides (n= 34, Figure 1b). The thick, organized, biofilm formation
seen in controls was patchy if not altogether absent after AuNP exposure. The

18
morphology differences were also seen in biofilms treated with 4 nm AuNPs. Similar to
the 18 nm treatments, a chain-like assemblage with little or no significant biofilm was
observed (Figure 1c). This morphological change was not seen when biofilms were
exposed to similar sized, polystyrene beads for the same time period (Figure 1d).
C20 µm
A B20 µm
DC20 µm
A B20 µm
DC20 µm
A B20 µm
D Figure 1. Confocal microscopy shows induction of morphology changes after exposure to AuNPs. a) control biofilm and b) 18 nm gold c) 4 nm gold and d) 20 nm polystyrene treated biofilms.
AuNPs were next evaluated for their influence on L. pneumophila viability. The
parameters measured in this assay included growth kinetics, biomass, and pigment
production. An indication of virulence was assessed by measuring pigment production,
as the ability to reduce iron comes from hemogentisic acid originally produced from the
legiolysin (Lly) locus, and this ability to reduce iron is needed to successfully infect a
host cell20,21.
Planktonic viability was evaluated by examining growth kinetics after exposure to
18nm and 4nm AuNPs. L. pneumophila from 3 day old BCYE plates were added to
sterile BYE broth and each flask was inoculated at 0.05 OD600. The initial culture was
plated for determination of initial concentration. The culture was then divided into 5
tubes (VWR, BD Sciences) with a control, AuNP concentration of 1 µg/L added at t=0,

19
and then three concentrations (1, 10, and 100µg/L) added in the delayed samples at
t=18h. Previous work provided time points specific to L. pneumophila along a typical
bacterial growth curve. In this experiment design, idiophase, the transition from
exponential and stationary phase, occurred at t=18 hr. L. pneumophila transitioned at this
point from a replicative, non-transmissive phase to an infective, non-replicating phase
that is capable of infection. T=30 hrs was equivalent to the entry into stationary phase
and 48 hrs was the beginning of the bacteria’s decline phase. This viability assay
evaluated the bacteria-AuNP interation and utilized the above mentioned time points
along various stages of bacterial growth.
At each time point supernatants of samples collected were measured at OD400 for
pigment analysis while pellets were resuspended in sterile water and measured at OD600
to measure growth kinetics. Pigment production was initially low for all treatments but
increased by t=30 hr, typical for L. pneumophila. Pigment increased in all samples and
was not statistically significantly different between controls and treatments (ANOVA,
p>0.05, Figure 2). The OD600 for 18 nm treatments at 18hr increased from the 0.05 initial
OD600 to 1.28 (±0.024) for all samples with no significant differences between treatments
and controls. Overall, 18 nm AuNP treatments showed no effect on L. pneumophila via
measurements of growth, biomass, or pigment production suggesting no anti-microbial
activity of 18nm AuNPs on L. pneumophila.

20
Figure 2. Planktonic viability assays with 18 nm AuNPs show no significant differences in growth kinetics (OD600) or pigment production (OD400). T=18 hr is represented by black bars, t=30 hr by gray, and t=48 hr by gray and white for both OD400 (top) and OD600 (bottom). Standard error bars (standard deviation) on each, and treatment concentrations located on x-axis and optic density readings on y-axis.
The 4nm planktonic viability assays were conducted using the same methods
discussed previously used for the 18 AuNPs. No statistically significant differences were
observed in OD during the 18 hr time point (Figure 3). At the 30 hr time of sampling, a
decreasing trend was seen in the biomass. As the AuNP concentrations increased,
biomass decreased and there were statistically significant differences between the highest
treatment compared to the control, t=0 hr addition of nanoparticles, and the 1µg/L
concentration (ANOVA p<.05, Figure 3). The 10µg/L delay treatment was not
statistically significant from the any of the treatments, however the biomass was less than

21
the control, t=0 hr and 1 µg/L treatments and it began the decreasing trend in biomass at
30 hrs (Figure 3). At 48 hr sampling time however, biomass had increased in the 10 and
100 µg/L delay concentrations so that they were no longer statistically different from the
other treatments (ANOVA p> 0.05). A slight decrease in pigment production in the 10
µg/L delay occurred as well as a significant decrease in the 100 µg/L delay treatment
(ANOVA, p<0.05) as compared to controls. All other treatments exhibited a trend of
increasing pigment production as concentration increased, with the 1 µg/L delay being
significantly higher than all other treatments (Figure 3).

22
Figure 3. Results from the 4 nm planktonic viability assay indicate slight decreases in biomass at 30hr indicated by 100µg/L treatment, and an asterisk showing a statistically significant difference from controls. OD400 represented by gray bars and OD600 represented by black bars. Standard error bars on each, and treatment located on x-axis and optic density readings on y-axis.
As the alteration in biofilm morphology did not appear to be due to anti-microbial
effects, nanoparticle-bacteria interactions were examined by transmission electron
microscopy (TEM) to determine the level of association nanoparticles exhibited with L.

23
pneumophila planktonic cells. Cells were imaged in both sectioned and whole mount
samples. Samples of planktonic L. pneumophila at the t=30 hr time point were collected
for TEM analyses. Briefly, samples for sectioning were collected and fixed in 3%
gluteraldehyde for at least 4 hours followed by overnight incubation in cacodylate rinsing
buffer (pH 7.4). Samples were then stained with 1% osmium tetroxide solution and rinsed
twice in rinsing buffer. An ethanol dehydration series was conducted followed by
embedding in LR White resin and samples allowed to polymerize at 60˚C oven overnight.
Samples were then sectioned using an ultramicrotome at a thickness of approximately
100 nm and placed onto a formvar coated copper grid (Electron Microscopy Sciences)
and coated with lead citrate and uranyl acetate. For whole cell analysis, samples of 10µL
from t=30 hr were taken and pipetted directly onto copper grids and fixed in 3%
gluteraldehyde vapors for 1 hour, then coated in lead citrate and uranyl acetate. All
sectioned and whole samples were then analyzed on a Hitachi 7600T (Electron
Microscope Facility, Clemson University). Gold content was confirmed in samples using
energy dispersive x-ray spectroscope (EDS, Inca Energy Program) as part of the TEM
analysis.
The overall integrity of the bacterial cells in the whole mount TEM images
appeared normal and cells overall appeared free of lethal abnormalities. However, we
were able to distinguish differences in the appearance of the cell surface due to the
association of AuNPs. The surfaces of treated cells had large dark aggregates indicative
of the AuNPs, while the controls did not (Figures 4a-c). Further imaging on sectioned
samples allowed the evaluation of the ultimate fate of the AuNPs. AuNPs were seen

24
both tightly associated to the outside of the cell as well as having been taken up into the
cell. Upon uptake into the cell AuNPs appeared to form aggregates of various sizes and
became compartmentalized into one area of the cell (Figures 4 d-f). Analysis of EPS
produced by the bacteria appeared similar in size and structure for treatments and
controls. AuNP exposure did however lead to adsorption of the particles into the matrix
of treated biofilms (Figure 4g,h). The presence of AuNPs in both whole mount and
sectioned samples was confirmed via EDS analysis. Control images contained no gold
(Table 1) using EDS analysis program for any metallic gold peak at 2.1, 9.7, and 11.8
keV. All negative values were reported as 0% gold. The weight percent of gold found
in treatments varied greatly by area selected for analysis (Table 1), but some amount of
gold was detected in all treatment fields viewed. EDS further confirmed the presence of
AuNPs adsorbed to the EPS with no gold detected in the controls.

25
Figure 4. Transmission electron micrographs of Control, 18nm and 4nm AuNP exposed L. pneumophila. Whole bacteria preparation of a) control and b-c) 18nm AuNP treatment; sectioned samples of d) control and e) 4nm AuNPs demonstrating uptake and f) 18nm AuNPs demonstrating sequestration of large aggregates; Extrapolymeric matrix of g) control and h) AuNP exposed samples.

26
Table 1. Elemental dispersion x-ray spectroscope average % wt. reports for gold in TEM samples
Sample
Timepoint
(hrs)
Average % gold
in field of view
18 nm 30 1.75
4 nm 30 1.60
Control 30 0
Changes in morphology were seen at both biofilm and planktonic stages. Biofilm
morphology observations indicated L. pneumophila formed thick and structured biofilms,
irreversibly attached to glass substrates. Upon treatment with AuNPs, biofilm
morphology changed with cells forming chain-like assemblages (Figure 1 b,c) and
filamentous, elongated cells occurred in planktonic cells at stationary phase (Figure 4.
e,f). Details on exact mechanisms behind this change and the benefit it may provide are
still unknown for L. pneumophila22. It has been reported that during late growth phase,
decreased expression of the global regulator CsrA may induce filamentation23, and it has
also been suggested that filamentation is involved in biofilm formation24. However, most
frequently, filamentation has been documented to occur as a result of environmental
stresses, changes, or as a result from treatment with antibiotics. Environmental change or
stress occurring with L. pneumophila has indicated a survival strategy by a change in
overall morphology24. At higher temperatures or when treated with the antibiotic
erythromycin, L. pneumophila cells were not their typical rod-shape, but had become

27
elongated and filamentous24,25, a pattern also seen only in our AuNP treatments. It was
also noted that filamentation occurred in other genera of bacteria such as E. coli and
Salmonella when physiological changes take place or in response to treatment with
antibiotics26,27. This suggests that this morphological response may be a conserved
mechanism evolved in response to stress or environmental change. When we examined
L. pneumophila cultures for expression of a heat shock protein, no increased expression
was seen in samples after exposure to AuNPs (data not shown). The exact cause for
elongation being unknown, we were unable to discern whether the response exhibited in
the AuNP treated biofilms is indicative of stress or triggered by the mere presence of the
AuNPs. An example of a non-stress response to gold, He et al.28 reported that upon
exposure of Bacillus subtilus to citrate capped AuNPs, the bacteria assembled end-to-end
while the AuNPs attached to their cellular surfaces where sulfur groups existed. The
explanation was that the attachment to the bacterial cells was not due to electrostatic
attraction, as both have negative charges, but to the attraction of gold to sulfur and
phosphorus groups, based on the theory of hard and soft acids and bases28. The electron
microscopy images revealing the presence of AuNPs on the surface of L. pneumophila
suggests that this effect could also be occurring in our system.
In addition to the morphology change seen in the bacteria and in the overall
biofilm structure, it was also observed that much less biofilm was present after exposure
to AuNPs. This decrease in biofilm mass suggests that AuNPs could induce structural
changes in the biofilm leading to dispersal events, a mechanism that could be capitalized
on in novel treatment and disinfection schemes. L. pneumophila biofilms were

28
morphologically altered and potentially destabilized as they were reduced in size and
structure. AuNPs may then provide new routes for biocidal attack on the cell and EPS,
therefore either result would thus increase the overall effectiveness of a biocide. If
AuNPs are capable of 1) destabilizing the matrix and/or 2) altering cellular morphology
to loosen the biofilm itself, then a biocide could be used to cause mortality to dispersed
planktonic cells. Results of an earlier study using silver ions to inhibit Staphylococcus
epidermidis biofilms also suggested using such a combination approach. In this study, the
EPS was found to be destabilized, but the bacteria themselves were not inhibited as the
silver was incapable of penetrating the extensive EPS29. Their conclusion that a
disinfection protocol of silver ions would work synergistically with a biocide would agree
with our findings here. If NPs can be used to cause a sublethal effect, as shown here with
the destabilization of biofilms and decrease in size and structural depth, a lower
concentration of the biocide may be sufficient to remove biofilms from the system.
The morphological change in the biofilm may not necessarily result in cell death
as neither growth nor pigment production (a factor associated with virulence in
Legionella) was effectively inhibited in our assays (Figure 2, 3 and data not shown).
Here, we used low concentrations in order to detect sublethal effects and found no growth
inhibition with 18 nm, but a slight yet statistically significant reduction in biomass using
4 nm citrate capped AuNPs. Pal et al.27 examined the antibacterial efficiency of silver
nanoparticles of different sizes and shapes and found that effect was inversely related to
particle size (increased surface area). Other studies have found AuNPs with various
surface chemistries, including hyperbranched poly(amidoamine) with terminal

29
dimethylamine groups ((HPAMAM-N(CH3)2)15 and polyvinyl composite films16
inhibited bacterial growth. However, the antimicrobial action of the surface chemistry
itself may have confounded conclusions drawn from these studies. Hence, AuNP size
and surface chemistry may potentially influence the microbial response. Further, core
particle chemistry may play a role since polystyrene spheres of similar size did not elicit
the same effects in our analyses as AuNPs (figure 1). A sublethal effect in planktonic
bacteria such as inhibition of growth or virulence caused by the gold particle may aid in
the permanent removal of L. pneumophila from a system, while the synergistic use of low
concentrations of AuNPs and biocide offer a potential strategy for pathogenic biofilm
control.
These results represent new routes for biocidal contact and actions for both
biofilms and planktonic bacterial stages. Not only were the 18 and 4nm AuNPs able to
adsorb to EPS but were also capable of entering the bacterial cell. The small size and
available surface area may allow the AuNPs to travel to dead end piping in distribution
systems, penetrate the complex niche where L. pneumophila survive, and reach the
previously considered resistant bacterial cell. An even greater effect may be seen using
AuNPs with additional geometric planes, an increased overall surface area or different
surface chemistry. To further evaluate the use of AuNPs in synergism with a biocidal
administration protocol, a variety of synthesized gold nanoparticles should be tested
along with higher concentration ranges. Not only will different sizes, concentration
ranges, shapes, and surface chemistries of AuNPs reveal the most efficacious at causing

30
sublethal changes in bacteria, but also illuminate the reasons prompting the
morphological changes and inhibition of growth in bacteria.
Finally, this research is the first to examine the interaction of nanoparticles with
biofilms. While single species biofilms do not occur in natural ecosystems, the results of
this research underscore the need for quantitative assessment of the potential impacts of
nanoparticles on biofilms. This is particularly critical since a likely fate of nanoparticles
in aquatic ecosystems is sedimentation onto surfaces almost always covered by biofilms.
We would like to thank Luke Bury and Marie Cappelle for their assistance in
planktonic viability assays. For TEM/SEM imaging I thank Aaron Edgington, Dr. JoAn
Hudson, Donald Mulwee, and Dayton Cash for their expertise and Clemson University
Electron Microscopy Lab for the use of the equipment. For the gold nanoparticle
synthesis we thank Dr. Catherine Murphy’s lab, specifically Stefano Boulos, Department
of Chemistry, University of South Carolina. Also we acknowledge the Jordan Hall
Imaging Facility, Clemson University for use of confocal microscopy, and Clemson
University for funding and support.
Supporting Information Available: The characterization procedure and micrographs of all nanomaterials used in this paper are included. Also, time series biofilm growth study methods along with image are available.

31
REFERENCES (1) Fields, B.; Benson, R.; Besser, R. Clin. Microbial. Rev. 2002, 15, 506-526. (2) Neil, K.; Berkelman, R. Clin. Infect. Dis. 2008, 47, 591-599. (3) Atlas, R.M. Environ. Microbial. 1999, 1, 283-293. (4) Garcia-Nunez, M.; Sopena, N.; Ragull, S.; Pedro-Botet, M.L.; Morera, J.; Sabria, M. FEMS Immunol. Med. Microbiol. 2007, 52, 202-206. (5) Rogers, J.; Keevil, C.W. Appl. Environ. Microbiol. 1992, 58, 2326-2330. (6) Wever, P.C.; Yzerman, E.P.F.; Kuijper, E.J.; Speelman, P.; Dankert, J. J. Clin. Microbiol. 2000, 38, 2738-2739. (7) Nguyen, T.M.N.; Iief, D.; Jarraud, S.; Rouil, L.; Campese, C.; Che, D.; Haeghebaert, S.; Ganiayre, F.; Marcel, F.; Etienne, J.; Desenclos, J. Infect. Dis. 2006, 193, 102-111. (8) Bentham, R.H.; Broadbent, C.R. Curr. Microbiol. 1995, 30, 167-172. (9) Temmerman, R.; Vervaeren, H.; Noseda, B.; Boon, N.; Verstraete, W. Appl. Environ. Microbiol. 2006, 72, 4323-4328. (10) Harrison, J.; Ceri, H.; Turner, R.J. Nature. 2007, 5, 928-938. (11) England, J.C.; Fraser, D.W.; Mallison, G.W.; Mackel, D.C.; Skaliy, P.; Gorman, G. Appl. Environ. Microbiol. 1982, 43, 240-244. (12)Silvestry-Rodriguez, N.; Bright, K.R.; Slack, D.C.; Uhlmann, D.R.; Gerba, C.P. Appl. Environ. Microbiol. 2008,74, 1639-1641. (13) Wery, N.; Bru, V.; Minervini, C.; Delgenes, J.P.; Garrelly, L.; Godon, J.J. Appl. Environ. Microbiol. 2006, 74, 3030-3037. (14) Daniel, M.C.; Astruc, D. Chem. Rev. (Washington, DC, U.S.) 2004, 104, 293-346. (15) Zhang, Y.; Peng, H.; Huang, W.; Zhou, Y.; Yan, D. J. Colloid Interface Sci. 2008, 325, 371-376. (16) Shanmugam, S.; Viswanathan, B.; Varadarajan, T.K. Mater. Chem. Phys. 2006, 95, 51-55. (17) Hwang E.T.; Lee J.H.; Chae Y.J.; Kim B.C.; Sang B.I.; Gu M.B. Analysis of nanoparticles’ toxic modes of actions by using recombinant bioluminescent bacteria. Abstracts, American Institute of Chemical Engineers Meeting, Salt Lake City, UT, USA, November 4–9, 2007, 60e. 18) Klaine, S.J; Alvarez, P.J.J; Batley, G.E; Fernandes, T.F; Handy, R.D; Lyon, D.Y; Mahendra, S; McLaughlin, M.J; and Lead, J.R. 2008. Nanomaterials in the environment: behavior, fate, bioavailability, and effects. Environmental toxicology and chemistry. 27(9):1825-1851. (19) EPA, U.S., Methods for measuring the acute toxicity of effluents and receiving waters to freshwater and marine organisms. In: EPA, U.S., 2002. Short-term methods for estimating the chronic toxicity of effluents and receiving waters to freshwater organisms. US EPA, p. 350. (20) Chatfield, C.H.; Cianciotto, N.P. Infect. Immun. 2007, 75, 4062-4070. (21) Steinert, M.; Flügel, M.; Schuppler, M.; Helbig, J.H.; Supriyono, A.; Proksch, P.; Lück, P.C. FEMS Microbiol. Lett. 2001, 203, 41-47. (22) Justice, S.S.; Hunstad, D.A.; Cegelski, L.; Hultgren, S.J. Nature Rev. Microbiol. 2008, 6, 162-168.

32
(23) Fettes, P.S.; Forsbach-Birk, V.; Lynch, D.; Marre, R. Int. J. Med. Microbiol. 2001, 5, 353-360. (24) Garduno, R.A.; Chong, A.; Faulkner, G. Developmental cycle- differentiation of Legionella pneumophila. In Legionella molecular microbiology,Heuner, K.; Swanson, M.; Horizon Scientific: U.K, 2008; 55-63. (25) Piao, Z.; Sze, C.C.; Barysheva, O.; Iida, K.; Yoshida, S. Appl. Environ. Microbiol. 2006, 72, 1613-1622. (26) Elliott, T.S.J.; Rodgers, F.G. J. Med. Microbiol. 1985, 19, 383-390. (27) Rico, A.I.; Garcia-Ovalle, M.; Mingorance, J.; Vicente, M. Mol. Microbiol. 2004, 53, 1359-1371. (28) Yoshida, S.; Udou, T.; Mizuguchi, Y.; Tanabe, T. J. Clin. Microbiol. 1986, 23, 192-194. (29) Pal, S.; Tak, Y.K.; Song, J.M. Appl. Environ. Microbiol. 2007, 73, 1712-1720. (30) He, Y.; Yuan, J.; Su, F.; Xing, X.; Shi, G. J. Phys. Chem. 2006, 110, 17813-17818. (31) Chaw, K.C.; Manimaran, M.; Tay, F.E.H. Antimicrob. Agents Chemother. 2005, 49, 4853-4859.

33
SUPPLEMENTAL MATERIAL
Nanoparticles: Gold nanoparticles were obtained from Dr. Catherine Murphy’s lab
(University of South Carolina). Gold nanoparticles (AuNP) were received in excess
citrate in a 0.22 mM solution. Gold nanoparticles were functionalized with citrate in
order to maintain a stable suspension and avoid aggregation. We also used 20 nm
polystyrene beads (Fluospheres, Invitrogen) that were carboxylate modified and coated
with a hydrophilic polymer with carboxylic acids. Potential aggregation was assessed
before and immediately after each experiment using transmission electron microscopy
(Hitachi 7600T, Clemson Electron Microscopy Lab, Figure 1). When aggregates in
samples were found (Figure 2) the sample was not used in any further assays. Here in
Figure 2 is a representation of the shortest time lapse where aggregation occurred.
Typical aggregation of AuNPs occurred after approximately 5 months, however here the
NPs aggregated after only 3 months from production date.
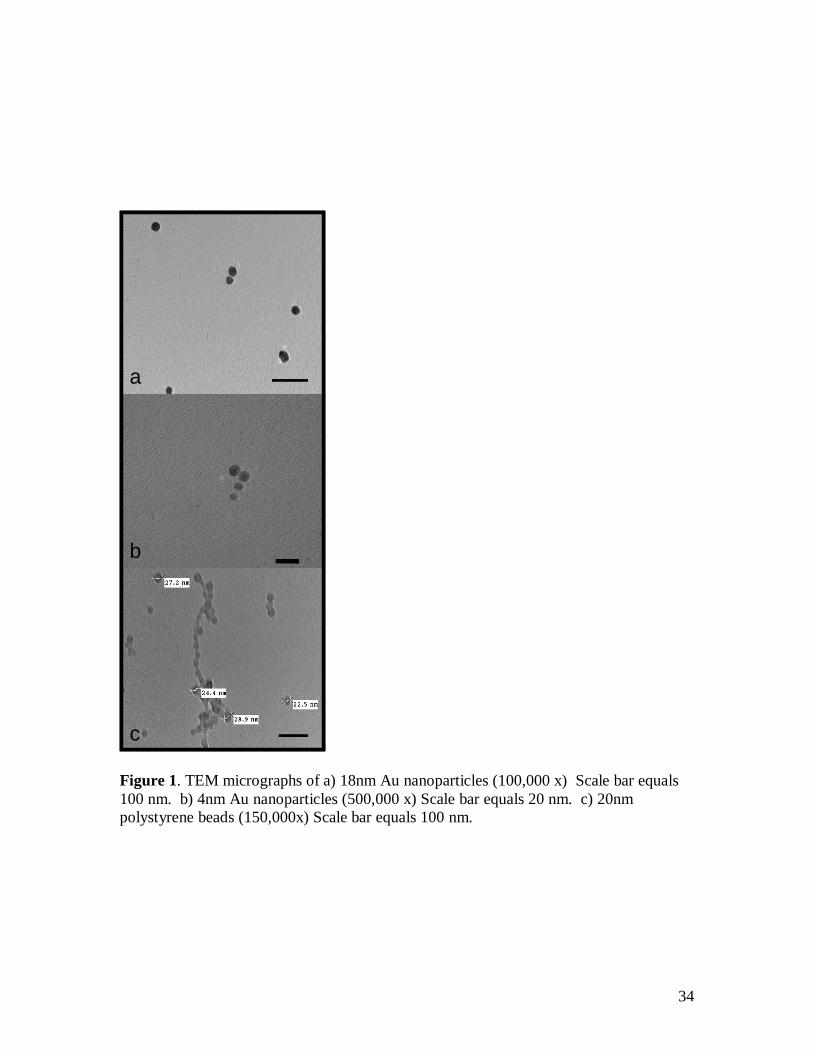
Imaging raw materials: A dilution series was plated initially for the AuNPs and
polystyrene nanoparticles for the ease of imaging. A 10-1 dilution (Figure 1 a, b) was
used for AuNPs in order to avoid over lapping particles during the drying process. No
dilution was used for polystyrene nanoparticles. Raw samples were taken from either the
dilution or directly from the stock solution, as for the polystyrene beads, and 10 µl was
pipetted directly onto formvar coated copper grids (Microscopy Sciences). The 18 and 4
nm nanoparticles appeared uniform in size (c.a 18 or 4 nm, figure 1 a,b), and stock
solutions of AuNPs were not used after 4 months, as aggregation was noted. Polystyrene
beads never aggregated over time, and were also found to be uniform in size (figure 1c).
34
a
b
c
a
b
c
Figure 1. TEM micrographs of a) 18nm Au nanoparticles (100,000 x) Scale bar equals 100 nm. b) 4nm Au nanoparticles (500,000 x) Scale bar equals 20 nm. c) 20nm polystyrene beads (150,000x) Scale bar equals 100 nm.

35
Figure 2. TEM micrograph of a) 4nm Au on January 15, 2009 scale= 20nm and b) 4 nm Au on March 19, 2009, scale = 500 nm. Biofilm growth over time series: Prior to the studies of the interaction of nanoparticles
with Legionella pneumophila, the allotted time required for a thick, structured biofilm
production was studied. L. pneumophila grown on BCYE agar for 3 days, and loopfulls
of L. pneumophila were added to BYE broth at 0.12 OD600. Ten-milliliters of the
innoculum was added to 90 ml of BYE broth into a rectangular Wheaton staining dish
with slides both horizontal (2) and vertical (2). Wheaton dishes were placed in an
incubator at 25˚C for either 2, 3, 5, 7, or 8 days. Media was removed on these days and
slides were rinsed three times with sterilized DI water. Slides were placed in methanol
for 10 min and then stained with Giemsa’s Solution for 15 min. Slides were imaged via
light microscopy and 5d were determined necessary for thick, structured, irreversibly-
attached biofilm growth.

36
Figure 3. Light microscopy image indicating sufficient L. pneumophila biofilm growth, stained with Geimsa dye, of a 5 day incubation in BCYE followed by 2 day incubation period in moderately hard water.


















