
FLUORIDE TOXICITY AND ITS EFFECTS ON GAMETOGENESIS … · Research report Fluoride 46(1)7–18...
12
Research report Fluoride 46(1)7–18 January-March 2013 Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi Beddard Casellato, Del Piero, Masiero, Covre 7 7 FLUORIDE TOXICITY AND ITS EFFECTS ON GAMETOGENESIS IN THE AQUATIC OLIGOCHAETE BRANCHIURA SOWERBYI BEDDARD Sandra Casellato, a Stefania Del Piero, a Luciano Masiero, a Valeria Covre a Padova, Italy SUMMARY: The purpose of this study was to examine the tolerance of the oligochaete Branchiura sowerbyi Beddard and its capacity to accumulate fluoride (F) by performing short-term (96-hr LC 50 ) and long-term experiments (18 days) at different temperatures (17ºC and 22ºC). LC 50 values showed that this aquatic worm is more resistant to F than other freshwater invertebrates, especially in the presence of sediment. Moreover, the effects of fluoride on gametogenesis were examined by exposing this oligochaete to concentrations 50 to 100 times above the normal of 0.1–0.3 mg F/L for five months from the end of February 2012 to the end of July 2012. The results revealed temporal differences in the gametogenesis phases of treated specimens, such as incomplete spermatozoa maturation when exposed to the highest fluoride concentration. At the end of the experiment, the exposed animals were thinner and shorter than the control, and normal cocoon deposition did not occur. Keywords: Abnormal gametogenesis; Branchiura sowerbyi Beddard; Fluoride accumulation; Oligochaeta. INTRODUCTION The inorganic fluoride (F) level in freshwater varies from ≤0.1 mg/L to more than 1 mg/L in volcanic areas, 1 but nowadays local levels of F are often increasing owing to the impact of human activities, such phosphate fertilizer production, aluminum smelting, bad waste disposal, and many others industrial processes. F is found in many common household products, including toothpastes, some vitamin supplements, and medicines, and it is also present in nearly every food. 2 Due to its ubiquity, F is ingested daily in varying amounts. However, in recent decades, serious health consequences of excessive F intake have been found, warranting further precautionary measures from its consumption in high dosages. Several studies on animal and plant phyla in aquatic environments have revealed that safe concentrations of F are significantly different between classes, families, genera, and species, and that aquatic animals living in freshwaters are more adversely affected by F contamination than those living in seawater because of the bioavailability of F ions (and, consequently, their toxic action) is reduced with increasing water hardness. 3 The risks to vertebrates are fairly well known, with the most significant effects on bone cells that can lead to the development of skeletal fluorosis. 4 It is also now recognised that F can affect soft tissue cells, such as renal, gonadal, and neurological cells, 5-7 and that its main toxic effect is due to its interaction with enzymes. 8,9 The effect of F on animal and human reproduction has likewise become an area of great interest, and several studies have reported toxic effects on the reproductive system, particularly in males. 10-18 Studies on rats revealed that NaF caused alterations in testicular seminiferous tubules, such as vacuolation and denudation a For correspondence: Sandra Casellato, Department of Biology, University of Padova, Via U. Bassi 58/A, 35129, Italy; E-mail: [email protected]
Transcript of FLUORIDE TOXICITY AND ITS EFFECTS ON GAMETOGENESIS … · Research report Fluoride 46(1)7–18...

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
77
FLUORIDE TOXICITY AND ITS EFFECTS ON GAMETOGENESIS IN THE AQUATIC OLIGOCHAETE BRANCHIURA
SOWERBYI BEDDARDSandra Casellato,a Stefania Del Piero,a Luciano Masiero,a Valeria Covrea
Padova, Italy
SUMMARY: The purpose of this study was to examine the tolerance of the oligochaeteBranchiura sowerbyi Beddard and its capacity to accumulate fluoride (F) by performingshort-term (96-hr LC50) and long-term experiments (18 days) at different temperatures(17ºC and 22ºC). LC50 values showed that this aquatic worm is more resistant to F thanother freshwater invertebrates, especially in the presence of sediment. Moreover, theeffects of fluoride on gametogenesis were examined by exposing this oligochaete toconcentrations 50 to 100 times above the normal of 0.1–0.3 mg F/L for five months fromthe end of February 2012 to the end of July 2012. The results revealed temporaldifferences in the gametogenesis phases of treated specimens, such as incompletespermatozoa maturation when exposed to the highest fluoride concentration. At the endof the experiment, the exposed animals were thinner and shorter than the control, andnormal cocoon deposition did not occur.
Keywords: Abnormal gametogenesis; Branchiura sowerbyi Beddard; Fluoride accumulation; Oligochaeta.
INTRODUCTION
The inorganic fluoride (F) level in freshwater varies from ≤0.1 mg/L to morethan 1 mg/L in volcanic areas,1 but nowadays local levels of F are often increasingowing to the impact of human activities, such phosphate fertilizer production,aluminum smelting, bad waste disposal, and many others industrial processes. F isfound in many common household products, including toothpastes, some vitaminsupplements, and medicines, and it is also present in nearly every food.2 Due to itsubiquity, F is ingested daily in varying amounts. However, in recent decades,serious health consequences of excessive F intake have been found, warrantingfurther precautionary measures from its consumption in high dosages.
Several studies on animal and plant phyla in aquatic environments have revealedthat safe concentrations of F are significantly different between classes, families,genera, and species, and that aquatic animals living in freshwaters are moreadversely affected by F contamination than those living in seawater because of thebioavailability of F ions (and, consequently, their toxic action) is reduced withincreasing water hardness.3 The risks to vertebrates are fairly well known, with themost significant effects on bone cells that can lead to the development of skeletalfluorosis.4 It is also now recognised that F can affect soft tissue cells, such as renal,gonadal, and neurological cells,5-7 and that its main toxic effect is due to itsinteraction with enzymes.8,9
The effect of F on animal and human reproduction has likewise become an areaof great interest, and several studies have reported toxic effects on the reproductivesystem, particularly in males.10-18 Studies on rats revealed that NaF causedalterations in testicular seminiferous tubules, such as vacuolation and denudation
aFor correspondence: Sandra Casellato, Department of Biology, University of Padova, Via U.Bassi 58/A, 35129, Italy; E-mail: [email protected]

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
88
of spermatogenic epithelium, which disrupted spermatogenesis, reducing spermcounts and serum testosterone levels.19 Changes in the epididymal secretoryepithelium also impaired its metabolic and functional status, inhibited spermmaturation and motility, and reduced the fertility of treated animals. Negativeeffects of F on gametogenesis have been reported in many animal species as wellas in humans,20 and hence prolonged exposure to F may have serious implicationsfor fertility.
Although aquatic oligochaetes have a long history for their use in pollutionmonitoring,21-27 no data appear to have been reported on the toxicity andaccumulation of F in them. Owing to its relatively large size and prominentposterior gills,28 Branchiura sowerbyi Beddard can be easily collected andidentified, and it has a low natural mortality rate in the laboratory compared withother tubificids.29 These worms feed in the sediment, a behaviour that involves theintake of large amounts of substrate.30 Because of its size, B. sowerbyi is moreappropriate for accumulation tests than smaller oligochaeta species.31 Finally, B.sowerbyi dominates other bottom macroinvertebrates in terms of abundance andbiomass,28 and it is also a significant part of the diet of some fish and crustaceans.As far as we are aware, no previous researchers have studied effects of F ongametogenic processes in aquatic oligochaeta, so our investigation will contributeto fill these gaps.
MATERIALS AND METHODS
The specimens of B. sowerbyi used in our study came from a lily tank at theBotanic Garden of Padua University, which has been monitored for twodecades.32,33 The tank is filled by a thermal spring, and the temperature fluctuatesbetween 17°C and 27°C throughout the year. Samples of the original tank waterand the original sediment were collected, together with the oligochaetes, to set upoptimal laboratory culture and test conditions. Two hundred worms were graduallyacclimatised for one week to the test temperatures in 1-L glass containerscontinuously aerated. To evaluate the toxic effects of sodium fluoride (NaF),different experiments were performed: four short-term tests (96-hr), two long-termtests (18 days), and one test on the gametogenesis for 5 months, starting from thebeginning of February 2012.
To determine whether the temperature has an influence on F toxicity andaccumulation, the short- and long-term experiments were performed at 17±0.5ºCand 22±0.5ºC, which are the temperatures at which B. sowerbyi has the lowest andthe highest growth rate, respectively.34 The vitality of the animals was frequentlychecked using soft tweezers, and they were considered dead when there was noresponse to physical stimulation. The dead specimens were removed and kept in4% formaldehyde for F accumulation analysis. At the end of the experiments, thesurviving worms were also placed in formaldehyde and analysed.
F accumulation in the soft tissues of B. sowerbyi (as total body) was determinedby the method of Malde et al.35 For F in the sediment, the alkali fusion method ofMcQuacker and Gurney36 was used. The analyses were conducted at pH 5.5 afteradding a total ionic strength adjustment buffer (TISAB III) and using a F ion-

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
99
selective electrode ISE F800 (0.02 mg F–/L to saturation, accuracy 0.1%, WTW).An analysis of variance (ANOVA) was carried out on F accumulation values(expressed in µg F–/g of dry weight) to determine the change in the responsevariable (Software R ver. 2.15.0). Data relationships were assessed using simplelinear regression.
Short-term experiments: In these experiments, performed with and withoutsediment, the worms were exposed to NaF at nominal concentrations of 80, 160,320, and 640 mg F–/L in glass containers covered with Plexiglas plates to preventF evaporation. The daily mortality percentage was used to estimate LC50 values atthe 95% confidence level using multifactor probit analysis (SAS softwarever.9.1.3).37
Without sediment: Fifteen worms (three replicates of five) were exposed to NaFin glass containers, filled only with the original water.
With sediment: The same number of worms were treated with NaF, introducing125 g of site sediment into each glass container.
Long-term experiments: In the long-term tests, three replicates of five worms foreach temperature were exposed to NaF at nominal concentrations of 80, 120, and160 mg F–/L, which are the concentrations at which B. sowerbyi showed thelowest mortality percentage in the short-term tests. To ensure the survival of theanimals, these long-term tests were performed only in the presence of sitesediment. Every week 10 mL of water was taken from each container andcombined with 10 mL of TISAB III for ion-selective electrode analysis. At the endof the experiment, F content analysis was performed on both the worms and thesediment.
Effects on gametogenesis: To mimic the conditions of the original sampling site,three small 10-L aquariums were prepared using the original water together withan 8-cm layer of the original sediment and the original population density (200worms/m2). Ninety specimens (30 in each aquarium), with an average length of 9–12 cm) were reared for five months and exposed to 0, 15 and 30 mg NaF/L at23±1ºC. This temperature was chosen for optimal gamete maturation.38 Thetreated and untreated specimens were checked weekly to monitor gametematuration in both male and female germinal lines. Every week some specimenswere removed and anaesthetised with 70% alcohol, after which segments from 5 to17, containing the entire sexual apparatus, were dissected using a micro-scissors.These segments were placed on a slide and gently compressed with a cover slip tolet the germinal cysts shed into the coelomatic fluid from the testes, ovaries, andseminal vesicles. In this manner, cytomorphological changes during thetransformation of spermatogonia to mature spermatozoans in males and thetransformation of oogonia to mature eggs in females could be observed under alight microscope (Leica Application Suite software with a Leica HQ DFC480camera) over the course of five months.
The nominal F concentrations were maintained by weekly renewal of the waterduring the entire treatment period. In addition, F accumulation in the sediment was

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
1010
periodically tested to quantify the real exposure level of the worms to F over time.The F concentration was determined from aliquots of water from the aquariums atthe beginning of the experiment and before every weekly renewal using the ion-selective electrode ISE F800 already described. The sediment was also monitoredby taking 0.5-g samples and analysing them by the alkali fusion method, asdescribed by McQuaker and Gurney.36 Chemical analyses were conductedperiodically (monthly) during the experiments to quantify organic matter content39
and the concentration of dissolved oxygen.40
RESULTS
Short-term experiments: B. sowerbyi demonstrated high tolerance to F,especially at 17ºC: at this temperature the mortality was lower than at 22ºC, bothwith and without sediment (Table). The mortality was also reduced in the presenceof sediment at both 17ºC and 22ºC, as evidenced by the LC50 values (Table).
Temperature influenced F accumulation, with worms treated at 22ºCaccumulating greater amounts of F in their bodies than those treated at 17ºC(Figure 1A, B). The presence of sediment enhanced the F accumulation values atboth temperatures: the animals treated with sediment accumulated double theamount of F (Figure 1A), and the accumulation trend increased with increasing Fconcentration (p<0.001).
Long-term experiments: The influence of temperature on F toxicity in B.sowerbyi was also evident also in the long-term tests. Although the same mortalitypercentage was found for all concentrations at 17ºC, it increased monotonically by1.2% for every 10 mg F–/L added (r = 0.94) at 22ºC (Figure 2).
F accumulation in B. sowerbyi showed a significant statistical relationship withthe increasing concentration of F in the water (p<0.001), and the highest valueswere observed in animals treated at 22ºC (Figure 3).
Sediment adsorbed part of the F dissolved in water, especially at 22ºC, and theaccumulation trend was directly proportional to the increasing F concentration inthe water (p<0.01) (Figure 4).
Table. LC50 values at 96 hr for B. sowerbyi at different temperatures (95% confidence limits in parenthesis)
Temperature
17±0.5°C 22±0.5°C
Sediment present
Sediment absent
Sediment present
Sediment absent
LC50 (mg F–/L) 267.63 (257.75–277.51)
91.28 (84.50–98.05)
80.07 (62.10–111.55)
61.68 (47.83–90.11)

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
1111
Figure 1. F accumulation in B. sowerbyi during short term experiments (96 hr) attemperatures of 17°C and 22°C. A) With sediment; B) Without sediment. (Error bars showthe standard deviations).
Figure 2. Mortality percentages in B. sowerbyi during long-term experiments (18 days) attemperatures of 17°C and 22°C.

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
1212
Water analyses showed that animals and sediment absorbed the most F duringthe first 6 days of the experiment, and that the highest percentage decrease of F inthe water was reached at 22ºC (Figure 5). F concentrations in the water wereclearly inversely related to increasing temperature.
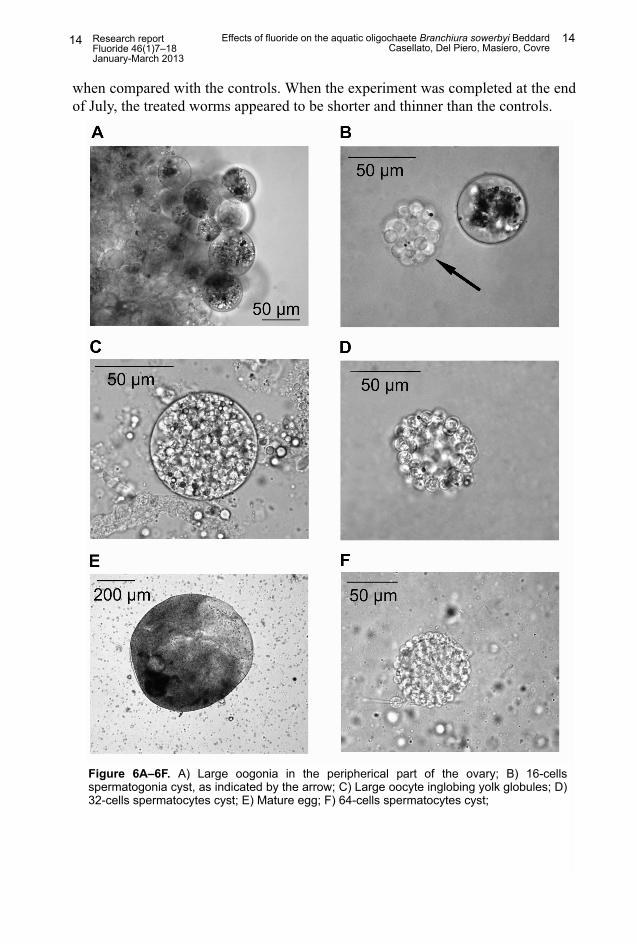
Effects on gametogenesis: When the specimens of B. sowerbyi were firstexposed to NaF at the beginning of February 2012, they showed normal gonadactivity, as indicated by the increased volume of the testes and ovaries, as well asby the appearance of four to eight spermatogonial cysts in the sperm sacs of malesand large oogonia in the ovaries of females (Figure 6A). None of the specimenshad yet developed a clitellum, which appeared only after one month of NaFexposure in both the treated animals and in the controls.
Figure 4. Sediment F content at the end of the long term experiments (18 days) at temperatures of 17°C and 22°C. (Error bars show the standard deviations).
Figure 3. F accumulation in B. sowerbyi during long term experiments (18 days) at temperatures of 17°C and 22°C. (Error bars show the standard deviations).

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
1313
At the beginning of March 2012, the first 16-cell spermatogonial cysts (46 µm)(Figure 6B) appeared in the controls, which occurred one month later in the treatedanimals. On the other hand, free oocytes in the coelom were found in all thetreated and the untreated specimens at the beginning of March (Figure 6C). Theyappeared greatly enlarged (70 µm), showing more inglobing yolk globules in theircytoplasm. The first 32-cell spermatocyte cysts (60 µm) appeared at the beginningof April in the control specimens, but they did not appear until one month later inthe worms treated with either 15 mg/L or 30 mg/L of NaF (Figure 6D). The firstmature eggs were observed in the control animals. These were observed only onemonth later for those treated with 15 and 30 mg NaF/L (Figure 6E). However,development of the male germinal line occurred normally in the control animals.The 64-cell spermatocytes (Figure 6F) and the 128-cells spermatid cysts (Figure6G) were present in large numbers in the coelom cavity, and free spermatozoanswere present in some of the controls at the end of May (Figure 6H). In the treatedworms, the first spermatid cysts were not observed until one month later, and theyshowed certain deformities (Figure 6I). No free spermatozoans were found inthese specimens. At the beginning of June, some spermatozeugmata could beobserved in the seminal funnels of the control specimens, and these were alsofound one month later into the spermathecae (Figure 6J), which confirmed theyhad copulated. These structures have already been described for other tubificidspecies as sperm bundles that are transferred into the spermathecae of thecopulation partner (Figure 6K).41,42 Mature spermatozoa and spermatozeugmatawere never observed in the treated specimens, which also had fewer mature eggs,
Figure 5. Decreasing water F content during long term experiments (18 days) attemperatures of 17°C and 22°C.
Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
1414
when compared with the controls. When the experiment was completed at the endof July, the treated worms appeared to be shorter and thinner than the controls.
Figure 6A–6F. A) Large oogonia in the peripherical part of the ovary; B) 16-cellsspermatogonia cyst, as indicated by the arrow; C) Large oocyte inglobing yolk globules; D)32-cells spermatocytes cyst; E) Mature egg; F) 64-cells spermatocytes cyst;

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
1515
The samples from the aquarium containing 15 mg NaF/L showed that theconcentration varied from 15 to 19 mg F–/L. In the aquarium that contained 30 mgNaF/L, the values ranged from 30 to 39.56 mg F–/L, suggesting that some of the Fwas absorbed by the sediment and was then released back into the water at eachweek of water renewal. The F concentration in the sediment increased from aninitial level of 2.59 ppm to 95.87 ppm (dry weight) in the aquarium filled with 15
Figure 6G–6K. G) 128-cells spermatid cyst; H) Free spematozoans; I) Abnormal spermatid cyst in a treated specimen; J) Spermatozeugma (sperm bundle) into the spermatheca of a specimen of B. sowerbyi after copulating, as indicated by the arrow; K) Spermatozeugma.

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
1616
mg/L of NaF and reached 122.01 ppm in the aquarium with 30 mg NaF/L,indicating that the worms were exposed to a higher concentration of F than thenominal level in the water during the experimental period. The amount ofdissolved oxygen varied between 3.9 and 5.3 mg/L, corresponding to an averagesaturation percentage of 68% at 25ºC. The mean organic matter content neverexceeded 4%, which is consistent with the level in the original sampling site.
DISCUSSION
As our data indicate, increased temperature amplified the toxicity of F: withhigher temperature, B. sowerbyi became less tolerant to this pollutant. Angelovicet al.43 demonstrated that higher temperature increases the rate of metabolism andtoxicity of F in rainbow trout, and this probably happened in our experiments,resulting in faster F absorption and faster poisoning of the enzyme systems at 22ºCthan at 17ºC. Camargo3 in his review reported similar findings of increased Ftoxicity in freshwater invertebrates, showing that the 96-hr LC50 ranges from 10.5mg F–/L for some species of crustaceans to 45 mg F–/L for the most resistantspecies of caddisfly (Chimarra marginata). B. sowerbyi showed a 96-hr LC50close to the 24-hr LC50 value of Daphnia magna, which is considered one of theless sensitive invertebrate species.44 In the absence of sediment, 50% of the wormpopulation died in water with 61.68 mg F–/L at 22ºC; the same value was reportedfor Dreissena polymorpha.45 The resistance of B. sowerbyi during short-termexposure was enhanced by the presence of the sediment. Chapman et al.,25
Casellato and Negrisolo,26 and Naqui46 used various toxicity bioassays todemonstrate that sediment reduces the bioavailability of contaminants, limiting thetoxic effects on organisms exposed to pollutants dissolved in water. This wasobserved for B. sowerbyi only in the short-term experiments. However, when theexposure time was extended to 18 days, these tubificids fed on the contaminatedsediment. Thus, their F accumulation values increased (Figure 4). Severalauthors47-49 have suggested that the contaminated sediments ingested by deposit-feeding animals should be considered the major source of the absorbedcontaminant. Therefore, sediment was an important source for the accumulation ofF in the long-term exposures because of its high adsorption capacity. We alsoobserved that absence of sediment represents a source of stress for this species,because then the worms were unable to feed.
Finally, as in studies on vertebrates showing a toxic effect of F onspermatogenesis, our results demonstrated that this toxicity of F is more commonthan commonly thought, being present in many animal groups. Prolongedexposure to F damages the gametogenic cycle in B. sowerbyi, particularly thespermatogenic one, even if the worms were exposed to concentrations that did notshow an effect from short- or medium-term exposure. Temporal differences wereobserved in the gametogenetic phases of the treated specimens when comparedwith the controls. The treated specimens did not complete spermatozoanmaturation at the highest NaF concentration used, and the number of mature eggsproduced was greatly reduced. The male line stopped maturation at the spermatidstage, which appeared abnormal and regressed. Moreover, the treated animals

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
1717
were thinner and shorter than the controls at the end of the experiment, and cocoondeposition did not occur.
Whereas F toxicity data for mammals and birds are fairly abundant, suchinformation for invertebrates has been completely lacking until now. Our researchon B. sowerbyi emphasises the importance of investigating other animal groups toexplain the exact mode of action of F and to monitor its toxic potential.
ACKNOWLEDGEMENTS
This work was supported by a grant from Padova University “Progetti di Ateneo(PRAT)–Bando 2010”.
REFERENCES1 Dobbs GG. Fluoride and the environment. Fluoride 1974;7(3):123-35.2 Waldbott GL. Fluoride in food. Am J Clin Nutr 1963;12:455-62.3 Camargo JA. Fluoride toxicity to aquatic organisms: a review. Chemosphere 2003;50:251-64.4 Dhar V, Bhatnagar M. Physiology and toxicity of fluoride. Indian J Dent Res 2009;20:350-5.5 National Research Council of the National Academies. Fluoride in drinking-water: a scientific
review of EPA’s standards. Washington, DC: The National Academies Press; 2006.6 Lantz O, Jouvin MH, De Vernejoul MC, Druet P. Fluoride-induced chronic renal failure. Am J
Kidney Dis 1987;10(2):136-9.7 Basha PM, Rai P, Begum S. Evaluation of fluoride-induced oxidative stress in rat brain: a
multigeneration study. Biol Trace Elem Res 2010;142(3):623-37.8 Casellato S, Masiero L, Ballarin L. Toxicity of fluoride to the freshwater mollusc Dreissena
polymorpha: effects on survival, histology, and antioxidant enzyme activity. Fluoride2012;45(1):35-46.
9 Barbier O, Arreola-Mendoza L, Del Razo LM. Molecular mechanisms of fluoride toxicity.Chem Biol Interact 2010;188:319-33.
10 Long H, Jin Y, Lin M, Sun Y, Zhang L, Clinch C. Fluoride toxicity in the male reproductivesystem. Fluoride 2009;42(4):260-76.
11 Hoffman DJ, Pattee OH, Wiemeyer SN. Effects of fluoride on screech owl reproduction:teratological evaluation, growth, and blood chemistry in hatchlings. Toxicol Lett1985;26(1):19-24.
12 Mehdi AWR, Al-Soudi KA, Al-Jiboori NAJ, Al-Hiti MK. Effect of high fluoride intake on chickenperformance, ovulation, spermatogenesis and bone fluoride content. Fluoride 1983;16(1):37-43.
13 El-lethey HS, Kamel MM, Shaheed IB. Perinatal exposure to sodium fluoride with emphasison territorial aggression, sexual behaviour and fertility in male rats. Life Sci J 2011;8(2):686-94.
14 Ghosh D, Das Sarkar S, Maiti R, Jana D, Das UB. Testicular toxicity in sodium fluoride treatedrats: association with oxidative stress. Reprod Toxicol 2002;16:385-90.
15 Reddy PS, Pushpalatha T, Sreenivasula Reddy P. Suppression of male reproduction in ratsafter exposure to sodium fluoride during early stages of development. Naturwissenschaften2007;94:607-11.
16 Kour K, Singh J. Histological finding of mice testes following fluoride ingestion. Fluoride1980;13(4):160-2.
17 Elbetieha A, Darmani H, Al-Hiyasat AS. Fertility effects of sodium fluoride in male mice.Fluoride 2000;33(3):128-34.
18 Susheela AK, Kumar A. A study of the effect of high concentrations of fluoride on thereproductive organs of male rabbits, using light and scanning electron microscopy. J ReprodFertil 1991;92:353-60.
19 Wan SX, Zhang JH, Wang JD. Effects of high fluoride on sperm quality and testicularhistology in male rats. Fluoride 2006;39(1):17-21.
20 Chinoy NJ, Narayana MV. In vitro fluoride toxicity in human spermatozoa. Reprod Toxicol1994;8(2):155-9.
21 Champman PM. Utility and relevance of aquatic oligochaetes in Ecological Risk Assessment.Hydrobiologia 2001;463:149-69.
22 Ingersoll CG, Ankley GT, Benoit DA, Brunson EL, Burton GA, Dwyer FJ, et al. Toxicity andbioaccumulation of sediment associated contaminants using freshwater invertebrates: areview of methods and applications. Environ Toxicol Chem 1995;14:1885-94.

Research reportFluoride 46(1)7–18January-March 2013
Effects of fluoride on the aquatic oligochaete Branchiura sowerbyi BeddardCasellato, Del Piero, Masiero, Covre
1818
23 Reynoldson TB, Thompson SP, Bamsey JL. A sediment bioassay using the tubificidoligochaete worm Tubifex tubifex. Environ Toxicol Chem 1991;10:1061-72.
24 Bhunia F, Saha NC, Kaviraj A. Toxicity of thiocyanate to fish, plankton, worm and aquaticecosystem. Bull Environ Contam Toxicol 2000;64:197-204.
25 Chapman PM, Farrell MA, Brinkhurst RO. Relative tolerances of selected aquaticoligochaetes to individual pollutants and environmental factors. Aquat Toxicol 1982;2:47-67.
26 Casellato S, Negrisolo P. Acute and chronic effects of an anionic surfactant on somefreshwater tubificid species. Hydrobiologia 1989;180:243-52.
27 Casellato S, Aiello R, Negrisolo PA, Seno M. Long-term experiment on Branchiura sowerbyiBeddard (Oligochaeta, Tubificidae) using sediment treated with LAS (Linear AlkylbenzeneSulphonate). Hydrobiologia 1992;232:169-73.
28 Ducrot V, Pèry ARR, Quèau H, Mons R, Lafont M, Garric J. Rearing and estimation of life-cycle parameters of the tubificid worm Branchiura sowerbyi: application to ecotoxicity testing.Sci Total Environ 2007;384:252-63.
29 Bonacina C, Pasteris A, Bonomi G, Marzuoli D. Quantitative observations on the populationecology of Branchiura sowerbyi (Oligochaeta, Tubificidae). Hydrobiologia 1994;278:267-74.
30 Wang X, Matisoff G. Solute transport in sediments by a large freshwater Oligochaete,Branchiura sowerbyi. Environ Sci Technol 1997;31:1926-33.
31 Chapman PM, Churchland LM, Thompson P, Michnowsky E. Heavy metal studies witholigochaetes. In: Brinkhurst RO, Cook DG, editors. Aquatic Oligochaete Biology. New York:Plenum Press; 1980.
32 Casellato S. Life-cycle and karyology of Branchiura sowerbyi Beddard (Oligochaeta,Tubificidae). Hydrobiologia 1984;115:65-9.
33 Casellato S, Martinucci G, Zoja M. Ultrastructural features of gametogenesis during the lifecycle in Branchiura sowerbyi Beddard (Oligochaeta, Tubificidae). Hydrobiologia1987;155:145-54.
34 Aston RJ, Sadler K, Milner AGP. The effects of temperature on the culture of Branchiurasowerbyi (Oligochaeta, Tubificidae) on activated sludge. Aquaculture 1982;29:137-45.
35 Malde MJ, Bjorvatn K, Julshamn K. Determination of fluoride in food by the use of alkali fusionand fluoride ion-selective electrode. Food Chem 2001;73:373-9.
36 McQuaker N, Gurney M. Determination of total fluoride in soil and vegetation using an alkalifusion-selective ion electrode technique. Anal Chem 1977;49:53-6.
37 Lee G. Multifactor probit analysis. Gulf Breeze: US Environmental Protection Agency, Officeof Research and Development, Environmental Research Laboratory; 1991.
38 Aston RJ. The effect of temperature on the life cycle, growth and fecundity of Branchiurasowerbyi (Oligochaeta: Tubificidae). J Zool 1968;154:29-40.
39 Gaudette HE, Wilson RF, Toner L, Folger DW. An inexpensive titration method for thedetermination of organic carbon in recent sediments. Journal Sedimentary Petrology1974;44:249-53.
40 Strickland JDH, Parsons TR. A practical handbook of seawater analysis. Ottawa: FisheriesResearch of Canada; 1968.
41 Braidotti P, Ferraguti M. Two sperm types in the spermatozeugmata of Tubifex tubifex(Annelida, Oligochaeta). J Morphol 1982;171:123-36.
42 Ferraguti M, Grassi G, Ersèus C. Different models of tubificid spermatozeugmata.Hydrobiologia 1989;180(1):73-82.
43 Angelovic JW, Sigler WF, Neuhold JM. Temperature and fluorosis in rainbow trout. WaterPollut Control 1961;33:371-81.
44 Dave G. Effects of fluoride on growth, reproduction and survival in Daphnia magna. CompBiochem Phys 1984;78C:425-31.
45 Del Piero S, Masiero L, Casellato S. Influence of temperature on fluoride toxicity andbioaccumulation in the non-indigenous mollusk Dreissena polymorpha Pallas, 1769. EnvironToxicol Chem 2012;31:1-5.
46 Naqui SMZ. Toxicity of twenty-three insecticides to a tubificid worm Branchiura sowerbyi fromMississippi Delta. J Econ Entomol 1973;66:70-4.
47 Lee BG, Griscom SB, Lee JS, Choi HJ, Kohn CH, Luoma SN, et al. Influences of dietaryuptake and reactive slides on metal bioavailability from aquatic sediments. Science2000;287:282-4.
48 Volparil IM, Mayer LM. Dissolution of sedimentary polycyclic aromatic hydrocarbons into thelugworm’s (Arenicola marina) digestive fluids. Environ Sci Technol 2000;34:1221-8.
49 Weston DP, Penry DL, Gulmann LK. The role of ingestion as a route of contaminantbioaccumulation in a deposit-feeding polychaete. Arch Environ Con Tox 2000;38(4):446-54.
Copyright © 2013 The International Society for Fluoride Research Inc. www.fluorideresearch.org www.fluorideresearch.com www.fluorideresearch.net
Editorial Office: 727 Brighton Road, Ocean View, Dunedin 9035, New Zealand.


















