
Feline Immunodeficiency Virus Envelope Glycoproteins ...jvi.asm.org/content/88/6/3255.full.pdf ·...
18
Feline Immunodeficiency Virus Envelope Glycoproteins Antagonize Tetherin through a Distinctive Mechanism That Requires Virion Incorporation James H. Morrison, a Rebekah B. Guevara, a * Adriana C. Marcano, a * Dyana T. Saenz, a Hind J. Fadel, a,b Daniel K. Rogstad, a,b Eric M. Poeschla a,b Departments of Molecular Medicine a and Medicine (Division of Infectious Diseases), b Mayo Clinic College of Medicine, Rochester, Minnesota, USA ABSTRACT BST2/tetherin inhibits the release of enveloped viruses from cells. Primate lentiviruses have evolved specific antagonists (Vpu, Nef, and Env). Here we characterized tetherin proteins of species representing both branches of the order Carnivora. Compari- son of tiger and cat (Feliformia) to dog and ferret (Caniformia) genes demonstrated that the tiger and cat share a start codon mutation that truncated most of the tetherin cytoplasmic tail early in the Feliformia lineage (19 of 27 amino acids, including the dual tyrosine motif). Alpha interferon (IFN-) induced tetherin and blocked feline immunodeficiency virus (FIV) replication in lymphoid and nonlymphoid feline cells. Budding of bald FIV and HIV particles was blocked by carnivore tetherins. However, infectious FIV particles were resistant, and spreading FIV replication was uninhibited. Antagonism mapped to the envelope gly- coprotein (Env), which rescued FIV from carnivore tetherin restriction when expressed in trans but, in contrast to known antag- onists, did not rescue noncognate particles. Also unlike the primate lentiviral antagonists, but similar to the Ebola virus glyco- protein, FIV Env did not reduce intracellular or cell surface tetherin levels. Furthermore, FIV-enveloped FIV particles actually required tetherin for optimal release from cells. The results show that FIV Envs mediate a distinctive tetherin evasion. Well adapted to a phylogenetically ancient tetherin tail truncation in the Felidae, it requires functional virion incorporation of Env, and it shields the budding particle without downregulating plasma membrane tetherin. Moreover, FIV has evolved dependence on this protein: particles containing FIV Env need tetherin for optimal release from the cell, while Env particles do not. IMPORTANCE HIV-1 antagonizes the restriction factor tetherin with the accessory protein Vpu, while HIV-2 and the filovirus Ebola use their envelope (Env) glycoproteins for this purpose. It turns out that the FIV tetherin antagonist is also its Env protein, but the mechanism is distinctive. Unlike other tetherin antagonists, FIV Env cannot act in trans to rescue vpu-deficient HIV-1. It must be incorporated specifically into FIV virions to be active. Also unlike other retroviral antagonists, but similar to Ebola virus Env, it does not act by downregulating or degrading tetherin. FIV Env might exclude tetherin locally or direct assembly to tetherin- negative membrane domains. Other distinctive features are apparent, including evidence that this virus evolved an equilibrium in which tetherin is both restriction factor and cofactor, as FIV requires tetherin for optimal particle release. T he type I interferon (IFN-I)-inducible factor BST2/tetherin/ CD317/HM1.24 inhibits cell surface release of a diverse set of enveloped viruses that include retroviruses (1–5), filoviruses (2, 6, 7), the arenavirus Lassa virus (7), and the DNA virus Kaposi’s sarcoma-associated herpesvirus (KSHV) (8). Viruses have evolved antagonists that sequester tetherin away from the cell sur- face or directly induce its degradation through proteasomal or lysosomal pathways (for reviews, see references 9, 10, and 11). For primate lentiviruses, three different viral proteins are known to antagonize tetherin restriction: Vpu (HIV-1, SIVmus/gsn/mon) (4, 5, 12–14), Nef (several simian immunodeficiency virus [SIV] strains) (13–16), and Env (HIV-2, SIVtan) (3, 17, 18). The antiviral phenotype that prompted the search for tetherin, restricted HIV-1 particle release from the plasma membrane, was observed over 20 years ago (19, 20). Tetherin contains a short cytoplasmic N terminus with dual Yxx motifs that mediate clathrin-dependent endocytosis (21), a single-pass transmem- brane domain, an ectodomain that contains two N-linked glyco- sylation sites (22) as well as three cysteines that participate in in- termolecular disulfide bonds (22, 23), and a C-terminal glycosylphosphatidylinositol (GPI) anchor (24). These distinctive features suggested that tetherin exerts its antiviral function by connecting cellular and viral membranes. This mechanism has been supported by immunoelectron microscopy (25–27) and by evidence that antiviral function is preserved if the component do- mains are substituted with structurally similar domains from other proteins (27). Recent experiments have clarified the topol- ogy involved: viral and plasma membranes are infiltrated by op- posite ends of a parallel tetherin homodimer, with a severalfold preference for insertion of the transmembrane domains into the Received 22 December 2013 Accepted 26 December 2013 Published ahead of print 3 January 2014 Editor: S. R. Ross Address correspondence to James H. Morrison, [email protected], or Eric M. Poeschla, [email protected]. * Present address: Rebekah B. Guevara, Dartmouth Graduate School, Hanover, New Hampshire, USA; Adriana C. Marcano, Ponce School of Medicine and Health Sciences, Ponce, Puerto Rico. Copyright © 2014, American Society for Microbiology. All Rights Reserved. doi:10.1128/JVI.03814-13 March 2014 Volume 88 Number 6 Journal of Virology p. 3255–3272 jvi.asm.org 3255 on July 13, 2018 by guest http://jvi.asm.org/ Downloaded from
-
Upload
truongngoc -
Category
Documents
-
view
219 -
download
0
Transcript of Feline Immunodeficiency Virus Envelope Glycoproteins ...jvi.asm.org/content/88/6/3255.full.pdf ·...

Feline Immunodeficiency Virus Envelope Glycoproteins AntagonizeTetherin through a Distinctive Mechanism That Requires VirionIncorporation
James H. Morrison,a Rebekah B. Guevara,a* Adriana C. Marcano,a* Dyana T. Saenz,a Hind J. Fadel,a,b Daniel K. Rogstad,a,b
Eric M. Poeschlaa,b
Departments of Molecular Medicinea and Medicine (Division of Infectious Diseases),b Mayo Clinic College of Medicine, Rochester, Minnesota, USA
ABSTRACT
BST2/tetherin inhibits the release of enveloped viruses from cells. Primate lentiviruses have evolved specific antagonists (Vpu,Nef, and Env). Here we characterized tetherin proteins of species representing both branches of the order Carnivora. Compari-son of tiger and cat (Feliformia) to dog and ferret (Caniformia) genes demonstrated that the tiger and cat share a start codonmutation that truncated most of the tetherin cytoplasmic tail early in the Feliformia lineage (19 of 27 amino acids, including thedual tyrosine motif). Alpha interferon (IFN-�) induced tetherin and blocked feline immunodeficiency virus (FIV) replication inlymphoid and nonlymphoid feline cells. Budding of bald FIV and HIV particles was blocked by carnivore tetherins. However,infectious FIV particles were resistant, and spreading FIV replication was uninhibited. Antagonism mapped to the envelope gly-coprotein (Env), which rescued FIV from carnivore tetherin restriction when expressed in trans but, in contrast to known antag-onists, did not rescue noncognate particles. Also unlike the primate lentiviral antagonists, but similar to the Ebola virus glyco-protein, FIV Env did not reduce intracellular or cell surface tetherin levels. Furthermore, FIV-enveloped FIV particles actuallyrequired tetherin for optimal release from cells. The results show that FIV Envs mediate a distinctive tetherin evasion. Welladapted to a phylogenetically ancient tetherin tail truncation in the Felidae, it requires functional virion incorporation of Env,and it shields the budding particle without downregulating plasma membrane tetherin. Moreover, FIV has evolved dependenceon this protein: particles containing FIV Env need tetherin for optimal release from the cell, while Env� particles do not.
IMPORTANCE
HIV-1 antagonizes the restriction factor tetherin with the accessory protein Vpu, while HIV-2 and the filovirus Ebola use theirenvelope (Env) glycoproteins for this purpose. It turns out that the FIV tetherin antagonist is also its Env protein, but themechanism is distinctive. Unlike other tetherin antagonists, FIV Env cannot act in trans to rescue vpu-deficient HIV-1. It mustbe incorporated specifically into FIV virions to be active. Also unlike other retroviral antagonists, but similar to Ebola virus Env,it does not act by downregulating or degrading tetherin. FIV Env might exclude tetherin locally or direct assembly to tetherin-negative membrane domains. Other distinctive features are apparent, including evidence that this virus evolved an equilibriumin which tetherin is both restriction factor and cofactor, as FIV requires tetherin for optimal particle release.
The type I interferon (IFN-I)-inducible factor BST2/tetherin/CD317/HM1.24 inhibits cell surface release of a diverse set of
enveloped viruses that include retroviruses (1–5), filoviruses (2, 6,7), the arenavirus Lassa virus (7), and the DNA virus Kaposi’ssarcoma-associated herpesvirus (KSHV) (8). Viruses haveevolved antagonists that sequester tetherin away from the cell sur-face or directly induce its degradation through proteasomal orlysosomal pathways (for reviews, see references 9, 10, and 11). Forprimate lentiviruses, three different viral proteins are known toantagonize tetherin restriction: Vpu (HIV-1, SIVmus/gsn/mon)(4, 5, 12–14), Nef (several simian immunodeficiency virus [SIV]strains) (13–16), and Env (HIV-2, SIVtan) (3, 17, 18).
The antiviral phenotype that prompted the search for tetherin,restricted HIV-1 particle release from the plasma membrane, wasobserved over 20 years ago (19, 20). Tetherin contains a shortcytoplasmic N terminus with dual Yxx� motifs that mediateclathrin-dependent endocytosis (21), a single-pass transmem-brane domain, an ectodomain that contains two N-linked glyco-sylation sites (22) as well as three cysteines that participate in in-termolecular disulfide bonds (22, 23), and a C-terminalglycosylphosphatidylinositol (GPI) anchor (24). These distinctive
features suggested that tetherin exerts its antiviral function byconnecting cellular and viral membranes. This mechanism hasbeen supported by immunoelectron microscopy (25–27) and byevidence that antiviral function is preserved if the component do-mains are substituted with structurally similar domains fromother proteins (27). Recent experiments have clarified the topol-ogy involved: viral and plasma membranes are infiltrated by op-posite ends of a parallel tetherin homodimer, with a severalfoldpreference for insertion of the transmembrane domains into the
Received 22 December 2013 Accepted 26 December 2013
Published ahead of print 3 January 2014
Editor: S. R. Ross
Address correspondence to James H. Morrison, [email protected], orEric M. Poeschla, [email protected].
* Present address: Rebekah B. Guevara, Dartmouth Graduate School, Hanover,New Hampshire, USA; Adriana C. Marcano, Ponce School of Medicine and HealthSciences, Ponce, Puerto Rico.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.03814-13
March 2014 Volume 88 Number 6 Journal of Virology p. 3255–3272 jvi.asm.org 3255
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

cell membrane and the GPI anchors into the virion (28). Thus,each protein monomer operates, within a dimer, as an end-to-endtether between cell and virus.
The domestic dog, cat, and ferret lack functional antiviralTRIM5alpha and TRIMCyp genes (29–31), leaving them without aversion of the important postentry capsid-targeting systems thatrestrict retroviruses in the cells of numerous primates (32–35);there is similarly no known carnivore equivalent to the mouse Fv1gene (36). The cat does have an effective APOBEC3 protein rep-ertoire, which is antagonized by feline immunodeficiency virus(FIV) Vif and feline foamy virus Bet (37–41). FIV particle release isalso inhibited by human tetherin (2), but FIV does not encodehomologues of Vpu or Nef.
Two different tetherin cDNAs generated from Felis catusmRNA transcripts have been tested (42, 43). The two predictedproteins have dissimilar N termini. Dietrich et al. reported a 186-amino-acid sequence for feline tetherin, which we here refer to asFelis catus tetherin long form (fcTetherinLF). The N-terminal 19amino acids align well with the N termini of primate tetherinproteins. In contrast, a truncated form, here termed Felis catustetherin short form (fcTetherinSF), lacks these 19 N-terminalamino acids (43, 44). The difference is potentially important be-cause N termini of primate tetherin proteins contain highly con-sequential motifs. For example, deletion of the N-terminal 20amino acids of human tetherin results in a protein with alteredpatterns of glycosylation and subcellular trafficking and absentantiviral activity against HIV-1 (4). Furthermore, rhesus macaquetetherin residues 14 to 18 determine Nef antagonism, and theabsence of these 5 amino acids in the human tetherin N terminusdrove the evolution of HIV-1 Vpu (13, 15, 16; see references 45and 10 for reviews). Additional debate concerns whether FIV en-codes a tetherin antagonist or has been otherwise selected to evadethe protein. A recent study reported that FIV Env can amelioratetetherin restriction (44), although this was not observed by Di-etrich et al. (42).
MATERIALS AND METHODSUse of primary carnivore cells. Tiger and ferret blood samples were pro-vided by outside collaborators (see Acknowledgments) in accordancewith their institutional guidelines. Felis catus blood was collected by us inaccordance with the Guide for the Care and Use of Laboratory Animalsunder Mayo Clinic Institutional Animal Care and Use Committee(AAALAC Accreditation No. 000717; OLAW Assurance No. A3291-01)Protocol A4210, approved 3 March 2010.
Cell lines. MCC cells are a feline large granular lymphoma cell line(46); Crandell feline kidney (CrFK) cells are an adherent cell line derivedfrom feline kidney (47). Mya-1 cells are an interleukin-2 (IL-2)-depen-dent T cell line (48). Cell lines were maintained in Dulbecco’s modifiedEagle medium (DMEM) or RPMI medium supplemented with 10% fetalcalf serum (FCS), penicillin-streptomycin, and L-glutamine. KE-R cellswere a gift of Carsten Münk. Mya-1 cells were additionally cultured with20% FCS, 10% D-glucose, 1% sodium bicarbonate, 1% sodium pyruvate,1% nonessential amino acids (NEAA), and 0.1% �-mercaptoethanol. Fe-line peripheral blood mononuclear cells (PBMCs) were purified by Ficollcentrifugation from whole blood and maintained in RPMI medium sup-plemented with 10% FCS, 2 mM glutamine, 1 mM sodium pyruvate,essential and nonessential amino acids, 10 mM HEPES, 0.05 mM �-mer-captoethanol, phytohemagglutinin E (PHA-E; 2 �g/ml), and human IL-2(in conditioned medium from murine L2.23 feeder cells, which were akind gift of T. Miyazawa). PHA-E was discontinued 48 h after isolation.For FIV replication assays in IFN-treated cells, 1 ng/ml human IFN-�
(hIFN-�; Sigma) was added 24 h before infection and maintained for theduration of the experiment.
Isolation of tetherin cDNAs and 5= RACE. A table that details alloligonucleotides used in this work is available on request. cDNAs cor-responding to fcTetherinLF, accession no. NM_001243085 (42), andfcTetherinSF, accession no. DDBJ AB564550 (43), were amplified by re-verse transcriptase PCR (RT-PCR) from mRNA of feline PBMCs pre-treated with 1,000 U/ml rat IFN-� (Sigma). A dog tetherin cDNA wasisolated by RT-PCR using mRNA of D17 cells, also pretreated with 1,000U/ml rat IFN-�. Total RNA was collected with the RNeasy Plus Minikit(Qiagen) and converted to cDNA with Transcriptor First Strand cDNASynthesis (Roche). Tetherin cDNAs were amplified using species-specificprimers, blunt ligated into pSC-B amp/kan (Agilent), and confirmed bysequencing. For 5= rapid amplification of cDNA ends (RACE), total RNAwas collected as described above from domestic cat, tiger, and ferretPBMCs as well as from canine D17 and feline K-ER and CrFK cells. 5= halfsegments of tetherin transcripts were determined using the FirstChoiceRLM-RACE kit (Applied Biosciences) according to the manufacturer’sinstructions. Tetherin-specific primers were used. Nested products wereisolated on agarose gels and blunt ligated using the StrataClone Blunt PCRcloning kit (Agilent). Six to 10 clones were sequenced for each species, andonly clones containing the intact 36-bp 5= RACE adapter were includedfor later comparison.
Plasmid constructions and generation of stable cell lines. Briefly,p1012 HIV-1 IN-myc-IRES-puro (49) was partially digested with KpnI,and a 1.5-kbp fragment containing IRESpuro was isolated and insertedinto the KpnI site in Tsin poly (50) to create Tsin-IRESpuro. Tsin-HA-tetherin-IRES-puro was created by PCR amplifying BglII/ClaI-flanked tetherin and cloning into a BamHI/ClaI-digested Tsin-IRESpuro.Tsin tetherin-HA was constructed by overlap extension PCR of a BamHI/ClaI (human and canine)- or BglII/ClaI (felineLF/SF)-flanked tetherin toinsert an HA tag internally within the tetherin open reading frame (ORF)(immediately after amino acid 154, 158, or 159 of human, canine, orfelineLF tetherin, respectively) before insertion in a BamHI/ClaI-digestedTsin-IRESpuro backbone. Stable cell lines were generated by transducingtargets and selecting in puromycin (3 �g/ml for 293T cells, 0.5 �g/ml forK-ER cells, or 5 �g/ml for CrFK cells). pHIV-1luc�Vpu was created byexchanging a SalI/KpnI fragment of pNL4.3 �Vpu containing a frame-shifted vpu into pNL4.3R-E-�426 (pHIV-1luc) (51). pCT5 (52, 53) en-codes replication-competent FIV34TF10 (54). The truncated OrfA of pCT5(43 amino acids) was repaired, changing a stop codon to Trp and resultingin the production of a 78-residue protein in pCT5A�, which produces thevirus FIV34TF10A� when transfected into human cells (55). Two pCT5-analogous constructs that allow production of FIV particles in humancells were created for the PPR and C36 clones of FIV. pCT-PPR was con-structed similarly to pCT5 (52, 53) from FIVPPR in order to replace the 5=FIV U3 element with the cytomegalovirus (CMV) immediate early pro-moter. pCT-C36A� was created in two steps. First, an Msc1-Eco47IIIfragment of FIVC36 Clone C was ligated into the Msc1-Eco47III-digestedpCT-PPR backbone. The intermediate from step 1 was then linearizedwith MscI, and an MscI-MscI fragment from FIVC36 Clone C was insertedand screened for proper orientation by sequencing. pCT-C36 thus con-tains PPR LTR elements but is otherwise entirely C36 from the 33rd Gagnucleotide to the middle of the 3= U3 element. Only one amino acid isdifferent from C36, a V-to-M change at amino acid 11 of Gag. FIV Enve-lope expression plasmids (pFE) were constructed by PCR amplification ofNotI/SalI-flanked Env from pCT-PPR or pCT-C36. This PCR extendedfrom the first codon of Env to the end of the RRE. PCR amplicons weredigested with NotI/SalI and cloned into a NotI/SalI-digested FP93 back-bone, which replaces Gag-Pol, but not Rev. pCT-C36 A� was created bysite-directed mutagenesis (QuikChange Lightning, Agilent), causing asingle-nucleotide insertion and frameshift of the OrfA open reading framein the fourth codon. The pCT-C36 A� furin cleavage site mutant wasconstructed by site-directed mutagenesis (606RQKR609 to 606RQAA609).pCT-C36 TM�43 was constructed by site-directed mutagenesis to gener-
Morrison et al.
3256 jvi.asm.org Journal of Virology
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

ate a stop codon immediately following Env amino acid 812. pCG-IRES-dsRed2 constructs for fcTetherinSF and Canis lupus familiaris tetherincDNA (cfTetherin) were made by swapping a SnaBI to PmlI fragment ofTsin-tetherin-IRESpuro into pCG-huTetherin-IRES-dsRed2.
Supernatant RT activity. Ten microliters of filtered supernatant wasaliquoted in triplicate on a 96-well U-bottom Falcon plate. Fifty microli-ters of reverse transcriptase (RT) cocktail [67 mM Tris, 100 mM KCl, 2.7mM dithiothreitol, 6.6 mM MgCl2, 6.7 �g/ml poly(A), 2.1 �g/ml oli-go(dT), 0.067% NP-40, and 13 nCi/ml [32P]dP] was added, and the platewas incubated at 37°C for 1.5 h. Five microliters of the mixture was thenpipetted onto a Millipore MultiscreenHTS DE plate and incubated untildry. Filter plates were attached to a vacuum and washed with 1.5 ml 2SSC (1 SSC is 0.15 M NaCl plus 0.015 M sodium citrate) and 0.4 ml ethylalcohol (EtOH) per well. Plates were dried for another 30 min before aback-seal adhesive was added. Thirty-five microliters of MicroScint-Oscintillation fluor was added before plates’ relative rads were measured ona TopCount plate reader. Reverse transcriptase counts are displayed as theaverages of the triplicates standard deviations (SD).
Single-cycle and replicating FIV stocks. Previously described meth-ods were used to generate viral stocks (56, 57) by 293T cell transfection.FIV34TF10 was produced in 293T cells by transfection of pCT5 (52, 53) orpCT5A�. Similarly, FIVPPR and FIVC36 were produced by 293T transfec-tion of pCT-PPR and pCT-C36A�, respectively. When indicated, viruseswere first-round vesicular stomatitis virus G glycoprotein (VSV-G) pseu-dotyped by cotransfection with pMD.G (VSV-G). Viral stocks were fil-tered (0.45 �m), and their titers were determined on CrFK cells by a focalinfectivity assay (52). Viral replication was monitored by RT activity (57).Mean RT activity SD from duplicate measurements for each sample wascalculated.
Real-time PCR quantification of tetherin cDNA. Total RNA was iso-lated using a Qiagen RNeasy Plus Minikit after treatment for 24 h withhIFN-� (1 ng/ml, Sigma) or control vehicle. cDNA was generated from 5�g of total RNA using Transcriptor First Strand cDNA Synthesis (Roche)for real-time PCR. Two microliters cDNA was quantified against a seriallydiluted plasmid standard using a Roche 480 LightCycler and RocheLCDA software. Feline glyceraldehyde-3-phosphate dehydrogenase gene(GAPDH) was quantified in parallel. LightCycler programs included aninitial denaturation step at 95°C for 10 min and a melting step after am-plification (65°C to 97°C; temperature transition rate, 0.11°C/s). Tetherinor IRF7 cDNA was amplified using 500 nM sense primer and 500 nMantisense primer. PCR amplification and analysis were performed as fol-lows: for tetherin, 95°C for 10 s, 57°C for 10 s, and 72°C for 17 s for 45cycles, with temperature transition rate of 4.4°C/s; for IRF7, 95°C for 10 s,57°C for 10 s, and 72°C for 6 s for 45 cycles, with temperature transitionrate of 4.4°C/s; for GAPDH, 95°C for 10 s, 53°C for 10 s, and 72°C for 17s for 45 cycles, with temperature transition rate of 4.4°C/s. Triplicate RNAisolations were performed for each treatment group of each cell line (n �1 for PBMCs), and samples was measured in triplicate (tetherin gene) orin duplicate (GAPDH, IRF7); a two-tailed t test was performed betweencontrol and treated groups.
NF-�B induction by tetherin. 293T cells (2.5 104) were seeded in a96-well plate and allowed to adhere overnight. Cells were polyethylenei-mine (PEI) transfected in triplicate with pNF�B(3x)-Firefly luciferase,pTAL-Gaussia luciferase, pCG-tetherin-IRES-dsRed2, and pCG-HIV-1M-vpu-IRES-eGFP at a 4:1:1.6:4 ratio with 150 ng total DNA in each well(58). Forty-eight hours after transfection, gaussia and firefly luciferaseactivities were measured in the supernatant and cells, respectively. Gaussialuciferase was used to control for transfection efficiency, and NF-�B in-duction was compared for tetherin-transfected versus control transfectedcells.
Indirect immunofluorescence, laser scanning confocal microscopy.Localization experiments were performed using previously describedmethods (50) with variations described below. Briefly, 1 105 cells wereplated in LabTek II chamber slides, allowed to adhere overnight, fixedwith 4% (wt/vol) paraformaldehyde, permeabilized with methanol (un-
less otherwise stated), stained with 4=,6-diamidino-2-phenylindole(DAPI), and imaged by laser scanning confocal fluorescence microscopy.Imaging of cell surface tetherin expression was performed by calciumphosphate transfection of 2 105 293T cells with 1 �g tetherin-HA ex-pression plasmids, 1.5 �g pCT5A�, pCT-PPR, or pCT-C36A�, and Tsinpoly (stuffer) to bring the total amount of DNA to 2.5 �g in each well.Cells were washed 16 h after transfection, and after an additional 24 h cellswere fixed, stained, and imaged as described above. Antibodies used wererat monoclonal anti-HA (Roche), Alexa Fluor 594-conjugated goat anti-rat (Invitrogen), cat anti-PPR serum, and fluorescein isothiocyanate(FITC)-conjugated goat anti-cat (Santa Cruz Biotech) antibodies.
Particle release assays. 293T cells were PEI transfected with 1.5 �g thepackaging constructs pHIV-1Luc (51), pHIV-1Luc �Vpu, pFP93 (FIV[59]), pCT-PPR, pCT-C36A�, or the indicated viral mutant. Cells werewashed 8 h later and harvested after an additional 48 h. Cell lysates werecollected in radioimmunoprecipitation assay (RIPA) buffer (150 mMNaCl, 0.5% deoxycholate, 0.1% sodium dodecyl sulfate, 1% NP-40, 150mM Tris-HCl, pH 8.0) plus protease inhibitors (complete-Mini; Boehr-inger). Supernatants were filtered (0.45 �m). Equivalent amounts of su-pernatant and cell lysates were immunoblotted for HIV-1 CA using the39/5.4A monoclonal antibody (MAb; Abcam) or FIV CA using felineserum reactive to PPR (a gift of Peggy Barr). Blots were stripped andreprobed with rat �-HA (Roche) and mouse anti-alpha-tubulin (Sigma)monoclonal antibodies. RT activity was also quantified as describedabove. CrFK particle release assays were performed as described above,except transduction by indicated single-round VSV-G-pseudotyped vec-tors produced in 293T cells was used in lieu of transfection. A total of2.5 104 cells were plated in a 24-well plate and allowed to adhere over-night before being transduced with equivalent volumes of vector. Cellswere washed 3 times after 24 h and then 72 h after transduction superna-tants and cell lysates were harvested as described above. Envelope antag-onism experiments were performed by 293T PEI transfection as describedabove with 750 ng viral packaging construct (pHIV-1Luc �Vpu orpCT5efs) and 750 ng VSV-G, pFE-PPRRRE, pFE-C36RRE, pEbolaGPmyc(myc-tagged Ebola virus glycoprotein [60]), or pCDNA3.1 (stuffer).Ebola virus glycoprotein (GP) expression was detected by immunoblot-ting with polyclonal rabbit anti-Ebola virus sera.
Pseudotyped lentiviral vector titers. Green fluorescent protein(GFP)-expressing HIV-1 vectors were produced by PEI cotransfection of3 105 293T cells with 600 ng HIV-1 packaging construct pCMV-�R8.9,600 ng HIV-1 GFP transfer construct TSinG (51), and 200 ng envelopeplasmid (pMD.G [VSV-G], pFE-PPR, or pFE-C36A�). GFP-expressingFIV vectors were similarly produced with 600 ng pFP93, 600 ng FIV GFPtransfer construct GiNSIN (61), and 200 ng envelope. Cells were washed 8h after transfection, and after an additional 48 h supernatant was filtered(0.45 �m), and titers were determined on CrFK cells expressing felineCD134 (fCD134). Titers are shown as transducing units (TU) per ml.Reverse transcriptase activity of vectors was also measured as describedabove.
Flow cytometry for intracellular and cell surface tetherin. For intra-cellular tetherin degradation experiments, CrFK cells were plated in a24-well plate, allowed to adhere overnight, and infected with FIV. Sixteenhours later cells were washed, and 48 h later cells were fixed with 4%(wt/vol) paraformaldehyde, permeabilized with methanol, and stainedfor FIV proteins (cat anti-PPR serum primary and goat anti-cat phyco-erythrin [PE]-conjugated [Santa Cruz Biotech] secondary antibodies)and tetherin (rat anti-HA primary and goat anti-rat Alexa Fluor 488-conjugated [Invitrogen] secondary antibodies). Tetherin and FIV expres-sion was then analyzed using a FACScan (BD Biosciences) and CellQuest(Becton, Dickinson) software. Fluorescence-activated cell sorting (FACS)for cell surface tetherin levels was performed on clonal 293T cells express-ing internally epitope-tagged tetherin. Cells were PEI transfected with 100ng pEGFP-N1 and 1.4 �g of pCDNA3.1(�), pCDNA3.1vphu (62), pFE-PPRRRE, pFE-C36RRE, pCT-PPR, or pCT-C36A�. Cells were washed 8 hafter transfection, and after an additional 48 h cells were trypsinized,
Mechanism of Tetherin Antagonism
March 2014 Volume 88 Number 6 jvi.asm.org 3257
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

washed with phosphate-buffered saline (PBS), and stained for HA (ratanti-HA primary and goat anti-rat Alexa Fluor 594-conjugated [Invitro-gen] secondary antibodies), and dual-color FACS was run on an LSRII(Becton, Dickinson). Analysis was performed by gating for GFP-positive cellsand comparing mean fluorescence intensity (MFI) of Alexa Fluor 594.
RESULTSEffects of type I IFN on FIV replication. We observed that humanIFN-� treatment blocked FIV replication in primary felinePBMCs (Fig. 1A). We then tested IFN-� antiviral effects in a panelof domestic cat cell lines and observed clear differences. Viral rep-lication was completely suppressed in the adherent cell line K-ERand in the IL-2-dependent T cell line Mya-1 (Fig. 1B and C). Incontrast, replication was inhibited but not abrogated in a secondadherent feline cell line, CrFK (Fig. 1D), and in the lymphoma cellline MCC peak viral production was nearly the same in the pres-ence and absence of IFN-� (Fig. 1E). Type I interferon inhibitionof HIV-1 replication (63) is due at least in part to upregulatedexpression of host cell restriction factors, including tetherin (4, 5,64). To determine whether feline tetherin induction could beplaying a role in the patterns of viral suppression shown in Fig. 1,we proceeded to assess the IFN-� inducibility of feline tetherinmRNA in different feline cells and to clone, sequence, and expresscDNAs for both reported Felis catus isoforms (42, 43) as well as forCanis familiaris tetherin. The purpose of characterizing the dogmRNA and protein was to encompass both main branches of theCarnivora (Caniformia and Feliformia).
Carnivore tetherins. cDNAs for fcTetherinLF and fcTetherinSF
(corresponding to cBST2* and cBST2504, respectively, in the studyby Celestino et al. [44]) were amplified by RT-PCR from mRNA offeline peripheral blood mononuclear cells (PBMCs) that we pre-treated with 1,000 U/ml rat IFN-� (rIFN-�). A Canis lupus famil-iaris tetherin cDNA (cfTetherin) was similarly isolated from rIFN-�-treated canine DK cell mRNA using primers designed fromBLAT searches of the dog genome. In Fig. 2A, the deduced proteinsequences are aligned with the human tetherin sequence.
Although the carnivore proteins have limited overall homol-
ogy with human tetherin (46% and 43% amino acid identity forfcTetherinLF and cfTetherin, respectively), residues known to beimportant for proper dimerization, glycosylation, and traffickingof the human protein to and from the cell surface are conserved,with the exception of a variance at one of the positions of N-linkedglycosylation and the absence of the dual-tyrosine-based endocy-tosis motif in fcTetherinSF. As expected, the two carnivore teth-erins share greater amino acid identity with each other (60%) thanwith human tetherin.
Interrogation of the domestic cat genome assembly did notidentify an open reading frame that corresponds directly to thesuggested (42) fcTetherinLF (Fig. 2B). However, splicing couldgenerate an mRNA that encodes this putative longer isoform. Toclarify the issue, we performed 5=RACE on mRNA from Felis catusPBMCs and cell lines, which identified only transcripts corre-sponding to fcTetherinSF (Fig. 2B). We also cloned and sequencedtiger (Panthera tigris) and ferret (Mustela putorius furo) tetherincDNAs. Alignment of tiger, cat, dog, and ferret tetherin sequences,the last two representing Caniformia (Canidae and Mustelidae,respectively), revealed a felid-specific single-nucleotide deletion atthe codon corresponding to the initiator methionine of fcTetherinLF
(Fig. 2B, arrow). The presence of this deletion in both the tiger andthe domestic cat genes suggests that it occurred after the Cani-formia-Feliformia split, around 43 to 60 million years ago (MYA)(65), but before the modern felid species radiation in the late Miocenebegan, approximately 11 MYA (66). The AG dinucleotide generatedby the T nucleotide deletion could theoretically act as a splice acceptorto bring an upstream ATG into the frame here, but we found noevidence for this in repeated RACE experiments using primary andimmortalized feline cells (data not shown).
Using primers that amplify across exon boundaries of the BST2gene transcript, quantitative real-time PCR was performed onmRNA collected from a variety of canine and domestic cat celllines. Consistent with the IFN-� inducibility of human tetherin(4), treatment of feline cells with human IFN-� resulted inmarked upregulation of tetherin transcripts in cells of lymphoid
FIG 1 Type I IFN inhibits FIV replication. (A) Domestic cat PBMCs treated or not treated with 1 ng/ml of recombinant human IFN-� were infected withFIV34TF10A� (OrfA-repaired version of FIV34TF10) and assayed for RT activity. (B to E) Mya1 (B), K-ER (C), CrFK (D), and MCC (E) cells were also infected underthe same conditions.
Morrison et al.
3258 jvi.asm.org Journal of Virology
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

FIG 2 Deduced amino acid sequences, mRNA expression profiles, and NF-�B activation of carnivore tetherins. (A) Human tetherin is aligned with Felis catusand Canis familiaris tetherins. Highlighting indicates residues implicated by prior studies in human tetherin trafficking and multimerization (21–24, 27, 95, 96).(B) mRNA sequences determined by 5= RACE. Underlined codons and numbering refer to putative fcTetherinLF (M1) and fcTetherinSF initiator methionines.(C to E) Analysis of tetherin mRNA expression and IFN upregulation was performed in domestic cat cell lines (C), cat PBMCs (D), and canine cell lines (E); dataare presented as copies of tetherin cDNA per 105 copies of GAPDH cDNA SD; n � 9 experiments except for PBMCs, where n � 3 for each donor. *, P 0.01;**, P 0.001. (F) Primers amplifying a conserved region of IRF-7 were used to analyze IRF-7 expression in samples collected as described for panels C and E.*, P 0.01; **, P 0.001. (G) Tetherin-induced NF-�B induction was determined in 293T cells by cotransfection of an NF-�B-driven (3 NF-�B binding sites)firefly luciferase construct, a control promoter-driven gaussia luciferase, tetherin expression constructs, and HIV-1 M Vpu or control plasmids. Fold inductionof NF-�B-driven firefly luciferase is displayed as the average of three independent transfections SD.
Mechanism of Tetherin Antagonism
March 2014 Volume 88 Number 6 jvi.asm.org 3259
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

(Mya-1, FeT-J) and nonlymphoid (CrFK, K-ER) origin (Fig. 2C).Feline PBMCs also exhibited IFN-�-inducible tetherin expressionat a level comparable to that seen in human PBMCs, yielding 5.9-and 13.6-fold induction, respectively, from two different donors(Fig. 2D). In contrast, IFN-� treatment did not result in tetherininduction in a canine osteosarcoma (D17) or kidney cell line (DK)(Fig. 2E). The MCC lymphoma cell line was the only feline cell linetested in which IFN-� did not substantially inhibit FIV replication(Fig. 1E), and it was also the cell line in which IFN-� induction oftetherin mRNA was least, although the absolute mRNA levelswere lower in CrFK cells. This failure to substantially induce teth-erin in MCC cells appears to reflect a deficit in the IFN-� signalingpathway because IRF-7, an IFN-�-inducible transcription factor(67) recently found to boost tetherin upregulation in mice (68),was also not upregulated by IFN-� treatment in these cells (Fig.2F). Likewise, the canine cell lines did not exhibit IRF-7 upregu-lation, correlating with a lack of tetherin mRNA induction (Fig. 2Eand F).
Before it was implicated in defense against enveloped viruses,human tetherin was identified in a large-scale cDNA library ex-pression screen for NF-�B-activating proteins (69). Recent evi-dence for discordant NF-�B signaling between human tetherinisoforms that differ only in cytoplasmic tail length suggests thattetherin may mediate complex signal transduction activities andact as an antiviral sensor that triggers proinflammatory gene ex-pression (70). Specifically, the YxY clathrin adaptor binding motiflocated in the cytoplasmic tail of the longer human isoform wasfound to be critical for NF-�B signaling (70–72). To test whethercarnivore tetherins induce NF-�B signaling and whether the cyto-plasmic tail truncation of feline tetherin may impact this, we com-pared cfTetherin and fcTetherinSF to human tetherin for theirability to activate a minimal NF-�B promoter in 293T cells. Nei-ther carnivore tetherin was found to activate NF-�B signaling un-der these conditions (Fig. 2G).
Subcellular localization of feline and canine tetherins. Weexpressed each of the four tetherins stably in 293T cells and per-
FIG 3 Subcellular localization of tetherin in human and feline cells. Puromycin selection was used to generate stable tetherin-expressing cell lines. Scanningconfocal microscopy was used for immunofluorescence analysis of tetherin localization (anti-HA, red) and DAPI (blue) for 293T (A), CrFK (B), and K-ER (C)cells. Boxed regions are shown at higher magnification alongside differential interference contrast (DIC) images. (D) Cell surface accumulation of tetherin inCrFK cells was determined by performing immunofluorescence as described for panel B, except without cell permeabilization by methanol. Cells expressed aninternally HA-tagged tetherin (diagrammed).
Morrison et al.
3260 jvi.asm.org Journal of Virology
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

formed confocal immunofluorescence microscopy (Fig. 3A). Weincluded fcTetherinLF here and subsequently in order to testwhether its 19 N-terminal amino acids, which we speculate mighthave formed the N terminus of an ancestral feline tetherin protein,confer altered bioactivity. Each protein displayed a similar intra-cellular distribution, with abundant cytoplasmic foci and detect-able plasma membrane-localized protein, which is consistent withthe pattern described previously for the human protein (4). Incontrast, in the feline cell line CrFK, tetherin was detected pre-dominantly in cytoplasmic inclusions, with little visible accumu-lation at the plasma membrane (Fig. 3B). The exception wasfcTetherinSF, which had slightly increased levels of diffuse andplasma membrane-localized tetherin, suggesting functional ca-pacity of the 19-amino-acid tail to mediate recycling from the cellsurface. Stable expression in a second feline cell line, KE-R (Fig.3C), revealed similar distributions for each tetherin.
Though little ectopically expressed tetherin was generally de-tected at the plasma membrane of feline cell lines in standardconfocal images, the protein may accumulate at the cell surface inamounts that are less easily discerned when abundant intracellulartetherin is coimaged. Therefore, we inserted an epitope tag intothe extracellular portion of the protein, according to the strategyof Perez-Caballero et al. (27), and analyzed feline cell surface ac-cumulation. While tetherin was again visible predominantly in
intracellular accumulations in permeabilized cells, in nonperme-abilized cells cell surface tetherin was clearly visible (Fig. 3D).These results indicate that the protein does indeed traffic to thesurface of these feline cells, even though the protein present therein the steady state may be a relatively small fraction of the totalcellular pool.
Carnivore tetherins restrict lentiviral budding. In order totest the antiviral properties of carnivore tetherins, we used lenti-viral vectors to generate stable epitope-tagged tetherin-expressinglines from 293T cells, which express negligible endogenous teth-erin. We derived single-cell clones and used two independentclones for each protein in further experiments (Fig. 4A). Viralexpression constructs used here and in subsequent experimentsare diagrammed in Fig. 4B and are denoted as pFIV when referringto direct expression from transfected plasmid and as FIV whenviral particles produced from these vectors are used to infect freshcells. Similar to the human protein, canine and feline tetherinsreduced the accumulation of Vpu-deficient HIV-1 virus-like par-ticles (VLPs) in supernatant to background levels (Fig. 4C). Con-sistent with the limited transmembrane domain conservation be-tween human and carnivore tetherins (Fig. 2A), Vpu was able torescue HIV-1 budding in cells expressing human tetherin but hadno effect at all in cells expressing feline or canine tetherin (Fig. 4C).Budding of FIV VLPs (53, 73) was similarly restricted by all of the
FIG 4 Carnivore tetherins restrict lentiviral particle release. 293T cells expressing HA-tagged human (hs), canine (cf), or either form of domestic cat tetherin(fcSF; fcLF) were single-cell cloned, and tetherin expression in multiple clones was confirmed by immunoblotting for HA (A). (B) Diagram of FIV and HIV-1packaging constructs. Intact open reading frames are shown in white, while truncated or frameshifted ORFs are in gray. Names correspond to plasmid name(first) and viruses produced by transfection of the plasmid in 293T cells (second). (C) 293T clones expressing tetherin were transfected with either pHIV-1Luc orpHIV-1Luc �Vpu. Viral protein expression and budding were confirmed by immunoblotting for HIV-1 CA in cell lysates and supernatants, respectively. Whilein this experiment processing of HIV-1 Gag appears inhibited in the presence of fcTetherinLF, this was not seen in other experiments and may mainly reflect lowerGag expression in these lanes. (D) Same as panel C, using the FIV packaging construct pFP93 (73) and immunoblotting with anti-FIV (serum from anFIVPPR-infected cat).
Mechanism of Tetherin Antagonism
March 2014 Volume 88 Number 6 jvi.asm.org 3261
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

tetherins, though canine tetherin was less effective than human orfeline tetherins (Fig. 4D). For both viruses, the shorter form offeline tetherin, which our evidence indicates is the natural one inpresent-day felids (Fig. 2B), and the artificially longer form hadsimilar antiviral properties, indicating that inclusion of the puta-tive ancestral N-terminal cytoplasmic tail including the endocyto-sis motif does not augment antiviral function.
Budding of infectious FIV is not affected by tetherin. To fur-ther characterize the effects of feline tetherin on replication-com-petent, wild-type FIV, we generated CrFK cell lines that stablyexpress the protein. Levels of endogenous tetherin mRNA in CrFKcells, which we assessed by GAPDH (glyceraldehyde-3-phosphatedehydrogenase) mRNA-normalized quantitative RT-PCR, wereextremely low, and the peak level after IFN induction was also thelowest in any line that we examined (Fig. 2C), making it a goodsubstrate for stable ectopic expression. Single-cell clones weregenerated, with two independent clonal lines for each tetherinused in further analyses (Fig. 5A). Cells that express fcTetherinSF
had substantially increased protein levels compared to fcTetherinLF
or cfTetherin despite the presence of similar numbers of mRNAtranscripts for each protein (Fig. 5B), suggesting that the N-ter-minal cytoplasmic tail of fcTetherinLF either reduces translationor, more likely, increases the protein’s turnover rate through en-hanced internalization.
These cell lines were infected with a �Vpu HIV-1 reportervirus, and the supernatants were assayed for de novo VLP accumu-lation. Similar to what was observed in the human cells, each of thetetherins effectively inhibited VLP accumulation (Fig. 5C). Vari-ation was evident in the supernatant p24 levels, which may reflectdifferences in tetherin expression levels between cell clones (thefirst Homo sapiens [hs] tetherin lane in Fig. 5A corresponds to thefirst H. sapiens tetherin lane in Fig. 5C).
CrFK cells do not express the primary FIV receptor CD134,which restricts spreading replication in these cells to laboratory-adapted strains that can use the coreceptor CXCR4 alone for en-try. However, VSV-G-pseudotyped CD134-dependent FIVs com-plete a single round of infection and subsequent viral assemblyunimpeded in these cells, which allows assessment of particle bud-ding under conditions where all viral proteins, including Env, areexpressed. We used VSV-G-pseudotyped preparations of full-length CD134 receptor-dependent FIVs to infect tetherin-ex-pressing CrFK cell lines and then examined viral accumulation inthe supernatant. Following infection with the clade A virusFIVPPR, intracellular and supernatant accumulation of viral pro-teins, including CA and Env, was observed in control CrFK cells(Fig. 5D). In contrast to their effects on HIV-1 virions and FIVVLPs, fcTetherinSF and fcTetherinLF only mildly inhibited FIVPPR
egress (Fig. 5D). FIVPPR was also only modestly inhibited by
FIG 5 FIV budding is not restricted by tetherin in domestic cat cells. Lentiviral vectors were used to express tetherin stably, followed by single-cell cloning andconfirmation of expression by immunoblotting for the HA tag (A) and qRT-PCR for BST-2 mRNA transcripts (B). qPCR data are presented as numbers of copiesof BST-2 cDNA per 105 copies of GAPDH cDNA SD, n � 3. (C to E) Cells expressing human (hs), canine (cf), or domestic cat (fcSF; fcLF) tetherin were infectedwith VSV-G pseudotyped HIV-1Luc �Vpu (C), FIVPPR (D), and FIVC36A� (E). Cells were then washed of input virus, and equivalent protein expression wasconfirmed by immunoblotting of whole-cell lysates for viral capsid. Accumulation of newly formed particles in supernatants (Sup) was assessed by immuno-blotting. FIVPPR-infected cat serum was used for the immunoblots in panels D and E.
Morrison et al.
3262 jvi.asm.org Journal of Virology
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

hsTetherin and was completely unaffected by cfTetherin. To de-termine whether this phenotype was strain/clade-specific or a uni-versal mechanism of FIV-mediated tetherin antagonism, a clade Cvirus, FIVC36, which is highly pathogenic in vivo (74, 75), was alsotested in single-round particle release assays. It was even less sen-sitive to tetherin than FIVPPR, with no budding differences in teth-erin-expressing cells compared to controls (Fig. 5E).
Feline tetherin minimally restricts spreading FIV replica-tion. We next examined spreading replication of FIV, usingFIV34TF10, which utilizes the entry receptor CXCR4 (52, 76). Weemployed versions of FIV34TF10 (Fig. 4B) with or without repair ofthe premature stop codon in the OrfA accessory gene (55).Whereas stable expression of the restriction factor rhesus TRIMCypreduced viral reverse transcriptase production at all postchallengetime points by approximately 2 log (Fig. 6A), ectopic tetherinexpression reduced peak RT production marginally and did not
delay the time course (Fig. 6B and C). In addition, syncytia formedequivalently in the presence of each of the tetherins (data notshown). OrfA, the only accessory gene yet identified in FIV besidesRev, is required for efficient FIV replication in primary PBMCs,but the mechanism remains unclear (55, 77–83). Replication ofthe parental OrfA(�) FIV34TF10 virus was similar to the OrfA(�)virus, which indicates that OrfA is not an antitetherin (Fig. 6Band C).
Tetherin antagonism by FIV maps to Env. Given the clearphenotypic disparities that we observed between budding of thefull-length viruses and FIV VLPs or FIV vectors that lack genesbesides Gag, Gag-Pol, and Rev, as well as the ability of FIV toundergo spreading replication in carnivore tetherin-expressingcells, we proceeded to map the source of tetherin antagonism inthe FIV genome. A series of viruses with different complements ofviral proteins were compared in single-cycle budding assays. Wecompared virions generated with pFP93 (which encodes onlyFIV34TF10-derived FIV Gag, Gag-Pol, and Rev) with those gener-ated by pCT5efs, which expresses all FIV34TF10 viral proteins ex-cept the envelope glycoprotein, due to a single frameshift in env(84). Additionally, we constructed analogous FIVPPR and FIVC36
expression plasmids similarly enabled for producing FIV in hu-man cells by substituting their 5= long terminal repeat (LTR) U3elements with the human cytomegalovirus immediate early genepromoter (Fig. 4B). Particle production and release were com-pared in transfected 293T cells. Normalized to tetherin-negativecells, virions produced by pFP93 and pCT5efs were each impaired,by all tetherins tested, for mature particle release (Fig. 7).
The env-intact full-length FIVs had markedly different pheno-types. Their particle formation was impaired in the presence ofhsTetherin, but enhanced viral budding was actually consistentlyobserved in the presence of cfTetherin and fcTetherinSF (Fig. 7A toC). As in the FIV34TF10 spreading replication experiments,OrfA(�) and OrfA(�) versions of FIVC36 displayed approxi-mately equivalent sensitivities to tetherin (Fig. 7A). The enhancedbudding of replication-competent FIV was dependent on the teth-erin being of carnivore origin; both hsTetherin and an artificialtetherin (artTetherin; see reference 27) blocked budding of FIVPPR
and FIVC36 (Fig. 7B and C). These results strongly implicated FIVEnv as the mediator of carnivore tetherin antagonism, so we nextdirectly tested this hypothesis by expressing Env in trans.
FIV Env antagonizes tetherin to enable FIV, but not HIV-1,particle production. We devised plasmids that coexpress Rev andthe env alleles of FIVPPR or FIVC36 by replacing the Gag-Pol genesof pFP93 with the respective env segments (Fig. 8A). Both PPR andC36 Env were processed similarly when expressed alone or in thecontext of the full-length viral clones in 293T cells (Fig. 8B). Ex-pression of both FIVPPR and FIVC36 Env caused syncytia in a pri-mary-receptor-dependent manner in 293T cells (Fig. 8C), dem-onstrating that the proteins produced are functional.
Viral release assays were performed by cotransfectingpCT5efsA� with different Env expression plasmids. Supernatantaccumulation of bald or VSV-G-pseudotyped particles was mark-edly reduced in cells expressing any of the three mammalian teth-erins (Fig. 8D). Particles pseudotyped with FIVPPR Env remainedsensitive to hsTetherin restriction, with substantially decreasedyields in mature particle formation and supernatant capsid accu-mulation. In contrast, neither cfTetherin nor fcTetherinSF had anyeffect on FIVPPR Env-pseudotyped virion egress. Moreover, su-pernatant CA yields were significantly higher for these Env-bear-
FIG 6 Effect of tetherin on FIV spreading replication. (A) FIV34TF10A� wasused to infect CrFK cells stably expressing Rhesus TrimCyp or control cells.CrFK cells stably expressing tetherin were infected with FIV34TF10A� (B) orFIV34TF10 (C), and viral replication was monitored by accumulation of RTactivity in the supernatant.
Mechanism of Tetherin Antagonism
March 2014 Volume 88 Number 6 jvi.asm.org 3263
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

ing vectors in the presence of either carnivore tetherin, demon-strating again the synergistic augmentation effect that FIV Envand tetherin confer to FIV particle release (Fig. 8D). FIVC36 Env-pseudotyped vector particles also displayed this pattern of in-creased supernatant capsid accumulation and equivalent reversetranscriptase activity in cfTetherin- and fcTetherinSF-expressingcells (Fig. 8D).
Primate lentiviral tetherin antagonists (Vpu, Nef, and Env)principally act to remove tetherin from sites of virus release at thecell surface, although the mechanisms may vary (10, 85). Thus,these antagonists can rescue particle release of other envelopedviruses as well as the virus that encodes them. Both SIVtan andHIV-2 Env can rescue HIV-1 particles in trans by downregulatingcell surface tetherin, primarily by sequestering it within intracel-lular compartments (3, 17, 18). To test whether FIV Env acts sim-ilarly, we examined HIV-1 budding. Surprisingly, in contrast totheir effects on FIV, the FIV envelope glycoproteins were unable torescue HIV-1 particle release (Fig. 8E). These results raise the in-teresting possibility that unlike HIV-2 Env (3, 18), incorporationof FIV Env into the particle is required to activate the tetherinevasion mechanism.
To confirm that FIV Env is not incorporated functionally intoHIV-1 particles, HIV-1 and FIV reporter viruses were pseu-dotyped and their titers were determined on feline cells that ex-press the FIV primary receptor CD134 (Table 1). These experi-ments yielded clear results that were moreover strongly consistentbetween the two FIV Env glycoproteins: PPR Env and C36 Envpseudotyping produced FIV particles that had 343-fold- and 338-fold-greater FIV Env/VSV-G infectivity ratios, respectively, thanHIV-1 particles (Table 1).
FIV Env does not modulate intracellular levels or cell surfacetrafficking of tetherin. We found that CrFK cells infected with
FIVPPR (introduced into these cells via VSV-G pseudotyping toestablish proviruses) expressed FIV well (flow cytometry data areshown in Fig. 9; immunoblotting is not shown), but changes intotal intracellular tetherin levels compared to uninfected cellswere not evident (Fig. 9A and B). FIVC36 produced a minor de-crease in bulk carnivore tetherin intracellular levels but did notaffect human tetherin (Fig. 9A and B). We considered that FIVC36
may specifically deplete the cell surface pool of tetherin or other-wise prevent its accumulation at the cell surface, as has been doc-umented for HIV-1 Vpu (5, 86–88) and HIV-2 Env (3). Therefore,we specifically imaged cell surface tetherin in nonpermeabilizedcells. However, in cells transfected with full-length FIV plasmids,normal trafficking to the cell surface was observed for tetherinproteins from all three species (Fig. 9C). Transient expression ofHIV-1 Vpu decreased cell surface levels of hsTetherin, whereasFIV Env, either alone or in the presence of all viral proteins, didnot alter the cell surface pool of fcTetherinSF (Fig. 9D).
For HIV-2 Env, tetherin antagonism requires endocytic mo-tifs in the envelope transmembrane (TM) domain cytoplasmictail, which is consistent with the cell-surface downregulationmechanism (3, 18). While the FIV TM cytoplasmic tail is sig-nificantly shorter than those found in the primate lentiviruses(89), it does contain the conserved GYXX� endocytosis motifalso present in all primate lentivirus Envs (90). The lack oftetherin downregulation that we observed suggests that FIVEnv endocytic sorting is dispensable for tetherin antagonism.To test directly the role of cytoplasmic tail elements, a prema-ture stop codon was introduced into FIVC36 Env (C36 TM�43),removing a majority of the cytoplasmic tail (43 of 53 aminoacids) including the GYTVI endocytosis motif. A virus with afurin cleavage site mutation was also constructed (C36 furinmutant). Wild-type and Env-mutant FIVC36 viruses were pro-
FIG 7 Env is required for efficient budding of FIV in the presence of tetherin. (A) 293T cells were transfected with the indicated FIV packaging constructs thateither encode an intact Env (pCT-PPR, pCT-C36A�, pCT-C36) or have a disrupted Env open reading frame (pFP93, pCT5A�efs). Cell lysates and supernatantswere immunoblotted as indicated. Supernatant RT activity is normalized to tetherin-negative cells. (B) The experiment was performed as described for panel A,with the addition of artificial tetherin (Art).
Morrison et al.
3264 jvi.asm.org Journal of Virology
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

duced in control and fcTetherinSF-expressing 293T cells, andthe resulting supernatants were tested for reverse transcriptaseactivity and capsid accumulation. Despite low intracellular Envlevels, removal of the TM cytoplasmic tail did not reduce Envprotection of FIVC36 budding in the presence of fcTetherinSF
(Fig. 10). Mutation of the furin cleavage site resulted in accu-
mulation of only the uncleaved Env precursor. Notably, how-ever, this virus budded as efficiently as wild-type virus in thepresence of tetherin (Fig. 10). These results demonstrate thatFIV Env antagonism of fcTetherin does not proceed via the cellsurface depletion mechanism used by primate lentiviral teth-erin antagonists. Moreover, neither the cytoplasmic tail endo-
FIG 8 Expression of FIV Env in trans rescues budding of FIV in tetherin-expressing cells. (A) Diagram of FIVPPR and FIVC36 Env expression constructs. (B)Immunoblot of 293T cells transfected with FIV packaging constructs or FIV Env expression constructs. (C) 293T cells were cotransfected with expressionconstructs for EGFP, FIV Env, and either empty vector or feline CD134. (D) Immunoblotting and RT activity of cell lysates and supernatants from 293T cellsstably expressing tetherin cotransfected with an Env-disrupted FIV packaging (pCT5efsA�) and envelope expression constructs. (E) Cells were treated asdescribed for panel D with an Env-disrupted HIV-1 packaging construct (pHIV-1Luc �Vpu) and anti-HIV-1 p24 antibodies.
TABLE 1 Infectivity comparisons for Env pseudotypes
Virus Envelope Titera (SD) RT activityb (SD)Infectivity/RT ratio
VSV-G/FIV Envinfectivity ratio
Fold infectivity ratiodeficit for HIV-1
HIV-1 None 0.0 (0.0) 8.3 106 (2.9 105) 0.0HIV-1 VSV-G 1.8 107 (4.5 106) 8.1 106 (1.1 106) 2.2HIV-1 FIVPPR 9.2 103 (3.1 103) 1.3 107 (1.1 106) 7.1 10�4 1,957 343HIV-1 FIVC36 1.9 104 (6.6 103) 8.5 106 (1.2 106) 2.2 10�3 947 338FIV None 0.0 (0.0) 1.1 107 (5.7 105) 0.0FIV VSV-G 2.4 106 (2.1 105) 9.2 106 (4.2 105) 2.6 10�1
FIV FIVPPR 4.2 105 (7.2 104) 8.9 106 (7.4 105) 4.7 10�2 5.7FIV FIVC36 8.7 105 (1.9 105) 8.7 106 (3.1 105) 1.0 10�1 2.8a Values are in transducing units (TU)/ml.b RT, reverse transcriptase; values are in cpm/ml.
Mechanism of Tetherin Antagonism
March 2014 Volume 88 Number 6 jvi.asm.org 3265
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

FIG 9 Analysis of tetherin antagonism by FIV. (A) The ability of FIV to perturb intracellular levels of tetherin was determined by infecting CrFK cells with FIVand performing dual-color FACS for both viral protein expression (anti-FIVPPR) and tetherin expression (anti-HA). (B) Average anti-HA mean fluorescenceintensity (MFI) of FIV-positive and -negative cells was calculated. (C) Immunofluorescence analysis of 293T cells transfected with indicated expressionconstructs and immunolabeled, without prior permeabilization, for tetherin (anti-HA, red), FIV Env (anti-FIVPPR, green), and DNA (DAPI, blue). TheDAPI-positive, red/green-negative cells demonstrate specificity of labeling. (D) Cell surface tetherin expression (anti-HA) is displayed for GFP� 293T cells stablyexpressing tetherin-HA transfected with pEGFP as well as empty vector or expression plasmids for codon-optimized HIV-1 vpu (vphu), FIV Env, or full-lengthFIV constructs.
Morrison et al.
3266 jvi.asm.org Journal of Virology
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
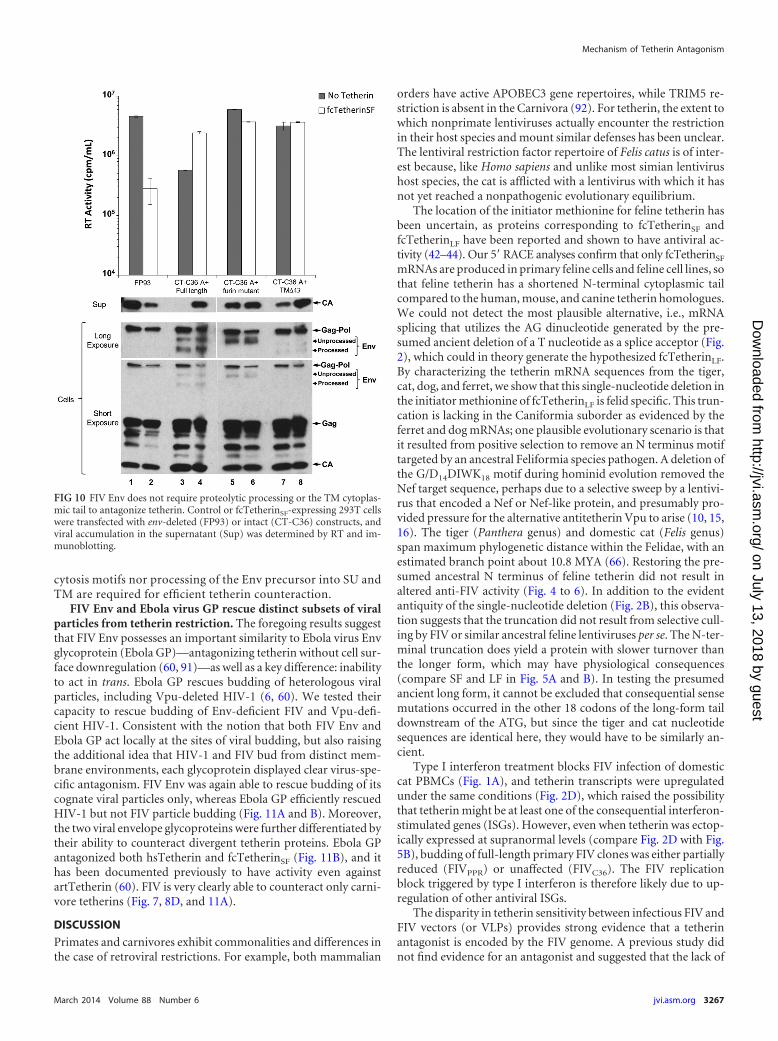
cytosis motifs nor processing of the Env precursor into SU andTM are required for efficient tetherin counteraction.
FIV Env and Ebola virus GP rescue distinct subsets of viralparticles from tetherin restriction. The foregoing results suggestthat FIV Env possesses an important similarity to Ebola virus Envglycoprotein (Ebola GP)—antagonizing tetherin without cell sur-face downregulation (60, 91)—as well as a key difference: inabilityto act in trans. Ebola GP rescues budding of heterologous viralparticles, including Vpu-deleted HIV-1 (6, 60). We tested theircapacity to rescue budding of Env-deficient FIV and Vpu-defi-cient HIV-1. Consistent with the notion that both FIV Env andEbola GP act locally at the sites of viral budding, but also raisingthe additional idea that HIV-1 and FIV bud from distinct mem-brane environments, each glycoprotein displayed clear virus-spe-cific antagonism. FIV Env was again able to rescue budding of itscognate viral particles only, whereas Ebola GP efficiently rescuedHIV-1 but not FIV particle budding (Fig. 11A and B). Moreover,the two viral envelope glycoproteins were further differentiated bytheir ability to counteract divergent tetherin proteins. Ebola GPantagonized both hsTetherin and fcTetherinSF (Fig. 11B), and ithas been documented previously to have activity even againstartTetherin (60). FIV is very clearly able to counteract only carni-vore tetherins (Fig. 7, 8D, and 11A).
DISCUSSION
Primates and carnivores exhibit commonalities and differences inthe case of retroviral restrictions. For example, both mammalian
orders have active APOBEC3 gene repertoires, while TRIM5 re-striction is absent in the Carnivora (92). For tetherin, the extent towhich nonprimate lentiviruses actually encounter the restrictionin their host species and mount similar defenses has been unclear.The lentiviral restriction factor repertoire of Felis catus is of inter-est because, like Homo sapiens and unlike most simian lentivirushost species, the cat is afflicted with a lentivirus with which it hasnot yet reached a nonpathogenic evolutionary equilibrium.
The location of the initiator methionine for feline tetherin hasbeen uncertain, as proteins corresponding to fcTetherinSF andfcTetherinLF have been reported and shown to have antiviral ac-tivity (42–44). Our 5= RACE analyses confirm that only fcTetherinSF
mRNAs are produced in primary feline cells and feline cell lines, sothat feline tetherin has a shortened N-terminal cytoplasmic tailcompared to the human, mouse, and canine tetherin homologues.We could not detect the most plausible alternative, i.e., mRNAsplicing that utilizes the AG dinucleotide generated by the pre-sumed ancient deletion of a T nucleotide as a splice acceptor (Fig.2), which could in theory generate the hypothesized fcTetherinLF.By characterizing the tetherin mRNA sequences from the tiger,cat, dog, and ferret, we show that this single-nucleotide deletion inthe initiator methionine of fcTetherinLF is felid specific. This trun-cation is lacking in the Caniformia suborder as evidenced by theferret and dog mRNAs; one plausible evolutionary scenario is thatit resulted from positive selection to remove an N terminus motiftargeted by an ancestral Feliformia species pathogen. A deletion ofthe G/D14DIWK18 motif during hominid evolution removed theNef target sequence, perhaps due to a selective sweep by a lentivi-rus that encoded a Nef or Nef-like protein, and presumably pro-vided pressure for the alternative antitetherin Vpu to arise (10, 15,16). The tiger (Panthera genus) and domestic cat (Felis genus)span maximum phylogenetic distance within the Felidae, with anestimated branch point about 10.8 MYA (66). Restoring the pre-sumed ancestral N terminus of feline tetherin did not result inaltered anti-FIV activity (Fig. 4 to 6). In addition to the evidentantiquity of the single-nucleotide deletion (Fig. 2B), this observa-tion suggests that the truncation did not result from selective cull-ing by FIV or similar ancestral feline lentiviruses per se. The N-ter-minal truncation does yield a protein with slower turnover thanthe longer form, which may have physiological consequences(compare SF and LF in Fig. 5A and B). In testing the presumedancient long form, it cannot be excluded that consequential sensemutations occurred in the other 18 codons of the long-form taildownstream of the ATG, but since the tiger and cat nucleotidesequences are identical here, they would have to be similarly an-cient.
Type I interferon treatment blocks FIV infection of domesticcat PBMCs (Fig. 1A), and tetherin transcripts were upregulatedunder the same conditions (Fig. 2D), which raised the possibilitythat tetherin might be at least one of the consequential interferon-stimulated genes (ISGs). However, even when tetherin was ectop-ically expressed at supranormal levels (compare Fig. 2D with Fig.5B), budding of full-length primary FIV clones was either partiallyreduced (FIVPPR) or unaffected (FIVC36). The FIV replicationblock triggered by type I interferon is therefore likely due to up-regulation of other antiviral ISGs.
The disparity in tetherin sensitivity between infectious FIV andFIV vectors (or VLPs) provides strong evidence that a tetherinantagonist is encoded by the FIV genome. A previous study didnot find evidence for an antagonist and suggested that the lack of
FIG 10 FIV Env does not require proteolytic processing or the TM cytoplas-mic tail to antagonize tetherin. Control or fcTetherinSF-expressing 293T cellswere transfected with env-deleted (FP93) or intact (CT-C36) constructs, andviral accumulation in the supernatant (Sup) was determined by RT and im-munoblotting.
Mechanism of Tetherin Antagonism
March 2014 Volume 88 Number 6 jvi.asm.org 3267
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

restriction to spreading FIV replication is due to enhanced cell-to-cell spread (42). Consistent with observations by Celestino et al.on fcTetherinSF (44), we determined that when supplied in trans,FIV Env antagonizes both fcTetherinSF and C. familiaris tetherin.However, we show that this is only for cognate (FIV) particles (Fig.8D). Celestino and colleagues also reported that fcTetherinSF iscoprecipitated from cells by antibody to FIV Env (44). A singleclade A strain (FIVPetaluma) was tested, and the mechanism for Envantagonism was not further explored. We demonstrate here thatthis Env-specific antagonism is conserved between clades, usingFIVPPR (clade A) and FIVC36 (clade C). While the absence of spe-cific antibodies presently hinders determination of the tissue ex-pression profile of tetherin in the cat in vivo, the observation by usand others (44) that FIV envelope glycoprotein can act as an ef-fective antagonist suggests that FIV indeed encounters and evadestetherin in vivo.
We additionally report here enhanced budding of env-intactbut not env-deficient FIV in the presence of tetherin (Fig. 7). Thisstriking phenomenon, which suggests that FIV has evolved to co-opt tetherin, is consistent with our finding that FIV does not evaderestriction by degrading the protein or removing it from the cellsurface. Instead, we show that evasion of tetherin restriction re-quires specific incorporation of FIV Env into FIV particles. Envcannot act in trans to rescue noncognate particles. We proposethat FIV envelope glycoproteins are triggered to engage tetherincoincident with or after virion incorporation, thereby shielding
the nascent particle. The degree of restriction antagonism by Envcorrelates with the efficiency of viral particle incorporation. Wenote that tetherin enhanced budding most dramatically for FIVsthat express their native Env proteins (Fig. 7). Pseudotyping ofFIV34TF10 particles with FIVPPR and FIVC36 Envs resulted in anintermediate phenotype whereby mature particle formation wascomparable to control (tetherin-absent) levels, and capsid levelsin supernatant were still increased in the presence of fcTetherin(Fig. 8D). In contrast, HIV-1 is minimally pseudotypeable by FIVenvelope glycoproteins (Table 1) and was not rescued in trans byFIVPPR or FIVC36 envelope (Fig. 8E). The envelope glycoproteinsof SIVtan, HIV-2, and Ebola virus are tetherin antagonists, butexpression of each of these envelope glycoproteins rescues bud-ding of HIV-1 (3, 6, 18, 60). This difference demarcates theseproteins from FIV Env, which rescues budding of cognate parti-cles only. Some main properties of FIV Env versus other antago-nists are summarized in Table 2.
Examining the interplay between FIV Env and tetherin alsorevealed an interesting negative impact of FIV Env expression onFIV budding. This phenomenon is most clearly seen in Fig. 10(compare lane 3 with lanes 1 and 5). Budding of env-intact FIVC36
was significantly reduced compared to env-deficient virions(FP93), virions defective for SU-TM cleavage, and virions with adeletion of the TM cytoplasmic domain (Fig. 10, lanes 1 and 7).Importantly, expression of tetherin restores budding of full-length FIVC36 to levels observed in the absence of Env, suggesting
FIG 11 Ebola virus GP and FIV Env display virus-specific tetherin counteraction. Control or tetherin-expressing 293T cells were cotransfected with either FP93(A) or pHIV-1Luc �Vpu (B) and the indicated viral glycoprotein expression plasmid or a control vector. Appropriate expression of viral Gag and Gag-Pol,glycoprotein, and tetherin was confirmed by immunoblotting the cell lysate, and viral budding was again determined by supernatant (Sup) RT and capsidaccumulation.
Morrison et al.
3268 jvi.asm.org Journal of Virology
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

that the two phenotypes are linked. We speculate that FIV Envmay constrain budding of FIV particles in situations where teth-erin expression is low, yet it may serve to enable efficient buddingin cells expressing high levels of tetherin. Tetherin would by thisreasoning act akin to a licensing factor for FIV, helping to main-tain a balance of cell-free and cell-associated particle formationthat promotes viral spread in the in vivo environment. In thisscenario, FIV Env has evolved to both evade and exploit this pro-tein, allowing us to suggest that tetherin is both a restriction factorand a cofactor for this lentivirus.
The observation that FIV is able to counteract both cfTetherinand fcTetherin but remains sensitive to hsTetherin and the com-pletely artificial tetherin (Fig. 7 and 8) implies that Env may inter-act directly or indirectly with fcTetherin, which is supported bythe previous observation that FIV Env coimmunoprecipitatedfrom cells with tetherin (44). Combined with the evidence thatFIV, unlike primate lentiviruses, does not significantly degradetetherin or downregulate it from the cell surface (Fig. 9), theseresults suggest that Env is acting locally on a per-particle basis atthe point of viral budding to affect tetherin sensitivity, rather thanexerting a cell-wide phenotype. FIV Env-directed assembly mayexclude tetherin from virions or virion assembly sites or directassembly to tetherin-negative membrane domains. This “local”point-of-assembly model may also apply to Ebola virus GP, whichhas previously been described to counteract tetherin without re-moving it from the cell surface (60, 91). There are important dif-ferences however, in that that Ebola virus GP and FIV Env selec-tively rescued only HIV-1 or FIV, respectively. While Ebola virusGP pseudotypes HIV-1 and FIV well (93, 94), FIV Env pseu-dotypes only FIV (Table 1). Thus, the mirrored tetherin antago-nism phenotypes of these two glycoproteins do not depend on thevariable of particle incorporation alone. Investigating how FIVEnv distinguishes and rescues only cognate particles while coop-erating in particle formation at the plasma membrane has thepotential to advance understanding of retroviral assembly andinnate immunity circumvention.
ACKNOWLEDGMENTS
We thank the Minnesota Zoo and T. Wolf for tiger blood, V. von Messlingfor ferret blood, J. Guatelli for a human tetherin cDNA, M. Peretz fortechnical assistance, K. Strebel for pNL4.3�Vpu, M. Barr and S. Vande-woude for feline sera, B. Willet for pDON-AI feline CD134 and cell lines,J. Elder and the NIH AIDS Research and Reference Reagent Program forFIVC36 and FIVPPR clones, P. Cannon for pEbolaGPmyc, P. Bates forEbola virus glycoprotein antiserum, P. Bieniasz for pLPCX-artTetherin, D. Sauter and F. Kirchhoff for human tetherin, HIV-M Vpu,and NF-�B reporter plasmids, and J. Yamamoto and C. Münk for celllines.
The work was supported by NIH grants AI77344 and AI100797 toE.M.P. An institutional R25 PREP grant (GM075148) helped supportR. B. Guevara’s stipend.
REFERENCES1. Groom HC, Yap MW, Galao RP, Neil SJ, Bishop KN. 2010. Suscepti-
bility of xenotropic murine leukemia virus-related virus (XMRV) to ret-roviral restriction factors. Proc. Natl. Acad. Sci. U. S. A. 107:5166 –5171.http://dx.doi.org/10.1073/pnas.0913650107.
2. Jouvenet N, Neil SJ, Zhadina M, Zang T, Kratovac Z, Lee Y, McNatt M,Hatziioannou T, Bieniasz PD. 2009. Broad-spectrum inhibition of ret-roviral and filoviral particle release by tetherin. J. Virol. 83:1837–1844.http://dx.doi.org/10.1128/JVI.02211-08.
3. Le Tortorec A, Neil SJ. 2009. Antagonism to and intracellular sequestra-tion of human tetherin by the human immunodeficiency virus type 2envelope glycoprotein. J. Virol. 83:11966 –11978. http://dx.doi.org/10.1128/JVI.01515-09.
4. Neil SJ, Zang T, Bieniasz PD. 2008. Tetherin inhibits retrovirus releaseand is antagonized by HIV-1 Vpu. Nature 451:425– 430. http://dx.doi.org/10.1038/nature06553.
5. Van Damme N, Goff D, Katsura C, Jorgenson RL, Mitchell R, JohnsonMC, Stephens EB, Guatelli J. 2008. The interferon-induced proteinBST-2 restricts HIV-1 release and is downregulated from the cell surfaceby the viral Vpu protein. Cell Host Microbe 3:245–252. http://dx.doi.org/10.1016/j.chom.2008.03.001.
6. Kaletsky RL, Francica JR, Agrawal-Gamse C, Bates P. 2009. tetherin-mediated restriction of filovirus budding is antagonized by the Ebola gly-coprotein. Proc. Natl. Acad. Sci. U. S. A. 106:2886 –2891. http://dx.doi.org/10.1073/pnas.0811014106.
7. Sakuma T, Noda T, Urata S, Kawaoka Y, Yasuda J. 2009. Inhibition ofLassa and Marburg virus production by tetherin. J. Virol. 83:2382–2385.http://dx.doi.org/10.1128/JVI.01607-08.
8. Mansouri M, Viswanathan K, Douglas JL, Hines J, Gustin J, Moses AV,Fruh K. 2009. Molecular mechanism of BST2/tetherin downregulation byK5/MIR2 of Kaposi’s sarcoma-associated herpesvirus. J. Virol. 83:9672–9681. http://dx.doi.org/10.1128/JVI.00597-09.
9. Douglas JL, Gustin JK, Viswanathan K, Mansouri M, Moses AV, FruhK. 2010. The great escape: viral strategies to counter BST-2/tetherin. PLoSPathog. 6:e1000913. http://dx.doi.org/10.1371/journal.ppat.1000913.
10. Evans DT, Serra-Moreno R, Singh RK, Guatelli JC. 2010. BST-2/tetherin: a new component of the innate immune response to envelopedviruses. Trends Microbiol. 18:388 –396. http://dx.doi.org/10.1016/j.tim.2010.06.010.
11. Gupta RK, Towers GJ. 2009. A tail of tetherin: how pandemic HIV-1conquered the world. Cell Host Microbe 6:393–395. http://dx.doi.org/10.1016/j.chom.2009.11.002.
12. Lim ES, Emerman M. 2009. Simian immunodeficiency virus SIVagmfrom African green monkeys does not antagonize endogenous levels ofAfrican green monkey tetherin/BST-2. J. Virol. 83:11673–11681. http://dx.doi.org/10.1128/JVI.00569-09.
13. Sauter D, Schindler M, Specht A, Landford WN, Munch J, Kim KA,Votteler J, Schubert U, Bibollet-Ruche F, Keele BF, Takehisa J, OgandoY, Ochsenbauer C, Kappes JC, Ayouba A, Peeters M, Learn GH, ShawG, Sharp PM, Bieniasz P, Hahn BH, Hatziioannou T, Kirchhoff F. 2009.Tetherin-driven adaptation of Vpu and Nef function and the evolution ofpandemic and nonpandemic HIV-1 strains. Cell Host Microbe 6:409 –421. http://dx.doi.org/10.1016/j.chom.2009.10.004.
14. Yang SJ, Lopez LA, Hauser H, Exline CM, Haworth KG, Cannon PM.2010. Anti-tetherin activities in Vpu-expressing primate lentiviruses. Ret-rovirology 7:13. http://dx.doi.org/10.1186/1742-4690-7-13.
15. Jia B, Serra-Moreno R, Neidermyer W, Rahmberg A, Mackey J, FofanaIB, Johnson WE, Westmoreland S, Evans DT. 2009. Species-specificactivity of SIV Nef and HIV-1 Vpu in overcoming restriction by tetherin/BST2. PLoS Pathog. 5:e1000429. http://dx.doi.org/10.1371/journal.ppat.1000429.
16. Zhang F, Wilson SJ, Landford WC, Virgen B, Gregory D, Johnson MC,Munch J, Kirchhoff F, Bieniasz PD, Hatziioannou T. 2009. Nef proteinsfrom simian immunodeficiency viruses are tetherin antagonists. Cell HostMicrobe 6:54 – 67. http://dx.doi.org/10.1016/j.chom.2009.05.008.
17. Gupta RK, Mlcochova P, Pelchen-Matthews A, Petit SJ, Mattiuzzo G,Pillay D, Takeuchi Y, Marsh M, Towers GJ. 2009. Simian immunode-ficiency virus envelope glycoprotein counteracts tetherin/BST-2/CD317by intracellular sequestration. Proc. Natl. Acad. Sci. U. S. A. 106:20889 –20894. http://dx.doi.org/10.1073/pnas.0907075106.
18. Hauser H, Lopez LA, Yang SJ, Oldenburg JE, Exline CM, Guatelli JC,Cannon PM. 2010. HIV-1 Vpu and HIV-2 Env counteract BST-2/tetherin
TABLE 2 Properties of FIV Env versus other viral tetherin antagonists
Antagonist
Species-specificaction
Cell surfacedownregulationof tetherin
Activity requirescognate particleincorporation
RescuesHIV-1budding
Dependency ontetherin foroptimal particlerelease
HIV-1 Vpu � � � � �SIV Nef � � � � �HIV-2 Env � � � � �FIV Env � � � � �Ebola Env � � � � �
Mechanism of Tetherin Antagonism
March 2014 Volume 88 Number 6 jvi.asm.org 3269
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

by sequestration in a perinuclear compartment. Retrovirology 7:51. http://dx.doi.org/10.1186/1742-4690-7-51.
19. Gottlinger HG, Dorfman T, Cohen EA, Haseltine WA. 1993. Vpuprotein of human immunodeficiency virus type 1 enhances the release ofcapsids produced by gag gene constructs of widely divergent retroviruses.Proc. Natl. Acad. Sci. U. S. A. 90:7381–7385. http://dx.doi.org/10.1073/pnas.90.15.7381.
20. Klimkait T, Strebel K, Hoggan MD, Martin MA, Orenstein JM. 1990.The human immunodeficiency virus type 1-specific protein vpu is re-quired for efficient virus maturation and release. J. Virol. 64:621– 629.
21. Rollason R, Korolchuk V, Hamilton C, Schu P, Banting G. 2007.Clathrin-mediated endocytosis of a lipid-raft-associated protein is medi-ated through a dual tyrosine motif. J. Cell Sci. 120:3850 –3858. http://dx.doi.org/10.1242/jcs.003343.
22. Andrew AJ, Miyagi E, Kao S, Strebel K. 2009. The formation of cysteine-linked dimers of BST-2/tetherin is important for inhibition of HIV-1 virusrelease but not for sensitivity to Vpu. Retrovirology 6:80. http://dx.doi.org/10.1186/1742-4690-6-80.
23. Ohtomo T, Sugamata Y, Ozaki Y, Ono K, Yoshimura Y, Kawai S,Koishihara Y, Ozaki S, Kosaka M, Hirano T, Tsuchiya M. 1999. Mo-lecular cloning and characterization of a surface antigen preferentiallyoverexpressed on multiple myeloma cells. Biochem. Biophys. Res. Com-mun. 258:583–591. http://dx.doi.org/10.1006/bbrc.1999.0683.
24. Kupzig S, Korolchuk V, Rollason R, Sugden A, Wilde A, Banting G.2003. Bst-2/HM1.24 is a raft-associated apical membrane protein with anunusual topology. Traffic 4:694 –709. http://dx.doi.org/10.1034/j.1600-0854.2003.00129.x.
25. Fitzpatrick K, Skasko M, Deerinck TJ, Crum J, Ellisman MH, GuatelliJ. 2010. Direct restriction of virus release and incorporation of the inter-feron-induced protein BST-2 into HIV-1 particles. PLoS Pathog.6:e1000701. http://dx.doi.org/10.1371/journal.ppat.1000701.
26. Habermann A, Krijnse-Locker J, Oberwinkler H, Eckhardt M, HomannS, Andrew A, Strebel K, Krausslich HG. 2010. CD317/tetherin is en-riched in the HIV-1 envelope and downregulated from the plasma mem-brane upon virus infection. J. Virol. 84:4646 – 4658. http://dx.doi.org/10.1128/JVI.02421-09.
27. Perez-Caballero D, Zang T, Ebrahimi A, McNatt MW, Gregory DA,Johnson MC, Bieniasz PD. 2009. Tetherin inhibits HIV-1 release bydirectly tethering virions to cells. Cell 139:499 –511. http://dx.doi.org/10.1016/j.cell.2009.08.039.
28. Venkatesh S, Bieniasz PD. 2013. Mechanism of HIV-1 virion entrapmentby tetherin. PLoS Pathog. 9:e1003483. http://dx.doi.org/10.1371/journal.ppat.1003483.
29. Fadel H, Saenz D, Guevara R, Peretz M, Poeschla E. 2012. Productivereplication and evolution of HIV-1 in ferret cells. J. Virol. 86:2312–2322.
30. McEwan WA, Schaller T, Ylinen LM, Hosie MJ, Towers GJ, Willett BJ.2009. Truncation of TRIM5 in the Feliformia explains the absence of ret-roviral restriction in cells of the domestic cat. J. Virol. 16:8270 – 8275. http://dx.doi.org/10.1128/JVI.00670-09.
31. Sawyer SL, Emerman M, Malik HS. 2007. Discordant evolution of theadjacent antiretroviral genes TRIM22 and TRIM5 in mammals. PLoS Pat-hog. 3:e197. http://dx.doi.org/10.1371/journal.ppat.0030197.
32. Sayah DM, Sokolskaja E, Berthoux L, Luban J. 2004. Cyclophilin Aretrotransposition into TRIM5 explains owl monkey resistance to HIV-1.Nature 430:569 –573. http://dx.doi.org/10.1038/nature02777.
33. Stremlau M, Owens CM, Perron MJ, Kiessling M, Autissier P, SodroskiJ. 2004. The cytoplasmic body component TRIM5alpha restricts HIV-1infection in Old World monkeys. Nature 427:848 – 853. http://dx.doi.org/10.1038/nature02343.
34. Virgen CA, Kratovac Z, Bieniasz PD, Hatziioannou T. 2008. Indepen-dent genesis of chimeric TRIM5-cyclophilin proteins in two primate spe-cies. Proc. Natl. Acad. Sci. U. S. A. 105:3563–3568. http://dx.doi.org/10.1073/pnas.0709258105.
35. Wilson SJ, Webb BL, Ylinen LM, Verschoor E, Heeney JL, Towers GJ.2008. Independent evolution of an antiviral TRIMCyp in rhesus ma-caques. Proc. Natl. Acad. Sci. U. S. A. 105:3557–3562. http://dx.doi.org/10.1073/pnas.0709003105.
36. Best S, Le Tissier P, Towers G, Stoye JP. 1996. Positional cloning of themouse retrovirus restriction gene Fv1. Nature 382:826 – 829. http://dx.doi.org/10.1038/382826a0.
37. Lochelt M, Romen F, Bastone P, Muckenfuss H, Kirchner N, Kim YB,Truyen U, Rosler U, Battenberg M, Saib A, Flory E, Cichutek K, MunkC. 2005. The antiretroviral activity of APOBEC3 is inhibited by the foamy
virus accessory Bet protein. Proc. Natl. Acad. Sci. U. S. A. 102:7982–7987.http://dx.doi.org/10.1073/pnas.0501445102.
38. Münk C, Beck T, Zielonka J, Hotz-Wagenblatt A, Chareza S, BattenbergM, Thielebein J, Cichutek K, Bravo IG, O’Brien SJ, Lochelt M, Yuhki N.2008. Functions, structure, and read-through alternative splicing of felineAPOBEC3 genes. Genome Biol. 9:R48. http://dx.doi.org/10.1186/gb-2008-9-3-r48.
39. Münk C, Zielonka J, Constabel H, Kloke BP, Rengstl B, Battenberg M,Bonci F, Pistello M, Lochelt M, Cichutek K. 2007. Multiple restrictionsof human immunodeficiency virus type 1 in feline cells. J. Virol. 81:7048 –7060. http://dx.doi.org/10.1128/JVI.02714-06.
40. Stern MA, Hu C, Saenz DT, Fadel HJ, Sims O, Peretz M, Poeschla EM.2010. Productive replication of Vif-chimeric HIV-1 in feline cells. J. Virol.84:7378 –7395. http://dx.doi.org/10.1128/JVI.00584-10.
41. Zielonka J, Marino D, Hofmann H, Yuhki N, Lochelt M, Munk C. 2010.Vif of feline immunodeficiency virus from domestic cats protects againstAPOBEC3 restriction factors from many felids. J. Virol. 84:7312–7324.http://dx.doi.org/10.1128/JVI.00209-10.
42. Dietrich I, McMonagle EL, Petit SJ, Vijayakrishnan S, Logan N, Chan CN,Towers GJ, Hosie MJ, Willett BJ. 2011. Feline tetherin efficiently restrictsrelease of feline immunodeficiency virus but not spreading of infection. J.Virol. 85:5840–5852. http://dx.doi.org/10.1128/JVI.00071-11.
43. Fukuma A, Abe M, Morikawa Y, Miyazawa T, Yasuda J. 2011. Cloningand characterization of the antiviral activity of feline tetherin/BST-2. PLoSOne 6:e18247. http://dx.doi.org/10.1371/journal.pone.0018247.
44. Celestino M, Calistri A, Del Vecchio C, Salata C, Chiuppesi F, PistelloM, Borsetti A, Palu G, Parolin C. 2012. Feline tetherin is characterized bya short N-terminal region and is counteracted by the feline immunodefi-ciency virus envelope glycoprotein. J. Virol. 86:6688 – 6700. http://dx.doi.org/10.1128/JVI.07037-11.
45. Kirchhoff F. 2009. Is the high virulence of HIV-1 an unfortunate coinci-dence of primate lentiviral evolution? Nat. Rev. Microbiol. 7:467– 476.http://dx.doi.org/10.1038/nrmicro2111.
46. Cheney CM, Rojko JL, Kociba GJ, Wellman ML, Di Bartola SP, Rez-anka LJ, Forman L, Mathes LE. 1990. A feline large granular lymphomaand its derived cell line. In Vitro Cell. Dev. Biol. 26:455– 463. http://dx.doi.org/10.1007/BF02624087.
47. Crandell RA, Fabricant CG, Nelson-Rees WA. 1973. Development,characterization, and viral susceptibility of a feline (Felis catus) renal cellline (CRFK). In Vitro 9:176 –185. http://dx.doi.org/10.1007/BF02618435.
48. Tokunaga K, Nishino Y, Oikawa H, Ishihara C, Mikami T, Ikuta K.1992. Altered cell tropism and cytopathicity of feline immunodeficiencyviruses in two different feline CD4-positive, CD8-negative cell lines. J.Virol. 66:3893–3898.
49. Vanegas M, Llano M, Delgado S, Thompson D, Peretz M, Poeschla E.2005. Identification of the LEDGF/p75 HIV-1 integrase-interaction do-main and NLS reveals NLS-independent chromatin tethering. J. Cell Sci.118:1733–1743. http://dx.doi.org/10.1242/jcs.02299.
50. Meehan AM, Saenz DT, Morrison JH, Garcia-Rivera JA, Peretz M,Llano M, Poeschla EM. 2009. LEDGF/p75 proteins with alternative chro-matin tethers are functional HIV-1 cofactors. PLoS Pathog. 5:e1000522.http://dx.doi.org/10.1371/journal.ppat.1000522.
51. Llano M, Saenz DT, Meehan A, Wongthida P, Peretz M, Walker WH,Teo W, Poeschla EM. 2006. An essential role for LEDGF/p75 in HIVintegration. Science 314:461– 464. http://dx.doi.org/10.1126/science.1132319.
52. Poeschla E, Looney D. 1998. CXCR4 is required by a non-primate lenti-virus: heterologous expression of feline immunodeficiency virus in hu-man, rodent and feline cells. J. Virol. 72:6858 – 6866.
53. Poeschla E, Wong-Staal F, Looney D. 1998. Efficient transduction ofnondividing cells by feline immunodeficiency virus lentiviral vectors. Nat.Med. 4:354 –357. http://dx.doi.org/10.1038/nm0398-354.
54. Talbott RL, Sparger EE, Lovelace KM, Fitch WM, Pedersen NC, LuciwPA, Elder JH. 1989. Nucleotide sequence and genomic organization offeline immunodeficiency virus. Proc. Natl. Acad. Sci. U. S. A. 86:5743–5747. http://dx.doi.org/10.1073/pnas.86.15.5743.
55. Fadel HJ, Saenz D, Poeschla E. 2012. Construction and testing of orfA�/� FIV reporter viruses. Viruses 4:184 –198. http://dx.doi.org/10.3390/v4010184.
56. Saenz DT, Barraza R, Loewen N, Teo W, Poeschla E. 2006. Productionand use of feline immunodeficiency virus (FIV)-based lentiviral vectors, p57–74. In Rossi J, Friedman T (ed), Gene transfer: a Cold Spring Harbor
Morrison et al.
3270 jvi.asm.org Journal of Virology
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

laboratory manual. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
57. Saenz DT, Teo W, Olsen JC, Poeschla E. 2005. Restriction of felineimmunodeficiency virus by Ref1, LV1 and primate TRIM5a proteins. J.Virol. 79:15175–15188. http://dx.doi.org/10.1128/JVI.79.24.15175-15188.2005.
58. Sauter D, Hotter D, Engelhart S, Giehler F, Kieser A, Kubisch C,Kirchhoff F. 2013. A rare missense variant abrogates the signaling activityof tetherin/BST-2 without affecting its effect on virus release. Retrovirol-ogy 10:85. http://dx.doi.org/10.1186/1742-4690-10-85.
59. Loewen N, Barraza R, Whitwam T, Saenz DT, Kemler I, Poeschla E.2003. FIV vectors. Methods Mol. Biol. 229:251–271. http://dx.doi.org/10.1385/1-59259-393-3:251.
60. Lopez LA, Yang SJ, Hauser H, Exline CM, Haworth KG, Oldenburg J,Cannon PM. 2010. Ebola virus glycoprotein counteracts BST-2/tetherinrestriction in a sequence-independent manner that does not require teth-erin surface removal. J. Virol. 84:7243–7255. http://dx.doi.org/10.1128/JVI.02636-09.
61. Saenz DT, Barraza R, Loewen N, Teo W, Poeschla EM. 2012. Production,harvest, and concentration of feline immunodeficiency virus-based lentiviralvector from cells grown in CF10 or CF2 devices. Cold Spring Harbor Protoc.2012:118–123. http://dx.doi.org/10.1101/pdb.prot067546.
62. Nguyen KL, Llano M, Akari H, Miyagi E, Poeschla EM, Strebel K, BourS. 2004. Codon optimization of the HIV-1 vpu and vif genes stabilizestheir mRNA and allows for highly efficient Rev-independent expression.Virology 319:163–175. http://dx.doi.org/10.1016/j.virol.2003.11.021.
63. Yamamoto JK, Barre-Sinoussi F, Bolton V, Pedersen NC, Gardner MB.1986. Human alpha- and beta-interferon but not gamma- suppress the invitro replication of LAV, HTLV-III, and ARV-2. J. Interferon Res. 6:143–152. http://dx.doi.org/10.1089/jir.1986.6.143.
64. Koning FA, Newman EN, Kim EY, Kunstman KJ, Wolinsky SM, MalimMH. 2009. Defining APOBEC3 expression patterns in human tissues andhematopoietic cell subsets. J. Virol. 83:9474 –9485. http://dx.doi.org/10.1128/JVI.01089-09.
65. Benton MJ, Donoghue PC. 2007. Paleontological evidence to date thetree of life. Mol. Biol. Evol. 24:26 –53. http://dx.doi.org/10.1093/molbev/msm017.
66. Johnson WE, Eizirik E, Pecon-Slattery J, Murphy WJ, Antunes A,Teeling E, O’Brien SJ. 2006. The late Miocene radiation of modern Feli-dae: a genetic assessment. Science 311:73–77. http://dx.doi.org/10.1126/science.1122277.
67. Au WC, Moore PA, LaFleur DW, Tombal B, Pitha PM. 1998. Charac-terization of the interferon regulatory factor-7 and its potential role in thetranscription activation of interferon A genes. J. Biol. Chem. 273:29210 –29217. http://dx.doi.org/10.1074/jbc.273.44.29210.
68. Bego MG, Mercier J, Cohen EA. 2012. Virus-activated interferon regu-latory factor 7 upregulates expression of the interferon-regulated BST2gene independently of interferon signaling. J. Virol. 86:3513–3527. http://dx.doi.org/10.1128/JVI.06971-11.
69. Matsuda A, Suzuki Y, Honda G, Muramatsu S, Matsuzaki O, Nagano Y,Doi T, Shimotohno K, Harada T, Nishida E, Hayashi H, Sugano S.2003. Large-scale identification and characterization of human genes thatactivate NF-kappaB and MAPK signaling pathways. Oncogene 22:3307-3318. http://dx.doi.org/10.1038/sj.onc.1206406.
70. Cocka LJ, Bates P. 2012. Identification of alternatively translated tetherinisoforms with differing antiviral and signaling activities. PLoS Pathog.8:e1002931. http://dx.doi.org/10.1371/journal.ppat.1002931.
71. Galao RP, Le Tortorec A, Pickering S, Kueck T, Neil SJ. 2012. Innatesensing of HIV-1 assembly by tetherin induces NFkappaB-dependent pro-inflammatory responses. Cell Host Microbe 12:633– 644. http://dx.doi.org/10.1016/j.chom.2012.10.007.
72. Tokarev A, Suarez M, Kwan W, Fitzpatrick K, Singh R, Guatelli J. 2013.Stimulation of NF-kappaB activity by the HIV restriction factor BST2. J.Virol. 87:2046 –2057. http://dx.doi.org/10.1128/JVI.02272-12.
73. Saenz DT, Barraza R, Loewen N, Teo W, Poeschla EM. 2012. Felineimmunodeficiency virus-based lentiviral vectors. Cold Spring HarborProtoc. 2012(1):71–76. http://dx.doi.org/10.1101/pdb.ip067579.
74. de Rozieres S, Mathiason CK, Rolston MR, Chatterji U, Hoover EA,Elder JH. 2004. Characterization of a highly pathogenic molecular cloneof feline immunodeficiency virus clade C. J. Virol. 78:8971– 8982. http://dx.doi.org/10.1128/JVI.78.17.8971-8982.2004.
75. de Rozieres S, Thompson J, Sundstrom M, Gruber J, Stump DS, deParseval AP, VandeWoude S, Elder JH. 2008. Replication properties of
clade A/C chimeric feline immunodeficiency viruses and evaluation ofinfection kinetics in the domestic cat. J. Virol. 82:7953–7963. http://dx.doi.org/10.1128/JVI.00337-08.
76. Willett BJ, Picard L, Hosie MJ, Turner JD, Adema K, Clapham PR.1997. Shared usage of the chemokine receptor CXCR4 by the feline andhuman immunodeficiency viruses. J. Virol. 71:6407– 6415.
77. Chatterji U, de Parseval A, Elder JH. 2002. Feline immunodeficiencyvirus OrfA is distinct from other lentivirus transactivators. J. Virol. 76:9624 –9634. http://dx.doi.org/10.1128/JVI.76.19.9624-9634.2002.
78. de Parseval A, Elder JH. 1999. Demonstration that orf2 encodes the felineimmunodeficiency virus transactivating (Tat) protein and characteriza-tion of a unique gene product with partial rev activity. J. Virol. 73:608 –617.
79. Gemeniano MC, Sawai ET, Leutenegger CM, Sparger EE. 2003. Felineimmunodeficiency virus ORF-Ais required for virus particle formationand virus infectivity. J. Virol. 77:8819 – 8830. http://dx.doi.org/10.1128/JVI.77.16.8819-8830.2003.
80. Gemeniano MC, Sawai ET, Sparger EE. 2004. Feline immunodeficiencyvirus Orf-A localizes to the nucleus and induces cell cycle arrest. Virology325:167–174. http://dx.doi.org/10.1016/j.virol.2004.05.007.
81. Hong Y, Fink E, Hu QY, Kiosses WB, Elder JH. 2010. OrfA downregu-lates feline immunodeficiency virus primary receptor CD134 on the hostcell surface and is important in viral infection. J. Virol. 84:7225–7232.http://dx.doi.org/10.1128/JVI.00434-10.
82. Sparger EE, Shacklett BL, Renshaw-Gegg L, Barry PA, Pedersen NC,Elder JH, Luciw PA. 1992. Regulation of gene expression directed by thelong terminal repeat of the feline immunodeficiency virus. Virology 187:165–177. http://dx.doi.org/10.1016/0042-6822(92)90305-9.
83. Sundstrom M, Chatterji U, Schaffer L, de Rozieres S, Elder JH. 2008.Feline immunodeficiency virus OrfA alters gene expression of splicingfactors and proteasome-ubiquitination proteins. Virology 371:394 – 404.http://dx.doi.org/10.1016/j.virol.2007.09.039.
84. Kemler I, Barraza R, Poeschla EM. 2002. Mapping of the encapsidationdeterminants of feline immunodeficiency virus. J. Virol. 76:11889 –11903.http://dx.doi.org/10.1128/JVI.76.23.11889-11903.2002.
85. Tokarev A, Skasko M, Fitzpatrick K, Guatelli J. 2009. Antiviral activity of theinterferon-induced cellular protein BST-2/tetherin. AIDS Res. Human Ret-roviruses 25:1197–1210. http://dx.doi.org/10.1089/aid.2009.0253.
86. Dube M, Roy BB, Guiot-Guillain P, Binette J, Mercier J, Chiasson A,Cohen EA. 2010. Antagonism of tetherin restriction of HIV-1 release byVpu involves binding and sequestration of the restriction factor in a peri-nuclear compartment. PLoS Pathog. 6:e1000856. http://dx.doi.org/10.1371/journal.ppat.1000856.
87. Goffinet C, Homann S, Ambiel I, Tibroni N, Rupp D, Keppler OT,Fackler OT. 2010. Antagonism of CD317 restriction of human immuno-deficiency virus type 1 (HIV-1) particle release and depletion of CD317 areseparable activities of HIV-1 Vpu. J. Virol. 84:4089 – 4094. http://dx.doi.org/10.1128/JVI.01549-09.
88. Mitchell RS, Katsura C, Skasko MA, Fitzpatrick K, Lau D, Ruiz A,Stephens EB, Margottin-Goguet F, Benarous R, Guatelli JC. 2009. Vpuantagonizes BST-2-mediated restriction of HIV-1 release via beta-TrCPand endo-lysosomal trafficking. PLoS Pathog. 5:e1000450. http://dx.doi.org/10.1371/journal.ppat.1000450.
89. Checkley MA, Luttge BG, Freed EO. 2011. HIV-1 envelope glycoproteinbiosynthesis, trafficking, and incorporation. J. Mol. Biol. 410:582– 608.http://dx.doi.org/10.1016/j.jmb.2011.04.042.
90. Hosie MJ, Klein D, Binley JM, Dunsford TH, Jarrett O, Neil JC, KnappE, Giannecchini S, Matteucci D, Bendinelli M, Hoxie JA, Willett BJ.2005. Vaccination with an inactivated virulent feline immunodeficiencyvirus engineered to express high levels of Env. J. Virol. 79:1954 –1957.http://dx.doi.org/10.1128/JVI.79.3.1954-1957.2005.
91. Kuhl A, Banning C, Marzi A, Votteler J, Steffen I, Bertram S, GlowackaI, Konrad A, Sturzl M, Guo JT, Schubert U, Feldmann H, Behrens G,Schindler M, Pohlmann S. 2011. The Ebola virus glycoprotein and HIV-1Vpu employ different strategies to counteract the antiviral factor tetherin.J. Infect. Dis. 204(Suppl 3):S850 –S860. http://dx.doi.org/10.1093/infdis/jir378.
92. Fadel H, Poeschla E. 2011. Retroviral restriction and dependency factorsin primates and carnivores. Vet. Immunol. Immunopathol. 143:179 –189.http://dx.doi.org/10.1016/j.vetimm.2011.06.002.
93. Medina MF, Kobinger GP, Rux J, Gasmi M, Looney DJ, Bates P, WilsonJM. 2003. Lentiviral vectors pseudotyped with minimal filovirus enve-
Mechanism of Tetherin Antagonism
March 2014 Volume 88 Number 6 jvi.asm.org 3271
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

lopes increased gene transfer in murine lung. Mol. Ther. 8:777–789. http://dx.doi.org/10.1016/j.ymthe.2003.07.003.
94. Sinn PL, Hickey MA, Staber PD, Dylla DE, Jeffers SA, Davidson BL,Sanders DA, McCray PB, Jr. 2003. Lentivirus vectors pseudotyped withfiloviral envelope glycoproteins transduce airway epithelia from the apicalsurface independently of folate receptor alpha. J. Virol. 77:5902–5910.http://dx.doi.org/10.1128/JVI.77.10.5902-5910.2003.
95. Masuyama N, Kuronita T, Tanaka R, Muto T, Hirota Y, Takigawa A,
Fujita H, Aso Y, Amano J, Tanaka Y. 2009. HM1.24 is internalized fromlipid rafts by clathrin-mediated endocytosis through interaction with al-pha-adaptin. J. Biol. Chem. 284:15927–15941. http://dx.doi.org/10.1074/jbc.M109.005124.
96. Yang H, Wang J, Jia X, McNatt MW, Zang T, Pan B, Meng W, WangHW, Bieniasz PD, Xiong Y. 2010. Structural insight into the mechanismsof enveloped virus tethering by tetherin. Proc. Natl. Acad. Sci. U. S. A.107:18428 –18432. http://dx.doi.org/10.1073/pnas.1011485107.
Morrison et al.
3272 jvi.asm.org Journal of Virology
on July 13, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from


















