
Food Restriction Prevents Advanced Glycation End...
10
Food Restriction Prevents Advanced Glycation End Product Accumulation and Retards Kidney Aging in Lean Rats LAURENT TEILLET,* ² PHILIPPE VERBEKE, ‡ SABINE GOURAUD,* HILAIRE BAKALA, ‡ CAROLINE BOROT-LALOI, ‡ DIDIER HEUDES, § PATRICK BRUNEVAL, § and BRUNO CORMAN* *Service de Biologie Cellulaire, Commissariat a ` l’Energie Atomique/Saclay, Gif-sur-Yvette, France; ² AP-HP, Ho ˆpital Sainte-Pe ´rine, Paris, France; ‡ Universite ´ Paris VII, Laboratoire de Biologie et Biochimie Cellulaire du Vieillissement, Paris, France; and § Institut National de la Sante ´ et de la Recherche Me ´dicale U430, Ho ˆpital Broussais, Paris, France. Abstract. Tissue content of advanced glycation end products (AGE) increases with age and contributes to the changes in structure and function of the renal and cardiovascular systems. The effect of chronic food restriction on this AGE accumulation was investigated in lean WAG/Rij rats. A 30% food restriction performed from 10 to 30 mo in female rats reduced their mean body weight from 240 6 7 to 160 6 12 g, but did not modify their survival. AGE collagen content increased from 14.3 6 5.5 to 104.7 6 13.0 arbitrary units per microgram (AU/mg) of hy- droxyproline (OHPro) in kidney between 10 and 30 mo, and from 9.7 6 1.2 to 310.6 6 34.6 AU/mg OHPro in the abdominal aorta. Food restriction reduced AGE accumulation to 21.4 6 3.3 and 74.6 6 16.5 AU/mg OHPro in kidney and aorta of 30-mo-old animals. Similar results were found for collagen prepared from isolated glomeruli (7.8 6 1.2, 81.2 6 16.1, and 10.3 6 4.3 AU/mg OHPro in 10-mo, 30-mo, and restricted 30-mo-old rats). Reduc- tion of intrarenal and arterial AGE accumulation by food restric- tion was confirmed by immunostaining in optical microscopy. Age-related changes in arterial and kidney structures as polyuria and proteinuria were mainly prevented by food restriction. These data indicate that chronic food restriction reduces the accumula- tion of AGE and preserves the structure and function of the renal and cardiovascular systems in lean rats, although it did not affect survival of the animals between 10 and 30 mo. Chronic food restriction usually slows the aging process and prevents age-related diseases reported in most species. In ro- dents, as a rule, it lowers body weight of the animals, increases mean and maximal survival, and postpones the morphologic and functional changes observed in the course of aging. Food restriction is efficient when started early in life or after the growing period, and its effects are more or less proportional to the extent of the restriction. Selective reduction of different diet components indicates that this beneficial effect is related to caloric restriction rather than to any constituent, although the underlying mechanisms are not firmly established. Several hypotheses have been proposed to explain the effects of food restriction, including prevention of overeating, reduction of oxidative stress, and decreased accumulation of advanced gly- cation end products (AGE). The influence of chronic food restriction on kidney structure and function has been documented in many studies. Caloric restriction prevents the glomerulosclerosis and proteinuria commonly reported in senescent rats. Protein restriction or substitution of animal for vegetable proteins in the diet also has a protective effect on age-related renal diseases in rodents. However, these studies have usually been performed in rats that gain weight throughout their life, and it was suggested that part of the beneficial effect of food restriction was due to the prevention of overeating frequently observed in rodents. Such a role for overeating in accelerated aging and diseases has been documented from comparison of survival in different strains of rodents. Sprague Dawley rats, which may reach 1000 to 1200 g at 24 mo, or Fisher 344 rats, which can grow up to 600 g when fed ad libitum, are short-living animals that develop severe renal diseases with age. Conversely, rats from the Brown- Norway, WAG/Rij, or Wistar/Lou strains, which remain lean, even when fed ad libitum, are long-living animals that are nearly free of glomerulosclerosis throughout their life (1– 6). Although food restriction may play a role in reducing over- eating-related diseases, it also slows the aging process of lean rats, indicating that the restriction affects some specific mo- lecular mechanisms (2,7,8). AGE accumulation is one of these mechanisms corresponding to the prolonged exposure of pro- teins to glucose due to changes in glucose tolerance and re- duced protein turnover with age (9 –11). The consequences of AGE accumulation on renal and cardiovascular systems have recently been documented by aminoguanidine administration in rodents. Treatment of Fischer 344 or Sprague Dawley rats from 6 to 24 mo reduced the AGE content of kidney, heart, and arteries (12). Concomitantly, age-related cardiac hypertrophy, vascular vasodilatory response, proteinuria, and glomerular sclerosis were all improved by aminoguanidine, suggesting a Received October 18, 1999. Accepted December 20, 1999. Correspondence to Dr. Bruno Corman, Commissariat a ` l’Energie Atomique (CEA)/Saclay, Service de Biologie Cellulaire, 91191 Gif-sur-Yvette, France. Phone: 133 1 69 08 63; 99; Fax: 133 1 69 08 35 70; E-mail: [email protected] 1046-6673/1108-1488 Journal of the American Society of Nephrology Copyright © 2000 by the American Society of Nephrology J Am Soc Nephrol 11: 1488 –1497, 2000
Transcript of Food Restriction Prevents Advanced Glycation End...

Food Restriction Prevents Advanced Glycation End ProductAccumulation and Retards Kidney Aging in Lean Rats
LAURENT TEILLET,* † PHILIPPE VERBEKE,‡ SABINE GOURAUD,*HILAIRE BAKALA, ‡ CAROLINE BOROT-LALOI,‡ DIDIER HEUDES,§
PATRICK BRUNEVAL,§ and BRUNO CORMAN**Service de Biologie Cellulaire, Commissariat a` l’Energie Atomique/Saclay, Gif-sur-Yvette, France;†AP-HP,Hopital Sainte-Pe´rine, Paris, France;‡UniversiteParis VII, Laboratoire de Biologie et Biochimie Cellulairedu Vieillissement, Paris, France; and§Institut National de la Sante´ et de la Recherche Me´dicale U430,Hopital Broussais, Paris, France.
Abstract. Tissue content of advanced glycation end products(AGE) increases with age and contributes to the changes instructure and function of the renal and cardiovascular systems.The effect of chronic food restriction on this AGE accumulationwas investigated in lean WAG/Rij rats. A 30% food restrictionperformed from 10 to 30 mo in female rats reduced their meanbody weight from 2406 7 to 1606 12 g, but did not modify theirsurvival. AGE collagen content increased from 14.36 5.5 to104.7 6 13.0 arbitrary units per microgram (AU/mg) of hy-droxyproline (OHPro) in kidney between 10 and 30 mo, and from9.76 1.2 to 310.66 34.6 AU/mg OHPro in the abdominal aorta.Food restriction reduced AGE accumulation to 21.46 3.3 and
74.6 6 16.5 AU/mg OHPro in kidney and aorta of 30-mo-oldanimals. Similar results were found for collagen prepared fromisolated glomeruli (7.86 1.2, 81.26 16.1, and 10.36 4.3 AU/mgOHPro in 10-mo, 30-mo, and restricted 30-mo-old rats). Reduc-tion of intrarenal and arterial AGE accumulation by food restric-tion was confirmed by immunostaining in optical microscopy.Age-related changes in arterial and kidney structures as polyuriaand proteinuria were mainly prevented by food restriction. Thesedata indicate that chronic food restriction reduces the accumula-tion of AGE and preserves the structure and function of the renaland cardiovascular systems in lean rats, although it did not affectsurvival of the animals between 10 and 30 mo.
Chronic food restriction usually slows the aging process andprevents age-related diseases reported in most species. In ro-dents, as a rule, it lowers body weight of the animals, increasesmean and maximal survival, and postpones the morphologicand functional changes observed in the course of aging. Foodrestriction is efficient when started early in life or after thegrowing period, and its effects are more or less proportional tothe extent of the restriction. Selective reduction of differentdiet components indicates that this beneficial effect is related tocaloric restriction rather than to any constituent, although theunderlying mechanisms are not firmly established. Severalhypotheses have been proposed to explain the effects of foodrestriction, including prevention of overeating, reduction ofoxidative stress, and decreased accumulation of advanced gly-cation end products (AGE).
The influence of chronic food restriction on kidney structureand function has been documented in many studies. Caloricrestriction prevents the glomerulosclerosis and proteinuriacommonly reported in senescent rats. Protein restriction orsubstitution of animal for vegetable proteins in the diet also has
a protective effect on age-related renal diseases in rodents.However, these studies have usually been performed in ratsthat gain weight throughout their life, and it was suggested thatpart of the beneficial effect of food restriction was due to theprevention of overeating frequently observed in rodents. Sucha role for overeating in accelerated aging and diseases has beendocumented from comparison of survival in different strains ofrodents. Sprague Dawley rats, which may reach 1000 to 1200 gat 24 mo, or Fisher 344 rats, which can grow up to 600 g whenfed ad libitum, are short-living animals that develop severerenal diseases with age. Conversely, rats from the Brown-Norway, WAG/Rij, or Wistar/Lou strains, which remain lean,even when fedad libitum, are long-living animals that arenearly free of glomerulosclerosis throughout their life (1–6).
Although food restriction may play a role in reducing over-eating-related diseases, it also slows the aging process of leanrats, indicating that the restriction affects some specific mo-lecular mechanisms (2,7,8). AGE accumulation is one of thesemechanisms corresponding to the prolonged exposure of pro-teins to glucose due to changes in glucose tolerance and re-duced protein turnover with age (9–11). The consequences ofAGE accumulation on renal and cardiovascular systems haverecently been documented by aminoguanidine administrationin rodents. Treatment of Fischer 344 or Sprague Dawley ratsfrom 6 to 24 mo reduced the AGE content of kidney, heart, andarteries (12). Concomitantly, age-related cardiac hypertrophy,vascular vasodilatory response, proteinuria, and glomerularsclerosis were all improved by aminoguanidine, suggesting a
Received October 18, 1999. Accepted December 20, 1999.Correspondence to Dr. Bruno Corman, Commissariat a` l’Energie Atomique(CEA)/Saclay, Service de Biologie Cellulaire, 91191 Gif-sur-Yvette, France.Phone:133 1 69 08 63; 99; Fax:133 1 69 08 35 70; E-mail: [email protected]
1046-6673/1108-1488Journal of the American Society of NephrologyCopyright © 2000 by the American Society of Nephrology
J Am Soc Nephrol 11: 1488–1497, 2000

causal link between AGE production and alterations in renaland cardiovascular systems. This hypothesis was strengthenedby the prevention of arterial stiffening and cardiac hypertrophyin aging rats treated with aminoguanidine in the last part of life,when AGE accumulation is maximal (13).
Several experiments have shown that food restriction bylowering plasma glucose may reduce the formation of inter-mediate glycation products and collagen fluorescence, which isconsidered an index of AGE content (10,14–19). This wouldserve to prevent the cross-linking of extracellular matrix andthe development of glomerulosclerosis. In the present study,we tested whether diet restriction also aids in the prevention ofAGE accumulation and kidney aging in lean rats, as observedin overeating animals. A 30% restriction was performed from10 to 30 mo in female WAG/Rij rats. AGE accumulation inextracellular matrix was assessed by competitive enzyme-linked immunosorbent assay (ELISA) with anti-AGE antibod-ies and visualized by immunolocalization in confocal andelectron microscopy. The structural consequences of food re-striction on kidney and arteries were determined by quantita-tive morphometry.
Materials and MethodsAnimals
Experiments were performed on female WAG/Rij rats that wereborn and raised in the animal care facilities of the Commissariat a`l’Energie Atomique, Centre d’Etudes de Saclay, Gif-sur-Yvette,France. Survival, growth rate, and tumor incidence of the inbredWAG/Rij strain have been published previously by Bureck (1). Meansurvival of female rats fedad libitum is close to 30 mo.
Rats were maintained on a 12/12 light-dark cycle at 50% humidityand a temperature of 21°C. From weaning to 3 mo, they were fed acommercial diet (DO3; UAR, Villemoisson, France) composed of 9%fish proteins, 16% vegetable proteins, and a total of 3500 kcal/kg. Atthe age of 3 mo, they were switched to a diet containing 2% fish and15% vegetable proteins, 0.71% phosphate, 0.78% calcium, 0.62%potassium, 0.27% sodium, 0.22% magnesium, and a total of 2900kcal/kg (DO4; UAR).
Restriction ProtocolMean food intake measured in female WAG/Rij rats fedad libitum
was 10.76 1.3 g (n 5 10) per day and was unchanged until 30 mo(10.36 1.1 g,n 5 10). A 40% food restriction, the usual protocol foroptimal survival in most experiments, was tested in 10-mo-old femalerats that were singly housed or grouped four per cage. In the twoconditions, body weight progressively decreased to 110 g, faster in thesingly housed than in the grouped rats, and the animals died within 7to 8 mo. In a second preliminary study, single or grouped rats werefood-restricted by 30%,i.e., 7 g/d per rat. The singly housed animalsreached a body weight equilibrium close to 130 g and survived. Thegrouped animals also lost weight, but to a lesser extent. Within a cage,individual body weights were very similar, indicating that each rat atea comparable amount of food. This latter protocol, a 30% restrictionin animals grouped four per cage, was followed from 10 to 30 mo, thetime at which rats fedad libitum or those that were food-restrictedwere sacrificed for morphologic and biochemical determinations.Animals (44 in control and 44 in restricted groups) were weighedweekly, and individual life spans were recorded to plot survival curve.
BP and Kidney FunctionSystemic BP and heart rate were measured by the tail-cuff method
in conscious 10-mo-old, 30-mo-old control, and 30-mo-old restrictedfemale rats. Urine volume was collected in metabolic cages for a 24-hperiod after a 2-d equilibration period. Urine osmolality was measuredwith a Roebling Automatik Osmometer (Berlin, Germany). Proteinconcentration in urine was assessed by the Bradford method withfraction V bovine serum albumin as a standard. Urine samples werefurther submitted to sodium dodecyl sulfate-polyacrylamide gel elec-trophoresis to determine the contribution of albumin to whole pro-teinuria.
Insulin Response to Glucose AdministrationInsulin secretion induced by a glucose load was determined in
10-mo-old, 30-mo-oldad libitum, and 30-mo-old restricted animals.The rats were fasted for 4 h, and a bolus of 0.54 g of glucose per 100 gbody wt was injected in the saphenous vein of anesthetized animals.Blood samples were sequentially collected on aprotinin at the caudalvein before glucose administration and at 5, 10, 15, 20, and 30 minafter glucose injection. Glycemia was immediately determined (One-Touch; Lifescan, Neckargemu¨nd, Germany), and the remaining sam-ples were stored at220°C. Plasma insulin was measured in thesesamples by RIA with the INSIK-5 kit (Sorin, Cedex, France).
Collagen ExtractionCollagen extracts were prepared from whole kidney, abdominal
aorta, and glomerular basement membranes. Rats were sacrificed bycervical dislocation. Lower abdominal aorta and kidneys were rapidlyremoved, frozen in liquid nitrogen, and stored at280°C. Aorta andkidneys were further homogenized with a Polytron (Ultra-Turrax;IKA Labortechnik) and suspended in phosphate-buffered saline, pH7.4. The resulting suspension was centrifuged at 40,0003 g for 30min at 4°C. Lipid extraction of the pellet was performed by additionof 5 ml chloroform/methanol (2:1 vol/vol) followed by gentle shaking,and left to stand overnight at 4°C. The upper layer was removed andthe pellet was washed three times with methanol and distilled water.Then the pellets were resuspended in 0.5 M acetic acid and 1 mg/mlpepsin, incubated for 18 h at 4°C to remove noncollagen proteins, andwashed three times with 0.1 M CaCl2, 0.02 M Tris-HCl (pH 7.5), and0.05% toluene. The pellets were digested with type VII collagenase(0.1 mg/ml; Sigma, St. Louis, MO) by gentle shaking at 37°C for 24 hand centrifuged at 40,0003 g for 30 min, 4°C. The resulting collagensupernatant was quantified from its hydroxyproline content accordingto Bergman and Loxley (20).
In a second series, the glomeruli were isolated from kidney cortexby the sieving method, as described by Krakower and Greenspon (21),and their basement membranes were prepared according to Meezanetal. (22). Collagen was extracted from the obtained glomerular base-ment membranes as described previously and quantified by hy-droxyproline determination (23).
Quantification of Renal and Arterial AGE Content byCompetitive ELISA
The used AGE polyclonal antibodies were raised against bovinepancreatic ribonuclease A (AGE-RNase) incubated for 60 d with 0.5M glucose in phosphate-buffered saline, pH 7.5, according to theprotocol of Makitaet al.(24). The specificity of the obtained antibodyhas been tested previously with soluble and structural glycoproteins,fibronectin, type IV collagen, laminin, and AGE-bovine serum albu-min (BSA) (23). ELISA was set up according to Papanastasiouet al.(25). The standard curve was established with AGE-BSA dilutions (1
J Am Soc Nephrol 11: 1488–1497, 2000 AGE Accumulation and Kidney Aging 1489

to 5000 ng) and AGE-RNase polyclonal antibody (1/500). The reac-tion was quantified by goat antibody directed against rabbit IgGcoupled to peroxidase (1/1000) (Sigma). The data were expressed asarbitrary units (AU)/mg hydroxyproline, with 1 AU corresponding tothe reactivity of 1 ng of AGE-BSA.
ImmunofluorescenceIntrarenal and vascular localization of AGE was performed by
immunocytochemistry, as described previously (23). Rats were anes-thetized with a 10 mg/100 g body wt intraperitoneal injection ofInactin (Byk-Gulden, Constance, Germany). A catheter, connected toa warmed tank (37°C) containing the Dubosq-Brazil solution (form-aldehyde, acetic acid, alcohol 80%, 4/1/10 vol/vol, and picric acid0.36%), was inserted in the abdominal aorta below the renal arteries.The vena cava was incised, and 50 to 100 ml of the fixative solutionwere perfused to the animal at a pressure of 120 mmHg. Kidney andaorta were embedded in paraffin and sliced in 5-mm-thick sections.These were incubated with anti-AGE antibodies (1/20) for 1 h andwith the fluorescence antibody FITC-labeled goat anti-rabbit IgG 1/20for an additional hour (Cappel, Cochranville, PA). The sections wereexamined with a confocal Argon-Krypton laser microscope LeicaTCS NT (Leica Microsystems, Rueil-Malmaison, France), using anexcitation wavelength at 488 nm, and an emission wavelength bandfrom 500 to 540 nm. The same intensity of laser energy and the samephotomultiplier setting was used to allow comparison between sectionstaining.
Electron Microscopy and AGE Localization byImmunogold Labeling
Small pieces of renal cortex were fixed by immersion in 4%paraformaldehyde/0.1 M sodium cacodylate buffer for 24 h, dehy-drated, and embedded in LR White. Ultrathin 600 Å sections wereprocessed for immunogold labeling with anti-AGE antibody (1/20)and protein A-gold complex (diameter, 15 nm) (1/80) (Biol Cell,Cardiff, United Kingdom) (23). The resulting preparations were ex-amined on a Zeiss 912 transmission electron microscope.
Renal and Arterial Quantitative MorphometryKidneys and abdominal aorta of 10-mo-old, 30-mo-oldad libitum,
and 30-mo-old restricted rats were fixed by perfusion as described forimmunofluorescence. The left kidney and the abdominal aorta wereexcised, stored for 24 h in the Dubosq-Brazil solution, and embeddedin paraffin. Sections of kidneys and aorta were stained by Marinozzisilver staining or elastic stain, respectively.
For each rat, quantitative morphometry analyses of the differentglomerular domains were performed as described previously (26). A5-mm-thick section was obtained from left kidney transversely cutthrough the hilus in each rat. On this section, 30 different superficialglomeruli were randomly sampled for morphometric analysis as fol-lows: The microscopic slide was scanned clockwise along the super-ficial cortex, and one glomerulus out of three was analyzed. Prelim-inary experiments indicated that convergence of the running mean andvariance of every tested parameter was reached for 30 glomerulimeasured in each animal. For every investigated glomerulus, thefollowing measurements were obtained with an automated imageanalysis system: (1) the total glomerular profile area limited by theinternal edge of Bowman’s capsule; (2) the glomerular tuft area; (3)the mesangial domain area defined as glomerular tuft area minus areasof the capillary lumens and the capillary-free walls; (4) the total areaof the capillary lumen profiles; and (5) the number of capillary lumenprofiles (26). Mean glomerular volume was calculated in each rat
from the glomerular section area according to Gil and Barba (27) andWeibel (28). In their model, the true mean glomerular diameterD 5(4/p) 3 d# , whered# is the mean glomerular diameter obtained fromglomerular profile area. Considering the glomerulus as a sphere, thecorresponding mean glomerular volume was calculated as:V 5 (4/3) 3 p 3 (D/2)3. Although the sphere hypothesis used for volumecalculation is not an unbiased method, a possible error would mostlikely affect all groups equally. The volumes of the different domainswere calculated from this mean glomerular volume and the areafraction of mesangial, capillary, and whole tuft sections. For abdom-inal aorta, circumference of the vessels, thickness of the arterial wall,and total area surface were determined on transverse sections (29).Total cross-sectional parietal area corresponded to the section area ofthe vessel minus the luminal area.
Renal HistologyParaffin sections of the kidney were stained with Masson’s
trichrome. The percentage of glomeruli with focal and segmentalsclerosis or total sclerosis (obsolescent glomeruli) was determined on100 glomeruli randomly sampled in a transverse renal section for eachrat.
Statistical AnalysesResults are presented as means6 SEM and were statistically
analyzed by ANOVA. Survival rates were compared with the log-ranktest. Significance was set atP , 0.05.
ResultsBody Weights and Survival
Body weights of female WAG/Rij rats fedad libitum andrestricted by 30% from 10 to 30 mo are shown in Figure 1.Mean values were 2136 3 g (n 5 44) for the 10-mo-old ratsand 2406 7 g (n 5 28) for the 30-mo-oldad libitum rats. Inrestricted female rats, body weight decreased during the first 3mo of treatment and reached a steady state close to the 160612 g value recorded at 30 mo (n 5 28).
At 30 mo, corresponding to the end of the experiment, 28adlibitum and 28 food-restricted animals were still alive. Whencompared with the initial cohorts of 44 animals in each group,
Figure 1. Body weight of female WAG/Rij rats that were fedadlibitum or were food-restricted by 30% from 10 to 30 mo. Results aregiven as mean6 SEM. The initial cohort included 44 animals in eachgroup.
1490 Journal of the American Society of Nephrology J Am Soc Nephrol 11: 1488–1497, 2000

this corresponded to 64% surviving in the two groups. The twosurvival curves were not statistically different.
Physiologic ParametersSystolic BP measured in conscious animals was comparable
in 10- and 30-mo-old rats (1286 3 mmHg,n 5 12 and 12964 mmHg, n 5 12, respectively), and was not significantlymodified by the restriction (1376 2 mmHg,n 5 12). Heartrate was similar in 10- and 30-mo-old animals fedad libitum(348 6 6 beats/min,n 5 12 and 3616 7 beats/min,n 5 12,respectively), but was significantly lowered in restricted 30-mo-old rats (2706 7 beats/min,n 5 12). Heart weight in-creased in proportion to body weight from 10 to 30 mo (0.6760.01 g,n 5 7 and 0.806 0.02 g,n 5 7, respectively) inadlibitum fed rats and was reduced in the restricted animals(0.546 0.02 g,n 5 7).
Plasma insulin concentration measured before glucose ad-
ministration was not statistically different in 10- and 30-mo-oldcontrol animals, but was significantly lowered in the 30-mo-old restricted rats (Figure 2). Injection of similar glucose loadwith respect to body weight increased plasma glucose concen-tration to the same extend in the three groups of rats (data notshown). This increase in plasma glucose concentration resultedin a transient release of insulin, the amplitude of which wassignificantly larger in thead libitum30-mo-old rats than in the10-mo-old animals (Figure 2). The glucose-induced insulinsecretion in 30-mo-old restricted animals was significantlylower than that obtained in the 10- and 30-mo-old rats fedadlibitum (Figure 2).
Values for daily water intake, urine volume, and osmolalityare shown in Table 1. Urine volume was larger in the 30-mo-old than in the 10-mo-old animals, and urine osmolality wasdecreased in proportion. Chronic food restriction preventedthis age-related change in water homeostasis. To check that themaintained urine osmolality in the restricted animals was notjust related to the reduced daily food intake, the effect of anacute 30% food restriction on urine volume and osmolality wasinvestigated in control 30-mo-old rats. In these additionalseries, urine volume and osmolality were 7.76 1.5 ml/24 hand 13616 210 mosmol/kg H2O (n 5 10) in 30-mo-oldadlibitum-fed animals and 4.96 0.7 ml/24 h and 13216 131mosmol/kg H2O (n 5 10) in the same animals acutely re-stricted by 30% for 1 wk.
Proteinuria was significantly increased from 10 to 30 mo inad libitum-fed rats (Table 1). Electrophoresis of the urineindicated that proteinuria was mainly due to an increase inalbumin excretion (data not shown). Food restriction preventedthe age-related increase in protein and albumin excretions,which were comparable in 10-mo-old and restricted 30-mo-oldrats (Table 1).
AGE AccumulationAGE content of collagen extracted from abdominal aorta
and whole kidney markedly increased from 10 to 30 mo in rats
Figure 2.Plasma insulin concentration after a 0.54 g/100 g body wtinjection of glucose in 10 and 30-mo-old female WAG/Rij rats fedadlibitum and 30-mo-old animals food-restricted by 30%. Results aregiven as mean6 SEM of six animals in each group.
Table 1. Kidney weight, water homeostasis, and proteinuriaa
Group Right Kidney(g)
Right Kidney(g/100 g body wt)
Left Kidney(g)
Left Kidney(g/100 g body wt)
10-mo-old 0.656 0.02 0.3186 0.004 0.646 0.02 0.3146 0.00530-mo-old 0.806 0.02b 0.3266 0.007 0.776 0.02b 0.3166 0.00930-mo-old restricted 0.556 0.01b,c 0.3686 0.013b,c 0.556 0.02b,c 0.3606 0.012b,c
Water Intake(ml/24 h)
Urinary Volume(ml/24 h)
Urine Osmolality(mosmol/kg H2O)
Proteinuria(mg/24 h)
10-mo-old 13.26 0.7 3.76 0.4 23896 112 3.56 0.330-mo-old 16.36 1.6 8.36 1.7b 13396 157b 13.86 2.4b
30-mo-old restricted 8.66 0.4b,c 3.06 0.4c 19606 154c 3.46 0.6c
a Kidney weight (n 5 7 in each group), daily water intake, urinary volume, urine osmolality, and proteinuria (n 5 10 in each group) in10- and 30-mo-old female WAG/Rij rats fedad libitumand 30-mo-old animals food-restricted by 30%. Results are given as mean6 SEM.
b P , 0.05, significantly different from 10-mo-old rats.c P , 0.05, significantly different from 30-mo-old rats fedad libitum.
J Am Soc Nephrol 11: 1488–1497, 2000 AGE Accumulation and Kidney Aging 1491

fed ad libitum (Figure 3, A and B). Chronic food restrictionmostly prevented this accumulation of AGE in both aorta andkidneys of 30-mo-old animals (Figure 3, A and B). Compara-ble determinations performed on collagen prepared from iso-lated glomerular basement membranes confirmed the rise ofAGE in ad libitum-fed animals and the beneficial effect of dietrestriction on intrarenal accumulation of AGE (Figure 3C).
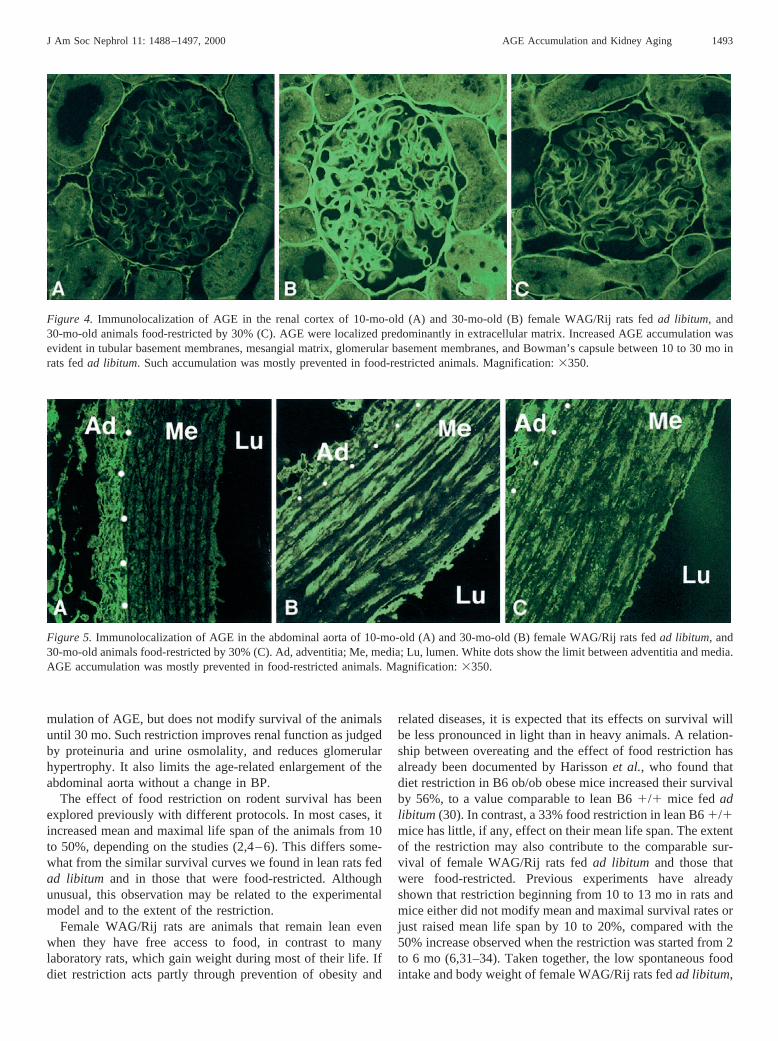
Immunolocalization of AGE in renal cortex by confocalmicroscopy showed a weak labeling in peritubular matrix andBowman’s capsule of 10-mo-old animals (Figure 4A). Thisimmunofluorescent staining of AGE was greatly increased inbasement membranes and Bowman’s capsule of 30-mo-old ratsfed ad libitum (Figure 4B). Accumulation of AGE was alsoevident on capillary walls and mesangium within the glomeruliof 30-mo-old rats. Food restriction markedly reduced the im-munofluorescent staining of AGE in the extracellular peritu-bular and glomerular matrix (Figure 4C). Immunolocalizationof AGE in abdominal aorta is shown in Figure 5. In 10-mo-oldanimals, AGE were localized in the extracellular matrix of theadventitia, whereas the media matrix was mostly negative. At30 mo, AGE accumulated in the matrix of both the adventitiaand the media. Food restriction reduced this accumulation(Figure 5).
Ultrastructural localization of AGE in the renal cortex wasperformed by immunogold staining. As shown in Figure 6,specific immunogold labeling of AGE was evident on tubularbasement membranes in 10- and 30-mo-old animals fedadlibitum, as well as in animals on a restricted diet. Within theglomerulus, AGE accumulation was localized mainly in theglomerular basement membrane in every group (data notshown).
MorphometryKidney weight increased by 20% between 10 and 30 mo in
ad libitum-fed rats, as did body weight (Table 1). In 30-mo-oldrestricted animals, kidney weight and body weight were re-duced, respectively, by 15 and 25% compared with 10-mo-oldanimals, and by 30 and 38% compared with 30-mo-old rats fedad libitum. The kidney to body weight ratio was significantlygreater in 30-mo-old restricted animals than in adult and se-nescent rats fedad libitum (Table 1). Examination of kidneysections failed to detect interstitial fibrosis, inflammation, ormarked vascular alterations whatever the age of the animals orthe diet. The proportion of glomeruli with evidence of segmen-tal or total sclerosis did not exceed 2% of the nephrons in10-mo-old rats. This proportion was not significantly modifiedin senescent rats fedad libitumor in those that were restricted.The lack of glomerulosclerosis in senescent WAG/Rij rats isconsistent with the previous observations performed in thisstrain (1,26).
Quantitative morphometry performed by automated imageanalysis showed an age-related increase in the volume of theglomeruli and of the glomerular tuft, a mesangial expansion,and a greater capillary surface in senescent than in adult rats(Table 2). The increase in capillary volume was proportional tothe glomerular enlargement, whereas the mesangial volumeincreased by 86% when glomerular volume was raised by 19%.
Chronic food restriction prevented glomerular hypertrophy andthe increase in capillary volume, but did not affect mesangialexpansion (Table 2). The abdominal aorta was enlarged from10 to 30 mo, as was the wall thickness and total parietal surface(Table 2). Food restriction partly prevented the increase in sizeand parietal surface of abdominal aorta (Table 2).
DiscussionThis study shows that a 30% food restriction initiated at 10
mo in lean female rats prevents intrarenal and vascular accu-
Figure 3.Advanced glycation end product (AGE) content in collagenextracted from abdominal aorta (A), kidneys (B), and glomeruli (C) in10- and 30-mo-old female WAG/Rij rats fedad libitum and 30-mo-old animals food-restricted by 30%. Results are given as mean6SEM of six animals in each group.
1492 Journal of the American Society of Nephrology J Am Soc Nephrol 11: 1488–1497, 2000
mulation of AGE, but does not modify survival of the animalsuntil 30 mo. Such restriction improves renal function as judgedby proteinuria and urine osmolality, and reduces glomerularhypertrophy. It also limits the age-related enlargement of theabdominal aorta without a change in BP.
The effect of food restriction on rodent survival has beenexplored previously with different protocols. In most cases, itincreased mean and maximal life span of the animals from 10to 50%, depending on the studies (2,4–6). This differs some-what from the similar survival curves we found in lean rats fedad libitum and in those that were food-restricted. Althoughunusual, this observation may be related to the experimentalmodel and to the extent of the restriction.
Female WAG/Rij rats are animals that remain lean evenwhen they have free access to food, in contrast to manylaboratory rats, which gain weight during most of their life. Ifdiet restriction acts partly through prevention of obesity and
related diseases, it is expected that its effects on survival willbe less pronounced in light than in heavy animals. A relation-ship between overeating and the effect of food restriction hasalready been documented by Harissonet al., who found thatdiet restriction in B6 ob/ob obese mice increased their survivalby 56%, to a value comparable to lean B61/1 mice fedadlibitum (30). In contrast, a 33% food restriction in lean B61/1mice has little, if any, effect on their mean life span. The extentof the restriction may also contribute to the comparable sur-vival of female WAG/Rij rats fedad libitum and those thatwere food-restricted. Previous experiments have alreadyshown that restriction beginning from 10 to 13 mo in rats andmice either did not modify mean and maximal survival rates orjust raised mean life span by 10 to 20%, compared with the50% increase observed when the restriction was started from 2to 6 mo (6,31–34). Taken together, the low spontaneous foodintake and body weight of female WAG/Rij rats fedad libitum,
Figure 4. Immunolocalization of AGE in the renal cortex of 10-mo-old (A) and 30-mo-old (B) female WAG/Rij rats fedad libitum, and30-mo-old animals food-restricted by 30% (C). AGE were localized predominantly in extracellular matrix. Increased AGE accumulation wasevident in tubular basement membranes, mesangial matrix, glomerular basement membranes, and Bowman’s capsule between 10 to 30 mo inrats fedad libitum. Such accumulation was mostly prevented in food-restricted animals. Magnification:3350.
Figure 5. Immunolocalization of AGE in the abdominal aorta of 10-mo-old (A) and 30-mo-old (B) female WAG/Rij rats fedad libitum, and30-mo-old animals food-restricted by 30% (C). Ad, adventitia; Me, media; Lu, lumen. White dots show the limit between adventitia and media.AGE accumulation was mostly prevented in food-restricted animals. Magnification:3350.
J Am Soc Nephrol 11: 1488–1497, 2000 AGE Accumulation and Kidney Aging 1493

the present moderate 30% food restriction, and the extent of thetreatment from 10 to 30 mo may be responsible for the similarsurvival curves reported forad libitum-fed and restricted ani-mals. It is not excluded, however, that an effect on maximallife span should have been evident if the experiment wasprolonged over 30 mo.
Although survival is comparable inad libitumand restrictedlean female WAG/Rij rats, the present data indicate that foodrestriction prevents the age-related accumulation of AGE incollagen extracted from the abdominal aorta and kidney, as incollagen isolated from glomerular basement membrane. Extra-cellular localization of AGE accumulation was confirmed byimmunofluorescence in confocal microscopy and by immuno-gold labeling in electron microscopy. This prevention of AGEaccumulation is in good agreement with the reduced pentosi-dine or furosine formation—two Amadori-derived products—and the lower fluorescence of skin, aortic, or tail collagenreported in food-restricted rats or mice (10,14,16–18). Differ-ent mechanisms have been proposed to explain the age-relatedaccumulation of AGE. A reduced glucose tolerance and aninsulin resistance, would tend to increase plasma glucose con-centration during the different feeding and fasting periods ofthe day (9,35). The resulting increase in glucose concentrationover time would favor the nonenzymatic reactions betweenproteins and sugars, and enhance production of AGE (9,11).According to this hypothesis, any sustained decrease in plasmaglucose concentration would reduce AGE formation. A lower-ing effect of food restriction on plasma glucose has beendocumented by Masoroet al., who measured the daily patternof plasma glucose and insulin concentrations in rats fedadlibitum and in those that were food-restricted (15). A reducedinsulin plasma concentration in baseline conditions or in re-sponse to a glucose load in restricted animals has also beenreported by several authors, as in this study, confirming theeffect of diet restriction on glucose homeostasis (17,19,36).
It has been hypothesized that AGE accumulation would
contribute to the age-related changes in renal and cardiovas-cular systems, the extent of which depends on the experimentalmodel (9–11). Conversely, prevention of AGE accumulationwould lower these alterations. This has been recently demon-strated in aging rats using aminoguanidine, a drug that inhibitsAGE formation without altering plasma glucose concentration(9,11–13). When administered from 6 to 24 mo or from 24 to30 mo, aminoguanidine did not modify survival of SpragueDawley, Fisher 344, or WAG/Rij rats, but prevented hearthypertrophy and arterial stiffening. In rats that exhibited glo-merulosclerosis, it also improved the development of chronicprogressive nephrosis.
The present food restriction in lean rats prevented AGEaccumulation, glomerular hypertrophy, proteinuria, and thereduced urine osmolality commonly reported in aging rats. Theenlargement in glomerular volume from 10 to 30 mo is con-sistent with the determinations already performed in femaleWAG/Rij rats by automated image analysis or in dissectednephrons (37,38). The effect of food restriction on glomerularmorphology may be partly due to a reduction in body weight ofthe animals, and partly to prevention of the AGE cross-linkingof extracellular matrix components and of the AGE-stimulatedcollagen synthesis by mesangial and interstitial cells (9,39,40).These mechanisms could also apply to the age-related enlarge-ment and thickening of the aorta and its prevention by foodrestriction. In the absence of any change in BP, a lower AGEaccumulation in the arterial media of food-restricted rats maycontribute to a decrease in age-related arterial stiffness andcardiac hypertrophy (11,13,26,41).
Protein excretion was increased with age. This was mostlydue to albuminuria and usually reflects a change in glomerularhemodynamics or sieving (42). The prevention of albuminuriaby diet restriction suggests that either filtration rate and pres-sure are modified by food intake, or that the molecular struc-ture of the glomerular basement membrane is preserved in30-mo-old food-restricted animals (43,44). In addition, AGE
Table 2. Glomerular and arterial morphometrya
Variable and Group Glomerular Volume(mm3 3106)
Glomerular Tuft(mm3 3106)
Mesangial Volume(mm3 3106)
Capillary Volume(mm3 3106)
Glomerulus10-mo-old 1.496 0.05 0.966 0.03 0.146 0.01 0.626 0.0230-mo-old 1.776 0.07b 1.266 0.05b 0.266 0.02b 0.756 0.03b
30-mo-old restricted 1.496 0.05c 1.016 0.03c 0.236 0.03b 0.586 0.02c
Luminal Circumference (mm) Wall Thickness (mm) Parietal Surface (mm2)
Abdominal aorta10-mo-old 4.556 0.05 0.0666 0.003 0.346 0.0230-mo-old 5.466 0.08b 0.0916 0.003b 0.556 0.02b
30-mo-old restricted 5.046 0.05b,c 0.0826 0.002b,c 0.476 0.01b,c
a Quantitative morphometry of superficial glomeruli and abdominal aorta of 10- and 30-mo-old female WAG/Rij rats fedad libitum and30-mo-old animals food-restricted by 30%. Measurements were performed in 30 superficial glomeruli randomly sampled in transversalsections for each rat. Results are given as means6 SEM of 10 animals per group.
b P , 0.05, significantly different from 10-mo-old rats.c P , 0.05, significantly different from 30-mo-old rats fedad libitum.
1494 Journal of the American Society of Nephrology J Am Soc Nephrol 11: 1488–1497, 2000

may have a direct effect on protein excretion by the kidney(40). Polyuria and decreased urine osmolality, frequently re-ported in aging rats, were prevented by food restriction. Thiswas not due to reduced daily food intake, because a 30% food
restriction for 1 wk in senescent animals did not alter theirurine osmolality. The maintained kidney concentrating abilityin restricted animals may be related either to an increase invasopressin secretion in 30-mo-old treated rats or to prevention
Figure 6.Ultrastructural localization of AGE by immunogold labeling in the tubular basement membrane of 10-mo-old (A) and 30-mo-old (B)female WAG/Rij rats fedad libitumand 30-mo-old animals food-restricted by 30% (C). Negative control in the absence of AGE antibody (D).TBM, tubular basement membrane; Epi, epithelial cells. Magnification:310,000.
J Am Soc Nephrol 11: 1488–1497, 2000 AGE Accumulation and Kidney Aging 1495

of the vasopressin resistance reported in the aging kidney (45).Vasopressin measurements and determination of V2 receptorand aquaporin expression would be necessary to establish themechanisms by which chronic diet restriction maintains waterhomeostasis.
In conclusion, the present data indicate that chronic foodrestriction in lean rats prevents the accumulation of AGE andinhibits structural and functional alterations of the renal andcardiovascular systems, yet does not affect survival of theanimals between 10 and 30 mo.
AcknowledgmentsThis work was funded by the Institut de Recherche International
Servier (IRIS). Laurent Teillet was supported by a grant from theAssistance Publique – Hoˆpitaux de Paris, Direction de la RechercheClinique (Paris, France). We thank Martine Douheret for technicalassistance in quantitative morphometry. Electronic micrography wasperformed at the CIME-Jussieu, 7 quai Saint-Bernard, 75005 Paris,France. We are grateful to Patrick Herry and Jean-Charles Robillardfor their outstanding work with animal care.
References1. Bureck JD:Pathology of Aging Rats, Boca Raton, FL, CRC
Press, 19782. Fernandes G, Venkatraman J, Turturro A, Attwood V, Hart R:
Effect of food restriction on life span and immune functions inlong-lived Fischer-3443 Brown Norway F1 rats.J Clin Immu-nol 17: 85–95, 1997
3. Keenan K, Smith P, Hertzog P, Soper K, Ballam G, Clark R: Theeffects of overeating and dietary restriction on Sprague-Dawleyrat survival and early pathology biomarkers of aging.ToxicolPathol 22: 300–315, 1994
4. McCay C, Crowell M, Maynard L: The effect of retarded growthupon the length of life span and upon the ultimate body size.JNutr 10: 63–79, 1935
5. Wyndham J, Everitt A, Everitt S: Effects of isolation and foodrestriction begun at 50 days on the development of age-associ-ated renal disease in the male Wistar rat.Arch Gerontol Geriatr2: 317–332, 1983
6. Yu B, Masoro E, Murata I, Bertrand H, Lynd F: Life span studyof SPF Fischer 344 male rats fed ad libitum or restricted diets:Longevity, growth, lean body mass and disease.J Gerontol37:130–141, 1982
7. Randerath K, Zhou G, Hart R, Turturro A, Randerath E: Biomar-kers of aging: Correlation of DNA I-compound levels withmedian lifespan of calorically restricted andad libitum fed ratsand mice.Mutat Res295: 247–263, 1993
8. Sonntag W, Xu X, Ingram R, D’Costa A: Moderate caloricrestriction alters the subcellular distribution of somatostatinmRNA and increases growth hormone pulse amplitude in agedanimals.Neuroendocrinol61: 601–608, 1995
9. Cerami A: Glucose as a mediator of aging.J Am Geriatr Soc33:626–634, 1985
10. Sell D, Lane M, Johnson W, Masoro E, Mock O, Reisier K,Fogarty J, Cutler R, Ingram D, Roth G, Monnier V: Longevityand the genetic determination of collagen glycoxidation kineticsin mammalian senescence.Proc Natl Acad Sci USA93: 485–490, 1996
11. Vlassara H, Bucala R, Striker L: Pathogenic effects of advanced
glycosylation: Biochemical, biologic, and clinical implicationsfor diabetes and aging.Lab Invest70: 138–151, 1994
12. Li YM, Steffes M, Donnelly T, Liu C, Fuh H, Basgen J, BucalaR, Vlassara H: Prevention of cardiovascular and renal pathologyof aging by the advanced glycation inhibitor aminoguanidine.Proc Natl Acad Sci USA93: 3902–3907, 1996
13. Corman B, Duriez M, Poitevin P, Heudes D, Bruneval P, TedguiA, Levy BI: Aminoguanidine prevents age-related arterial stiff-ening and cardiac hypertrophy.Proc Natl Acad Sci USA95:1301–1306, 1998
14. Cefalu W, Bell-Farrow A, Wang Z, Sonntag W, Flu M, BaynesJ, Thorpe S: Caloric restriction decreases age-dependent accu-mulation of the glycoxidation products, N-(carboxymethyl) ly-sine and pentosidine, in rat skin collagen.J Gerontol Biol Sci50:B337–B341, 1995
15. Masoro E, McCarter R, Katz M, McMahan A: Dietary restrictionalters characteristics of glucose fuel use.J Gerontol47: B202–B208, 1992
16. Miksik I, Struzinsky R, Deyl Z: Change with age of UV absor-bance and fluorescence of collagen and accumulation of epsilon-hexosyllysine in collagen from Wistar rats living on differentfood restriction regimens.Mech Ageing Dev57: 163–174, 1991
17. Novelli M, Masiello P, Bombara M, Bergamini E: Protein gly-cation in the aging male Sprague Dawley rat: Effects of antiagingdiet restriction.J Gerontol53: 94–101, 1998
18. Reiser K: Influence of age and long-term dietary restriction onenzymatically mediated crosslinks and nonenzymatic glycationof collagen in mice.J Gerontol49: 71–79, 1994
19. Wang Z, Bell-Farrow A, Sonntag W, Cefalu W: Effect of age andcaloric restriction on insulin receptor binding and glucose trans-porter levels in aging rats.Exp Gerontol32: 671–684, 1997
20. Bergman J, Loxley R: The determination of hydroxyproline inurine hydrolysates.Clin Chim Acta27: 347–349, 1970
21. Krakower C, Greenspon S: The isolation of basement mem-branes. In:Biology and Chemistry of Basement Membranes,edited by Kefalides NA, New York, Academic, 1978, pp 1–16
22. Meezan E, Hjell J, Brendel K, Carlson E: A simple, versatile,non-disruptive method for the isolation of morphologically andchemically pure basement membranes for several tissues.Life Sci17: 1721–1732, 1976
23. Verbeke P, Perichon M, Borot-Laloi C, Shaeverbeke J, BakalaH: Accumulation of advanced glycation end products in the ratnephron: Link with circulating AGEs during aging.J HistochemCytochem45: 1059–1068, 1997
24. Makita Z, Vlassara H, Cerami A, Bucala R: Immunochemicaldetection of advanced glycosylation end products in vivo.J BiolChem267: 5133–5138, 1992
25. Papanastasiou P, Grass L, Rodela H, Patikarea A, Oreopoulos D,Diamandis E: Immunological quantification of advanced glyco-sylation end products in the serum of patients on hemodialysis orCAPD. Kidney Int46: 216–222, 1994
26. Heudes D, Michel O, Chevalier J, Scalbert E, Ezan E, Bariety J,Zimmerman A, Corman B: Effect of chronic ANG I-convertingenzyme inhibition on the aging processes. I. Kidney structure andfunction.Am J Physiol266: R1038–R1051, 1994
27. Gil J, Barba J: Principles of stereology. In:Image Analysis: APrimer for Pathologists, edited by Marchevski AM, Bartels PH,New York, Raven, 1994, pp 79–124
28. Weibel ER: Practical methods for biological morphometry (Vol-ume 1) and theoretical foundations (Volume 2). In:StereologicalMethods, New York, Academic, 1979
29. Michel JB, Heudes D, Michel O, Potevin P, Philippe M, Scalbert
1496 Journal of the American Society of Nephrology J Am Soc Nephrol 11: 1488–1497, 2000

E, Corman B, Le´vy BI: Effect of chronic ANG I-convertingenzyme inhibition on the aging processes. II. Large arteries.Am JPhysiol267: R124–R135, 1994
30. Harisson D, Archer J, Astle C: Effects of food restriction onaging: Separation of food intake and adiposity.Proc Natl AcadSci USA81: 1835–1838, 1984
31. Beauchene R, Bales C, Bragg C, Hawkins S, Mason R: Effect ofage of initiation of feed restriction on growth, body composition,and longevity of rats.J Gerontol41: 13–19, 1986
32. Cheney K, Liu R, Smith G, Meredith P, Mickey M, Walford R:The effect of dietary restriction of varying duration on survival,tumor patterns, immune function, and body temperature inB10C3F1 female mice.J Gerontol38: 420–430, 1983
33. Weindruch R, Walford R: Dietary restriction in mice beginningat 1 year of age: Effect on life span and spontaneous cancerincidence.Science215: 1415–1418, 1982
34. Goodrick C, Ingram D, Reynolds M, Freeman J, Cider N: Effectsof intermittent feeding upon body weight and lifespan in inbredmice: Interaction of genotype and age.Mech Ageing Dev55:69–87, 1990
35. Barzilai N, Rosetti L: Age-related changes in body compositionare associated with hepatic insulin resistance in conscious rats.Am J Physiol270: E930–E936, 1996
36. Bergamini E, Cacallini G, Del Roso A: Different circadianvariation of plasma glucose and insulin concentration in ratssubmitted to 60% food restriction or intermittent feeding. In:Protein Metabolism in Aging, edited by Segal HL, Rothstein M,Bergamini E, New York, Wiley-Liss, 1990, pp 295–297
37. Corman B, Pratz J, Poujeol P: Changes in anatomy, glomerular
filtration and solute excretion in aging rat kidney.Am J Physiol248: R282–R287, 1985
38. Francois VD, Heudes J, Bariety P, Bruneval B, Corman B:Glomerular capillary network of cortical nephron is reduced inmale but not in female aging rats.Mech Ageing Dev91: 11–22,1996
39. Cohen MP, Hud E, Wu VY, Ziyadeh FN: Albumin modified byAmadori glucose adducts activates mesangial cell type IV col-lagen gene transcription.Mol Cell Biochem151: 61–67, 1995
40. Vlassara H, Striker L, Teichberg S, Fuh H, Li Y, Steffes M:Advanced glycation end products induce glomerular sclerosisand albuminuria in normal rats.Proc Natl Acad Sci USA91:11704–11708, 1994
41. Brownlee M, Vlassara H, Kooney A, Ulrich P, Cerami A: Ami-noguanidine prevents diabetes-induced arterial wall proteincross-linking.Science232: 1629–1632, 1986
42. Baylis C, Corman B: Kidney aging: Insights from experimentalstudies.J Am Soc Nephrol9: 699–709, 1998
43. Corman B, Chami-Khazraji S, Schaeverbeke J, Michel JB: Effectof feeding on glomerular filtration rate and proteinuria in con-scious aging rats.Am J Physiol255: F250–F256, 1988
44. Van Liew J, Davis F, Davis P, Noble B, Bernardis L: Calorierestriction decreases microalbuminuria associated with aging inbarrier-raised Fisher 344 rats.Am J Physiol263: F554–F561,1992
45. Geelen G, Corman B: Relationship between vasopressin and renalconcentrating ability in aging rats.Am J Physiol262: 826–833,1992
J Am Soc Nephrol 11: 1488–1497, 2000 AGE Accumulation and Kidney Aging 1497


![5 Leaving on a jet plane Vuong - currytbcenter.ucsf.edunid]/5_leaving_on_a_jet... · Do Not Board (DNB) This travel restriction prevents people who meet specific criteria from obtaining](https://static.fdocuments.us/doc/165x107/5d54c90a88c9934e2b8b5fbf/5-leaving-on-a-jet-plane-vuong-nid5leavingonajet-do-not-board-dnb.jpg)















