
Fluorescent Tagging and Cellular Distribution of the ...jvi.asm.org/content/88/21/12839.full.pdf ·...
14
Fluorescent Tagging and Cellular Distribution of the Kaposi’s Sarcoma-Associated Herpesvirus ORF45 Tegument Protein Shir Bergson, a Inna Kalt, a Inbal Itzhak, a Kevin F. Brulois, b Jae U. Jung, b Ronit Sarid a Mina and Everard Goodman Faculty of Life Sciences, Bar Ilan University, Ramat-Gan, Israel a ; Department of Molecular Microbiology and Immunology, Keck School of Medicine, University of Southern California, Los Angeles, California, USA b ABSTRACT Kaposi’s sarcoma-associated herpesvirus (KSHV), also known as human herpesvirus 8 (HHV-8), is a cancer-related human vi- rus, classified as a member of the Gammaherpesvirinae subfamily. We report here the construction of a dual fluorescent-tagged KSHV genome (BAC16-mCherry-ORF45), which constitutively expresses green fluorescent protein (GFP) and contains the tegu- ment multifunctional ORF45 protein as a fusion protein with monomeric Cherry fluorescent protein (mCherry). We confirmed that this virus is properly expressed and correctly replicates and that the mCherry-ORF45 protein is incorporated into the viri- ons. Using this labeled virus, we describe the dynamics of mCherry-ORF45 expression and localization in newly infected cells as well as in latently infected cells undergoing lytic induction and show that mCherry can be used to monitor cells undergoing the lytic viral cycle. This virus is likely to enable future studies monitoring the dynamics of viral trafficking and tegumentation dur- ing viral ingress and egress. IMPORTANCE The present study describes the construction and characterization of a new recombinant KSHV genome BAC16 clone which ex- presses mCherry-tagged ORF45. This virus enables the tracking of cells undergoing lytic infection and can be used to address issues related to the trafficking and maturation pathways of KSHV virions. K aposi’s sarcoma-associated herpesvirus (KSHV), also known as human herpesvirus 8 (HHV-8), is a cancer-related human virus which is classified as a member of the Gammaherpesvirinae subfamily (1–3). Like all other herpesviruses, KSHV exhibits two alternative infection cycles, lytic and latent; both cycles are neces- sary for long-term persistence of KSHV and for its pathogenesis. Infection with KSHV begins with the attachment and entry of KSHV virions into the cell, while fusion of the viral envelope with endocytic vesicles releases the tegumented capsids into the cyto- plasm. Incoming nucleocapsids then utilize motor proteins to reach the nuclear pores and release the viral genome into the nu- cleoplasm, where transcription of viral genes and viral genomic replication take place (4). The virus may enter a productive lytic or nonproductive latent infection. The lytic cycle is characterized by a temporally regulated cascade of viral gene expression and viral DNA replication that culminates in the assembly, maturation, and release of newly synthesized virions. Latent infection, the com- mon in vitro default replication program of KSHV, which involves the expression of a small set of viral genes and presence of viral episomes, can be established following the expression of a unique set of viral genes under nonpermissive cellular conditions (5). Nevertheless, under conditions that induce the expression of the virally encoded regulatory protein RTA, the latent viral genome may reactivate and switch the viral genetic program toward pro- ductive infection (6–8). All mature herpesvirus particles have a characteristic multilay- ered architecture, including (i) an inner core containing the linear double-stranded viral genome, (ii) an icosahedral protein shell called the capsid, (iii) an outer lipid bilayer envelope spiked with viral glycoproteins, and (iv) a thick proteinaceous electron-dense layer designated the tegument, which is located between the nu- cleocapsid and the envelope. Herpesviral assembly is a multistage event consisting of the formation of capsids within the nucleus, packaging of the replicated viral DNA into the capsids, and exit from the nucleus to the cytoplasm via the acquisition of the pri- mary envelope by budding through the inner nuclear membrane and its subsequent loss at the outer nuclear membrane. During primary envelopment, a portion of the tegument proteins are al- ready bound to the nucleocapsid. In the cytoplasm, tegument pro- teins join the partially tegumented nucleocapsids, and the capsids are enveloped in the trans-Golgi apparatus. Final envelopment, including acquisition of additional tegument proteins, the lipid bilayer envelope, and viral glycoproteins, occurs through the bud- ding into Golgi vesicles. Ultimately, virion-containing vesicles fol- low the secretory pathway to the cell membrane, and mature viral particles are released into the extracellular environment by exocy- tosis (9, 10). The entire assembly course is regulated mainly by tegument proteins that sequentially interact with capsid, enve- lope, and cellular proteins at different intracellular locations dur- ing virus egress. Yet the molecular mechanisms that enable re- cruitment of tegument proteins to the nucleocapsid are poorly understood (11, 12). In addition to their role during virus assem- bly and maturation, tegument proteins play important roles in various aspects of the virus lytic replication cycle, at the very early Received 17 April 2014 Accepted 19 August 2014 Published ahead of print 27 August 2014 Editor: R. M. Longnecker Address correspondence to Ronit Sarid, [email protected]. S.B. and I.K. contributed equally to this article. Supplemental material for this article may be found at http://dx.doi.org/10.1128 /JVI.01091-14. Copyright © 2014, American Society for Microbiology. All Rights Reserved. doi:10.1128/JVI.01091-14 November 2014 Volume 88 Number 21 Journal of Virology p. 12839 –12852 jvi.asm.org 12839 on June 8, 2018 by guest http://jvi.asm.org/ Downloaded from
Transcript of Fluorescent Tagging and Cellular Distribution of the ...jvi.asm.org/content/88/21/12839.full.pdf ·...

Fluorescent Tagging and Cellular Distribution of the Kaposi’sSarcoma-Associated Herpesvirus ORF45 Tegument Protein
Shir Bergson,a Inna Kalt,a Inbal Itzhak,a Kevin F. Brulois,b Jae U. Jung,b Ronit Sarida
Mina and Everard Goodman Faculty of Life Sciences, Bar Ilan University, Ramat-Gan, Israela; Department of Molecular Microbiology and Immunology, Keck School ofMedicine, University of Southern California, Los Angeles, California, USAb
ABSTRACT
Kaposi’s sarcoma-associated herpesvirus (KSHV), also known as human herpesvirus 8 (HHV-8), is a cancer-related human vi-rus, classified as a member of the Gammaherpesvirinae subfamily. We report here the construction of a dual fluorescent-taggedKSHV genome (BAC16-mCherry-ORF45), which constitutively expresses green fluorescent protein (GFP) and contains the tegu-ment multifunctional ORF45 protein as a fusion protein with monomeric Cherry fluorescent protein (mCherry). We confirmedthat this virus is properly expressed and correctly replicates and that the mCherry-ORF45 protein is incorporated into the viri-ons. Using this labeled virus, we describe the dynamics of mCherry-ORF45 expression and localization in newly infected cells aswell as in latently infected cells undergoing lytic induction and show that mCherry can be used to monitor cells undergoing thelytic viral cycle. This virus is likely to enable future studies monitoring the dynamics of viral trafficking and tegumentation dur-ing viral ingress and egress.
IMPORTANCE
The present study describes the construction and characterization of a new recombinant KSHV genome BAC16 clone which ex-presses mCherry-tagged ORF45. This virus enables the tracking of cells undergoing lytic infection and can be used to addressissues related to the trafficking and maturation pathways of KSHV virions.
Kaposi’s sarcoma-associated herpesvirus (KSHV), also knownas human herpesvirus 8 (HHV-8), is a cancer-related human
virus which is classified as a member of the Gammaherpesvirinaesubfamily (1–3). Like all other herpesviruses, KSHV exhibits twoalternative infection cycles, lytic and latent; both cycles are neces-sary for long-term persistence of KSHV and for its pathogenesis.Infection with KSHV begins with the attachment and entry ofKSHV virions into the cell, while fusion of the viral envelope withendocytic vesicles releases the tegumented capsids into the cyto-plasm. Incoming nucleocapsids then utilize motor proteins toreach the nuclear pores and release the viral genome into the nu-cleoplasm, where transcription of viral genes and viral genomicreplication take place (4). The virus may enter a productive lytic ornonproductive latent infection. The lytic cycle is characterized bya temporally regulated cascade of viral gene expression and viralDNA replication that culminates in the assembly, maturation, andrelease of newly synthesized virions. Latent infection, the com-mon in vitro default replication program of KSHV, which involvesthe expression of a small set of viral genes and presence of viralepisomes, can be established following the expression of a uniqueset of viral genes under nonpermissive cellular conditions (5).Nevertheless, under conditions that induce the expression of thevirally encoded regulatory protein RTA, the latent viral genomemay reactivate and switch the viral genetic program toward pro-ductive infection (6–8).
All mature herpesvirus particles have a characteristic multilay-ered architecture, including (i) an inner core containing the lineardouble-stranded viral genome, (ii) an icosahedral protein shellcalled the capsid, (iii) an outer lipid bilayer envelope spiked withviral glycoproteins, and (iv) a thick proteinaceous electron-denselayer designated the tegument, which is located between the nu-cleocapsid and the envelope. Herpesviral assembly is a multistageevent consisting of the formation of capsids within the nucleus,
packaging of the replicated viral DNA into the capsids, and exitfrom the nucleus to the cytoplasm via the acquisition of the pri-mary envelope by budding through the inner nuclear membraneand its subsequent loss at the outer nuclear membrane. Duringprimary envelopment, a portion of the tegument proteins are al-ready bound to the nucleocapsid. In the cytoplasm, tegument pro-teins join the partially tegumented nucleocapsids, and the capsidsare enveloped in the trans-Golgi apparatus. Final envelopment,including acquisition of additional tegument proteins, the lipidbilayer envelope, and viral glycoproteins, occurs through the bud-ding into Golgi vesicles. Ultimately, virion-containing vesicles fol-low the secretory pathway to the cell membrane, and mature viralparticles are released into the extracellular environment by exocy-tosis (9, 10). The entire assembly course is regulated mainly bytegument proteins that sequentially interact with capsid, enve-lope, and cellular proteins at different intracellular locations dur-ing virus egress. Yet the molecular mechanisms that enable re-cruitment of tegument proteins to the nucleocapsid are poorlyunderstood (11, 12). In addition to their role during virus assem-bly and maturation, tegument proteins play important roles invarious aspects of the virus lytic replication cycle, at the very early
Received 17 April 2014 Accepted 19 August 2014
Published ahead of print 27 August 2014
Editor: R. M. Longnecker
Address correspondence to Ronit Sarid, [email protected].
S.B. and I.K. contributed equally to this article.
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JVI.01091-14.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.01091-14
November 2014 Volume 88 Number 21 Journal of Virology p. 12839 –12852 jvi.asm.org 12839
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

phases of infection, during progression of the infection, and at thelate phases. Thus, tegument proteins are expressed at various cel-lular compartments, in which they mediate unique functions.
The assembly and egress mechanism of the gammaherpesvi-ruses, in particular that of the KSHV virions, has only begun to beelucidated; however, it is assumed to share a course similar to thatof other herpesviruses (13–16). Proteomic studies using massspectrometry identified the ORF11, -21, -33, -45, -50, -63, -64, and-75 gene products as components of the KSHV tegument andtogether with a systematic investigation of the interactions be-tween the KSHV virion proteins enabled schematic modeling ofthe outer and inner tegument organization (13, 17). KSHV ORF45is an immediate early tegument protein which has homologuesonly in the gammaherpesviruses. During infection, ORF45 isthought to be involved in several functions, including the antiviralresponse, regulation of the lytic cycle, and migration of capsids onmicrotubules. Disruption of ORF45 drastically reduces KSHV in-fectivity and progeny virion yields, though viral gene expressionand DNA replication remain unaffected. This indicates a vital rolefor ORF45 in events leading to viral ingress and egress (18).
In the present study, we used the full-length KSHV genomebacterial artificial chromosome 16 (BAC16) clone to construct arecombinant KSHV containing the monomeric Cherry fluores-cent protein (mCherry) fused to the amino terminus of ORF45.We show that the new recombinant virus undergoes reactivationand produces infectious viruses upon reconstitution in humancells. Incorporation of mCherry-tagged ORF45 into assemblingvirions was as efficient as that of wild-type virus. We investigatedthe cellular compartmentalization of ORF45 and found that un-like that of the ectopically expressed protein, the localization ofmCherry-ORF45 is dynamic along the infection process. Finally,we show that expression of mCherry-ORF45 can be used to trackcells undergoing lytic infection.
MATERIALS AND METHODSCells. HEK-293T, SLK, and iSLK-PURO cells (kindly provided by DonGanem and Rolf Renne) (19) were maintained in Dulbecco’s modifiedEagle’s medium (DMEM) supplemented with 10% fetal calf serum, 50IU/ml penicillin, and 50 �g/ml streptomycin (Biological Industries, Kib-butz Beit Haemek, Israel). Tet system-approved certified fetal bovine se-rum (Biological Industries, Kibbutz Beit Haemek, Israel), 250 �g/mlG418, and 1 �g/ml puromycin (A.G. Scientific, Inc.) were added to iSLK-PURO growth medium to maintain the Tet-On transactivator (Clontech)and RTA expression cassette, respectively.
Plasmids. The expression plasmids encoding green fluorescent pro-tein (GFP)-ORF45 and hemagglutinin (HA)-ORF45 were previously de-scribed (20).
Construction of BAC16 mCherry-ORF45 recombinant virus.BAC16 containing a recombinant full-length KSHV genome containing aGFP cassette under the control of EF-1� promoter has been previouslydescribed (21). The construction of the mCherry-ORF45-expressing vi-rus, BAC16-mCherry-ORF45, was accomplished using the GS1783 strainof Escherichia coli, which encodes inducible Red and I-Sce-I activities, witha two-step recombination protocol, as described previously (22). To con-struct an mCherry cassette for recombination into the BAC16 genome(pEP-mCherry), we first generated a universal transfer plasmid based on ared fluorescent protein (RFP) cassette (pEP-mRFP1), which was kindlyprovided by Nikolaus Osterrieder (22). Specifically, mCherry was ampli-fied by PCR using 5=-AAAGAATTCATGGTGAGCAAGGGCGAGGAATGGTGAGCAAGGGCGAGGA-3= and 5=-AAAGGATCCCTTGTACAGCTCGTCCATGCCGGCATGGACGAGCTGTACAAG-3= primerscontaining EcoRI and BamHI restriction sites and pmCherry-C1 (Clon-
tech) as a template. The resulting PCR product was cloned into the pBlue-script-KS plasmid following digestion with EcoRI and BamHI to producethe pBluescript-mCherry plasmid. A PstI fragment, containing a kanamy-cin resistance (Kanr) cassette and an I-SceI restriction site, was then iso-lated from the pEP-mRFP1 plasmid and inserted into the PstI site ofpBluescript-mCherry, resulting in a plasmid carrying a kanamycin resis-tance cassette and an I-SceI restriction site, flanked with mCherry con-taining a small internal duplication. The resulting plasmid was confirmedby nucleotide sequence analysis. This plasmid was then used as a PCRtemplate to obtain a linear DNA fragment encompassing mCherry, a ka-namycin resistance cassette, and an I-SceI restriction site and the flanking47 bp of viral sequences from nucleotides (nt) 68393 to 68439 locatedupstream of the KSHV orf45 start codon. The primer set used to amplifythis fragment was 5=-TGTGCTTTGTAAATTTCCGCCCCTAGCGGTCAACCCCGTACAAGGCCATGGTGAGCAAGGGCGAGGAGGATAAC-3= and 5=-ATTCTCTCTTCATCGTGTGTGCTAGACGAGGTCCTCACAAACATCGCCTTGTACAGCTCGTCCATGC-3= (the mCherry startmethionine codon is in boldface; the sequence homologous to that ofmCherry is underlined). The methylated plasmid template was then di-gested by DpnI, and the agarose-purified PCR product of 1,736 bp wassubsequently introduced by electroporation (0.1-cm cuvette, 1.8 kV, 200�, 25 �F) into the GS1783 E. coli strain harboring BAC16, while thetransient expression of gam, bet, and exo gene products required for ho-mologous recombination was induced by incubation at 42°C. Cells werethen plated on LB agar plates containing both chloramphenicol (17 �g/ml) and kanamycin (15 �g/ml). Integration of the mCherry/Kanr/I-SceIcassette was verified by PCR and restriction enzyme digestion of the pu-rified BAC16 DNA. Primers A and B located upstream of the orf45 gene(nt 68459 to 68440; 5=-GCAGTAAAGGCAATAACTCG-3=) and withinthe orf45 coding sequence (nt 67895 to 67912; 5=-AACCCATCCCATGGACGC-3=), respectively, were used to amplify a diagnostic DNA fragmentof 564 bp, whereas a DNA fragment of 2,256 bp was obtained, as pre-dicted, after insertion of the mCherry/Kanr/I-SceI cassette. The integratedcassette was then cleaved upon treatment with 1% L-arabinose; a secondrecombination event between the duplicated sequences resulted in theloss Kanr/I-SceI cassette and recircularization of the BAC16 DNA, yield-ing kanamycin-sensitive colonies that were screened by replica plating,followed by diagnostic PCR as well as restriction enzyme digestion of thepurified DNA. BAC DNA was purified by using a large-construct kit(Qiagen) according to the manufacturer’s instructions. BAC DNA wasdigested with HindIII and SnaB1 and run on a 0.4% agarose gel. Theinserted DNA fragment was sequenced to confirm the junction sequenceat the insertion site.
Transfection of KSHV BAC16 DNA and virus reconstitution. ForBAC16 transfection and reconstitution, �70% confluent HEK-293T cellswere grown in a 24-well plate followed by transfection with 500 ng of BACDNA via Lipofectamine 2000 transfection reagent (Life Technologies, In-vitrogen). Infected cells were selected in a medium which also contained200 �g/ml hygromycin B (A.G. Scientific, Inc.). Following selection ofHEK-293T cells that carry recombinant viruses, these cells were subcul-tured and mixed at a 1:1 ratio with iSLK-PURO cells. After 24 h, the lyticvirus cycle was induced by 20 ng/ml 12-O-tetradecanoylphorbol-13-ace-tate (TPA) and 1 mM sodium butyrate (Sigma). Selection medium con-taining 250 �g/ml G418, 1 �g/ml puromycin, and 1.2 mg/ml hygromycinB was added 4 days later, and iSLK-PURO cells infected with the recom-binant viruses were established. KSHV-infected iSLK-PURO cells weretreated with 1 �g/ml doxycycline and 1 mM sodium butyrate or 1 mMvalproic acid (Sigma), in the absence of hygromycin, puromycin, andG418, to induce RTA transgene expression and lytic cycle reactivation. Toprepare virus stocks, 4 days after lytic induction, supernatant was col-lected and cleared of cells and debris by centrifugation (700 � g for 10 minat 4°C) and filtration (0.45-�m cellulose acetate filters; Corning). Virusparticles were pelleted by centrifugation (40,000 � g for 2 h at 4°C).
Quantification of infectious virus production. Cells weretrypsinized, washed, and suspended in 0.5 ml phosphate-buffered saline
Bergson et al.
12840 jvi.asm.org Journal of Virology
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

(PBS); 0.5 ml 2% formaldehyde was added, and the cells were incubated at4°C for 20 min. The cells were then washed with PBS, suspended in 1 ml of70% ethanol, and incubated for 30 min at 4°C. Finally, cells were washedonce and suspended in PBS. GFP- and mCherry-positive cells were deter-mined using a fluorescence-activated cell sorter (FACS) (BD FACSAriaIII; BD Biosciences). Data analysis was performed with FlowJo software.This approach was used to measure virus reactivation and to quantitateinfectious virions. Various volumes of cell-free virus were used to infectSLK cells that were seeded at approximately 105 cells/well in 24-well plates24 h prior to infection. Cells were collected 48 h later and washed twicewith cold PBS. The titer for each inoculum size, assuming that one infec-tious particle generates a single GFP-positive cell, was calculated as a per-centage of the uninfected cells, according to Poisson’s law, and expressedas infectious units/ml.
Purification and quantification of genomic DNA from infected cellsand virions. High-molecular-weight (HMW) DNA was extracted using amammalian genomic DNA miniprep kit (Sigma). Purified cell-free virusfrom supernatants was treated with TurboDNase (Ambion, Inc.) for 1 h at37°C to eliminate nonencapsidated genomes. Virion DNA was extractedfollowing proteinase K treatment by using a DNeasy blood and tissue kit(Qiagen). KSHV DNA was quantitated using a previously described Taq-Man PCR employing primers and a probe targeting orf K6 (23, 24) withduplicates of 12 ng HMW DNA or 0.5, 1, and 5 �l from each virion DNApreparation. A second quantitative TaqMan-based real-time PCR, detect-ing human endogenous RNaseP (TaqMan human RNaseP control kit,Applied Biosystems), was used as a normalization tool for HMW and toconfirm DNA quality. The number of KSHV genomes in each sample wasdetermined by comparison of the obtained threshold cycle (CT) value tothe corresponding value of the standard curve. All PCRs were performedusing an Opticon2 real-time PCR (MJ Research, Inc., MA).
Antibodies and Western blot analysis. Cells were washed twice incold PBS, suspended in radioimmunoprecipitation assay (RIPA) lysis buf-fer, and incubated on ice for 30 min. Cell debris was removed by centrif-ugation at 12,000 � g for 15 min at 4°C. Virion proteins were extracted bysuspension of the viral particles in sodium-dodecyl sulfate (SDS) loadingbuffer followed by 10 min of boiling. Protein lysates were resolved bySDS-PAGE and transferred to nitrocellulose membranes using a Trans-Blot Turbo RTA Midi nitrocellulose transfer kit (Bio-Rad). The proteincontent of different samples was verified by Ponceau S staining. The ni-trocellulose membranes were blocked with 5% dry milk in Tris-bufferedsaline (TBS) and subsequently incubated with primary mouse antibodiesto GFP (Covance Research Products), mCherry (catalog no. 632496;Clontech), glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (SC-25778; Santa Cruz), ORF45 (17), ORF K8 (25) (kindly obtained from YanYuan), ORF65 (26) (kindly obtained from Shou Jiang Gao), or RTA (27)(kindly obtained from Keiji Ueda). Specific reactive bands were detectedusing anti-mouse conjugated to horseradish peroxidase. Immunoreactivebands were visualized using an EZ-enhanced chemiluminescence (ECL)detection kit (Biological Industries, Israel).
Fluorescence microscopy. Cells were seeded on coverslips and wereeither induced to undergo lytic reactivation or infected with purified vi-rions by spinoculation (1,500 � g, 1 h, 4°C). To reduce fluorescent back-ground, medium without phenol red was used. At the indicated timepoints, cells were washed with PBS and fixed by incubation in 4% form-aldehyde for 20 min at room temperature. The cells were then washedtwice in PBS and permeabilized in PBS containing 0.2% Triton X-100 and1% bovine serum albumin (BSA) at room temperature for 30 min. Cellswere probed with primary antibody (mouse anti-HA [Covance ResearchProducts], anti-ORF45, anti-ORF65, or anti-�-tubulin [Sigma]) at 4°C,and a conjugated secondary antibody (anti-mouse Alexa Fluor 488, anti-mouse rhodamine red, or anti-mouse Alexa Fluor 647 [Jackson Labora-tory]) was then applied for detection. To stain the nuclei, the cells wereincubated for 30 min with 0.05 �g/ml Hoechst dye (Sigma). For virionimaging, poly-L-lysine (Sigma)-coated coverslips were placed in a 24-wellplate and overlaid with purified virions, and the plates were centrifuged at
1,500 � g at 4°C for 1 h. Adhered virions were fixed with 4% formalde-hyde, permeabilized by treatment with PBS containing 0.1% Triton X-100and 1% BSA, and then probed with anti-ORF65 and stained with Hoechstdye. Cells and virions were examined and photographed under a confocallaser scanning microscope (Zeiss LSM 510 Meta). Cells for live imagingwere plated on 35-mm-diameter glass-bottom dishes and induced to un-dergo lytic reactivation. Twenty-four hours later, dishes were placed in ahumidified chamber supplied with 5% CO2 and heated to 37°C that wasmounted on a Zeiss Axio Observer.Z1 inverted fluorescence microscope.Cells were subsequently monitored for the appearance of mCherry, andindividual cells were chosen for serial photodocumentation over a periodof 26 h. Images at individual time points were taken, at an excitationwavelength of 582 nm and with a 595-nm emission filter with an exposureof 700 ms, by using an EC Plan-Neofluar 40� objective with a 0.75 nu-merical aperture and an AxioCam HSm digital camera controlled by ZeissAxioVision software. Differential interference contrast (DIC) images werealso collected.
RESULTSConstruction of a recombinant KSHV BAC16 expressing N-ter-minal mCherry-tagged ORF45. To construct a virus expressingthe mCherry protein fused to the 5= end of KSHV ORF45, we tookthe advantage of the complete KSHV BAC16 clone, which enablesgenetic manipulation of the KSHV genome in E. coli. As previ-ously described (21, 22) (see Materials and Methods), a BAC-based two-step Red-mediated recombination system was used.The final virus, designated BAC16-mCherry-ORF45, was testedfor the presence of the engineered insertion by diagnostic PCR andrestriction enzyme analysis, with HindIII and SnaBI revealingsimilar DNA fragmentation patterns, with the exceptions of the5,838- and 9,925-bp fragments predicted to undergo recombina-tion, respectively. BAC16-mCherry-ORF45 contained the ex-pected 6,546- and 10,633-bp restriction fragments compared tothe wild-type BAC16, respectively. No additional differences wereobserved between the BAC16 and BAC16-mCherry-ORF45 viralDNAs following restriction enzyme digestion (Fig. 1). To furtherconfirm that faithful recombination had occurred, we PCR am-plified and sequenced genomic DNA, including the boundary ofthe targeting sequence used for allelic exchange. These experi-ments established that the recombinant genome contains the cor-rect insertion, with no evident rearrangements.
The BAC16-mCherry-ORF45 exhibits functional propertiessimilar to those of wild-type BAC16. To reconstitute infectiousviruses from the recombinant BAC16-mCherry-ORF45 DNA,HEK-293T cells were transfected with BAC16 or BAC16-mCherry-ORF45 DNA, selected with hygromycin, and coculti-vated with iSLK following TPA and sodium butyrate treatment.All further experiments employed the iSLK cells, which permitefficient virus reactivation and virion production following doxy-cycline induction of RTA and sodium butyrate treatment (19, 28).To assess the replication of BAC16-mCherry-ORF45 in compari-son to that of the BAC16 wild type, we first induced the lytic cyclein infected iSLK cells. Proteins were extracted 24, 48, and 72 h afterinduction and analyzed by Western blotting. As shown in Fig. 2and in line with previous reports, expression of ORF45 was in-duced following lytic induction. ORF45 migrated at a molecularmass corresponding to �80 kDa in cell extracts harboring BAC16wild-type virus, while an �100-kDa band was detected for ex-tracts from cells infected with BAC16-mCherry-ORF45. Similarly,anti-mCherry detected a band of �100 kDa in extracts fromBAC16-mCherry-ORF45-infected cells, which was induced upon
Fluorescent Tagging of the KSHV ORF45 Tegument Protein
November 2014 Volume 88 Number 21 jvi.asm.org 12841
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

reactivation. No cleavage products that reacted with mCherry an-tibody were evident, thereby enabling the tracking of this fluores-cence as a marker for ORF45. Further incubation of this blot withantibodies to RTA, ORF K8, and GFP revealed similar levels ofexpression of these proteins following lytic induction, suggestingthat the efficiency of lytic reactivation of BAC16-mCherry-ORF45was similar to that of the BAC16 wild type. Moreover, quantitativereal-time PCR, to test for viral DNA replication, revealed similarincreases in KSHV DNA following induction of iSLK cells thatwere infected with either BAC16-mCherry-ORF45 or BAC16
(Fig. 2B). In addition, TaqMan PCR that quantitated the relativeamounts of encapsidated viral genomes found in pellets from su-pernatants that were collected from iSLK cells containing BAC16and BAC16-mCherry-ORF45 revealed similar KSHV genomecopy numbers. In agreement, we found that BAC16 and BAC16-mCherry-ORF45-infected iSLK cells produced similar numbersof infectious virions as measured by FACS analysis of the percent-age of GFP-positive cells (data not shown). Taken together, ouranalyses indicate that BAC16-mCherry-ORF45 replicates withsimilar kinetics and to the same titers as its parental BAC16 strain,
FIG 1 Construction and analysis of a recombinant BAC16 containing mCherry protein fusion to the orf45 gene. (A) Schematic illustration of the wild-typeKSHV genome encoding ORF45. The nucleotide positions (GenBank accession number GQ994935.1) are indicated above the diagram, as are the ORF45initiation methionine (ATG) and stop codon. Primers A and B, used to confirm the insertion of mCherry by PCR, are also shown. Genomic positions of HindIIIand SnaBI restriction sites are indicated. Below the wild-type genome (BAC16) is a schematic of the designed mCherry-tagged ORF45. (B) Agarose gelelectrophoresis of wild-type BAC16 and recombinant BAC16-mCherry-ORF45. DNAs were digested and resolved on a 0.4% agarose gel stained with ethidiumbromide. As predicted from HindIII and SnaB1 restriction patterns, the introduction of mCherry into BAC16 led to an increase in size from 5.8- and 9.9-kbp to6.5- and 10.6-kbp fragments, respectively. Altered bands are indicated by asterisks. Molecular weight markers (M) (in thousands) are shown on the left. (C)Sequence analysis of the recombinant BAC16-mCherry-tagged ORF45 at the orf45-mCherry junctions (top, orf45 upstream and mCherry N terminus sequence;bottom, mCherry C terminus and orf45 coding sequence missing the original initiation codon following recombination).
Bergson et al.
12842 jvi.asm.org Journal of Virology
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

indicating that the fluorescent tag does not interfere with the virallife cycle.
To determine whether mCherry-ORF45 is incorporated intoKSHV virions, extracellular virions were harvested from the cellsupernatant 96 h postinduction, and purified virions were lysed,resolved by SDS-PAGE, and analyzed by Western blotting with ananti-ORF45 monoclonal antibody (Fig. 3A). Eighty kilodaltonand �100-kDa proteins were detected in virion lysates fromBAC16 and BAC16-mCherry-ORF45, respectively, consistentwith the expected molecular masses of ORF45 and mCherry-ORF45. Polyclonal antibodies to the small capsid protein ORF65,used as a loading control, detected a protein of 18 kDa in bothvirion types. Quantitative analysis of the signals obtained with thetwo antibodies by using ImageJ revealed a similar ORF45/ORF65ratio for the two virion types, suggesting that the level of mCherry-ORF45 that is incorporated to virions is similar to that for theuntagged ORF45. In addition, virion particles were bound to aglass coverslip and stained for ORF65. mCherry-ORF45 fluores-cence was observed to overlap ORF65 and Hoechst staining inmost particles, thus revealing that mCherry-ORF45 is incorpo-rated into virions (Fig. 3B).
mCherry-ORF45 is first expressed in the nucleus and then inthe cytoplasm during virus reactivation. Previous studies exam-ined the cellular localization of ectopically expressed ORF45 inHEK-293T and BCBL-1 cells and found ORF45 to be expressedpredominantly in the cytoplasm (20, 29, 30). To determine thecellular localization of ORF45 in iSLK cells, we first introducedexpression plasmids for GFP- or HA-tagged ORF45 into cells. Asshown in Fig. 4, transiently expressed GFP- or HA-tagged ORF45demonstrated nuclear localization, or localization to both the nu-cleus and cytoplasm. A similar cellular distribution of ORF45 wasevident in iSLK cells that were treated with doxycycline to inducethe expression of RTA, suggesting that RTA does not affect thecellular distribution of ORF45.
To determine the cellular localization of ORF45 following vi-rus reactivation, we induced lytic reactivation in BAC16-mCherry-ORF45-infected iSLK cells by treatment with doxycy-cline and sodium butyrate. Cells were fixed 24, 48, and 72 hpostinduction, and the localization of ORF45 was examined bytracking the expression of mCherry. As shown in Fig. 5A, at 24 hfollowing induction, mCherry-ORF45 was localized predomi-nantly to the nucleus with a dot-like pattern. At 48 h postinduc-
FIG 2 Comparison of the levels of KSHV gene expression in iSLK cells infected with BAC16 and BAC16-mCherry-ORF45. (A) iSLK cells harboring BAC16 andBAC16-mCherryORF45 were induced to undergo lytic virus reactivation using sodium butyrate and doxycycline. At 0, 24, 48, and 72 h following induction,whole-cell lysates were prepared, and protein expression of viral genes was analyzed by immunoblotting with the indicated antibodies against viral proteins aswell as GFP. The same blot was also reprobed with anti-GAPDH as a loading control. (B) High-molecular-weight DNA was extracted at 0, 24, 48, and 72 hpostinduction, and the copy number of KSHV genomes relative to cellular DNA was determined by quantitative TaqMan PCR. All values were normalizedrelative to noninduced BAC16-infected cells that were set as 1. The KSHV DNA level was normalized to that of purified host cellular DNA.
Fluorescent Tagging of the KSHV ORF45 Tegument Protein
November 2014 Volume 88 Number 21 jvi.asm.org 12843
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

tion, mCherry-ORF45 appeared as fluorescent puncta that werelocalized in the nuclear margins, possibly inside or near the nu-clear envelope, and within a structure adjacent to the nucleuswhich probably corresponds to the trans-Golgi network. At 72 h
postinduction, mCherry-ORF45 appeared as punctum structureswithin the cytoplasm and at the cell periphery. The pattern ofmCherry expression matched the fluorescent staining that wasobtained with anti-ORF45, indicating that the localization of
FIG 3 Extracellular progeny of BAC16-mCherry-ORF45 virions contain recombinant mCherry-ORF45. (A) Extracellular progeny KSHV virions were collectedin pellets and then dissolved in SDS-loading buffer and analyzed by Western blotting with antibodies to KSHV virion proteins, as indicated. (B) Visualization offluorescent virion particles. BAC16-mCherry-ORF45 virions were bound to a glass coverslip, fixed, and immunostained with anti-ORF65 followed by anti-mouse Alexa Fluor 647-conjugated secondary antibodies. The corresponding staining of virion DNA by Hoechst is also displayed. White arrowheads highlightoverlapping signals and colocalization of mCherry-ORF45, ORF65, and Hoechst staining.
FIG 4 Ectopically expressed HA- and GFP-tagged ORF45 demonstrate both nuclear and cytoplasmic localization. iSLK cells were transfected with plasmidsencoding EGFP-ORF45 or HA-ORF45. RTA expression was induced 24 h posttransfection, and cells were fixed 24 h later. HA-transfected cells were stained withmouse monoclonal anti-HA, while Alexa Fluor 488-conjugated anti-mouse IgG was used as a secondary antibody. The corresponding staining of nuclear DNAby Hoechst, and differential interference contrast (DIC) images are also displayed. �RTA, no expression of RTA; �RTA, induction of RTA expression.
Bergson et al.
12844 jvi.asm.org Journal of Virology
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

FIG 5 Cellular distribution of mCherry-ORF45 and ORF45 in iSLK cells at different time points following lytic induction. (A) BAC16-mCherryORF45-infectediSLK cells were induced to undergo lytic virus reactivation with sodium butyrate and doxycycline. Cells were fixed at the indicated time points postinduction andstained with mouse monoclonal anti-ORF45 followed by anti-mouse Alexa Fluor 647-conjugated secondary antibodies. Direct visualization of mCherry-ORF45is shown on the top row. The corresponding staining of nuclear DNA by Hoechst and DIC images are also displayed. (B) Cellular distribution of ORF45 in iSLKcells at different time points postinduction. BAC16-infected iSLK cells were induced and stained with anti-ORF45 as described for panel A followed byanti-mouse rhodamine red-conjugated secondary antibodies.
Fluorescent Tagging of the KSHV ORF45 Tegument Protein
November 2014 Volume 88 Number 21 jvi.asm.org 12845
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
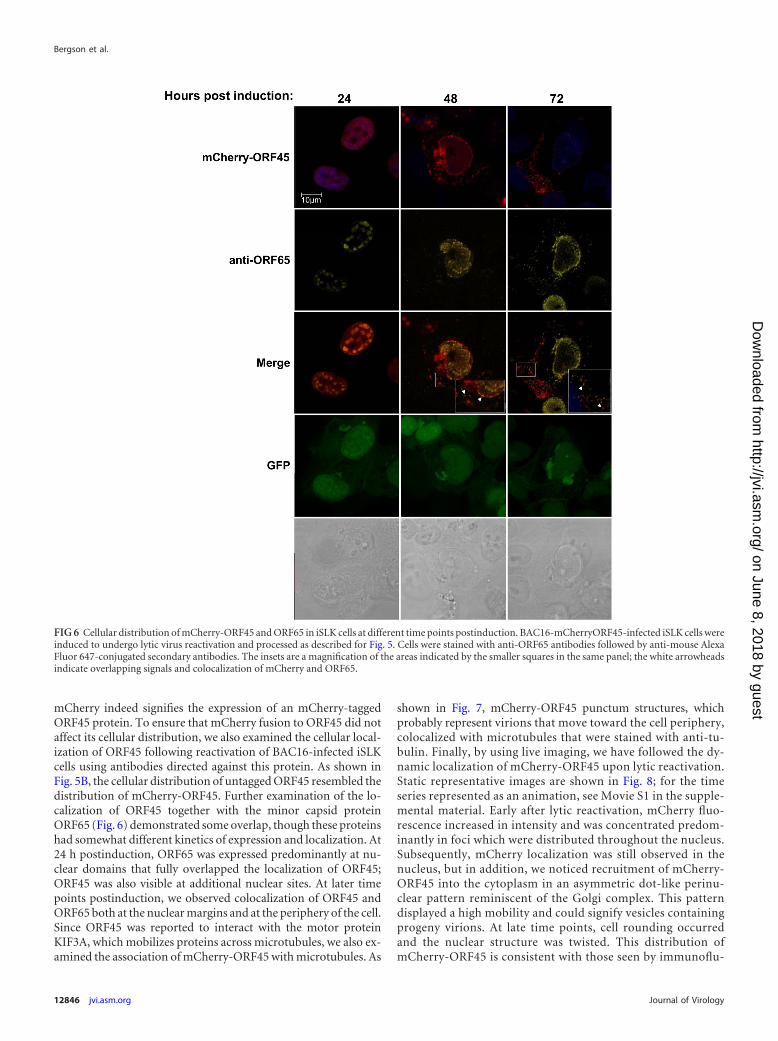
mCherry indeed signifies the expression of an mCherry-taggedORF45 protein. To ensure that mCherry fusion to ORF45 did notaffect its cellular distribution, we also examined the cellular local-ization of ORF45 following reactivation of BAC16-infected iSLKcells using antibodies directed against this protein. As shown inFig. 5B, the cellular distribution of untagged ORF45 resembled thedistribution of mCherry-ORF45. Further examination of the lo-calization of ORF45 together with the minor capsid proteinORF65 (Fig. 6) demonstrated some overlap, though these proteinshad somewhat different kinetics of expression and localization. At24 h postinduction, ORF65 was expressed predominantly at nu-clear domains that fully overlapped the localization of ORF45;ORF45 was also visible at additional nuclear sites. At later timepoints postinduction, we observed colocalization of ORF45 andORF65 both at the nuclear margins and at the periphery of the cell.Since ORF45 was reported to interact with the motor proteinKIF3A, which mobilizes proteins across microtubules, we also ex-amined the association of mCherry-ORF45 with microtubules. As
shown in Fig. 7, mCherry-ORF45 punctum structures, whichprobably represent virions that move toward the cell periphery,colocalized with microtubules that were stained with anti-tu-bulin. Finally, by using live imaging, we have followed the dy-namic localization of mCherry-ORF45 upon lytic reactivation.Static representative images are shown in Fig. 8; for the timeseries represented as an animation, see Movie S1 in the supple-mental material. Early after lytic reactivation, mCherry fluo-rescence increased in intensity and was concentrated predom-inantly in foci which were distributed throughout the nucleus.Subsequently, mCherry localization was still observed in thenucleus, but in addition, we noticed recruitment of mCherry-ORF45 into the cytoplasm in an asymmetric dot-like perinu-clear pattern reminiscent of the Golgi complex. This patterndisplayed a high mobility and could signify vesicles containingprogeny virions. At late time points, cell rounding occurredand the nuclear structure was twisted. This distribution ofmCherry-ORF45 is consistent with those seen by immunoflu-
FIG 6 Cellular distribution of mCherry-ORF45 and ORF65 in iSLK cells at different time points postinduction. BAC16-mCherryORF45-infected iSLK cells wereinduced to undergo lytic virus reactivation and processed as described for Fig. 5. Cells were stained with anti-ORF65 antibodies followed by anti-mouse AlexaFluor 647-conjugated secondary antibodies. The insets are a magnification of the areas indicated by the smaller squares in the same panel; the white arrowheadsindicate overlapping signals and colocalization of mCherry and ORF65.
Bergson et al.
12846 jvi.asm.org Journal of Virology
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

orescence examination of fixed cells infected with the BAC16wild type and BAC16-mCherry-ORF45.
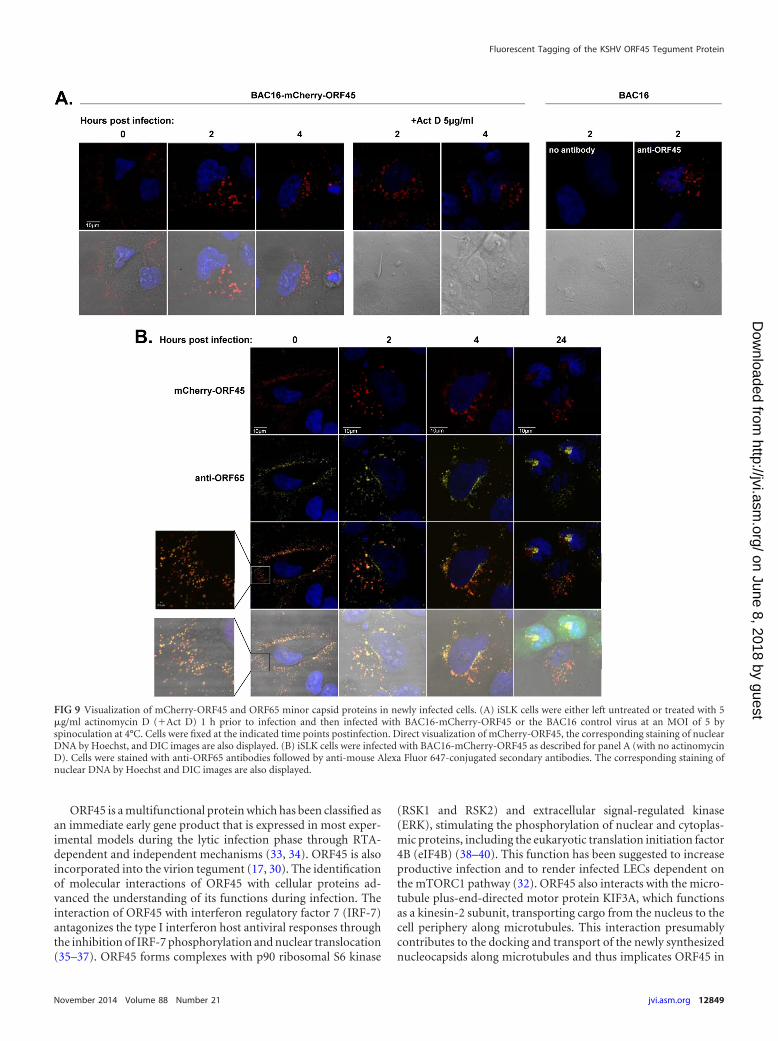
Cellular localization of mCherry-ORF45 following infec-tion. To determine the cellular localization of ORF45 at differenttime points postinfection, we infected iSLK cells with BAC16-mCherry-ORF45 at a multiplicity of infection (MOI) of 5 by spi-noculation at 4°C and fixed the cells at 0, 2, 4, and 24 h postinfec-tion. The localization of ORF45 was examined by tracking theexpression of mCherry while the minor capsid protein ORF65 wasidentified with anti-ORF65. To control for incoming virion-asso-ciated mCherry-ORF45 and potential de novo expression ofmCherry-ORF45, target cells were first treated with 5 �g/ml acti-nomycin D 1 h prior to infection and during the infection course.The specificity of mCherry fluorescence was also confirmed incells that were infected with BAC16. As shown in Fig. 9A, a similardistribution of mCherry was evident in infected cells that wereeither left untreated or treated with actinomycin D, thus indicat-ing the virion source of mCherry fluorescence. No fluorescentsignal was evident when cells were infected with the control virusBAC16, indicating the specificity of the mCherry fluorescence. Asshown in Fig. 9B, extracellular virions, identified by anti-ORF65staining and mCherry-ORF45 fluorescence, appeared as discretefluorescent puncta and were mostly localized adjacent to the cellsurface immediately postspinoculation. Of note, although virusinfection was carried at an MOI of 5, numerous particles sur-rounded the cells, thus emphasizing the high ratio of particles toinfectious units for the KSHV stocks (19). At 2 h postinfection,fluorescent puncta were detected in the cytosol and at the nuclearrim. The majority of ORF65-stained capsids in the cytosol con-tained mCherry-ORF45, whereas capsids that reached the nuclearrim lacked mCherry fluorescence, suggesting the dissociation ofORF45 from incoming virions. By 4 h postinfection, most capsidscould be seen in the cytosol or at the nuclear rim. At 24 h postin-fection, faint mCherry puncta, independent of ORF65, were de-
tected in the cytosol, suggesting possible degradation of freemCherry-ORF45. These findings provide evidence for the associ-ation of ORF45 tegument protein with incoming capsids in thecytosol and its dissociation as the capsids move toward the nu-cleus.
Evaluation of KSHV lytic reactivation levels by quantifica-tion of mCherry-positive cells. To demonstrate the ability ofmCherry-ORF45 to identify lytic infection and to be used as amarker for this phase, BAC16-mCherryORF45-infected iSLK cellswere treated with doxycycline alone, or combined with TPA, so-dium butyrate, or valproic acid, for 24 and 48 h. Cells were col-lected, and the percentage of GFP- and mCherry-positive cells wasdetermined by FACS analysis. As shown in Fig. 10 and in line withprevious results (19), under standard growth conditions, almostno mCherry-expressing cells were observed. Roughly 11% of cellsexpressed mCherry 24 and 48 h after induction of RTA expressionby treatment with doxycycline. The effect of doxycycline was fur-ther enhanced when combined with chemical inducers, as therewere up to 87% mCherry-positive cells 48 h after combined treat-ment with doxycycline and sodium butyrate. These results are inagreement with previous results in which the red fluorescent pro-tein (RFP) gene from the PAN RNA promoter was used as amarker for the inducibility of iSLK cells (19).
DISCUSSION
In most experimental cell cultures, KSHV infection results in theestablishment of a latent infection in which infected cells cannotbe readily distinguished from uninfected ones. Furthermore, cellsundergoing lytic reactivation cannot be distinguished based ontheir morphology, in particular during the early phases. Recom-binant viruses that express fluorescent proteins commonly serveas important tools to track viral infection and to elucidate theintracellular trafficking of virions and subviral particles. Indeed, arecombinant dual-color KSHV (rKSHV.219), designed to consti-
FIG 7 Colocalization of mCherry-ORF45 with microtubules. BAC16-mCherryORF45-infected iSLK cells were induced to undergo lytic reactivation andprocessed as described for Fig. 5. Cells were stained with mouse monoclonal anti-tubulin followed by anti-mouse Alexa Fluor 647-conjugated secondaryantibodies. White arrowheads indicate overlapping signals and colocalization of mCherry and tubulin.
Fluorescent Tagging of the KSHV ORF45 Tegument Protein
November 2014 Volume 88 Number 21 jvi.asm.org 12847
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

tutively express GFP as a marker of infection and RFP under thecontrol of the strong early lytic promoter PAN, enables the iden-tification of KSHV-infected cells and the discrimination of cellsundergoing lytic infection by using fluorescence microscopy orFACS analysis (31). BAC16, a full-length KSHV genome BAC-containing clone, was derived from rKSHV.219 by replacing theGFP and RFP cassettes; this construct can be stably propagatedin bacteria and produces infectious virions upon reconstitution inmammalian cells (21). This clone constitutively expresses GFP inmammalian cells. Here, we report the construction of a recombi-nant dual-color KSHV genome, which constitutively expressesGFP and contains the fluorescent protein mCherry that tags thetegument protein ORF45.
Several assays were used to ensure that the BAC16-mCherry-ORF45 was properly expressed. First, the presence of the fusion pro-tein in lysates from cells that were induced into lytic phase was as-sessed by Western blot analysis using an antibody directed againstmCherry. Second, structural incorporation of the mCherry-ORF45fusion to KSHV virions was confirmed by Western blot analysis and
confocal fluorescent imaging. BAC16-mCherry-ORF45 DNA estab-lished a latent infection in iSLK cells, expressed the viral lytic proteinsupon reactivation, and produced infectious virions at a level similarto that of its parental BAC16 clone. Furthermore, the cellular local-ization of mCherry-ORF45 fully matched that of the authentic un-tagged ORF45, and an absolute overlap between the mCherry fluo-rescence and the anti-ORF45 fluorescent signal was documented.Therefore, our results suggest that BAC16-mCherry-ORF45 can beused to monitor ORF45 during infection and to follow infected cellsthat undergo lytic infection with KSHV. As such, the expression ofGFP marks infected cells, while induction of early viral lytic geneexpression can be followed by mCherry expression. However, thisapproach cannot be implemented for all KSHV-infected cell types,and the classification of ORF45 as a lytic gene product must be con-firmed in each experimental setting. In fact, mTORC1 activation andrapamycin sensitivity of KSHV-infected primary dermal and lunghuman lymphatic endothelial cells (LECs) was recently demon-strated and attributed to the latent expression of ORF45 in these cells(32).
FIG 8 Time-lapse microscopy of live iSLK cells following lytic induction. BAC16-mCherryORF45-infected iSLK cells seeded on a glass-bottom dish wereinduced to undergo lytic virus reactivation with sodium butyrate and doxycycline and transferred to a heated chamber at 24 h postinduction. Images werecollected every 20 min. Patterns representing nuclear accumulation, cytoplasmic dots, and morphological changes are shown. For the time series represented asan animation, see Movie S1 in the supplemental material.
Bergson et al.
12848 jvi.asm.org Journal of Virology
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
ORF45 is a multifunctional protein which has been classified asan immediate early gene product that is expressed in most exper-imental models during the lytic infection phase through RTA-dependent and independent mechanisms (33, 34). ORF45 is alsoincorporated into the virion tegument (17, 30). The identificationof molecular interactions of ORF45 with cellular proteins ad-vanced the understanding of its functions during infection. Theinteraction of ORF45 with interferon regulatory factor 7 (IRF-7)antagonizes the type I interferon host antiviral responses throughthe inhibition of IRF-7 phosphorylation and nuclear translocation(35–37). ORF45 forms complexes with p90 ribosomal S6 kinase
(RSK1 and RSK2) and extracellular signal-regulated kinase(ERK), stimulating the phosphorylation of nuclear and cytoplas-mic proteins, including the eukaryotic translation initiation factor4B (eIF4B) (38–40). This function has been suggested to increaseproductive infection and to render infected LECs dependent onthe mTORC1 pathway (32). ORF45 also interacts with the micro-tubule plus-end-directed motor protein KIF3A, which functionsas a kinesin-2 subunit, transporting cargo from the nucleus to thecell periphery along microtubules. This interaction presumablycontributes to the docking and transport of the newly synthesizednucleocapsids along microtubules and thus implicates ORF45 in
FIG 9 Visualization of mCherry-ORF45 and ORF65 minor capsid proteins in newly infected cells. (A) iSLK cells were either left untreated or treated with 5�g/ml actinomycin D (�Act D) 1 h prior to infection and then infected with BAC16-mCherry-ORF45 or the BAC16 control virus at an MOI of 5 byspinoculation at 4°C. Cells were fixed at the indicated time points postinfection. Direct visualization of mCherry-ORF45, the corresponding staining of nuclearDNA by Hoechst, and DIC images are also displayed. (B) iSLK cells were infected with BAC16-mCherry-ORF45 as described for panel A (with no actinomycinD). Cells were stained with anti-ORF65 antibodies followed by anti-mouse Alexa Fluor 647-conjugated secondary antibodies. The corresponding staining ofnuclear DNA by Hoechst and DIC images are also displayed.
Fluorescent Tagging of the KSHV ORF45 Tegument Protein
November 2014 Volume 88 Number 21 jvi.asm.org 12849
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

viral egress (41). The above functions are most likely exerted invarious cellular compartments and appear to be important forefficient productive infection, for establishment of latency, and inmediating robust lytic reactivation.
Our study found mCherry-ORF45 predominantly in the nu-cleus at early time points post-lytic reactivation, with a small frac-tion of the protein present in the cytoplasm. At later time points,mCherry-ORF45 was localized to the periphery of the nucleus andclose to the cell plasma membrane, as well as to a structure near thenucleus, which probably represents the trans-Golgi network. Theauthenticity of this cellular distribution was confirmed by usingimmunofluorescent staining with anti-ORF45 antibodies. We andothers reported previously that KSHV ORF45 is found predomi-nantly in the cytoplasm (20, 29, 35); however, accumulation ofORF45 in the nucleus after inhibition of CRM1-dependent nu-clear export and shuttling of ORF45 between the nucleus and thecytoplasm during infection were previously reported as well (42).Nuclear localization of ORF45, which was documented with aviral replication compartment, in primary effusion lymphomacells has been reported (40). Interestingly, recombinant viruses inwhich ORF45 is restricted to the nucleus behave normally,whereas recombinant KSHV in which ORF45 is restricted to thecytoplasm behaves similarly to ORF45-null mutant viruses, sug-gesting that ORF45 has an essential nuclear function (42). Giventhe nuclear localization of ORF45 homologs encoded by other
members of the Gammaherpesvirinae, they are likely to share com-mon functions in the nucleus. The present study demonstratesdynamic localization of ORF45 during lytic reactivation. In con-trast, ectopically expressed protein maintained a steady cellularlocalization that was not affected by the expression of RTA. Theseresults highlight the advantage of studying viral proteins withinthe context of infection. The dynamics are likely to reflect thevarious functions of ORF45 and possibly its unique protein inter-actions and posttranslational modifications during the differentphases of infection.
In summary, this study presents the assembly and initial char-acterization of a new dual-color fluorescently labeled KSHV virus,which can be used to track cells that undergo lytic infection. Aswas already done for other viruses (43), the BAC16-mCherry-ORF45 recombinant virus can also be used to address issues re-lated to the trafficking and maturation pathways of KSHV virionsfollowing de novo infection or during reactivation. Specifically, itmay allow the identification of the phases of infection at whichORF45 assembles or disassembles and thus may confirm its can-didacy as an effector of capsid transport during ingress and egress.Furthermore, as shown in the present study, this virus can serve asa platform for digital time-lapse microscopy experiments withliving cells. These experiments are expected to provide insightsinto the dynamics of intracellular transport of KSHV capsids andtheir tegumentation and detegumentation, thereby highlighting
FIG 10 Evaluation of KSHV lytic reactivation levels by quantification of mCherry-positive cells. (A) BAC16-mCherryORF45-infected iSLK cells were leftuninduced (SLK) or induced by 1 �g/ml doxycycline alone (Dox) or in combination with either TPA (Dox�TPA; 20 ng/ml), sodium butyrate (Dox�SB; 1 mM),or valproic acid (Dox�Val; 1 mM) for the indicated duration. GFP and mCherry expression was analyzed by flow cytometry. The x axis shows GFP, and the yaxis indicates the mCherry signal. (B) Graph showing the percentage of GFP and GFP�mCherry-positive cells at 24 and 48 h post-lytic induction.
Bergson et al.
12850 jvi.asm.org Journal of Virology
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

key pathways that are activated during the various phases of KSHVinfection.
ACKNOWLEDGMENTS
We gratefully acknowledge Don Ganem, Rolf Renne, Nikolaus Osterrie-der, Yan Yuan, Shou Jiang Gao, and Keiji Ueda for reagents and ThomasSchulz for his help in the implementation of the BAC system. We alsothank the Bar Ilan Faculty of Life Sciences Scientific Equipment Centerstaff for excellent technical assistance.
This work was supported by a grant from the Israel Science Founda-tion (grant no. 709/10).
REFERENCES1. Mesri EA, Cesarman E, Boshoff C. 2010. Kaposi’s sarcoma and its asso-
ciated herpesvirus. Nat. Rev. Cancer 10:707–719. http://dx.doi.org/10.1038/nrc2888.
2. Kalt I, Masa SR, Sarid R. 2009. Linking the Kaposi’s sarcoma-associatedherpesvirus (KSHV/HHV-8) to human malignancies. Methods Mol. Biol.471:387– 407. http://dx.doi.org/10.1007/978-1-59745-416-2_19.
3. Ganem D. 2010. KSHV and the pathogenesis of Kaposi sarcoma: listeningto human biology and medicine. J. Clin. Invest. 120:939 –949. http://dx.doi.org/10.1172/JCI40567.
4. Chandran B. 2010. Early events in Kaposi’s sarcoma-associated herpesvi-rus infection of target cells. J. Virol. 84:2188 –2199. http://dx.doi.org/10.1128/JVI.01334-09.
5. Krishnan HH, Naranatt PP, Smith MS, Zeng L, Bloomer C, ChandranB. 2004. Concurrent expression of latent and a limited number of lyticgenes with immune modulation and antiapoptotic function by Kaposi’ssarcoma-associated herpesvirus early during infection of primary endo-thelial and fibroblast cells and subsequent decline of lytic gene expression.J. Virol. 78:3601–3620. http://dx.doi.org/10.1128/JVI.78.7.3601-3620.2004.
6. Miller G, El-Guindy A, Countryman J, Ye J, Gradoville L. 2007. Lyticcycle switches of oncogenic human gammaherpesviruses. Adv. CancerRes. 97:81–109. http://dx.doi.org/10.1016/S0065-230X(06)97004-3.
7. Deng H, Liang Y, Sun R. 2007. Regulation of KSHV lytic gene expression.Curr. Top. Microbiol. Immunol. 312:157–183.
8. Staudt MR, Dittmer DP. 2007. The Rta/Orf50 transactivator proteins ofthe gamma-herpesviridae. Curr. Top. Microbiol. Immunol. 312:71–100.
9. Johnson DC, Baines JD. 2011. Herpesviruses remodel host membranesfor virus egress. Nat. Rev. Microbiol. 9:382–394. http://dx.doi.org/10.1038/nrmicro2559.
10. Kelly BJ, Fraefel C, Cunningham AL, Diefenbach RJ. 2009. Functionalroles of the tegument proteins of herpes simplex virus type 1. Virus Res.145:173–186. http://dx.doi.org/10.1016/j.virusres.2009.07.007.
11. Mettenleiter TC, Muller F, Granzow H, Klupp BG. 2013. The way out:what we know and do not know about herpesvirus nuclear egress. Cell.Microbiol. 15:170 –178. http://dx.doi.org/10.1111/cmi.12044.
12. Diefenbach RJ, Miranda-Saksena M, Douglas MW, Cunningham AL.2008. Transport and egress of herpes simplex virus in neurons. Rev. Med.Virol. 18:35–51. http://dx.doi.org/10.1002/rmv.560.
13. Sathish N, Wang X, Yuan Y. 2012. Tegument proteins of Kaposi’s sar-coma-associated herpesvirus and related gamma-herpesviruses. Front.Microbiol. 3:98. http://dx.doi.org/10.3389/fmicb.2012.00098.
14. Guo H, Wang L, Peng L, Zhou ZH, Deng H. 2009. Open reading frame33 of a gammaherpesvirus encodes a tegument protein essential for virionmorphogenesis and egress. J. Virol. 83:10582–10595. http://dx.doi.org/10.1128/JVI.00497-09.
15. Shen S, Guo H, Deng H. 2014. Murine gammaherpesvirus-68 ORF38encodes a tegument protein and is packaged into virions during secondaryenvelopment. Protein Cell 5:141–150. http://dx.doi.org/10.1007/s13238-013-0005-0.
16. Desai PJ, Pryce EN, Henson BW, Luitweiler EM, Cothran J. 2012.Reconstitution of the Kaposi’s sarcoma-associated herpesvirus nuclearegress complex and formation of nuclear membrane vesicles by coexpres-sion of ORF67 and ORF69 gene products. J. Virol. 86:594 –598. http://dx.doi.org/10.1128/JVI.05988-11.
17. Zhu FX, Chong JM, Wu L, Yuan Y. 2005. Virion proteins of Kaposi’ssarcoma-associated herpesvirus. J. Virol. 79:800 – 811. http://dx.doi.org/10.1128/JVI.79.2.800-811.2005.
18. Zhu FX, Li X, Zhou F, Gao SJ, Yuan Y. 2006. Functional characteriza-
tion of Kaposi’s sarcoma-associated herpesvirus ORF45 by BAC-basedmutagenesis. J. Virol. 80:12187–12196. http://dx.doi.org/10.1128/JVI.01275-06.
19. Myoung J, Ganem D. 2011. Generation of a doxycycline-inducible KSHVproducer cell line of endothelial origin: maintenance of tight latency withefficient reactivation upon induction. J. Virol. Methods 174:12–21. http://dx.doi.org/10.1016/j.jviromet.2011.03.012.
20. Abada R, Dreyfuss-Grossman T, Herman-Bachinsky Y, Geva H, MasaSR, Sarid R. 2008. SIAH-1 interacts with the Kaposi’s sarcoma-associatedherpesvirus-encoded ORF45 protein and promotes its ubiquitylation andproteasomal degradation. J. Virol. 82:2230 –2240. http://dx.doi.org/10.1128/JVI.02285-07.
21. Brulois KF, Chang H, Lee AS, Ensser A, Wong LY, Toth Z, Lee SH, LeeHR, Myoung J, Ganem D, Oh TK, Kim JF, Gao SJ, Jung JU. 2012.Construction and manipulation of a new Kaposi’s sarcoma-associatedherpesvirus bacterial artificial chromosome clone. J. Virol. 86:9708 –9720.http://dx.doi.org/10.1128/JVI.01019-12.
22. Tischer BK, von Einem J, Kaufer B, Osterrieder N. 2006. Two-stepred-mediated recombination for versatile high-efficiency markerless DNAmanipulation in Escherichia coli. Biotechniques 40:191–197. http://dx.doi.org/10.2144/000112096.
23. Guttman-Yassky E, Abada R, Kra-Oz Z, Sattinger J, Perelman A,Bergman R, Sarid R. 2007. Relationship between human herpesvirus 8loads and disease stage in classic Kaposi sarcoma patients. Diagn. Micro-biol. Infect. Dis. 57:387–392. http://dx.doi.org/10.1016/j.diagmicrobio.2006.09.012.
24. de Sanjosé S, Marshall V, Sola J, Palacio V, Almirall R, Goedert JJ,Bosch FX, Whitby D. 2002. Prevalence of Kaposi’s sarcoma-associatedherpesvirus infection in sex workers and women from the general popu-lation in Spain. Int. J. Cancer 98:155–158. http://dx.doi.org/10.1002/ijc.10190.
25. Wang Y, Li H, Tang Q, Maul GG, Yuan Y. 2008. Kaposi’s sarcoma-associated herpesvirus ori-Lyt-dependent DNA replication: involvementof host cellular factors. J. Virol. 82:2867–2882. http://dx.doi.org/10.1128/JVI.01319-07.
26. Ye F, Zhou F, Bedolla RG, Jones T, Lei X, Kang T, Guadalupe M, GaoSJ. 2011. Reactive oxygen species hydrogen peroxide mediates Kaposi’ssarcoma-associated herpesvirus reactivation from latency. PLoS Pathog.7:e1002054. http://dx.doi.org/10.1371/journal.ppat.1002054.
27. Ueda K, Ishikawa K, Nishimura K, Sakakibara S, Do E, Yamanishi K.2002. Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8)replication and transcription factor activates the K9 (vIRF) gene throughtwo distinct cis elements by a non-DNA-binding mechanism. J. Virol.76:12044 –12054. http://dx.doi.org/10.1128/JVI.76.23.12044-12054.2002.
28. Stürzl M, Gaus D, Dirks WG, Ganem D, Jochmann R. 2013. Kaposi’ssarcoma-derived cell line SLK is not of endothelial origin, but is a contam-inant from a known renal carcinoma cell line. Int. J. Cancer 132:1954 –1958. http://dx.doi.org/10.1002/ijc.27849.
29. Sander G, Konrad A, Thurau M, Wies E, Leubert R, Kremmer E, DinkelH, Schulz T, Neipel F, Sturzl M. 2008. Intracellular localization map ofhuman herpesvirus 8 proteins. J. Virol. 82:1908 –1922. http://dx.doi.org/10.1128/JVI.01716-07.
30. Zhu FX, Yuan Y. 2003. The ORF45 protein of Kaposi’s sarcoma-associated herpesvirus is associated with purified virions. J. Virol. 77:4221– 4230. http://dx.doi.org/10.1128/JVI.77.7.4221-4230.2003.
31. Vieira J, O’Hearn PM. 2004. Use of the red fluorescent protein as amarker of Kaposi’s sarcoma-associated herpesvirus lytic gene expression.Virology 325:225–240. http://dx.doi.org/10.1016/j.virol.2004.03.049.
32. Chang HH, Ganem D. 2013. A unique herpesviral transcriptional pro-gram in KSHV-infected lymphatic endothelial cells leads to mTORC1activation and rapamycin sensitivity. Cell Host Microbe 13:429 – 440.http://dx.doi.org/10.1016/j.chom.2013.03.009.
33. Zhu FX, Cusano T, Yuan Y. 1999. Identification of the immediate-earlytranscripts of Kaposi’s sarcoma-associated herpesvirus. J. Virol. 73:5556 –5567.
34. Chang PJ, Wang SS, Chen LY, Hung CH, Huang HY, Shih YJ, Yen JB,Liou JY, Chen LW. 2013. ORF50-dependent and ORF50-independentactivation of the ORF45 gene of Kaposi’s sarcoma-associated herpesvirus.Virology 442:38 –50. http://dx.doi.org/10.1016/j.virol.2013.03.023.
35. Zhu FX, King SM, Smith EJ, Levy DE, Yuan Y. 2002. A Kaposi’ssarcoma-associated herpesviral protein inhibits virus-mediated inductionof type I interferon by blocking IRF-7 phosphorylation and nuclear accu-
Fluorescent Tagging of the KSHV ORF45 Tegument Protein
November 2014 Volume 88 Number 21 jvi.asm.org 12851
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

mulation. Proc. Natl. Acad. Sci. U. S. A. 99:5573–5578. http://dx.doi.org/10.1073/pnas.082420599.
36. Zhu FX, Sathish N, Yuan Y. 2010. Antagonism of host antiviral responsesby Kaposi’s sarcoma-associated herpesvirus tegument protein ORF45.PLoS One 5:e10573. http://dx.doi.org/10.1371/journal.pone.0010573.
37. Sathish N, Yuan Y. 2011. Evasion and subversion of interferon-mediatedantiviral immunity by Kaposi’s sarcoma-associated herpesvirus: an over-view. J. Virol. 85:10934 –10944. http://dx.doi.org/10.1128/JVI.00687-11.
38. Kuang E, Wu F, Zhu F. 2009. Mechanism of sustained activation ofribosomal S6 kinase (RSK) and ERK by Kaposi sarcoma-associated her-pesvirus ORF45: multiprotein complexes retain active phosphorylatedERK AND RSK and protect them from dephosphorylation. J. Biol. Chem.284:13958 –13968. http://dx.doi.org/10.1074/jbc.M900025200.
39. Kuang E, Fu B, Liang Q, Myoung J, Zhu F. 2011. Phosphorylation ofeukaryotic translation initiation factor 4B (EIF4B) by open reading frame45/p90 ribosomal S6 kinase (ORF45/RSK) signaling axis facilitates proteintranslation during Kaposi sarcoma-associated herpesvirus (KSHV) lytic
replication. J. Biol. Chem. 286:41171– 41182. http://dx.doi.org/10.1074/jbc.M111.280982.
40. Kuang E, Tang Q, Maul GG, Zhu F. 2008. Activation of p90 ribosomalS6 kinase by ORF45 of Kaposi’s sarcoma-associated herpesvirus and itsrole in viral lytic replication. J. Virol. 82:1838 –1850. http://dx.doi.org/10.1128/JVI.02119-07.
41. Sathish N, Zhu FX, Yuan Y. 2009. Kaposi’s sarcoma-associated herpes-virus ORF45 interacts with kinesin-2 transporting viral capsid-tegumentcomplexes along microtubules. PLoS Pathog. 5:e1000332. http://dx.doi.org/10.1371/journal.ppat.1000332.
42. Li X, Zhu F. 2009. Identification of the nuclear export and adjacentnuclear localization signals for ORF45 of Kaposi’s sarcoma-associatedherpesvirus. J. Virol. 83:2531–2539. http://dx.doi.org/10.1128/JVI.02209-08.
43. Brandenburg B, Zhuang X. 2007. Virus trafficking: learning from single-virus tracking. Nat. Rev. Microbiol. 5:197–208. http://dx.doi.org/10.1038/nrmicro1615.
Bergson et al.
12852 jvi.asm.org Journal of Virology
on June 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from


















