BROWSER COMPATIBILITY, DOMAINS & HOSTING ART340. BROWSER COMPATIBILITY.

1/18/2012
1
POPULATION GENOMICS OF
158 GENOMES OF DROSOPHILA MELANOGASTER
Antonio Barbadilla Bioinformatics of Genetic Diversity
Genomics, Bioinformatics and Evolution research group Departament de Genètica i Microbiologia
Universitat Autònoma de Barcelona
The problematic of population genetics is the description and explanation of genetic variation within and between populations
Theodosious Dobzhansky
Population genetics
The golden age of the study of genetic variation
Internacional Project (US National Human Genome Research Institute): Sequencing of a genetic reference panel of 192 wild lines of D. melanogaster
Drosophila Genetic Reference Panel
(DGRP): A Community Resource for the Study of Genotypic and Phenotypic Variation in model organism
Trudy Mackay
Stephen Richards
Richard Gibbs

1/18/2012
2
The Drosophila Genetic Reference Panel – DGRP
Sequencing of a D. melanogaster genetic reference panel of 192 wild-type inbred lines from a single natural population (Raleigh, North Carolina, USA)
The Drosophila Genetic Reference Panel – DGRP
192 lines which extensive information on complex trait phenotypes has been collected
DGRP – Sequencing strategies
Long reads (~500 bp)
Primary error: homopolymer
length determination
Short reads (~45 bp), Paired-
end. Primary error: substit. at
the ends of the reads
De-novo assembly
Identify large polymorphic
features (>20 bp)
Identify small polymorphic
features (<10 – 20 bp)
Mapping reads to the
Drosophila reference genome
(Mosaik)

1/18/2012
3
DGRP – Main Goals
1. Create a community resource for the association mapping of quantitative trait loci (QTLs)
2. Create a community resource containing the common sequence polymorphisms of Drosophila melanogaster (SNPs and structural variation)
3. Create a “test bench” for statistical methods used in QTL association mapping studies for traits affecting human diseases
Last Name First Name Email Address Proposed Analysis
Andolfatto Peter [email protected] Recombination rates
Anholt Robert [email protected] Olfactory behavior (mulitple odorants); molecular population genetics of odorant binding proteins
Arnosti David [email protected] Mathematical modeling of enhancers, testing quantitative predictions
Awadalla Philip [email protected]
Genome wide analysis of historical rates of recombination and gene conversion
Barbadilla Antonio [email protected] Create and maintain a Web server containing the high resolution sequence polymorphism map of each strain; participate in genome wide molecular population genetics analyses
Bergmann Casey [email protected]
TE insertion sites; Wolbachia infection status
Caellarts Patrick [email protected]
Quantitative morphological analyses of nervous system
Carbone Ignacio [email protected] Whole genome phylogenetic analyses
Clark Andrew [email protected] Immunity, metabolism
DeLuca Maria [email protected] Energy storage, metabolic rate, mitochondrial respiration
Dworkin Ian [email protected] Wing morphology
Eyre-Walker Adam [email protected]
Distribution of fitness effects of deleterious mutations
Fanara Juan Jose [email protected]
Larval olfactory behavior, development time, adult body size
Gleason Jennifer [email protected] Courtship song
Hasson Esteban [email protected]
Development time, body size and related traits, male genital morphology, oviposition preference
Houle David [email protected] Wing morphology (108 parameters)
Hughes Kimberly [email protected] Sperm precedence and male fertility
Jiggins Frank [email protected] Susceptibility/resistance to virus infection
Kreitman Martin [email protected] Molecular evolutionary genetics of trancription factor binding sites
Lazarro Brian [email protected] Resistance to bacterial infection (mulitple bacteria), endogenous microbiomes, metabolism, reproduction
Leips Jeff [email protected] Immune responses at different ages
Levine Joel [email protected] Circadian rhythms, cuticular hydrocarbons, social behaviors
Mackay Trudy [email protected]
Abdominal and sternopleural bristle number, pigmentation, locomotion (startle response, open field activity), sleep, aggression, EtOH sensitivity and tolerance, copulation latency, phototaxis, oxidative stress resistance, life span, starvation stress resistance, chill coma recovery, competitive fitness, geotaxis, modifiers of mutations, mutation accumulation, whole genome transcript profiling, eQTLs and systems genetics of complex traits analyses; most traits in multiple environments
Mackey Aaron [email protected]
Genome assemblies, SNP/indel identification
Mahaffey James [email protected] Appendage morphometrics
Mery Frederic [email protected]
Learning and memory
Mezey Jason [email protected]
Wing morphology
Otto Sarah [email protected] Signatures of disequilibrium and other signs of selective interference among loci (Hill-Robertson effects)
Petrov Dmitri [email protected]
TE insertion sites
Promislow Daniel [email protected]
Somatic mutation rates
Rollmann Stephanie [email protected] Molecular population genetics of ororant and taste receptor gene families; gustatory perception of sugars and aversive behavior to noxious food stimuli
Rozas Julio [email protected] Molecular evolutionary analyses of odorant binding protein cis-regulatory sequences using phylogenetic footprinting/shadowing methodologies
Sokolowski Marla [email protected]
larval and adult food related behaviors including food related locomotion, food intake, lipid, carbohydrate and protein levels, glucose and amino acid uptake, sucrose responsiveness and social feeding; all in environments varying in food deprivation, food quality and composition
Stephan Wolfgang [email protected]
Molecular population genomics of selective sweeps, signatures of adaptive evolution
Stone Eric [email protected] eQTL and systems genetics of complex traits analyses
Uyenoyama Marcy [email protected] Analysis of site frequency spectrum (SNPs and CNVs); analysis of whole genome linkage disequilibria; local scale variation in recombination rates
Wayne Marta [email protected] Ovariole number, sigma virus resistance and transmission, flight under field conditions
Wolfner Mariana [email protected]
variation in sperm competition (both males RNAi'd for individual Acps in mated to DGRP females, and DGRP males in mated to females RNAi'd for specific female reproductive tract proteins
Wu Louisa [email protected] Susceptibility/resistance to virus infection (Drosophila X virus, Drosophila C virus); differences in ability to phagocytose bacteria
Yarali Ayse [email protected]
Associative learning (reinforcement)
DGRP – The Consortium T. Mackay, S. Richards and R. Gibbs
DGRP – Our Task
1. Create and maintain a Genome
Browser containing the high-
resolution sequence
polymorphism map of D.
melanogaster
2. Genome-wide molecular
population genetic analyses
Population genomics
Description and explanation of patterns of nucleotide
variation at a large scale
Population genetics studies have been based on fragmentary and
non-random samples of the genome, providing a partial view, often
biased, of the population genetic processes

1/18/2012
4
Questions that can be solved with
a genome-wide perspective
• X vs autosomes. X-fast evolution?
Yes (Baines et al 2008)
No (Connalon 2007, Thornton et al. )
Regional Effect Centromere – Middle - Telomere on nucleotide variation
Role recombination, mutation, gene density on nucleotide variation and adaptive evolution
% genomes underlying natural selection, positive, balanced and negative according functional or structural region of genomes
X
Population Genomics Analyses
Raw data: 158 genomes D. melanogaster DGRP project (Freeze 1).

1/18/2012
5
Raw data: Outgruoup species
Data visualization: The Population Drosophila Browser (PopDrowser)
Data quality: Julien Ayroles

1/18/2012
6
0
200000
400000
600000
800000
1000000
1200000
2L 2R 3L 3R X
SNPs
#SNPs (all)
#SNPs (no-singletons)
#SNPs (all) #SNPs (no-singletons)
% no-singletons % singletons
2L 1073340 787498 73,37% 26,63%
2R 891243 645133 72,39% 27,61%
3L 1095397 772713 70,54% 29,46%
3R 1050901 741908 70,60% 29,40%
X 699821 454374 64,93% 35,07%
TOTAL 4810702 3401626 70,71% 29,29%
Eric Stone’s genotyper (JGIL)
Data analysis approaches
• Differently-sized non-overlapping sliding windows along chromosome arms (ranging from 50pb to 100kb)
•Specific and general patterns that follows genetic diversity (polymorphism and divergence) along the chromosome arms •Correlating them to other variables such as recombination rate, gene density, linkage disequilibrium, or structural regions
•Coding genes centred approach: Site (functional) classes of coding gene: coding (synonymous, non-synonymous) and non-coding (5’ and 3’UTR, intron, 5’ and 3’ intergenic).
•Nucleotide variation •Evidence of natural selection acting on each site class.
Polymorphism and divergence

1/18/2012
7
Patterns polymorphism and divergence along chromosome arms
PopDrowser -> http://popdrowser.uab.cat;
2L 2R 3L 3R X
Tel Cen Cent Tel Cen Cen Cen Tel Tel Cen
Patterns polymorphism and divergence along chromosome arms
• centromeric vs. non-centromeric regions within autosome arms • autosomes vs. X chromosome PopDrowser -> http://popdrowser.uab.cat;
Polymorphism / divergence and recombination
• centromeric vs. non-centromeric regions within autosome arms • autosomes vs. X chromosome PopDrowser -> http://popdrowser.uab.cat;
and the rate of recombination Site class Spearman’s ρ Prob
rec < 2 cM/Mb 0.471 p < 2.2 e-16
rec ≥ 2 cM/Mb -0.0044 p = 0.987
Gene centred approach: Polymorphism and divergence by site classes

1/18/2012
8
•Consistent pattern among and within chromosomes for both polymorphism and divergence as previously observed on smaller data set.
Polymorphism: π_Syn 4-fold> π _Intron > π _Intergenic > π _5’-3’UTR > π _Non-syn 0-fold Divergence: k_Syn 4-fold> k _Intron > k _Intergenic > k _5’-3’UTR > k _Non-syn 0-fold
Outgroup: D. yakuba
Polymorphism (π)
and
Divergence (k)
by site class
among regions
within
chromosome arms
and
X vs. Autosomes
Sum of squares
df F-value p-value
Autosomes
Recombination rate
0.00637 1 199.03 < 2.2e-16 ***
Region 0.00224 1 69.88 < 2.2e-16 ***
Divergence 0.0034 1 106.21 < 2.2e-16 ***
Gene density 0.0003 1 8.43 0.0037 **
Residuals 0.0591 1848
X
Recombination rate
0.0007 1 37.32 5.844e-10***
Divergence 0.0001 1 4.80 0.0290 * Region 0.0000 1 0.87 0.3522 Gene density 0.0000 1 0.61 0.4363 Residuals 0.0073 415
Anova of the regression model to test the effect of different genomic
variables on nucleotide variation. •Dependent variables: recombination rate, genomic region (centromere or
non-centromere), 4-fold divergence to D. yakuba and gene density
•Independent variable is π 4fold

1/18/2012
9
Allele
frequency
Time
1
0
Dinamics of substitutions of neutral mutations
Motoo Kimura
i
neut =
ki kneut
Ratio divergence = ω =
i neut
Ratio polymorphism =
Expected ratio according
neutral theory
ki kneut
= 4Nμ
k = μ
McDonald-Kreitman test (MKT)
μ i μ neut
=
4N μi 4N μneut
=
Allele
frequency
Time
1
0
Non-neutral substitutions
i neut
=
Expected ratio according
neutral theory
ki kneut
i neut
< ki kneut
Adaptive fixation (α ,
fraction of sites fixed
by adaptive selection)
i neut
> ki kneut
Weakly negative
selection
= 4Nμ
k = μ
McDonald-Kreitman test (MKT)
1/18/2012
10
i neut
=
Expected ratio according
neutral theory
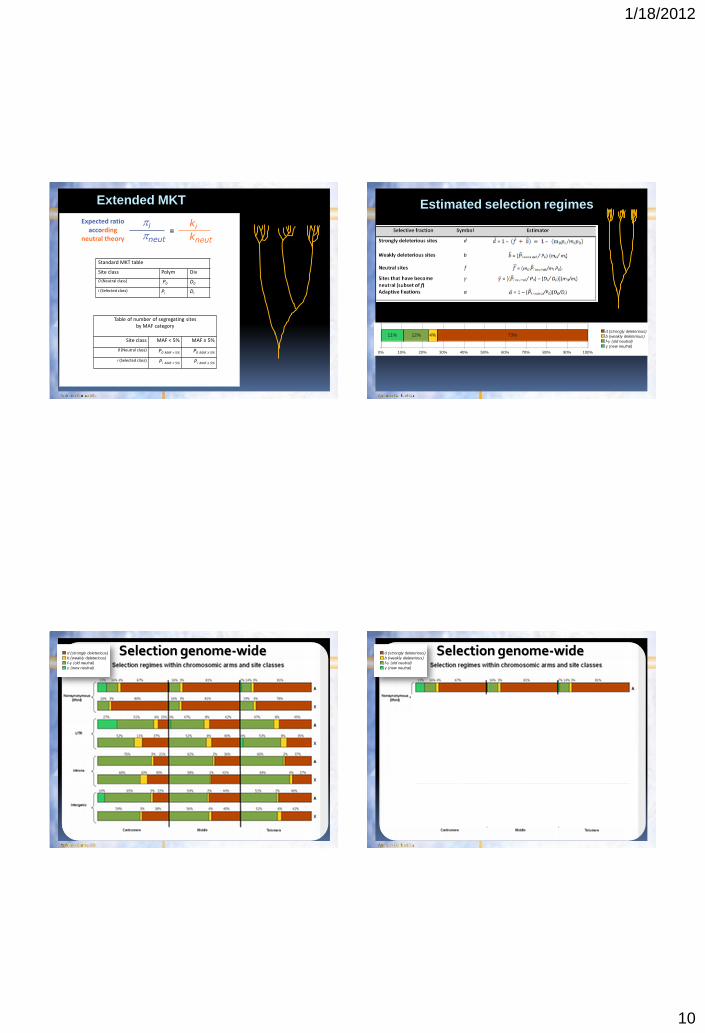
Extended MKT
ki kneut
Standard MKT table
Site class Polym Div
0 (Neutral class) P0 D0
i (Selected class) Pi Di
Table of number of segregating sites by MAF category
Site class MAF < 5% MAF ≥ 5%
0 (Neutral class) P0 MAF < 5% P0 MAF ≥ 5%
i (Selected class) Pi MAF < 5% Pi MAF ≥ 5%
Estimated selection regimes
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
11% 12% 4% 73%d (strongly deleterious)
b (weakly deleterious)
f-γ (old neutral)
γ (new neutral)
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
11% 12% 4% 73%d (strongly deleterious)
b (weakly deleterious)
f-γ (old neutral)
γ (new neutral)
Selection genome-wide
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
11% 12% 4% 73%d (strongly deleterious)
b (weakly deleterious)
f-γ (old neutral)
γ (new neutral)
Selection genome-wide

1/18/2012
11
●●●●●
●
●●●
●
●
●●
●
●●
●
●
●
●●
●●
●
●
●
●●●
●●●
●
●●
●●
●●
●
●●
●●●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●●
●
●
●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●●
●
●
●
●●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●●
●
●●●●
●
●●●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●●●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●●●●●
●
●
●●
●●
●●●
●
●
●●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●●
●
●●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●●●●
●●
●
●
●
●●●
●●
●
●
●
●
●
●
●●
●●
●●●
●●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●●
●
●
●
●●
●●
●
●●
●●
●
●●
●
●●●
●●
●●
●
●●●
●
●●
●●●
●
●
●
●●●
●●●
●●●●●●
●●
●
●
●
●
0.0
00
0.0
04
0.0
08
0.0
12
p −
Nu
cle
otid
e D
ive
rsity
a)
●●●●●●●●●●●●●●●●●
●
●●●●
●
●
●●
●
●
●
●●●●
●
●
●
●●●●●
●
●
●
●
●
●●●
●●●●
●●
●
●●
●●●●●
●
●
●●●●●●
●
●●
●
●●
●●●
●
●●●
●
●
●
●●
●
●
●●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●●
●
●
●●●●●
●
●
●
●
●
●
●●
●
●
●
●●●
●
●
●●●
●
●
●●●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●●
●
●●●
●●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●●
●
●
●●●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●●●●●●
●
●
●●
●
●
●●●●●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●●●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●●●
●●
●
●●●
●
●●
●
●●●
●
●
●
●
●
●●
●
●
●●
●
●
●●
●
●
●●
●●
●
●
●●●
●●●
●●
●
●
●
●
●●●
●●
●
●
●●●
●
●
●●
●
●●●●
●
●
●
●●●
●
●●
●●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●●●●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●●●
●
●●
●
●
●●●●
●
●
●●●●
●●
●
●
●●
●●
●
●●
●
●
●
●●●
●
●
●
●
●
●
●●
●●●
●
●
●
●●
●●
●
●
●
●
●●●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●●
●
●
●
●
●●
●
●
●●
●
●●●
●
●
●●
●●●
●●●
●●
●
●
●
●
●●●
●●●
●
●●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●●
●
●●
●●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●●
●
●●
●
●●●
●
●
●
●
●●
●●●
●
●●
●●●
●●●●
●
●●
●●●
●
●●●●●●●
●
●●
●●●●●
●●
●
●●●●●●●
●
●●●
●●
●●
●●●●●●
●
●
●●
●
●
●●●●●●
●
●●
●●●
●
●●
●
●●●●●●●●●●
●
●●●●●●●●●●●●
●●●●●●●
●●●●●●●●●
●●
●
●●
●●●●
●●
●●●●●●●
●●
●
●●
●●
●●●
●●●●●●●●
●●
●●●●
●
●●●●●
●●●●●●●●
●
●●
●●
●
●●●
●●●
●●●●●
●●●●
●
●
●●
●
●
●●
●
●
●●
●
●
●
●●
●●●
●●
●
●
●●
●●●
●●
●●●●●
●●
●
●●
●●
●●●●
●
●
●
●
●●
●●
●●●
●●
●●
●
●●●●
●
●
●●
●
●
●●
●●
●
●●●
●
●
●●●●
●
●
●
●●●●●
●
●
●
●●●
●
●
●●●●
●
●●
●●
●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●●
●●
●●●●●●●
●●
●●●
●
●
●
●
●●
●
●
●
●
●
●●●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●●●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●●
●
●
●
●●
●
●●
●●●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●●●●
●
●●●
●●●●●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●●
●
●
●
●●
●●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●●●
●
●
●●
●
●●
●
●
●
●
●●
●●
●
●
●
●●
●●●
●
●●
●●
●
●
●
●
●●
●●●
●
●●
●
●
●
●
●
●●●●
●
●●●●●●●
●
●
●
●●
●●●●
●
●●
●
●●
●●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●●
●●●
●
●
●●
●
●
●●
●●●
●
●
01
23
4
Re
co
mb
ina
tion
(cM
/Mb
)
●●●●●●
●●●
●
●●●
●
●
●
●
●
●●
●
●
●
●●
●●●●●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●●
●
●
●●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●●●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●●
●
●
●●
●
●
●●
●
●●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●●
●
●
●
●
●
●
●
●●
●
●●
●●
●●
●●
●●
●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●●
●●●●●●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●●
●
●●
●
●
●●
●
●
●
●
●
●●
●●
●
●●●●●
●
●
●
●
●●
●
●
●●
●●
●●●
●
●
●●
●●
●●●
●
●
●
●
●●
●
●●
●
●
●
●
●●●●●●
●
●●●●
●●
●
●●●●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●●●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●
●●
●●
●
Recombinationpi
●
●●●●●●●●●●●●●●●●●
●●●●●●
●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●
●●●●
●
●
●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●
●●●●●●●
●
●●●●●●
●●●●●
●●●
●●
●●
●
●●●●●
●
●
●●
●
●
●●●
●
●●
●●●●●●●●●●●●●●
●●●●●●●●●
●
●
●
●●●●●●●●●●●
●●
●●●●●
●●●●
●
●●●●●
●●
●
●●
●
●●
●●●●
●●
●●
●●●●
●
●●●●●●●
●●
●
●●●●●●●
●
●
●
●●●●●●●●
●●●●●●
●
●●●●●●
●●●●●●●●●●●●●●●●●●
●
●●●
●●
●●
●●
●●
●
●
●
●●●
●●
●●●
●
●
●●
●
●
●●●●●●
●
●●
●
●●●●●●●●●●●●●
●●
●●●●●●●●●●●●●●●●
●
●●●●●●●●
●●●●
●
●●●●
●
●
●●●●●●
●●●●●●
●
●●●●●●●●●●●●●●
●●●●
●●●●●●●
●
●●●●●●●
●
●●●●●●●●●●●●●●●●
●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●
●●●
●●
●●●
●●●
●
●●
●●●●●
●
●●●●●●
●●●
●●●●●●●●●
●●●●●●●●
●
●●●●●
●●●●●●●●●●●●●●●
●
●●●●●●●●
●●
●●
●●●
●●●●
●
●●●●
●
●
●
●●●●
●
●●●
●
●
●
●
●
●●●●
●
●
●
●
0.0
0.1
0.2
0.3
0.4
0.5
k −
Div
erg
en
ce
b)
●
●
●
●
●
●●●●
●●●
●
●●●●
●●●●
●●●●●●●●●●
●
●●●●●●●●
●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●
●
●●●●●●
●●●●●
●●
●●●●●●●●●●●
●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●
●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●
●●●●●●●●●●●●●●●●●●●●●
●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●
●●●
●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●
●●
●
●
●
●
●●●
●●●
●
●
●●●
●
●●
●
●
●●●●●●
●●●●●
●●●●●●
●●
●
●●●●●●●●●
●●●●●
●●●●
●
●
●●●
●
●●●●
●
●●
●●●●●●
●●
●
●●●
●●●
●●
●●
●
●●●●●●●●
●●●●●●
●
●●●●●●
●●●
●●●●●●
●●●
●●●●
●
●●●●●●●
●
●●●●●●●●●●●●●
●
●
●●
●
●
●
●●●●●
●●
●●
●
●
●
●●●●●●●●
●●●●●●●●●●●
●●●●●●●
●●●
●●●
●
●
●
●●
●
●●●●●
●●●●●●●●●
●
●●●●●●●●●●●●●
●●●
●
●
●
●●●●●●
●●●●●●
●
●●●
●
●●
●●●●●●●●●●●●●●●●●●●●●●
●●●
●●
●●●●●●●●●
●●●
●
●●●
●
●●●
●●●●●●●●
●●●
●●
●●●●●●●●●●●●●●●●●●●●
●●●
●●●●
●●●
●●●●●●●●●●
●●●
●
●
●●●●●●
●●●●●
●●
●●
●●●●●●●●
●●●●
●
●
●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●●
●●●●●●●●●●●
●
●●●●●●
●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●●
●●●●●●
●●●●●●●●●●●●●
●●●
●●●
●●●●●●●●
●●●●●●●●●●●●●●
●●
●●
●●●●●
●
●●
●
●●●●●
●●●
●
●
●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●
●
●●●●●●●●●●●●●●●
●
●●●●
●●●●●●●●●
●
●●
●●●●●●●●●
●●●●
●
●
●
●●●●●●●●●●
●●
●●●●●●●
●●●●●●●●●●●●●●●●●●
●●
●
●●●●●●●
●
●●●●●●●●
●●●●●●●●
●
●●●
●
●●●
●●
●●●●●●●●●●●●●●●
●●●●●●●●
●
●●●
●●●
●
●●●●●●●●
●●
●
●●●●●●●●●●●●●●●
●●
●
●
●●●●●●●●●
●●●●
●
●●
●●●●●
●●●●
●●●●●●●●●
●●●
●
●●
●●
●
●
●●●●
●
●
●
●
●
●●
●●●
●●●●
●●●●
●●
●●●●●●
●●
●
●
●
●●●
●
●●●●●
●
●●
●●●●●●●
●
●●●●
●●●
●●●●
●
●●
●
●●●●●
●
●
●
●●●●
●●●●
●●
●●
●●●●●●●●●
●
●●
●●●
●
●
●
●
●●
●●
●
●
●●
●●●
●●
●
●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●
●●●
●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●
●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●
●●●
●
●
●
●
●
●
●
●●●
●●●●●●●
●●●●
●
●●●●●●●●●
●
●●
●●●●
●
●●●●
●●●●●●●●
●●●●
●
●●
●●●
●
●●
●
●
●
●
●●●●●●●●●
●
●●●●
●●●●●●●●●●●●●●●●●●●●●
●
●●
●●●●●●●●●
●●●●●●
●●●●●●
●
●●●●
●●●●●
●●●●
●●
●●
●●●●●●
●
●●●●●●●●●
●●●
●●●●●●●●●●●●
●●●●
●●
●●●●●●
●●
●
●
●
●
●●●●●●●●●●
●●●●●
●
●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●
●
●●
●●
●●●●●●●●●
●●●●●
●●●●●●●
●●
●●●●●●
●●●●●
●●●●●
●●●●●●●
●●●
●●●●
●●●
●●●
●
●●
●●●●
●●●
●●●●●●●●●●●●●
●
●●
●●●●●●●●●
●
●
●
●
●●●●
●
●
●
●●●
●
●●●●
●●
●●
●●●●
●
●●●●●●
●●●●●●
●
●●●●●
●
●●●●●●●
●●●●●●●●●
●●
●
●
●●●●●●●
●
●●●
●
●●●
●
●●●●●●●●●
●●●●●●●●●●
●●
●
●●●●●●●●●●●
●
●●●●●●●●●
●
●●●●●
●●●●●●●●
●●●●●●
●●
●●●●●●●●●●●●●●
●
●●●●●●
●
●●●●●
●●●
●●
●●●
●●●●
●●
●●●●●●●●●●●
●●●●
●●●●●
●●
●
●
●
●●
●●
●●●●●●●●
●
●●●●
●
●●●●●●●
●●●
●●●●●●
●●
●
●●
●●
●
●●
●●●●●●●●●
●●●●●●●●●●●
●●●●
●●●
●●●●●●●●●●●
●●●●
●
●●●
●●●●●●●●●
●
●
●
●●●●
●
●●●●●●
●●●●
●●
●●
●
●●●●●●
●
●●●●●●●●●●
●●●●●●●●●●●●●●●●
●●●●
●
●●●●●
●
●●●
●●●●●●●●
●
●●●
●
●●●
●
●●●●●●●●●●●●●
●
●
●●
●
●●●●●●
●
●●●●●
●
●
●●●●●●
●
●
●●●●●●●●
●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●
●●●
●
●●
●
●●●●●●●●
●●●●
●●●●●
●●●●●●
●●
●●●
●●
●●●
●●●●●●●●●●
●
●
●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●
●●●●●●●●●
●
●●●●●●●●●●●●●●●●
●
●
●●●●
●
●●
●●●
●●●●●●●●●●●●●●●●●
●
●●●●
●●
● ●●
●
●
●
●●●
●
●
●
●
●●
●●●●
●
●
●
●●●●
●
●●●●●●●●●
●●●●
●●
●
●●
●
●
●
●
●●
●
●
●●
●●●●●
●●●●
●●
●●
●
●●●●●●●●●●●
●●●
●●●●●●
●●
●
●●
●●
●●●
●
●●●●
●●●●
●
●
●
●
●●
●●●●●●
●●●●●●
●
●●
●●●●●●●●●●
●
●
●●
●
●●●
●●●●●●●
●●
●●●●●●
●●●
●
●●
●
●
●
●●●
●●
●●●
●●●
●●
●
●●●●
●●●●●
●●
●●●●●●●●●
●●
●
●●
●●●
●●
●
●●●●●●●
●●●●
●
●●●●●
●
●
●●
●●●●●●●
●●
●●
●●●
●●
●●●●●
●●
●●●●●
●
●●
●
●
●●●
●●●●●●●●
●●●●●●●
●
●●●●●●
●
●●●●●●
●
●
●
●●●
●●●
●●
●●
●
●
●
●
●●
●●●●
●●●●●●●
●
●
●
●●●●●●
●●●●●
●
●●●●
●
●
●●●
●
●●●●
●
●●●●●●●
●●●●●
●
●●
●●
●●●●
●
●●●●●
●
●●●●
●
●
●
●
●
●
●●●●
●
●●●●●●●●●
●●●●●
●●●
●
●
●●●
●●
●
●
●
●
●●
●
●
●
●
k dyakk dsim
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●●
●
●●
●
●
●
●
●●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●●
●●
●●
●●
●●
●
●
●●●
●
●
●
●
●●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●●
●
●
●
●
●
●
●●●●●●
●
●●●
●
●
●
●
●
●
●●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●●
●●●●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●●
●
●●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
−5
−4
−3
−2
−1
01
Ratio P
ol/D
iv
2LTel Cent
c)
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●●
●
●
●
●
●●
●●
●
●
●●●●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●
●●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●●●●
●
●●●
●
●●
●●
●
●●●
●
●
●
●
●●
●
●
●
●
●●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●●●
●
●
●●
●
●
●●●●●●●
●
●●
●
●
●
●●
●●
●
●
●
●●
●
●●●●●●●●●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●●●
●
●●●●
●
●
●●
●●●
●●
●
●
●
●
●
●
●●●
●
●●●
●●●
●
●
●
●●
●
●●●●
●
●
●●●
●
●●●●●●
●
●●●●●●
●
●
●
●
●●●●
●
●●
●●●●
●
●●●
●●●●
●
●
●
●●●
●●●
●
●
●
●
●
●●
●
●●
●●●
●●
●
●
●
●
●
●●●●
●●●●●●●●●●●●●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
2R Tel Cent
●
●
●
●
●
●●
●●●
●●●
●
●
●●
●
●●
●
●●●●●
●
●
●
●●●●
●●
●
●
●●●
●
●
●
●
●
●●●
●
●●●
●
●●
●
●
●
●●
●
●●
●
●●●●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●●●
●●
●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●●●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●●●
●
●
●●●●
●●
●●
●
●●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●●●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●●●●
●●
●
●
●
●
●
●
●
●
●
●●●●
●
●●●●●●●●●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●●●●●
●●
●
●
●●●
●
●
●●
●
●
●
●●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●●●●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●
3L Tel Cent
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●●
●
●
●●●
●●
●●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●●
●●●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●●
●
●
●
●
●
●
●
●
●●
●●
●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●●
●
●●
●●
●
●
●
●●
●●
●●
●●
●
●●
●
●
●
●●●
●
●
●
●
●
●●
●
●●●
●
●●●
●
●●
●
●
●
●
●
●●
●●
●●●
●
●
●●
●●
●
●
●
●
●
●
●●●●●
●●●
●●●
●●
●
●
●
●●
●●
●
●
●●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●●●●
●
●●
●
●●
●
●
●
●●
●
●
●
●●●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
3R Tel Cent
●
●
●
●●
●
●
●
●
●●●●
●●●
●
●
●●
●●
●●
●●
●●
●●●●●●●
●
●
●●
●●●●●●●
●
●●●●●
●●
●
●
●
●●
●
●
●●
●●
●●●●
●●●
●
●●
●
●●
●●
●
●●●
●●
●
●●●●
●
●
●●●●●●
●●
●
●
●●
●●
●
●●
●
●●●
●
●●
●
●●●●●●●●●●●●
●
●●●
●
●
●●●●
●
●
●●●●●●●●●●●
●
●●●●
●
●
●●
●
●●●
●●●●●●●●●●●●●●●●●●●●
●●●
●
●
●●●●●
●
●●●●
●
●●
●●●
●●●●
●
●
●
●●●●●●●
●●●●●●
●
●
●
●●●●●●
●
●
●
●
●●●
●●
●
●●●
●●
●
●
●
●●●●●
●●
●
●●●
●
●
●●
●●
●
●
●●●●
●
●●●●
●
●●●●●●●●
●
●
●
●
●
●
●●
●●
●
●
●●●●
●●●●●●●●
●●●●●
●
●
●
●
●
●
●●●●
●
●●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●●●●
●●●
●
●●●●
●●●●●●●●●●●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●●●●●●●
●●●●
●
●●●●
●
●
●
●●
●
●
●
X Tel Cent
α calculated aggregating for windows along chromosome arms on non-
synonymous sites (0fold)
Adaptive selection
2L 2R 3L 3R X
Tel Cen Cent Tel Cen Cen Cen Tel Tel Cen
α = 1 neutral region
α > 1 adaptive selection
α < 1 negative selection
α = 1
Significant MKT on individual coding genes (syn vs. non-syn)
Adaptive propensity and recombination Conclusions
The 158 genomes has allowed the
unprecedented opportunity to perform the most comprehensive nucleotide variation study done so far in any species
By means of a battery of comparative methods for polymorphism and divergence data we try to answer questions of fundamental interest in population genomics

1/18/2012
12
Conclusions
(1) The genome patterns of polymorphism and divergence differs manifestly
(i) centromeric vs. non-centromeric regions within autosome arms
(ii) autosomes vs. X chromosome
(2) Natural selection is pervasive along the D. melanogaster genome, and the relative importance of different selection regimes depends on both the site classes and the genome regions considered
(3) There exists a threshold value of recombination rate which defines the efficiency of selection for any genome region
(4) All evidence together supports the fast X-hypothesis Bioinformatics of Genetic Diversity Genomics, Bioinformatics and Evolution
Research group