Fish diversity of the Bear Gulch Limestone, Namurian, Lower Carboniferous of Montana, USA
11
RICHARD L U N D & CI~CILE P O P L I N FISH DIVERSITY OF THE BEAR GULCH LIMESTONE, NAMURIAN, LOWER CARBONIFEROUS OF MONTANA, USA LUND R. & POPLIN C. 1999. Fish diversity of the Bear Gulch Limestone, Namurian, Lower Carboniferous of Montana, USA. [La diversit~ des poissons de Bear Gulch, Namurien, Carbonifere inf~rieur du Montana, USA]. GEO- BIOS, 32, 2: 285-295. Villeurbanne, le 30.04.1999. Manuscrit d~pos~ le 25.08.1998; accept~ d~finitivement le 19.10.1998. ABSTRACT - Quarrying operations for fossil fish from the Bear Gulch Limestone (Heath Formation, Namurian, Montana, USA) have occurred from 1968 to 1997 and continue. The Bear Gulch Limestone lens (14 x 9 km) thins to a shore line on all edges except the buried eastern boundary, and contains an entirely marine flora and fauna. Eighty outcrops have been sampled, and 4 547 identifiable fish of 113 taxa have been recorded. This study details aspects of the diversity and distribution of the fishes of the Bear Gulch Bay analyzed on the basis of geographic areas, following facies and sedimentary criteria. The total diversity of the fish fauna is 21.08 (Simpson index) and 12.07 (Margalef index). Diversity decreases from east to west and towards the southern margin. These trends mask dramatic diffe- rences in species occurrences and dominant species in each area. The high diversity of the "Depocenter" area can be accounted for by its proximity to the mouth of the bay and access to the epicontinental seaway to the east. The sou- theast region is comprised of the Blacktail, Allen, and Buchek areas. Diversity declines greatly from the algae - and sponge - rich Blacktail area southwestward to the marginal Buchek area. This region contains several dominant spe- cies that are adapted to lurk in or maneuver among plants, feed from the bottom, or burrow. High numbers of larval paleoniscoids and the coelacanth Caridosuctor lower the diversity of the near-shore North Shore area. The western end of the bay is dominated by Acanthodes, scales of Listracanthus and Strepsodus, and large cladodont sharks. Fish distributions, preliminary assessments of morphological adaptations, and facies all support the concept of a heteroge- neous set of habitats within this fossil bay. KEYWORDS: BIODIVERSITY, FISH, MARINE, CARBONIFEROUS, PALEOENVIRONMENT, NORTH AMERICA. Rt~SUMI~ - Des poissons fossiles ont ~t~ r~colt~s de 1968 h 1997 (et continuent de l'~tre) dans le Calcaire de Bear Gulch (Formation de Heath, Namurien, Montana, USA). La lentille de calcaire (14 x 9 km) s'amincit jusqu'h une ligne de rivage sur tout son pourtour, sauf le long de sa limite est qui est enfouie. Elle contient une flore et une faune enti~- rement marines. Quatre - vingt affleurements ont ~t~ ~chantillonn~s et 4 547 poissons identifiables appartenant 113 taxons ont ~t~ enregistr~s. Les donn~es sur la r~partition des poissons ont ~t~ analys~es par aires g~ographiques d~finies en fonction de crit~res s~dimentaires et de facies. La pr~sente ~tude d~taille les aspects de la diversit6 des poissons de la Baie de Bear Gulch. La diversit~ totale de la faune ichthyologique est 21,08 (indice de Simpson) et 12,07 (indice de Margalefl. La diversit~ d~cro~t d'Est en Ouest et vers la marge sud. Ces gradients masquent des diff,- rences importantes dans la r~partition des esp~ces, en particulier des esp~ces dominantes, dans chaque aire g~ogra- phique. La forte diversit~ du "Depocenter" peut 6tre expliqu~e par sa proximit~ du d~bouch~ de la baie sur le chenal ~picontinental vers l'Est. La r~gion du Sud-Est comprend les aires de Blacktail, Allen et Buchek: depuis Blacktail, riche en algues et ~ponges, la diversit~ d~croit fortement en direction du Sud-Est jusqu'h l'aire marginale de Buchek. Cette r~gion comporte plusieurs esp~ces dominantes qui sont adapt~es pour s'embusquer ou manoeuvrer parmi les plantes, ou encore fouir le fond pour chercher leur nourriture ou s'enfouir. Dans l'aire de North Shore, pros du riva- ge, le grand nombre de larves de pal~onisco~des et du coelacanthe Caridosuctor font baisser la diversitY. L'extr~mit~ ouest de la baie est domin~e par Acanthodes, les ~cailles de Listracanthus et de Strepsodus et de grands requins cla- dodontes. Les r~partitions des poissons, les estimations pr~liminaires sur les adaptations morphologiques, et enfin les facies ~tayent tousle concept d'une s~rie h~t~rog~ne d'habitats dans cette baie fossile. MOTS-CLI~S:BIODIVERSITI~, POISSONS, MARIN, CARBONIFt~RE,PALI~OENVIRONNEMENT, AMI~RIQUE DU NORD. INTRODUCTION The Bear Gulch Limestone is a Konservat-Lagers- t~tte that contains an excellently preserved record of occurrences of Carboniferous fishes of a tropical marine bay, deposited through a very brief interval of geological time (estimated at 1 000 - 2 000 years, Williams 1980) at the end of the Upper Chesterian. This study is based upon quarrying operations for fossils that were conducted from 1968 to 1997, and, during these years, 80 outcrops were sampled within the Bear Gulch Limestone and 4 547 iden- tifiable fish of 113 taxa have been recorded from these sites.
-
Upload
richard-lund -
Category
Documents
-
view
212 -
download
0
Transcript of Fish diversity of the Bear Gulch Limestone, Namurian, Lower Carboniferous of Montana, USA

RICHARD L U N D & CI~CILE P O P L I N
FISH DIVERSITY OF THE BEAR GULCH LIMESTONE, NAMURIAN, LOWER
CARBONIFEROUS OF MONTANA, USA
LUND R. & POPLIN C. 1999. Fish diversity of the Bear Gulch Limestone, Namurian, Lower Carboniferous of Montana, USA. [La diversit~ des poissons de Bear Gulch, Namurien, Carbonifere inf~rieur du Montana, USA]. GEO- BIOS, 32, 2: 285-295. Villeurbanne, le 30.04.1999.
Manuscrit d~pos~ le 25.08.1998; accept~ d~finitivement le 19.10.1998.
ABSTRACT - Quarrying operations for fossil fish from the Bear Gulch Limestone (Heath Formation, Namurian, Montana, USA) have occurred from 1968 to 1997 and continue. The Bear Gulch Limestone lens (14 x 9 km) thins to a shore line on all edges except the buried eastern boundary, and contains an entirely marine flora and fauna. Eighty outcrops have been sampled, and 4 547 identifiable fish of 113 taxa have been recorded. This study details aspects of the diversity and distribution of the fishes of the Bear Gulch Bay analyzed on the basis of geographic areas, following facies and sedimentary criteria. The total diversity of the fish fauna is 21.08 (Simpson index) and 12.07 (Margalef index). Diversity decreases from east to west and towards the southern margin. These trends mask dramatic diffe- rences in species occurrences and dominant species in each area. The high diversity of the "Depocenter" area can be accounted for by its proximity to the mouth of the bay and access to the epicontinental seaway to the east. The sou- theast region is comprised of the Blacktail, Allen, and Buchek areas. Diversity declines greatly from the algae - and sponge - rich Blacktail area southwestward to the marginal Buchek area. This region contains several dominant spe- cies that are adapted to lurk in or maneuver among plants, feed from the bottom, or burrow. High numbers of larval paleoniscoids and the coelacanth Caridosuctor lower the diversity of the near-shore North Shore area. The western end of the bay is dominated by Acanthodes, scales of Listracanthus and Strepsodus, and large cladodont sharks. Fish distributions, preliminary assessments of morphological adaptations, and facies all support the concept of a heteroge- neous set of habitats within this fossil bay.
KEYWORDS: BIODIVERSITY, FISH, MARINE, CARBONIFEROUS, PALEOENVIRONMENT, NORTH AMERICA.
Rt~SUMI~ - Des poissons fossiles ont ~t~ r~colt~s de 1968 h 1997 (et continuent de l'~tre) dans le Calcaire de Bear Gulch (Formation de Heath, Namurien, Montana, USA). La lentille de calcaire (14 x 9 km) s'amincit jusqu'h une ligne de rivage sur tout son pourtour, sauf le long de sa limite est qui est enfouie. Elle contient une flore et une faune enti~- rement marines. Quatre - vingt affleurements ont ~t~ ~chantillonn~s et 4 547 poissons identifiables appartenant 113 taxons ont ~t~ enregistr~s. Les donn~es sur la r~partition des poissons ont ~t~ analys~es par aires g~ographiques d~finies en fonction de crit~res s~dimentaires et de facies. La pr~sente ~tude d~taille les aspects de la diversit6 des poissons de la Baie de Bear Gulch. La diversit~ totale de la faune ichthyologique est 21,08 (indice de Simpson) et 12,07 (indice de Margalefl. La diversit~ d~cro~t d'Est en Ouest et vers la marge sud. Ces gradients masquent des diff,- rences importantes dans la r~partition des esp~ces, en particulier des esp~ces dominantes, dans chaque aire g~ogra- phique. La forte diversit~ du "Depocenter" peut 6tre expliqu~e par sa proximit~ du d~bouch~ de la baie sur le chenal ~picontinental vers l'Est. La r~gion du Sud-Est comprend les aires de Blacktail, Allen et Buchek: depuis Blacktail, riche en algues et ~ponges, la diversit~ d~croit fortement en direction du Sud-Est jusqu'h l'aire marginale de Buchek. Cette r~gion comporte plusieurs esp~ces dominantes qui sont adapt~es pour s'embusquer ou manoeuvrer parmi les plantes, ou encore fouir le fond pour chercher leur nourriture ou s'enfouir. Dans l'aire de North Shore, pros du riva- ge, le grand nombre de larves de pal~onisco~des et du coelacanthe Caridosuctor font baisser la diversitY. L'extr~mit~ ouest de la baie est domin~e par Acanthodes, les ~cailles de Listracanthus et de Strepsodus et de grands requins cla- dodontes. Les r~partitions des poissons, les estimations pr~liminaires sur les adaptations morphologiques, et enfin les facies ~tayent tousle concept d'une s~rie h~t~rog~ne d'habitats dans cette baie fossile.
MOTS-CLI~S: BIODIVERSITI~, POISSONS, MARIN, CARBONIFt~RE, PALI~OENVIRONNEMENT, AMI~RIQUE DU NORD.
I N T R O D U C T I O N
The Bear Gulch Limestone is a Konservat -Lagers- t~tte tha t contains an excellently preserved record of occurrences of Carboniferous fishes of a tropical mar ine bay, deposited th rough a very brief interval of geological t ime (es t imated at 1 000 - 2 000 years,
Williams 1980) at the end of the Upper Chester ian. This s tudy is based upon quar ry ing operat ions for fossils tha t were conducted from 1968 to 1997, and, dur ing these years, 80 outcrops were sampled within the Bear Gulch Limestone and 4 547 iden- tifiable fish of 113 taxa have been recorded from these sites.

286
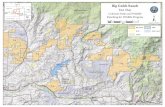
FIGURE 1 - Namurian paleogeographic setting of the Bear Gulch Limestone, Heath Formation, Big Snowy Group, Fergus County Montana. BGL, loca- tion of Bear Gulch Limestone outcrop area; CMT, Central Montana Trough; EQ, 10 °, equator and 10 ° north latitu- de; M, miogeosyndine; N, paleomagne- tic North; WB, Williston Basin. Shaded areas, emergent land; solid lines, cur- rent Canadian and US political boun- daries; dashed line, approximate limit of continental plate. (After Witzke 1990). Situation paldoggographique au Namurien du Calcaire de Bear Gulch, Heath Formation, Big Snowy Group, Fergus County, Montana. BGL, locali- sation de l'affleurement du Calcaire de Bear Gulch; CMT, Bassin Central du Montana; IEQ, 10, dquateur et 10 ° de latitude nord; M, miogdosynclinal; N, Nord paldomagndtique; WB, bassin de Williston. Grisd: terres dmergges; lignes pleines: fronti~res politiques actuelles du Canada et des Etats-Unis; ligne pointillge: limite approximative du pla- teau continental (d'apr~s Witzke 1990).
The Bear Gulch Limestone Konservat-Lagerst~itte is unique among Carboniferous fish faunas in the high number of species, the preservation of the fish and the varied bay habitats, and in the complete absence of freshwater macrofaunal elements. By comparison, Schultze and Maples (1992), in discus- sing Upper Carboniferous fish faunas, list 13 gene- ra for Essex, 12 for Braidwood, 10 for Kinney, 8 for Hamilton, Robinson and Linton (where there are 11-12 species), 5 for Montceau-les-Mines, and 4 for N~r~ny, each of which have a high percentage of freshwater faunal elements. After Moy-Thomas & Bradley Dyne (1938) 32 species of fish are known from the marine Visean of Glencartholm.
The Bear Gulch Limestone fish fauna contrasts str- ongly with modern fish faunas in its species com- position and taxonomic distribution, being compo- sed of about 60% chondrichthyans, about 34% acti- nopterygians, and 5% coelacanths. On the contrary, in terms of number of individuals, the most abun- dant Bear Gulch species is a coelacanth, followed by a few actinopterygians and a chondrichthyan. By comparison, about 96% of modern fish are acti- nopterygians, 3% are chondrichthyans (Elasmo- branchii), and there are trivial percentages of agna- thans, holocephalans, and a coelacanth; in modern faunas only actinopterygians can be considered to be abundant. This study details aspects of the bio- diversity of the fishes of the Bear Gulch Limestone.
The Bear Gulch Limestone lens of the Bear Gulch member of the Heath Formation (Upper Cheste- rian, Namurian E2b, Montana, USA) is 14 km x 9 km in extent, and thins to a shore line on all edges except the eastern boundary, which is buried (Horner 1985; Williams 1983; Feldman et a]. 1994).
O
All indicators suggest that the Bear Gulch Limest- one was deposited in a bay whose narrow mouth opened to the epicontinental seaway along the nor- theastern edge of the deposit. Paleomagnetic evi- dence indicates that the Bear Gulch Limestone was located at about 12 ° N latitude in the Upper Namu- rian, that the North American continent was rota- ted about 45 ° clockwise relative to today's north, while paleogeographical and sedimentary evidence indicates a tropical, semi-arid climate in the region (Williams 1983; Horner 1985; Witzke 1990) (Fig. 1). Climatological evidence indicates that seasonal monsoonal conditions would occur at these lati- tudes, with prevailing wind conditions normally blowing almost directly across the bay: in the win- ter from the east and northeast, and in the summer from the west and southwest (Grogan & Lund, ms.).
The Bear Gulch limestone lens was very rapidly deposited in the narrow and tectonically active Central Montana Trough. It shallowed upward into terrestrial beds and an unconformity that marks the local Mississippian-Pennsylvanian boundary. Direct marine access to the Pacific Ocean through the Central Montana Trough was blocked immedia- tely west of the Bear Gulch Limestone during its time of deposition, but access to the Pacific ocean was possible through the miogeosyncline south of the Williston basin, and access was also available to the eastern North American epicontinental seaway (Fig. 1).
Several facies have been described within the Bear Gulch Limestone on the basis of sedimentological and paleontological data: a basinal facies indicating a clear bottom of fine mud with little or no sessile benthos; an algal facies characterized by large

287
concentrations of fine filamentous floating algae with small spiny productid brachiopods suspended in the algae; an Arborispongia facies characterized by the dominance of up to l-meter- diameter arbo- rescent sponges along with many associated epi- zootic and epiphytic forms and many forms of al- gae; and a marginal facies of dark, irregularly bed- ded, high-organic-content shales and limestones, showing wave action and poor preservation of most vertebrates (Williams 1983; Horner & Lund 1985; Lurid et al. 1993; Feldman et al. 1994) (Fig. 2). The sessile benthic invertebrate fauna of the Bear Gulch Limestone lens is limited in variety and hig- hly restricted in area of occurrence. Cephalopods, crustaceans, and worms dominate the mobile inver- tebrate fauna. There are no significant freshwater or terrestr ial macrofossils in the Bear Gulch Limestone.
MATERIALS A N D M E T H O D S
All identifiable specimens from the Bear Gulch lens are included in the data base for the initial analy- sis; this information is identified as "all Bear Gulch" in the text and tables. Unpublished species are referred to by "code names"; their taxonomic assignments are listed in Table 1.
Data from outcrops have been grouped into seve- ral areas on the basis of geographic and facies cri- teria, and the data from all outcrops within each area are merged for this analysis. One area, in the extreme southeast, has been excluded from fur- ther analyses because it lacks sufficient numbers of specimens, and specimens not attributable to a specific outcrop or area have also been excluded from fur ther consideration.
Data are corrected against sampling bias for the taxa that are only represented by isolated scales or
Class Chondrichthyes
"Subclass - - E1as-mo-br ancl~ii-[ - -E-uc 1~ on~t r oc e p-h-a~-"
Paraselachii Heteropetalus deganhdus
Inf-raclass Holocephali
Psephodus sp. Iarpagofi~tutor volsellorhinu:
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Cladodontida Iniopterygii large stethacanthid "Iniop spl"
O r d e r Falcatusfalcatus "Iniop sp3" Incertae sedis
Listracanthus sp.
TABLE :[ - S y s t e m a t i c d i s t r i b u - t i o n of t h e t a x a r e f e r r e d to in t h i s paper . Place systdmatique des taxons citds clans le texte.
teeth, Strepsodus sp., Psephodus sp., large clado- dont (stethacanthid) sharks, and Listracanthus sp., by only admitting one specimen at any outcrop in any one quarrying day into the analysis. Con- cerning the fragmentary remains, while there is no uncertainty about the identity of any chondrich- thyan species because they are based on gross mor- phological characteristics such as tooth and spine morphology, there are several difficulties inherent in the identification of the actinopterygian taxa, few of which present readily identifiable isolated bones. Many larval actinopterygians may not be identifiable to genus or to species at present, but comprise a significant part of the fish fauna and are treated as distinct ecomorphological units for the determination of species diversity. These larval actinopterygians can only be divided into two groups at present, those that may be attributable to the taxon "Yogo" and those that do not pertain to this taxon. The taxon '~ogo" itself contains two sis- ter species, but few of the specimens can be identi- fied to species at this time. Additionally, about 15 actinopterygian specimens cannot currently be firmly assigned to known species: these are entered into the data base as three tentative unique spe- cies. The number of actinopterygian taxa used for the analyses is therefore considered to be conserva- tive.
The species occurrence data in each area have been analyzed in two manners, first using the absolute numbers, and second as standardized to percent of 100% occurrence at any area to correct for unequal sample sizes. However, unequal sample sizes from area to area, as well as preservational differences, reflect substantive aspects of bay ecology and depo- sition as well as retrieval rates per person per day of quarrying effort.
Acanthodii Acanthodes hmdi
Osteichthyes
Ac~nopterygii Crossopterygii "Yogo" Strepsodus sp.
Wendykhthys dicksoni "Fcop" "Prol"
'Tube snout" "Long pelvic"
"Ae" "Yogo" larvae "Garden Eel"
"Disc" actinopterygian larvae
Coelacanthiformes Caridosuctor populosmn Hadrcalector donbairdi Allenypterus montanus

288
Diversity and dominance indices are calculated using the Species Diversity and Richness software program; equations for the indices and references are given in the accompanying manual (Henderson & Seaby 1997). While there are many different diversity indices comparing the numbers of indivi- duals with the numbers of taxa, the MargalefD and Simpson indices have been used here because they offer different but comparable views of the data. The Berger-Parker dominance index compares the abundance of the most dominant taxa to those of the remaining taxa (Henderson & Seaby 1997). In the figures and tables, however, the most dominant taxa are arbitrarily designated as those containing 3.5% or greater of the individuals at any one area of outcrops.
The map is drawn with a CADD program using a base of U.S. Geological Survey 7 1/2' topographic maps, employing both aerial photographs and ground observations as checks upon the maps. Graphical representation of the standardized occu- rrences at each outcrop are drawn with the Corel Draw vector graphics program, and arranged along
Forest Grove Rose Canyon
Acanthodii
Adult Actinopterygii
Coelacanthiformes
Elasmobranchii
Pa ra se lachii
Holocephali Strepsodus
N l't4
0 5 10
KM
three transects. These transects cover all areas, and facies, for which adequate occurrence data exists.
All data are archived at the Section of Vertebrate Paleontology, Carnegie Museum of Natural History, Pittsburgh, Pa., USA.
RESULTS
FAUNAL COMPOSITION
Data from the entire Bear Gulch Limestone fish fauna show that Acanthodii occur at 1.6% of the total individuals sampled, Actinopterygii compri- se 62% (50% adults and 12% larvae), Coela- canthiformes occur as 14%, and Chondrichthyes as 19% (8% elasmobranchs and 11% Euchondro- cephali) (Table 2). Considerable variabili ty in the percent composition of the major taxonomic groups is evident between the regions of the basin and along transects within the regions (Fig. 2). Acanthodii increase to abundance only toward the western margin of the bay. The greatest percent of adult actinopterygians occurs along the northeast
North Shore Depocenter Big Spring Buchek Allen Blacktail
100
75
50
25
CIES (HABITATS)
446'4/` 4"~80EI~4,£I/v M4RGfN4. O/VO/4 L
FIGURE 2 - Transects of percent occurrences of major taxonomic groups at each outcrop, across outcrop areas of the Bear Gulch Limestone. N, present day North; NN, North during the Namurian (after Witzke 1990). Profils des pourcentages des groupes taxo- nomiques majeurs ~t chaque affleurement, le long des aires d'affleurements du Calcaire de Bear Gulch. N, Nord actuel; NN, Nord au Namurien (d'apr~s Witzke 1990).

289
to southwest transect in the southeastern region (comprised of the Blacktail, Allen-Surprize, and Buchek areas) where larval actinopterygians are scarce; actinopterygians are scarce toward the wes- tern margin of the bay. Coelacanths are at their highest percentages in the central eastern region of the bay (comprised of the Big Spring, Depocenter, and North Shore areas) along the South to North transect. Both elasmobranchian and euchondroce- phalan (= Iniopterygii + Paraselachii + Holoce- phali, Lund & Grogan 1997) Chondrichthyes are at their most abundant toward the southern and wes- tern margins of the basin. The crossopterygian Strepsodus sp., represented only by scales and teeth, is most common at the southern and western margins of the basin. Generalizing by major taxo- nomic subdivision, the acanthodian Acanthodes lundi dominates toward the western extent of the Bear Gulch Limestone, along with Elasmobranchii, Euchondrocephali and Strepsodus sp., while the percentages of adult actinopterygians, larval acti- nopterygians and coelacanths diminish westward. Coelacanths as well as adult and larval actinopte- rygians dominate in the North Shore area.
Data from the entire Bear Gulch Limestone fish fauna show also that, of the 113 taxonomic units, only seven are present at greater than 3.5% of the total: the coelacanth Caridosuctor populosum, lar- val actinopterygians, the actinopterygians Wen- dyichthys dicksoni and "Fcop", the stethacanthid elasmobranch Falcatus falcatus, and the actinopte- rygians "Yogo" and "Prol" (Table 3).
In the northeastern region (Fig. 3) of the Bear Gulch Limestone, the Big Spring area differs pri- marily in adding the actinopterygian Long Pelvic to the dominant fish. The North Shore area differs in the high percentage of larval actinopterygians (29%) collected, followed by C. populosum, the abundance of the actinopterygian "Garden Eel" and the lower percentage of"Fcop".
In the southeastern region (Fig. 3) of the Bear Gulch Limestone, larval actinopterygians are not among the dominant elements of the fauna. C. populosum only occurs among the dominant ele- ments of the fauna at Allen-Surprise, F. falcatus only in the Blacktail area, while at the Buchek area the only dominant elements of the fauna that are shared in common with the fauna as a whole are W. dicksoni and "Fcop". The actinopterygians "Tube snout" and "Long pelvic" appear among the domi- nants in both Blacktail and Allen-Surprise areas. The "Garden Eel" occurs among the dominants at Blacktail area, along with "Ae".
In the western region (Fig. 3) of the Bear Gulch Limestone, the percentages of C. populosum and the numbers of actinopterygian taxa decline, while the numbers of chondrichthyan taxa and the per- cent ofAcanthodes lundi increase to the westward. "Yogo" continues to be a dominant element of the fauna in this region. The Rose Canyon area is simi- lar in composition to the northeastern region but includes "Yogo" larvae and the holocephalan Harpagofututor volseUorhinus among the domi- nants.
TABLE 2 - Percent composition of Bear Gulch Limestone fish fauna by major taxonomic subdivision for the entire deposit (ALLBG) and by outcrop areas along tran- sects. Composition en pourcen- tages de la faune ichthyologique de Bear Gulch pour les groupes taxonomiques majeurs, pour la totalitd du dgpSt (ALLBG) et par aires d'affleurement selon les axes d'orientation.
NE to S W South to North East to West
A g n a t h a 0.07 0.20 0.07
A c a n t h o d i i 1.61 0.76 0.69 0.00 1.78 0.87 1.14 9.80 28.57
E l a s m o b r a n c h i i 8.01 9.09 5.52 6.90 5.92 8.99 5.85 17 .65 19.48
E u c h o n d r o c e p h a l i 11.59 15,91 8.28 24 .14 10.26 11.59 9.27 17 .65 20.78
Chondr ichthyes 19.60 25.00 13.79 31.03 16.17 20.58 15.12 35.29 40.26
A c t i n o p t e r y g i a n A d u l t 50.19 59.85 69.66 43.38 46,94 53.20 35.61 43.14 20.78
A c f i n o p t e r y g i a n L a r v a e 12.38 3,79 4.14 3,45 13 .81 9 . 9 3 29.59 3.92 0.00
Act inopterygi i 62.57 63.64 73.79 46.83 60.75 63.13 65.20 47.06 20.78
C o e l a c a n t h i f o r m e s 14.34 7.58 11,72 10.34 19.13 13.80 16.91 5.88 2.60
S t r e p s o d u s 0.o4 3.03 1.38 6.90 1.97 1.56 1.63 1.96 7.79

290
Forest Grove Rose Canyon
Acanthodes
Prol W. dicksoni
Yogo C. populosum
Ctadodont E falcatus
Iniop 1 INop 3
hi. volsellorhinus Strepsodua sp.
N iiii!iiiiii!iiiiiii! l . . . . . .
I
North Shore Depocenter Big Spring
N Buchek Allen Blacktail
N
:':'::+:': : :" 50 . • '
Larval Actinop
-
!i!!!:!i!i!i!i!i!i~::x ~\',~ ......... ~,z~i! H. elegantulus -:':+:+1";5':" +:':+. ~ "~ ' ; 0
FIGURE 3 - Transects of dominant species (>3.5%) across outcrop areas of the Bear Gulch Limestone. N, present day North; NN, North during the Namurian (after Witzke 1990). See Table 1 for systematic assignments of species. Published species in italics, unpublished species designated by identification codes. Profils des nombres d'esp~ces dominantes (>3,5%) le long des aires d'af- fleurement du Calcaire de Bear Gulch. N, Nord actuel; NN, Nord au NaTnurien (d'apr~s Witzke 1990). Voir le Tableau 1 pour la place systdmatique des esp~ces. Esp~ces publiges en italiques, esp~ces non publides ddsignges par des codes.
DIVERSITY
The total diversity of all Bear Gulch Limestone fish is calculated as 21.08 (Simpson index) and 12.07 (Margalef index) (Table 4).
Diversity indices of each area, plotted on a map of the bay, show trends of decreasing diversity along transects westward away from the eastern margin. Diversity of the fish fauna is the smallest in the two areas located in the marginal facies, Buchek and Forest Grove (Fig. 4).
The southeast region is fringed along its eastern border by Arborispongia (Rigby 1985) facies, parti- cularly rich in filamentous algae centrally, and extends into the marginal facies in the Buchek area. There is a distinct decrease in diversity in the southeastern region along the transect from Blacktail through Allen to Buchek, corresponding to this facies transition. Blacktail has the highest Simpson number in the Bear Gulch, 23.2, but it is ranked significantly lower by the Margalef index (4.45). This region contains a high number of domi- nant species that are adapted to lurk in or maneu- vet among plants, feed from the bottom, or burrow. Thus the elongate, diminutive paraselachian
Heteropetalus elegantulus and the eel-like holoce- phalan Harpagofututor volsellorhinus accompany actinopterygians with elongate, eel-like, deep, or flattened-venter body morphologies. Unusually, the coelacanth Caridosuctor populosum is very rare in this area, as are larval actinopterygians. This region was clearly sheltered from a high-energy flow regime but the individual areas within it are quite heterogeneous in habitat.
The Allen - Surprize area (Simpson - 16.6, Margalef - 3.8) is noteworthy principally for its low Margalef diversity index, but is unremarkable in terms of the relative abundance of its fish. The Buchek area has a sufficiently low sample number that any statis- tics calculated here are very uncertain. Notwiths- tanding, three rare to uncommon species are among the principal species found, the eel-like holocepha- lan chondrichthyan Harpagofututor volsellorhinus, the discoidal actinopterygian "Disc" and the short- bodied coelacanth Hadronector donbairdi.
The three areas of the northeast region, Big Spring, Depocenter, and North Shore, lay along a south-to- north transect starting in a thin area of the basin facies, passing through the thickest part of the

291
TABLE 3 - Percent occurrences of dominan t species (>3.5 %), n u m b e r of taxa, and n u m b e r s of specimens, for all f ish and for outcrop a reas of the Bear Gulch Limestone. Publi- shed species in italics, unpub l i shed species des ignated by identifica- t ion codes. Composition en pourcen- tages des esp~ces dominantes (>3,5°/o), et pour les aires d'affleure- ments du calcaire de Bear Gulch. Esp~ces publides en italiques, esp~ces non publides ddsigndes par des codes.
S p e c i e s
Acanthodes hmdi
Cladodont sp
Falcatus falcatus
Listracanthus histrix
Heteropetalus elegantulus
Harpagofututor volsellorhinus
Iniop spl
Iniop sp2
Yogo gulo
Wendyichthys dicksoni
F cop
F prolatus
Tube snout
Long pelvic
Ae erin
Garden eel
Disc pec
Larval actinop
Strepsodus sp.
Caridosuctor populosum
Hadronector donbairdi
NO taxa:
N:
Bear Gulch Limestone lens, and ending in Arbo- rispongia facies extremely close to the northern limit of the Bear Gulch Limestone lens. They pre- sent a faunal continuum in terms of their species content as well as in terms of their relative diversi- ty profiles. The Big Spring area (Simpson - 15.4, Marga le f - 6.84) is characterized by its high per- centages of Caridosuctor populosum, larval acti- nopterygians, and the unusual actinopterygian "Long Pelvic". The Depocenter area (Simpson - 22.3, Margalef - 10.5) has the highest number of species in all groups. There are fewer, and lower-ranking, dominant species here than elsewhere in the bay; two of these are the coelacanth C. populosum and large full - bodied larval actinopterygians.
Extremely high numbers of small slim larval acti- nopterygians, and the abundant coelacanth C. po- pulosum lower the overall diversity of the reef- like North Shore area (Simpson - 8.36, Margalef- 7.3).
NE to SW South to North East to West
"~ r ,
= -~ ~ .~ ca ~ -=
9.80 28.57
12.99
5.32 6.12 11.76 5.19
6.90
8.33
10.34 5.88
5.19
5.19
5.08 5.52 5.40 4.55 9.80 7.79
6.91 11.36 8.28 10.34 5.92 7.44 7.84
6.71 15.17 6.90 9 .07 7.37
4.31 5.30 4.53 5.53 5.88
5.52
4.55 8.28 4.93
4.55
4.55 4.07
6.90
9.39 4.14 !13.81 9.93 29.59
6.90 7.79
11.50 8.28 16.37 11.38 14.47
6.90
113 38 33 20 54 84 58 26 17
4547 132 145 29 507 2891 615 51 77
The shallow western end of the bay, Forest Grove (Simpson - 8.7, Margalef - 2.2) and Rose Canyon (Simpson - 16.8, Margalef - 2.7) areas, has far fewer species and far fewer specimens than other areas, as well as poorer preservation. Acanthodes lundi, rare elsewhere in the fauna and usually represen- ted by poorly preserved or fragmentary individuals, is the dominant member of the fish fauna at Forest Grove. Scales of Listracanthus and Strepsodes sp., large cladodont shark carcasses and teeth, and the remains of two species ofiniopterygians, "Iniop spl" and "Iniop sp3", are also common at the western margin of the bay.
DOMINANCE
The diversity indices conceal dramatic differences in percentages of species occurrences, and domi- nant species, in each habitat (Tables 3,4). Two areas in particular, North Shore and Forest Grove, are

292
A S A S A S A S A S A S A S A S A S
H 3.58 3.552 3.392 3.42 3.165 3.165 2.901 2.901 3.307 3.323 3.584 3.568 2.982 2.982 2.994 3.032 2.628 2.628 S H A N N O N - W E I N E R -. i . . . . . . . . . . .
Var iance H 0,0003 0,0002 0,0059 0,0001 0,0065 0,0001 0,0181 0 0,0031 0.0602 0,0005 0.0001 0,0041 0.0002 0,0139 0 i0,0141 0.0001 i
S I g N 21.726 21.079 27.041 23.187 18.643 16.652 36.909 16.52 15.717 15.419 22.608 22.344 8.457 8.366 23.558 16.807 9.657 : 8.689
M A R G A L E F D 13.298 12.073 8.218 4.451 7.033 3.8 5.643 2.063 9.8 6.84 13.05 10.535 10.434 7.274 6.133 2.714 4.604 2.171
E Q U I T A B I L I T Y 0.757 0.753 0,634 0,723 0,591 0,67 0,542 0.614 0.618 0.703 AREA826 AREA836 AREA834 AREA827
0,67 0.755 0,557 0.631 0,359 0.641 0,491 0.556
D O M I N A N C E - B E R G E R - P A R K E R 0.115 0.117 0.115 0.114 0.152 0.152 0.103 0.103 0.164 0.164 0.114 0.114 0.293 0.293 0.12 0.118 0.286 0.286
TABLE 4 - Diversity, equitability and Berger-Parker dominance indices for all Bear Gulch fishes and for outcrop areas of the Bear Gulch Limestone. Data are calculated for absolute numbers of individuals at outcrop areas (A = Absolute) and for numbers of indi- viduals standardized (S = Standardized) to percent of the total individuals at each outcrop area. Indices de diversitY, "~quitabilitd" et de dominance Berger-Parker pour tous les poissons de Bear Gulch et pour les aires d'affleurement du calcaire de Bear Gulch. Calculs faits pour le nombre absolu d' individus dans les aires (A = Absolute) et pour les nombres d ' individus ramen~s au pourcen- tage du nombre total d ' individus dans chaque aire (S = Standardized).
FOREST GROVE 4 0 - -
3O
20
ROSE CANYON NORTH SHORE DEPOCENTER BIG SPRING BUCHEK ALLEN BLACKTAIL 4O
30
2O
10 10
N N ~ l l k OUTCROP *
o ~
0
N / ' .1" ~ s,MPso. FACIES (HABITATS) ~ MARGALEF
UAtG'/4
FIGURE 4 - Transects of Simpson and Margalef D diversity indices across outcrop areas of the Bear Gulch Limestone, using percent occurrences of individuals in species at each outcrop. N, present day North; NN, North during the Namurian (after Witzke 1990). Profils des indices de diversitg de Simpson et de Margalef D le long des aires d'affleurement du Calcaire de Bear Gulch, calculgs & par- tir des pourcentages d'individus des esp~ces & chaque affleurement. N, Nord actuel; NN, Nord au Namurien (d'apr~s Witzke 1990).
e x t r a o r d i n a r i l y d o m i n a t e d by one t a x o n o m i c uni t , and h a v e B e r g e r - P a r k e r d o m i n a n c e i n d e x scores in t he .28 to .29 r a n g e , wh i l e B e r g e r - P a r k e r scores for m o s t a r e a s of t h e B e a r Gu lch L i m e s t o n e is in
t he r a n g e of .1O to .16. D i f f e r e n t f i sh d o m i n a t e a t t he se two a reas : 29.5% of t h e f i sh a t N o r t h Sho re a re l a r v a l a c t i n o p t e r y g i a n s , w h i l e 28.5% of t h e spe- c imens a t Fo re s t Grove a re A c a n t h o d e s l u n d i . A .

293
lundi is a member of the dominant fish group at only one other area, Rose Canyon, where it occurs at 9.8%, and is present at other areas only as a tri- vial component of the fauna. The Buchek area has a low dominance index of .10, reflecting the collec- tion of 29 specimens from 20 different taxa and again, but numbers of specimens found for each day of quarrying effort were also extraordinarily low in and near this area.
D I S C U S S I O N
The fishes of the Bear Gulch Limestone present a faunal continuum throughout their bay environ- merit. There are, however, clear gradients of decrea- sing diversity among the Bear Gulch fishes, with the highest diversity occurring closest to the eas- tern margin and the entrance from the epiconti- nental seaway, and the lowest diversity occurring in the near-shore marginal facies to the south and west. A south-to-north transect close to and paral- lel with the eastern margin of the deposit traverses the deepest, thickest, most speciose, and most diverse area of the bay to terminate close to the northern margin of the Bear Gulch lens in an area dominated by sponges, algae, brachiopods, bivalves, larval actinopterygians, and the coelacanth C. populosum. High concentrations of sponges, bra- chiopods, and other associated invertebrates at the North Shore and along the eastern edge of the Bear Gulch Limestone indicate a relative]y high energy environment and strong nutrient supply (Grogan & Lund, ms), in association with the direction and
timing of the seasonally fluctuating wind-driven hydrodynamic and nutrient flow conditions near the mouth of the bay. This shallow, high productivi- ty zone provided nutrients as well as shelter for large numbers of larval fish, and proximity to the mouth of the bay and the epicontinental seaway would have provided a source of refreshment of the bay waters and a route of immigration for plankto- nic, opportunistic, and migratory fish species. These conditions would support the levels of diversity found in the eastern reaches of the Bear Gulch bay.
The southeastern area of the bay is almost as high in diversity by the Simpson index as is its northern counterpart, although it has far fewer species and individuals. It is most noteworthy, however, that a large proportion of the species and the individual fish in this area have body forms that deviate from those of normal swimming, maneuvering fish towards either the eellike or the deepened body form and that neither larval actinopterygians nor C. populosum are dominant parts of the local com- munity.
The east-west diversity gradient culminates to the westward in a marginal facies area dominated by A. lundi, with lesser components of several chon- drichthyans and scales and teeth of Strepsodus sp. A. lundi is a toothless filter-feeder that is known to have bred in fresh-brackish water lagoons to the immediate west of the Bear Gulch lens (Zidek 1980; Lund et al. 1993). The increase in abundance of A. lundi to the west can be interpreted to mean a higher level of the fine prey particles necessary for
Forest Grove North Shore Depocenter Buchek Blacktail
Larval Actinops W. dicksoni W. dicksonJ W. dicksoni
H. elegantulus niop sp 3 Yogo Yogo
~ ~ ~ Larval ~ Disco ~
F. falcatus F Prol Actinops ~ ~ /~)i~ Ae
1 C. populosum C. populosum ~ F cop
Long Pelvic
H. volsellorhinus Garden eel F. cop H. volsellorhinus Garden eel
FIGURE 5 - Body forms of dominant species at selected areas (see Table 1 for sys temat ic a s s ignmen t s of species). Acanthodes drawn from A. bridgei after Zidek 1980. Silhouettes des esp~ces dominantes dans quelques aires gdographiques (voir le Tableau I pour la place systdmatique des esp~ces). Acanthodes dessinds selon Acanthodes bridgei d'apr~s Zidek 1980.

294
their maintenance as well as increased seasonal fresh water input from the west and southwest, possibly signifying true estuarine conditions. Strepsodus sp., however, a large - toothed predator, is most common in shallow water marginal deposits regardless of presumed salinity or the availability of fine particulate prey.
The trend in diversity indices from the eastern margin, the most diverse area, to the least diverse at the southern and western margins of the Bear Gulch Limestone are accompanied by distinctive differences in faunal composition. These distinc- tions among the dominant members of each area agree with the habitat patchiness indicated by pre- viously assigned geofacies and biofacies, supporting the concept that different patterns of resource uti- lization were responsible for the existence of dis- tinctive subcommunities within the bay.
It is possible to extract some preliminary ecomor- phological information from the fossil fishes of the Bear Gulch Limestone with which to examine some aspects of resource utilization in this bay, albeit in a tentative and preliminary manner. There is, for instance, ample evidence from the contrast between the abundance of larval actinopterygians and the record of life stage histories among the chondrich- thyans to indicate that the actinopterygians were R-selected, and that the coelacanths as well as many of the chondrichthyans, as at present, were clearly K-selected and utilized strong ]ife-stage segregation (Pianka 1994; Lurid 1990; Lund & Lund 1984). The distribution of the fish also dem- onstrates distinctive differences in body form am- ong the areas. Body form relates directly to swim- ruing function and thus may most reasonably be equated to habitat preference as a measure of niche utilization (Webb 1984; Pianka 1994). In the broa- dest sense of body and fin form and function, larval actinopterygians and all but one of the coelacanths (Allenypterus montanus LUND & LUND, 1984), with their rounded, low aspect ratio tails, share a com- mon suite of maneuvering adaptations (Webb 1984), with the coelacanths specialized to the highest degree for fine positional adjustments (Lund et al. 1985). The actinopterygians W. dickso- ni and '~Yogo", by contrast, typify more open-water swimming adaptations, while "Disc" and "Ae" exem- plify shelter - dwelling maneuvering forms. Among the chondrichthyans, the elasmobranchs, that also demonstrate strong size-based resource partitio- ning (Schoener 1974), typify the body forms of acti- ve predatory swimmers, while most of the euchon- drocephalans and particularly the holocephalans possess body forms with maneuvering, top-water, benthic, or near-benthic adaptations (Fig. 5).
Feeding morphology is intimately related to prey preferences and thus to prey availability. The five
species of coelacanths also epitomize fine niche par- titioning on the basis of gape size (prey size) as well as body form (Lund et al. 1985). Acanthodes lundi was an obligate filter-feeder. There is a great mor- phological differentiation of dentitions, suspenso- ria, dental histologies, and therefore feeding adap- tations among the subclasses of chondrichthyans (Lund 1990; Lund & Grogan 1997). While many of the actinopterygians remain to be described in detail, the most prevalent actinopterygian across all but the most marginal areas of the Bear Gulch Limestone is W. dicksoni, with a unique combina- tion of feeding-mechanism characters of a keyhole- shaped notch in the snout, an extralateral bone in the gular series, and fine, forwardly-pointed teeth (Lund & Poplin 1997).
CONCLUSION
Over 113 species of fish have been identified from the Namurian Bear Gulch Limestone of Montana, rendering it the largest Paleozoic fish fauna, and one of the largest fossil fish faunas known. This study of the occurrence and the diversity of the fishes of this Konservat-Lagerst~itte has revealed intrabasinal gradients between areas with distinct differences in species composition within the faunal continuum of the bay. These differences correspond to geofacies and biofacies differences in the bay, and are related to seasonal wind-driven hydrodynamics and the consequent patterns of nutrient flow, and can be further correlated to both morphological and life-history adaptations of the fish found in the bay. A preliminary assessment of the morphological adaptations of some of the dominant fish within each area presents ample evidence to support the hypothesis that these adaptations in swimming mechanisms, feeding mechanisms, and life history strategies were critical determinants of resource utilization during the Namurian. Further study of these factors, and finer-grained study of the distri- bution of the fish within areas, may further clarify the basis for the diversity and distribution of the fishes of the Bear Gulch Limestone. Anyhow, even if only a preliminary study, the present work shows how, in its main lines, the ecology of a marine tro- pical bay could be organized during Namurian times.
Acknowledgments - We are grateful to Henri Lavina for edi- t ing the tables, to Eileen Grogan for advice and help, and also to all those who have helped in the excavations through the years, and par t icu lar ly Ron Bogosh, Carol DeBell, Bill Hopkins, Br ian Kleinhaus, Myra Moore and Bryan Snyder.
R E F E R E N C E S
FELDMAN H., LUND l:{., MAPLES C. • ARCHER A. 1994 - Origin of the Bear Gulch beds (Namurian, Montana, USA). Geobios, M.S. 16: 283-291.

295
HENDERSON P.A. & SExY R. 1997 - Species diversity and richness. Pisces Conservation Ltd., Pennington, UK, 10 p., plus softL ware.
HORNER J.R. 1985 - The stratigraphic position of the Bear Gulch Limestone (Lower Carboniferous) of central Montana. Comp- te Rendu, Neuvi~me Congr~s International de Stratigraphie et de Gdologie du Carbonil~re, Urbana (Illinois), 5: 427-436.
HORNER J.R. & LUND R. 1985 - Biotic distribution and diversity in the Bear Gulch limestone. Comptes Rendus Neuvi~me Congr~s International de Stratigraphie et de Ggologie du Carbonif~re, Urbana (Illinois), 5: 437-442.
LUND R. 1990 - Chondrichthyan life history styles as revealed by the 320 million years old Mississippian of Montana. Environmental Biology of Fishes, 27, 19 p.
LUND R., FELDMAN H., LUND W. & MAPLES C. 1993 - The depositio- hal environment of the Bear Gulch Limestone. In Montana Geological Society Guidebook 1993, Energy and Mineral Resources of Central Montana. Fergus County Montana: 87- 96.
LUND R. & GROGAN E.D. 1997 - Relationships of the Chimae- riformes and the basal radiation of the Chondrichthyes. Reviews ofFish Biology and Fisheries, 7, 59 p.
LUNI~ R. & LUND W.L. 1984 - New genera and species of coela- canths from the Bear Gulch Limestone (Lower Carbonife- rous) of Montana (U.S.A.). Geobios, 17, 2: 237-244.
LUND R. & POPLIN C. 1997 - The rhadinichthyids (paleoniscoid actinopterygians) from the Bear Gulch Limestone of Monta- na. Journal of Vertebrate Paleontology, 17: 466-486.
LUND W.L., LUND R. & KLEIN G. 1985 - Coelacanth feeding mecha- nisms and the ecology of the Bear Gulch coelacanths. Comp- tes Rendus Neuvi~me Congr~s International de Stratigraphie et de Ggologie du Carboni~re, Urbana (Illinois), 5: 492-500.
MoY-THOMAS J.A. & BRADLEY DYNE B.A. 1938 - The actinoptery- gian fishes from the Lower Carboniferous of Glencartholm, Eskdale, Dumfriesshire. Transactions of the Royal Society of Edinburgh, 59: 437-480.
PI~KA E.R. 1994 - Evolutionary ecology, 5th ed.. Harper Collins, New York, 486 p.
RIGBY J.K. 1985 - The sponge fauna of the Mississippian Heath Formation of Central Montana. Comptes Rendus Neuvi~me Congr~s International de Stratigraphie et de Gdologie du Carbonil~re, Urbana (Illinois), 5: 443-456.
SCHOENER. T.W. 1974 - Resource partitioning in ecological com- munities. Science, 185: 27-39.
SCHULTZE. H - P. & C.G. MAPLES 1992 - Comparison of the Late Pennsylvanian faunal assemblage of Kinney Brick Company quarry, New Mexico, with other Late Pennsylvanian lagers- t~tten. New Mexico Bureau of Mines and Mineral Resources Bulletin, 138: 231-235.
WEBB P.W. 1984 - Body form, locomotion and foraging in aquatic vertebrates. American Zoologist, 24: 107-120.
WILLIAMS L.A. 1981 - The sedimentational history of the Bear Gulch Limestone (Middle Carboniferous, Central Montana). Ph.D. dissertation, Princeton University, 242 p. (unpublished).
WILLL~MS L.A. 1983 - Deposition of the Bear Gulch Limestone: a Carboniferous Plattenkalk from Central Montana. Sedimen- tology, 30: 843-860.
WITZK~ B.J. 1990 - Paleoclimatic constraints for Paleozoic Paleolatitudes of Laurentia and Euramerica. In Mc KERROW W.S. & SCOTESE C.R. (eds), Paleozoic Paleogeography and bio- geography. Geological Society Memoirs, 12: 57-73.
ZIDEK J. 1980 - Acanthodes lundi, new species (Acanthodii) and associated coprolites from uppermost Mississippian Heath Formation of Central Montana. Annals of the Carnegie Museum, 49: 49-78.
R. L U N D Biology Department, Adelphi University
Garden City New York 11530, U.S.A.
E.mail: [email protected]
C. P O P L I N Laboratoire de Pal4ontologie, UMR 8569 CNRS
8 rue Buffon F - 75005 Paris