
Freeman 1e: How we got there -...
53
• Bakterijska ćelija - ćelijski rast uključuje: - veliki broj raznorodnih hemijskih reakcija, - transformacija energije i biosinteza u najširem smislu RAST MIKROORGANIZAMA mali molekuli neophodni za sintezu makromolekula kofaktori i koenzimi neophodni za enzimatske reakcije polimerizacija sastavljanje struktura za nove ćelije
Transcript of Freeman 1e: How we got there -...

• Bakterijska ćelija - ćelijski rast uključuje:
- veliki broj raznorodnih hemijskih reakcija,
- transformacija energije i biosinteza u najširem smislu
RAST MIKROORGANIZAMA
mali molekuli
neophodni za sintezu
makromolekula
kofaktori i koenzimi
neophodni za
enzimatske reakcije
polimerizacija sastavljanje struktura
za nove ćelije

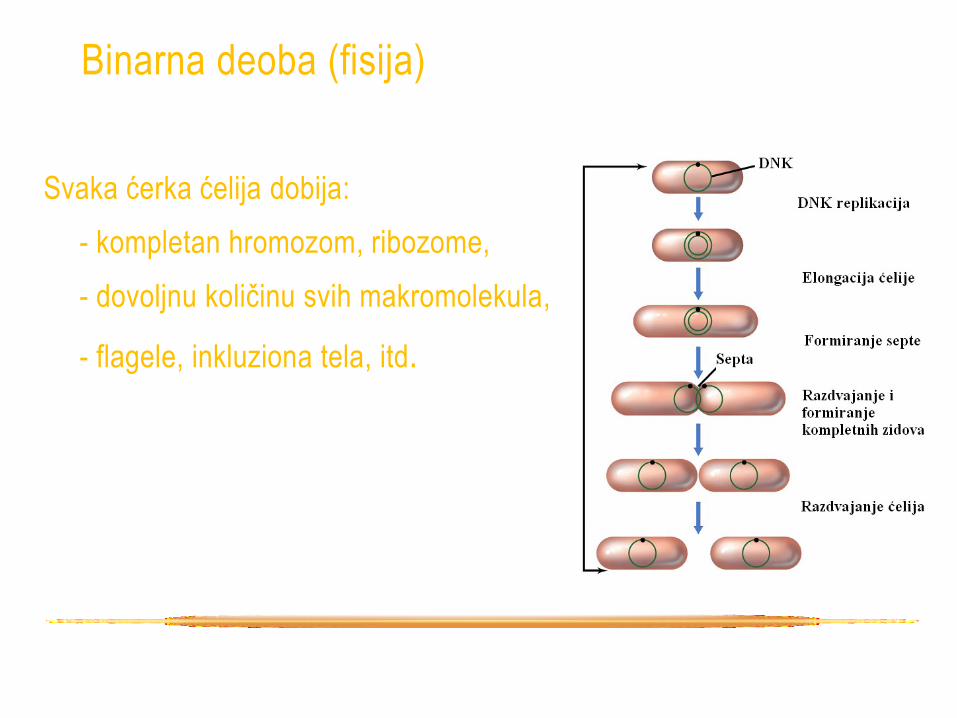
Binarna deoba (fisija)
• U bakteriologiji rast - povećanje broja ćelija (binarna
deoba = 2 ćelije od 1).
• Rast individue prethodi podeli na dve ćerke ćelije (E. coli
se izdužuje 2 puta pre deobe).
• Formiranje septe - rast citoplazmatične mebrane i
ćelijskog zida sa suprotnih strana ka unutrašnjosti, sve
dok se ćelije ne razdvoje
• Jedna ćelija se podeli na dve = 1 generacija
• Vreme potrebno za to = vreme generacije
Binarna deoba (fisija)
Svaka ćerka ćelija dobija:
- kompletan hromozom, ribozome,
- dovoljnu količinu svih makromolekula,
- flagele, inkluziona tela, itd.
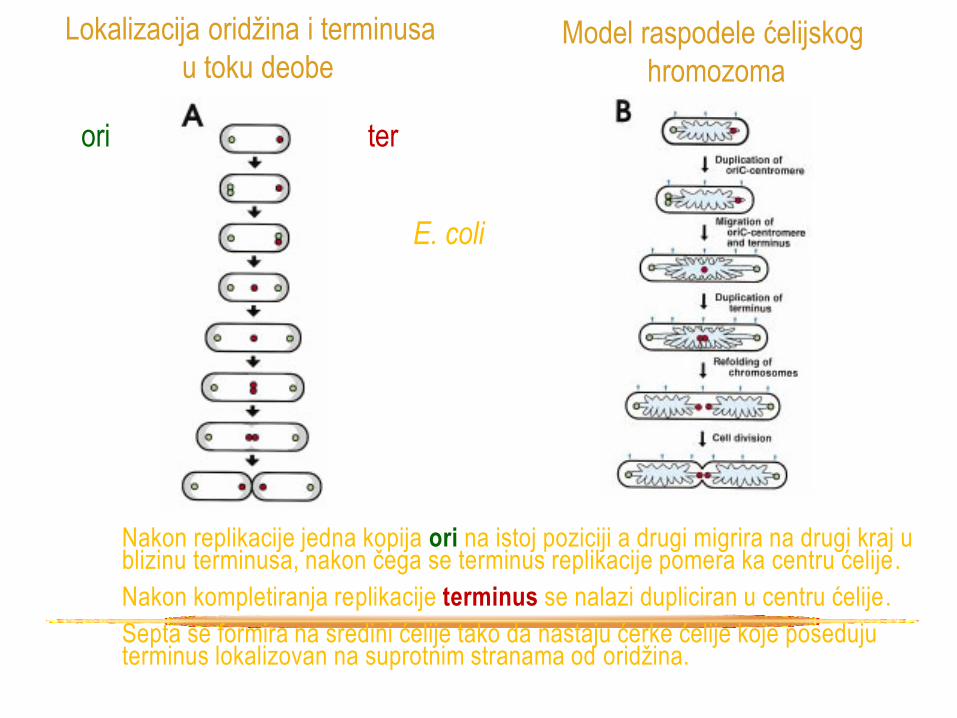
Nakon replikacije jedna kopija ori na istoj poziciji a drugi migrira na drugi kraj u blizinu terminusa, nakon čega se terminus replikacije pomera ka centru ćelije .
Nakon kompletiranja replikacije terminus se nalazi dupliciran u centru ćelije.
Septa se formira na sredini ćelije tako da nastaju ćerke ćelije koje poseduju terminus lokalizovan na suprotnim stranama od oridžina.
Lokalizacija oridžina i terminusa
u toku deobe Model raspodele ćelijskog
hromozoma
E. coli
ori ter

Fts proteini i ćelijska deoba
• Fts proteini (filamentous temperature sensitive)
• FtsZ – ključni protein grupe, dobro proučen kod E. coli
• univerzalnan kod prokariota, čak i u hloroplastima i
mitohondrijama
• strukturna sličnost sa tubulinom eukariota
• "divisom" - "aparat za deobu"
• FtsZ definiše ravan deobe
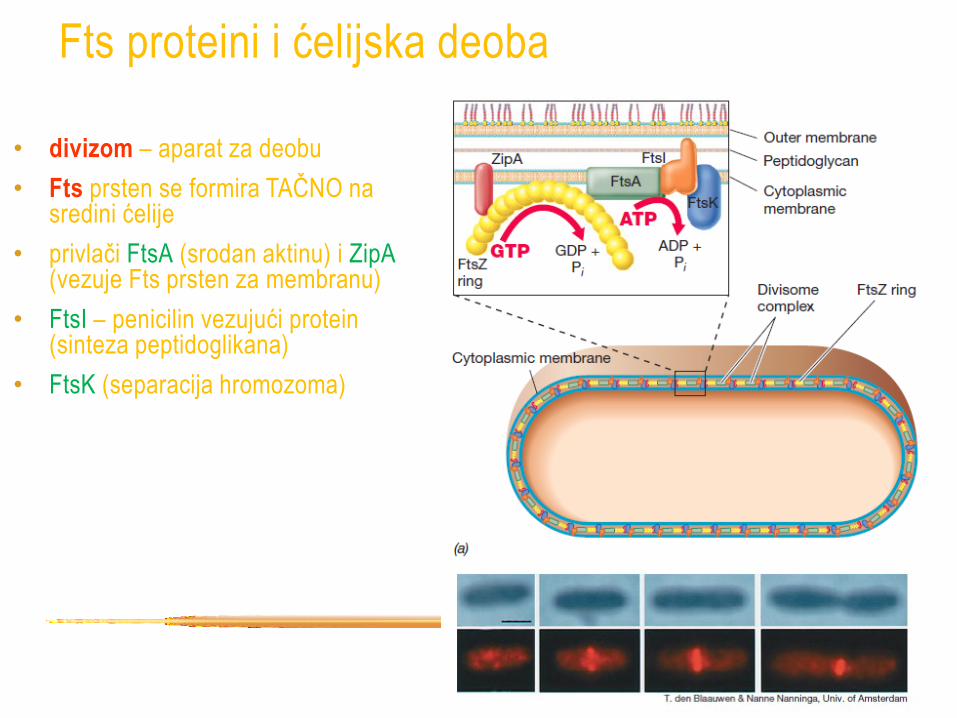
• divizom – aparat za deobu
• Fts prsten se formira TAČNO na sredini ćelije
• privlači FtsA (srodan aktinu) i ZipA (vezuje Fts prsten za membranu)
• FtsI – penicilin vezujući protein (sinteza peptidoglikana)
• FtsK (separacija hromozoma)
Fts proteini i ćelijska deoba
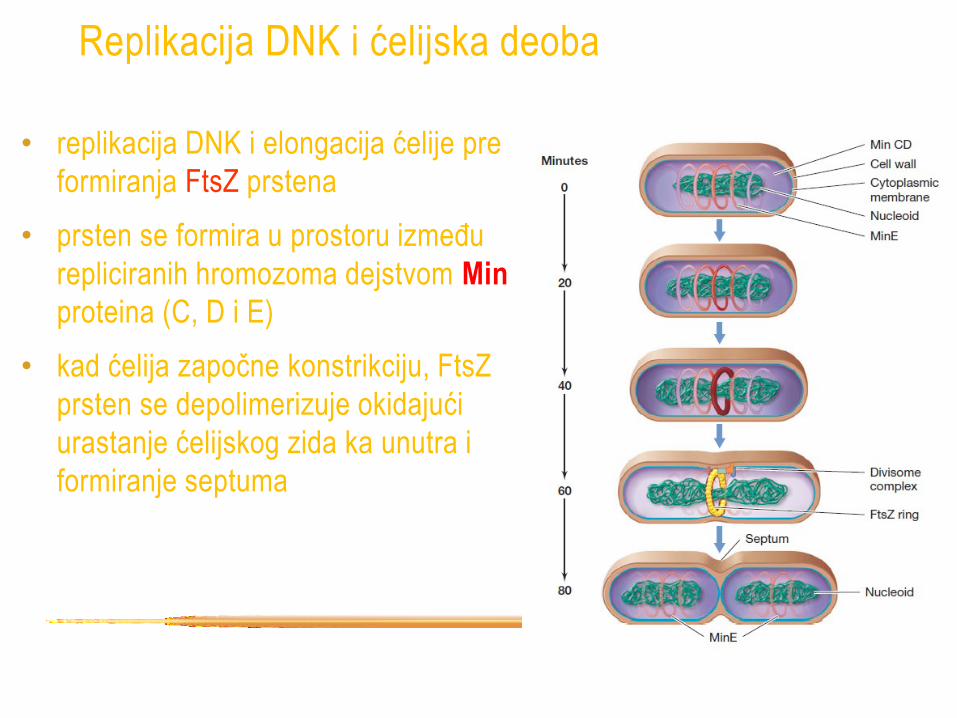
Replikacija DNK i ćelijska deoba
• replikacija DNK i elongacija ćelije pre
formiranja FtsZ prstena
• prsten se formira u prostoru između
repliciranih hromozoma dejstvom Min
proteina (C, D i E)
• kad ćelija započne konstrikciju, FtsZ
prsten se depolimerizuje okidajući
urastanje ćelijskog zida ka unutra i
formiranje septuma
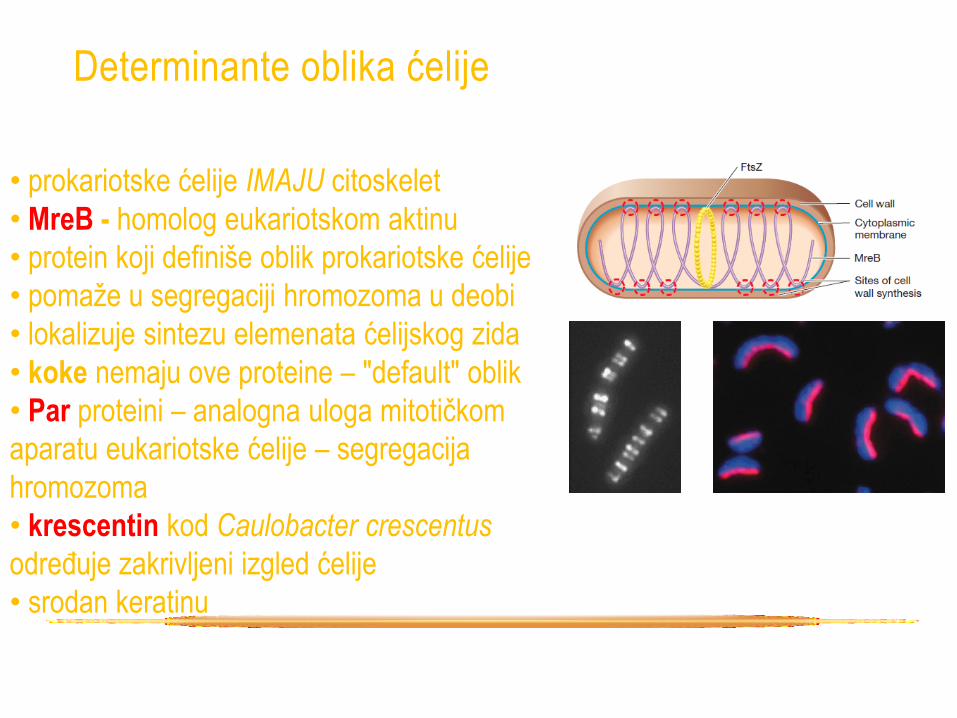
Determinante oblika ćelije
• prokariotske ćelije IMAJU citoskelet
• MreB - homolog eukariotskom aktinu
• protein koji definiše oblik prokariotske ćelije
• pomaže u segregaciji hromozoma u deobi
• lokalizuje sintezu elemenata ćelijskog zida
• koke nemaju ove proteine – "default" oblik
• Par proteini – analogna uloga mitotičkom
aparatu eukariotske ćelije – segregacija
hromozoma
• krescentin kod Caulobacter crescentus
određuje zakrivljeni izgled ćelije
• srodan keratinu

• MreB = aktin (mikrofilamenti eukariota – citoskelet, citokineza)
• FtsZ = tubulin (mikrotubule eukariota – mitoza)
• krescetin = keratin (intermedijarni filamenti eukariota – citoskelet)
Evolucioni koreni citoskeleta eukariota

Sinteza ćelijskog zida i ćelijska deoba
• Autolizini - formiraju otvore u zidu raskidajući β-1,4 glikozidne veze
• Kontrolisano isecanje delova starog zida i simultano "krpljenje" rupa prekursorima peptidoglikana
• Baktoprenol - hidrofobni nosač dodaje disaharid pentapeptid (prekursori ćelijskog zida) kroz membranu
• Transglikozilaze povezuju prekursore sa ostatkom zida na odgovarajućem mestu formiranjem glikozidne veze
• Transpeptidacija – finalni korak u sintezi zida
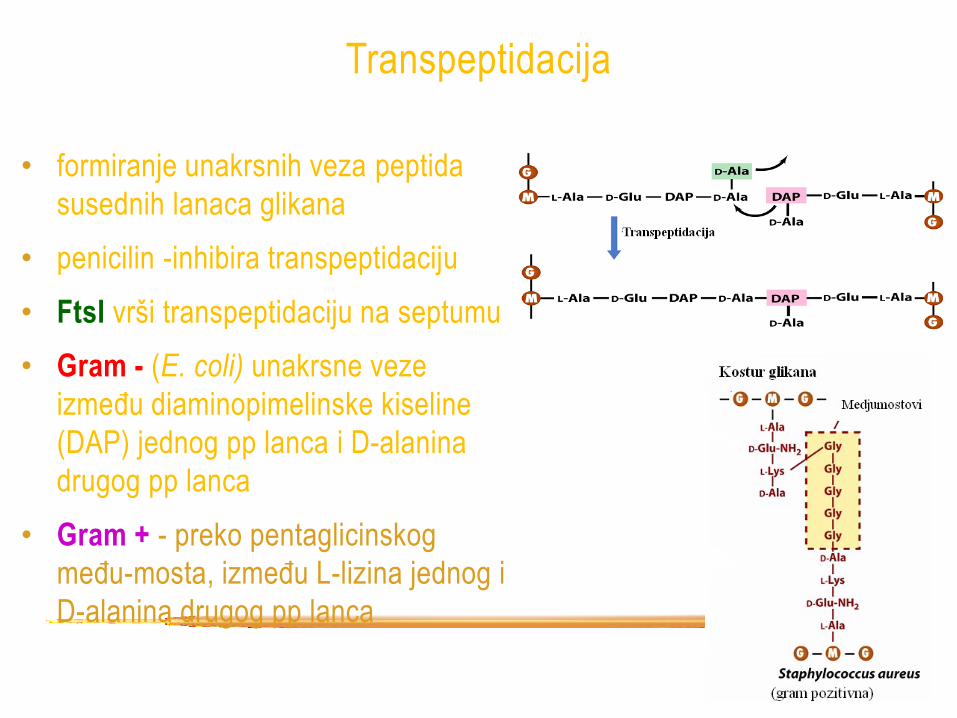
Transpeptidacija
• formiranje unakrsnih veza peptida
susednih lanaca glikana
• penicilin -inhibira transpeptidaciju
• FtsI vrši transpeptidaciju na septumu
• Gram - (E. coli) unakrsne veze
između diaminopimelinske kiseline
(DAP) jednog pp lanca i D-alanina
drugog pp lanca
• Gram + - preko pentaglicinskog
među-mosta, između L-lizina jednog i
D-alanina drugog pp lanca
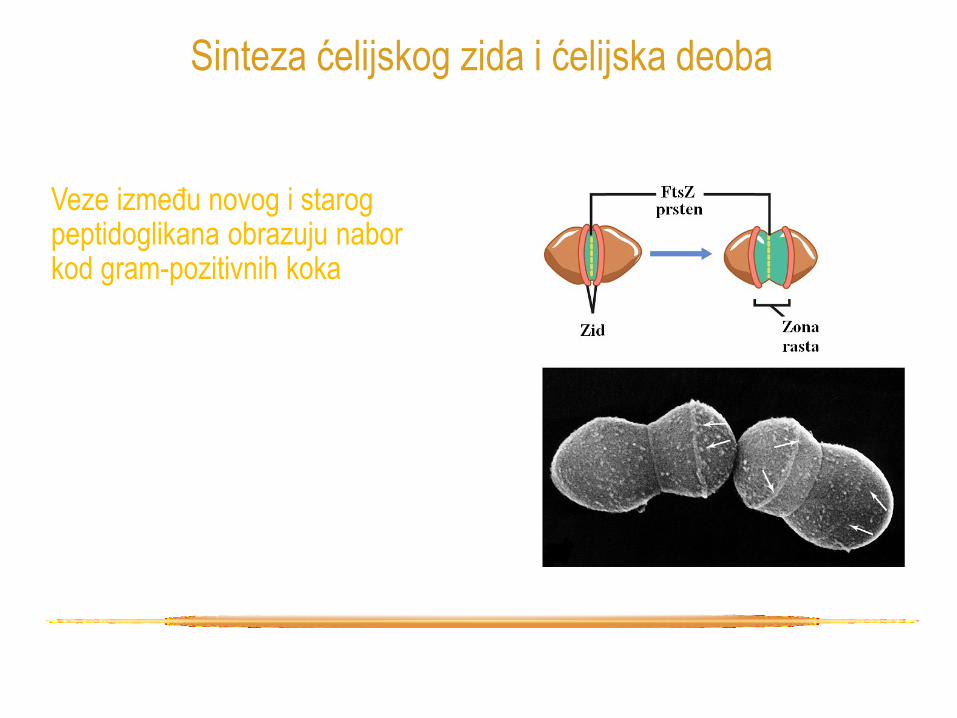
Veze između novog i starog peptidoglikana obrazuju nabor kod gram-pozitivnih koka
Sinteza ćelijskog zida i ćelijska deoba

Rast bakterijske populacije
• Rast - povećanje broja ćelija m.o. u populaciji
• Može se meriti i kao povećanje mase mikroorganizama
• Brzina rasta - promena u broju ili masi ćelija u jedinici vremena
• Generacija - interval za koji se formiraju dve ćelije od jedne
• Vreme generacije – vremenski period za koji se duplira ćelijska populacija (0.5 - 6h)
• Različito u lab uslovima i u prirodi

Rast bakterijske populacije
• Populacije mikroorganizama pokazuju
eksponencijalni rast
• Povećanje broja ćelija u funkciji vremena
• Vreme neophodno za kompletan životni ciklus
bakterija je veoma varijabilno i zavisi od uslova
spoljašnje sredine i genotipa
• U optimalnim uslovima životni ciklus E. coli se
odvija za 20 minuta
• Praktične implikacije eksponencijalnog rasta

Određivanje vremena generacije

• Tipična kriva = četiri faze
Kriva rasta bakterija

• Lag faza se uočava:
- ako se ćelije iz stare kulture (stacionarna faza) prenesu u istu vrstu medijuma pod istim uslovima, iako sve ćelije u inokulumu mogu biti vijabilne
- ako nedostaju esencijalni faktori (neophodno vreme za sintezu)
- ako su ćelije oštećene različitim tretmanima (temperatura, zračenje ili toksični agensi) potrebno je vreme da se isprave oštećenja
• Nema lag faze ukoliko se eksponencijalna kultura inokulira u istu vrstu medijuma pod istim fizičkim uslovima.
Lag faza

Log faza
• Faza intenzivnih deoba ćelija
• Brzina rasta zavisi od:
- genetičkih karakteristika
samog organizma,
- uslova spoljašnje sredine
(temperatura, pH, osmotski
pritisak, količina kiseonika,
sastav hranljivog medijuma)
Vreme generacije
• Izračunava se iz dela krive sa
eksponencijalnim rastom
• Pokazatelj fiziološkog stanja
populacije
Eksponencijalna faza (log) i vreme generacije

• Stacionarna faza - nema promene u broju ćelija.
• U zatvorenom sistemu (kulturi) rast nije neograničen, utrošene
su neophodne hranljive materije a neki produkti metabolizma
mogu inhibirati rast.
• Neke ćelije se dele, neke umiru.
• Faza smrti – nastavljanjem inkubacije ćelija u stacionarnoj
fazi može doći do smrti većine ćelija.
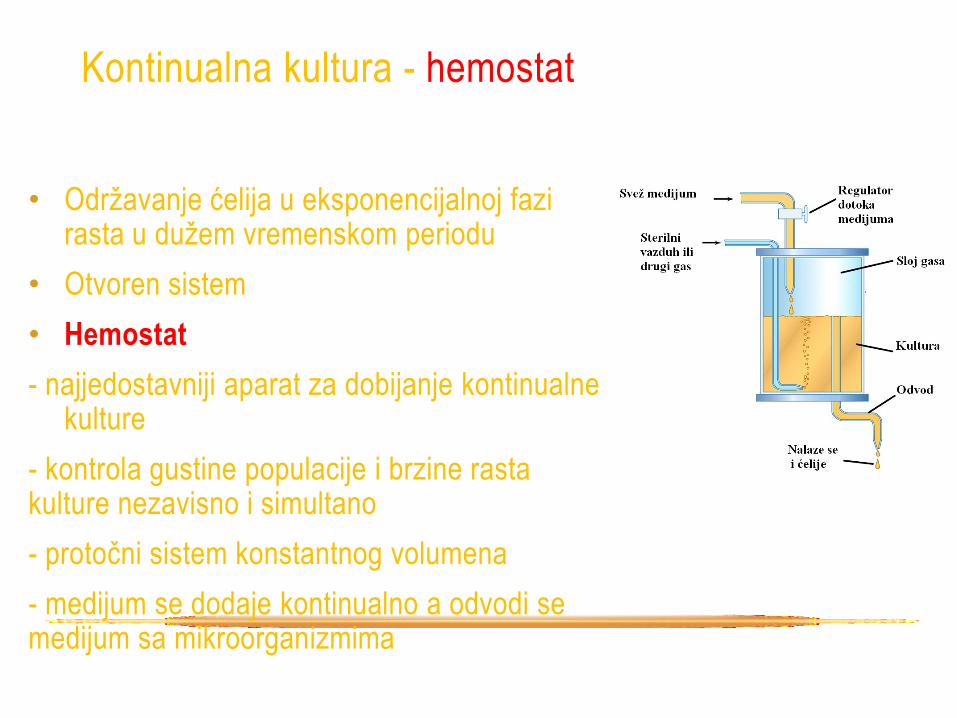
Kontinualna kultura - hemostat
• Održavanje ćelija u eksponencijalnoj fazi rasta u dužem vremenskom periodu
• Otvoren sistem
• Hemostat
- najjedostavniji aparat za dobijanje kontinualne kulture
- kontrola gustine populacije i brzine rasta kulture nezavisno i simultano
- protočni sistem konstantnog volumena
- medijum se dodaje kontinualno a odvodi se medijum sa mikroorganizmima

Elementi u kontroli rasta kulture u hemostatu
• Kontrola brzine rasta i prinosa je
nezavisna:
- brzina rasta - podešavanjem razređenja
- prinos - variranjem koncentracija
hranljivih materija prisutnih u limitirajućoj
količini
• Koncentracija hranljivih materija (izvori
ugljenika i azota)
• Ukoliko se ove materije nalaze u niskoj
koncentraciji rast je redukovan

Fermentori
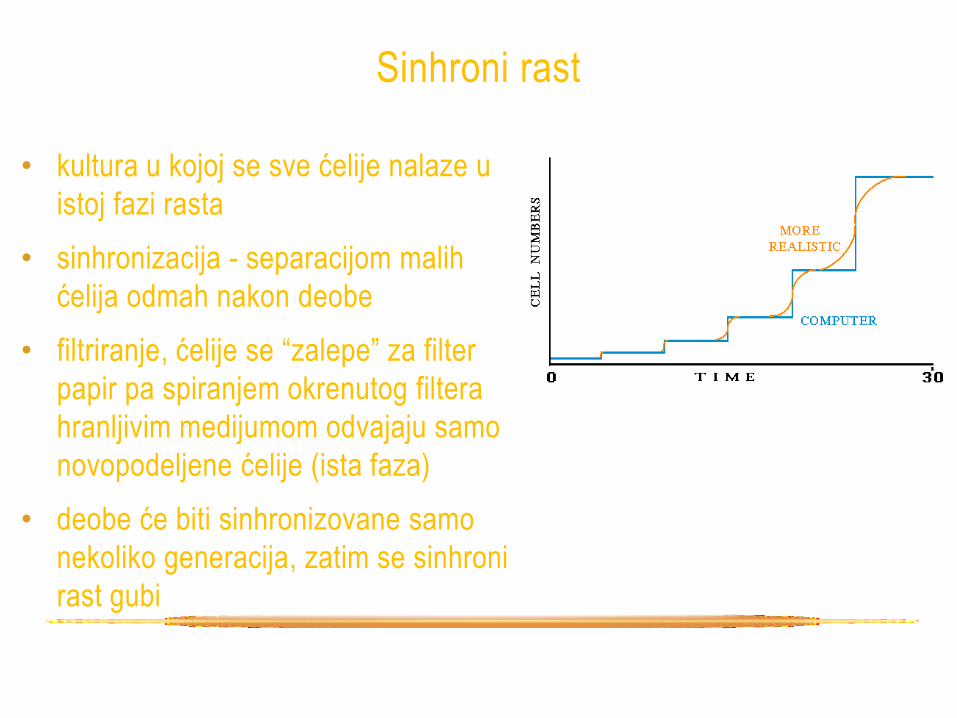
Sinhroni rast
• kultura u kojoj se sve ćelije nalaze u
istoj fazi rasta
• sinhronizacija - separacijom malih
ćelija odmah nakon deobe
• filtriranje, ćelije se “zalepe” za filter
papir pa spiranjem okrenutog filtera
hranljivim medijumom odvajaju samo
novopodeljene ćelije (ista faza)
• deobe će biti sinhronizovane samo
nekoliko generacija, zatim se sinhroni
rast gubi

Merenje rasta mikroorganizama
• Merenje rasta populacije:
- praćenje broja ili mase ćelija
• Veći broj metoda za određivanje broja ili mase ćelija
prilagođenih za različite organizme
• Direktne i indirektne metode
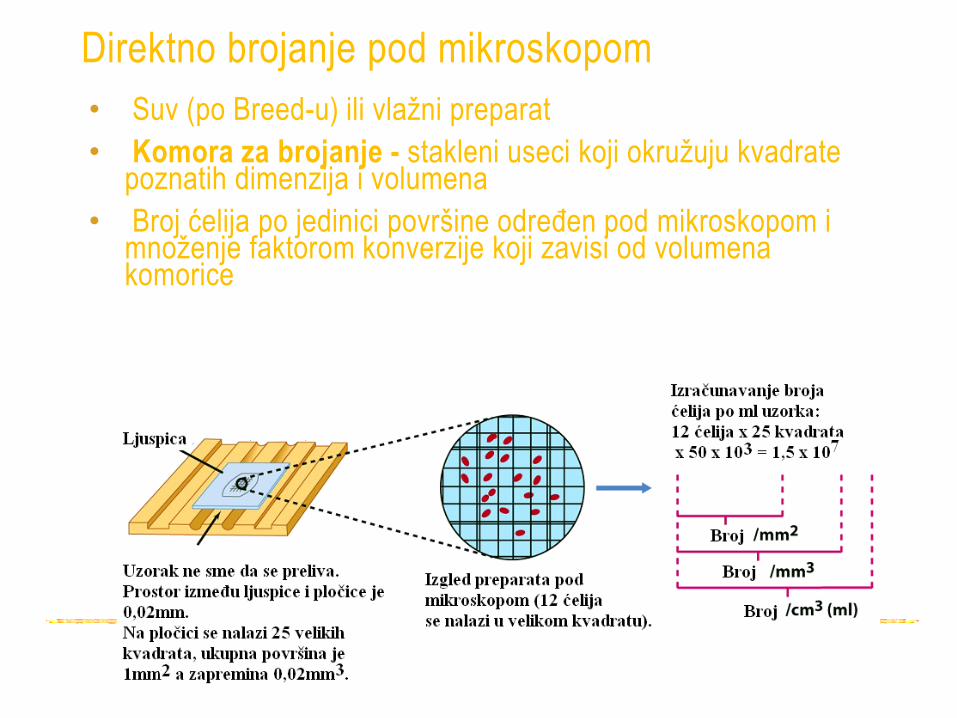
Direktno brojanje pod mikroskopom
• Suv (po Breed-u) ili vlažni preparat
• Komora za brojanje - stakleni useci koji okružuju kvadrate poznatih dimenzija i volumena
• Broj ćelija po jedinici površine određen pod mikroskopom i množenje faktorom konverzije koji zavisi od volumena komorice

Direktno brojanje pod mikroskopom
• prednost – brzo se dobijaju rezultati
• mane – broje se i mrtve ćelije; male ćelije je teško videti; teško je postići preciznost; kad uzorak nije obojen potreban je fazno-kontrasni mikroskop; nije pogodan za uzorke male brojnosti (ispod milion po ml); pokretne ćelije se moraju prethodno imobilizovati
• DNK-specifično bojenje = ukupni broj u mikrobijalnoj ekologiji (fluorescentni mikroskop)
• filogenetsko bojenje
• hemocitometri
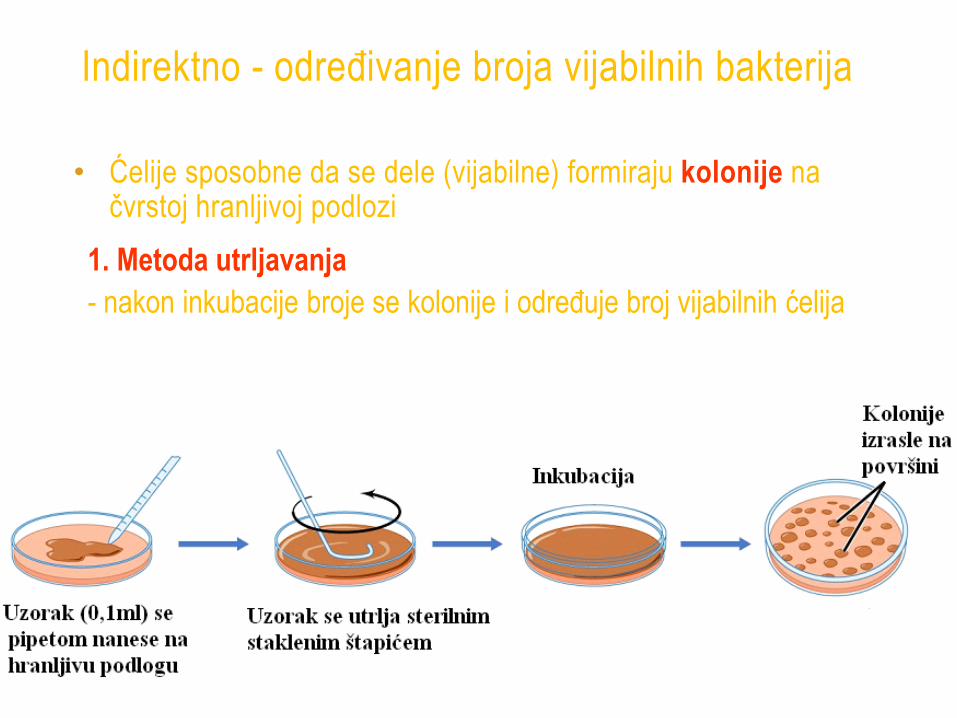
Indirektno - određivanje broja vijabilnih bakterija
• Ćelije sposobne da se dele (vijabilne) formiraju kolonije na čvrstoj hranljivoj podlozi
1. Metoda utrljavanja
- nakon inkubacije broje se kolonije i određuje broj vijabilnih ćelija
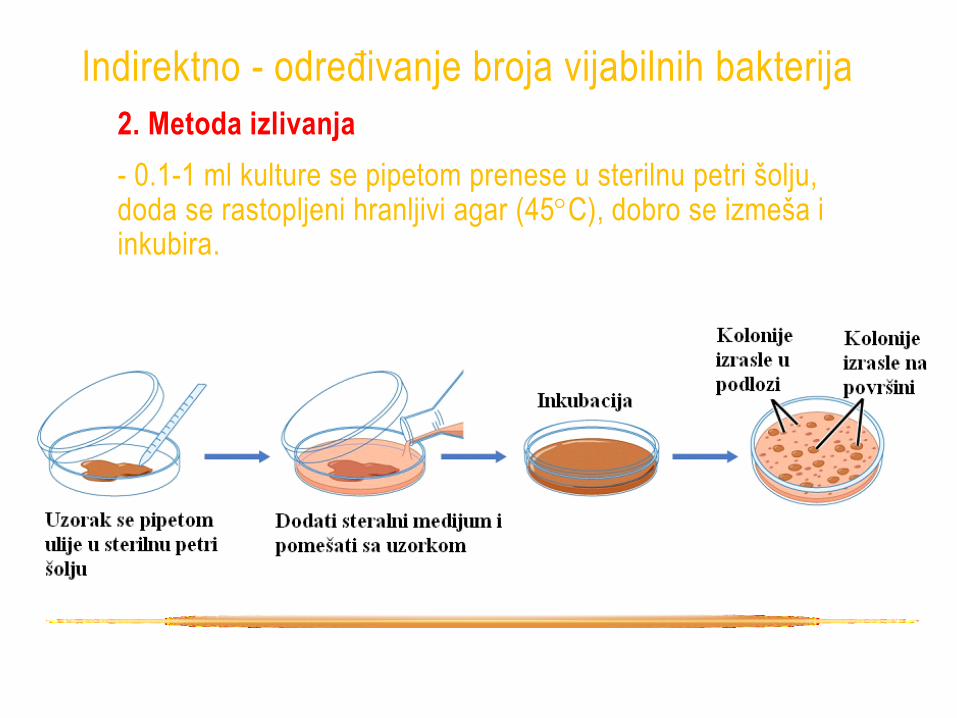
2. Metoda izlivanja
- 0.1-1 ml kulture se pipetom prenese u sterilnu petri šolju, doda se rastopljeni hranljivi agar (45C), dobro se izmeša i inkubira.
Indirektno - određivanje broja vijabilnih bakterija

• Da bi se dobio odgovarajući broj kolonija uzorak skoro uvek mora biti razređen (napravljena dilucija).
• Retko se zna približan broj vijabilnih ćelija u uzorku pa je neophodno napraviti veći broj različitih razređenja.
• Koriste se decimalna razređenja
• Najčešće je neophodan serija decimalnih razređenja.
Razređenja (dilucije)
10 x = 10-1
0.5 ml uzorka i 4.5 ml fr ili
1 ml uzorka i 9 ml fr
100 x = 10-2
0.05ml uzorka i 4.95 fr ili
0.1 ml i 9.9 ml fr
Milion x = 10-6
3 sukcesivna 10-2 ili
6 sukcesivnih 10-1
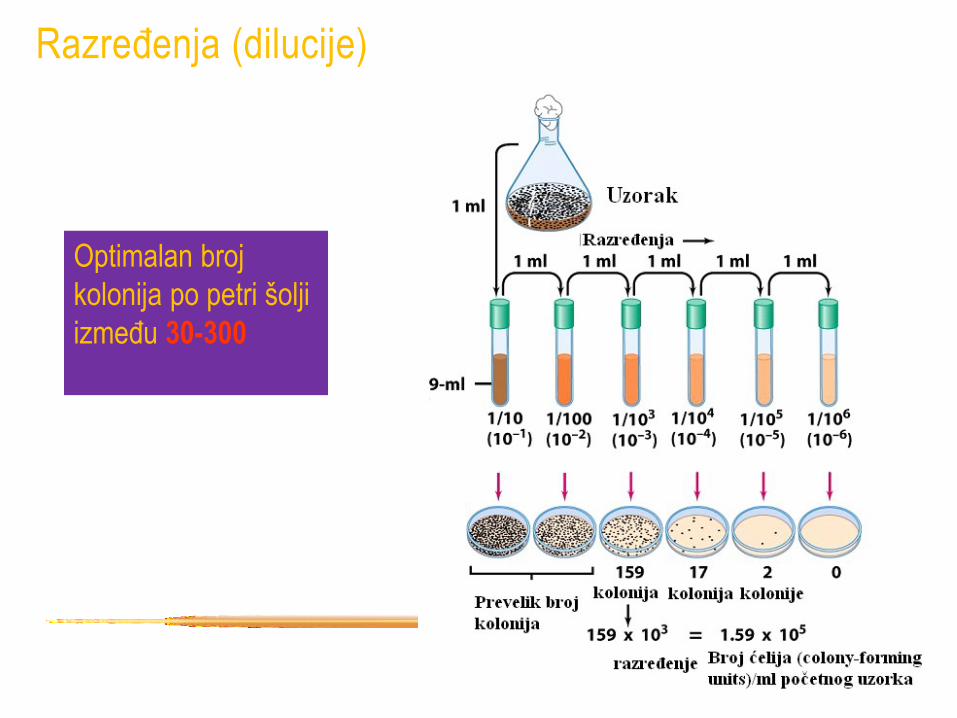
Razređenja (dilucije)
Optimalan broj
kolonija po petri šolji
između 30-300

• bez obzira na sve teškoće pri određivanju broja vijabilnih ćelija ova
procedura daje najbolju informaciju i široko se koristi
• Colony Forming Unit CFU/ml umesto ćelija/ml
• metod je veoma senzitivan pa se u uzorcima sa nekoliko ćelija može
odrediti broj (detekcija mikroorganizama koji kontaminiraju različite
produkte)
• na visoko selektivnim medijumima i specifičnim uslovima rasta mogu
se iz mešovite kulture izolovati (precizno prebrojati) određene vrste
• Velika anomalija -potcenjivanje broja ćelija u prirodnim uzorcima
(vrlo različite potrebe za resursima)
• totalni broj ćelija vs. ciljani broj ćelija
Indirektno - određivanje broja vijabilnih bakterija

Merenje ćelijske mase
• Masa ćelija proporcionalna broju, određivanje jednog
parametra koristi se za procenu drugog.
• Masa ćelija može biti određena koncentrovanjem (npr.
centrifugiranjem) poznatog volumena i merenjem dobijenog
taloga.
• Uobičajeno se određuje suva težina nakon sušenja taloga na
90-110C preko noći (10-20% od vlažne mase).

Indirektna metoda –merenje zamućenje
• metod za dobijanje podataka o broju i masi
ćelija je određivanje turbiditeta
• ćelije rasipaju svetlo koje prolazi kroz
suspenziju, više ćelija više svetlosti se
odbije, veći je turbiditet suspenzije
• spektrofotometar – aparat koji određuje
koja količina emitovane svetlosti je
propuštena kroz suspenziju
• izlazna vrednost – OD (optička gustina)
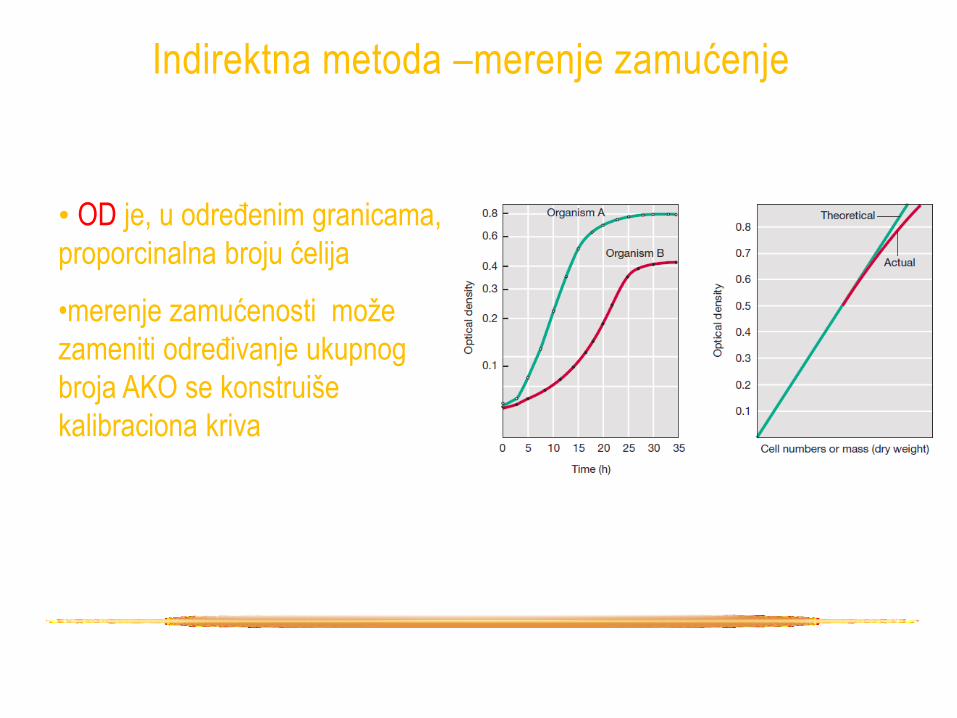
• OD je, u određenim granicama,
proporcinalna broju ćelija
•merenje zamućenosti može
zameniti određivanje ukupnog
broja AKO se konstruiše
kalibraciona kriva
Indirektna metoda –merenje zamućenje

Ključni faktori sredine koji utiču na rast svih
mikroorganizama
• temperatura
• pH
• dostupnost vode
• kiseonik
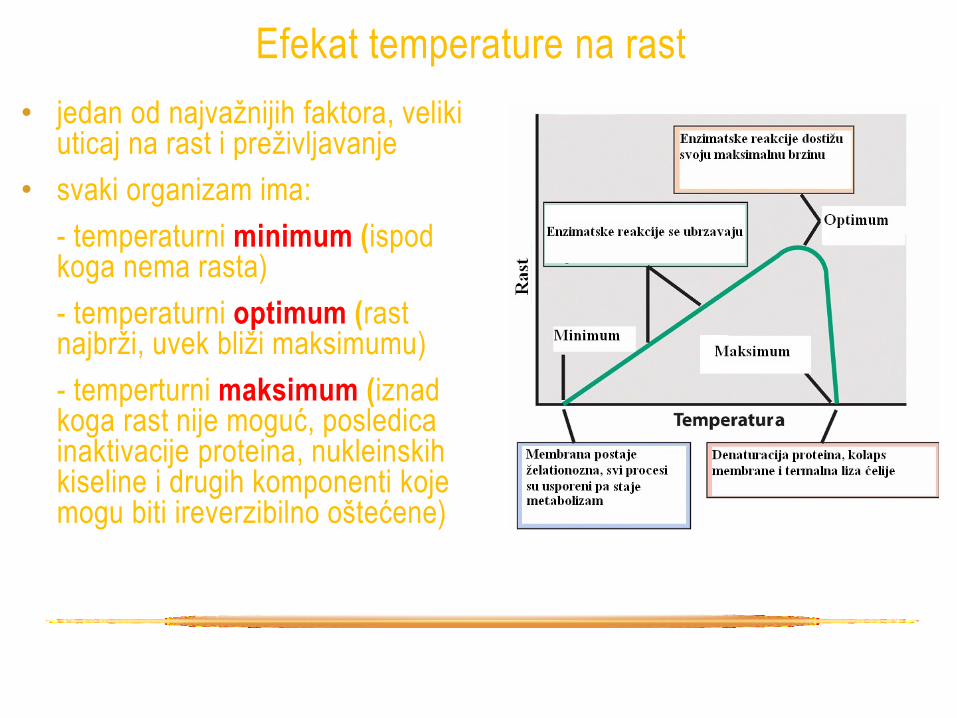
Efekat temperature na rast
• jedan od najvažnijih faktora, veliki uticaj na rast i preživljavanje
• svaki organizam ima:
- temperaturni minimum (ispod koga nema rasta)
- temperaturni optimum (rast najbrži, uvek bliži maksimumu)
- temperturni maksimum (iznad koga rast nije moguć, posledica inaktivacije proteina, nukleinskih kiseline i drugih komponenti koje mogu biti ireverzibilno oštećene)
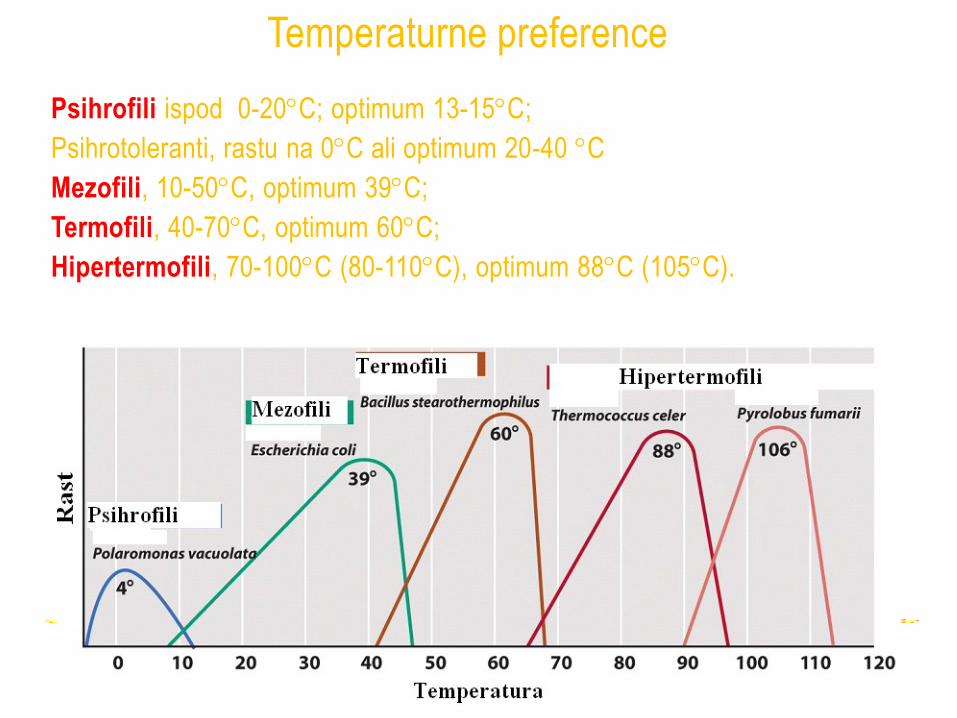
Psihrofili ispod 0-20C; optimum 13-15C;
Psihrotoleranti, rastu na 0C ali optimum 20-40 C
Mezofili, 10-50C, optimum 39C;
Termofili, 40-70C, optimum 60C;
Hipertermofili, 70-100C (80-110C), optimum 88C (105C).
Temperaturne preference

Psihrofili
• ispod 0-20C, optimum 13-15C
• okean, glečeri, hladna jezera, permafrost
• uslov – tečna voda
• Polaromonas 4-12C
• Psychromonas -12C
• minimum -20C?
• Psihrotolerantni mikroorganizmi - rasprostranjeniji od psihrofila (u umerenom pojasu, hrana u frižideru (4C).
Chlamydomonas nivalis
Polaromonas
zelene alge i dijatomeje
jezero u Suvoj dolini na
Antarktiku

Adaptacije psihrofila
• Enzimi funkcionalni na niskim temperaturama
- više α- heliksa = fleksibilnost (- ploče formiraju rigidnije strukture)
- više polarnih i manje hidrofobnih AK = fleksibilnost
- manje slabih veza i manje interakcija između domena
• Citoplazmatična membrana - semifluidna na niskim temperaturama
- visok % nezasićenih masnih kiselina (polinezasićenih) kraćeg lanca
• „cold-shock“ proteini i krioprotektanti
• Psihrotolerantni mikroorganizmi - rasprostranjeniji od psihrofila (u umerenom pojasu, hrana u frižideru (4C)

Termofili i hipertermofili
• termofili (optimum ˃45C) i hipertermofili (optimum ˃80C)
• zemljišta izložena neprekidno sunčevoj svetlosti zagreju se u toku dana do 50C - 70C
• temperatura fermentirajućeg materijala 60-65C
• topli izvori - 150-500C
- najveća koncentracija toplih izvora na jednom mestu - Yellowstone sa izvorima 92-93C ili 99-100C
• hidrotermalni izvori na dnu okeana 350C i više
• iznad 65C žive samo prokarioti a iznad 95ºC samo Archaea
• Methanopyrus na 122ºC
• nefototrofni organizmi mogu da rastu na višim temperaturama od fototrofnih
Hipertermofili,
(Yellowstone 97-99C)
Termofilnih cijanobakterija
(Yellowstone 70-74C)
Hidrotermalni ventili

Adaptacije termofila
• Enzimi
- zamena jedne ili nekoliko amino kiselina
- povećan broj jonskih veza između pozitivnih i negativnih naelektrisanja različitih AK
- gusto spakovana hidrofobna unutrašnjost proteina
• Citoplazmatična membrana
- bogata zasićenim masnim kiselinama (mnogo jače hidrofobne veze), membrana stabilna na visokim temperaturama.
• Hipertermofilne Archaea
- ne sadrže uopšte MK
- fitanil povezan etarskom vezom sa glicerofosfatom
- membrana u monosloju
• Stabilnost DNK („stabilizujuća jedinjenja“, reverzna DNK giraza, histoni)

• Enzimi termofila i hipertermofila katalizuju reakcije na visokim
temperaturama.
• DNK polimeraza Thermus aquaticus (Taq) koristi se za PCR,
amplifikaciju sekvenca DNK
• industrijski procesi
Značaj termofila i hipertermofila

Temperaturni maksimumi

• Neutrofili - optimalni pH 6-8, (najčešća staništa u prirodi pH 5-9).
• Acidofili – ispod pH 5.5 (gljive i obligatni acidofili); ispod pH 3 (Acidithiobacillus): ispod pH 2
- stabilnost citoplazmatične membrane
• Alkalifili - optimalni rast pH 10-11 (visokobazne sredine, slana jezera i zemljišta bogata karbonatima)
- aerobne bakterije slatkih voda, Bacillus firmus, Archaea
- enzimi u deterdžentima
• Intracelularni pH može varirati samo za 1- 1.5 pH jedinica od neutralne.
• Izuzeci: Picrophilus oshimae (pH 4.6) i ekstremni alkalifili (pH 9.5)
Efekat pH na rast

Puferi
• U toku rasta mikroorganizama u medijumu dolazi do promene
vrednosti pH usled metaboličkih reakcija
• Puferi se dodaju radi održanja konstantne pH

• Većina mikroorganizama ne može da preživi uslove sa niskom vodenom aktivnošću, dehidriraju, prelaze u dormantno stanje ili umiru (plazmoliza), sredina analogu suvom staništu.
• Halofili
– morski organizmi 1-4 % NaCl
- organizmi u hiperslanim sredinama 3-12 % NaCl
• Ekstremni halofili, 15-30 % NaCl za optimalni rast
• Halotoleranti tolerišu donekle redukciju dostupnosti vode
• Osmofili – staništa sa visokom koncentracijom šećera
• Kserofili – zaista suva staništa
Efekat osmotskog pritiska na rast

Efekat osmotskog pritiska na rast
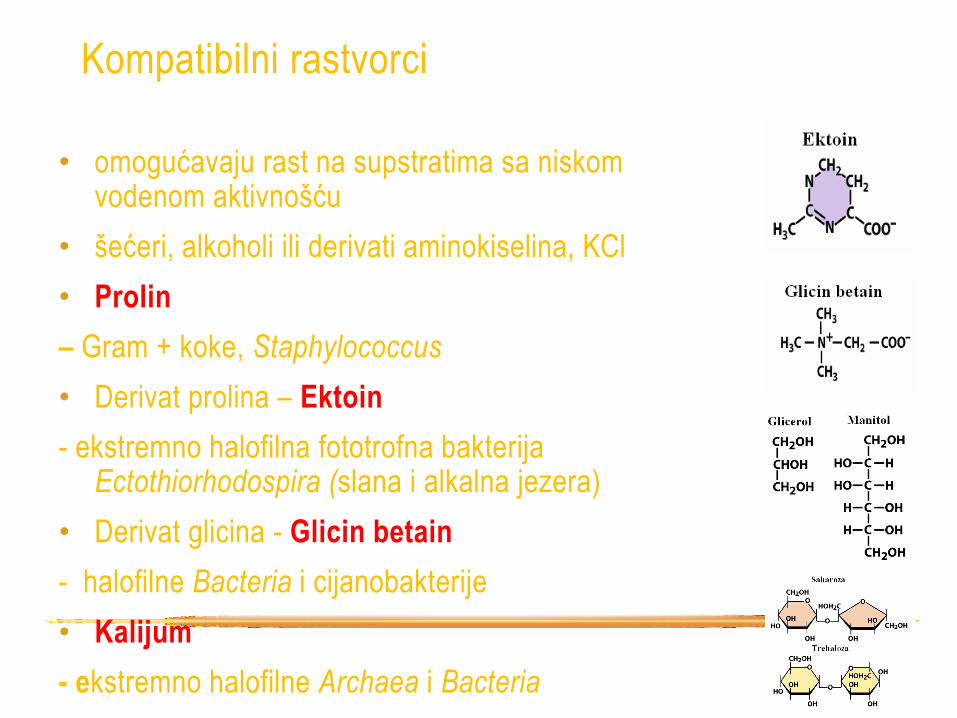
Kompatibilni rastvorci
• omogućavaju rast na supstratima sa niskom vodenom aktivnošću
• šećeri, alkoholi ili derivati aminokiselina, KCl
• Prolin
– Gram + koke, Staphylococcus
• Derivat prolina – Ektoin
- ekstremno halofilna fototrofna bakterija Ectothiorhodospira (slana i alkalna jezera)
• Derivat glicina - Glicin betain
- halofilne Bacteria i cijanobakterije
• Kalijum
- ekstremno halofilne Archaea i Bacteria

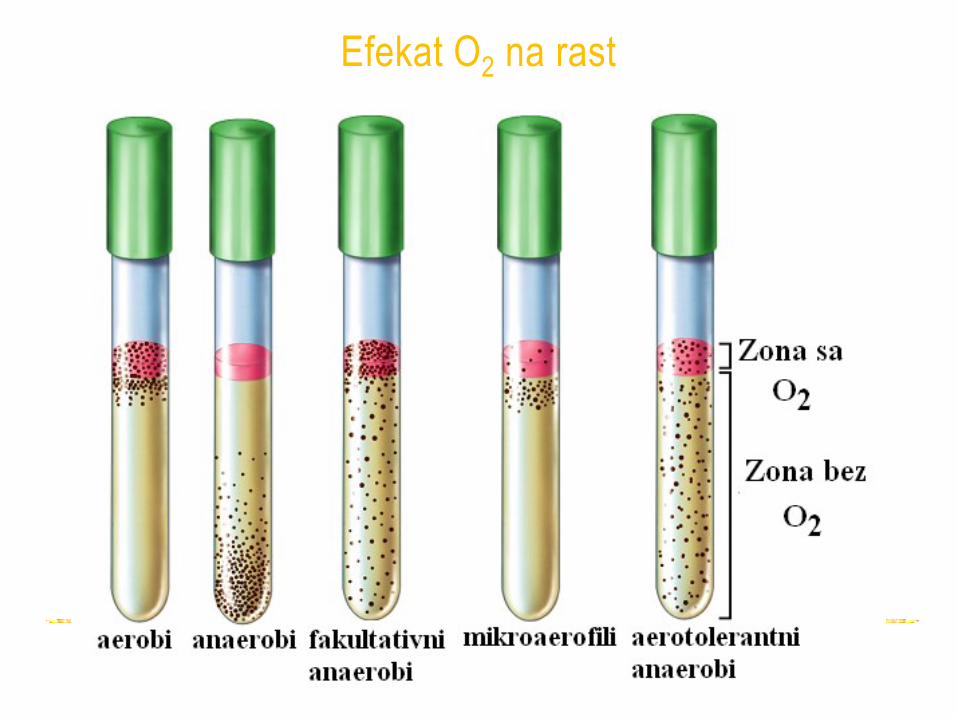
Efekat O2 na rast
• Aerobi - 21% O2, mogu tolerisati i povećanu koncentraciju (hiperbarični kiseonik).
• Fakultativni anaerobi - u aerobnim i anaerobnim uslovima
• Mikroaerofili - O2 u koncentraciji manjoj od 21%, (limitirani kapacitet za respiraciju ili enzimi osetljivi na O2)
• Anaerobi - ne koriste O2 kao terminalni akceptor elektrona
- aerotolerantni anaerobi - rastu u prisustvu O2
- obligatni (striktni) anaerobi - O2 je toksičan, nemaju enzime za uklanjanje toksičnih produkata kiseonika
- bakterije, arhee i nekoliko vrsta gljiva i protozoa

Grupa O2 Metabolizam Vrsta Stanište
Aerobi
Obligatni
Fakultativni
Mikroaerofili
neophodan
bolji rast
< 21%
Aerobna respiracija
Aerobna i anaerobna
respiracija, fermentacija
Aerobna respiracija
Micrococcus
E. coli
Spirillum
koža,
debelo crevo
jezera
Anaerobi
Aerotolerantni
Obligatni
bez efekta
štetan ili
letalan
Fermentacija
Fermentacija ili anaerobna
respiracija
Streptococcus
Methano-
bacterium
respiratorni
trakt
sedimenti
Efekat O2 na rast
Efekat O2 na rast

• enzimi za uklanjanje toksičnih
produkata aerobne respiracije
• Katalaza - H2O2
• Peroksidaza - H2O2 (zahteva
reduktanta NADH)
• Superoksid dismutaza - od 2
molekula superoksid O2- formira se
H2O2 i jedan molekul kiseonika
• superoksid dizmutaza i katalaza
deluju zajedno
• Superoksid reduktaza – ne nastaje
O2 kao nusprodukt
- Pyrococcus furiosus- obligatni anaerob
Efekat O2 na rast

Anaerobni mikroorganizmi
• obligatni anaerobni: prokarioti, neke gljive i protozoe
• anaerobne bakterije Clostridium, grupa Gram + štapićastih bakterija koje formiraju endospore
• nalaze se u zemljištu, sedimentima jezera, intestinalnom traktu, odgovorne za kvarenje konzervirane hrane
• metanogeni (Archaea), sulfat-redukujuće i homoacetogene bakterije, bakterije rumena, creva


















