
Getting a Grip on Prions: Oligomers, Amyloids, and Pathological ...bio156/Papers/PDFs/Annu. Rev....
31
Getting a Grip on Prions: Oligomers, Amyloids, and Pathological Membrane Interactions ∗ Byron Caughey, 1 Gerald S. Baron, 1 Bruce Chesebro, 1 and Martin Jeffrey 2 1 Laboratory of Persistent Viral Diseases, Rocky Mountain Laboratories, National Institute for Allergy and Infectious Diseases, National Institutes of Health, Hamilton, Montana 59840; email: [email protected]; [email protected]; [email protected] 2 Veterinary Laboratories Agency, Lasswade, Pentlands Science Park, Penicuik, Midlothian, EH26 0PZ, United Kingdom; email: [email protected] Annu. Rev. Biochem. 2009. 78:177–204 First published online as a Review in Advance on February 20, 2009 The Annual Review of Biochemistry is online at biochem.annualreviews.org This article’s doi: 10.1146/annurev.biochem.78.082907.145410 Copyright c 2009 by Annual Reviews. All rights reserved 0066-4154/09/0707-0177$20.00 ∗ The U.S. Government has the right to retain a nonexclusive, royalty-free license in and to any copyright covering this paper. Key Words exosomes, glycophosphatidylinositol anchor, protein misfolding diseases, tunneling nanotubes Abstract The prion (infectious protein) concept has evolved with the discovery of new self-propagating protein states in organisms as diverse as mammals and fungi. The infectious agent of the mammalian transmissible spongi- form encephalopathies (TSE) has long been considered the prototypi- cal prion, and recent cell-free propagation and biophysical analyses of TSE infectivity have now firmly established its prion credentials. Other disease-associated protein aggregates, such as some amyloids, can also have prion-like characteristics under certain experimental conditions. However, most amyloids appear to lack the natural transmissibility of TSE prions. One feature that distinguishes the latter from the former is the glycophosphatidylinositol membrane anchor on prion protein, the molecule that is corrupted in TSE diseases. The presence of this anchor profoundly affects TSE pathogenesis, which involves major membrane distortions in the brain, and may be a key reason for the greater neu- rovirulence of TSE prions relative to many other autocatalytic protein aggregates. 177 Annu. Rev. Biochem. 2009.78:177-204. Downloaded from www.annualreviews.org by California Institute of Technology on 02/10/12. For personal use only.
Transcript of Getting a Grip on Prions: Oligomers, Amyloids, and Pathological ...bio156/Papers/PDFs/Annu. Rev....

ANRV378-BI78-08 ARI 5 May 2009 13:45
Getting a Grip on Prions:Oligomers, Amyloids,and Pathological MembraneInteractions∗
Byron Caughey,1 Gerald S. Baron,1 Bruce Chesebro,1
and Martin Jeffrey2
1Laboratory of Persistent Viral Diseases, Rocky Mountain Laboratories, National Institutefor Allergy and Infectious Diseases, National Institutes of Health, Hamilton,Montana 59840; email: [email protected]; [email protected]; [email protected] Laboratories Agency, Lasswade, Pentlands Science Park, Penicuik,Midlothian, EH26 0PZ, United Kingdom; email: [email protected]
Annu. Rev. Biochem. 2009. 78:177–204
First published online as a Review in Advance onFebruary 20, 2009
The Annual Review of Biochemistry is online atbiochem.annualreviews.org
This article’s doi:10.1146/annurev.biochem.78.082907.145410
Copyright c© 2009 by Annual Reviews.All rights reserved
0066-4154/09/0707-0177$20.00∗ The U.S. Government has the right to retain anonexclusive, royalty-free license in and to anycopyright covering this paper.
Key Words
exosomes, glycophosphatidylinositol anchor, protein misfoldingdiseases, tunneling nanotubes
AbstractThe prion (infectious protein) concept has evolved with the discovery ofnew self-propagating protein states in organisms as diverse as mammalsand fungi. The infectious agent of the mammalian transmissible spongi-form encephalopathies (TSE) has long been considered the prototypi-cal prion, and recent cell-free propagation and biophysical analyses ofTSE infectivity have now firmly established its prion credentials. Otherdisease-associated protein aggregates, such as some amyloids, can alsohave prion-like characteristics under certain experimental conditions.However, most amyloids appear to lack the natural transmissibility ofTSE prions. One feature that distinguishes the latter from the former isthe glycophosphatidylinositol membrane anchor on prion protein, themolecule that is corrupted in TSE diseases. The presence of this anchorprofoundly affects TSE pathogenesis, which involves major membranedistortions in the brain, and may be a key reason for the greater neu-rovirulence of TSE prions relative to many other autocatalytic proteinaggregates.
177
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
Contents
INTRODUCTION . . . . . . . . . . . . . . . . . . 178IN VITRO PROPAGATION
OF PRIONS . . . . . . . . . . . . . . . . . . . . . . 179PrP Conversion . . . . . . . . . . . . . . . . . . . . 179“Synthetic Prions” . . . . . . . . . . . . . . . . . 179Protein Misfolding Cyclic
Amplification . . . . . . . . . . . . . . . . . . . 180Recombinant PrP-PMCA
and Related AmplificationReactions . . . . . . . . . . . . . . . . . . . . . . . 181
STRUCTURAL STUDIES OF PrPOLIGOMERS AND FIBRILS . . . . . 182Electron Crystallography
and Modeling . . . . . . . . . . . . . . . . . . . 182Other Probing of Fibril Structure . . . 184The Most Infectious Particle . . . . . . . 184
BIOGENESIS OF PrPres . . . . . . . . . . . . . 184FIBRILLAR VERSUS
NONFIBRILLAR PrPres
DEPOSITS . . . . . . . . . . . . . . . . . . . . . . . 185TSE PRION TRANSMISSION,
UPTAKE, AND TRANSPORTMECHANISMS. . . . . . . . . . . . . . . . . . . 187
PrPres Uptake and Acute Inductionof PrPC Conversion . . . . . . . . . . . . . 187
CELL-TO-CELL SPREADOF TSE PRIONS . . . . . . . . . . . . . . . . . 188Exosomes . . . . . . . . . . . . . . . . . . . . . . . . . 188Tunneling Nanotubes . . . . . . . . . . . . . . 189
PrP-MEMBRANE ASSOCIATIONAND PATHOGENESIS. . . . . . . . . . . 190How Might Membrane-Associated
PrPres Be Neurotoxic? . . . . . . . . . . . 191A Pathological Role for
GPI-Anchored PrPC
on Neurons? . . . . . . . . . . . . . . . . . . . . 192Direct and Indirect Mechanisms
of PrPres Neurotoxicity . . . . . . . . . . 193STRETCHING THE REALM
OF PRIONS . . . . . . . . . . . . . . . . . . . . . . 193AA Amyloidosis . . . . . . . . . . . . . . . . . . . . 194Apolipoprotein AII Amyloidosis . . . . 195Aβ Amyloidoses . . . . . . . . . . . . . . . . . . . 195Cytoplasmic Polyadenylation
Element-Binding Protein . . . . . . . 195Amyloid Spreading Mechanisms? . . . 195
Prion: an infectiousprotein or self-propagating protein-based element ofepigenetic inheritancewith the capacity tospread between hosts
TSE: transmissiblespongiformencephalopathy
INTRODUCTION
In the 1960s, the notion that corrupted hostproteins could act as infectious pathogens wasproposed by Griffith (1) as an explanation forthe mysterious sheep disease called scrapie. In1982, Prusiner (2) coined the term prion forsuch proteinaceous infectious agents that ap-parently lack their own specific nucleic acidgenome. Shortly thereafter, Prusiner discov-ered a host protein, prion protein (PrP), whosealteration is critical in the pathogenesis ofthe transmissible spongiform encephalopathies(TSEs) of mammals. However, it was excep-tionally difficult to show unequivocally thatprions are composed solely of modified PrPwithout any prion-specific nucleic acid. Thus,for a long time it remained uncertain whetherprions exist as infectious pathogenic proteinsin mammals. Only recently has there been a
report of the propagation of robust mammalianprions in vitro using semipurified molecularconstituents. This directly implicates a PrPconformational change in prion propagationand seems to rule out the need for an agent-specific protein-encoding nucleic acid (3, 4).Meanwhile, in 1994, Wickner (5) argued com-pellingly that mysterious epigenetic elements ofyeast could also be explained by the transfer ofaggregated proteins from one cell to another.In doing so, Wickner proposed that the prionconcept be extended to include these proteina-ceous entities that could act as infectious agentsand/or convey heritable changes in phenotypewithout mediation by nucleic acids. Many di-verse biological phenomena have now been de-scribed as being prions or prion like. Thereare several basic requirements for a prion (6).First, there must be a self-propagating state of a
178 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
protein (the prion) that is biologically accessiblebut rarely formed spontaneously. Second, pri-ons must replicate themselves by acting on theirnonprion substrate protein. And third, prionsmust spread to naive hosts and find new sub-strate pools for replication. Typically, prionsalso cause phenotypic changes in the host.
In this review, we first describe the challeng-ing hunt for molecularly defined mammalianTSE prions. We then consider recent insightsinto their three-dimensional structure, biogen-esis, and neuropathogenic effects. Emphasisis given to a unique aspect of mammalianTSE diseases, that is, the anchoring of theprotein of interest, PrP, to membranes by aglycophosphatidylinositol (GPI) moiety. Thisfeature strongly influences TSE prionpathogenicity. Moreover, neuronal mem-brane abnormalities figure prominently inthe pathognomonic lesions of TSE diseases,prompting us to also consider the extent towhich membrane tethering plays a role indistinguishing TSE prion biology from thebiology of other types of prions or prion-likephenomena in mammals. A number of differentproteins of mammals and fungi can be foundin altered states with at least some propertiesof prions. These examples extend the prionconcept and help to define its fundamentalfeatures and useful limits.
IN VITRO PROPAGATIONOF PRIONS
The most compelling way to demonstrate thecomposition of prions would be to propagatethem in vitro using defined biochemical con-stituents. Such a demonstration has long beenelusive in the case of TSE prions. However,striking progress toward this goal has beenmade in recent years.
PrP Conversion
Mammalian prion propagation is usually asso-ciated with the posttranslational conversion ofthe host’s normal, soluble, protease-sensitivePrP (PrPC or PrP-sen) to a less soluble and
GPI: glycophos-phatidylinositol
PrPC: normal cellularisoform of the prionprotein (PrP)
PrPres: abnormal,partially protease-resistant, priondisease-associated isoform ofPrP; largelysynonymous withPrPSc
Amyloid: proteinfibrils exhibiting across-β-core structureand specific stainingcharacteristics withdiagnostic dyes(thioflavins S and T,Congo red)
more proteinase K-resistant state (PrPres orPrPSc) (7). Initial attempts to propagate pri-ons in extensively purified cell-free systemsshowed that PrPres can cause PrPC to con-vert to a PrPres-like protease-resistant statein a highly species- and sequence-dependentmanner consistent with known prion transmis-sion barriers in vivo (8–11). PrPres with PrPC-converting activity is oligomeric, and newlyconverted PrP molecules become associatedwith PrPres oligomers (12–14). These and otherfeatures of the conversion reaction are consis-tent with either noncatalytic or autocatalytic(templated) seeded polymerization mechanismsinitially outlined by Gadjusek (15) and Lans-bury and coworkers (12, 16) (Figure 1). PrPres
oligomers of different prion strain-associatedconformations are capable of imposing thosedistinct conformations on PrPC molecules dur-ing conversion (8). This provides a potentialmolecular basis for the propagation of prionstrains without the need for a prion-specificnucleic acid. Although the presence of cell-free PrPC-converting activity correlates withthe presence of prion infectivity (17), the yieldsof the initial cell-free PrPC conversion reactionswere usually substoichiometric relative to thePrPres seed, and the generation of prion infec-tivity was not demonstrable (18).
“Synthetic Prions”
More recently, synthetic amyloid fibrils com-posed of a truncated fragment of recombinantPrPC (PrP residues 90–231) were shown to ac-celerate clinical disease when inoculated intotransgenic mice that overexpress PrP90-231by ∼16-fold (19, 20). Furthermore, brain tis-sue from these sick mice contained prions thatcaused TSE disease when inoculated into wild-type mice. These striking results suggested thatamyloid preparations of recombinant PrP aloneinstigated transmissible prion disease, albeit inan unnatural host that, by virtue of PrP over-expression, was likely to be strongly primedfor PrP aggregation and susceptibility to bothinduced and spontaneous prion disease. Im-portantly, the initial synthetic fibrils had no
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 179
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
a Seeded polymerization (autocatalytic or templated)
b Seeded polymerization (noncatalytic)
PrPres
Rapid
PrPC
[ ]n
[ ]n
Figure 1Seeded PrP polymerization mechanisms. (a) In the autocatalytic model, the conformational conversion ofPrPC to PrPres is rare unless catalyzed by contact with an existing PrPres multimer. (b) In the noncatalyticmodel, the conformational interchange between the PrPC and PrPres conformations is rapid, but the PrPres
conformer is poorly populated unless stabilized by binding to an existing PrPres multimer.
PMCA: proteinmisfolding cyclicamplification
activity in wild-type mice, implying that these“synthetic prions” were many orders of magni-tude less infectious than bona fide scrapie pri-ons. There are several possible explanations forthese results:
1. Recombinant PrP90-231 amyloid maybe infectious but with extremely poorefficiency.
2. The infectious entity might not be theamyloid itself but a different form ofPrP-containing particle that is present atonly trace quantities in the synthetic amy-loid preparation.
3. Robust recombinant PrP prions mightrequire another molecular constituent[e.g., polyanions such as glycosaminogly-cans (GAGs) or RNA (4)].
4. The amyloid preparations might not beinfectious at all but may accelerate aneurodegenerative process that occursspontaneously in transgenic mice overex-pressing truncated PrP90–231.
In this latter case, the infectivity for wild-type mice could have arisen spontaneously inthe transgenic mice, as has been reported inother transgenic mice that overexpress PrPmutants (21). Further studies are required todiscriminate between these possibilities.
Protein Misfolding CyclicAmplification
In a major breakthrough, brain-homogenate-based reactions were developed that allowedunlimited amplification of both PrPres andprions that were infectious for wild-type ro-dents (3, 22, 23). This amplification proto-col, called PMCA for protein misfolding cyclicamplification, involves introducing minuteamounts of an infectious PrPres-containing seedinto a homogenate of brain from an unin-fected animal, which provides PrPC and anycofactors that might be needed. The suspen-sion is subjected to cycles of incubation, which
180 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.
ANRV378-BI78-08 ARI 5 May 2009 13:45
a
Normal brain homogenateor
PrPC + polyanions
PrPres
seedAmplified
PrPres
Polyanion
Sonication
Sonication-incubation
cycles
Dilution intofresh PrPC
etc.
b
PrPres
PrPC
[ ]n
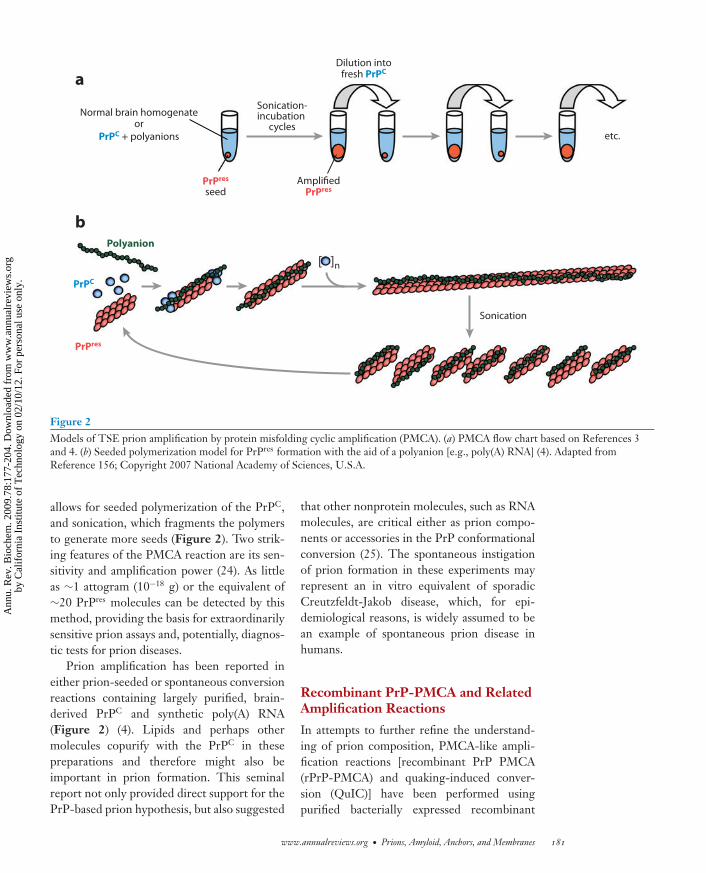
Figure 2Models of TSE prion amplification by protein misfolding cyclic amplification (PMCA). (a) PMCA flow chart based on References 3and 4. (b) Seeded polymerization model for PrPres formation with the aid of a polyanion [e.g., poly(A) RNA] (4). Adapted fromReference 156; Copyright 2007 National Academy of Sciences, U.S.A.
allows for seeded polymerization of the PrPC,and sonication, which fragments the polymersto generate more seeds (Figure 2). Two strik-ing features of the PMCA reaction are its sen-sitivity and amplification power (24). As littleas ∼1 attogram (10−18 g) or the equivalent of∼20 PrPres molecules can be detected by thismethod, providing the basis for extraordinarilysensitive prion assays and, potentially, diagnos-tic tests for prion diseases.
Prion amplification has been reported ineither prion-seeded or spontaneous conversionreactions containing largely purified, brain-derived PrPC and synthetic poly(A) RNA(Figure 2) (4). Lipids and perhaps othermolecules copurify with the PrPC in thesepreparations and therefore might also beimportant in prion formation. This seminalreport not only provided direct support for thePrP-based prion hypothesis, but also suggested
that other nonprotein molecules, such as RNAmolecules, are critical either as prion compo-nents or accessories in the PrP conformationalconversion (25). The spontaneous instigationof prion formation in these experiments mayrepresent an in vitro equivalent of sporadicCreutzfeldt-Jakob disease, which, for epi-demiological reasons, is widely assumed to bean example of spontaneous prion disease inhumans.
Recombinant PrP-PMCA and RelatedAmplification Reactions
In attempts to further refine the understand-ing of prion composition, PMCA-like ampli-fication reactions [recombinant PrP PMCA(rPrP-PMCA) and quaking-induced conver-sion (QuIC)] have been performed usingpurified bacterially expressed recombinant
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 181
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
Protofilament:fibrillar proteinpolymers that formamyloid fibrils bylateral association
PrPC as a substitute for brain-derived PrPC
(26, 27). PrPres-seeded rPrP-PMCA and QuICreactions produce a protease-resistant, largelyβ-sheet, fibrillar PrP recombinant conversionproduct that serves as a highly amplified markerof subfemptogram amounts of PrPres. Further-more, these and related reactions can be muchfaster than conventional PMCA reactions, im-proving prospects for the development of prac-tical prion assays and diagnostic tests (26–28).However, inoculations of hamster rPrP-PMCAproduct into hamsters have not yet revealedclear evidence of prion infectivity (R. Atarashi,G. Raymond, & B. Caughey, unpublished data).Thus, like the truncated recombinant PrP amy-loid fibrils called synthetic prions (19), the in-fectivity of the rPrP-PMCA product appears tobe much lower than either PrPres itself or theproduct of PrPres amplification in PMCA reac-tions using brain-derived PrPC, if it proves tobe infectious at all. It should be revealing to de-termine whether the apparently vast differencein infectivity between these various PrP aggre-gates is a matter of molecular composition, con-formation, protease resistance, ultrastructure,cofactors, and/or aggregation conditions. Twonotable features of natural PrPC that are lackingon the recombinant PrPC are N-linked glycansand the GPI anchor. Interestingly, these post-translational modifications do not appear to beessential for the generation of infectious pri-ons as indicated by studies of scrapie-infectedtransgenic mice that express PrPC moleculeslacking these modifications (29–31). These ob-servations suggest that conformational detailsand/or non-PrP molecular cofactors are morecritical determinants of prion infectivity.
STRUCTURAL STUDIES OF PrPOLIGOMERS AND FIBRILS
One of the greatest hurdles in understand-ing PrPres is the lack of biophysical techniquessuitable for determining the high-resolutionstructures of noncrystalline fibrillar protein as-semblies. The preparations of PrPres that areof the highest purity have most often con-tained amyloid-like fibrils, which have been
characterized in terms of fibril dimensions(32–34) and secondary structure composition(35–38). Visualization of the protein core ofwild-type prion fibrils can be obscured by heavyglycosylation and GPI anchors (Figure 3a).However, plaques and fibrils containing mostlyunglycosylated and anchorless PrPres have beenisolated from scrapie-infected transgenic miceexpressing only anchorless PrPC (34). Thesefibrils are composed of protofilaments 3.0–3.5 nm in width as measured by negative-staintransmission electron microscopy (Figure 3a).The protofilament widths and twist periodic-ities can vary significantly with scrapie strain,indicating strain-dependent fibril ultrastruc-tures. This expands the list of strain-dependentfeatures of PrPres, which includes secondarystructures (37, 38), proteolytic sensitivities (39),glycoform patterns (40–43), stabilities (44), andconformational templating activities (8).
Electron Crystallographyand Modeling
So far, the highest resolution information de-rived from infectious PrPres preparations hascome from analyses of two-dimensional crystalsthat appeared along with fibrils in certain PrPres
preparations (45) (Figure 3b). Image process-ing and electron crystallography of these struc-tures revealed that they were likely composedof PrP trimers. On the basis of molecular mod-eling and mutational analyses, a β-helix modelwas proposed (46) (Figure 3c). In this model, aflexibly disordered domain and helix 1 of PrPC
are coiled into a β-helix, which then aligns withthe β-helices of two other PrPres monomersto form a trimer. N-linked glycans are dis-played on the outside of the trimer. An alter-native model has been developed on the basisof molecular dynamics simulations and dockingprocedures (47, 48) (Figure 3c). In the result-ing spiral model, monomers are joined togethervia intermolecular β-sheets and can be readilyassembled into continuous twisting filaments.Both the β-helix and spiral models are close tobeing consistent with the protofilament dimen-sions measured in the anchorless PrPres fibrils
182 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
Wild type Anchorless
Spiral model
Glycan
β-helix model
ba
c
d
α-helix
β-strand
Figure 3Ultrastructures and models of PrPres fibrils. (a) Negative-stained transmission electron micrographs ofproteinase K-treated wild-type or GPI-anchorless PrPres amyloid fibrils (34). (b) Electron micrographs andcrystallographically refined images of hexagonal two-dimensional (2D) crystals and of proteinase K-treatedPrPres. On the left, the 2D crystals are shown together with fibrils. Rows of dense stain accumulations suggeststacked subunits (arrowheads). Scale bar = 100 nm. Adapted from Reference 45; Copyright 2007 NationalAcademy of Sciences, U.S.A. (c) Top-down view of PrPres trimers according to the spiral (47, 48) and β-helix(46) models. Adapted from Reference 48. (d ) Parallel, in-register β-sheet model of synthetic fibrils of humanPrP residues 120–231. Adapted from Reference 50; Copyright 2007 National Academy of Sciences, U.S.A.
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 183
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
(34); however, the spiral model is more consis-tent with a variety of other biochemical charac-teristics of PrPres (48).
Other Probing of Fibril Structure
Recent studies of synthetic recombinant PrP90–231 amyloids using hydrogen/deuteriumexchange and electron spin resonance pro-vided evidence that residues ∼160–220 couldform parallel, in-register β-sheet structuresthat were unlike either the spiral or β-helixmodels (49, 50) (Figure 3d ). In contrast, solidstate NMR studies of fibrils of recombinant hu-man PrP residues 23–144, which correspond toa Gerstmann-Straussler-Scheinker syndrome(GSS)-associated mutant PrP (Y145stop) in hu-mans, showed that residues 112–144 could forma compact, highly ordered core, whereas the re-mainder of the residues toward the N terminuswere largely unordered (51). Earlier studies ofdisease-inducing fibrils of murine PrP residues89–143 showed that residues 112–124 formedan extended β-sheet conformation without theβ-strands being in a parallel, in-register align-ment (52). These and other recent develop-ments in the analysis of a variety of other prionproteins and peptide amyloids (53–64) are lay-ing the foundation for more detailed inves-tigation of the PrPres structures. Particularlynoteworthy are the first high-resolution X-raydiffraction-based structures of amyloid fibrilsof short peptides determined by the Eisen-berg group (65, 66). One important theme thatemerges from these structures is that amyloidscan be stabilized by short stretches of aminoacid residues that align to form β-sheets, which,in turn, are welded to adjacent sheets by theclose interdigitation of side chains, forming adehydrated “steric zipper.”
The Most Infectious Particle
Assessments of the relative infectivity of PrPres
aggregates of various sizes indicated that themost infectious particle per unit protein is∼300–600 kDa, or the mass equivalent of 14–24PrP monomers, and the smallest oligomer with
cell-free PrPC-converting activity is larger thana PrP pentamer in the presence of a low con-centration of sodium dodecyl sufate (13). Thesestudies provide evidence that as long as the in-fectious particles are above a certain size, pre-sumably related to a stability threshold of somesort, it is the concentration of particles ratherthan the concentration of PrPres molecules perse that correlates best with an infectivity titer.Still, the multimeric nature and heterogeneityof infectious PrP oligomers, as well as uncer-tainties about the role of other potential molec-ular constituents, greatly complicate moredetailed analyses of their structures by conven-tional techniques of structural biology.
BIOGENESIS OF PrPres
One of the difficulties in reconstituting prionpropagation with defined molecular con-stituents may be the recapitulation of the mem-brane microenvironment that supports PrPres
formation in vivo. The PrPC polypeptide isfirst synthesized as a nascent GPI-anchored gly-coprotein in the endoplasmic reticulum (ER),subjected to glycan modifications in the Golgiapparatus, and then transported to the cell sur-face (Figure 4), where it is associated primar-ily with cholesterol-rich raft membrane subdo-mains. In the case of familial TSE-associatedmutant PrPC molecules, spontaneous foldingabnormalities can occur in the ER and/orGolgi apparatus (reviewed in Reference 67).In scrapie-infected cells, the conversion of ma-ture PrPC to PrPres occurs on the cell surfaceand/or in endosomal vesicles that are internal-ized from the cell surface (68–70). In contrast toPrPC, PrPres is resistant to detergent solubiliza-tion, proteinase K, and phosphatidylinositol-specific phospholipase C, which cleaves theGPI anchor. PrPres, with the apparent assis-tance of cofactors, directly induces PrPC con-version via an ill-defined templating or seedingmechanism, which results in PrPC joining thePrPres oligomer/polymer. These apparent co-factors include sulfated GAG-containing pro-teoglycans (71–73) and a laminin receptor andits precursor (LRP/LR) (74). Although the
184 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
Cu2+ N
GPI
Glycans
ERAD
Proteasome
ER
Golgi
Aggregation,neurotoxicity
Endosome
Lysosome
Fibrils, plaques
PrPres
Nucleus
PrPC
Autophagosome
Escape fromvesicle?
Aggresome
Autophagy?
Plasmamembrane
Cytoplasm
a
b
Plasma membrane
Cell surface
Cytoplasm
Figure 4Model of biogenesis and accumulation of PrPres in scrapie-infected cells. As a glycophosphatidylinositol (GPI)-anchored plasmamembrane glycoprotein (a), PrPC is first synthesized in the endoplasmic reticulum (ER), processed in the Golgi apparatus, andtransported to the cell surface (b). PrPres, together with apparent cofactors, directly induces the conversion of GPI-anchored PrPC onthe cell surface and/or in endosomes. PrPC that is released from the cell may be converted on extracellular deposits such as amyloidfibrils. Once PrPres is made, it can accumulate on the cell surface, in intracellular vesicles (e.g., lysosomes) and aggresomes, or inextracellular deposits. Under conditions of mild proteasome inhibition, cytotoxic cytoplasmic PrP aggregates (e.g., aggresomes) can befound (157, 158). Scrapie infection alone can inhibit proteasomes, apparently owing to the presence of cytoplasmic PrP oligomers (140).
mechanistic role of these accessory moleculesis unclear, there is evidence that each can in-teract directly with PrP molecules and mighttherefore (a) prime PrPC conformationally forconversion, (b) preorganize or orient PrPC
and PrPres molecules for efficient conversion,and/or (c) help sequester PrPC and PrPres
together in the membrane microdomain orsubcellular compartment, where efficient con-version occurs. Another possible “cofactor” inconversion could be membrane surfaces, whichlike GAGs, can be polyanionic and induceconformational changes in PrP molecules (see
below). The importance of membrane domainstructure is indicated by the influence of al-terations in raft membrane components, suchas cholesterol (75), cholesterol esters (76), andsphingolipids (77) on PrPres formation, but themechanism of such effects is not clear.
FIBRILLAR VERSUSNONFIBRILLAR PrPres DEPOSITS
Once PrPres is made, it can accumulate onthe cell surface, in intracellular vesicles (e.g.,lysosomes), or in extracellular deposits such as
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 185
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
amyloid plaques (Figure 4). Amyloid plaquesare composed of bundles of fibrillar proteinaggregates with a core β-sheet secondary struc-ture that typically gives a cross-β X-ray diffrac-tion pattern with characteristic staining byCongo red and thioflavins S and T. In mosttypes of TSE diseases in natural hosts express-ing GPI-anchored PrPC, amyloid plaques areuncommon or absent altogether. More typi-cal PrPres deposits, such as those described atthe light microscopy level as granular or dif-fuse punctate patterns (Figure 5), are usuallyassociated with membranous ultrastructuresthat lack visible fibrils (78). It is unclearwhether these PrPres deposits are structurallydistinct from amyloids or simply amyloid fib-rils whose visualization is impaired by theirsmall size, irregular length, or association withmembranes and/or other structures. It hasbeen reported that amyloid fibrils of PrPres
(scrapie-associated fibrils or prion rods) that arerecovered in purified preparations typicallyarise only when PrPres is extracted from mem-branes with detergent (79). Because the evi-dence that PrPres can exist in a stable, bio-logically relevant monomeric state is tenuous,such a result could be consistent with PrPres
existing as a small oligomer or aggregate inits membrane-bound state and assembling intofibrils upon extraction from membranes. A ra-diation inactivation target size of ∼55 kDa sug-gests that a dimeric form of PrPres might be thekey infectious unit in liposomes (80). By con-trast, detergent-extracted PrPres preparationscan have regular nonfibrillar two-dimensionalarrays of PrPres trimers (45, 46), whereas size-based fractionations of PrPres particles in deter-gents indicate that PrPC-converting activity isassociated with oligomers larger than PrP pen-tamers (13). It is clear that PrPres can also existin much larger highly stable ordered aggregatesunder a wide variety of conditions both in vitroand in vivo, with amyloid fibrils and plaquesthe most obvious examples in brain tissue(Figure 5). Further study is required to deter-mine the nature of membrane-bound states ofPrPres in vivo and whether they include individ-ual PrPres monomers or small oligomers.
Wild type
Anchorless
Figure 5Different patterns of PrPres deposition in braintissue of scrapie-infected mice expressing wild-type(top) versus anchorless PrPC (bottom) (29). Thesections shown are from the hippocampus in thevicinity of the dentate gyrus. Adapted fromReference 104.
Although rare in many TSE diseases, amy-loid plaques are especially prominent in humanswith familial GSS and kuru and also in scrapie-infected transgenic mice expressing onlymutated PrPC that lacks a GPI anchor and issecreted from cells (29). In GSS patients, theabnormal PrP plaque amyloid that accumulatesis composed primarily of truncated internalPrP fragments (e.g., residues 82–153) that lackthe GPI anchor. In scrapie-infected anchor-less PrP transgenic mice, PrPres accumulatesalmost exclusively in prominent extracellularamyloid plaques that assemble around brainblood vessels (Figure 5). These perivascularplaques have close morphologic similarity tothe cerebrovascular plaques of Alzheimer’s dis-ease (AD), including initial fibrilization withincapillary basement membranes. This suggests
186 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
that both the concentration of PrPres multimersand extracellular matrix components within theendothelial basement membranes are impor-tant for the conversion of diffusible forms ofPrP into fibrillar amyloid (71, 73, 81, 82). Be-cause plaques are formed in several tissues ofthe scrapie-infected anchorless PrP transgenicmice, the anchorless PrPC substrate must be ca-pable of being secreted by a number of differentcell types and diffusing to the amyloid plaquesfor conversion at the growing tips of fibrils.
TSE PRION TRANSMISSION,UPTAKE, AND TRANSPORTMECHANISMS
TSE prions can be naturally or experimen-tally transmitted to new individuals via in-oculation into several peripheral sites withsubsequent and critical spread to the centralnervous system (CNS). After oral challenge,infectivity can be taken up by gut mu-cosa and Peyer’s patches and then be trans-ferred somehow to intestinal nerve end-ings (83–85). However, orally acquired priondiseases do not invariably affect lymphoidtissues or the peripheral nervous system.In such situations, hematogenous neuroin-vasion, most probably involving specializedcapillaries within the circumventricular or-gans of the brain, is a likely alterna-tive route (86). Hematogenous neuroinvasion
can also occur following blood transfu-sion from infected donors (87). With intra-tongue inoculations, transport to the CNSoccurs efficiently via cranial nerves (88).Clearly, the spread of prion infections to andwithin the CNS requires the transfer of prionswithin and between cells.
PrPres Uptake and Acute Inductionof PrPC Conversion
A variety of cultured cell types are capa-ble of sustaining prion infections. Neuronalcells can endocytose PrPres aggregates andtransport them (Supplemental video; fol-low the Supplemental Material link fromthe Annual Reviews home page at http://www.annualreviews.org) via acidic transportvesicles throughout the cytoplasm and alongneuritic projections to points of contact withother cells (Figure 6) (89). Concurrently, newPrPres formation and chronic scrapie infectionscan be established (89, 90). The uptake mech-anism does not appear to be specific to GPI-anchored PrPres because similar uptake andvesicular transport can occur with other typesof amyloid fibrils, such as those composed ofAlzheimer’s β-peptide or anchorless recombi-nant PrP. Uptake of exogenous PrPres can de-pend upon heparan sulfate-containing proteo-glycans [(72, 91), but also consider Reference92] and the 37/67-kDa laminin receptor (93),
Figure 6Neuritic transport of PrPres during acute infection. SN56 mouse neuroblastoma cells were treated withAlexa Fluor 568-labeled mouse PrPres and imaged at 4 d postinfection. Arrows (left panel ) indicate examplesof fluorescent PrPres particles transported within neuritic processes that contact other neurites and cells (89).The left panel shows Alexa Fluor 568 fluorescence. (right panel ) Alexa Fluor 568 fluorescence issuperimposed on a differential interference contrast image.
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 187
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
but not PrPC (89, 90, 92). However, cell surfacePrPC expression is obviously required for newPrPres formation and the maintenance of scrapieinfections. Presumably, contacts between in-coming PrPres and the endogenous PrPC canbegin on the cell surface, but the subcellu-lar sites at which the PrPC is converted tonew PrPres remain unclear. Virtually all of thePrPres produced in cultured N2a neuroblas-toma cells is internalized and rapidly truncatedat the N terminus by endolysosomal proteases(69). Evidence for similar events can be found inscrapie brain and lymphoid tissues (94). How-ever, much of the PrPres is only visualized in vivoat the cell membrane, where it usually remainsfull length, suggesting that it may be formed andinitially accumulated on the cell surface withoutbeing endocytosed and exposed to intracellularproteases.
Although isolated detergent-extractedPrPres can instigate PrPC conversion andchronic infection in neuroblastoma cells,PrPres that is still associated with membranousvesicles is much more efficient at doing so perunit of PrPres (90, 95). Cell-free conversionreactions have shown that, when PrPC is GPIanchored to membranes, PrPres can much morereadily induce PrPC conversion when the twoisoforms are attached to the same membrane(96, 97). By contrast, if the GPI anchor of PrPC
is cleaved by a phospholipase, then it is freeto be converted by PrPres in a separate mem-brane. Interestingly, PrPC need not be fullydissociated from its membrane to be convertedby PrPres on a separate membrane. For exam-ple, GPI-anchorless PrPC can bind to certainmembranes with the help of residues 34–94in the flexible N-terminal domain and, in thisconfiguration, can be converted by PrPres onseparate membrane vesicles (96). Altogether,these findings support the view that the GPIanchoring of PrPC to membranes constrains itsinteractions with PrPres such that conversionis only efficient when PrPres is anchored in cisto the same membrane. However, in apparentcontradiction to this conclusion are studiesshowing that TSE infectivity tightly bound
to steel surfaces can readily induce infectionsin vivo (98). At present, it is not clear howto reconcile these observations unless cellscan extract PrPres/infectivity from the wires.Another possibility is that a distinct orientationof steel-bound PrPres, perhaps with exposedGPI anchors, allows it to induce the conversionof membrane-bound PrPC without dissociatingfrom the steel.
CELL-TO-CELL SPREADOF TSE PRIONS
Once an individual cell is propagating prions,a central unresolved issue is how the infec-tion then spreads to other cells. Cell proxim-ity appears to be important for efficient spreadin cell culture (98a, 99). The greater appar-ent specific infectivity of infectivity associatedwith membrane-bound PrPres (95), as well asthe greater efficiency of conversion when GPI-anchored PrPC and PrPres are inserted intothe same membranes (96, 97), led us to pro-pose that the cell-to-cell spread of infectionmight occur primarily via transfer of PrPres-containing membrane particles (Figure 7) (97).Fusion of such particles into recipient cellscould insert PrPres into the plasma membranein a topology compatible with conversion ofGPI-anchored PrPC molecules.
Exosomes
Consistent with this idea are observations thatPrPC, PrPres, and scrapie infectivity can be re-leased from cultured scrapie-infected cells in as-sociation with small (50–90 nm) vesicles calledexosomes (100). Exosomes are assembled in cy-toplasmic organelles known as multivesicularbodies and secreted via exocytosis (Figure 7).Released exosomes can then fuse with othercells and deliver their cargos. Appropriately,the topology of exosomes is such that exofa-cial membrane proteins, such as PrPres, wouldmaintain an exofacial topology upon fusion withthe plasmalemma of a new cell. Both nonneu-ronal and neuronal prion-infected cell lines canrelease exosomes that are infectious for mice,
188 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
PrPC
PrPres
Exosome
MVB y x
z
Top view(DIC)
Top view(merged)
Side view(merged)
a b
Green fluorescent protein
mCherry fluorescent protein
13
2
Figure 7Potential mechanisms of intercellular spread of prions. (a) Exosomes/membrane particles. Prion-infectedcells release PrPres-containing membrane vesicles that deliver PrPres (red squares) to membranes ofuninfected cells to initiate new PrPres formation (blue squares) in recipient cells (95–97). These includeexosomes, vesicles generated by invagination of the limiting membranes of multivesicular bodies (MVBs)and released by MVB fusion with the plasma membrane (100). Delivery of membrane vesicles could occur atthe cell surface (not shown) or in an endocytic compartment. (b) Tunneling nanotubes (TNTs). TNTs arevisualized as thin intercellular projections (number 1) through space (number 2) in a coculture of two cell linesexpressing either a GPI-anchored protein tagged with green fluorescent protein or one tagged withmCherry fluorescent protein. Number 3 (white area) shows a TNT that has facilitated the transfer ofGFP-tagged protein onto the surface of the mCherry-expressing cell. Side view corresponds to a side view ofa three-dimensional volume rendering of the area indicated by the dotted rectangle in top view (merged ).DIC, differential interference contrast image.
supporting the concept that exosomes may bea significant mechanism for the spread of theseinfections in vivo (100, 101). However, exten-sive efforts have failed so far to visualize ex-osomes in brain tissue (78). Exosomes can befound in germinal centers of lymphoid tissues,where they derive from B cells and not folliculardendritic cells, the cell type that putatively am-plifies infectivity. In scrapie infected follicles,B cell–derived exosomes lack demonstrableabnormal PrP (102).
Tunneling Nanotubes
Another potential spreading mechanism is thetransfer of membrane-associated PrPres be-tween cells via recently described entities called
tunneling nanotubes (89, 103, 104). Tunnelingnanotubes are thin membranous bridges thatcan form between cells and mediate the transferof organelles, plasma membrane components(including GPI-anchored proteins), cytoplas-mic molecules, and pathogens (Figure 7) (105).Interestingly, tunneling nanotubes can trans-port vesicles that, like those responsible for neu-ritic transport of PrPres (89), are small, acidicand of endosomal/lysosomal origin. Althoughtheir existence remains to be documented invivo, tunneling nanotubes have been observedin a variety of cultured cell types (105). For ex-ample, murine retroviruses are known to exploittunneling nanotube-like cell bridges to spreadbetween cells (105a). These viruses can inducethe outgrowth of filapodia from uninfected cells
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 189
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
PrPd: disease-specificisoform of PrPdetected byimmunologicalstaining of tissueswithout implying anyother biochemical orinfectious properties
to virus-infected cells and then migrate alongthe bridge back to the target cell. It is no-table that murine retroviruses are also knownto enhance the release of scrapie prions intothe medium of cultured cells (106). In the latterstudy, the prions were found in exosome-likestructures; however, it is possible that vesicularstructures can also be derived from the shearingof tunneling nanotubes.
PrP-MEMBRANE ASSOCIATIONAND PATHOGENESIS
The usual proximity between PrP moleculesand cellular membranes raises the issue of theinfluence of these interactions in the neurode-generative pathology of TSE diseases. Theanchorless PrP transgenic mouse model pro-vides striking evidence that, although the lackof GPI-anchored PrP does not prevent PrPres
and prion accumulation, it profoundly altersthe manifestations of scrapie infections (29).Indeed, most of the typical clinical and neu-ropathogical characteristics of scrapie are eitherabsent or greatly reduced in these mice despitethe accumulation of brain PrPres to levels com-parable to those in scrapie-infected wild-typemice. This reduced brain damage could be dueeither to a need for anchored PrPC on braincells for toxicity induced by PrPres and/or to alower pathogenicity of PrPres amyloid plaquescompared to the more dispersed, amorphous,and membrane-associated PrPres deposits seenin most other TSE/prion diseases. This lattersituation would be consistent with the emerg-ing view that, in other protein-misfolding dis-eases, such as AD, it is not amyloid plaques butsmaller misfolded protein oligomers that arethe most neuropathologic (107, 108). It is likelythat potentially toxic protein aggregates are rel-atively benign when deposited in the extracel-lular space as plaques with relatively low sur-face area-to-mass ratios. Moreover, microgliatend to surround amyloid plaques and may sealthem off from exposure to the rest of the braintissue while attempting to degrade them [e.g.,(109, 110)]. This is not to say that PrPres amy-loid deposits are innocuous because they clearly
can be associated with severe pathogenic lesionsand clinical disease. However, these pathogeniceffects are different and appear to be slowerto result in clinical deficits than those associ-ated with diffuse types of PrPres deposits (29; B.Chesebro, unpublished data).
In TSE-infected wild-type hosts expressingGPI-anchored PrPC, a number of ultrastruc-tural neuropathological lesions of neuronaland/or astrocyte membranes are consideredpathognomonic. These include membrane pro-liferation and microfolding, highly abnormalcoated pits, tubulovesicular bodies, and spi-ral membrane inclusions of axons (111). Otherfindings that are typical, but not specific forTSEs, include vacuolation, gliosis, degenerateprocesses, dystrophic axons, autophagic vac-uoles, clumped synaptic vesicles, and neuronaldeath. TSE-associated pathology often involvesmembrane distortions in proximity to the ac-cumulation of what has been aptly describedas disease-associated PrP (PrPd) because it canbe labeled specifically in histological sectionsof infected hosts without determination of thevarious biochemical properties that are usuallyused to identify PrPres. PrPd accumulation cor-responds ultrastructurally with abnormal en-docytosis, increased endolysosomes, and bothfinger-like extensions and invaginations (i.e.,microfolding) of plasma membranes (Figure 8)(78). Extracellular accumulations of PrPd arealso observed, as well as suggestions of intercel-lular transfer of PrPd between neurons and/orglia. PrPd can appear to fill the narrow gap(∼10 nm) between the closely opposed plas-malemmas (Figure 8a). Some aberrant mem-brane structures, notably tubulovesicular struc-tures, appear to lack PrPd (78). Tubulovesicularbodies are small (20–35 nm), irregular sphericalor short tubular structures that are packed intodendrites and occasionally axon terminals (112,113). Tubulovesicular body clusters have someresemblance to multivesicular bodies but differin their smaller size, lack of a limiting mem-brane, and absence of PrPd, which suggests thatthey are not the same as the PrPres-containingexosomes that may mediate transfer of infectiv-ity between cultured cells.
190 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
a
b
c
d e
***
Figure 8Abnormal PrP (PrPd) accumulation and membrane distortions on the surface of neurons and astrocytes inscrapie-infected sheep brain tissue. PrPd is labeled with immunogold particles. (a) PrPd on the plasmamembrane of a neuron without morphological changes. (b) Complex dendrite membrane disturbances(microfolding shown by asterisks) associated with the formation of abnormal pits (not immunolabeled).(c) PrPd accumulation associated with the formation of excess coated pits (arrows) and with the transfer tomembranes of adjacent processes (arrow heads). (d ) Coated membrane invaginations, one of which shows aspiral, twisted neck (not immunolabeled). (e) PrPd on the plasma membrane of astrocytic processes,containing abundant glial filaments. Parts of the plasmalemma of some processes have linear segments(arrows) suggesting increased membrane rigidity and incipient fibril formation. Adapted from Reference 78.
How Might Membrane-AssociatedPrPres Be Neurotoxic?
The importance of GPI anchoring of PrPmolecules in TSE pathogenesis is likely con-nected mechanistically to the distortions of
cellular membranes and membrane traffickingassociated with TSE disease. When PrPC is an-chored to membranes by a GPI (or GPI-like)moiety, it typically assumes the α-helical con-formation of native soluble PrPC (114, 115).The anchoring and heavy glycosylation may
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 191
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
stabilize wild-type PrPC in this conformation,reducing the likelihood of spontaneous conver-sion to PrPres during the life span of the host.Although isolated, glycosylated and GPI-linkedPrPC can oligomerize spontaneously into aform with higher β-sheet content when ex-posed to raft-like membranes in vitro (116),it is not clear whether the GPI anchors actu-ally penetrate the lipid bilayer in this experi-mental system. In the absence of GPI anchors,full-length PrPC and PrP90–231 can bind tolipid membranes and undergo major changesin conformation, protease resistance, and ag-gregation state (96, 117–122). In doing so, suchPrP molecules can disrupt the lipid bilayer (120,122) and, in the case of the GSS-associatedmutant PrP fragment 82–146, form channels(118). Moreover, oligomers of full-length an-chorless PrP can be neurotoxic when added toculture media or inoculated into the brain (123).When anchorless PrP is artificially expressedin the cytoplasm rather than in the secretorypathway, its interaction with membranes cor-relates with cytotoxicity (119). Despite thesemultiple observations of seemingly unhealthyinteractions between anchorless PrP moleculesand membranes, the fact remains that trans-genic mice that express anchorless PrP are nor-mally healthy and when infected with scrapiehave neuropathological manifestations that arequite different from those of wild-type mice.This again highlights the GPI anchor as a keyarbiter in TSE pathogenesis in vivo.
Given that PrPres tends to be oligomeric,membrane-associated, and difficult for cellsto degrade, it is possible that the mere ac-cumulation of PrPres aggregates on mem-branes compromises fundamental membranefunctions. Because most PrPres molecules haveGPI moieties (124), the GPI anchoring ofPrPres oligomers to membranes should be mul-tivalent. Multivalent membrane interactionsshould constrain the quaternary interactionsbetween PrPres molecules within the plane ofthe membrane and may promote sheet-like lat-eral structures (125) compatible with the two-dimensional PrPres crystals visualized by Willeet al. (45). By contrast, whole-cell atomic force
microscopy suggests that fibrillar PrPres aggre-gates might be present on the surface of scrapie-infected cells in culture (126).
Whatever the preferred PrPres quaternarystructure(s), it is likely that the accumulationof any relatively large PrPres aggregate that ismultiply anchored to membranes could dis-tort its local structure, composition, flexibility,fluidity, dynamics, integrity, and, hence, func-tionality. The presence of PrPd in abnormalspiral clefts at the cell surface and in endosome-derived subcellular structures points to aber-rant attempts by cells to internalize membranescontaining PrPd aggregates (Figure 8). Mem-branes containing intense PrPd accumulationcan also be unusually linear and close to shortextracellular fibrils (Figure 8e). This suggeststhat PrPd accumulation might impart an in-creased rigidity to membranes, which could af-fect PrPd internalization and degradation aswell as normal endocytic processes. PrPd of-ten appears to be associated with ubiquitin andclathrin on opposite sides of lipid bilayers ofspiral clefts and internalized coated membra-nous structures (78). Thus, ubiquitin taggingand the membrane-inverting forces of clathrincould play roles in the generation of these dis-torted structures. The packing of PrPd betweenclosely opposed membranes of adjacent cells(Figure 8a) and into narrow spiral clefts (78)also suggests that PrPd, alone or in conjunc-tion with other molecules, may cause abnormaladhesion between membrane surfaces.
A Pathological Role for GPI-AnchoredPrPC on Neurons?
Following infection, transgenic PrP knockoutmice that lack PrP altogether (127) or turnoff PrPC expression in adulthood (128) do notexhibit the symptoms of TSE disease. Thus,although other recent data suggest that someaspects of neurological disease associated withinherited PrP mutations might be due todeficits in the apparent neuroprotective activ-ities of PrPC (129), TSE pathogenesis doesnot appear to be due simply to a loss of PrPC
function. On the contrary, there is evidence
192 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
that clinical disease may require the presenceof PrPC as well as PrPres. For instance, wild-type brain tissue grafts in scrapie-infected PrPknockout mice produce PrPres that does notcause neuropathology in surrounding tissuethat lacks PrPC (130). Moreover, Mallucci andcolleagues (131) have shown that cessation ofPrPC expression in the neurons of adult scrapie-infected transgenic mice reverses spongiosisand delays clinical disease long term withoutstopping further accumulation of PrPres pro-duced in other cell types (131). In addition,transgenic mice expressing PrPC exclusively inneurons are susceptible to scrapie (132). Theseresults suggest that neuronal PrPC expressionis needed to mediate most PrPres-induced neu-ropathogenesis. However, this is not alwaystrue because Raeber and colleagues (133) haveshown that TSE disease can develop in scrapie-infected transgenic mice that express PrPC onlyin astrocytes and not to any detectable extent inneurons. The reason for the apparent discrep-ancy between the Mallucci and Raeber resultsis not clear but could relate to differences inthe strain and species of prion, the sequenceof expressed PrPC, and/or the levels of PrPC
expression in the respective transgenic mousemodels.
Direct and Indirect Mechanismsof PrPres Neurotoxicity
Collectively, these studies leave open the possi-bilities of both direct and indirect mechanismsof PrPres neurotoxicity, which may vary in im-portance between various TSE disease mod-els or neuroanatomical sites within hosts. Di-rect PrPres toxicity might be mediated by themembrane disturbances described above, whichcould have profound effects on processes suchas neuronal homeostasis, intercellular contacts,synaptogenesis, synaptic functions, and axonaltransport. Indirect effects of PrPres could be me-diated by perturbations of glial functions that,in turn, cause neuronal lesions (112, 134, 135).Alternatively PrPres could corrupt, rather thanknock out, PrPC function (136). Transgenicmice expressing various mutant PrPC molecules
have revealed that perturbations of PrPC struc-ture, expression level, and/or functions alonecan cause neurological disease, even withoutTSE infection (reviewed in Reference 136). Itfollows then that the accumulation of PrPres
might cause disease by altering the metabolismor activities of PrPC. To decipher these pos-sibilities, it would be helpful to understand thephysiological function of PrPC, but this remainselusive (104, 137). Nonetheless, a number ofstudies have provided evidence that PrPC hascytoprotective, antiapoptotic activities againstvarious cellular stresses (reviewed in References104, 136, and 137). At the same time, injec-tions of PrP antibodies into the brain can leadto neuronal apoptosis, providing evidence thatPrPC cross-linking or interference with PrPC-ligand binding can have adverse consequences(138). Thus, it is possible that contact betweenPrPres and PrPC could have similar effects inTSE-infected individuals. It has been proposedthat PrPC interacts with a physiological ligandin two sites and that disruption of one of thoseinteractions can elicit cell death (139). Still an-other possibility is that an intermediate or by-product of PrPres formation is more neurotoxicthan mature PrPres itself. This scenario couldaccount for the enhanced neuropathogenesisobserved in scrapie-infected hosts expressingGPI-anchored PrPC in neurons. Interestingly,small amounts of PrPres have been detectedin the cytosol and can inhibit the ubiquitin-proteosome system, with oligomeric aggrega-tion intermediates being the most inhibitoryin this system (140). At this point, it is un-clear which of these direct and indirect mech-anisms of PrPres-associated neuropathogenesis,or some combination thereof, is most impor-tant in TSE diseases. It seems unlikely that asingle discrete mechanism is at play in all TSEdiseases, given their phenotypic diversity.
STRETCHING THE REALMOF PRIONS
Since Prusiner’s coining of the term prionand Wickner’s expansion of the prion conceptto fungal epigenetics, a number of biological
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 193
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
entities have been described as prions or prionlike. These include both functional (reviewed inReferences 141 and 142) and disease-associatedamyloids in a variety of organisms. The pri-ons of Saccharomyces cerevisiae and other fungihave become the best characterized both genet-ically and biochemically (reviewed in Reference143). The [URE3], [PSI+], [PIN+], and [Het-s]prions are amyloid forms of the normallysoluble proteins Ure2p, Sup35p, Rnq1p, andHETs, respectively. The prion forms of Ure2pand Sup35p are inactive and result in loss-of-function phenotypes for the cell. Rnq1p amy-loid primes yeast for [PSI+] formation (144),but the function of soluble Rnq1p is unknown.The HETs protein function is also unknown,but its amyloid (prion) form is active in mediat-ing heterokaryon incompatibility in Podosporaanserina (145). Thus, unlike most prions, the[Het-s] prion may play a positive physiologicalrole in the host organism. Wickner & Roberts(145a) have also reported evidence for a novelnonamyloid prion of S. cerevisiae, [β], whichtakes the form of a self-activating vacuolar pro-tease B that is necessary for meiosis and aidssurvival in the stationary phase of the cell cycle.Even a self-activating protein regulatory net-work, as opposed to an altered form of an indi-vidual protein, has been shown to share manyproperties with fungal prions. In this case, a self-activating MAP kinase cascade appears to be re-sponsible for a non-Mendelian hereditary unitcalled [C], which is responsible for inheritanceand possibly the cell-to-cell spread of the crip-pled growth phenotype in P. anserina (146).
Whether in fungi or mammals, the ability ofamyloids to nucleate or seed their own growthgives them self-propagating activity that is atleast partly analogous to that of prions. In-deed, some experimental amyloidoses in mam-mals have shown evidence of being transmis-sible in the sense that amyloid taken from anamyloidotic individual can sometimes greatlyaccelerate amyloidosis in another individual.However, it is important to emphasize thatsuch “transmissions” have only been shownunder unnatural, experimental conditions inwhich the recipient animal is highly primed for
susceptibility to amyloidosis and would be ex-pected to develop amyloidosis spontaneouslygiven time.
AA Amyloidosis
One prototypic example of an apparently trans-missible amyloid disease is AA amyloidosis, amalady of humans and animals involving thewidespread accumulation of serum amyloid A(AA) protein amyloid, especially in the kidney,liver, and spleen. Most cases of AA amyloido-sis in mammals appear to occur spontaneouslyowing to chronic inflammation or genetic pe-culiarities that elevate blood levels of AA andpredispose the organism to AA amyloid for-mation. However, it has long been known thatinoculation of extracts of material from amy-loidotic mice (a.k.a., amyloid-enhancing fac-tor) into other mice can rapidly induce amy-loidosis as long as the recipients are themselvesstrongly primed for amyloidosis by proinflam-matory treatments such as silver nitrate injec-tions (147). Evidence now points to aggregatedAA as the amyloid-enhancing factor (148). Aswith the synthetic prion experiments discussedabove (19), these experiments raise the questionof whether the amyloid inoculum initiates dis-ease, as is clearly the case with natural TSE pri-ons, or merely enhances an ongoing pathogenicprocess.
AA amyloidosis is wreaking havoc in thecaptive cheetah population, complicatingefforts to rescue this endangered species fromextinction (149, 150). Amyloidotic cheetahsshed AA amyloid in the feces, and this fecalamyloid can accelerate AA amyloidosis wheninjected intravenously into silver nitrate–primed mice (151). Given that murine AAamyloid can accelerate amyloidosis in micewhen administered orally, these results suggesta plausible fecal-oral route for the naturaltransmission of AA amyloidosis in cheetahs.More study is required to demonstrate theactuality of such a transmission cycle. Ifproven to be true, cheetah AA amyloidosismight become the first known naturally trans-missible, non-PrP prion in mammals. Even
194 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
so, the question of whether the transmittedamyloid initiates or enhances amyloidosis innaive cheetahs will remain. Moreover, onestriking characteristic of captive cheetahsrelative to those in the wild is their chronicinflammation and high blood AA levels (150).These factors could make them susceptible tospontaneous amyloidosis, as is the case withsilver nitrate–treated mice. If so, then the highincidence of amyloidosis in captive cheetahsmight be due to spontaneous disease rather thaninfections with amyloid from other animals.
Apolipoprotein AII Amyloidosis
Apolipoprotein AII (ApoAII) is an amy-loidogenic protein that causes mouse senileamyloidosis and a hereditary human diseaseowing to a mutation in the APOA2 gene. Likeexperimental AA amyloidosis, mouse senileamyloidosis affects visceral organs and can bestimulated by the experimental administrationof exogenous amyloid via the gastrointestinaltract, possibly after consumption of ApoAIIamyloid in the feces of amyloidotic cage mates(152). ApoAII amyloid accumulation alsooccurs spontaneously in aged mice, raisingthe possibility that inoculated ApoAII amyloidmerely accelerates systemic amyloidosis.
Aβ Amyloidoses
In AD, cerebral Aβ angiopathy, and, to some ex-tent, in many aged humans who are cognitivelynormal, Aβ, a small proteolytic fragment ofthe Aβ precursor protein (βAPP), accumulatesin amyloid deposits in the brain. Whether theamyloid form of Aβ is the primary pathogenicentity in AD remains a matter of debate, withrecent evidence leaning toward smaller sub-amyloid Aβ oligomers being more neurotoxic(107, 108). Although there is no evidence thatAD per se is transmissible, studies in transgenicmouse models of AD have suggested that Aβ
amyloidosis can be stimulated by intracerebralinjections of brain extracts from AD patients oramyloidotic βAPP transgenic mice (153). Onceagain, as was the case with the AA and ApoAII
amyloidosis inoculation studies noted above,it is unclear whether the Aβ amyloid causedamyloidosis or merely accelerated an ongo-ing pathogenic process because the transgenicrecipients ultimately develop deposits sponta-neously without inoculation exogenous of theAβ amyloid extracts. Another major unresolvedissue, when comparing this amyloid-inducingactivity with a TSE prion disease, is the extentto which the induced Aβ-amyloidotic lesionscan spread from localized sites of seeding bythe inoculum to other sites within the host.
Cytoplasmic PolyadenylationElement-Binding Protein
Another protein that has been describedas having prion-like properties is cytoplas-mic polyadenylation element-binding protein(CPEB) (154). The behavior of ectopically ex-pressed neuronal CPEB or its N-terminal do-main in yeast led Si and coworkers to pro-pose that distinct CPEB states in neuronsmight serve as a mechanism for stabilizingplastic changes in synapses. In their model,a self-perpetuating aggregated state of CPEBis induced in activated synapses, altering lo-cal mRNA translation and helping to sustainlong-term facilitation. Although this is an in-triguing potential mechanism for synaptic reg-ulation, it would be difficult to consider it aprion-like phenomenon in neurons because ofthe lack of CPEB-mediated transmission of thephenotypic change to other cells and organisms.
Amyloid Spreading Mechanisms?
Consideration of the above examples of prionor prion-like entities raises the key issue ofspreading mechanisms. It is now commonplaceto find proteins that can exist in two or morestates, with at least one of those states beingable to perpetuate itself via self-seeding or au-tocatalytic activity. Indeed, most proteins arepredicted to be capable of forming amyloidsunder certain conditions (142). As noted above,the ability of a self-perpetuating protein stateto spread between cells and organisms is a
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 195
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
hallmark of prions. In yeast and at least someother fungi, the spread of cytoplasmic prionscan occur through cytoplasmic exchange ortransfer during cell division, mating (in whichcells fuse), or cytoduction (a largely experi-mental abortive mating procedure in whichcytoplasmic mixing occurs while nuclear fu-sion is blocked). In mammals, horizontal prionspreading mechanisms would tend to be muchmore complex, involving host-to-host transfersthrough the environment and tissue barriers, aswell as cell-to-cell transfers without overt cy-toplasmic mixing. With most mammalian cells,mechanisms for the horizontal exchange of cy-toplasmic molecules larger than 1 kDa are notreadily apparent, except perhaps in some casesfor exosomes and tunneling nanotubes, as de-scribed above. The apparent lack of such mech-anisms would argue against the likelihood offinding purely cytoplasmic prions in mammals.
As noted above, many mammalian diseaseslead to the accumulation of extracellular, ratherthan cytoplasmic, amyloid, which might morereadily migrate between cells to spread withintissues and hosts. With inducible systemic amy-loidoses, such as AA and ApoAII amyloidoses,hematogenous spread appears to occur. Blood-
borne amyloid is particularly capable of pene-trating some tissues via fenestrated capillaries,which allow passage of large molecules suchas proteins (155). By contrast, access of suchblood-borne amyloids to the CNS is severelylimited. Furthermore, it is not yet clear whetherextracellular amyloids, such as Aβ amyloid, thatare produced within the brain are capable ofspreading effectively from their sites of initialdeposition to other regions. If it turns out to betrue that various brain amyloids have little ca-pacity to propagate throughout the CNS, or toinvade the CNS from peripheral tissues, thenthe hazards of localized amyloid inoculationswill likely be small. However, if amyloids canpropagate throughout the CNS like TSE pri-ons, then there might be real reason to fear thepossibility of amyloid infections. For the timebeing, it appears that, relative to other self-propagating protein aggregates, TSE prionsare unusually adept at spreading between andwithin mammalian hosts by practical routes.TSEs are also the only prion diseases knownto involve a protein that is normally GPI an-chored, a feature that accounts at least in partfor the unique biology and neurovirulence ofthese devastating transmissible diseases.
SUMMARY LIST
1. Cell-free propagation of TSE prions appears to rule out the need for an exogenous,pathogen-encoding nucleic acid, but polyanions, such as poly(A) RNA, might play animportant role in prion replication.
2. The three-dimensional structure of TSE prions remains largely a mystery, but recenttechnical advances have greatly enhanced the understanding of the molecular architectureof yeast prions and other amyloid fibrils.
3. The ability to propagate within and between hosts is a key characteristic of prions, whichdistinguishes them from other autocatalytic protein states.
4. The GPI anchor distinguishes mammalian prion protein from other amyloidogenic pro-teins and has strong influences on TSE pathogenesis.
5. TSE pathogenesis involves major membrane disturbances in the brain, which may inpart be mediated by the accumulation of abnormal GPI-anchored PrP molecules.
6. Several non-PrP-based mammalian amyloidoses appear to be transmissible under experi-mental conditions. Further studies are required to determine if transmission of such amy-loids between hosts under practical circumstances poses a risk to humans or to animals.
196 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
FUTURE ISSUES
1. What is the complete molecular composition of TSE prions, and what is the reason forthe poor infectivity of amyloid fibrils derived from recombinant PrPC?
2. What are the three-dimensional structures of prions and their different strains?
3. What are the most neurotoxic entities in TSE diseases and other protein misfoldingdiseases?
4. What are the critical neuropathogenic pathways in TSE diseases?
5. How do TSE prions spread from cell to cell and tissue to tissue?
6. What are the natural spreading capacities of disease-associated non-PrP amyloids andamyloid-like protein aggregates? Do they pose risks of infection?
7. How can TSE prion infections be detected early and treated effectively?
8. How widespread and diverse are prions in biology?
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
We thank Drs. Valerie Sim, Kelly Barton, Winslow Caughey, Leah Christensen, and KimHasenkrug for helpful comments on this manuscript. Gary Hettrick provided excellent graph-ics assistance. This work was supported by the Intramural Research Program of the NIAID, NIH.
LITERATURE CITED
1. Griffith JS. 1967. Self-replication and scrapie. Nature 215:1043–442. Prusiner SB. 1982. Novel proteinaceous infectious particles cause scrapie. Science 216:136–443. Castilla J, Saa P, Hetz C, Soto C. 2005. In vitro generation of infectious scrapie prions. Cell 121:195–2064. Deleault NR, Harris BT, Rees JR, Supattapone S. 2007. Formation of native prions from minimal
components in vitro. Proc. Natl. Acad. Sci. USA 104:9741–465. Wickner RB. 1994. [URE3] as an altered URE2 protein: evidence for a prion analog in Saccharomyces
cerevisia. Science 264:566–696. Chien P, Weissman JS, DePace AH. 2004. Emerging principles of conformation-based prion inheri-
tance. Annu. Rev. Biochem. 73:617–567. Prusiner SB, Groth DF, Bolton DC, Kent SB, Hood LE. 1984. Purification and structural studies of a
major scrapie prion protein. Cell 38:127–348. Bessen RA, Kocisko DA, Raymond GJ, Nandan S, Lansbury PT Jr, Caughey B. 1995. Nongenetic
propagation of strain-specific phenotypes of scrapie prion protein. Nature 375:698–7009. Kocisko DA, Come JH, Priola SA, Chesebro B, Raymond GJ, et al. 1994. Cell-free formation of
protease-resistant prion protein. Nature 370:471–7410. Kocisko DA, Priola SA, Raymond GJ, Chesebro B, Lansbury PT Jr, Caughey B. 1995. Species specificity
in the cell-free conversion of prion protein to protease-resistant forms: a model for the scrapie speciesbarrier. Proc. Natl. Acad. Sci. USA 92:3923–27
11. Raymond GJ, Hope J, Kocisko DA, Priola SA, Raymond LD, et al. 1997. Molecular assessment of thetransmissibilities of BSE and scrapie to humans. Nature 388:285–88
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 197
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
12. Caughey B, Kocisko DA, Raymond GJ, Lansbury PT. 1995. Aggregates of scrapie-associated prionprotein induce the cell-free conversion of protease-sensitive prion protein to the protease-resistantstate. Chem. Biol. 2:807–17
13. Silveira JR, Raymond GJ, Hughson AG, Race RE, Sim VL, et al. 2005. The most infectious prionprotein particles. Nature 437:257–61
14. Callahan MA, Xiong L, Caughey B. 2001. Reversibility of scrapie-associated prion protein aggregation.J. Biol. Chem. 276:28022–28
15. Gadjusek DC. 1988. Transmissible and nontransmissible amyloidoses: autocatalytic post-translationalconversion of host precursor proteins to beta-pleated configurations. J. Neuroimmunol. 20:95–110
16. Jarrett JT, Lansbury PT Jr. 1993. Seeding “one-dimensional crystallization” of amyloid: A pathogenicmechanism in Alzheimer’s disease and scrapie? Cell 73:1055–58
17. Caughey B, Raymond GJ, Kocisko DA, Lansbury PT Jr. 1997. Scrapie infectivity correlates withconverting activity, protease resistance, and aggregation of scrapie-associated prion protein in guanidinedenaturation studies. J. Virol. 71:4107–10
18. Hill AF, Antoniou M, Collinge J. 1999. Protease-resistant prion protein produced in vitro lacks de-tectable infectivity. J. Gen. Virol. 80:11–14
19. Legname G, Baskakov IV, Nguyen HO, Riesner D, Cohen FE, et al. 2004. Synthetic mammalianprions. Science 305:673–76
20. Legname G, Nguyen HO, Baskakov IV, Cohen FE, DeArmond SJ, Prusiner SB. 2005. Strain-specifiedcharacteristics of mouse synthetic prions. Proc. Natl. Acad. Sci. USA 102:2168–73
21. Hsiao KK, Groth D, Scott M, Yang SL, Serban H, et al. 1994. Serial transmission in rodents ofneurodegeneration from transgenic mice expressing mutant prion protein. Proc. Natl. Acad. Sci. USA91:9126–30
22. Saborio GP, Permanne B, Soto C. 2001. Sensitive detection of pathological prion protein by cyclicamplification of protein misfolding. Nature 411:810–13
23. Weber P, Giese A, Piening N, Mitteregger G, Thomzig A, et al. 2006. Cell-free formation of misfoldedprion protein with authentic prion infectivity. Proc. Natl. Acad. Sci. USA 103:15818–23
24. Saa P, Castilla J, Soto C. 2006. Ultra-efficient replication of infectious prions by automated proteinmisfolding cyclic amplification. J. Biol. Chem. 281:35245–52
25. Geoghegan JC, Valdes PA, Orem NR, Deleault NR, Williamson RA, et al. 2007. Selective incorporationof polyanionic molecules into hamster prions. J. Biol. Chem. 282:36341–53
26. Atarashi R, Wilham JM, Christensen L, Hughson AG, Moore RA, et al. 2008. Simplified ultrasensitiveprion detection by recombinant PrP conversion with shaking. Nat. Methods 5:211–12
27. Atarashi R, Moore RA, Sim VL, Hughson AG, Dorward DW, et al. 2007. Ultrasensitive detection ofscrapie prion protein using seeded conversion of recombinant prion protein. Nat. Methods 4:645–50
28. Colby DW, Zhang Q, Wang S, Groth D, Legname G, et al. 2007. Prion detection by an amyloidseeding assay. Proc. Natl. Acad. Sci. USA 104:20914–19
29. Chesebro B, Trifilo M, Race R, Meade-White K, Teng C, et al. 2005. Anchorless prion protein resultsin infectious amyloid disease without clinical scrapie. Science 308:1435–39
30. Tuzi NL, Cancellotti E, Baybutt H, Blackford L, Bradford B, et al. 2008. Host PrP glycosylation: amajor factor determining the outcome of prion infection. PLoS Biol. 6:e100
31. Neuendorf E, Weber A, Saalmueller A, Schatzl H, Reifenberg K, et al. 2004. Glycosylation deficiencyat either one of the two glycan attachment sites of cellular prion protein preserves susceptibility tobovine spongiform encephalopathy and scrapie infections. J. Biol. Chem. 279:53306–16
32. Merz PA, Somerville RA, Wisniewski HM, Iqbal K. 1981. Abnormal fibrils from scrapie-infected brain.Acta Neuropathol. 54:63–74
33. Prusiner SB, McKinley MP, Bowman KA, Bendheim PE, Bolton DC, et al. 1983. Scrapie prionsaggregate to form amyloid-like birefringent rods. Cell 35:349–58
34. Sim VL, Caughey B. 2008. Ultrastructures and strain comparison of under-glycosylated scrapie prionfibrils. Neurobiol. Aging doi:10.1016/j.neurobiolaging.2008.02.016
35. Caughey BW, Dong A, Bhat KS, Ernst D, Hayes SF, Caughey WS. 1991. Secondary structure analysisof the scrapie-associated protein PrP 27-30 in water by infrared spectroscopy. Biochemistry 30:7672–80
198 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
36. Pan KM, Baldwin M, Nguyen J, Gasset M, Serban A, et al. 1993. Conversion of alpha-helices into beta-sheets features in the formation of the scrapie prion proteins. Proc. Natl. Acad. Sci. USA 90:10962–66
37. Caughey B, Raymond GJ, Bessen RA. 1998. Strain-dependent differences in beta-sheet conformationsof abnormal prion protein. J. Biol. Chem. 273:32230–35
38. Spassov S, Beekes M, Naumann D. 2006. Structural differences between TSEs strains investigated byFT-IR spectroscopy. Biochim. Biophys. Acta 1760:1138–49
39. Bessen RA, Marsh RF. 1994. Distinct PrP properties suggest the molecular basis of strain variation intransmissible mink encephalopathy. J. Virol. 68:7859–68
40. Somerville RA, Hamilton S, Fernie K. 2005. Transmissible spongiform encephalopathy strain, PrPgenotype and brain region all affect the degree of glycosylation of PrPSc. J. Gen. Virol. 86:241–46
41. Kascsak RJ, Rubenstein R, Carp RI. 1991. Evidence for biological and structural diversity amongscrapie strains. In Transmissible Spongiform Encephalopathies: Scrapie, BSE and Related Human Disorders,ed. B Chesebro, pp. 139–52. Berlin /Heidelberg: Springer-Verlag
42. Parchi P, Castellani R, Capellari S, Ghetti B, Young K, et al. 1996. Molecular basis of phenotypicvariability in sporadic Creutzfeldt-Jakob disease. Ann. Neurol. 39:767–78
43. Collinge J, Sidle KCL, Meads J, Ironside J, Hill AF. 1996. Molecular analysis of prion strain variationand the aetiology of “new variant” CJD. Nature 383:685–90
44. Safar J, Wille H, Itri V, Groth D, Serban H, et al. 1998. Eight prion strains have PrP(Sc) moleculeswith different conformations. Nat. Med. 4:1157–65
45. Wille H, Michelitsch MD, Guenebaut V, Supattapone S, Serban A, et al. 2002. Structural studies ofthe scrapie prion protein by electron crystallography. Proc. Natl. Acad. Sci. USA 99:3563–68
46. Govaerts C, Wille H, Prusiner SB, Cohen FE. 2004. Evidence for assembly of prions with left-handedbeta-helices into trimers. Proc. Natl. Acad. Sci. USA 101:8342–47
47. Demarco ML, Daggett V. 2004. From conversion to aggregation: protofibril formation of the prionprotein. Proc. Natl. Acad. Sci. USA 101:2293–98
48. Demarco ML, Silveira J, Caughey B, Daggett V. 2006. Structural properties of prion protein protofibrilsand fibrils: an experimental assessment of atomic models. Biochemistry 45:15573–82
49. Lu X, Wintrode PL, Surewicz WK. 2007. Beta-sheet core of human prion protein amyloid fibrils asdetermined by hydrogen/deuterium exchange. Proc. Natl. Acad. Sci. USA 104:1510–15
50. Cobb NJ, Sonnichsen FD, McHaourab H, Surewicz WK. 2007. Molecular architecture of human prionprotein amyloid: a parallel, in-register beta-structure. Proc. Natl. Acad. Sci. USA 104:18946–51
51. Helmus JJ, Surewicz K, Nadaud PS, Surewicz WK, Jaroniec CP. 2008. Molecular conformation anddynamics of the Y145Stop variant of human prion protein in amyloid fibrils. Proc. Natl. Acad. Sci. USA105:6284–89
52. Lim KH, Nguyen TN, Damo SM, Mazur T, Ball HL, et al. 2006. Solid-state NMR structural studiesof the fibril form of a mutant mouse prion peptide PrP89-143(P101L). Solid State Nucl. Magn. Reson.29:183–90
53. Baxa U, Wickner RB, Steven AC, Anderson DE, Marekov LN, et al. 2007. Characterization of beta-sheet structure in Ure2p1-89 yeast prion fibrils by solid-state nuclear magnetic resonance. Biochemistry46:13149–62
54. Shewmaker F, Ross ED, Tycko R, Wickner RB. 2008. Amyloids of shuffled prion domains that formprions have a parallel in-register beta-sheet structure. Biochemistry 47:4000–7
55. Shewmaker F, Wickner RB, Tycko R. 2006. Amyloid of the prion domain of Sup35p has an in-registerparallel beta-sheet structure. Proc. Natl. Acad. Sci. USA 103:19754–59
56. Wickner RB, Dyda F, Tycko R. 2008. Amyloid of Rnq1p, the basis of the [PIN+] prion, has a parallelin-register beta-sheet structure. Proc. Natl. Acad. Sci. USA 105:2403–8
57. Krishnan R, Lindquist SL. 2005. Structural insights into a yeast prion illuminate nucleation and straindiversity. Nature 435:765–72
58. Luhrs T, Ritter C, Adrian M, Riek-Loher D, Bohrmann B, et al. 2005. 3D structure of Alzheimer’samyloid-beta(1–42) fibrils. Proc. Natl. Acad. Sci. USA 102:17342–47
59. Ritter C, Maddelein ML, Siemer AB, Luhrs T, Ernst M, et al. 2005. Correlation of structural elementsand infectivity of the HET-s prion. Nature 435:844–48
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 199
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
60. Wasmer C, Lange A, Van Melckebeke H, Siemer AB, Riek R, Meier BH. 2008. Amyloid fibrils of theHET-s(218–289) prion form a beta solenoid with a triangular hydrophobic core. Science 319:1523–26
61. Toyama BH, Kelly MJ, Gross JD, Weissman JS. 2007. The structural basis of yeast prion strain variants.Nature 449:233–37
62. Tanaka M, Collins SR, Toyama BH, Weissman JS. 2006. The physical basis of how prion conformationsdetermine strain phenotypes. Nature 442:585–89
63. Sun Y, Breydo L, Makarava N, Yang Q, Bocharova OV, Baskakov IV. 2007. Site-specific conformationalstudies of prion protein (PrP) amyloid fibrils revealed two cooperative folding domains within amyloidstructure. J. Biol. Chem. 282:9090–97
64. Anderson M, Bocharova OV, Makarava N, Breydo L, Salnikov VV, Baskakov IV. 2006. Polymorphismand ultrastructural organization of prion protein amyloid fibrils: an insight from high resolution atomicforce microscopy. J. Mol. Biol. 358:580–96
65. Nelson R, Sawaya MR, Balbirnie M, Madsen AØ, Riekel C, et al. 2005. Structure of the cross-betaspine of amyloid-like fibrils. Nature 435:773–78
66. Sawaya MR, Sambashivan S, Nelson R, Ivanova MI, Sievers SA, et al. 2007. Atomic structures of amyloidcross-beta spines reveal varied steric zippers. Nature 447:453–57
67. Chiesa R, Harris DA. 2001. Prion diseases: What is the neurotoxic molecule? Neurobiol. Dis. 8:743–6368. Caughey B, Raymond GJ. 1991. The scrapie-associated form of PrP is made from a cell surface precursor
that is both protease- and phospholipase-sensitive. J. Biol. Chem. 266:18217–2369. Caughey B, Raymond GJ, Ernst D, Race RE. 1991. N-terminal truncation of the scrapie-associated
form of PrP by lysosomal protease(s): implications regarding the site of conversion of PrP to theprotease-resistant state. J. Virol. 65:6597–603
70. Borchelt DR, Taraboulos A, Prusiner SB. 1992. Evidence for synthesis of scrapie prion protein in theendocytic pathway. J. Biol. Chem. 267:16188–99
71. Wong C, Xiong L-W, Horiuchi M, Raymond L, Wehrly K, et al. 2001. Sulfated glycans and elevatedtemperature stimulate PrPSc-dependent cell-free formation of protease-resistant prion protein. EMBOJ. 20:377–86
72. Horonchik L, Tzaban S, Ben-Zaken O, Yedidia Y, Rouvinski A, et al. 2005. Heparan sulfate is a cellularreceptor for purified infectious prions. J. Biol. Chem. 280:17062–67
73. Shaked GM, Meiner Z, Avraham I, Taraboulos A, Gabizon R. 2001. Reconstitution of prion infectiv-ity from solubilized protease-resistant PrP and nonprotein components of prion rods. J. Biol. Chem.276:14324–28
74. Leucht C, Simoneau S, Rey C, Vana K, Rieger R, et al. 2003. The 37 kDa/67 kDa laminin receptor isrequired for PrP(Sc) propagation in scrapie-infected neuronal cells. EMBO Rep. 4:290–95
75. Taraboulos A, Scott M, Semenov A, Avraham D, Laszlo L, Prusiner SB. 1995. Cholesterol depletionand modification of COOH-terminal targeting sequence of the prion protein inhibit formation of thescrapie isoform. J. Cell. Biol. 129:121–32
76. Pani A, Norfo C, Abete C, Mulas C, Putzolu M, et al. 2007. Antiprion activity of cholesterol esteri-fication modulators: a comparative study using ex vivo sheep fibroblasts and lymphocytes and mouseneuroblastoma cell lines. Antimicrob. Agents Chemother. 51:4141–47
77. Naslavsky N, Shmeeda H, Friedlander G, Yanai A, Futerman AH, et al. 1999. Sphingolipid depletionincreases formation of the scrapie prion protein in neuroblastoma cells infected with prions. J. Biol.Chem. 274:20763–71
78. Jeffrey M, McGovern G, Goodsir CM, Gonzalez L. 2009. Strain-associated variations in abnormal PrPtrafficking of sheep scrapie. Brain Pathol. 19:1–11
79. McKinley MP, Meyer RK, Kenaga L, Rahbar F, Cotter R, et al. 1991. Scrapie prion rod formation invitro requires both detergent extraction and limited proteolysis. J. Virol. 65:1340–51
80. Bellinger-Kawahara CG, Kempner E, Groth D, Gabizon R, Prusiner SB. 1988. Scrapie prion liposomesand rods exhibit target sizes of 55,000 Da. Virology 164:537–41
81. Deleault NR, Geoghegan JC, Nishina K, Kascsak R, Williamson RA, Supattapone S. 2005. Protease-resistant prion protein amplification reconstituted with partially purified substrates and syntheticpolyanions. J. Biol. Chem. 280:26873–79
200 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
82. Ben-Zaken O, Tzaban S, Tal Y, Horonchik L, Esko JD, et al. 2003. Cellular heparan sulfate participatesin the metabolism of prions. J. Biol. Chem. 278:40041–49
83. Mabbott NA, MacPherson GG. 2006. Prions and their lethal journey to the brain. Nat. Rev. Microbiol.4:201–11
84. Aguzzi A, Sigurdson C, Heikenwaelder M. 2008. Molecular mechanisms of prion pathogenesis. Annu.Rev. Pathol. 3:11–40
85. Prinz M, Heikenwalder M, Junt T, Schwarz P, Glatzel M, et al. 2003. Positioning of follicular dendriticcells within the spleen controls prion neuroinvasion. Nature 425:957–62
86. Siso S, Jeffrey M, Gonzalez L. 2009. Neuroinvasion in sheep TSEs: the role of the haematogenousroute. Neuropathol. Appl. Neurobiol. 35:232–46
87. Houston F, McCutcheon S, Goldmann W, Chong A, Foster J, et al. 2008. Prion diseases are efficientlytransmitted by blood transfusion in sheep. Blood 112:4739–45
88. Bartz JC, Kincaid AE, Bessen RA. 2003. Rapid prion neuroinvasion following tongue infection. J. Virol.77:583–91
89. Magalhaes AC, Baron GS, Lee KS, Steele-Mortimer O, Dorward D, et al. 2005. Uptake and neuritictransport of scrapie prion protein coincident with infection of neuronal cells. J. Neurosci. 25:5207–16
90. Greil CS, Vorberg IM, Ward AE, Meade-White KD, Harris DA, Priola SA. 2008. Acute cellular uptakeof abnormal prion protein is cell type and scrapie-strain independent. Virology 379:284–93
91. Hijazi N, Kariv-Inbal Z, Gasset M, Gabizon R. 2005. PrPSc incorporation to cells requires endogenousglycosaminoglycan expression. J. Biol. Chem. 280:17057–61
92. Paquet S, Daude N, Courageot MP, Chapuis J, Laude H, Vilette D. 2007. PrPc does not mediateinternalization of PrPSc but is required at an early stage for de novo prion infection of Rov cells. J.Virol. 81:10786–91
93. Gauczynski S, Nikles D, El-Gogo S, Papy-Garcia D, Rey C, et al. 2006. The 37-kDa/67-kDa lamininreceptor acts as a receptor for infectious prions and is inhibited by polysulfated glycanes. J. Infect. Dis.194:702–9
94. Jeffrey M, Martin S, Gonzalez L. 2003. Cell-associated variants of disease-specific prion protein im-munolabelling are found in different sources of sheep transmissible spongiform encephalopathy. J. Gen.Virol. 84:1033–45
95. Baron GS, Magalhaes AC, Prado MA, Caughey B. 2006. Mouse-adapted scrapie infection of SN56cells: greater efficiency with microsome-associated versus purified PrP-res. J. Virol. 80:2106–17
96. Baron GS, Caughey B. 2003. Effect of glycosylphosphatidylinositol anchor-dependent and - indepen-dent prion protein association with model raft membranes on conversion to the protease-resistantisoform. J. Biol. Chem. 278:14883–92
97. Baron GS, Wehrly K, Dorward DW, Chesebro B, Caughey B. 2002. Conversion of raft associated prionprotein to the protease-resistant state requires insertion of PrP-res (PrPSc) into contiguous membranes.EMBO J. 21:1031–40
98. Flechsig E, Hegyi I, Enari M, Schwarz P, Collinge J, Weissmann C. 2001. Transmission of scrapie bysteel-surface-bound prions. Mol. Med. 7:679–84
98a. Kanu N, Imokawa Y, Drechsel DN, Williamson RA, Birkett CR, et al. 2002. Transfer of scrapie prioninfectivity by cell contact in culture. Curr. Biol. 12:523–30
99. Paquet S, Langevin C, Chapuis J, Jackson GS, Laude H, Vilette D. 2007. Efficient dissemination ofprions through preferential transmission to nearby cells. J. Gen. Virol. 88:706–13
100. Fevrier B, Vilette D, Archer F, Loew D, Faigle W, et al. 2004. Cells release prions in association withexosomes. Proc. Natl. Acad. Sci. USA 101:9683–88
101. Vella LJ, Sharples RA, Lawson VA, Masters CL, Cappai R, Hill AF. 2007. Packaging of prions intoexosomes is associated with a novel pathway of PrP processing. J. Pathol. 211:582–90
102. McGovern G, Jeffrey M. 2007. Scrapie-specific pathology of sheep lymphoid tissues. PLoS ONE 2:e1304103. Rustom A, Saffrich R, Markovic I, Walther P, Gerdes HH. 2004. Nanotubular highways for intercellular
organelle transport. Science 303:1007–10104. Caughey B, Baron GS. 2006. Prions and their partners in crime. Nature 443:803–10
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 201
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
105. Gerdes HH, Carvalho RN. 2008. Intercellular transfer mediated by tunneling nanotubes. Curr. Opin.Cell Biol. 20:470–75
105a. Sherer NM, Lehmann MJ, Jimenez-Soto LF, Horensavitz C, Pypaert M, Mothes W. 2007. Retrovirusescan establish filopodial bridges for efficient cell-to-cell transmission. Nat. Cell Biol. 9:310–15
106. Leblanc P, Alais S, Porto-Carreiro I, Lehmann S, Grassi J, et al. 2006. Retrovirus infection stronglyenhances scrapie infectivity release in cell culture. EMBO J. 25:2674–85
107. Caughey B, Lansbury PT. 2003. Protofibrils, pores, fibrils, and neurodegeneration: separating theresponsible protein aggregates from the innocent bystanders. Annu. Rev. Neurosci. 26:267–98
108. Walsh DM, Selkoe DJ. 2007. Aβ oligomers—a decade of discovery. J. Neurochem. 101:1172–84109. Jeffrey M, Goodsir CM, Bruce ME, McBride PA, Farquhar C. 1994. Morphogenesis of amyloid plaques
in 87V murine scrapie. Neuropathol. Appl. Neurobiol. 20:535–42110. Bolmont T, Haiss F, Eicke D, Radde R, Mathis CA, et al. 2008. Dynamics of the microglial/amyloid
interaction indicate a role in plaque maintenance. J. Neurosci. 28:4283–92111. Belichenko PV, Brown D, Jeffrey M, Fraser JR. 2000. Dendritic and synaptic alterations of hippocampal
pyramidal neurones in scrapie-infected mice. Neuropathol. Appl. Neurobiol. 26:143–49112. Jeffrey M, Goodsir CM, Race RE, Chesebro B. 2004. Scrapie-specific neuronal lesions are independent
of neuronal PrP expression. Ann. Neurol. 55:781–92113. Liberski P. 2008. The tubulovesicular structures—the ultrastructural hallmark for all prion diseases.
Acta Neurobiol. Exp. 68:113–21114. Eberl H, Tittmann P, Glockshuber R. 2004. Characterization of recombinant, membrane-attached
full-length prion protein. J. Biol. Chem. 279:25058–65115. Hicks MR, Gill AC, Bath IK, Rullay AK, Sylvester ID, et al. 2006. Synthesis and structural characteri-
zation of a mimetic membrane-anchored prion protein. FEBS J. 273:1285–99116. Elfrink K, Ollesch J, Stohr J, Willbold D, Riesner D, Gerwert K. 2008. Structural changes of membrane-
anchored native PrP(C). Proc. Natl. Acad. Sci. USA 105:10815–19117. Morillas M, Swietnicki W, Gambetti P, Surewicz WK. 1999. Membrane environment alters the con-
formational structure of the recombinant human prion protein. J. Biol. Chem. 274:36859–65118. Bahadi R, Farrelly PV, Kenna BL, Kourie JI, Tagliavini F, et al. 2003. Channels formed with a mutant
prion protein PrP(82–146) homologous to a 7-kDa fragment in diseased brain of GSS patients. Am. J.Physiol. Cell Physiol. 285:C862–72
119. Wang X, Wang F, Arterburn L, Wollmann R, Ma J. 2006. The interaction between cytoplasmic prionprotein and the hydrophobic lipid core of membrane correlates with neurotoxicity. J. Biol. Chem.281:13559–65
120. Wang F, Yang F, Hu Y, Wang X, Wang X, et al. 2007. Lipid interaction converts prion protein to aPrPSc-like proteinase K-resistant conformation under physiological conditions. Biochemistry 46:7045–53
121. Kazlauskaite J, Sanghera N, Sylvester I, Venien-Bryan C, Pinheiro TJ. 2003. Structural changes of theprion protein in lipid membranes leading to aggregation and fibrillization. Biochemistry 42:3295–304
122. Sanghera N, Pinheiro TJ. 2002. Binding of prion protein to lipid membranes and implications for prionconversion. J. Mol. Biol. 315:1241–56
123. Simoneau S, Rezaei H, Sales N, Kaiser-Schulz G, Lefebvre-Roque M, et al. 2007. In vitro and in vivoneurotoxicity of prion protein oligomers. PLoS Pathog. 3:e125
124. Stahl N, Baldwin MA, Burlingame AL, Prusiner SB. 1990. Identification of glycoinositol phospholipidlinked and truncated forms of the scrapie prion protein. Biochemistry 29:8879–84
125. Caughey B. 2001. Interactions between prion protein isoforms: The kiss of death? Trends Biochem. Sci.26:235–42
126. Wegmann S, Miesbauer M, Winklhofer KF, Tatzelt J, Muller DJ. 2008. Observing fibrillar assemblieson scrapie-infected cells. Pflugers Arch. 456:83–93
127. Bueler H, Fischer M, Lang Y, Bluethmann H, Lipp H-P, et al. 1992. Normal development and behaviorof mice lacking the neuronal cell-surface PrP protein. Nature 356:577–82
128. Mallucci GR, Ratte S, Asante EA, Linehan J, Gowland I, et al. 2002. Post-natal knockout of prion proteinalters hippocampal CA1 properties, but does not result in neurodegeneration. EMBO J. 21:202–10
202 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
129. Li A, Piccardo P, Barmada SJ, Ghetti B, Harris DA. 2007. Prion protein with an octapeptide insertionhas impaired neuroprotective activity in transgenic mice. EMBO J. 26:2777–85
130. Brandner S, Isenmann S, Raeber A, Fischer M, Sailer A, et al. 1996. Normal host prion protein necessaryfor scrapie-induced neurotoxicity. Nature 379:339–43
131. Mallucci G, Dickinson A, Linehan J, Klohn PC, Brandner S, Collinge J. 2003. Depleting neuronal PrPin prion infection prevents disease and reverses spongiosis. Science 302:871–74
132. Race R, Oldstone M, Chesebro B. 2000. Entry versus blockade of brain infection following oral orintraperitoneal scrapie administration: role of prion protein expression in peripheral nerves and spleen.J. Virol. 74:828–33
133. Raeber AJ, Race RE, Brandner S, Priola SA, Sailer A, et al. 1997. Astrocyte-specific expression of hamsterprion protein (PrP) renders PrP knockout mice susceptible to hamster scrapie. EMBO J. 16:6057–65
134. Marella M, Gaggioli C, Batoz M, Deckert M, Tartare-Deckert S, Chabry J. 2005. Pathological prionprotein exposure switches on neuronal mitogen-activated protein kinase pathway resulting in microgliarecruitment. J. Biol. Chem. 280:1529–34
135. Giese A, Brown DR, Groschup MH, Feldmann C, Haist I, Kretzschmar HA. 1998. Role of microgliain neuronal cell death in prion disease. Brain Pathol. 8:449–57
136. Harris DA, True HL. 2006. New insights into prion structure and toxicity. Neuron 50:353–57137. Linden R, Martins VR, Prado MA, Cammarota M, Izquierdo I, Brentani RR. 2008. Physiology of the
prion protein. Physiol. Rev. 88:673–728138. Solforosi L, Criado JR, McGavern DB, Wirz S, Sanchez-Alavez M, et al. 2004. Cross-linking cellular
prion protein triggers neuronal apoptosis in vivo. Science 303:1514–16139. Watts JC, Westaway D. 2007. The prion protein family: diversity, rivalry, and dysfunction. Biochim.
Biophys. Acta 1772:654–72140. Kristiansen M, Deriziotis P, Dimcheff DE, Jackson GS, Ovaa H, et al. 2007. Disease-associated prion
protein oligomers inhibit the 26S proteasome. Mol. Cell 26:175–88141. Fowler DM, Koulov AV, Balch WE, Kelly JW. 2007. Functional amyloid—from bacteria to humans.
Trends Biochem. Sci. 32:217–24142. Chiti F, Dobson CM. 2006. Protein misfolding, functional amyloid, and human disease. Annu. Rev.
Biochem. 75:333–66143. Wickner RB, Edskes HK, Shewmaker F, Nakayashiki T. 2007. Prions of fungi: inherited structures and
biological roles. Nat. Rev. Microbiol. 5:611–18144. Derkatch IL, Bradley ME, Hong JY, Liebman SW. 2001. Prions affect the appearance of other prions:
the story of [PIN(+)]. Cell 106:171–82145. Coustou-Linares V, Maddelein ML, Begueret J, Saupe SJ. 2001. In vivo aggregation of the HET-s
prion protein of the fungus Podospora anserina. Mol. Microbiol. 42:1325–35145a. Roberts BT, Wickner RB. 2003. A class of prions that propagate via covalent auto-activation. Genes
Dev. 17:2083–87146. Kicka S, Bonnet C, Sobering AK, Ganesan LP, Silar P. 2006. A mitotically inheritable unit containing
a MAP kinase module. Proc. Natl. Acad. Sci. USA 103:13445–50147. Kisilevsky R. 2005. Preparation and propagation of amyloid-enhancing factor. Methods Mol. Biol.
299:237–41148. Lundmark K, Westermark GT, Nystrom S, Murphy CL, Solomon A, Westermark P. 2002. Transmis-
sibility of systemic amyloidosis by a prion-like mechanism. Proc. Natl. Acad. Sci. USA 99:6979–84149. Papendick RE, Munson L, O’Brien TD, Johnson KH. 1997. Systemic AA amyloidosis in captive chee-
tahs (Acinonyx jubatus). Vet. Pathol. 34:549–56150. Munson L, Terio KA, Worley M, Jago M, Bagot-Smith A, Marker L. 2005. Extrinsic factors significantly
affect patterns of disease in free-ranging and captive cheetah (Acinonyx jubatus) populations. J. Wildl.Dis. 41:542–48
151. Zhang B, Une Y, Fu X, Yan J, Ge F, et al. 2008. Fecal transmission of AA amyloidosis in the cheetahcontributes to high incidence of disease. Proc. Natl. Acad. Sci. USA 105:7263–68
152. Xing Y, Nakamura A, Chiba T, Kogishi K, Matsushita T, et al. 2001. Transmission of mouse senileamyloidosis. Lab. Investig. 81:493–99
www.annualreviews.org • Prions, Amyloid, Anchors, and Membranes 203
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-08 ARI 5 May 2009 13:45
153. Meyer-Luehmann M, Coomaraswamy J, Bolmont T, Kaeser S, Schaefer C, et al. 2006. Exogenousinduction of cerebral beta-amyloidogenesis is governed by agent and host. Science 313:1781–84
154. Si K, Lindquist S, Kandel ER. 2003. A neuronal isoform of the aplysia CPEB has prion-like properties.Cell 115:879–91
155. Walker LC, LeVine H III, Mattson MP, Jucker M. 2006. Inducible proteopathies. Trends Neurosci.29:438–43
156. Lee KS, Caughey B. 2007. A simplified recipe for prions. Proc. Natl. Acad. Sci. USA 104:9551–52157. Ma J, Wollmann R, Lindquist S. 2002. Neurotoxicity and neurodegeneration when PrP accumulates
in the cytosol. Science 298:1781–85158. Kristiansen M, Messenger MJ, Klohn PC, Brandner S, Wadsworth JD, et al. 2005. Disease-related
prion protein forms aggresomes in neuronal cells leading to caspase activation and apoptosis. J. Biol.Chem. 280:38851–61
204 Caughey et al.
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

AR378-FM ARI 7 May 2009 15:43
Annual Review ofBiochemistry
Volume 78, 2009
ContentsPreface � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �v
Prefatory Articles
FrontispieceE. Peter Geiduschek � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �xii
Without a License, or Accidents Waiting to HappenE. Peter Geiduschek � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
FrontispieceJames C. Wang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �30
A Journey in the World of DNA Rings and BeyondJames C. Wang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �31
Biochemistry and Disease Theme
The Biochemistry of Disease: Desperately Seeking SyzygyJohn W. Kozarich � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �55
Biosynthesis of Phosphonic and Phosphinic Acid Natural ProductsWilliam W. Metcalf and Wilfred A. van der Donk � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �65
New Antivirals and Drug ResistancePeter M. Colman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �95
Multidrug Resistance in BacteriaHiroshi Nikaido � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 119
Conformational Pathology of the Serpins: Themes, Variations,and Therapeutic StrategiesBibek Gooptu and David A. Lomas � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 147
Getting a Grip on Prions: Oligomers, Amyloids, and PathologicalMembrane InteractionsByron Caughey, Gerald S. Baron, Bruce Chesebro, and Martin Jeffrey � � � � � � � � � � � � � � � � � 177
Ubiquitin-Mediated Protein Regulation
RING Domain E3 Ubiquitin LigasesRaymond J. Deshaies and Claudio A.P. Joazeiro � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 399
Regulation and Cellular Roles of Ubiquitin-SpecificDeubiquitinating EnzymesFrancisca E. Reyes-Turcu, Karen H. Ventii, and Keith D. Wilkinson � � � � � � � � � � � � � � � � � � � � 363
vi
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

AR378-FM ARI 7 May 2009 15:43
Recognition and Processing of Ubiquitin-Protein Conjugatesby the ProteasomeDaniel Finley � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 477
Degradation of Activated Protein Kinases by UbiquitinationZhimin Lu and Tony Hunter � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 435
The Role of Ubiquitin in NF-κB Regulatory PathwaysBrian Skaug, Xiaomo Jiang, and Zhijian J. Chen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 769
Biological and Chemical Approaches to Diseases of ProteostasisDeficiencyEvan T. Powers, Richard I. Morimoto, Andrew Dillin, Jeffery W. Kelly,and William E. Balch � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 959
Gene Expression
RNA Polymerase Active Center: The Molecular Engineof TranscriptionEvgeny Nudler � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 335
Genome-Wide Views of Chromatin StructureOliver J. Rando and Howard Y. Chang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 245
The Biology of Chromatin Remodeling ComplexesCedric R. Clapier and Bradley R. Cairns � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 273
The Structural and Functional Diversity of Metabolite-BindingRiboswitchesAdam Roth and Ronald R. Breaker � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 305
Lipid and Membrane Biogenesis
Genetic and Biochemical Analysis of Non-Vesicular Lipid TrafficDennis R. Voelker � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 827
Cholesterol 24-Hydroxylase: An Enzyme of Cholesterol Turnoverin the BrainDavid W. Russell, Rebekkah W. Halford, Denise M.O. Ramirez, Rahul Shah,and Tiina Kotti � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �1017
Lipid-Dependent Membrane Protein TopogenesisWilliam Dowhan and Mikhail Bogdanov � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 515
Single-Molecule Studies of the Neuronal SNARE Fusion MachineryAxel T. Brunger, Keith Weninger, Mark Bowen, and Steven Chu � � � � � � � � � � � � � � � � � � � � � � � 903
Mechanisms of EndocytosisGary J. Doherty and Harvey T. McMahon � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 857
Contents vii
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.

AR378-FM ARI 7 May 2009 15:43
Recent Advances in Biochemistry
Motors, Switches, and Contacts in the ReplisomeSamir M. Hamdan and Charles C. Richardson � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 205
Large-Scale Structural Biology of the Human ProteomeAled Edwards � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 541
Collagen Structure and StabilityMatthew D. Shoulders and Ronald T. Raines � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 929
The Structural and Biochemical Foundations of Thiamin BiosynthesisChristopher T. Jurgenson, Tadhg P. Begley, and Steven E. Ealick � � � � � � � � � � � � � � � � � � � � � � � � 569
Proton-Coupled Electron Transfer in Biology: Results fromSynergistic Studies in Natural and Model SystemsSteven Y. Reece and Daniel G. Nocera � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 673
Mechanism of Mo-Dependent NitrogenaseLance C. Seefeldt, Brian M. Hoffman, and Dennis R. Dean � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 701
Inorganic Polyphosphate: Essential for Growth and SurvivalNarayana N. Rao, Marıa R. Gomez-Garcıa, and Arthur Kornberg � � � � � � � � � � � � � � � � � � � � � 605
Essentials for ATP Synthesis by F1F0 ATP SynthasesChristoph von Ballmoos, Alexander Wiedenmann, and Peter Dimroth � � � � � � � � � � � � � � � � � � 649
The Chemical Biology of Protein PhosphorylationMary Katherine Tarrant and Philip A. Cole � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 797
Sphingosine 1-Phosphate Receptor SignalingHugh Rosen, Pedro J. Gonzalez-Cabrera, M. Germana Sanna, and Steven Brown � � � � 743
The Advent of Near-Atomic Resolution in Single-Particle ElectronMicroscopyYifan Cheng and Thomas Walz � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 723
Super-Resolution Fluorescence MicroscopyBo Huang, Mark Bates, and Xiaowei Zhuang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 993
Indexes
Cumulative Index of Contributing Authors, Volumes 74–78 � � � � � � � � � � � � � � � � � � � � � � � � � �1041
Cumulative Index of Chapter Titles, Volumes 74–78 � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �1045
Errata
An online log of corrections to Annual Review of Biochemistry articles may be found athttp://biochem.annualreviews.org/errata.shtml
viii Contents
Ann
u. R
ev. B
ioch
em. 2
009.
78:1
77-2
04. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
02/
10/1
2. F
or p
erso
nal u
se o
nly.


















