
Glycan variation and evolution in the eukaryotes -...
20
Assigned Reading: Glycan Variation and Evolution in the Eukaryotes Genome Analyses Highlight the Different Biological Roles of Cellulases
Transcript of Glycan variation and evolution in the eukaryotes -...

Assigned Reading: Glycan Variation and Evolution in the Eukaryotes Genome Analyses Highlight the Different Biological Roles of Cellulases

Glycan Variation and Evolution in the Eukaryotes

Special Issue: The Magic of the Sugar Code
Glycan variation and evolution in theeukaryotesAnthony P. Corfield1 and Monica Berry2
1 Mucin Research Group, University of Bristol, School of Clinical Sciences, Bristol Royal Infirmary, Bristol, BS2 8HW, UK2 University of Bristol, School of Physics, Centre for Nanoscience and Quantum Information, Tyndall Avenue, Bristol, BS8 1FD, UK
Review
In this review, we document the evolution of commonglycan structures in the eukaryotes, and illustrate theconsiderable variety of oligosaccharides existing inthese organisms. We focus on the families of N- andO-glycans, glycosphingolipids, glycosaminoglycans,glycosylphosphatidylinositol (GPI) anchors, sialic acids(Sias), and cytoplasmic and nuclear glycans. We alsooutline similar and divergent aspects of the glycansduring evolution within the groups, which include inter-and intraspecies differences, molecular mimicry, viralglycosylation adaptations, glycosyltransferase specifici-ty relating to function, and the natural dynamism power-ing these events. Finally, we present an overview of thepatterns of glycosylation found within the groups com-prising the Eukaryota, namely the Deuterostomia, Fungi,Viridiplantae, Nematoda, and Arthropoda.
Glycan structure: why it mattersIt is well known that the surface of cells at all levels of life iscoated with an array of glycans, often termed the ‘glyco-calyx’ [1–4]. The presence of this sugar coat is linked withbiological functions that are fundamental to the normalbehavior of the organisms throughout their life. Suchfunctions range from imparting protein stability and func-tion [5], to mediating the interaction of cells with theextracellular matrix [6–9], to sperm–egg binding processes[10], and host–pathogen interactions [11–16]. The biologi-cal choice of glycans as targets for such interactions reflectsboth their diversity and specificity. It also places demandson glycosylation to maintain and adapt to dynamic biologi-cal environments.
Here, we consider glycan structure and glycoconjugatefunction within the eukaryotes. Details for the individualphyla can be found in supplemental material online. Aphylogenetic tree for eukaryotic species that have had theirgenomes sequenced is shown in Figure 1. The CAZy data-base (http://www.cazy.org) provides an invaluable source ofinformation regarding all proteins that manipulate gly-cans in all their forms and is widely used as a primarytarget for genomic screening of species-specific glycomes
0968-0004/
� 2015 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tibs.2015.04.004
Corresponding authors: Corfield, A.P. ([email protected]);Berry, M. ([email protected]).Keywords: glycan; glycosylation; eukaryotes; evolution; glycosyltransferase;molecular mimicry.
[17]. The Consortium for Functional Glycomics websitealso provides a valuable tool for glycomic research(http://www.functionalglycomics.org). A consequence ofthe increased biological interest in glycans has been afocus on the chemistry–glycobiology frontier and the needto understand chemical and physical approaches for glycananalysis [3,4,18,19].
Sugars chains face the outside world and can occupymost of the hydrodynamic volume of any molecules thatcarry them. Together with cognate molecules, they createbonds and assist intercellular recognition. Through spatialorganization and concentration, they confer a further de-gree of selectivity to the formation of these bonds. Glyco-sylation is the default protein modification acrossprokaryotes [20] and eukaryotes; it affects fundamentaland conserved aspects of the protein life cycle, includingfolding, intracellular trafficking, and co-translational qual-ity control. The processes that enable protein maturationare homologous and occur in all kingdoms of life. Relevantto this discussion, the assembly of an oligosaccharide onthe cytoplasmic side of the plasma membrane by a set ofspecific glycosyltransferases and the subsequent translo-cation of the lipid-linked oligosaccharide is a unifyingscheme in all systems.
Changes to glycosylation in tumor cells [21] occur toboth core and outer carbohydrate structures. Among thefirst are: (i) overexpression of b1–6 branching of N-glycan[22]; (ii) greatly enhanced presence of truncated T, Tn, andsialyl-Tn glycans on mucins [23,24]; and (iii) loss of GPIanchors [25]. Increased Sia content [26,27], and enrich-ment in hyaluronan in the stroma surrounding a tumor[28–30] are examples of peripheral glycan changes inmalignancies.
Here, we focus on eukaryotic glycosylation, whichrepresents only an instance of glycosylation, given thatcontinuous glycan evolution is traceable through toEubacteria and Archaea. A more comprehensive reviewof bacterial glycosylation can be found in the article by Tanet al. in this special issue of TiBS [20]. Several earlierpublications have addressed the variation of eukaryoticglycosylation compared with other organisms and theirrole in evolution (e.g., [31–38]). Furthermore, the scope ofeukaryotic glycosylation is too broad to review in fulldetail; here, we highlight the fundamental patterns andconcepts of glycan structure and evolution, illustratedwith examples.
Trends in Biochemical Sciences, July 2015, Vol. 40, No. 7 351

Arthropoda
DeuterostomiaNematoda
ViridiplantaeFungi
Fruit fly(Drosophila melanogaster )
Roundworm(Caenorhabdi�selegans)
Mustard weed(Arabidopsisthaliana) Yeast
(Saccharomycescerevisiae)
Trypanosome
Sea urchin
Fish
Frog
Chicken
Mouse
Human
First fossilanimals
Last common ancestor800 mya 400 mya Present
Chimpanzee
Eukaryota
Slime mold Candida
First fossilland plants
Hydra
Sponge
TiBS
Figure 1. Eukaryote phylogenetic tree. Phylogenetic tree of the multicellular
Eukaryota, showing the main groups discussed in this review. Reproduced from
Essentials of Glycobiology, Chapter 19 page 282, with permission of Cold Spring
Harbor Press, P. Gagneux and TD Pollard. Abbreviation: mya, million years ago.
Review Trends in Biochemical Sciences July 2015, Vol. 40, No. 7
Glycan variation in the eukaryotesExamination of glycoconjugate properties immediatelyuncovers a multiplicity of structures that require appraisalin terms of their function within these molecules. Table 1
Table 1. Major groups of proteins manipulating carbohydrates in
Protein family Main categories
Glycosyltransferases Transfer of mono- or oligosaccharides to acce
nucleotide sugars and dolichol phosphate-mo
A large family of enzymes catalyzing the transfe
N-acetylgalactosamine, N-acetylmuramic acid
Also includes transfer of sulfate, phosphate, a
Glycosidases Catalyze the hydrolytic cleavage of the glycos
cellular processes
Over 100 families identified
Polysaccharide lyases Cleave polysaccharides containing uronic acid
cleavage site
Nine families identified
Carbohydrate esterases Remove acetyl groups from polysaccharides,
glycan linked Sias
16 or more families identified
Carbohydrate-binding
modules
Noncatalytic proteins, that bind soluble and c
catalytic domains in carbohydrate-active enzy
Found in glycoside hydrolases, polysaccharid
wall expansins
14 families identified
Glycan-binding protein
(GBP) molecules
GBPs enable binding of the carbohydrate-activ
enzyme and enhancing substrate degradation
Examples of proteins containing GBPs: C-type
collectins, selectins, dectins, mannose recepto
Sugar transporters Transport of sugars across the plasma memb
Hexose transporters
Energy-independent, facilitated diffusion, gluc
mammals.
Energy dependent; e.g., sodium-dependent gl
aA brief overview of eukaryote proteins that interact with carbohydrates. More detail can
and on The Consortium for Functional Glycomics website (http://www.fuctionalglyc
databases.
352
shows examples of the major groups of proteins that ma-nipulate glycoconjugates, highlighting the broad range ofevents that are involved in glycobiological metabolism.
Initial consideration of glycan structures must includethe metabolic pathways that lead to their formation. Themonosaccharides that form the building blocks are derivedfrom diverse sources. Some are obtained from extracellularorigins via salvage pathways from both external and in-tracellular sources, together with the major intracellularglycolytic pathways that feed monosaccharides into glycansynthesis and degradation.
A series of interlinked metabolic pathways produces theprecursors of the nucleotide sugars, while sugar transpor-ters facilitate the transport of glucose across the plasmamembrane [34] in either an energy-independent (facilitateddiffusion transporters, the GLUT family), or an energy- andsodium-dependent way (SGLT family). Monosaccharidesare activated to high-energy nucleotide sugars (UDP-Glc,UDP-Gal, UDP-GlcNAc, UDP-GalNAc, UDP-GlcA, UDP-Xyl, GDP-Man, GDP-Fuc, and CMP-NeuAc), which act asuniversal donors in glycosyltransferase reactions for eachglycosynthetic pathway [39,40]. Glycosulfotransferases uti-lize 30-phosphoadenosyl-50-phosphosulfate (PAPS) as theactive sulfate donor. Phosphorylated sugars, such as glu-cose-6-phosphate, mannose-6-phosphate, and glucosamine-6-phosphate, are similarly formed through the action ofkinases with ATP as the phosphate donor [39,40].
Pathways leading to the nucleotide sugars are regulatedthrough feedback inhibition of the key enzymatic stepsinitiating each route [39]. The entry of glucose into the
eukaryotesa
ptors, mono- and oligosaccharides, proteins, lipids, and DNA using
no and oligosaccharides
r of fucose, mannose, glucose, galactose, xylose, N-acetylglucosamine,
s, glucuronic acid, and Sias
cetyl, methyl, pyruvate, and ethanolamine to glycans
idic bond in all glycoconjugates; function in biosynthetic and catabolic
through a b-elimination mechanism forming a new reducing end at the
including pectin, xylan, galactoglucomannan, rhamnogalacturan and
rystalline carbohydrates; usually present in proteins together with
mes
e lyases, polysaccharide oxidases, glycosyltransferases, and plant cell
e enzyme to its substrate, thus increasing the local concentration of the
lectins, proteoglycan core proteins, Type II membrane receptors,
rs, layilin, galectins, and siglecs
rane into cells
ose transporters (GLUT protein family; SLC2 gene) in yeast and
ucose transporters (SGLT proteins; SLC5A gene)
be found on the CAZy website (http://www.cazy.org), at http://www.cazypedia.org
omics.org). All three sites provide further links to other carbohydrate-relevant

Table 2. Main types of glycan structurea,b
Glycan group Glycan structure
Proteoglycans
Hyaluronan
Glycoproteins
(N-glycans)
Mannose-6-phosphate glycans
Glycoproteins
(O-glycans)
Glycoproteins
(O-GlcNAcylation)
Review Trends in Biochemical Sciences July 2015, Vol. 40, No. 7
pathways leading to the formation of the N-acetylhexosa-mines is regulated by glucosamine-6-phosphate synthase,which is feedback inhibited by the target nucleotide sugarUDP-GlcNAc [41,42]. This same nucleotide sugar is fur-ther metabolized to yield Sias and CMP-b-O-Sias. Thispathway is subject to inhibition at the level of the initialenzyme UDP-GlcNAc epimerase/kinase [43]. Further inte-gration of glycosylation with monosaccharide biologyrequires the presence of sugar transporters. The transportof monosaccharides and activated sugars as nucleotidephosphate or dolichol phosphate into the cell is a signifi-cant event, enabling optimal efficiency of glycosylationprocesses in general.
In addition to de novo biosynthetic routes, salvage path-ways add monosaccharides released from glycoconjugatesduring normal cellular turnover. Most are released duringlysosomal degradation. The impact of these pathwaysdepends on the organ and tissue. The liver is a major siteof glycoconjugate degradation and the recovery of mono-saccharide products makes a significant contribution to thebiosynthetic potential of new glycoconjugates. Integrationof these pathways and their valuable monosaccharideproducts between tissues and organs constitutes a centralfacet of whole-body eukaryotic glycobiology. Examples ofthe variety of glycan structures found in eukaryotes arelisted in Table 2.
Molecular mimicry
Natural and synthetic mechanisms utilize glycan struc-tures to mimic a variety of cellular structures. Pathogenicorganisms synthesize antigens that are analogs of natu-rally occurring biotargets and, in this way, induce cellularresponses that enhance the survival of the pathogen, butdisadvantage the host [44]. Glycoengineering can be usedin the same manner to create glycotopes that can be usedtherapeutically. Currently, there is keen interest in theapplication of glycoengineering in recombinant glycopro-tein technology. Correct glycosylation for optimal biologi-cal relevance is paramount in such research [45–48].
N-glycans
The N-glycan class represents a range of glycans found inb-N-glycosidic linkage of an N-acetyl-D-glucosamine resi-due with a peptide and/or protein asparagine. Targeting ofN-glycans to appropriate asparagine residues in proteins isachieved through the recognition of a tri-amino acid sequon:asparagine-X-serine/threonine. ‘X’ may be any other aminoacid, barring proline. There are several common features ofthe N-glycans: first, the core comprises Mana1-6(Mana1-3)Manb1-4GlcNAcb1-4GlcNAcb1Asn-X-Ser/Thr and gener-ates two antennae. Second, the core is extended to formthree groups of N-glycan: oligomannose forms only containmannose; complex type have sialylated N-acetylactosaminetrisaccharides on each antenna and a fucose on the GlcNAclinked to asparagine; and hybrid types have a combinationof oligomannose and complex units on the Mana1-6 andMana1-3 antennae, respectively.
The process of protein N-glycosylation is conservedacross eukaryotes. The precursor glycan is assembled bythe stepwise glycosyl transfer of the individual sugarswhile attached to a lipid carrier (dolichol pyrophosphate)
353

Table 2 (Continued )
Glycan group Glycan structure
Glycoproteins
(GPI) anchor)
Glycoproteins
(C-mannose)
Glycosphingolipids
aExamples of the general types of glycan, largely drawn from animal examples,
are shown. Key: yellow circles, D-galactose; yellow squares, N-acetyl-D-galactos-
amine; blue circles, D-glucose; blue squares, N-acetyl-D-glucosamine; blue/white
squares, D-glucosamine; green circles, D-mannose; red triangles, L-fucose; purple
diamonds, N-acetyl-D-neuraminic acid (Neu5Ac); light blue diamonds, N-glycolyl-
D-neuraminic acid (Neu5Gc); blue/white diamonds D-glucuronic acid; orange/
white diamonds, L-iduronic acid; orange stars, D-xylose; white diamonds, myo-
inositol. All glycosidic linkages are shown as a or b, with the corresponding
position; for example, b4, b1-4 linkage.
bAbbreviations: 2S, 2-O-sulfate; 3S, 3-O-sulfate; 4S, 4-O-sulfate; 6S, 6-O-sulfate;
2P, 2-O-phosphate; 6P, 6-O-phosphate; Asn, asparagine; CH2CH2NH2, ethanol-
amine; FA, fatty acid, predominantly palmitate; Hyd, hydroxylysine; Hyp, hy-
droxyproline; NS, N-sulfate; Tryp, tryptophan; R, various glycan substitutions
occur at the initial mannose in GPI anchors.
Review Trends in Biochemical Sciences July 2015, Vol. 40, No. 7
on the cytoplasmic face of the endoplasmic reticulum (ER).Addition of the first sugar, GlcNAc, to dolichol phosphateinvolves transfer of a phosphate group and the sugar toform dolichyl pyrophosphate-GlcNAc. This product is ex-tended to yield dolichyl pyrophosphate-GlcNAc2Man5 be-fore being translocated to the lumenal surface of the ERmembrane by a flippase. Here, further mannose residuesare transferred and the Mana1-3 antenna is elongatedwith two mannoses terminated by a triglucosyl unitGlca1-2Glca1-3Glca1-3-. These glycosylation steps usedolichol-P-mannose and dolichol-P-glucose as donors rath-er than UDP-Glc and GDP-Man, the corresponding nucle-otide sugar donors, which are substrates for the glycosyltransfer reactions on the cytoplasmic side of the ER. Thefinal dolichol-linked glycan, Glc3Man9GlcNAc2-, is thenrecognized by an oligosaccharyltransferase in the ER lu-men that mediates attachment to protein asparagines toyield the b-N-glycosidic linkage [49,50]. This glycan servesas a substrate for trimming and processing reactions toyield the range of N-glycans found in nature. The biosyn-thetic pathway is found in all metazoans, plants, and fungi.
354
The trimming and processing events occur in the ERand have many features that are conserved in eukar-yotes. These link with crucial regulatory functions, nota-bly the correct folding of the glycoprotein to ensureoptimal biological activity. The folding mechanism isgoverned by the chaperones calnexin and calreticulin,which regulate the exit of glycoproteins from the ER.This cycle of events involves the monitoring of the glyco-protein after trimming of the triglucosyl unit by a-gluco-sidases I and II. If the glycoprotein is incompletely folded,an alpha-glucosyl-glycoprotein glucotransferase acts onthe terminal a1-2 linked mannose on the a1-3 mannosylantenna. The glycoprotein is then recycled through thecalnexin/calreticulin complex to reassess the folding effi-ciency. Glycoproteins that fail to fold properly are elimi-nated after translocation to the ER cytoplasm, where theN-glycans are removed and the protein is degraded in aseries of events termed ‘ER-associated degradation’(ERAD) [49,51]. Correctly folded glycoproteins are fur-ther trimmed and glycosylated to generate the three mainclasses of N-glycan in addition to the mannose-6-phos-phate-terminated N-glycans, which bind to the Man-6-P-receptor and are delivered to endosomes and lysosomes.This series of events occurs in the Golgi apparatus.Distinct patterns of peripheral N-glycosylation are foundin different phyla from plants and invertebrates to mam-mals [49,51,52].
O-glycans
O-glycosylation has also been termed ‘mucin-type O-glyco-sylation’. Mucins represent the major examples of glyco-proteins that carry this type of glycan. Glycans areattached to the protein backbone through a-linkage ofN-acetyl-D-galactosamine to the side-arm hydroxyl groupsof serine and threonine residues. The addition of theGalNAc residue to generate the initial residue of mucin-type glycans, in addition to proteins that carry singleGalNAc moieties, is catalyzed by a family of GalNActransferases that has been extensively researched by thegroup of H. Clausen [53]. The mucins are a family ofglycoproteins having serine-threonine-proline-rich vari-able number tandem repeat (VNTR) domains, which arethe sites of multiple O-glycan substitution. They are foundcommonly at mucosal surfaces in mammals, where theyhave a protective role [1]. The O-glycosylation patternsfound in the different mucins have formed the basis forreview of the range of O-glycan structures and O-glycosyl-ation pathways [1,40,54]. In addition to mucins, manyother glycoproteins have peptide domains with mucin-likeproperties, but without the VNTR patterns that define themucins themselves. The glycosylation of these domainsfollows the same biosynthetic routes reported for themucins.
The mucins have a series of core structures that form abase for extended glycans. These show linear Galb1-3Gal-NAc- (core 1) and GlcNAcb1-3GalNAc- (core 3) structuresand branched Galb1-3(GlcNAcb1-6)GalNAc- (core 2) andGlcNAcb1-3(GlcNAcb1-6)GalNAc- (core 4) structures. Ex-tension may occur via the addition of backbone repeat unitsof N-acetyllactosamine types 1 and 2 (Galb1-3GlcNAc- andGalb1-4GlcNAc-, respectively), or as the branched I and

Ser/Thr- polypep�de
Core 3Core 1
Core 2 Core 4
β4 β4 β6
α3 β3
α3
α3
β3β6
β4
β3β6
α-Ser /Thr α-Ser /Thr
α-Ser /Thr
β6
β6β3
β3
β3α3α-Ser /Thr
α-Ser /Thr β3
α6
α-Ser /Thr
α-Ser /Thr
α-Ser /Thrβ3
α-Ser /Thr α-Ser /Thr
TiBS
Figure 2. The biosynthetic pathways for the formation of a linear and branched
mucin-type O-glycans. Steps leading to the formation of the core 1–4 di- and
trisaccharides are shown. Enzymatic synthesis of two core 1-based sialylated
glycans is shown, linked by black arrows, to illustrate some of the routes to
individual glycans. Possible extensions of the core 3 and 4 structures are indicated
by red arrows. Key: yellow circles, D-galactose; yellow squares, N-acetyl-D-
galactosamine; blue circles, D-glucose; blue squares, N-acetyl-D-glucosamine;
blue/white squares, D-glucosamine; green circles, D-mannose; red triangles, L-
fucose; purple diamonds, N-acetyl-D-neuraminic acid (Neu5Ac); light blue
diamonds, N-glycolyl-D-neuraminic acid (Neu5Gc); blue/white diamonds D-
glucuronic acid; orange/white diamonds, L-iduronic acid; orange stars, D-xylose;
white diamonds, myo-inositol.
Review Trends in Biochemical Sciences July 2015, Vol. 40, No. 7
linear i antigens (Galb1-4GlcNAcb1-3Galb1-4-). Subse-quent peripheral substitution leads to complex oligosac-charides with ABO and Lewis blood group structures and arange of sialylated, fucosylated, and sulfated glycans[1,40,54]. The biosynthetic pathways leading to the forma-tion of the mucin type O-glycans are well known, and anexample is shown in Figure 2.
A large group of nuclear and cytoplasmic proteins carrya single sugar, O-b-N-acetyl-D-glucosamine (O-b-GlcNAc),attached to serine or threonine hydroxyl residues[55,56]. Most of these proteins have multiple O-b-GlcNAcadditions at different sites. The common serine or threo-nine sites identified for O-b-GlcNAc substitution are alsocommon sites for O-phosphorylation. Alternate O-b-GlcNAcylation and phosphorylation has been identifiedfor several nuclear and cytoplasmic proteins. This modifi-cation is never extended and, furthermore, is a dynamicglycosylation event, with cyclic patterns mediated by twospecific enzymes, an O-GlcNAc transferase (OGT) and anN-acetyl-D- glucosaminidase (OGlcNAcase). The latterrecognizes and removes the protein-bound O-GlcNAc. Thispost-translational modification is abundant and commonto all metazoans [55,56].
Several other O-glycans exist and, although these maynot be as abundant or widely distributed as the mucins,they all have significant biological relevance.
O-Mannosylation comprises a range of glycans withmannose alpha linked to serine or threonine found inmetazoans [57,58]. Major sites of expression are skeletalmuscle and the nervous system, particularly the brain.Most structures reported relate to Neu5Aca2-3Galb1-4GlcNAcb1-2ManaSer/Thr, which is the major glycan ina-dystroglycan, a skeletal muscle glycoprotein. However,other glycans have been found, including fucosylated andglucuronic acid-3-sulfated forms and branched chains[57]. The Drosophila melanogaster O-glycan repertoirecontains two genes encoding O-mannosyltransferase
(POMT1 and POMT2), which catalyze this reaction. O-b-glucose and O-a-fucose are both found in the epidermalgrowth factor (EGF)-like repeats of Notch and Cripto/FRL/Critic proteins. These monosaccharides are linked to ser-ine or threonine.
O-fucosylation shows a consensus sequence C2X4-5(Ser/Thr)C3, with C2 and C3 being the second and third con-served cysteines in the EGF-like repeats. Two specificprotein-O-fucosyltransferases (POFUT-I and POFUT-II)are responsible for this transfer, with GDP fucose as thedonor substrate [59]. POFUT-I catalyzes transfer to EGFdomains, while POFUT-II shows specificity for thrombos-pondin type-1 repeats [60]. The largest structure found is atetrasaccharide, Neu5Aca2-3/6Galb1-4GlcNAcb1-3Fuc-a-O-Ser/Thr. Urokinase, factor XII, Cripto factor IX, Throm-bospondin type-1 repeats, Notch, Delta, and Serrate,among others, have EGF-like repeats with both singleO-fucose and tetrasaccharide glycosylation [59,60].
O-b-glucosylation occurs at serine and threonine resi-dues in the EGF-like repeats, with the consensus sequenceC1XSerXPC2 distinct from the O-fucosylation sites. Themost common glycan identified is a trisaccharide with twoxylose residues (Xyla1-3Xyla1-3Glcb-O-). Some EGF-likerepeats also have single O-b-glucose additions, whereasfew proteins show this modification: only factor VII, factorIX, and Notch [61,62].
O-b-galactosylation of collagen has been known for sometime. This modification occurs together with the creation ofhydroxylysine and hydroxyproline in the collagen proteinand the addition of a disaccharide Glca1-2Galb-O-Hyl/Hyp[63].
C-mannosylation
This modification entails the attachment of a single man-nose residue, in a C-linkage, to the indole ring of trypto-phan. It is found in most eukaryotes, except yeasts, and isabsent in bacteria. A peptide consensus sequence, W-X-X-W, carries the mannose residue on the initial tryptophan[64,65]. It also occurs in the CYS domains of MUC5AC andMUC5B [65]. The biosynthesis involves dolichol-phos-phate-mannose as the donor [64]. C-mannosylation isthought to have a role in protein folding.
GPI anchors
GPI anchors are a common means of tethering proteins,through their carboxyl terminal, to the outer leaf of the cellmembrane lipid bilayer, and presenting them to the exter-nal environment. GPI anchors are found throughouteukaryotes, including protozoa, plants, fungi, inverte-brates, and mammals [66]. The membrane-mediated bio-synthesis of GPI anchors is complex, but proceeds throughconventional pathways and involves ten steps to yield amembrane GPI anchor-linked protein. A final step cleavesthe inositol-bound palmitate from the anchor and enablesphosphatidylinositol phospholipase C (PI-PLC) to releasethe protein from the membrane [66].
The general structure of the GPI anchors includes acommon core comprising ethanolamine-phosphate-6Ma-na1-2Mana1-6Mana1-4GlcNa1-6myo-inositol-1-phosphate-lipid. Proteins are attached to the amino group of the etha-nolamine through their C-terminal carboxyl groups.
355

Table 3. Variety of Sias in eukaryotes
Sia feature Examples
Chemical
modifications
Mono-O-acetylation at positions 4, 7, 8, and 9;
di-O-acetyl at 4,9, 7,9, 7,8, and 8,9; and possibly
tri –O-acetyl form at 7,8,9.
O-sulfate at position 8
O-methyl at position 8
O-lactyl at position 9
O-phosphate at position 9
Dehydro forms, with double bond in ring at position 2-3
Lactones and lactams
Glycosidic
linkage
a2-3, a2-6 and a2-8 are most common
Adjacent
sugars as
a carrying
glycan
Glycan type N-linked; O-linked; glycosphingolipid; and polysaccharide
Cellular
organisation
Cell surface plasma membranes; glycocalyx;
glycosynapse
Subcellular membranes: rough ER; Golgi apparatus;
secretory granules; endosomes; and lysosomes
Subcellular compartments: cytoplasm; nucleus;
lysosomes; endosomes; and secretory granules
The Sias exist in nature as terminal sugars in many glycans and also in mono-
saccharide form as their dehydro analogs. A range of molecular characteristics
leads to the abundance and variety of Sias. Key: yellow circles, D-galactose;
yellow squares, N-acetyl-D-galactosamine; blue circles, D-glucose; blue squares,
N-acetyl-D-glucosamine; blue/white squares, D-glucosamine; green circles, D-
mannose; red triangles, L-fucose; purple diamonds, N-acetyl-D-neuraminic acid
(Neu5Ac); light blue diamonds, N-glycolyl-D-neuraminic acid (Neu5Gc); blue/
white diamonds D-glucuronic acid; orange/white diamonds, L-iduronic acid;
orange stars, D-xylose; white diamonds, myo-inositol.
Review Trends in Biochemical Sciences July 2015, Vol. 40, No. 7
356
Different GPI anchors arise through substitutions to thiscore structure. Commonly found additions are the attach-ment of: (i) alkyl or alkenyl groups to the lipid terminus; (ii)palmitate to the C2 of myo-inositol; (iii) ethanolamine phos-phate on C2 of Mana1-4 or C6 of Mana1-6; or (iv) glycans oneach of the core mannoses. GPI-tethered proteins may bereleased from the plasma membrane by the action of PI-PLC:this property has significant biological relevance in cellsurface protein expression. The substitution of palmitateon the C2 of myo-inositol blocks the action of PI-PLC andprovides a significant regulatory modification. Many pro-teins with GPI anchors have been identified and the biologi-cal impact of this modification is evident in examples whereboth GPI-anchor tethered and soluble forms exist, relating totissue distribution or developmental programmes; see [66]for a summary of examples.
Sias
The Sias represent a family of over 50 derivatives withunique diversity, wide occurrence, and miscellaneous bio-logical functions [67]. They are not found in plants, pro-karyotes, arthropods, and most invertebrates. An overviewof the molecular and biological features of the Sias is givenin Table 3.
The Echinodermata are the earliest clade identified tohave Sias of various types [67,68]. The mammals express aplethora of Sias whose biology has attracted a large body ofresearch.
Sia is a nine-carbon monosaccharide, comprising a six-membered ring with a three-carbon C7-C9 glycerol chainattached at ring carbon 6. Chemically, this is 5-acetamido-2-keto-3,5-dideoxy-D-glycero-D-galactononic acid, an un-wieldy and impractical name that is effectively shortenedto Sia, and abbreviated as ‘Neu5Ac’ or ‘Sia’ as a generalform. A carboxyl group is attached at ring carbon 2 and anamino group is substituted on ring carbon 5. The aminogroup exists as acetyl (Neu5Ac) and glycolyl (Neu5Gc)forms. These two Sias are parent forms and both acceptall of the modifications found in this family of monosac-charides. In brief, O-acetylation may occur at positions 4, 7,8, and 9; O-methyl and O-sulfate at position 8, and O-lactyl,phosphate, and O-sulfate at position 9. The glycosidichydroxyl at position 2 is in the a or b configuration forthe free monosaccharide and forms the basis of glycosidiclinkage to other sugars. Thus, a2-3, a2-6, and a2-8 (to otherSias) are common structures. Monosaccharide anhydroforms also exist, where a double bond is formed betweenC2 and C3, yielding, for example, Neu2en5Ac [67].
Biosynthesis of the Sias is initiated de novo from UDP-GlcNAc by the action of the UDP-GlcNAc epimerase/kinasecomplex and culminates in the formation of CMP-b-O-Neu5Ac. The de novo pathway also connects with salvagepathways, where Neu5Gc may be derived from the diet,and progresses through the activation mechanisms togenerate CMP-b-O-Neu5Gc. The CMP-b-O-Sias are thedonors in the sialyltransferase reactions leading to theformation of the library of sialoglycoconjugates found innature. In line with the broad range of potential acceptors,the sialyltransferase family mediates the biosynthesis ofglycoconjugates in the Golgi. Some eukaryotic sialyltrans-ferases exhibit substrate specificity towards different

Review Trends in Biochemical Sciences July 2015, Vol. 40, No. 7
acceptors, glycoproteins, gangliosides (glycolipids), andpolysaccharides, including polysialic acid chains. Thesetransferases contain peptide sequences termed ‘sialylmo-tifs’ and are found in most eukaryote enzymes, apart fromthose in bacteria, suggesting different evolutionary routes[69]. Several Sia-related monosaccharides, such as 2-keto-3-deoxynonic acid (Kdn), are found at lower levels in mosteukaryotes. Transfer of these analogs is also catalyzedeffectively by the sialyltransferases. A novel form of sialyl-transfer has been identified in the protozoa: trypanosomeshave a trans-sialidase, which recognizes cell surface-boundSias, cleaves them, and transfers them to their own cellsurface acceptors to evade host immune screening [70].
The presence of a CMP-b-O-sialate transporter in theGolgi apparatus, but not in the ER, confirms the specificityof subcellular sialylation in eukaryotic cells [39].
In nonhuman species, the conversion of Neu5Ac toNeu5Gc is catalyzed by a specific CMP-b-O-Neu5Ac hy-droxylase, encoded by the CMAH gene, which has beeninactivated in humans [67]. O-acetylation of the 4-O andC7-C9-O positions is carried out enzymatically. Evidencefor enzymatic O-acetyl transfer to a single position withsubsequent migration of the acetyl esters to the nonoccu-pied positions has been proposed [67]. Enzymatic transferof methyl groups by S-adenosylmethionine and sulfate byPAPS has been confirmed [67], but the mechanism oflactosylation has not been established.
Sialidases (neuraminidases) that release a-linked Siasfrom sialoglycoconjugates are known as NEU enzymes(NEU1–4). These show specificity for the type of sialogly-coconjugate and also have a characteristic tissue and cellexpression pattern. By contrast, other eukaryotes, such asthe fungi and invertebrates, have sialidases with broadersubstrate specificities, reflecting the strategy required tomanipulate the sialoglycoconjugates encountered in theirenvironment. Transport of Sias out of the lysosomes to thecytoplasm requires the action of sialin, a Sia exporter, butno plasma membrane-located Sia transporter has beenidentified so far.
The only Sia found in beta-linkage is CMPb-O-Sia. Thissialoside can be cleaved enzymatically by CMP-b-O-sialatehydrolase, a b-sialidase. Although this enzyme has beendetected, there is only limited information concerning suchactivity and this remains an area for further study relativeto the regulation of the sialome. The removal of Sia O-acetyl esters is governed by sialate O-acetyl esterases,which have selective or broad specificities for the site ofO-acetylation. Release of 4-O-acetyl, but not C7-C9, ormono 9-O-acetyl, but not 7,9-di-O-acetyl implies roles fordifferent sialate O-acetylation patterns.
In accord with the range of sialoglycoconjugates andtheir sialylation patterns, Sia-binding lectins have beenidentified from many sources and are features of glyco-biology throughout the eukaryotes. The vertebrates havetwo main classes of lectin: (i) the C-type lectins, theselectins; and (ii) the I-type lectins, the siglecs. Theseproteins have important roles in cell–cell recognition pro-cesses. Accordingly, the glycans that carry the Sias have arole in recognition. Furthermore, the various modifica-tions of the Sias, especially O-acetylation, may mediatethe interactions between cells [67]. The specific binding
properties of these lectins have led to their use as tools inmonitoring Sia expression and events facilitated by Sias[71–73].
Glycosphingolipids and glycosaminoglycans are de-scribed in other reviews in this Special Issue and so arenot discussed here.
Evolutionary variation of glycansThis final section emphasizes some of the main issues thatrelate to the study of glycans. These are essentially generalcomments drawn from the glycobiological literature andrelate to evolutionary aspects of this work.
General diversity
Monosaccharides are particularly well suited to con-struct a range of different sequences, due to their ringand open chain patterns with equatorial and axial ar-rangement of the hydroxyl groups. In addition, theanomeric hydroxyl group enables the formation of alphaand beta glycosides for each monosaccharide. Thus,when the complete range of glycan structures found inthe eukaryotes is screened, the initial impression is oneof enormous scope and diversity. Closer examinationshows that, although many monosaccharides exist chem-ically, only a relatively small selection of the total hasbeen adopted to build biological sequences, and thisreflects evolutionary selection.
It is striking how many glycan structures are shared bydifferent groups of eukaryote. This can be explained by theevolution of proteins that control glycan metabolism. Theglycosyltransferases, glycosyl hydrolases, sugar transpor-ters, and lectins are organized in each tissue and cell todeliver the required glycosylation of the glycoconjugates,providing the structural and functional needs in each case.All of the families of glycans are represented in eukaryotes:N-glycans, the different O-glycans, C-mannose, glycolipids,and glycosaminoglycans. Each glycan type exhibits a coreunit that is common to all. Extension of these core unitsalso follows shared processes. However, completion of theglycans through peripheral additions results in strain- andphylum-specific structures. These are the moieties thatface the outer world.
Natural forces powering glycan evolution
The glycan diversity found in individual environments isformed through known metabolic pathways. Therefore,many features of glycan structure influence phenotypicvariation and serve to emphasize the flexibility of utilizingsugar-based sequences [18,23,37,38,51,54,74–78]. The gly-cobiological process is powerful because it enables a varietyof possibilities to be selected in exclusive and appropriateconditions. Evolutionary analysis shows that unicellularorganisms create glycans from a range of monosaccharides,while the eukaryotes use a smaller number [79]. Structuralselection occurs by mutation or through exclusion path-ways [80]. The basis of glycan structure and recognition atchemical and biochemical levels is robust. Glycoforms aregenerated through the action of the glycosyltransferasesand degraded by the glycosidases. The pattern of expres-sion of these enzymes, combined with their spatial andtemporal properties, leads to a predictable range of glycan
357

Review Trends in Biochemical Sciences July 2015, Vol. 40, No. 7
structures, which can be matched with those detected bychemical methods in vivo. A further level of recognitionarises due to spatial clustering and ligand density.
Concluding remarksGlycans are generated through the action of glycosyltrans-ferases and degraded by glycosidases. The pattern of ex-pression of these enzymes, combined with their spatial andtemporal properties, leads to a predictable and dynamicrange of glycan structures, which share common features ofmonosaccharide composition, sequence, and conformation,and also confer cell, tissue, and clade specificity. Recogni-tion relies on not only ligand presence, but also liganddensity and its spatial organization; that is, clustering.These molecular patterns form a basis for the recognitionof self and non-self.
AcknowledgmentsWe wish to thank Hans-Joachim Gabius for his guidance and encourage-ment during the preparation of this review. We are also grateful to PascalGagneux and P.T. Pollard for permission to reproduce the phylogenetictree in Figure 1. Due to editorial constraints, it is not possible to cite all ofthe relevant literature linked with the broad topic of eukaryoteglycosylation. Therefore, we acknowledge the contributions made byauthors whose work it was not possible to reference here.
Appendix A. Supplementary dataSupplementary data associated with this article can be found, in the onlineversion, at http://dx.doi.org/10.1016/j.tibs.2015.04.004.
References1 Corfield, A.P. (2015) Mucins: a biologically relevant glycan barrier in
mucosal protection. Biochim. Biophys. Acta 1850, 236–2522 Linden, S.K. et al. (2008) Mucins in the mucosal barrier to infection.
Nat. Mucosal Immunol. 1, 183–1973 Solis, D. et al. (2015) A guide into glycosciences: how chemistry,
biochemistry and biology cooperate to crack the sugar code.Biochim. Biophys. Acta 1850, 186–235
4 Gabius, H-J. (2015) The magic of the sugar code. Trends Biochem. Sci.40, 341
5 Tran, D.T. and Ten Hagen, K.G. (2013) Mucin-type O-glycosylationduring development. J. Biol. Chem. 288, 6921–6929
6 Bartling, B. et al. (2009) Age-associated changes of extracellularmatrix collagen impair lung cancer cell migration. FASEB J. 23,1510–1520
7 Bassaganas, S. et al. (2014) Cell surface sialic acid modulatesextracellular matrix adhesion and migration in pancreaticadenocarcinoma cells. Pancreas 43, 109–117
8 Styriak, I. et al. (2003) Binding of extracellular matrix molecules byprobiotic bacteria. Lett. Appl. Microbiol. 37, 329–333
9 Zhang, L. and Ten Hagen, K.G. (2011) The cellular microenvironmentand cell adhesion: a role for O-glycosylation. Biochem. Soc. Trans. 39,378–382
10 Cortes, P.P. et al. (2004) Sperm binding to oviductal epithelial cellsin the rat: role of sialic acid residues on the epithelial surface andsialic acid-binding sites on the sperm surface. Biol. Reprod. 71,1262–1269
11 Albenberg, L.G. et al. (2012) Food and the gut microbiota ininflammatory bowel diseases: a critical connection. Curr. Opin.Gastroenterol. 28, 314–320
12 Alemka, A. et al. (2012) Defense and adaptation: the complex inter-relationship between Campylobacter jejuni and mucus. Front. Cell.Infect. Microbiol. 2, 15
13 Chow, J. et al. (2011) Pathobionts of the gastrointestinal microbiotaand inflammatory disease. Curr. Opin. Immunol. 23, 473–480
14 Feng, T. and Elson, C.O. (2011) Adaptive immunity in the host-microbiota dialog. Mucosal Immunol. 4, 5–21
358
15 Hajishengallis, G. et al. (2012) The keystone-pathogen hypothesis. Nat.Rev. Microbiol. 10, 717–725
16 Hormannsperger, G. et al. (2012) Gut matters: microbe-hostinteractions in allergic diseases. J. Allergy Clin. Immunol. 129,1452–1459
17 Cantarel, B.L. et al. (2009) The Carbohydrate-Active EnZymesdatabase (CAZy): an expert resource for glycogenomics. NucleicAcids Res. 37, D233–D238
18 Imperiali, B. (2012) The chemistry-glycobiology frontier. J. Am. Chem.Soc. 134, 17835–17839
19 Gabius, H.J. and Kayser, K. (2014) Introduction to glycopathology: theconcept, the tools and the perspectives. Diagn. Pathol. 9, 4
20 Tan, F. et al. (2015) Sweetening up the opposition: glycans in bacterial-host interaction. Trends Biochem. Sci. 40, 342–350
21 Hennet, T. and Cabalzar, J. (2015) Congenital disorders ofglycosylation: a concise chart of glycocalyx dysfunction. TrendsBiochem. Sci. 40, 377–384
22 Peracaula, R. et al. (2008) Altered glycosylation in tumours focused tocancer diagnosis. Dis. Markers 25, 207–218
23 Cummings, R.D. (2009) The repertoire of glycan determinants in thehuman glycome. Mol. Biosyst. 5, 1087–1104
24 Ju, T. et al. (2011) The Tn antigen-structural simplicity and biologicalcomplexity. Angew. Chem. Int. Ed Engl. 50, 1770–1791
25 Paulick, M.G. and Bertozzi, C.R. (2008) The glycosylphosphatidylinositolanchor: a complex membrane-anchoring structure for proteins.Biochemistry 47, 6991–7000
26 Varki, A. (2011) Evolutionary forces shaping the Golgi glycosylationmachinery: why cell surface glycans are universal to living cells. ColdSpring Harb. Perspect. Biol. 3, a005462
27 Bull, C. et al. (2014) Sweet escape: sialic acids in tumor immuneevasion. Biochim. Biophys. Acta 1846, 238–246
28 Aaltomaa, S. et al. (2002) Strong stromal hyaluronan expression isassociated with PSA recurrence in local prostate cancer. Urol. Int. 69,266–272
29 Anttila, M.A. et al. (2000) High levels of stromal hyaluronan predictpoor disease outcome in epithelial ovarian cancer. Cancer Res. 60,150–155
30 Auvinen, P. et al. (2014) Hyaluronan synthases (HAS1-3) in stromaland malignant cells correlate with breast cancer grade and predictpatient survival. Breast Cancer Res. Treat. 143, 277–286
31 Reuter, G. and Gabius, H.J. (1999) Eukaryotic glycosylation: whim ofnature or multipurpose tool? Cell. Mol. Life Sci. 55, 368–422
32 Schachter, H. (2010) Mgat1-dependent N-glycans are essential for thenormal development of both vertebrate and invertebrate metazoans.Semin. Cell Dev. Biol. 21, 609–615
33 Tabak, L.A. (2010) The role of mucin-type O-glycans in eukaryoticdevelopment. Semin. Cell Dev. Biol. 21, 616–621
34 Liu, L. et al. (2010) The role of nucleotide sugar transportersin development of eukaryotes. Semin. Cell Dev. Biol. 21,600–608
35 Dell, A. et al. (2010) Similarities and differences in the glycosylationmechanisms in prokaryotes and eukaryotes. Int. J. Microbiol. 2010,148178
36 Lauc, G. et al. (2014) Glycans – the third revolution in evolution. Front.Genet. 5, 145
37 Mengerink, K.J. and Vacquier, V.D. (2001) Glycobiology of sperm-egginteractions in deuterostomes. Glycobiology 11, 37R–43R
38 Varki, A. (2006) Nothing in glycobiology makes sense, except in thelight of evolution. Cell 126, 841–845
39 Freeze, H.H. and Elbein, A.D. (2009) Glycosylation precursors. InEssentials of Glycobiology (2nd edn) (Varki, A. et al., eds), pp. 47–61,Cold Spring Harbor Laboratory Press
40 Schachter, H. and Brockhausen, I. (1992) The biosynthesis of serine(threonine)-N-acetylgalactosamine-linked carbohydrate moieties. InGlycoconjugates (Allen, H.J. and Kisailus, E.C., eds), pp. 263–332,Marcel Dekker
41 Milewski, S. (2002) Glucosamine-6-phosphate synthase: the multi-facets enzyme. Biochim. Biophys. Acta 1597, 173–192
42 Miszkiel, A. et al. (2011) Long range molecular dynamics study ofregulation of eukaryotic glucosamine-6-phosphate synthase activity byUDP-GlcNAc. J. Mol. Model. 17, 3103–3115
43 Keppler, O.T. et al. (1999) UDP-GlcNAc 2-epimerase: a regulator of cellsurface sialic acid. Science 284, 1372–1376

Review Trends in Biochemical Sciences July 2015, Vol. 40, No. 7
44 Carlin, A.F. et al. (2009) Molecular mimicry of host sialylated glycansallows a bacterial pathogen to engage neutrophil Siglec-9 and dampenthe innate immune response. Blood 113, 3333–3336
45 Castilho, A. et al. (2013) Generation of biologically active multi-sialylated recombinant human EPOFc in plants. PLoS ONE 8, e54836
46 Loos, A. and Steinkellner, H. (2012) IgG-Fc glycoengineering in non-mammalian expression hosts. Arch. Biochem. Biophys. 526, 167–173
47 Piirainen, M.A. et al. (2014) Glycoengineering of yeasts from theperspective of glycosylation efficiency. Nat. Biotechnol. 31, 532–537
48 Toth, A.M. et al. (2014) A new insect cell glycoengineering approachprovides baculovirus-inducible glycogene expression and increaseshuman-type glycosylation efficiency. J. Biotechnol. 182–183, 19–29
49 Aebi, M. (2013) N-linked protein glycosylation in the ER. Biochim.Biophys. Acta 1833, 2430–2437
50 Kelleher, D.J. and Gilmore, R. (2006) An evolving view of theeukaryotic oligosaccharyltransferase. Glycobiology 16, 47R–62R
51 Freeze, H.H. et al. (2009) Glycans in glycoprotein quality control. InEssentials of Glycobiology (2nd edn) (Varki, A. et al., eds), pp. 513–521,Cold Spring Harbor Laboratory Press
52 Wilson, I.B. (2002) Glycosylation of proteins in plants andinvertebrates. Curr. Opin. Struct. Biol. 12, 569–577
53 Bennett, E.P. et al. (2012) Control of mucin-type O-glycosylation: aclassification of the polypeptide GalNAc-transferase gene family.Glycobiology 22, 736–756
54 Patsos, G. and Corfield, A. (2009) O-Glycosylation: structural diversityand functions. In The Sugar Code. Fundamentals of Glycoscience(Gabius, H-J., ed.), pp. 111–137, Wiley-VCH Verlag GmbH & Co
55 Butkinaree, C. et al. (2010) O-linked beta-N-acetylglucosamine (O-GlcNAc): extensive crosstalk with phosphorylation to regulatesignaling and transcription in response to nutrients and stress.Biochim. Biophys. Acta 1800, 96–106
56 Ma, J. and Hart, G.W. (2014) O-GlcNAc profiling: from proteins toproteomes. Clin. Proteomics 11, 8
57 Panin, V.M. and Wells, L. (2014) Protein O-mannosylation in metazoanorganisms. Curr. Protoc. Protein Sci. 75, 12
58 Lommel, M. and Strahl, S. (2009) Protein O-mannosylation: conservedfrom bacteria to humans. Glycobiology 19, 816–828
59 Takeuchi, H. and Haltiwanger, R.S. (2014) Significance of glycosylationin Notch signaling. Biochem. Biophys. Res. Commun. 453, 235–242
60 Freeze, H.H. and Haltiwanger, R.S. (2009) Other classes of ER/Golgi-derived glycans. In Essentials of Glycobiology (2nd edn) (Varki, A. et al.,eds), pp. 163–173, Cold Spring Harbor Laboratory Press
61 Gebauer, J.M. et al. (2008) O-glucosylation and O-fucosylation occurtogether in close proximity on the first epidermal growth factor repeatof AMACO (VWA2 protein). J. Biol. Chem. 283, 17846–17854
62 Takeuchi, H. et al. (2012) Site-specific O-glucosylation of the epidermalgrowth factor-like (EGF) repeats of notch: efficiency of glycosylation is
affected by proper folding and amino acid sequence of individual EGFrepeats. J. Biol. Chem. 287, 33934–33944
63 Schegg, B. et al. (2009) Core glycosylation of collagen is initiated by twobeta(1-O)galactosyltransferases. Mol. Cell. Biol. 29, 943–952
64 Furmanek, A. and Hofsteenge, J. (2000) Protein C-mannosylation:facts and questions. Acta Biochim. Pol. 47, 781–789
65 Perez-Vilar, J. et al. (2004) C-Mannosylation of MUC5AC and MUC5Bcys subdomains. Glycobiology 14, 325–337
66 Shams-Eldin, H. et al. (2009) Glycosylphosphatidylinositol anchors:structure, biosynthesis and functions. In The Sugar Code (Gabius, H-J.,ed.), pp. 155–173, Wiley-VCH Verlag GmbH & Co
67 Varki, A. and Schauer, R. (2009) Sialic acids. In Essentials ofGlycobiology (2nd edn) (Varki, A. et al., eds), pp. 199–217, ColdSpring Harbor Laboratory Press
68 Corfield, A.P. and Schauer, R. (1982) Occurrence of sialic acids. InSialic Acids. Chemistry, Metabolism and Function (Schauer, R., ed.),pp. 5–50, Springer-Verlag
69 Harduin-Lepers, A. et al. (2005) The animal sialyltransferases andsialyltransferase-related genes: a phylogenetic approach. Glycobiology15, 805–817
70 Schenkman, S. et al. (1994) Structural and functional properties ofTrypanosoma trans-sialidase. Annu. Rev. Microbiol. 48, 499–523
71 Sharon, N. (1993) Lectin-carbohydrate complexes of plants andanimals: an atomic view. Trends Biochem. Sci. 18, 221–226
72 Sharon, N. (1994) When lectin meets oligosaccharide. Nat. Struct. Biol.1, 843–845
73 Smits, S.L. et al. (2005) Nidovirus sialate-O-acetylesterases: evolutionand substrate specificity of coronaviral and toroviral receptor-destroying enzymes. J. Biol. Chem. 280, 6933–6941
74 Bertozzi, C.R. and Rabuka, D. (2009) Structural basis of glycandiversity. In Essentials of Glycobiology (2nd edn) (Varki, A. et al.,eds), pp. 23–36, Cold Spring Harbor Laboratory Press
75 Cohen, M. and Varki, A. (2010) The sialome: far more than the sum ofits parts. Omics 14, 455–464
76 Gagneux, P. and Varki, A. (1999) Evolutionary considerations inrelating oligosaccharide diversity to biological function. Glycobiology9, 747–755
77 Springer, S.A. and Gagneux, P. (2013) Glycan evolution in responseto coll-aboration, conflict, and constraint. J. Biol. Chem. 288,6904–6911
78 Varki, A. (1993) Biological roles of oligosaccharides: all of the theoriesare correct. Glycobiology 3, 97–130
79 Adibekian, A. et al. (2011) Comparative bioinformatics analysis of themammalian and bacterial glycomes. Chem. Sci. 2, 337–344
80 Sletten, E.M. and Bertozzi, C.R. (2009) Bioorthogonal chemistry:fishing for selectivity in a sea of functionality. Angew. Chem. Int. EdEngl. 48, 6974–6998
359

Genome Analyses Highlight the Different Biological Roles of Cellulases

Cellulose is the most abundant form of photosyntheti‑cally fixed carbon in the biosphere. It is a fibrous poly‑mer of glucose units that are linked by β‑1,4‑glycosidic bonds, and it occurs naturally in plants and in several other organisms, including certain bacteria, some fungi, protozoa (such as Naegleria gruberi1, Dictyostelium discoideum2 and Acanthamoeba castellanii 3) and in one group of animals, the urochordates or ‘tunicates’ (REF. 4). Natural cellulose chains are aggregated laterally by means of hydrogen bonding and van der Waals’ forces to generate microfibrils of parallel chains5 that crystal‑lize as two forms, Iα and Iβ6,7. Plant cellulose is further surrounded by a network of other polymers, including hemicelluloses, pectins and lignin8. Hemicelluloses are branched polysaccharides with a backbone of neutral sugars that form hydrogen bonds to the surface of cel‑lulose fibrils, whereas the major component of pectins is galacturonic acid. The simplest pectin is homogalactu‑ronan, an unbranched polymer of α‑1,4‑d‑galacturonic acid5, but some pectins can exhibit considerable chemi‑cal complexity, such as the rhamnogalacturonans, which display numerous and varied carbohydrate side chains. Lignin fills the space between the cellulose, hemi‑cellulose and pectin in the plant cell wall; it consists of a highly crosslinked matrix of aromatic residues.
The capacity to convert cellulose in complex plant cell wall substrates is of crucial importance for the carbon cycle9, reflecting the abundance of cellulose in nature. However, cellulose is notoriously difficult to hydrolyse enzymatically because it contains resilient glycosidic bonds10, is crystalline and is tightly associated with
other polysaccharides (as mentioned above). The com‑plete digestion of cellulose is therefore largely restricted to a specific group of cellulolytic microorganisms that produce complex combinations of enzymes (cellulases, hemicellulases and pectinases) which act synergisti‑cally to break down cellulose and its associated cell wall components. These enzymes belong to different sequence‑based families of glycoside hydrolases (GHs) — which include cellulases that cleave the β(1→4) bonds of cellulose, making them attractive candidates for the sustainable production of cellulosic ethanol from ligno‑cellulosic materials10–12 — carbohydrate‑binding mod‑ules (CBMs), polysaccharide lyases and carbo hydrate esterases, as classified in the Carbohydrate‑Active Enzyme database (CAZy)13,14. Cellulolytic bacteria have traditionally been searched for in a wide variety of envi‑ronmental and industrial niches, including soil, marine sediments and, more recently, the gut microbiota of ani‑mals and humans. Currently, the prevailing dogma is that cellulolytic bacteria live a saprophytic lifestyle.
However, the view that cellulases act only in the domain of plant cell wall degradation has recently been challenged by the demonstration that Mycobacterium tuberculosis, one of the pathogens responsible for ani‑mal and human tuberculosis, has three active cellulose‑targeting proteins, encoded by Rv0062 (also known as celA1 (REF. 15); belonging to family GH6), Rv1090 (also known as celA2b; belonging to family GH12) and Rv1987 (REFS 16,17) (belonging to family CBM2). Similarly, the pathogen Legionella pneumophila secretes an endoglucanase18 (belonging to family GH5). These
1Unité de Recherche sur les Maladies Infectieuses et Tropicales Emergentes, Centre National de la Recherche Scientifique (CNRS), Institut de Recherche pour le Développement, Faculté de Médecine, Aix-Marseille Université, 27 Bd Jean Moulin, 13005 Marseille, France.2Architecture et Fonction des Macromolécules Biologiques, CNRS, Aix-Marseille Université, 163 Avenue de Luminy, 13288 Marseille, France.3York Structural Biology Laboratory, Department of Chemistry, University of York, York YO10 5DD, UK.Correspondence to B.H. e-mail: [email protected]:10.1038/nrmicro2729Published online 23 January 2012
Genome analyses highlight the different biological roles of cellulasesFelix Mba Medie1,2, Gideon J. Davies3, Michel Drancourt1 and Bernard Henrissat2
Abstract | Cellulolytic enzymes have been the subject of renewed interest owing to their potential role in the conversion of plant lignocellulose to sustainable biofuels. An analysis of ~1,500 complete bacterial genomes, presented here, reveals that ~40% of the genomes of sequenced bacteria encode at least one cellulase gene. Most of the bacteria that encode cellulases are soil and marine saprophytes, many of which encode a range of enzymes for cellulose hydrolysis and also for the breakdown of the other constituents of plant cell walls (hemicelluloses and pectins). Intriguingly, cellulases are present in organisms that are usually considered as non-saprophytic, such as Mycobacterium tuberculosis, Legionella pneumophila, Yersinia pestis and even Escherichia coli. We also discuss newly emerging roles of cellulases in such non-saprophytic organisms.
Saprophytic lifestyleReferring to the lifestyle of an organism that feeds on dead organic matter of plant origin.
ANALYSIS
NATURE REVIEWS | MICROBIOLOGY VOLUME 10 | MARCH 2012 | 227
© 2012 Macmillan Publishers Limited. All rights reserved

Crystalline region Crystalline region
Lytic oxidasesa
b
Amorphous region Crystalline region Crystalline regionAmorphous region
Nature Reviews | Microbiology
Endoglucanases Cellobiohydrolases Cellobiose
Oxidized cellobiose
observations demonstrate that the analysis of complete genome sequences can reveal previously unconsidered metabolic traits and that the prevailing view that cellu‑lases are restricted to saprophytic organisms is incorrect. It is becoming clear that cellulases are also involved in cellulose biosynthesis, as originally described for the bacterium Gluconacetobacter xylinus (formerly known as Acetobacter xylinum)19 and perhaps best character‑ized subsequently for the plant protein KORRIGAN, which is required for the assembly of cellulose and hemicellulose networks in Arabidopsis thaliana20. In this Analysis article, we review some of the more unusual and indeed counter‑intuitive roles for cellulases that are now emerging from wide‑scale analyses of bacterial genomes.
Cellulose and its enzymatic breakdownTwo main types of cellulase enzyme activity are well char‑acterized. Endoglucanases (ENZYME entry EC 3.2.1.4) hydrolyse internal bonds at random positions of the less ordered (or amorphous) regions of cellulose. These enzymes generate chain ends for the processive action of the second type of cellulases, the cellobiohydrolases (which are exoglucanases). These act in a unidirectional man‑ner from either the non‑reducing (ENZYME entry EC 3.2.1.91) or the reducing (ENZYME entry EC 3.2.1.176) ends of cellulose polysaccharide chains, liberating cello‑biose as the major product. β‑d‑glucosidases (ENZYME entry EC 3.2.1.21) further hydrolyse cellobiose and thus relieve the system from end product inhibition10. Various
types of synergies have been observed between different sorts of cellulase during the breakdown of crystal line cellulose21, but convincing explanations at the molecular level have yet to be found, especially for the cooperation between cellobiohydrolases. The cooperation between endo‑ and exo‑acting cellulases is intuitively easier to understand, but the degree of synergy observed is usually less than that expected for a system in which the cello biohydrolase depends entirely on the endo‑glucanase for the provision of free chain ends. In an illustration of the poor understanding of the role of cellobiohydrolases, a recent investigation has shown that the two major cellobiohydrolases of Clostridium thermocellum can be deleted without severely affecting cellulose breakdown22.
Other enzymes besides the endo‑acting and exo‑acting cellulases participate in the breakdown of cel‑lulose. Some of these enzymes use oxidative chemistry, as occurs in the cleavage of chitin (a fibrous, crystalline polymer of β‑1,4‑N‑acetylglucosamine residues) by the bacterial enzyme chitin‑binding protein 21 (Cbp21); this mode opens up the crystalline polysaccharide material that is inaccessible for hydrolysis by other GHs. Furthermore, the reductant ascorbic acid boosts the efficiency of Cbp21 (REF. 23). Recently, a Streptomyces coelicolor homologue of Cbp21 was shown to act syner‑gistically with cellulases in the digestion of cellulose24. Similarly, GH61 enzymes25, which have structural and active centre similarity to Cbp21 and its homologues, also facilitate the degradation of cellulose through an
Figure 1 | An overview of enzymatic cellulose breakdown in the presence of lytic oxidative enzymes. a | Endoglucanases (hydrolases) catalyse hydrolytic chain cleavage, resulting in non-oxidized chain ends (blue hexagons), whereas lytic oxidases catalyse oxidative chain cleavage to result in oxidized chain ends (red hexagons). Cellobiohydrolases hydrolyse cellulose chains in a processive manner to produce cellobiose or oxidized cellobiose, depending on the oxidation state of the chain ends. The processive action of cellobiohydrolases generates a majority of cellobiose, which can be further cleaved to glucose by b-d-glucosidases. b | In the absence of oxidative cleavage, the hydrolysis of cellulose proceeds only through endoglucanases and processive cellobiohydrolases.
A N A LY S I S
228 | MARCH 2012 | VOLUME 10 www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved

Nature Reviews | Microbiology
GH5 CBM2 EXPN CBM63Clavibacter michiganensis subsp. sepedonicus (AAK16222)
CBM3 GH48 FN3 CBM2Acidothermus cellulolyticus (ABK52390)
GH6 CBM3 FN3 GH12 CBM2Acidothermus cellulolyticus (ABK52388)
GH74Thermotoga maritima (AAD35393)
GH74 CBM3 CBM3 GH48Caldicellulosiruptor bescii (ACM60948)
CalX CalX GH18 CalX CBM2 CalX CBM2 CalX GH5 CalXMycobacterium vanbaalenii (ABM14300)
GH26 GH5 CBM11 DOC1Clostridium thermocellum (AAA23225.1)
GH5 SLH SLH SLHCBM28Caldicellulosiruptor saccharolyticus (ABP66297)
GH5CE3 FN3 CBM2Amycolatopsis mediterranei (ADJ49127)
XylophagesOrganisms that feed on wood.
oxidative mechanism. In GH61 enzymes, copper is the most likely redox centre26 and, given the conservation of the catalytic metal‑binding site, it seems probable that members of both the GH61 and Cbp21 families are oxidative enzymes that target recalcitrant plant cell wall material such as cellulose. The possible cooperation of lytic oxidative enzymes with classical hydrolases during the breakdown of cellulose is shown in FIG. 1, although further dissection of the enzymatic mechanisms is neces‑sary given the range of both oxidized and non‑oxidized products that are produced by GH61 enzymes.
Architecture of cellulolytic enzymesEnzymes that digest plant cell walls, and in particular those that degrade cellulose and hemicellulose, are often modular, with a variable number of ancillary modules appended to the catalytic module9. In these modular structures, the ancillary module often binds com‑plex carbohydrates (such an ancillary module is then called a CBM) and sometimes binds other proteins or components of the bacterial cell surface (FIG. 2; see Supplementary information S1 (table)). In some cases, proteins contain more than one catalytic module and more than five different modules. Information on the molecular structure and catalytic mechanisms of these enzymes revealed that they have evolved convergently and several times from different ancestral scaffolds (BOX 1). CBMs are thought to potentiate the activity of the catalytic modules by targeting the enzyme to a specific component of the cell wall. However, given that cellulose‑binding CBMs may also be found in non‑cellulolytic enzymes, it is clear that the role of CBMs is probably much more complex than this. It should also be mentioned that sequence comparisons have found catalytic domains appended to protein domains with
no similarity to any other known enzyme27, suggesting that our knowledge of the complete enzymatic para‑phernalia involved in plant cell wall digestion is still incomplete.
Each type of structural polysaccharide‑degrading enzyme is found in multiple families and represented by myriads of modular architectures. Large efforts are under way worldwide to develop renewable energy sources as alternatives to fossil fuels, and the enzymatic conversion of plant cell walls to fermentable sugars is one of the most attractive routes. The discovery of new cellulose‑degrading enzymes will be an important facet of these efforts. Research into the use of cellulolytic bac‑teria and fungi for the production of cellulosic biofuels has been the subject of several recent reviews11,28 and is not discussed here.
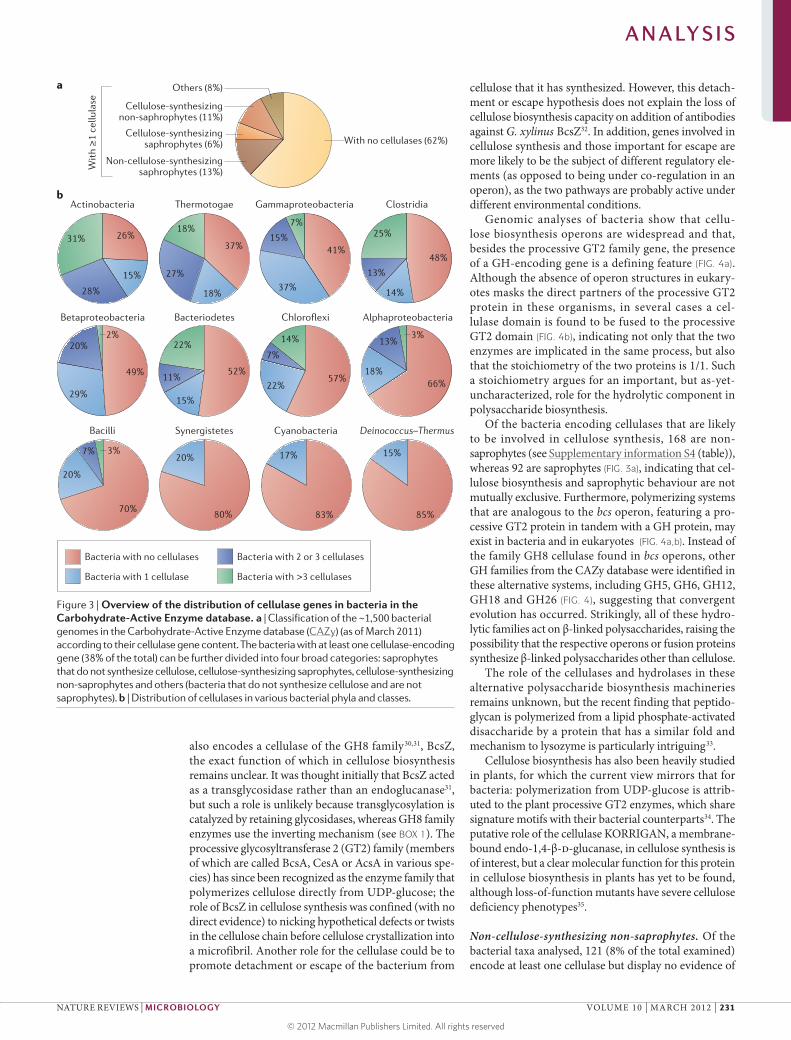
Phylogenetic distribution of cellulases By virtue of the abundance of cellulose in land plants, cellulases are commonly found in organisms that feed on plant cell walls, including herbivores, xylophages and, more generally, saprophytic organisms. To determine how common these enzymes are in bacteria, we analysed the genomes of the 1,495 bacteria listed in the CAZy database in March 2011 and, for each, we searched the genes encoding cellulases, hemicellulases and pectinases. Most bacteria (920 genomes, representing 62% of the total) do not contain cellulase genes, whereas 575 (38%) of all sequenced genomes contain at least one enzyme involved in cellulose cleavage. The bacteria that encode one cellulase or more can be divided into four categories: saprophytes that do not synthesize cellulose, cellulose‑synthesizing saprophytes, cellulose‑synthesizing non‑saprophytes and those that are neither saprophytic nor cellulose producing (FIG. 3a).
Figure 2 | Modularity of cellulolytic enzymes. Shown are some examples of bacterial cellulases that contain different domains. The Protein database accessions are given in brackets for each protein. CalX, Calx-β motif; CBM, carbohydrate- binding module; CE3, carbohydrate esterase family 3; DOC1, dockerin; EXPN, expansin; FN3, fibronectin type III; GH, glycoside hydrolase; SLH, surface layer homology.
A N A LY S I S
NATURE REVIEWS | MICROBIOLOGY VOLUME 10 | MARCH 2012 | 229
© 2012 Macmillan Publishers Limited. All rights reserved

Most bacteria that encode a cellulase have only one cellulase gene in their genome; such organisms were present in all phyla analysed. Bacteria with two or three cellulases genes were present in all phyla except for Synergistetes, Cyanobacteria and Deinococcus–Thermus (phyla for which all bacteria have only one or no cellulase) (FIG. 3b). Interestingly, few bacteria have more than three cellulases (FIG. 3b); these bac‑teria are mostly found in the phyla Actinobacteria, Bacteroidetes, Thermotogae and Chloroflexi and in the class Clostridia, and also include some members of the classes Gammaproteobacteria, Alphaproteobacteria, Bacilli and Betaproteobacteria (FIG. 3b). Bacteria that digest plant cell walls often encode multiple cellulases, together with hemicellulolytic and pectinolytic enzymes (TABLE 1). Thus, the co‑occurrence of genes encoding cellulolytic enzymes and those encoding hemicellulo‑lytic and/or pectinolytic enzymes accompanies a true saprophytic lifestyle.
Saprophytic bacteria. The saprophytic bacteria can be divided into two groups. The first group, referred to here as cellulase‑ and hemicellulase‑containing sapro phytes, was defined on the basis of the presence of at least one cellulase gene and three or more hemi cellulases or pectin‑ases. The 287 bacteria in this group (see Supplementary information S2 (table)) represent about half of the bacteria that encode cellulases. Of these, 92 can also
produce cellulose (see below). The second group con‑sists of 42 bacteria and is referred to as hemicellulase‑ containing saprophytes or non‑cellulolytic saprophytes, as the genomes contain hemicellulases and pectinases but lack cellulase (see Supplementary information S3 (table)). This group includes human gut bacteria such as Bacteroides thetaiotaomicron, Bacteroides vulgatus, Parabacteroides distasonis and Faecalibacterium prausnitzii str. SL3/3. The group also contains members of the genus Yersina, which includes major pathogens such as the plague agent, Yersinia pestis. All the Yersinia spp. genomes analysed contained cellulases and pectinases and were consequently assigned to the ‘cellulase‑ and hemicellulase‑ containing saprophytes’ category (see Supplementary information S2 (table)). This suggests that these bacteria have hitherto undescribed contact with plant material.
Cellulose-synthesizing bacteria. Approximately 11% (168 taxa) of the ~1,500 bacteria analysed are cellulose‑synthesizing non‑saprophytes (FIG. 3a). Numerous bacte‑ria produce cellulose as a biofilm matrix polymer using the enzymes encoded by the bacterial cellulose synthesis (bcs) operon. For example, in G. xylinus19,29 the cellulose synthesis pathway is encoded by the bcsA, bcsB, bcsC and bcsD genes, which together form an operon that is conserved in other cellulose‑producing bacteria such as Escherichia coli and Salmonella enterica subsp. enterica serovar Typhimurium30. Interestingly, the bcs operon
Box 1 | Structural and mechanistic features of cellulases
Sequence families and three-dimensional structuresThe Carbohydrate-Active Enzyme database (CAZy) sequence classification13 describes ~125 families of glycoside hydrolases. One of the notable features of cellulases is that they populate many different glycoside hydrolase families (around a dozen at the time of writing). What is particularly exciting, from a structural perspective, is that many of these families have completely different three-dimensional folds, indicating that cellulases have evolved convergently from diverse ancestors44,45.
Surface topographiesClassically, although not without controversy, cellulases have been considered as endo- or exo-acting and termed endoglucanases or cellobiohydrolases, respectively. The first three-dimensional structures of cellulases revealed that endoglucanases have an ‘open groove’ topography in which the substrate chain is accommodated in a canyon running across the enzyme surface, allowing access to the ‘internal’ glycosidic bonds of the substrate. The cellobiohydrolases, which are often related in sequence to endoglucanases, instead possess extended surface loops that generate a tunnel-shaped topography to facilitate a processive action on the substrate44,45. Loop breathing can probably impart an occasional endo-activity to the processive enzymes and, depending on whether the initial attack occurs solely at chain ends or randomly, cellobiohydrolases may be subdivided into exo-processive and endo-processive categories. In contrast to the cellobiohydrolases, true exo-enzymes (such as β-glucosidases) with an absolute specificity for the end of a substrate chain tend to have active centres that are buried in pockets or caves, and these enzymes have little, if any, activity on cellulose polymers.
Reaction mechanismsWith the exception of the emerging oxidative enzymes (see below), cellulases use acid–base chemistry to promote the hydrolysis of glycosidic bonds in cellulose. Two classical mechanisms exist, leading to inversion or retention of the configuration of the anomeric carbon. Transglycosylation events are restricted to retaining enzymes. Such mechanisms are beyond the scope of this Analysis article, and the reader is directed to recent reviews on the topic44,46,47.
Oxidative enzymes for polysaccharide degradationOther enzymes may participate in cellulose breakdown, using oxidative chemistry25,26. The first hint of these activities was given by a study reporting the cleavage of chitin (a fibrous, crystalline polymer of β-1,4-N-acetylglucosamine residues) by a bacterial enzyme that uses an oxidative step in tandem with an ascorbate cofactor23 to break down chitin and thus open up the crystalline polysaccharide material. Similar results have been observed with a family of fungal enzymes25,26 and, given the strong similarities in the three-dimensional structures and the binding sites of metal ions (most probably copper ions) for both enzyme classes, it seems likely that together they form a new group of lytic polysaccharide oxidases, many of which will be active on cellulose.
A N A LY S I S
230 | MARCH 2012 | VOLUME 10 www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved
Nature Reviews | Microbiology
Actinobacteria
Others (8%)
Cellulose-synthesizingnon-saphrophytes (11%)
Cellulose-synthesizingsaphrophytes (6%)
Non-cellulose-synthesizingsaphrophytes (13%)
With no cellulases (62%)
Wit
h ≥
1 ce
llula
sea
b
26%18%
37%15%
37%
41%
25%
13%
14%
48%
7%
18%
27%15%
28%
31%
20%
29%
49%
70%
Bacteria with no cellulases
Bacteria with 1 cellulase
Bacteria with 2 or 3 cellulases
Bacteria with >3 cellulases
80%
20% 17%
83% 85%
15%
20%
7% 3%
22%
11%
15%
52%22%
57% 66%18%
13%3%14%
7%
2%
Thermotogae Gammaproteobacteria Clostridia
BacteriodetesBetaproteobacteria Chloroflexi Alphaproteobacteria
Bacilli Synergistetes Cyanobacteria Deinococcus–Thermus
also encodes a cellulase of the GH8 family30,31, BcsZ, the exact function of which in cellulose biosynthesis remains unclear. It was thought initially that BcsZ acted as a transglycosidase rather than an endoglucanase31, but such a role is unlikely because transglycosylation is catalyzed by retaining glycosidases, whereas GH8 family enzymes use the inverting mechanism (see BOX 1). The processive glycosyltransferase 2 (GT2) family (members of which are called BcsA, CesA or AcsA in various spe‑cies) has since been recognized as the enzyme family that polymerizes cellulose directly from UDP‑glucose; the role of BcsZ in cellulose synthesis was confined (with no direct evidence) to nicking hypothetical defects or twists in the cellulose chain before cellulose crystallization into a microfibril. Another role for the cellulase could be to promote detachment or escape of the bacterium from
cellulose that it has synthesized. However, this detach‑ment or escape hypothesis does not explain the loss of cellulose biosynthesis capacity on addition of antibodies against G. xylinus BcsZ32. In addition, genes involved in cellulose synthesis and those important for escape are more likely to be the subject of different regulatory ele‑ments (as opposed to being under co‑regulation in an operon), as the two pathways are probably active under different environmental conditions.
Genomic analyses of bacteria show that cellu‑lose biosynthesis operons are widespread and that, besides the processive GT2 family gene, the presence of a GH‑encoding gene is a defining feature (FIG. 4a). Although the absence of operon structures in eukary‑otes masks the direct partners of the processive GT2 protein in these organisms, in several cases a cel‑lulase domain is found to be fused to the processive GT2 domain (FIG. 4b), indicating not only that the two enzymes are implicated in the same process, but also that the stoichiometry of the two proteins is 1/1. Such a stoichiometry argues for an important, but as‑yet‑uncharacterized, role for the hydrolytic component in polysaccharide biosynthesis.
Of the bacteria encoding cellulases that are likely to be involved in cellulose synthesis, 168 are non‑ saprophytes (see Supplementary information S4 (table)), whereas 92 are saprophytes (FIG. 3a), indicating that cel‑lulose biosynthesis and saprophytic behaviour are not mutually exclusive. Furthermore, polymerizing systems that are analogous to the bcs operon, featuring a pro‑cessive GT2 protein in tandem with a GH protein, may exist in bacteria and in eukaryotes (FIG. 4a,b). Instead of the family GH8 cellulase found in bcs operons, other GH families from the CAZy database were identified in these alternative systems, including GH5, GH6, GH12, GH18 and GH26 (FIG. 4), suggesting that convergent evolution has occurred. Strikingly, all of these hydro‑lytic families act on β‑linked polysaccharides, raising the possibility that the respective operons or fusion proteins synthesize β‑linked polysaccharides other than cellulose.
The role of the cellulases and hydrolases in these alternative polysaccharide biosynthesis machineries remains unknown, but the recent finding that peptido‑glycan is polymerized from a lipid phosphate‑activated disaccharide by a protein that has a similar fold and mechanism to lysozyme is particularly intriguing33.
Cellulose biosynthesis has also been heavily studied in plants, for which the current view mirrors that for bacteria: polymerization from UDP‑glucose is attrib‑uted to the plant processive GT2 enzymes, which share signature motifs with their bacterial counterparts34. The putative role of the cellulase KORRIGAN, a membrane‑bound endo‑1,4‑β‑d‑glucanase, in cellulose synthesis is of interest, but a clear molecular function for this protein in cellulose biosynthesis in plants has yet to be found, although loss‑of‑function mutants have severe cellulose deficiency phenotypes35.
Non-cellulose-synthesizing non-saprophytes. Of the bacterial taxa analysed, 121 (8% of the total examined) encode at least one cellulase but display no evidence of
Figure 3 | Overview of the distribution of cellulase genes in bacteria in the Carbohydrate-Active Enzyme database. a | Classification of the ~1,500 bacterial genomes in the Carbohydrate-Active Enzyme database (CAZy) (as of March 2011) according to their cellulase gene content. The bacteria with at least one cellulase-encoding gene (38% of the total) can be further divided into four broad categories: saprophytes that do not synthesize cellulose, cellulose-synthesizing saprophytes, cellulose-synthesizing non-saprophytes and others (bacteria that do not synthesize cellulose and are not saprophytes). b | Distribution of cellulases in various bacterial phyla and classes.
A N A LY S I S
NATURE REVIEWS | MICROBIOLOGY VOLUME 10 | MARCH 2012 | 231
© 2012 Macmillan Publishers Limited. All rights reserved

a saprophytic lifestyle or the ability to synthesize cellu‑lose (see Supplementary information S5 (table)). The existence of cellulase genes in these bacteria is extremely puzzling, as it is unclear what the function of these genes might be in these organisms. One example of such an organism is M. tuberculosis, which encodes two active cellulases and one active cellulose‑binding protein regardless of the strain15,17. Two of these three proteins have secretion signal peptides. Likewise, all strains of the pathogenic bacterium L. pneumophila encode a secreted cellulase that degrades the artificial substrate carboxy‑methyl cellulose as well as microcrystalline cellulose18. Neither L. pneumophila nor M. tuberculosis has any known relationship with plants, and both species lack a more complete cellulose‑degrading machinery and have no hemicellulase nor pectinase genes. The cellulase genes that are present in these species may be remnants that are slowly decaying. For example, the M. tuberculosis cellulase‑related protein encoded by Rv1090 is truncated, as it lacks the first 35 amino acids (including the signal sequence16), and is unstable when produced in E. coli, perhaps indicative of decay. However, this protein has
been conserved since the speciation events that gave rise to M. tuberculosis and the six other, closely related myco‑bacterial species that also cause tuberculosis in humans and animals (collectively, these seven mycobacteria are called the tuberculous mycobacteria). Furthermore, this protein and the two other proteins of M. tuberculosis that putatively target cellulose — encoded by Rv0062 (REF. 15) and Rv1987 (REF. 17) — displayed the expected activity on production in E. coli, and the other six tuberculous mycobacteria also encode these three cellulose‑targeting proteins16. Together, these findings indicate that there has been evolutionary pressure to maintain the genes encoding these proteins.
L. pneumophila is an intracellular pathogen36,37 that lives in water‑borne free‑living amoebae38 and has evolved to resist macrophages after intra‑amoebal ‘train‑ing’ (REFS 18,38). M. tuberculosis is also an intracellular pathogen that can reside within host macrophages39,40, and it was recently shown to survive within amoebae following experimental infection41. Although the role of intra‑amoebal survival in the life cycle of tuberculous mycobacteria remains purely hypothetical, it is possible that the cellulases in M. tuberculosis and L. pneumo phila target the cellulose produced when amoebae encyst. Contrary to the non‑tuberculous species Mycobacterium avium, the tuberculous mycobacteria M. tuberculosis and Mycobacterium bovis bypass the amoebal cyst through an unknown mechanism after internalization into the amoebal trophozoite41. The amoebal cyst wall is a simple structure consisting of an outer, proteinaceous ectocyst and an inner, cellulose‑containing endocyst, and bacteria do not require a complex arsenal of enzymes to degrade this wall. Although an obvious hypothesis is that the cyst cellulose is broken down by the bacteria to generate glu‑cose, more intricate roles for this breakdown cannot be excluded; for example, this degradation may enable the bacteria to detect amoebal cellulose synthesis at an early stage to trigger escape mechanisms before encystment.
Other bacteria besides M. tuberculosis and L. pneumophila can survive or multiply in amoebae, including Vibrio cholerae42, related Vibrio spp. and Listeria monocytogenes43. Together, these examples suggest a novel role for bacterial cellulases in the interactions of bacte‑ria with the cellulose derived from amoebal encystment. The data discussed here could prompt additional studies on bacterium–amoeba relationships for bacterial species with one or more cellulases. However, for many bacteria, interactions with amoebae have not been documented in the published literature, and the role of cellulases in these organisms remains enigmatic (see Supplementary information S5 (table)).
ConclusionsWe have tried to show that, contrary to popular accept‑ance, the presence of a cellulase‑encoding gene in the genome of an organism does not automatically imply that the organism is involved in plant cell wall degrada‑tion. Until now, cellulases of saprophytic organisms have received attention mostly for their potential in industrial processes — most recently, biomass conversion — but the role of cellulases in non‑saprophytic organisms is a novel
Table 1 | The bacteria encoding the most cellulolytic enzymes in various taxa*
Name (GenBank accession) Number of cellulase genes
Number of hemicellulase genes
Phylum Actinobacteria
Streptomyces bingchenggensis str. BCW-1 (CP002047)
28 36
Amycolatopsis mediterranei str. U32 (CP002000)
21 30
Streptomyces scabiei str. 87.22 (FN554889)
20 33
Catenulispora acidiphila str. DSM 44928 (CP001700)
19 23
Actinosynnema mirum str. DSM 43827 (CP001630)
17 20
Phylum Gammaproteobacteria
Teredinibacter turnerae str. T7901 (CP001614)
29 28
Saccharophagus degradans str. 2-40 (CP000282)
24 29
Cellvibrio japonicus str. Ueda 107 (CP000934)
21 30
Phylum Firmicutes (class Clostridia)
Clostridium thermocellum str. ATCC 27405 (CP000568)
32 19
Clostridium thermocellum str. DSM 1313 (CP002416)
32 20
Clostridium cellulovorans str. 743B (CP002160)
29 24
Ruminococcus albus str. 7 (CP002403) 27 30
Ruminococcus sp. 18P13 (FP929052) 20 12
Phylum Bacteroidetes
Cytophaga hutchinsonii str. ATCC 33406 (CP000383)
19 8
*Only bacteria with 15 or more cellulase genes are included.
A N A LY S I S
232 | MARCH 2012 | VOLUME 10 www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved

Nature Reviews | Microbiology
Escherichia coli (ACR63025) GH8
GH5
GH18Bacteria
GH6Streptosporangium roseum (ACZ86824)
Paenibacillus sp. (ACX64320)
Nocardioides sp. (ABL80248)
Other genes of the operon (bcsB, bcsC, etc.) GH gene GT2 gene (bcsA, yhjO)
Signal peptide Transmembrane domain GT2 module GH module
GH12
GH8
a
b
GH6 Animal
Bacterium
Bacterium
Fungus
GH26
Thielavia terrestris (AEO69079)
Anaeromyxobacter dehalogenans (ACL65274)
Oikopleura dioica (BAJ65326)
Chitinophaga pinensis (ACU60596)
problem, the answer to which is emerging from our analy‑ses of genomic data. These analyses show that many non‑ saprophytic organisms encode a cellulase which, counter‑intuitively, plays a part in cellulose biosynthesis, revealing that cellulases are not restricted to cellulose degradation. The exact role of cellulase in this process remains unclear, despite the potential interest in controlling or engineering cellulose synthesis to produce materials with novel prop‑erties or enhanced biodigestibility. Furthermore, the cel‑lulases that are present in human pathogens may reflect an intra‑amoebal lifestyle and a requirement to degrade amoebal cellulose. In light of this genomic analysis, we
suggest that cellulolytic bacteria can currently be clas‑sified into four categories: those that use cellulases to feed on plant cell walls, those that use cellulases for cel‑lulose biosynthesis, those that do both and those, like L. pneumo phila, that may at one stage in their life cycle reside inside amoebae (FIG. 3a). Given the important soci‑etal and health issues related to the persistence of both mycobacteria and Legionella spp., the observation that cellulases may be implicated in the mechanisms of per‑sistence for these organisms is thought‑provoking and will perhaps lead to the generation of novel strategies to combat these pathogens.
Figure 4 | Polysaccharide biosynthesis operons and cellulase fusion proteins. a | Polysaccharide biosynthesis operons in bacterial genomes, including the bacterial cellulose synthesis (bcs) operon of Escherichia coli str. K12 and analogous operons in other species. The glycoside hydrolase (GH)-encoding gene of the bcs operon, bcsZ, is from the GH8 family; other GH families are represented in analogous operons. Similarly, each operon contains a processive glycosyl-transferase 2 (GT2) family-encoding gene, which in the bcs operon is bcsA. GenBank, Gene or Protein database accessions are given in brackets for each GH family gene (or the encoded protein). b | Cellulase fusion proteins observed in bacteria and eukaryotes. GenBank, Gene or Protein database accessions are given in brackets.
1. Chavez-Munguia, B. et al. Ultrastructure of cyst differentiation in parasitic protozoa. Parasitol. Res. 100, 1169–1175 (2007).
2. Matthysse, A. G. et al. A functional cellulose synthase from ascidian epidermis. Proc. Natl Acad. Sci. USA 101, 986–991 (2004).
3. Tomlinson, G., Jones, E. A. & Kahne, D. Isolation of cellulose from the cyst of a soil amoeba. Biochim. Biophs. Acta 63, 194–200 (1962).
4. Lynd, L. R., Weimer, P. J., van Zyl, W. H. & Pretorius, I. S. Microbial cellulose utilization: fundamentals and biotechnology. Microbiol. Mol. Biol. Rev. 66, 506–577 (2002).
5. Somerville, C. et al. Toward a systems approach to understanding plant cell walls. Science 306, 2206–2211 (2004).
6. Nishiyama, Y., Langan, P. & Chanzy, H. Crystal structure and hydrogen-bonding system in cellulose Iβ from synchrotron X-ray and neutron fiber diffraction. J. Am. Chem. Soc. 124, 9074–9082 (2002).
7. Nishiyama, Y., Sugiyama, J., Chanzy, H. & Langan, P. Crystal structure and hydrogen bonding system in cellulose Iα from synchrotron X-ray and neutron fiber diffraction. J. Am. Chem. Soc. 125, 14300–14306 (2003).
8. Yarbrough, J. M., Himmel, M. E. & Ding, S. Y. Plant cell wall characterization using scanning probe microscopy techniques. Biotechnol. Biofuels 2, 17 (2009).
9. Herve, C. et al. Carbohydrate-binding modules promote the enzymatic deconstruction of intact plant cell walls by targeting and proximity effects. Proc. Natl Acad. Sci. USA 107, 15293–15298 (2010).
10. Himmel, M. E. et al. Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science 315, 804–807 (2007).
11. Atsumi, S., Higashide, W. & Liao, J. C. Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde. Nature Biotech. 27, 1177–1180 (2009).
12. Doi, R. H. & Kosugi, A. Cellulosomes: plant-cell-wall-degrading enzyme complexes. Nature Rev. Microbiol. 2, 541–551 (2004).
13. Cantarel, B. L. et al. The Carbohydrate-Active EnZymes database (CAZy): an expert resource for glycogenomics. Nucleic Acids Res. 37, D233–D238 (2009).
14. Henrissat B. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 280, 309–316 (1991).
15. Varrot, A. et al. Mycobacterium tuberculosis strains possess functional cellulases. J. Biol. Chem. 280, 20181–20184 (2005).
16. Mba Medie, F., Ben Salah, I., Drancourt, M. & Henrissat, B. Paradoxical conservation of a set of three cellulose-targeting genes in Mycobacterium tuberculosis complex organisms. Microbiology 156, 1468–1475 (2010).
17. Mba Medie, F., Vincentelli, R., Drancourt, M. & Henrissat, B. Mycobacterium tuberculosis Rv1090 and Rv1987 encode functional β-glucan-targeting proteins. Protein Expr. Purif. 75, 172–176 (2011).
18. Pearce, M. M. & Cianciotto, N. P. Legionella pneumophila secretes an endoglucanase that belongs to the family-5 of glycosyl hydrolases and is dependent upon type II secretion. FEMS Microbiol. Lett. 300, 256–264 (2009).
19. Wong, H. C. et al. Genetic organization of the cellulose synthase operon in Acetobacter xylinum. Proc. Natl Acad. Sci. USA 87, 8130–8134 (1990).
20. Nicol, F. et al. A plasma membrane-bound putative endo-1,4-β-d-glucanase is required for normal wall assembly and cell elongation in Arabidopsis. EMBO J. 17, 5563–5576 (1998).
21. Henrissat, B., Driguez, H., Viet, C. & Schulein, M. Synergism of cellulase from Trichoderma reesei in the degradation of cellulose. Nature Biotech. 3, 722–726 (1985).
A N A LY S I S
NATURE REVIEWS | MICROBIOLOGY VOLUME 10 | MARCH 2012 | 233
© 2012 Macmillan Publishers Limited. All rights reserved

22. Olson, D. G. et al. Deletion of the Cel48S cellulase from Clostridium thermocellum. Proc. Natl Acad. Sci. USA 107, 17727–17732 (2010).
23. Vaaje-Kolstad, G. et al. An oxidative enzyme boosting the enzymatic conversion of recalcitrant polysaccharides. Science 330, 219–222 (2010).
24. Forsberg, Z. et al. Cleavage of cellulose by a CBM33 protein. Protein Sci. 20, 1479–1483 (2011).
25. Harris, P. V. et al. Stimulation of lignocellulosic biomass hydrolysis by proteins of glycoside hydrolase family 61: structure and function of a large, enigmatic family. Biochemistry 49, 3305–3316 (2010).
26. Quinlan, R. J. et al. Insights into the oxidative degradation of cellulose by a copper metalloenzyme that exploits biomass components. Proc. Natl Acad. Sci. USA 108, 15079–15084 (2011).
27. Vincent, F., Molin, D. D., Weiner, R. M., Bourne, Y. & Henrissat, B. Structure of a polyisoprenoid binding domain from Saccharophagus degradans implicated in plant cell wall breakdown. FEBS Lett. 584, 1577–1584 (2010).
28. Rubin, E. M. Genomics of cellulosic biofuels. Nature 454, 841–845 (2008).
29. Ross, P., Mayer, R. & Benziman, M. Cellulose biosynthesis and function in bacteria. Microbiol. Rev. 55, 35–58 (1991).
30. Zogaj, X., Nimtz, M., Rohde, M., Bokranz, W. & Romling, U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 39, 1452–1463 (2001).
31. Matthysse, A. G., Thomas, D. L. & White, A. R. Mechanism of cellulose synthesis in Agrobacterium tumefaciens. J. Bacteriol. 177, 1076–1081 (1995).
32. Koo, H. M., Song, S. H., Pyun, Y. R. & Kim, Y. S. Evidence that a β-1,4-endoglucanase secreted by Acetobacter xylinum plays an essential role for the formation of cellulose fiber. Biosci. Biotechnol. Biochem. 62, 2257–2259 (1998).
33. Yuan, Y. et al. Crystal structure of a peptidoglycan glycosyltransferase suggests a model for processive
glycan chain synthesis. Proc. Natl Acad. Sci. USA 104, 5348–5353 (2007).
34. Pear, J. R., Kawagoe, Y., Schreckengost, W. E., Delmer, D. P. & Stalker, D. M. Higher plants contain homologs of the bacterial celA genes encoding the catalytic subunit of cellulose synthase. Proc. Natl Acad. Sci. USA 93, 12637–12642 (1996).
35. Robert, S. et al. An Arabidopsis endo-1,4-β-d-glucanase involved in cellulose synthesis undergoes regulated intracellular cycling. Plant Cell 17, 3378–3389 (2005).
36. Molmeret, M. et al. Temporal and spatial trigger of post-exponential virulence-associated regulatory cascades by Legionella pneumophila after bacterial escape into the host cell cytosol. Environ. Microbiol. 12, 704–715 (2010).
37. Weissenmayer, B. A., Prendergast, J. G., Lohan, A. J. & Loftus, B. J. Sequencing illustrates the transcriptional response of Legionella pneumophila during infection and identifies seventy novel small non-coding RNAs. PLoS ONE 6, e17570 (2011).
38. Molmeret, M., Horn, M., Wagner, M., Santic, M. & Abu, K. Y. Amoebae as training grounds for intracellular bacterial pathogens. Appl. Environ. Microbiol. 71, 20–28 (2005).
39. Daniel, J., Maamar, H., Deb, C., Sirakova, T. D. & Kolattukudy, P. E. Mycobacterium tuberculosis uses host triacylglycerol to accumulate lipid droplets and acquires a dormancy-like phenotype in lipid-loaded macrophages. PLoS Pathog. 7, e1002093 (2011).
40. Harding, C. V. & Boom, W. H. Regulation of antigen presentation by Mycobacterium tuberculosis: a role for Toll-like receptors. Nature Rev. Microbiol. 8, 296–307 (2010).
41. Mba Medie, F., Ben Salah, I., Henrissat, B., Raoult, D. & Drancourt, M. Mycobacterium tuberculosis complex mycobacteria as amoeba-resistant organisms. PLoS ONE 6, e20499 (2011).
42. Abd, H., Saeed, A., Weintraub, A. & Sandstrom, G. Vibrio cholerae O139 requires neither capsule nor
LPS O side chain to grow inside Acanthamoeba castellanii. J. Med. Microbiol. 58, 125–131 (2009).
43. Thomas, V., McDonnell, G., Denyer, S. P. & Maillard, J. Y. Free-living amoebae and their intracellular pathogenic microorganisms: risks for water quality. FEMS Microbiol. Rev. 34, 231–259 (2009).
44. Davies, G. J., Tolley, S. P., Henrissat, B., Hjort, C. & Schulein, M. Structures of oligosaccharide-bound forms of the endoglucanase V from Humicola insolens at 1.9 Å resolution. Biochemistry 34, 16210–16220 (1995).
45. Henrissat, B. & Davies, G. Structural and sequence-based classification of glycoside hydrolases. Curr. Opin. Struct. Biol. 7, 637–644 (1997).
46. Zechel, D. L. & Withers, S. G. Glycosidase mechanisms: anatomy of a finely tuned catalyst. Acc. Chem. Res. 33, 11–18 (2000).
47. Vocadlo, D. J. & Davies, G. J. Mechanistic insights into glycosidase chemistry. Curr. Opin. Chem. Biol. 12, 539–555 (2008).
AcknowledgementsF.M.M. was funded by La Fondation Infectiopole Sud, France. G.J.D. is a Royal Society Wolfson Research Merit Award recipient.
Competing interests statementThe authors declare no competing financial interests.
FURTHER INFORMATIONCAZy: http://www.cazy.org/ENZYME: http://enzyme.expasy.org/GenBank: http://www.ncbi.nlm.nih.gov/genbank/Gene: http://www.ncbi.nlm.nih.gov/gene/Protein: http://www.ncbi.nlm.nih.gov/protein/
SUPPLEMENTARY INFORMATIONSee online article: S1 (table) | S2 (table) | S3 (table) | S4 (table) | S5 (table)
ALL LINKS ARE ACTIVE IN THE ONLINE PDF
A N A LY S I S
234 | MARCH 2012 | VOLUME 10 www.nature.com/reviews/micro
© 2012 Macmillan Publishers Limited. All rights reserved


















