
A futile metabolic cycle activated in adipocytes by antidiabetic agents
Upload
evangeline-shepardCategory
view
30download
0description

• Fat from diet (chylomicrons) is immediately stored (adipocytes)
• excess of replenishing glycogen are made into FA (liver, adypocytes) and sent to tissues for use and to adipocytes for storage.
• Most of FA synthesis in animals occurs in adipocytes and liver.

• The products of FA biosynthesis (stage I anabolic processes).– can be exported into blood (where they
travel bound to albumin) to serve as fuel for other tissues (through beta-oxidation: stage II catabolic) or
– they are esterified • with glycerol to result in TAG for storage; or • with phosphatidic acid to make
phospholipids (membrane constituents).

• When circulating glucose levels are elevated, as after a meal rich in carbs, insulin secretion from the pancreas is stimulated.
• Insulin activates hormone sensitive glucose transporter (GLUT4) by stimulating its translocation from cytosol to plasma membrane.
• Glucose enters the cells (liver and adipocytes) and following glycolysis plenty AcetylCoA is made which will be STORED as fatty acids through the FA biosynthetic pathway.
• Note that some tissues (heart, resting muscle, liver) prefer fats as a source of energy. Recall that fats generate more energy than carbs of same carbon content.

• Ac-CoA is a common metabolite in CARB and FA pathways.
• Ac-CoA is an intermediate in FA biosynthesis and degradation BUT it can only be a product of carbohydrate metabolism.
• Ac-CoA cannot be used for de novo glucose synthesis in animals because the reaction catalyzed by Pyr-DH complex is irreversible and catabolism through TCA cycle fully oxidizes both acetyl carbons to CO2.

1. Degradation of fatty acids takes place in mitochondria Synthesis of fatty acids occurs in the cytosol
2. Intermediates in fatty acid breakdown are bound to the -SH group of coenzyme A
Intermediates in fatty acid synthesis are linked covalently to the sulfhydryl groups of acyl carrier proteins
3. Degradative enzymes are not associated as a complex Enzymes of fatty acid synthesis (in animals) are components of
one long polypeptide chain, the fatty acid synthase (plants and bacteria have separate enzymes that form the fatty acid synthase)
4. In fatty acid degradation the coenzyme for the oxidation-reduction reactions involves NAD+/NADH
In fatty acid synthesis the coenzyme for the oxidation-reduction reactions is NADP+/NADPH
Fatty acid biosynthesis is not a simple reversal of fatty acid degradation

Transport of 2C units (acetyl CoA) out of the mitochondria
Production of e- (NADPH) for FA synthesis

NADPH Sources for FA Synthesis

8
Transport of 2C units and Reducing Transport of 2C units and Reducing EquivalentsEquivalents
• OAA cannot pass inner mitochondrial membrane either malate or pyruvate are exchanged

same process different diagram…same process different diagram…
MRC
AcetylCoA is made in the mitochondrion; FA biosynthesis occurs in cytosol; and inner mitochondrial membrane is impermeable to AcetylCoA or OAA.Net transport of acetyl-units (C2) can result by transporting citrate (C6) into the cytosol and returning C4 units as malate (C4) or as pyruvate (C3) and CO2. In the process reducing equivalents are also transported.
MITOCHONDRION CYTOSOL

10
Transport of Acetyl units Transport of Acetyl units
Enzymes (locations) and reactions involved: Citrate synthase( mit) Acetyl CoA + OAA citrate + CoASH
Citrate lyase (cyt): citrate + ATP + CoASH acetylCoA +ADP +Pi + OAA
Malate DH (cyt) OAA + NADH + H+ malate + NAD+
Malic enzyme (cyt) malate + NADP+ pyruvate + CO2 + NADPH + H+
Pyr carboxylase (mit) pyruvate + CO2 + ATP + H2O OAA + ADP + H+
--------------------------------------------------------------------
ATP is expended.
In addition to transporting one acetyl unit from mit to cyt, the process results in:
NAD+ NADH and NADH NAD+ (malate DH)
(malate DH) NADP+ NADPH (malic E)
(mitochondria) (cytosol)
Inner mitochondrial membrane transporters used: • Citrate: tricarboxylic acid transporter (citrate out)• Malate: dicarboxylic acid transporter (malate in)• Pyruvate: pyruvate transporter (pyruvate in)
GP_Fall2007

Fatty Acid Metabolism
Degradation Biosynthesis
Compartment Mitochondria, Peroxisomes
Cytosol, Chloroplasts
Oxidants/ reductants
NAD+, FAD NADPH
Carrier coenzyme A acyl carrier protein(ACP)
Intermediates acetyl CoA malonyl CoA
Enzymes acetyl CoA synthase, carnitine acyltransferase, acyl CoA dehydrogenase, enoyl CoA hydratase, L-3-hydroxyacyl CoA DH ß-ketothiolase
acetyl CoA carboxylase, acetyl transacylase, malonyl transacylase, acyl-malonyl ACP cond. enz., ß-ketoacyl ACP reductase, 3-hydroxyacyl ACP dehydratase enoyl ACP reductase

StrategyStrategyActivation: specific carrier• To store energy from carbs as FAT, the common intermediate,
AcetylCoA, is directed into the Fatty Acid Synthesis pathway by exchanging the acyl-carrier CoASH with the acyl-carrier ACP at the expense of one ATP.This represents the ‘activation’ step in Fatty Acid Synthesis pathway
Fatty Acid Synthesis• FAS pathway is a spiral stage II anabolic pathway where 2C units are
added sequentially to a growing chain of acyl-ACP to form C16 chains; same chemical strategy as beta-oxidation (in reverse):
• Condensation (SYNTHASE)• Reduction (REDUCTASE) with NADPH• Dehydration (DEHYDRATASE)• Reduction (REDUCTASE) with NADPH
Specific structural elements are introduced later:
• C16 Fatty Acids are elongated as necessary (to C18; C20; etc)
• Double bonds are introduced into saturated Fatty Acids as necessary.

– 1. Condensation:CO2 is eliminated from malonyl group. Net effect is extension of the acyl chain by two carbons. The beta group is then reduced in three more steps nearly identical to the reactions of beta oxidation, but in the reverse sequence.
– 2. The beta-keto group is reduced to an alcohol.
– 3. The elimination of H2O creates a double bond, and
– 4. The double bond is reduced to form the corresponding saturated f.acyl group.
• The f.a chain grows by two-carbon units donated by activated malonate, with loss of CO2.
• After each two-carbon addition, reductions convert the growing chain to a saturated fatty acid of four, then 6,8, and so on. The final product is palmitate (16:0)

FA Biosynthesis: First Committed Step
• Acetyl-CoA + HCO3— + ATP
Malonyl-CoA + ADP + Pi– Principal point of regulation for FA synthesis.– Catalyzed by acetyl CoA carboxylase.

Carboxylation of acetyl CoA to form malonyl CoA by Acetyl-CoA carboxylase
• Acetyl-CoA carboxylase has 3 functional regions:• 1. Biotin carrier protein• 2. Biotin carboxylase, which activates CO2 by attaching it to a
nitrogen in the biotin ring in an ATP-dependent reaction.• 3. Transcarboxylase, which transfers activated CO2 from
biotin to acetyl-CoA producing malonyl CoA. The long-flexible biotin arm makes this.

Reaction #1: synthesis of malonyl-CoA by acetyl CoA carboxylase
Acetyl CoA carboxylase has three functional regions
1. Biotin carrier protein
2. Biotin carboxylase which activates CO2 by attaching it to a nitrogen in the biotin ring in an ATP dependent reaction
3. Transcarboxylasewhich transfers activated CO2 from biotin to acetyl-CoA producing malonyl CoA
21:15


Fatty Acid Synthesis Overview
• The two carbon atoms at the methyl terminus in each FA molecule derive from acetyl-CoA.
• Malonyl-CoA supplies the remaining carbons. • No carbon atoms in FA derive from bicarbonate.• FA are assembled two carbon atoms at a time
by the fatty acid synthase complex.

• The stage is set for lipogenesis by two reactions: –(1) transfer of an acetyl group from acetyl-
SCoA to an acyl carrier protein (ACP)–(2) conversion of acetyl- SCoA to malonyl-
SCoA in a reaction that requires investment of energy from ATP. The malonyl-SCoA is then transferred to the acyl carrier protein (ACP).

Fatty acid synthesis requires ACP
• acetyl carrier protein
(ACP)– Small protein 77 kda – Contain the prosthetic group
(4’-phosphopantetheine)

21
ACP is the Carrier of Acyl ACP is the Carrier of Acyl Groups for FA BiosynthesisGroups for FA Biosynthesis
The use of separate carriers:• prevents futile cycles; • prevents recognition of intermediates by enzymes used for the opposite pathway; • allows for independent flux regulation.
business endof carriers
will be swinging arm in FA Synthase
ACYL-CARRIER PROTEIN:

22
Entry into FA Synthesis: Entry into FA Synthesis: Acetyl-CoA Carboxylase
Activation of C2 units for FA biosynthesis
• activation step (ATP) : metabolic principle• Commits acetyl-CoA to FA synthesis; • Bicarbonate requirement• Major control point: metabolic principle • Hormonal control: insulin activation by de-phosphorylation
Acetyl-CoA Carboxylase
Both transacylases (2, 3) are activities on the FA Synthase complex

Formation of acetyl ACP and malonyl ACP
1. acetyl CoA + ACP acetyl ACP + CoA
2. malonyl CoA + ACP malonyl ACP + CoA
acetyl transacylase
malonyl transacylase

Multifunctional enzyme complex• A dimer of 260 kd subunits
• 3 domains– Domain 1 : the substrate entry and condensation unit
» Acetyl transferase, malonyl transferase, -ketoacyl synthase
– Domain 2: the reduction unit» ACP, -ketoacyl reductase, 3-Hydroxylacyl-ACP
dehydratase, enoyl reductase– Domain 3: the palmitate release unit
» Thioesterase
– These enzymes are covalently linked..

25
FA Synthase ComplexFA Synthase Complex
1 a-transacylase2 synthase3 m-transacylase4 k-reductase5 dehydrase6 e-reductase P-pantheteine swinging arm
cycle begins
GP_Fall2007

Fatty Acid SynthaseFatty Acid SynthaseFatty acid synthase is the name of the complex of six enzymatic activities that performs biosynthesis of
fatty acids in cytosol. The enzyme is composed of two multifunctional polypeptide chains, which contain the six enzymatic activities below:
(2) Malonyl-CoA-ACP Transacylase: swaps ACP with CoA in malonyl-CoA
(3) Acetyl-CoA-ACP Transacylase: swaps ACP with CoA in acetyl-CoA
(4) -Ketoacyl-ACP Synthase: C2 is added to malonyl-ACP ketoacyl-ACP
(5) -Ketoacyl-ACP Reductase: reduces D-3-keto D-3-hydroxy, stereospecific
(6) 3-Hydroxylacyl-ACP Dehydrase: dehydration trans-2-enoyl
(7) Enoyl-ACP Reductase: reduction with NADPH


28
REACTIONSREACTIONSsynthase; reductase; dehydrase; synthase; reductase; dehydrase;
reductasereductase
GP_Fall2007

FA Synthase: Cycle of 4 rxns.The 2C from Malonyl-CoA are added to the carboxy terminus of the FA.

Synthesis of 16:0 via 7 cycles by Fatty Acid Synthase.

subunit = 3 activities
ß-subunit = 4 activities
1 polypeptide = 7 activities
MW= 240,00, functions as a dimer arranged head-to-tail.
7 separate proteins
Comparison of fatty acid synthase systems
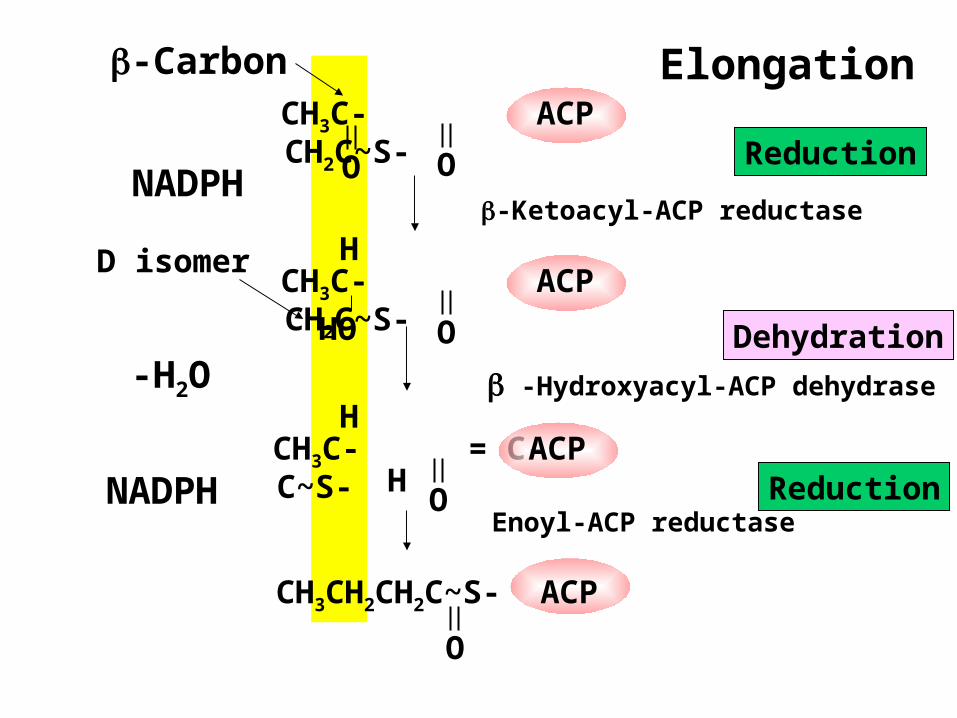
CH3C-
O
CH2C~S-
O
ACP
NADPH
CH3CH2CH2C~S-
O
ACP
CH3C- CH2C~S-
O
ACP
HO
H
CH3C- = C- C~S-
O
ACPH
H
-H2O
NADPH
-Carbon Elongation
D isomer
Reduction
Dehydration
Reduction
-Ketoacyl-ACP reductase
-Hydroxyacyl-ACP dehydrase
Enoyl-ACP reductase

Condensation and Reduction
In reactions 1 and 2 of fatty acid synthesis:
• Condensation by a synthase combines acetyl-ACP with malonyl-ACP to form acetoacetyl-ACP (4C) and CO2 (reaction 1).
• Reduction converts a ketone to an alcohol using NADPH (reaction 2).

Dehydration and Reduction
In reactions 3 and 4 of fatty acid synthesis:
• Dehydration forms a trans double bond (reaction 3).
• Reduction converts the double bond to a single bond using NADPH (Reaction 4).

Lipogenesis Cycle Repeats
Fatty acid synthesis continues:
• Malonyl-ACP combines with the four-carbon butyryl-ACP to form a six-carbon-ACP.
• The carbon chain lengthens by two carbons each cycle.

Lipogenesis Cycle Completed
• Fatty acid synthesis is completed when palmitoyl ACP reacts with water to give palmitate (C16) and free ACP.

Summary of Lipogenesis

ACP
KS -SH
HSAcetyl-CoA
CoA-SH
-C-CH3
OS
KS S-C-CH3
OKS -SH
SH
CoA-SH
Malonyl-CoA
S -C-CH2-COO-
O
CO2C=O
CH2
C=O
CH3
S
O
CH3-CH -CH2-C-S
OH
OCH3-CH=CH-C-S
OCH3-CH2-CH2-C-S
S-C-CH2-CH2-CH3
O
KS
KS
NADP+
NADPH H+
NADPH H+
NADP+
H2O
Initiation or priming
Elongation
Fatty Acid SynthaseFatty Acid Synthase
-Ketoacyl-ACP reductase
-Ketoacyl-ACP reductase
-Hydroxyacyl-ACP dehydrase
-Hydroxyacyl-ACP dehydrase
Enoyl-ACP reductase
Enoyl-ACP reductase
-Keto-ACP synthase (condensing enzyme)
-Keto-ACP synthase (condensing enzyme)
Malonyl-CoA-ACP transacylase
Malonyl-CoA-ACP transacylase
Acetyl-CoA-ACP transacylase
Acetyl-CoA-ACP transacylase
-Ketoacyl-ACP synthase
-Ketoacyl-ACP synthase

Step 4:Acetyl and Malonyl condense to form -ketoacyl-ACP bound to ACP
reaction is catylzed by-ketoacyl-ACP synthase
in this reaction the acetyl group is transferred to the malonyl group simultaneously a molecule of CO2 is produced
The decarboxylation facilitates the nucleophilic attack of the methylene carbon on the thioester linking the acetyl group to -ketoacyl-ACP synthase
Coupling the condensation to the decarboxylation makes the overall reaction highly exergonic
21:19

Step5: -ketoacyl-ACP is reduced (at the carbonyl group at C-3) to D-3-hydroxyacyl-ACP
the enzyme is -keotacyl-ACP reductasethe electron donor is NADPH
note that in fatty acid oxidation, the 3-hydroxyacyl-CoAs produced have the L configuration
Step 6: dehydration of D-3-hyroxyacly-ACP to trans-∆2-enoyl-ACP
water is removed from C-2 and C-3 to yield a double bond in the product
the enzyme is 3-hydroxyacyl-ACP dehydrase (dehyratase)
step #7: the synthesis of butyryl-ACPthe double bond generated in the last reaction is reduced the enzyme is enoly-CoA reductaseNADPH is the electron donor
21:20

One round of synthesis through the fatty acid synthase complex is complete
A 4 carbon butyryl-ACP has been generatedThe butyryl-ACP is now transferred from ACP to a Cys-SH of the -ketoacyl-ACP synthase portion of fatty acid synthase complex(just like an acetyl group was transferred in the first round)
To start a second cycle (of the last four reactions) in order to lengthen the chain by two more carbons, butyryl-ACP condenses with another molecule of malonyl-ACP
butyryl acts just like acetyl did in the first cycle, and CO2 is lossed
Now there would be six carbons total here, with the first two from malonyl CoA and the next four from butyryl
the product of this second round of reactions is hexanoly-ACP; the pattern continues till seven cycles of condensation and reduction produce the 16 carbon saturated palmitoyl group (still bound to ACP)
21:21

for reasons not well understood, chain elongation generally stops at this point and free palmitate is released from the ACP molecule by the action of a hydrolytic activity in the synthase complex
small amounts of longer fatty acids such as stearate (18:0) are also formed
in certain plants (cococut and palm) chain termination occurs earlier and up to 90% of the fatty acids in the oils of these plants are between 8 and 14 carbons long
21:22

21:23

-KS
CO2
-S-ACP
TERMINATION Ketoacyl ACPSynthase
Free to bindMalonyl-CoA
Transfer to KS
Split out CO2
Transfer to Malonyl-CoA
-CH2CH2CH2C~S-
O
ACP
When C16 stage is reached, instead of transferring to KS,the transfer is to H2O and the fatty acid is released

After 7* Cycles
Synthase-Cys SH
ACP-PPant SH
Synthase-Cys SH
ACP-PPant S C
O
(CH2)14 CH3
palmityl ACP
H2O
hydrolase
palmitate
.
.

FA Synthesis Stoichiometry
• Review the requirements for NADPH and ATP.
• Be able to identify where all carbon atoms in 16:0 come from.

Synthesis of palmitate• Part 1)
7 acetyl CoA + 7 CO2 + 7 ATP
7 malonyl CO2 + 7 AMP + 7 Pi
• Part 2)Acetyl CoA + 7 malonlyl CoA + 14 NADPH + 14 H+
palmitate + 7 CO2 + 8 CoA + 14 NADP + + 6 H2 0
• Overall process 8 Acetyl CoA + 14 NADPH + 14 H +
palmitate + 8 CoA + 6 H2 0 + 7 ADP + 7 Pi + 14 NADP +

Regulation of fatty acid biosynthesis occurs at the ACCase rxn
• ACCase (in animals)– rate limiting step for FA synthesis– dephosphorylated (polymerized,
active form)– phosphorylated
(depolymerized, inactive form)
• ACCase (plants, bacteria) – Not regulated by citrate or by
phosphorylation.– Activated in plants by elevated
[Mg2+] and lower pH.
• Reciprocal regulation of FA synthesis and -oxidation.– Elevated [Malonyl-CoA] inhibits
carnitine acyltransferase I and thus blocks -oxidation of FA at the level of transport.
Rxn2
Rxn1

Long chain fatty acid biosynthesis
• End product of FA biosynthesis in animals:– Palmitate (16:0)
• Fatty acid elongation systems: – in smooth ER, mitochondria– ER mechanism similar to general FA synthesis
• Condensing, reduction, dehydration, reduction• malonyl-CoA as 2 carbon donor• different enzymes and coenzyme A instead of ACP

50
ElongationElongation synthase, reductase; dehydrase; reductasesynthase, reductase; dehydrase; reductase
• The fatty acid synthesis pathway involving cytosolic FA synthase leads to palmitate• Synthesis of longer FA involves elongation; in EK this process occurs in mitochondria
and in ER membranes (microsomes)• Microsomal elongation is prevalent; it involves:
– acyl-CoA intermediates and – separate enzymes
• similar enzymatic complex as cytosolic FA Synthase; several condensing enzymes and one set of the rest three activities; can act on unsaturated FA
GP_Fall2007

51
Essential Fatty AcidsEssential Fatty Acids• Most common mono unsaturated FA in animals are :
– oleic (18:1c9) and palmitoleic (16:1c9)• Microsomes contain four Fatty Acid Desaturase enzymes which can introduce
desaturation at C4, C5, C6 or C9 (only up to C9).• Linoleic acid 18:2c9,12 or linolenic acid 18:3c9,12,15 cannot be synthesized as
they require desaturation beyond C9.• These must be provided in diet: are essential fatty acids
• Linoleic acid: is especially important in that it required for the synthesis of arachidonic acid: precursor for prostaglandins and thromboxanes (required for growth, wound healing and epidermal health)
• Evidence is accumulating for many other beneficial effects of poly-unsatturated fatty acids (PUFA) particularly those with 3 and 6 double bonds:
• linolenic acid (18:3) • stearidonic acid (18:4) • eicosatetraenoic acid (20:4) • EPA (20:5) • DHA (22:6)
• linoleic acid (18:2) • gamma-linolenic acid (18:3) • dihomo-gamma-linolenic acid (20:3) • ARA (20:4)

52
Fatty Acid DesaturationFatty Acid Desaturation• Fatty Acid Desaturase: introduces cis double bonds at specific C-positions
(oxidase: uses molecular oxygen)
• Reaction avoids generating H2O2 in the vicinity of the double bond by bringing additional e- through cyt b5 (Fe2+) and cyt b5 reductase (FAD) from NADH.
GP_Fall2007

Desaturase
Cyt b5
reductase
Cyt b5
C18-stearoly-CoA + O2 + 2H+
C18 9-oleyl-CoA + 2H2O
2 cyt b5 Fe2+ 2 cyt b5 Fe2+
2H+ + cyt b5 reductaseFAD
cyt b5 reductase FADH2
NADH + H+NAD+
Desaturase

Mammalian Fatty Acid Desaturation
CH3(CH2)16C
O
S CoA CH3(CH2)16C
O
SE
9 desaturase(acyltransferase)
CoASH
9 desaturase(hydroxylase)
O2 H2O
cyt b5
NADPHor NADH
+ H+
NADP+
or NAD+
(CH2)7C C
O OH
CS (CH2)7CH3E
H H
H
(CH2)7C C
O
CS (CH2)7CH3E
HH9 desaturase(hydratase)
H2O
E
(CH2)7C C
O
CS (CH2)7CH3E
H H 9 desaturase(acyltransferase)
CoASH
(CH2)7CC
O
C SCoA(CH2)7CH3
HH
.
.

Elongation of Chain (two systems)
HS-CoA
R-CH2CH2CH2C~SCoAO
OOC-CH2C~SCoA
OCO2
Malonyl-CoA* (cytosol)
R-CH2CH2CH2CCH2C~SCoAO O
O R-CH2CH2CH2CH2CH2C~SCoA
NADPH NADH
1
- H2O2
NADPH3
Elongation systems arefound in smooth ER andmitochondria
CH3C~SCoA
OAcetyl-CoA(mitochondria)

DesaturationRules:The fatty acid desaturation system is in the smooth membranes of the endoplasmicreticulum
There are 4 fatty acyl desaturase enzymes in mammals designated 9 , 6, 5, and 4 fattyacyl-CoA desaturase
Mammals cannot incorporate a double bondbeyond 9; plants can.
Mammals can synthesize long chain unsaturated fatty acids using desaturation and elongation

The Desaturase System requires O2 andresembles an electron transport system
Rule:
NADHCyt b5 reductase
Cyt b5O2
Saturated FA-CoA
(FAD)
NOTE:
1. System is in ER membrane
2. Both NADH and the fatty acid contribute electrons
3. Fatty acyl desaturase is considered a mixed function oxidase
2
2
3
1

Palmitate
Stearate
Oleate
Linoleate
-Linolenate-Linolenate
Eicosatrienoate
Arachidonate
18:3(9,12,15)
18:2(9,12)
18:3(6,9,12)
16:0
18:0
Elongase
18:1(9)
Palmitoleate
16:1(9)Desaturase
Desaturase
Desaturase
Desaturase
Desaturase
Desaturase
Elongase
20:3(8,11,14)
20:4(5,8,11,14)
Other lipids
Permittedtransitionsin mammalsEssential
fatty acid

59
Control of FA SynthesisControl of FA Synthesis• Largely hormonal control: stimulated by INSULIN
Glc uptake from blood: • GLUT4 is hormone sensitive glucose transporter (adipocytes, muscle); translocates from cytosol to plasma
membrane; allows Glc import by passive diffusion
• Hexokinase sequesters Glc as Glc6P and maintains low Glc intracellularly to allow more Glc import
• Glc6P provides NADPH by the PPP pathway (Glc6P dehydrogenase) and acetylCoA by glycolysis and PDC
Pyruvate Dehydrogenase Complex (de-phosphorylation, active) [acetyl-CoA] AcetylCoA Carboxylase (de-phosphorylation, active) [malonyl-CoA]
• Substrate conc increase in FA Synthesis (plenty of malonyl-CoA)
• Allosteric inhibition of carnitine-transport system (FAS)
• Substrate-level
[acetylCoA] and [malonyl-CoA]– [NADPH] may be limiting; abundant Glc results in NADPH (from the PPP) and favors FAS
• Acetyl-CoA carboxylase (first committed step):
– allosteric inhibition by long-chain Fatty Acyl-CoAs: promote monomeric form– active form is a polymeric filament; – inactive form is monomer;
GP_Fall2007

it polymerizes into a filamentous form which is the active species
the equilibrium between the monomers and the filamentous form is regulated
the final product of fatty acid biosynthesis (palmitoyl-CoA) shifts the equilibrium toward the inactive monomers
citrate an allosteric activator of the enzyme shifts the equilibrium toward the active filamentous form
the primary physiological regulator is probably long chain fatty acyl-CoAs
Acetyl CoA carboxylase (in eukaryotes) is a single protein with two identical polypeptide chains of MW 230kD each
21:16

Mammalian Fatty Acid Desaturation
CH3(CH2)16C
O
S CoA CH3(CH2)16C
O
SE
9 desaturase(acyltransferase)
CoASH
9 desaturase(hydroxylase)
O2 H2O
cyt b5
NADPHor NADH
+ H+
NADP+
or NAD+
(CH2)7C C
O OH
CS (CH2)7CH3E
H H
H
(CH2)7C C
O
CS (CH2)7CH3E
HH9 desaturase(hydratase)
H2O
E
(CH2)7C C
O
CS (CH2)7CH3E
H H 9 desaturase(acyltransferase)
CoASH
(CH2)7CC
O
C SCoA(CH2)7CH3
HH
.
.

















![Microarray analysis of adipocytes treated with protease ...€¦ · Web view] and lipid metabolism [43] in non-ARV-treated adipocytes. This finding is supported by evidence from](https://static.fdocuments.us/doc/165x107/5e27dbd605c1826b8578f3b6/microarray-analysis-of-adipocytes-treated-with-protease-web-view-and-lipid.jpg)
