
FACULTE DES SCIENCES DE LA NATURE ET DE LA VIE THESE · 2019-07-04 · Remerciements Je remercie...
159
Année universitaire 2017/2018 FACULTE DES SCIENCES DE LA NATURE ET DE LA VIE THESE EN VUE DE L’OBTENTION DU DIPLOME DE DOCTORAT EN SCIENCES Biologie Spécialité : Parasitologie Présentée par ALLAL-IKHLEF Amina Bouchra Intitulée Soutenue le 13 Décembre 2018 Devant le Jury : Président Mr. A. BENSAHLA TALET Professeur, Université Oran 1 Directeur de thèse Mme. K. SENOUCI Professeur, Université Oran 1 Examinateur Mme. S. BERCHI Professeur, Ecole Supérieure de Biotechnologie, Constantine Examinateur Mr. N. KARAM Professeur, Université Oran 1 Examinateur Mr. A. BOUDJEMAA Professeur, Université Mohamed Boudiaf, Oran Examinateur Dr. Z. HARRAT Directeur de Recherche, Institut Pasteur d’Algérie Invité Mr. R. TRIKI Professeur, Université de Blida 1 Contribution à l’étude du rôle vecteur des phlébotomes dans un foyer à leishmanioses à Draa El Mizan (Kabylie, Algérie)
Transcript of FACULTE DES SCIENCES DE LA NATURE ET DE LA VIE THESE · 2019-07-04 · Remerciements Je remercie...

Année universitaire 2017/2018
FACULTE DES SCIENCES DE LA NATURE ET DE LA VIE
THESE
EN VUE DE L’OBTENTION DU DIPLOME DE
DOCTORAT EN SCIENCES
Biologie
Spécialité : Parasitologie
Présentée par
ALLAL-IKHLEF Amina Bouchra
Intitulée
Soutenue le 13 Décembre 2018
Devant le Jury :
Président Mr. A. BENSAHLA TALET Professeur, Université Oran 1
Directeur de thèse Mme. K. SENOUCI Professeur, Université Oran 1
Examinateur Mme. S. BERCHI Professeur, Ecole Supérieure de
Biotechnologie, Constantine
Examinateur Mr. N. KARAM Professeur, Université Oran 1
Examinateur Mr. A. BOUDJEMAA Professeur, Université Mohamed
Boudiaf, Oran
Examinateur Dr. Z. HARRAT Directeur de Recherche, Institut
Pasteur d’Algérie
Invité Mr. R. TRIKI Professeur, Université de Blida 1
Contribution à l’étude du rôle vecteur des phlébotomes dans un foyer à
leishmanioses à Draa El Mizan (Kabylie, Algérie)

Dédicace
Ce travail est dédié à mon père Mohammed Fethi, décédé trop tôt, qui m'avait
toujours encouragée et motivée dans mes études.
J'espère que, du monde qui est sien maintenant, il apprécie cet humble geste
comme preuve de reconnaissance de la part sa fille qui l’aime et prie pour le salut
de son âme.
Puisse Dieu, le tout puissant, lui accorder Sa sainte Miséricorde.

Remerciements
Je remercie Dieu le tout puissant qui m’a donné la santé, la force et la volonté
pour mener à terme ce travail de thèse de Doctorat
Je remercie Monsieur le Professeur Ahmed BENSAHLA-TALET, Doyen de la
Faculté des Sciences de la Nature et de la vie pour l’honneur qu’il me fait en
acceptant de présider le jury de cette thèse malgré ses nombreuses obligations.
Qu’il trouve ici l’expression de mon profond respect et de ma reconnaissance.
Je tiens à remercier vivement ma Directrice de thèse,Madame le Professeur
Kheira SENOUCI, pour la confiance qu’elle m’a témoignée en acceptant de
diriger ce travail qui n’aurait pas abouti sans sa présence et son soutien. Je lui
suis reconnaissante de m’avoir fait bénéficier de sa compétence, de sa rigueur
intellectuelle, de son dynamisme, de sa générosité et de son efficacité certaine
que je n’oublierai jamais. J'aimerais également lui dire à quel point j’ai apprécié
sa grande disponibilité, ses recommandations et les nombreuses discussions
scientifiques que nous avons eues. J’ai été extrêmement sensible à ses qualités
humaines d'écoute et de compréhension, merci de votre patience lors de la
rédaction de cette thèse. Soyez assurée de mon attachement et de ma profonde
gratitude.
J’exprime mes sincères remerciements avec toute ma gratitude à Madame
Selima BERCHI, Professeur à l’Ecole Supérieure Biotechnologie de Constantine,
Monsieur Nourredine KARAM, Professeur à l’Université Oran 1 et Monsieur
Abdellah BOUDJEMAA, Professeur à l’Université Mohamed Boudiaf USTO, pour
l’honneur qu’ils m’ont fait en acceptant de juger ce travail.
Je tiens aussi à remercier vivement et particulièrement Docteur Zoubir
HARRAT, Directeur Général de l’Institut Pasteur d’Algérie et Chef de
Laboratoire d’éco-épidémiologie parasitaire et génétique de populations du
Centre National de référence de leishmania de l’Institut Pasteur d’Algérie qui
m’honore en acceptant d’examiner ce travail malgré ses grandes obligations, je le
remercie pour ses encouragements, ses conseils et sa disponibilité. Grand merci
à son équipe Kamel, Rafik, Ihcen, Sihem, Chahra,…..de m’avoir non seulement
initiée à l’identification microscopique des phlébotomes mais aussi pourles
discussions et conseils qui m’ont accompagnée et soutenue dans tout ce que j’ai
entrepris.

Je n’oublie pas de témoigner également ma sincère reconnaissance à Monsieur
Rachid TRIKI, Professeur à l’Université de Blida pour l’intérêt qu’il a manifesté
en participant en qualité de membre invité à ce jury.
Un grand MERCI adressé à Monsieur le Professeur Djamel SAIDI, Directeur de
l’Ecole des Sciences de la Nature et de la Vie d’Oran, pour ses précieux conseils,
ses encouragements et sa générosité.
Mes remerciements vont à toute l’équipe du service Parasitologie-Mycologie de
l’Hôpital Avicenne de Paris pour l’aide précieuse dont j’ai bénéficiée.
Je tiens aussi à remercier Monsieur le Professeur Rémi CHARRELde l’Université
d’Aix Marseille et son équipe du Laboratoire des Virus émergeants UMR_D
190de la Faculté de médecine, La Timone - Marseille (France) de m’avoir
accueillie au sein du Laboratoire et m’ont permis de finaliser ce travail de thèse.
Mon travail de recherche sur le terrain a été particulièrement facilité par
l’équipe d’hygiène de Draa El Mizan (Tizi-Ouzou) et les autorités locales. Je tiens
à leur exprimer ma reconnaissance pourl'intérêt dont elles ont fait preuve
envers ma recherche, les moyens qu'ellesont mis en œuvre pour medonner accès
au terrain.
Un grand MERCI au staff collègue de Parasitologie de l’Université Oran 1,
Madame Dounia Marzoug, Madame Lahouaria Bourguig et Touria Hadj Slimane
pour la bonne ambiance et leur soutien moral. Un MERCI particulier à Madame
Meryiem Berrazeg pour son aide sans faille et sa présence au bon moment, elle a
su me remonter le moral à chaque que j’en ai eu besoin.
Une petite pensée et un grand MERCI à l’équipe de l’association écologique
BARBAROUSS. Mon adhésion à cette association est venue au bon moment. Je
lui souhaite une longue vie pleine de succès.
Enfin, ma reconnaissance va à ceux qui ont plus particulièrement assuré le
soutien affectif à ce travail, ma famille et mes amis.
Mille MERCI à ma chère maman « MAMI » qui est toujours présente avec le
souci de nous réunir et de s’assurer de notre bonheur.
Un grand MERCI à mon frère ainé Aziz qui a su assurer son rôle de frère à
chaque fois qu’on a besoin de lui et son épouse Hasna. A mes sœurs Chafika

« KIKA » et Souad « SOUSSOU » pour leur générosité et leur bonne humeur. A
mon beau frère Ghouti « Titi » pour sa joie et son savoir vivre.
Un MERCI particulier à mon frère Hakim et sa charmante épouse Yasmine qui
m’ont bien accueillie lors de mes déplacements à Paris et qui m’ont toujours
encouragée dans mon travail.
A toutes mes nièces : Sana, Khadija, Soraya, Lyna et Sara et mes neveux :
Mohammed, Adel, Mustapha et Wahib à qui je souhaite beaucoup de succès et
de bonheur.
Merci à mon oncle Chawky Ghaouti qui me fait l’honneur de venir de loin pour
assister à ma soutenance. Grand merci aussi à son adorable petite famille. Ils ont
toujours été présents pour nous et nous ont soutenus dans les moments les plus
difficiles.
Merci aussi à ma belle famille qui m’encourage et me soutient.
Je ne peux pas finir d’écrire cette page, sans adresser mes remerciements
incommensurables à mon cher époux MOUSSA et mes chers joyaux Sheima et
Yousra. Sans eux la vie n’aurait pas de goût et je n’aurais pas pu aller à l’avant vu
toutes les difficultés que j’ai rencontrées durant mon parcours….Je ne m’en suis
sortie que plus forte, grâce à eux.
Longue vie à toute ma famille
Un grand MERCI à tous ceux qui ont partagé avec moi leurs connaissances, leur
diplomatie et leurs passions.

Table des matières
Résumés
Liste des tableaux
Liste des figures
INTRODUCTION
Introduction………………………………………………………………………......... 1
CHAPITRE I SYNTHESE BIBLIOGRAPHIQUE
1. Les phlébotomes……………………………………………………………………. 4
1.1. Taxonomie………………………………………………………………………... 4
1.1.1. Historique de la taxonomie……………………………………………………... 4
1.1.2. Position systématique des phlébotomes………………………………………… 6
1.2. Morphologie…………………………………………………………………….... 6
1.3. Biologie et éco-éthologie…………………………………………………………. 10
1.4. Répartition géographique…………………………………………………………. 13
1.4.1. Dans le monde………………………………………………………………….. 13
1.4.2. En Algérie………………………………………………………………………. 13
1.4.2.1. Espèces recensées…………………………………………………………….. 13
1.4.2.2. Répartition des espèces selon les étages bioclimatiques……………………... 15
1.5. Epidémiologie…………………………………………………………………….. 16
1.5.1. Facteurs bioclimatiques………………………………………………………… 16
1.5.2. Facteurs anthropiques…………………………………………………………... 16
2. Rôle vecteur des phlébotomes……………………………………………………… 17
2.1. Leishmanioses…………………………………………………………………….. 17
2.1.1. Historique………………………………………………………………………. 17
2.1.2. Généralités……………………………………………………………………… 18
2.1.3. Leishmanioses en Algérie………………………………………………………. 22
2.1.3.1. Leishmaniose viscérale……………………………………………………….. 22
2.1.3.2. Leishmaniose cutanée sporadique du Nord…………………………………... 23
2.1.3.3. Leishmaniose cutanée zoonotique……………………………………………. 23
2.1.4. Circulation du parasite………………………………………………………….. 24
2.1.5. Techniques diagnostiques………………………………………………………. 25
2.1.6. Le parasite………………………………………………………………………. 26
2.1.6.1. Taxonomie……………………………………………………………………. 26
2.1.6.2. Morphologie………………………………………………………………….. 27
2.1.6.3. Cycle biologique……………………………………………………………… 27
2.1.7. Leishmanies et réactions immunitaires…………………………………………. 29
2.2. Arbovirus…………………………………………………………………………. 31
2.2.1. Généralités……………………………………………………………………… 31
2.2.2. Bunyaviridae……………………………………………………………………. 33
2.2.2.1. Généralités……………………………………………………………………. 33
2.2.2.2. Structure………………………………………………………………………. 34

2.2.2.3. Dissémination virale et réponse immunitaire………………………………… 35
2.2.3. Phlébovirus……………………………………………………………………... 36
2.2.3.1. Généralités……………………………………………………………………. 36
2.2.3.2 Découverte des phlébovirus en Algérie……………………………………….. 40
2.2.4. Virus Toscana (Toscana virus ou TOSV)………………………………………. 42
2.2.4.1. Introduction…………………………………………………………………… 42
2.2.4.2. Structure et propriétés biologiques…………………………………………… 43
2.2.4.3. Découverte et pathogénicité de TOSV……………………………………….. 44
2.2.4.4. Syndromes cliniques………………………………………………………….. 44
2.2.4.5. Diagnostic…………………………………………………………………….. 45
CHAPITRE II MATERIEL ET METHODES
1. Caractéristiques de la zone d’étude………………………………………………… 46
2. Etude entomologique……………………………………………………………….. 47
2.1. Méthodes d’échantillonnage des phlébotomes adultes…………………………… 47
2.2. Identification des phlébotomes…………………………………………………… 50
2.2.1. Identification à frais…………………………………………………………….. 50
2.2.2. Identification après éclaircissement et montage………………………………... 51
3. Etude parasitologique………………………………………………………………. 51
3.1. Recherche des formes promastigotes chez le phlébotome……………………….. 51
3.2. Détection de cas humains de leishmaniose viscérale par Western blot…………... 52
4. Recherche et caractérisation moléculaire de phlébovirus…………………………... 53
4.1. Broyage et homogénéisation des pools de phlébotomes…………………………. 53
4.2. Extraction de l’ARN……………………………………………………………… 53
4.2.1. Extraction par le robot (Well Bloch, Eppendorf ep motion 5075)……………... 54
4.2.2. Extraction manuelle à partir du Kit Quiagen EZ1®. Virus MiniKit V2.0……... 54
4.3. Amplification génique……………………………………………………………. 54
4.3.1. Amorces………………………………………………………………………… 55
4.3.2. Préparation des mix…………………………………………………………….. 57
4.3.3. Programmation du thermocycleur……………………………………………… 58
4.4. Lecture des résultats de PCR par électrophorèse sur gel d’agarose……………… 59
4.5. Purification des produits de PCR………………………………………………… 60
4.6. Séquençage……………………………………………………………………….. 60
4.6.1. Préparation de la réaction du séquençage………………………………………. 61
4.6.2. Purification au Sephadex®……………………………………………………... 61
5. Lecture des résultats………………………………………………………………... 62
6. Mise en culture des phlébovirus……………………………………………………. 62
6.1.Préparation des cellules…………………………………………………………… 62
6.2.Trypsinisation des cellules………………………………………………………… 62
6.3. Inoculation du virus………………………………………………………………. 63
7. Séquençage du génome complet……………………………………………………. 63
7.1. Distances génétiques et analyse phylogénétique…………………………………. 64
8. Séroneutralisation…………………………………………………………………... 64

CHAPITRE III RESULTATS
1. Etude entomologique……………………………………………………………….. 67
1.1. Résultats des piégeages…………………………………………………………… 67
1.2. Inventaire taxonomique…………………………………………………………... 67
1.3. Aspect écologique………………………………………………………………… 75
1.3.1. Abondance relative selon le taxon des phlébotomes piégés sur papiers
Huileux………………………………………………………………………………...
75
1.2.3. Abondance relative selon le taxon des phlébotomes capturés par pièges
lumineux…………………………………………………………………………….....
77
2. Etude parasitologique………………………………………………………………. 79
2.1. Recherche des leishmanies chez le vecteur………………………………………. 79
2.2. Diagnostic sérologique de cas humains de leishmaniose viscérale………………. 80
3. Etude virologique…………………………………………………………………… 82
3.1. Isolement et caractérisation du virus Toscana par analyse moléculaire………….. 82
3.1.1. Recherche du virus……………………………………………………………... 82
3.1.2. Isolement du virus………………………………………………………………. 83
3.1.3. Séquençage du génome complet………………………………………………... 83
3.1.4. Analyse phylogénétique………………………………………………………… 84
3.2. Séroneutralisation………………………………………………………………… 86
4. Co-infections Leishmania-Toscana………………………………………………… 88
CHAPITRE IV DISCUSSION
Discussion……………………………………………………………………………... 90
CONCLUSION ET PERSPECTIVES
Conclusion et perspectives……………………………………………………………. 98
Références bibliographiques…………………………………………………………... 101
Annexes……………………………………………………………………………….. 114

Résumé
Les leishmanioses, maladies parasitaires observées chez les mammifères dont l’Homme,
sont causées par un protozoaire du genre Leishmania, transmis par un insecte piqueur, le
phlébotome femelle. Deux formes cliniques de leishmanioses sévissent en Algérie, la
leishmaniose viscérale et les leishmanioses cutanées. Récemment, des arbovirus du genre
Phlebovirus, les virus Sicilien et de Naples ayant le phlébotome comme vecteur commun
avec Leishmania, ont été signalés dans le Nord de l’Algérie. Notre travail a été consacré à
la recherche en Algérie, du virus Toscana et à la mise en évidence de l’existence de son lien
épidémiologique avec Leishmania, durant la période de 2012 à 2014. L’étude a été menée
dans la région de Draa El Mizan (Kabylie), un foyer endémique à leishmanioses. Un total
de 25781 phlébotomes a été capturé dont 4283 ont fait l’objet d’une étude taxonomique
pour déterminer la composition de la population phlébotomienne. Huit espèces du genre
Phlebotomus et Sergentomyia ont été recensées avec la prédominance de P. perniciosus. La
recherche de Leishmania chez les phlébotomes femelles s'est révélée négative. Une analyse
moléculaire, basée sur la technique de RT-PCR suivie de celle de la PCR nichée, a été
utilisée pour la détection des phlébovirus chez ces insectes. Ces techniques ont permis
d’identifier et de caractériser pour la première fois en Algérie, le virus Toscana (TOSV).
Après la culture du virus sur des cellules Vero (ATCC CCL81), le séquençage complet du
génome viral a été réalisé par la technique de séquençage de nouvelle génération (Next-
Generation- Sequencing, NGS). La souche obtenue, nommée «Toscana Virus Algérie,
souche 189», et classée phylogénétiquement dans la lignée A, est proche de la souche
TOSV isolée en Tunisie. L’étude a été complétée par deux tests sérologiques, Western Blot
et séroneutralisation, effectués sur 370 sérums provenant de personnes choisies
aléatoirement parmi la population de Draa El Mizan. Ces tests ont détecté respectivement
86 cas positifs à Leishmania et des anticorps neutralisants contre TOSV dans 171 sérums en
utilisant une dose de virus à 1000 DICT50. Ces résultats, qui indiquent que TOSV circule à
Draa El Mizan, sont en adéquation avec la présence du virus chez les phlébotomes
sévissant dans cette région. Par ailleurs, la séropositivité, aux deux tests, de 11 sérums
indique l’existence d’infections virales à TOSV concomitantes à une leishmaniose viscérale
parmi la population de Draa El Mizan.
Mots clés : Leishmania, virus Toscana, phlébotome, épidémiologie, Draa El Mizan,
Kabylie, Algérie.

Abstract:
A protozoan of the genus Leishmania, transmitted by a biting insect, the female sandfly,
causes Leishmaniasis, parasitic diseases observed in mammals including humans. Two
clinical forms of leishmaniasis occur in Algeria, visceral leishmaniasis and cutaneous
leishmaniasis. Recently, arboviruses of the genus Phlebovirus, the Sicilian and Naples viruses
with sandfly as a common vector with Leishmania, have been reported in northern Algeria.
Our work was devoted to search in Algeria, the virus Toscana and to demonstrate the
existence of its epidemiological link with Leishmania, during the period of 2012 - 2014. The
study was conducted in the region of Draa El Mizan (Kabylia), a leishmaniasis endemic
outbreak. A total of 25781 sandflies were captured, of which 4283 were the subject of a
taxonomic study to determine the composition of the phlebotomian population. Eight species
of the genus Phlebotomus and Sergentomyia were recorded with the predominance of P.
perniciosus. The search for Leishmania in female sandfly was negative. A molecular analysis,
based on the RT-PCR followed by the nested PCR, was used for the detection of phlebovirus
in these insects. These techniques have identified and characterized, the Toscana virus (TOSV)
for the first time in Algeria. After virus culture on Vero cells (ATCC CCL81), next
Generation Sequencing (Next Generation Sequencing) performed complete sequencing of the
viral genome. The strain obtained, called "Toscana Algeria Virus Strain 189" and
phylogenetically classified in line A, is close to the TOSV strain isolated in Tunisia. The
study was completed by two serological tests, Western blot and neutralization, performed on
370 serums from people randomly selected from the population of Draa El Mizan. These tests
respectively detected 86 positive cases of Leishmania and neutralizing antibodies against
TOSV in 171 serums, using a dose of 1000 TCID50 virus. These results, which indicate that
TOSV circulates in Draa El Mizan, are in line with the presence of the virus in the sandflies
found in this region. Moreover, HIV status for both tests of 11 serums indicates the presence
of viral infections TOSV concomitant with visceral leishmaniasis among the population of
Draa El Mizan.
Key words: Leishmania, Toscana virus, sandfly, epidemiology, Draa El Mizan, Kabylia,
Algeria.

ملخص
عن والناجمة البشر، ذلك في بما الثدييات في لوحظت التي الطفيلية واألمراض الليشمانيات، داء ينتقل
لداء شكلين هناك. اإلناث الرمل وذبابة القارضة، الحشرات طريق عن الليشمانيا، جنس من البروتوزوا
عن اإلبالغ تم مؤخرا،. الجلدي الليشمانيا وداء الحشوي الليشمانيا داء: الجزائر في يحدثان الليشمانيا
النابلس صقليةال فيروس( Phlebovirusمن نوع فيروس الفليبو )( arboviruses) فيروس األربو
الجزائرلبحث في ل عملنا جسد وقد. الجزائر شمال في الليشمانيا مع مشترك كما الرمل ذبابة ناقالتلها
الفترة خالل الليشمانيا مع وبائية عالقة وجود على الضوء تسليطو Toscana توسـانـا فيروس عن
العثور تم ،(القبائل منطقة) لميزاناع ادر منطقة في الدراسـة أجريت .2014 إلى 2012 عام من الممتدة
4283 منهاو الرمل ذبابة 25781 منالليشامانيـا دء ة المصابين بلمجموع المركز المستوطـن على
الفصد سكان تركيبة لتحديد تصنيفية لدراسة موضوعا كانت
. P. perniciosus هيمنة مع Sergentomyia و Phlebotomus جنس من أنواع ثمانية تسجيل تم
الرمل ذباب في الليشمانيا عن البحث كان RT-PCR تقنية على القائم ، الجزيئي التحليل استخدم. سالفا
سمحت. الحشرات هذه في الفليبرو فيروسات عن للكشف ، المتداخل المتسلسل البلمرة بتفاعل متبوعة
.الجزائر في مرة ألول (TOSV) توسكانا فيروسقدر اإلمكـان ووصف تحديد التقنيات هذه
الفيروسي للجينوم الكامل التسلسل تنفيذ تم ،( ATCC CCL81) فيرو خاليا على الفيروس بعد زراعة
فيروس توسكانا " ، عليها الحصول تم التي ساللةتسمى (. NGS) التالي الجيل تسلسل بواسطة
. تونس في المعزولة TOSV ساللة من قريبة ، A الخط في جينيا فيلو وتصنف ،" 189 ساللة ،"الجزائر
، Sero-neutralization و Western Blot هما ، مصليين اختبارين بإجراء الدراسة استكملت وقد
اختيارهم تم أشخاص من مصل 370 على إجراؤهما تم الميزان درعة سكان في عشوائيا
ضد المضادة األجسام وتحييد الليشمانيا مع إيجابية حالة 86 عن التوالي على االختبارات هذه كشفت قد
TOSV 171 في sera 1000 عند الفيروس من جرعة باستخدام TCID50 ، تشير التي ، النتائج وهذه
. المنطقة هذه في ذبابة الرمل في. الفيروس وجود مع كافية تكوندراع الميزان في تنتشر TOSV أن إلى
TOSV فيروسية عدوى وجود إلى االختبارين كال في مصل 11 إيجابية تشير ، ذلك إلى باإلضافة
.الميزان درعة سكان بين الحشوي الليشمانيات لداء مصاحبة
، القبايل ، الميزاندراع ، األوبئة علم ، الرمل ذبابة ، توسكانا فيروس ، الليشمانية: المفتاحية الكلمات
الجزائر

Liste des tableaux
Tableau I Phlébotomes d’Algérie………………………………………………………... 14
Tableau II Répartition des espèces de phlébotomes en Algérie en fonction des étages
bioclimatiques………………………………………………………………… 15
Tableau III Répartition selon le sous-genre des principaux complexes
du genre Leishmania………………………………………………………...... 18
Tableau IV Principales espèces de Leishmania pathogènes pour l’Homme et formes de
Leishmanioses……………………………………………………………….... 20
Tableau V Principales espèces de phlébotomes vectrices de leishmanioses dans l’Ancien
Monde………………………………………………………………………… 20
Tableau VI Liste des virus du genre Phlebovirus…………………………………………. 38
Tableau VII Séquences des amorces pour la RT-PCR……………………………………... 56
Tableau VIII Séquences des amorces pour la PCR nichée………………………………….. 56
Tableau IX Mix de la RT-PCR……………………………………………………………. 57
Tableau X Mix de la PCR nichée………………………………………………………… 57
Tableau XI Cycle de la RT-PCR…………………………………………………………... 58
Tableau XII Cycle de la PCR nichée………………………………………………………. 59
Tableau XIII Produits du mix du séquençage………………………………………………. 61
Tableau XIV Programme du thermocycleur………………………………………………… 61
Tableau XV Distribution des sérums et des contrôles sur une plaque de micro-titration
utilisée dans le test de séroneutralisation virale……………………………… 66
Tableau XVI Abondance relative selon le sous-genre des phlébotomes capturés par pièges
huileux………………………………………………………………………… 76
Tableau XVII Abondance relative par sexe des espèces recensées………………………….. 77
Tableau XVIII Abondance relative selon le sous-genre des phlébotomes capturés par pièges
lumineux………………………………………………………………………. 78
Tableau XIX Abondance relative par sexe des espèces recensées………………………….. 79
Tableau XX Caractéristiques de la population étudiée…………………………………….. 80
Tableau XXI Répartition des anticorps neutralisants du virus Toscana selon le sexe et l’âge
pour 1000 TCDI50……………………………………………………………………………………. 87
Tableau XXII Répartition des anticorps neutralisants du virus Toscana selon le sexe et l’âge
pour 100 TCDI50……………………………………………………………………………………… 87
Tableau XXIII Résultats du Western Blot et de la séroneutralisation………………………… 88
Tableau XXIV Résultats du Western Blot et de la séroneutralisation selon l’âge et le
sexe…………………………………………………………………………… 89

Liste des figures
Figure 1 Morphologie générale d’un phlébotome adulte…………………………… 7
Figure 2 Tête d’un phlébotome adulte………………………………………………. 7
Figure 3 Pièces buccales impaires (à l’exception du labium)
chez P. sergenti femelle……………………………………………………
8
Figure 4 Thorax d’un phlébotome………………………………………………....... 8
Figure 5 Aile de phlébotome………………………………………………………... 9
Figure 6 Génitalia mâle……………………………………………………………... 9
Figure 7 Génitalia femelle…………………………………………………………... 10
Figure 8 Cycle de vie du phlébotome……………………………………………….. 11
Figure 9 Exemples de gites de repos………………………………………………... 12
Figure 10 Répartition des leishmanioses, viscérale et cutanée en Algérie…………… 24
Figure 11 Formes de Leishmania…………………………………………………….. 27
Figure 12 Cycle biologique de Leishmania…………………………………………... 29
Figure 13 Illustration de l’interaction
entre les deux types de réponse Th1 et Th2………………………………..
31
Figure 14 Structure du génome et stratégie de codage des Bunyaviridae……………. 35
Figure 15 Arbre phylogénétique des phlébovirus transmis par les phlébotomes dans
l’Ancien Monde (Neighbour-Joining)……………………………………..
37
Figure 16 Distribution des phlébovirus transmis par
les phlébotomes en Afrique………………………………………………...
40
Figure 17 Analyse phylogénique du virus SFS, Algérie, basée sur la séquence 207-
nt (A) ou 67aa (B) du gène de la polymérase……………………………
41
Figure 18 Analyse phylogénétique de séquences de phlébovirus obtenues à partir de
phlébotomes algériens, basée sur la séquence 67-AA du gène de la
polymérase…………………………………………………………………
42
Figure 19 Situation géographique de Draa El Mizan………………………………… 46
Figure 20 Commune de Draa El Mizan………………………………………………. 47
Figure 21 Exemples de sites de capture des phlébotomes……………………………. 48

Figure 22 Lieux d’emplacement des pièges adhésifs………………………………… 48
Figure 23 Pièges lumineux CDC accrochés………………………………………….. 49
Figure 24 Capture des phlébotomes aux pièges lumineux CDC. ……………….........
50
Figure 25 Dissection de phlébotome femelle pour la détection de promastigotes……
52
Figure 26 Incubation des sérums avec les bandelettes du Western blot……………… 53
Figure 27 Principe de la réaction de le séroneutralisation……………………………. 65
Figure 28 Phlébotome adulte mâle (A). Phlébotome adulte femelle gorgée (B)……..
67
Figure 29 Critères d’identification du sous-genre Larroussius ……………………
68
Figure 30 Critères d’identification morphologique de P. perniciosus………………..
69
Figure 31 Critères d’identification morphologique de P. longicuspus…………………
70
Figure 32 Critères d’identification morphologique de P. perfiliewi………………….
71
Figure 33 Critères d’identifications morphologiques de P. papatasi…………………
72
Figure 34 Critères d’identification morphologique de P. sergenti. …………………..
73
Figure 35 Critères d’identification morphologique de S. minuta …………………….
74
Figure 36 Abondance relative selon le genre des phlébotomes capturés par pièges
huileux ……………………………………………………………………..
75
Figure 37 Abondance relative par sexe des espèces recensées………………………. 77
Figure 38 Abondance relative selon le genre des phlébotomes capturés par pièges
lumineux……………………………………………………………………
78
Figure 39 Abondance relative des espèces recensées………………………………… 79
Figure 40 Résultats du test Western-Blot…………………………………………….. 80
Figure 42 Répartition des cas positifs à Leishmanias selon le sexe………………….. 81
Figure 43 Résultat du gel d’agarose pour la RT-PCR en utilisant les amorces
SFNV-NP1…………………………………………………………………
82
Figure 44 Résultat du gel d’agarose pour la PCR nichée en utilisant les amorces
SFNV-NP2…………………………………………………………………
82
Figure 45 Blast du gène N codant pour la nucléocapside du pool 189 comparé à
celui de la souche tunisienne TOSV T152 (JX867536)……………………
83

Figure 46 Structure génomique du Virus TOSV-Algérie 189………………………... 84
Figure 47 Analyse phylogénétique des souches de phlébovirus en utilisant les
séquences d’acides nucléiques des segments L (1), M (2) et S (3)………..
85-86
Figure 48 Moyenne géométrique des titres pour les anticorps neutralisants de TOSV
selon les groupes d’âges (1000 TCID50)…………………………………...
87
Figure 49 Moyenne géométrique des titres pour les anticorps neutralisants de TOSV
selon les groupes d’âges (100 TCID50)…………………………………….
88

INTRODUCTION

Introduction
1
Les phlébotomes sont des insectes diptères largement répandus à travers le monde; du fait de
leur adaptation à différents climats, leur distribution englobe les zones tropicales et tempérées
chaudes. Ils sont signalés pour la première fois en Algérie en 1912 par Foley et Leduc; leur
statut taxonomique a été défini et leur pouvoir vectoriel démontré grâce aux travaux réalisés à
l’Institut Pasteur par les frères Sergent et Parot (Dedet et al., 1984). Seules les femelles sont
hématophages et impliquées dans la transmission de leishmanioses à l’Homme et à divers
mammifères, par inoculation d’un protozoaire flagellé du genre Leishmania, parasite du
système monocytes-macrophages.Une vingtaine d’espèces de ce genre peut infecter
l’Homme. Les leishmanioses, objet d’un grand nombre d’études écobiologiques, sont pour la
plupart des zoonoses liées à l’état de l’environnement et aux conditions climatiques; leur
répartition géographique étant superposable à celle de leurs vecteurs. Elles constituent chez
l’Homme un groupe de pathologies classées en quatre formes cliniques: leishmaniose cutanée
localisée (forme spontanément guérissable), leishmaniose cutanée diffuse (forme rebelle à
toute thérapeutique), leishmaniose cutanéo-muqueuse (forme mutilante) et leishmaniose
viscérale (forme grave). A l’échelle mondiale, la prévalence est estimée à 12 millions de cas
avec une incidence annuelle de 1 à 1,5 million de cas de leishmaniose cutanée et de 300 000
à500 000 cas de leishmaniose viscérale. Le nombre annuel de décès par leishmaniose
viscérale est de 70 000 et 350 millions d’individus sont exposés au risque d’infection (OMS,
2010). Un tiers seulement des nouveaux cas sont officiellement déclarés (OMS, 2017).
Les leishmanioses sévissent à l’état endémique sous forme de foyers dans près de 90 pays et
quatre continents (Afrique, Europe, Asie, Amérique). Il existe dans chaque foyer une ou
plusieurs formes cliniques dues à une ou à plusieurs espèces de Leishmania avec comme
réservoirs des mammifères sauvages et domestiques et comme vecteurs des phlébotomes
spécifiques. Les vecteurs démontrés sont les espèces du genre Phlebotomus dans l’Ancien
Monde (Sud de l’Europe, Afrique, Proche-Orient et Asie) et celles du genre Lutzomyia dans le
Nouveau Monde (Amériques du Nord, du Sud et Centrale) (Quitterie et al., 2005; Samake,
2006; Dedet, 2009; Estevez, 2009).
En Algérie, l’un des pays du pourtour méditerranéen et du Maghreb les plus concernés, ces
affections sont endémiques sous trois entités cliniques: la leishmaniose viscérale, la
leishmaniose cutanée sporadique du nord et la leishmaniose cutanée zoonotique (Harrat et
Belkaïd, 2002). La Kabylie est connue depuis longtemps comme une région à foyers très
actifs de la leishmaniose viscérale, regroupant près de 50% des cas (Harrat et al., 1995) puis
de la leishmaniose cutanée sporadique (Dedet et Addadi, 1977; Achour et Madiou, 2008 ).
Cette région présente une grande géodiversité, avec des zones très contrastées de points de vue

Introduction
2
bioclimatique, géomorphologique et végétationnel, offrant ainsi des biotopes très diversifiés
pour les phlébotomes (Mouloua et al. 2017). Ces insectes sont également impliqués dans la
transmission de la bartonellose et d'arboviroses. La bartonellose, causée par une rickettesie,
Bartonella bacilliformis, est une affection restreinte à certaines régions d’Amérique latine.
Elle apparait chez l’Homme sous deux formes cliniques distinctes, la verruga péruvienne
(forme caractérisée par une éruption cutanée d’éléments verruqueux) et la fièvre d’Oroya
(forme souvent fatale caractérisée par le syndrome hémolytique (Leger et Depaquit, 2001).
Les arboviroses sont dues à un ensemble hétérogène de virus à ARN qui sont transmis à des
vertébrés par des arthropodes hématophages (phlébotomes, moustiques, tiques) dans les
différentes régions du monde avec une répartition liée à celle de leurs vecteurs.
Les arbovirus transmis par les phlébotomes appartiennent à trois genres de familles
différentes: Phlebovirus, Orbivirus et Vesiculovirus. Dans la région méditerranéenne, seuls les
Phlebovirus ont un impact en santé humaine (Izri et al., 2006). Ce genre est composé de 9
espèces dont les virus des fièvres à phlébotomes (ou virus des fièvres de trois jours):virus
Sandfly Fever Naple (SFNV) (incluant le virus Toscana) et le virus Sandfly Fever Sicilian
(SFSV).
Ces virus se rencontrent dans toutes les régions du monde, Asie centrale, Amérique, Sud de
l’Europe, Afrique (Afrique du Nord, Maroc, Tunisie et Egypte) (Cusi et al., 2010). Les
résultats d’enquêtes épidémiologiques ont montré que la diversité des arbovirus dans le bassin
méditerranéen est plus importante que celle initialement soupçonnée et que les populations
vivant au sud et à l'est de la mer méditerranée ont un risque d’infection élevé (Sanbonmatsu-
G'amez et al., 2005; Papa et al., 2006; Konstantinou et al, 2007; Carhan et al., 2010; Bahri et
al., 2011; Ergunay et al., 2011, 2012; KocakTufan et al., 2011).
En Algérie, des cas documentés d’infections dues aux phlébovirus sont totalement absents et
les données épidémiologiques sur ces pathogènes quasi inexistantes. Seules deux études
sérologiques et moléculaires ont été menées dans le Nord de l’Algérie et rapporté la présence
des virus SFS et SFN parmi la population et de deux virus distincts mais apparentés au virus
SFS, chez les phlébotomes (Izri et al., 2008; Moureau et al., 2010). Ces résultats ont montré
que la population est fortement exposée au risque d’infections par les phlébovirus
Le virus Toscana est connu comme pathogène pour l’Homme, l’affectant avec la fréquence la
plus élevée en Europe par rapport aux autres phlébovirus et il est le seul à présenter un
tropisme pour le système nerveux. Il a été découvert en 1971 chez des phlébotomes en Italie
et sa pathogénicité vis-à-vis de l’Homme prouvée en 1980 (Calisher et al., 1987; Verani et al.,
1982). Dans l’ancien monde, il circule dans plusieurs pays du sud de l’Europe (Portugal,

Introduction
3
Espagne, France, Italie, Croatie et Turquie) (Alkan et al., 2013; Bichaud et al., 2013; Kay et
al., 2009) et en Tunisie (Bahri et al., 2010) et au Maroc (Es-Sette et al., 2016). Il représente
un réel problème de santé publique dans le bassin méditerranéen où il est à l’origine
d’infections restreintes à la période d’activité de son vecteur et dont les plus sévères sont les
formes neuro-invasives (Dionisio et al., 2001; Sanchez-Seco et Navarro, 2005; Bahri et al.,
2010). Ce virus est méconnu à l’échelle nationale avec particulièrement l’absence de données
épidémiologiques.
Par ailleurs, le lien épidémiologique entre les leishmanioses humaines due à Leishmania
infantum et les infections par le virus Toscana, a été statistiquement établi dans le sud-est de
la France (Bichaud et al., 2011).
Sur la base de ces données bibliographiques, il nous a paru important d’apporter d’autres
éléments de connaissances sur les phlébovirus en Algérie et de démontrer l’existence d’un
lien épidémiologique entre les infections dues à ces derniers et les leishmanioses. Pour ce
faire, nous avons mené de pair, une étude épidémiologique prospective de la leishmaniose
viscérale dans la région de Draa El Mizan, connue comme foyer actif de leishmanioses en
Kabylie et une étude virologique pour éventuellement détecter et identifier les phlébovirus qui
y circulent.
Ce travail est présenté en quatre chapitres dont le premier fait la synthèse des connaissances
acquises sur les aspects morpho-anatomiques et bioécologiques des phlébotomes et sur deux
pathologies transmises par ces derniers, les leishmanioses et les arboviroses.
Le deuxième chapitre décrit la zone d’étude, le matériel et la méthodologie expérimentale.
Les résultats sont présentés en trois parties (études entomologique, parasitologique et
virologique) dans le troisième chapitre et discutés dans le quatrième chapitre.
Une conclusion générale reprend l’essentiel des résultats et met l’accent sur les perspectives.

CHAPITRE I
SYNTHESE BIBLIOGRAPHIQUE

Chapitre I Synthèse bibliographique
4
1. Les phlébotomes
1.1. Taxonomie
1.1.1. Historique de la taxonomie
La classification des phlébotomes n’a jamais fait l’unanimité parmi les spécialistes de la
taxonomie car plusieurs systèmes declassifications ont été proposés. Ce qui a donné lieu à de
nombreuses controverses ce sont essentiellement les critères morphologiques utilisés dans la
diagnose de ces insectes, qui sont parfois d’application et d’interprétation difficiles.
Parmi les conceptions sur la classification des phlébotomes qui reflètent les divergences
d’opinion des auteurs:
- Walker (1851) regroupe les phlébotomes dans la famille des Phlebotomidae
- Kertesz (1902-1904) distingue deux sous-familles, les Psychodinae et les Phlebotominae et
les classe dans la famille des Psychodidae
- França et Parrot (1920) retiennent le taxon Phlebotomus comme genre et le subdivisent en
cinq sous-genres: Phlebotomus, Paraphlebotomus, Brumptomyia, Lutzomyia et Sergentomyia.
- Adler et Theodor (1929) suggèrent que les phlébotomes forment une famille.
- Theodor (1948) divise la famille des Psychodidae en 4 sous-familles dont la sous-famille des
Phlebotominae avec deux genres: Phlebotomus et Sergentomyia. Cette systématique est basée
sur les travaux de Sinton (1927) et Nitzulescu (1931) qui ont porté sur la variabilité de la
morphologie de l'armature cibariale, du pharynx et des spermathèques.
- Rhodendorf (1964) valide la famille des Phlebotomidae
- Theodor (1965) valide la sous famille des Phlebotominae
- Perfiliev (1966) inclut tous les phlébotomes dans la famille des Phlebotomidae
- Lewis (1973) retient le système de classification de Theodor (1948).
- Abonnenc et Léger (1976) proposent une nouvelle classification dans laquelle la famille des
Phlebotomidae (Walker, 1851) est divisée en 3 sous-familles et 10 genres:
* Sous-famille des Euphlebotominae avec 4 genres: Spelaeophlebotomus (Theodor, 1948),
Idiophlebotomus (Quate et Fairchild, 1961), Phlebotomus (Rondani, 1843) et Sergentomyia
(França et Parrot, 1921).
* Sous-famille des Neophlebotominae avec 4 genres: Grassomyia (Theodor, 1958), Parvidens
(Theodor et Mesghali, 1964), Brumptomyia (França et Parrot, 1921) et Lutzomyia (França,
1921).
* Sous-famille des Disphlebotominae avec 2 genres, Warileya (Hertig, 1948) et Hertigia
(Fairchild, 1949).

Chapitre I Synthèse bibliographique
5
- Lewis (1982), dans sa revue taxonomique du genre Phlébotomes propose 12 sous genres:
Adlerius (Nitzulescu,1931), Anaphlebotomus (Theodor, 1948), Australophlebotomus
(Theodor, 1946), Euphlebotomus (Theodor, 1948), Idiophlebotomus (Quate et Fairchid,
1961), Kasaulius (Lewis, 1982), Larroussius (Nitzulescu, 1931), Paraphlebotomus (Theodor,
1948), Phlebotomus (Rondani, 1840), Spelaeophlebotomus (Theodor,1948), Synphlebotomus
(Theodor, 1948), Transphlebotomus (Artemiev, 1948).
- Leng (1987) crée le genre Chinius.
- Artemiev (1991) propose 24 genres en en créant de nouveaux et en élevant certains sous-
genres au rang de genre.
La classification, basée essentiellement sur la biogéographie et adoptée par commodité,
place les Phlebotominae au rang de sous-famille avec cinq genres, Phlebotomus et
Sergentomyia pour l’Ancien Monde et Lutzomyia, Warileya et Brumptomyia pour le
Nouveau Monde (Locksley et Louis,1992).
Seuls les genres Sergentomyia et Phlébotomes sont présents dans la région méditerranéenne et
comportent chacun quatre sous-genres.
Le genre Phlebotomus se distingue par des soies des tergites abdominaux toutes dressées,
armature buccale (cibarium) absente ou rudimentaire, spermathèques complètement ou
incomplètement segmentées et style des génitalia comportant 4 à 5 épines.
Le genre Sergentomyia se caractérise par des soies abdominales couchées dans la plupart des
espèces, cibarium bien développé, spermathèques tubulaires, capsuliformes ou segmentées,
style des génitalia des mâles à quatre longues épines avec une petite soie sur le côté interne
(França et Parrot, 1921).
Actuellement, près de 900 espèces de phlébotomes sont décrites et largement réparties dans
les régions tropicales et tempérées (Izri, 2006). La taxonomie de ces espèces inclue
uniquement des critères morphologiques avant l’apport d’approches phylogénétiques. Les
critères utilisés, sont les soies antennaires, les dents cibariales, les dents du pharynx, la
nervation alaire, l’implantation des soies au niveau du thorax et de l’abdomen et les génitalia
(Abonnenc, 1972; Leger et al., 1983; Killick-Kendrick et al., 1991).
Lorsque les caractères morphologiques ne permettent pas de distinguer deux espèces, le
taxonomiste fait appel à l'analyse des isoenzymes (Ward et al., 1981). Des études plus
récentes ont contribué à une systématique moléculaire de ces insectes avec l’analyse des
séquences d’ADN (Depaquit et al., 2000a; Depaquit et al., 2000b; Aransay et al. 2000;
Depaquit, 2014).

Chapitre I Synthèse bibliographique
6
Outre l’approche morphologique et moléculaire, de nouveaux outils d’identification sont
désormais employés telle la spectrométrie de masse de type MALDI-TOF. Cette méthode
consiste à caractériser un échantillon à partir du spectre de masse moléculaire des différentes
protéines présentes, permettant d’identifier des arthropodes vecteurs au niveau de l’espèce
(C.N.E.V. 2014). Il est cependant nécessaire de disposer d’une banque de spectres des espèces
qui pourront ainsi être identifiées. L’efficacité du MALDI-TOF a pu être prouvée récemment
sur les phlébotomes dont le profilage de protéines représente un premier pas vers
l'établissement d'une base de données de spectres de protéines, permettant une identification
rapide et fiable des espèces (Dvorak et al., 2014).
1. 1. 2.Positionsystématiquedesphlébotomes
La position systématique des genres Phlebotomus et Sergentomyia selon Dolmatova et
Demina (1971), actuellement adoptée par la majorité des spécialistes est la suivante:
Règne: Animalia
Phylum: Arthropoda
Sous-Phylum: Hexapoda
Classe: Insecta
Sous-Classe: Pterygota
Super-Ordre : Endopterygota
Ordre : Diptera
Famille : Psychodidae (Bigot, 1845)
Sous-Famille: Phlebotominae (Kertesz, 1904)
Genres : Phlebotomus (Rondani, 1843)
Sergentomyia (França et Parrot, 1920)
1.2. Morphologie
Les phlébotomes sont des insectes de très petite taille, de 2 à 3 mm de long, à peine visibles à
l’œil nu, de couleur claire, jaune pâle à brun et au corps grêle et allongé (Izri, 2006; Lewis,
1973) et couvert d’une vestiture épaisse qui leur permet un vol silencieux (Léger et Depaquit,
2001). La tête, formant un angle de 45°avec le thorax, donne une allure bossue au phlébotome
(Léger et Depaquit, 2002) (Fig.1).

Chapitre I Synthèse bibliographique
7
Fig.1.Morphologie générale d’un phlébotome adulte (Niang et al., 2000).
La tête est formée en grande partie par une capsule chitineuse (épicrane), limitée de
chaque côté par un grand œil composé. Elle porte les pièces buccales dont l’ensemble forme
le proboscis, une trompe courte, le clypeus, les fosses tentoriales, le front et l’occiput (Killick-
Kendrick, 1990). Sur la région frontale sont attachées deux antennes formées chacune de seize
segments dont deux basaux et quatorze, beaucoup plus longs et minces, constituant le
flagellum (Lewis, 1982) (Fig.2).
Chez la femelle, les pièces buccales sont de type « piqueur » : un labium, deux maxilles, un
labre-épipharynx (Killick-Kendrick, 1990) et des mandibules dentelées (Léger et Depaquit,
2001) (Fig.3).
Fig.2.Tête d’un phlébotome adulte (Leger et Depaquit, 2001)

Chapitre I Synthèse bibliographique
8
Fig.3. Pièces buccales impaires (à l’exception du labium) chez P.sergenti femelle (Depaquit,
1997)
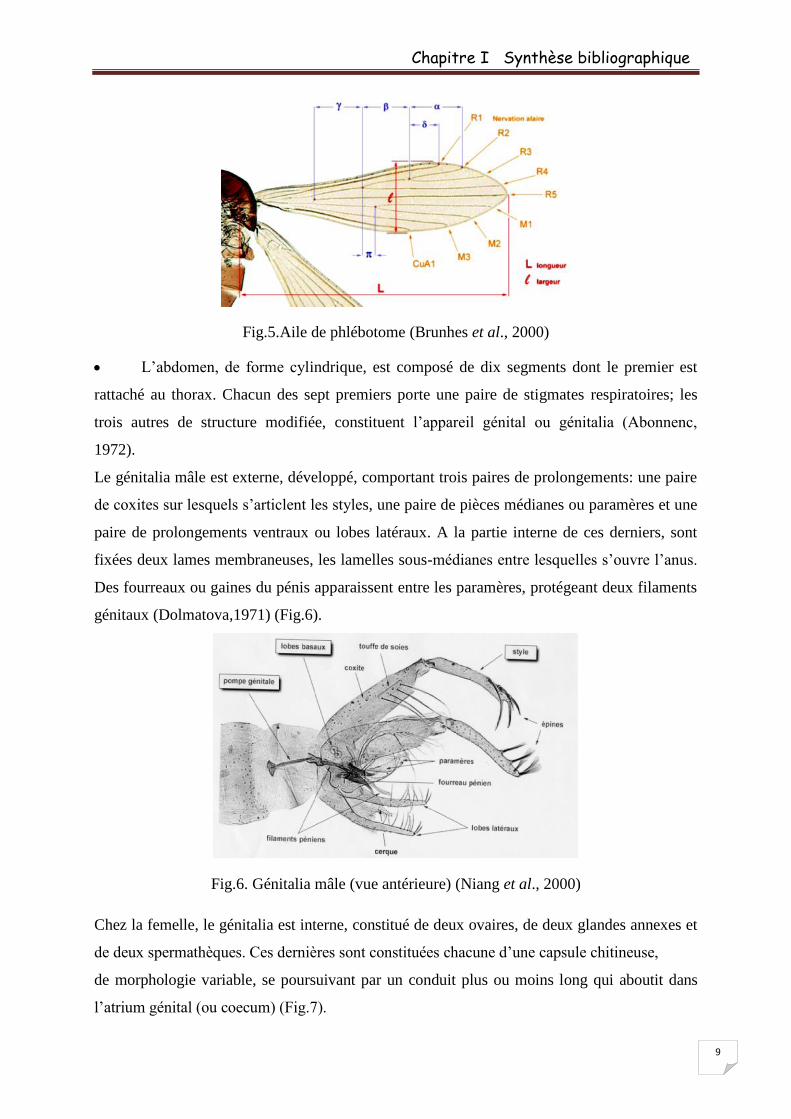
Le thorax est convexe et bien développé comme chez tous les diptères (Léger et
Depaquit, 2001). Il porte une paire d’ailes lancéolées et typiquement relevées en V au repos et
des balanciers ou altères ayant pour rôle d’assurer l’équilibration de l’insecte durant son vol
(Dedet et al., 1984). Les ailes comprennent neuf nervures, sept longitudinales et deux
transversales. Sur chacun des trois segments thoraciques est fixée une paire de pattes
articulées, longues, fines et couvertes de soies (Léger et Depaquit, 2001) (Fig.1, 4 et 5).
Fig.4. Thorax d’un phlébotome (Niang et al., 2000)
Chapitre I Synthèse bibliographique
9
Fig.5.Aile de phlébotome (Brunhes et al., 2000)
L’abdomen, de forme cylindrique, est composé de dix segments dont le premier est
rattaché au thorax. Chacun des sept premiers porte une paire de stigmates respiratoires; les
trois autres de structure modifiée, constituent l’appareil génital ou génitalia (Abonnenc,
1972).
Le génitalia mâle est externe, développé, comportant trois paires de prolongements: une paire
de coxites sur lesquels s’articlent les styles, une paire de pièces médianes ou paramères et une
paire de prolongements ventraux ou lobes latéraux. A la partie interne de ces derniers, sont
fixées deux lames membraneuses, les lamelles sous-médianes entre lesquelles s’ouvre l’anus.
Des fourreaux ou gaines du pénis apparaissent entre les paramères, protégeant deux filaments
génitaux (Dolmatova,1971) (Fig.6).
Fig.6. Génitalia mâle (vue antérieure) (Niang et al., 2000)
Chez la femelle, le génitalia est interne, constitué de deux ovaires, de deux glandes annexes et
de deux spermathèques. Ces dernières sont constituées chacune d’une capsule chitineuse,
de morphologie variable, se poursuivant par un conduit plus ou moins long qui aboutit dans
l’atrium génital (ou coecum) (Fig.7).

Chapitre I Synthèse bibliographique
10
Le génitalia mâle, les spermathèques et l’armature buccale de la femelle, dont la morphologie
diffère d’une espèce de phlébotome à une autre, sont utilisés comme critères taxonomiques.
Fig.7.Génitalia femelle (vue antérieure) (Niang et al., 2000)
1.3. Biologie et éco-éthologie
Les phlébotomes sont des insectes holométaboles, se nourrissent essentiellement de sucs
floraux et les femelles sont de plus hématophages, type telmophage.
L’accouplement a lieu peu de temps après leur émergence, souvent au crépuscule et dure
environ une quinzaine de minutes.Une fois fécondée, la femelle se nourrit sur un vertébré
(plusieurs espèces de mammifères dont l’Homme, oiseaux, reptiles et amphibiens) pour
assurer le développement de ses œufs. Le sang est absorbé d’un lac hémolymphatique qu’elle
crée en dilacérant les tissus de l’hôte avec ses mandibules. Puis, pour digérer son repas, elle
prend une période de repos dans un endroit ombragé et protégé du vent. Le temps de
maturation des œufs dépend de l’espèce et de la température ambiante. L’accouplement peut
se produire aussi dans le gîte de repos après la prise du repas de sang.
Les œufs pondus un à un, donnent à l’éclosion des larves vermiformes, de 0,5 à 4 mm de
long, composées de trois parties, tête, thorax et abdomen dont le 8ème
segment porte les
stigmates respiratoires et l’extrémité caudale, deux paires de longues soies. Ces larves vivent
dans les gites de ponte qui varient selon les espèces mais qui sont en général riches en
matières organiques, humides et obscurs. Elles se nourrissent de matières organiques au
moyen de pièces buccales broyeuses, subissent 4 mues avant de se transformer chacune en
une nymphe. En saison froide ou lors de conditions défavorables, les larves du stade 4 entrent
en diapause et s’enfoncent dans le sol (Dolmatova et Demina, 1971; Abonnenc, 1972).
La nymphe est de couleur blanc-jaunâtre; l’extrémité de l’abdomen porte les soies caudales et
l’exuvie du dernier stade larvaire qui lui permet de se fixer sur son support. Elle vit dans les
mêmes types de niches écologiques que ceux de la larve, ne se nourrit pas, entre en état de vie

Chapitre I Synthèse bibliographique
11
ralentie puis effectue la mue imaginale donnant l’adulte. Celui-ci émerge à partir d’une fissure
située sur le côté dorsal de la cuticule (Abonnenc, 1972)(Fig.8).
La durée du développement post-embryonnaire varie selon les conditions climatiques, de 20 à
90 jours (Izri et al., 2006). La longévité des adultes est de quelques mois et dépend des
conditions d’humidité et de température optimales. Le mâle a une durée de vie plus courte que
celle de la femelle. Celle-ci vit en moyenne deux mois au cours desquels elle peut s’accoupler
plusieurs fois mais ne prend qu’un repas de sang par cycle gonotrophique qui se répète tous
les 3 à 10 jours (Leger et Depaquit, 1999).
Fig.8. Cycle de vie du phlébotome (Garnotel, 2011)
Les phlébotomes sont présents toute l’année dans les régions tropicales mais seulement
pendant la saison chaude dans les régions tempérées. La période de leur vie active diffère
selon le climat (Dolmatova et al., 1971). Dans le bassin méditerranéen, ils sortent
massivement au printemps et en été par des temps pluvieux (Rioux et al., 1969).
Ces insectes se déplacent par des vols lents, silencieux et courts avec un rayon de
déplacement qui n’atteint généralement pas 1,5 km. Leur activité est nocturne, commence au
crépuscule par des temps calmes et sous des conditions de température et d’humidité
satisfaisantes. Dans la journée, ils se réfugient dans des endroits calmes, à l’abri de la lumière
et du vent (terriers de petitsmammifères sauvages, clapiers, niches, creux d’arbres,
anfractuosité de rochers…). En général, ils s’éloignent peu de ces abris où les femelles
trouvent les hôtes vertébrés sur lesquels elles doivent se gorger; ces abris peuvent constituer
des gîtes de repos ou de ponte favorables au développement larvaire (Fig.9).
Les phlébotomes prolifèrent dans des terriers de rongeurs sauvages qui constituent une niche
écologique naturelle ou primaire. La disparition des hôtes entraine l’extinction de la niche et le
déplacement des insectes vers un autre abri, par exemples:

Chapitre I Synthèse bibliographique
12
- Phlebotomus papatasi est retrouvé près des terriers habités par Psammomys obesus mais qu’il
quitte dès qu’ils sont abandonnés par le rongeur.
-P. perniciosus n’est rencontré qu’autour des clapiers habités (Izri et al., 1990; 2006).
A partir de cette niche, les phlébotomes se dispersent car les femelles vont pondre ailleurs
mais dans une aire limitée car elles se déplacent sur de courtes distances. Si la ponte se
produit dans un abri d’animaux domestiques (clapiers, étables..), les larves trouvent de bonnes
voire de meilleures conditions de développement que dans la niche écologique primaire. Ces
deux espèces de phlébotomes abondent donc dans les abris d’animaux domestiques (Izri et
Bellazoug, 1993).
Cette niche écologique secondaire ou niche anthropique prend le relais de la niche primaire
sauvage. Elle permet l’introduction et la multiplication des phlébotomes dans le milieu péri-
domestique ou domestique.
Les préférences trophiques des phlébotomes pour les différents vertébrés varient selon
l’espèce, on distingue des espèces anthropophiles, herpétophiles, ornithophiles et simiophiles.
Le genre Phlebotomus très ubiquiste pique plusieurs espèces de mammifères à l’exception de
P. papatasi qui est essentiellement anthropophile.
Les espèces du genre Sergentomyia sont herpétophiles mais S. minuta parroti peut
occasionnellement piquer l’Homme (Dedet et Addadi, 1977).
Le comportement de l’adulte femelle distingue :
- des espèces à activité diurne et des espèces à activité nocturne;
- des espèces endophages (espèces qui piquent à l’intérieur des bâtiments) et des espèces
exophages (espèces qui piquent à l’extérieur)
- des espèces endophiles (repos à l’intérieur après piqûre, pour la digestion du repas sanguin)
et des espèces exophiles (repos à l’extérieur après piqûre).
Fig.9. Exemples de gites de repos, barbacane (A), enclos (B) et chenil (C)
(Prudhomme, 2015)

Chapitre I Synthèse bibliographique
13
1.4. Répartition géographique
1.4.1. Dans le monde
Les phlébotomes ont une aire de distribution très étendue, ils sont présents sur tous les
continents mais sans dépasser certaines latitudes. Ils n’ont pas été signalés en Europe du
Nord ; leur présence est rare en Amérique du Nord et en Australie, ils abondent dans les zones
tropicales et équatoriales de l’Afrique et de l’Amérique orientale (Leger et Depaquit, 2002).
Ils sont fréquents dans le bassin méditerranéen, Afrique du Nord (Dedet et al., 1984;
Belazzoug, 1991).
La répartition géographique des genres Phlebotomus et Sergentomyia est comme suit (Rioux
et al., 1991):
Phlebotomus ariasi (Tonnoir, 1921): Sous-région méditerranéenne occidentale
Phlebotomus longicuspis (Nitzulescu, 1930): Méditerranée Sud-occidentale: Maghreb;
Espagne.
Phlebotomus perfiliewi (Parrot, 1930): Méditerranée orientale et sub-occidentale.
Phlebotomus perniciosus (Newstead, 1911): Bassin méditerranéen occidental
Phlebotomus papatasi (Scopoli, 1786): de l’Inde au Bassin méditerranéen.
Phlebotomus alexandri (Sinton, 1928): de la Chine au Maghreb et à l’Espagne
Phlebotomus kazeruni (Theodor et Mesgali, 1964): zonesirano-touranienne et saharo-
sindienne.
Phlebotomus sergenti (Parrot, 1917): de l’Inde et la Russie à la Méditerranée occidentale
Sergentomyia antennata (Newstead, 1912): du Moyen Orient à l’Afrique.
Sergentomyia fallax (Parrot, 1912): Afrique du Nord
Sergentomyia minuta (Rondani, 1843): Pourtour méditerranéen
Sergentomyia lewisi (Parrot, 1948): espèce transsaharienne.
1.4.2. En Algérie
1.4.2.1. Espèces recensées
Les phlébotomes sont répartis à travers tout le pays, de l’étage humide jusqu’à l’étage
saharien (Dedet et al.,1984; Belazzoug, 1991; Berchi, 1990; Izri, 1994). A l’heure actuelle, 24
espèces sont connues dont 14 appartiennent au genre Phlebotomus et 10 au genre
Sergentomyia (Tableau I).

Chapitre I Synthèse bibliographique
14
Tableau I. Phlébotomes d’Algérie (Dedet et al., 1984; Bellazoug, 1991; Bounamous et al.,
2008; Berdjane-Brouk et al., 2011)
Sous-famille des Phlebotominae
Genre Phlebotomus Genre Sergentomyia
Phlebotomus (Phlebotomus) papatasi (Scopoli,
1786)
Sergentomyia (Sergentomyia) antennata (Newstead,
1912)
Phlebotomus (Phlebotomus) bergeroti (Parrot,
1934)*
Sergentomyia (Sergentomyia) fallax(Parrot, 1921)
Phlebotomus (Paraphlebotomus) sergenti (Parrot,
1917)
Sergentomyia (Sergentomyia) minuta (Alder et Theodor,
1927)
Phlebotomus (Paraphlebotomus) alexandri
(Sinton, 1928)
Sergentomyia (Sergentomyia) schwetzi (Alder, Theodor
et Parrot, 1929) *
Phlebotomus (Paraphlebotomus) kazeruni
(Theodor et Mesghali, 1964) **
Sergentomyia (Parrotomyia) africana (Newstead, 1912)
Phlebotomus (Paraphlebotomus) chabaudi (Croset,
Abonnenc et Rioux, 1970)
Sergentomyia (Parrotomyia) eremitis (Parrot et De
Jolinière, 1945) *
Phlebotomus (Paraphlebotomus) Riouxi (Depaquit,
Killick-Kendrick, Leger, 1998)
Sergentomyia (Parrotomyia) Lewisi (Parrot,1948)
Phlebotomus (Transphlebotomus), Mascittii
(Grassi, 1908)
Sergentomyia (Grassomyia) dreyfussi (Parrot, 1933)
Phlebotomus (Larroussius) perniciosus (Newstead,
1911)
Sergentomyia (Sintonius) christophersi (Sinton, 1927)
Phlebotomus (Larroussius) ariasi(Tonnoir, 1921) Sergentomyia (Sintonius) clydei (Sinton, 1928)
Phlebotomus (Larroussius) perfiliewi
(Parrot, 1930)
Phlebotomus (Larroussius) langeroni
(Nitzulescu, 1930)
Phlebotomus (Larroussius) chadlii
(Rioux, Juminer et Gibily ,1966)
Phlebotomus (Larroussius) longicuspis
(Nitzulescu, 1930)
*Espèces recensées au Sahara Central.
**Espèce recensée pour la première fois en Algérie par Berchi et al. (1986) à partir de
phlébotomes récoltés à Mila (Nord-Est algérien).

Chapitre I Synthèse bibliographique
15
1.4.2.2. Répartition des espèces selon les étages bioclimatiques
Le tableau suivant indique la composition de la faune phlébotomienne propre à chaque étage
bioclimatique.
Tableau II. Répartition des espèces de phlébotomes en Algérie en fonction des étages
bioclimatiques (Dedet et al., 1984; Bellazoug, 1991, Berchi, 1993).
Etage
Humide
Etage
sub-humide
Etage
semi-aride
Etage
aride
Etage
Saharien
Sergentomyia
Minuta (Alder
et Theodor,
1927)*
Sergentomyia
minuta (Alder et
Theodor, 1927)*
Sergentomyia
minuta (Alder et
Theodor, 1927)
Sergentomyia
minuta (Alder et Theodor,
1927)
Sergentomyia
minuta (Alder et
Theodor, 1927)
Phlebotomus
Perniciosus
(Newstead,
1911)
Phlebotomus
Perniciosus
(Newstead, 1911)
Sergentomyia
Fallax (Parrot,
1921)
Segentomyia
Fallax (Parrot, 1921)*
Sergentomyia
Fallax (Parrot,
1921)
Phlebotomus
ariasi
(Tonnoir, 1921)
Phlebotomus
langeroni
(Nitzulescu,
1930)
Phlebotomus
Perniciosus
(Newstead, 1911)*
Sergentomyia
antennata
(Newstead, 1912)
Sergentomyia
christophersi
(Sinton, 1927)
Phlebotomus
perfiliewi
(Parrot, 1930)
Phlebotomus
perfiliewi
(Parrot, 1930)
Phlebotomus
langeroni
(Nitzulescu, 1930)
Sergentomyia
dreyfussi
(Parrot, 1933)
Sergentomyia
Dreyfussi
(Parrot,1933)
Phlebotomus
Sergenti
(Parrot, 1917)
Phlebotomus
Sergenti (Parrot,
1917)
Phlebotomus
perfiliewi
(Parrot, 1930)
Phlebotomus
perniciosus
(Newstead, 1911)
Phlebotomus
perfiliewi
(Parrot, 1930)
Phlebotomus
Chadlii (Rioux,
Juminer et
Gibily ,1966)
Phlebotomus
Chadlii (Rioux,
Juminer et
Gibily,1966)
Phlebotomus
Sergenti (Parrot,
1917)
Phlebotomus
langeroni
(Nitzulescu, 1930)
Phlebotomus
Sergenti (Parrot,
1917)
Phlebotomus
longicuspis
(Nitzulescu,
1930)
Phlebotomus
longicuspis
(Nitzulescu,
1930)
Phlebotomus
Chadlii (Rioux,
Juminer et
Gibily,1966)
Phlebotomus
perfiliewi
(Parrot, 1930)
Phlebotomus
Chadlii (Rioux,
Juminer et Gibily,
1966)
Phlebotomus
papatasi
(Scopoli, 1786)
Phlebotomus
Papatasi
(Scopoli, 1786)
Phlebotomus
longicuspis
(Nitzulescu, 1930)
Phlebotomus
Sergenti (Parrot, 1917)
Phlebotomus
longicuspis
(Nitzulescu,
1930)
Phlebotomus
Papatasi
(Scopoli, 1786)
Phlebotomus
Chadlii (Rioux,
Juminer et Gibily, 1966)
Phlebotomus
papatasi
(Scopoli, 1786)*
Phlebotomus
Chabaudi (Croset,
Abonnenc et Rioux,
1970)
Phlebotomus
Longicuspis
(Nitzulescu, 1930)
Phlebotomus
Papatasi (Scopoli, 1786)
P.alexandri (Sinton, 1928)
* Espèce prédominante.

Chapitre I Synthèse bibliographique
16
1.5. Epidémiologie
1.5.1. Facteurs bioclimatiques
La prolifération et la répartition des différentes espèces de phlébotomes sont influencées par
les facteurs bioclimatiques et en particulierpar la température et la pluviosité. Ces insectes sont
présents toute l’anné dans les régions tropicales où leur multiplication est favorisée par une
température de 30°C et un degré d’humité élévé (Abonnenc, 1972). En régions tempérées et du
fait des conditions climatiques plus variables, ils ne prolifèrent qu’à la saison estivale, quand la
température est élevée (20°C et plus) et en présence d’une humdité suffisante. Ces paramètres
expliquent leur absence à une altitude supérieure ou égale à 1000-1500 m (Ripert et
Ladier, 2005).
Le climat a un effet sur la nature de la végétation et par conséquent sur la répartition des
espèces animales. Les phlébotomes se distribuent selon le type de végétation auquel sont
adaptés leurs hôtes notamment les rongeurs sauvages. Parmi les espèces de phlébotomes,
Phlebotomus ariasi abonde dans la partie Nord de la méditerranée et P. papatasi dans la
partie Sud où se rencontre le rongeur Psammomys obesus. Ces régions se caractérisent par une
végétation respectivement hydrophile et xérophile.
La distribution des espèces animales peut être inégale au sein d’une même région car elle
dépend du milieu et des conditions géographiques. Psammomys obesus vit dans les régions
septiques où il se nourrit de chénopodiacées. Il construit ses terriers dans les terrains sableux
où poussent ces plantes xérophiles dont le développement est favorisé par la présence des
nappes phréatiques ou de points d’eau. L’existence de niches écologiques sauvages des
phlébotomes est de la sorte liée à la nature du sol, de la flore, de la faune et du climat (Le
Houerou, 1977).
Les variations météorologiques peuvent modifier aussi les conditions locales avec par
conséquent des répercussions sur la végétation et la répartition des populations animales. Des
précipitations intenses contribuent au développement de la végétation et à la pulullation des
rongeurs et des phlébotomes inféodés à leurs terriers. Une sécheresse prolongée a un effet
inverse (Izri et Bellazoug, 1993).
1.5.2.Facteurs anthropiques
Les activités humaines, principalement celles liées au développement agricole, ont des effets
sur l’environnement et les conditions naturelles de prolifération des espèces animales. La mise
en valeur de terres sauvages, a perturbé les biotopes des rongeurs, source de repas sanguins
des phlébotomes. Les rongeurs sont refoulés à la périphérie des cultures où ils reconstruisent
leurs terriers (Izri et al., 2006). La création de ces nouvelles niches écologiques a entrainé une

Chapitre I Synthèse bibliographique
17
multiplication des phlébotomes à proximité des espaces habités. Ce phénomène a été observé
notamment dans les régions steppiques avec l’exploitation de zones arides qui a bouleversé
les sites naturels de Psammomys obesus et où prolifère Phlebotomus papatasi. De plus, ces
activités agricoles sont souvent suivies d’élevages de bétail et d’installation d’abris pour les
animaux domestiques, pour ne citer que ces exemples, qui offrent aux phlébotomes les
conditions propices à leur développement.
Par ailleurs, l'augmentation des déchets domestiques rejetés par l'Homme modifie les milieux
et permet la création de terriers d’autres rongeurs qui attirent ces insectes.
De telles modifications environnementales contribuent à la pullulation d’animaux, hôtes
réservoirs potentiels de pathogènes et des phlébotomes vecteurs. Ceci a comme conséquence
une extension de maladies vers des régions jusque-là indemnes avec l’apparition de nouveaux
foyers.
2. Rôle vecteur des phlébotomes
De nombreux travaux, qui s’inscrivent dans le cadre d’enquête sur les maladies à transmission
vectorielle, ont montré le rôle des phlébotomes dans la transmission des arboviroses et des
leishmanioses, objets de notre étude, et de la bartonellose.
2.1.Leishmanioses
2.1.1. Historique
Selon Theodorides (1997), Cox (2002) et Bari (2006) :
- Les premières descriptions des leishmanioses sous ses formes cutanées datent de l’antiquité
- Au Xème siècle, des médecins arabes dont Al Bukhari font la description d’une affection
cutanée appelée Balkh douloureux, du nom d’une région de l’Afghanistan.
- En 1756, la leishmaniose cutanée est nommée « Bouton d’Alep » par Alexander Russel
après l’avoir décrite chez un patient turc.
- En 1885, le parasite est mis en évidence dans une lésion cutanée et décrit par David
Cunningham.
- En 1898, Peter Borovsky décrit le kinétoplaste du parasite et incrimine ce protozoaire
comme responsable de la leishmaniose cutanée.
- La leishmaniose viscérale est décrite par des médecins indiens sous le nom de Kala azar.
- William Leishman (1900) et Charles Donovan (1903) découvrent séparément le parasite
dans des ponctions spléniques de malades présentant un Kala-azar et l’incriminent comme
agent étiologique de cette maladie.
- Ronald Ross, en 1903, donne à cet agent le nom de Leishmania donovani.

Chapitre I Synthèse bibliographique
18
- En 1908, Charles Nicolle et ses collaborateurs mettent en évidence l’agent responsable de la
leishmaniose viscérale infantile à Tunis et le nomment L. infantum. Ils démontrent par ailleurs
le rôle du chien comme réservoir de la maladie.
- A l’Institut Pasteur d’Alger et en 1921, les frères Sergent et leurs collaborateurs apportent
expérimentalement la preuve et sur leurs propres personnes le rôle vecteur de ces derniers. Ils
montrent ainsi la transmission du parasite L. major, l’agentdu «Clou de Biskra» par le vecteur
Phlebotomus papatasi.
2.1. 2. Généralités
Les leishmanioses sont dues à des protozoaires flagellés, du genre Leishmania, transmis lors
d’une piqûre d’un phlébotome femelle infecté. Les parasites se multiplient dans les cellules du
système des phagocytes mononucléés de divers mammifères tels les rongeurs, les canidés et
l’Homme. Le genre Leishmania est divisé en deux sous-genres: Leishmania sensu stricto et
Viannia; les espèces de ces deux sous-genres sont regroupées en complexes (Tableau III).
Chez le phlébotome, les espèces du sous-genre Leishmania se développent à la jonction du
mésentéron (intestin moyen) et du proctodeum (intestin postérieur) et celles du sous-genre
Viannia dans ces deux portions intestinales.
Tableau III. Répartition selon le sous-genre des principaux complexes du genre Leishmania
(Dedet, 2001)
Sous-genre Leishmania Sous-genre Vannia
Ancien monde L. donovani
L. infantum
L. tropica, L. major
L. killicki,
L.aethiopica
Nouveau monde L. chagasi L. Mexicana
L. amazonensis
L. venezuelensis
L. guyanensis
L. panamensis
L. shawi, L. naiffi
L. lainsoni
L. peruviana
L. brasiliensis
Environ 20 espèces du genre Leishmania sont pathogènes pour l’Homme (OMS, 2010) et
selon leur tropisme, elles déterminent plusieurs groupes de pathologies avec des
manifestations cliniques différentes; celles-ci incluent principalement des formes viscérales,

Chapitre I Synthèse bibliographique
19
des formes cutanées et des formes cutanéo-muqueuses (Tableau IV). La majorité des cas
restent asymptomatiques.
La leishmaniose viscérale (LV) est la forme la plus grave; son pronostic est lié à la rapidité du
diagnostic et à la mise en place du traitement. Elle se manifeste par une fièvre irrégulière, une
hépatosplénomégalie, une anémie et une perte de poids. Leishmania infantum est l’agent
étiologique de la LV humaine et de la leishmaniose canine dans les pays du pourtour
méditerranéen, au Moyen-Orient et en Amérique du Sud. Il touche principalement les enfants
en bas âge et les sujets immunodéprimés (notamment ceux infectés par le VIH). Un nombre
important de zymodèmes (variants enzymatiques) a été identifié indiquant un polymorphisme
enzymatique important. Leishmania donovani, espèce anthroponotique et responsable de la
LV en Afrique de l’Est et dans le sous-continent indien, affecte des individus de tout âge
(Pearson et Sousa, 1996; Alvar et al., 2012).
La leishmaniose cutanée (LC), maladie très fréquente, se présente sous divers aspects
cliniques selon l’espèce de Leishmania en cause et la réponse immunitaire de l’hôte. Elle
provoque le plus souvent des lésions ou ulcères uniques ou multiples sur les parties
découvertes du corps laissant des cicatrices indélébiles (Reithinger et al., 2007). Elle est
connue sous plusieurs noms vernaculaires: Bouton d'Orient ou Clou de Biskra sur le pourtour
méditerranéen et Bouton d'Alep au Proche-Orient. Dans l’ancien monde, cette maladie est due
principalement à L. tropica et L. major (Alvar et al., 2012; Reithinger et al., 2007); la
première responsable d’une leishmaniose anthroponotique et la seconde d’une leishmaniose
zoonotique (Reithinger et al., 2007).
La leishmaniose cutanéo-muqueuse (LCM) est la forme mutilante. Les lésions cutanées
s’étendent aux muqueuses du nez, des oreilles et de la gorge, entrainant leur destruction de
manière partielle ou totale.
Sur les 900 espèces de phlébotomes décrites, 70 ont été identifiées comme vecteurs potentiels
de leishmanioses (Ready, 2013); les espèces des genres Phlebotomus et Lutzomyia en sont les
vecteurs exclusifs respectivement dans l’Ancien et le Nouveau Monde (Killick-Kendrick,
1990) (Tableau V).

Chapitre I Synthèse bibliographique
20
Tableau IV. Principales espèces de Leishmania pathogènes pour l’Homme et formes de
leishmanioses (OMS, 2010).
Espèces de
Leishmania de
l’Ancien
monde
Espèces de
Leishmania du
Nouveau
monde
Formes de
leishmanioses
Aspects cliniques Réservoirs
L.donovani Viscérale
anthroponotique
Kala azar Homme
L. infantum Viscérale zoonotique Kala azar Chien
L. chagasi Viscérale zoonotique Kala azar Chien, renard
L. tropica C cutanée simple L Lésion sèche
( (Bouton d’Orient)
Homme
L. major Idem Lésion humide Rongeurs
L.mexicana Cutanée simple Ulcère du chiclero Rongeurs,marsupiaux
L. peruviana Idem Uta Chien
L. guyanensis Idem Pian-bois Fourmiliers,Paresseux
L. panamensis Idem Ulcère unique Paresseux, singe
L. aethiopica Cutanée diffuse Diffuse et grave Daman
L. amazonensis Cutanée diffuse Idem Homme
L. pifanoi Idem Idem Rongeurs
L. brasiliensis Cutanéo-muqueuse Espundia Rongeurs, chien
Tableau V. Principales espèces de phlébotomes vectrices de leishmanioses dans l’Ancien
Monde (Dolmatova et Demina, 1971; Léger et Depaquit, 2001).
Genre Sous/Genres Espèces vectrices
Phlebotomus
Phlebotomus P. papatasi, P. dubosqi
Paraphlebotomus P.sergenti, P.alexandri,
Synphlebotomus P.martini, P.guggisbergi, P.ansarii
Larroussisus P.ariasi,P.langeroni,P.neglectus,P.perfiliewi
P. perniciosus, P. tobbi, P.longicuspis, P.pedifer
Adlerius P.chinensis
Euphlebotomus P. argentipes

Chapitre I Synthèse bibliographique
21
Les leishmanioses, toutes formes cliniques confondues, se répartissent sur quatre continents
(Afrique, Amérique, Asie et Europe) et sont endémiques dans près de 90 pays de l’Ancien
monde et du Nouveau monde (OMS, 2010).
La LV est présente en Asie du Sud-Est, Afrique de l’Est, Amérique centrale et du Sud et dans
le bassin méditerranéen. En 2015, plus de 90% des cas sont notifiés au Brésil, Éthiopie, Inde,
Kenya, Somalie et Soudan (OMS, 2017). Dans les pays du pourtour méditerranéen, cette
maladie est principalement infantile mais en association avec l'infection à VIH, elle affecte la
population adulte avec un risque de moralité élevé (Lachaud et al., 2002, Alvar et Desjeux,
2003).
La plupart des cas de LC surviennent dans le bassin méditerranéen, au Moyen-Orient, en Asie
centrale et dans les Amériques. Les pays les plus touchés, enregistrant les 2/3 des cas en 2015,
sont l’Afghanistan, l’Algérie, la Syrie, l’Iran, le Brésil et la Colombie (OMS, 2017).
Plus de 90% des cas de LCM sont enregistrés au Brésil, en Bolivie et au Pérou (OMS, 2017).
La population exposée au risque d’infection par les leishmanioses est estimée à 350 millions
de personnes à l’échelle mondiale, avec une prévalence de 12 millions. L'estimation de
l'incidence annuelle mondiale se situe entre 1,5 et 2 millions de nouveaux cas, se distribuant
entre 1 à 1,5 millions de cas de leishmanioses cutanées, souvent bénignes mais parfois
invalidantes et 300.000 à 500.000 cas de leishmanioses viscérales, mortelles en l’absence de
traitement et responsables de 70 000 décès. (OMS, 2010). Les chiffres enregistrés seraient
sous-estimés en raison des nombreux malades n’ayant pas accès aux soins et/ou des cas non
déclarés (OMS, 2007).
La transmission des leishmanioses est liée aux facteurs environnementaux (altitue, versant ) et
climatiques (humidité, température …). Ces derniers ont un effet sur le développement du
parasite chez le vecteur et dans le changement de comportement et de distribution des vecteurs
ou des réservoirs. L’expansion de ces affections sont conditionnées donc par les changements
climatiques mais également par l’augmentation de facteurs de risque que Desjeux (1999) a
résumé comme suit:
- Installation de populations dans des zones d’endémie du fait de problèmes démographiques
ou de manque de terres agricoles;
- mouvements de populations pour raison professionnelle ou autres;
- développement de nouveaux projets agricoles ou industriels conduisant des populations non
immunisées dans des zones d’endémie,
- urbanisation accélérée et/ou non planifiée;

Chapitre I Synthèse bibliographique
22
- détérioration du tissu économique et social dans les banlieues des villes où s’amassent des
populations à faibles revenus en raison d’un exode rural massif;
- modifications de l’environnement par l’homme (construction de barrages et de systèmes
d’irrigation, augmentation des déchets domestiques ou industriels rejetés).
2.1.3. Leishmanioses en Algérie
Quatre formes cliniques de leishmanioses sévissent à l’état endémique dans des zones
bioclimatiques distinctes. Au nord du pays sont rencontrées la forme viscérale (LV) et la
forme cutanée sporadique du nord (LCN); au sud la forme cutanée zoonotique (LCZ) et la
forme cutanée signalée plus récemment à Ghardaïa (Harrat et al., 2009) et à Constantine
(Boubidi et al., 2011), due respectivement à L.killicki et L. tropica (Fig.10).
2.1.3.1. Leishmaniose viscérale
La LV s’étend sur toute la partie nord du pays, sous un climat subhumide à humide. Elle
provoque plusieurs nouveaux cas chaque année avec une fréquence qui varie d’une région à
une autre. Sa transmission est attribuée principalement à l’espèce Phlébotomes (Larroussius)
perniciosus (Izri et al., 1990). Le principal réservoir est le chien domestique (Canis
familiaris) (Belazzoug 1986b, Dereure, 1993)
Les cas humains et canins sont dus essentiellement à Leishmania infantumMON-1 (Harrat et
al., 1996). D’autres zymodèmes peuvent être responsables de la LV humaine en particulier
chez des patients coinfectés par le VIH (Pratlong et al., 1995).
Le foyer de la Grande Kabylie regroupe à lui seul près de 50 % de cas humains et où on
dénombre environ 400 nouveaux cas par an (Harrat et al.,1995).
Cette affection est responsable de 6 % de décès dans le foyer de Tizi-Ouzou, qui reste le plus
actif dans le Maghreb (Harrat et al., 1995; Djerbouh et al., 2005). Elle touche habituellement
des enfants mal nourris, vivant en zone rurale, mais affecte depuis quelques années de plus en
plus de sujets n’ayant jamais quitté les grandes zones urbaines. Ce phénomène d’urbanisation
de la maladie, constaté dans la région d’Alger même, serait lié, d’une part, à l’exode rurale, à
cause de l’insécurité de milliers de citoyens campagnards, d’autre part, à la dégradation de
l’environnement, à la prolifération de chiens malades errants et enfin, à la multiplication des
gites de phlébotomes. Par ailleurs, l’expansion de la métropole vers la banlieue entrainerait un
rapprochement de citoyens des foyers sauvages d’infection, augmentant ainsi le risque de
contamination (Harrat et al., 1996).

Chapitre I Synthèse bibliographique
23
2.1.3.2. Leishmaniose cutanée sporadique du Nord
La LCN, causée par Leishmania infantum Mon-24, occupe la même aire géographique que la
LV. Elle est transmise par Phlebotomus (Larroussius) perfiliewi; le réservoir est inconnu, bien
que l’agent pathogène ait été isolé du chien (Izri et al.,1992).
Les cas observés sont signalés principalement en Kabylie et dans l’algérois, quelques cas ont
été notifiés dans l’Ouest du pays (Tlemcen) (Boudghene et al., 1991). Les lésions sont
généralement uniques, infiltrées, rarement ulcérées; leur évolution vers la guérison spontanée
est assez lente et peut durer parfois plus de deux ans (Dedet, 1999).
2.1.3.3. Leishmaniose cutanée zoonotique
Cette forme de leishmaniose est provoquée par Leishmania major zymodème MON-25
(Harrat et al., 1995) (Pratlong et al.,1990) et transmise par Phlebotomus (Phlebotomus)
papatasi (Sergent et al., 1926; Izri et al., 1992). Deux rongeurs sauvages, Psammomys obesus
et Meriones shawi constituent le réservoir de la maladie (Belazzoug, 1983; Belazzoug,
1986a). Le rôle vecteur des phlébotomes a été décrit pour la première fois à Biskra et Abadla,
foyers historiques du Clou de Biskra (Sergent et al.,1921).
Cette affection se manifeste par des lésions uniques ou multiples se caractérisant par une
ulcération qui évolue spontanément vers la guérison en 3 à 6 mois en laissant une cicatrice
indélébile. Elle sévit à l'état endémo-épidémique au niveau des zones arides et semi-arides de
l’Afrique du Nord. Elle était cantonnée à la région de Biskra jusqu’au début des années 1980
où une épidémie a éclaté dans la région voisine de M’Sila (Izri, 1994). Depuis, elle n’a cessé
de s’étendre à toutes les régions steppiques du sud maghrébin et depuis vers le nord
(Boudrissa, 2012).
Selon les conditions climatiques, des épidémies apparaissent périodiquement, frappant des
milliers de personnes et à chaque épisode, la maladie se déplace vers le nord menaçant un
territoire comptant les 2/3 de la population algérienne. En 2004, la barrière de l’Atlas tellien a
été franchie (Boudrissa et al., 2012). Cette extension s’explique principalement par la
présence du vecteur, P. papatasi, qui s’adapte bien au climat plus humide du nord. Cependant,
la prolifération de cet insecte est limitée par l’altitude, il est peu présent au-dessus de 600
mètres (Léger et Depaquit, 1999; Rioux et al., 1967).
La troisième forme de LC, identifiée dans la région de Ghardaïa par Harrat et al. (2009), a été
décrite pour la première fois au Maghreb en 1986 dans le sud tunisien (Bouratbine, 2005).
L’agent pathogène est Leishmania killicki MON-301 avec comme vecteur Phlebotomus
sergenti. Le réservoir reste inconnu, cependant Ctenodactylus gundi en est fortement suspecté

Chapitre I Synthèse bibliographique
24
(Boubidi et al., 2011). Elle a été découverte avec un nouveau variant enzymatique,
Leishmania killicki MON-306 (Mansouri et al., 2012).
C’est une forme bénigne, responsable de lésions sèches pouvant durer plus de deux ans
(Boubidi et al., 2011). La guérison spontanée ou après traitement laisse une cicatrice
indélébile. La récidive après guérison est possible.
Fig.10. Répartition des leishmanioses, viscérale et cutanée en Algérie (Eddaikra, 2016)
2.1.4. Circulation du parasite
Dans le milieu naturel, seuls les phlébotomes peuvent assurer la transmission et donc la
pérennité des leishmanies. Pour être vecteur, l'insecte doit:
- se nourrir sur un animal vertébré porteur de parasites,
- permettre le développement de ces derniers,
- vivre suffisamment longtemps pour assurer, lors d’une prochaine piqûre, leur passage sur un
autre vertébré pouvant devenir à son tour réservoir de parasites.
La spécificité parasitaire semble bien établie avec les phlébotomes (Killick-Kendrick, 1985).
Elle peut être particulièrement étroite, comme dans le cas du couple P. sergenti-L.tropica
(Rioux et al., 1992), ou étendue au sous-genre, comme pour les leishmanies du complexe L.
infantum transmises par les phlébotomes du sous-genre Laroussius. Cette notion de spécificité
vecteur-parasite permet de définir des espaces à risque correspondant aux régions où se
rencontre le vecteur. Ces espaces sont déterminés par l'étude de la chorologie vectorielle.

Chapitre I Synthèse bibliographique
25
Diverses espèces de mammifères constituent les hôtes réservoirs du genre Leishmania
(O.M.S, 1990). La circulation des parasites entre ces foyers se fait par les déplacements des
vecteurs ou des animaux réservoirs. On distingue trois types de foyers.
- Le foyer primaire : la leishmaniose est une zoonose où seuls des mammifères sauvages sont
infectés et constituent le réservoir. Les parasites sont transmis d'hôte à hôte par des
phlébotomes zoophiles (Izri, 1996).
- Le foyer secondaire: la leishmaniose est une zoonose des mammifères sauvages et
domestiques (les chiens pour la LV). L’Homme est infecté occasionnellement par
l’intermédiaire de l’insecte vecteur (Chang et al., 1985).
- Le foyer tertiaire, dit anthroponotique : le réservoir est strictement humain et la
leishmaniose se propage sur un mode endémo-épidémique. Ce type de foyer tertiaire est
retrouvé pour la LV principalement en Inde et en Afrique orientale (Chang et al., 1985).
Pour les leishmanioses tégumentaires, les réservoirs animaux les plus courants dans l’Ancien
Monde sont des rongeurs (Psammomys obesus, Meriones shawi) pour L. major, des damans
(Procavia habessinica) pour L. aethiopica.
Dans le Nouveau Monde, des paresseux (Choloepus hoffmanni et C. didactylus), des
fourmiliers (Tamandua tetradactyla), des marsupiaux, des primates et des rongeurs sont les
réservoirs de diverses espèces de Leishmania responsables de formes cutanées.
Pour la LV, les réservoirs animaux habituels sont divers canidés, souvent le chien (Canis
familiaris), le chacal (Canis aureus) en Asie centrale, les renards (Vulpes vulpes) en Europe
méditerranéenne et Lycalopex vetulus et Cerdocyon thous en Amérique du Sud ainsi que le
chien viverrin (Nyctereutes procyonoides) en Chine. D'autres mammifères, principalement
des félidés et des rongeurs, trouvés naturellement infectés, peuvent être impliqués dans les
cycles sauvages de la LV (Desjeux, 1991; Dedet, 1994).
2.1.5. Techniques diagnostiques
- Examen microscopique direct: c’est l’examen le plus spécifique pour un diagnostic de
certitude. Il repose sur la mise en évidence du parasite à partir de ponction ganglionnaire ou
de moelle osseuse pour la LV et de prélèvement de la bordure de la lésioncutanée ou
muqueusedans le cas des leishmanioses tégumentaires (LC et LCM). Des frottis sont pratiqués
à partir de ces échantillons, colorés au May-Grünwald-Giemsa (MGG) et observés en
microscopie optique.
- Examen après culture : constitue un complément pour le diagnostic parasitologique avec
l’obtention de formes promastigotes. Les prélèvements sont incubés à une température entre
21 et 27 C°, dans un milieu de culture additionné d’antibiotiques pour éviter le risque de

Chapitre I Synthèse bibliographique
26
contamination. Les milieux de Novy-MacNeal-Nicolle (NNN), de Brain Heart Infusion(BHI),
d’Evans et de Schneider sont les plus utilisés (Pan et al, 1993).
- Techniques indirectes (techniques immunologiques): la sérologie est utilisée dans le
diagnostic de la LV; elle est inutile dans les autres formes de leishmanioses.
Les tests les plus employés sont l’Enzyme Linked ImmunoSorbent Assay (ELISA),
l’immunofluorescence indirecte (IFI), l’électrosynérèse, l’hémagglutination et le western blot
(WB); ce dernier étant le test le plus sensible.
- Techniques de biologie moléculaire: complètent les cultures et les techniques sérologiques
par détection de l’ADN parasitaire. La Polymerase Chaine Reaction (PCR), réalisable à partir
de tous les types de prélèvements (moelle osseuse, biopsies ganglionnaires, sang…), permet
de détecter les acides nucléiques du parasite, par amplification d’une séquence donnée. Cette
technique présente plusieurs avantages, applicable sur des prélèvements peu invasifs (sang
périphérique, urines), plus rapide, plus sensible et plus spécifique que les autres méthodes
directes. Elle présente aussi un intérêt dans le typage moléculaire des souches, le suivi
évolutif des sujets traités et comme marqueur précoce de rechute chez l’immunodéprimé
(Lachaud et al., 2000).
2.1.6. Le parasite
2.1.6.1.Taxonomie
La position de Leishmania Ross, 1903 dans la classification de Levine et al. (1980) est la
suivante :
-Règne: Protista (Haeckel, 1866)
- Sous-Règne: Protozoa (Goldfuss, 1817, Siebold, 1848)
- Phylum: Sarcomastigophora (Honigberg et Balamuth, 1963)
- Sous-Phylum: Mastigophora (Diesing, 1866)
- Classe: Zoomastigophorea (Calkins, 1909)
- Ordre: Kinetoplastida (Honigberg, 1963; Emend. Vickerman, 1976)
- Sous-Ordre: Trypanosomatina (Kent, 1880)
- Famille: Trypanosomatidae (Doflein, 1901 Emend. Grobben, 1905)
- Genre: Leishmania Ross, 1903
Une classification fondée sur l’étude du profil électrophorétique de certaines isoenzymes
(zymodèmes) du parasite a été proposée par Rioux et al. (1990) et révisée en 1993 (Rioux et
Lanotte, 1993). Chaque électromorphe est identifié par rapport à une souche étalon utilisée

Chapitre I Synthèse bibliographique
27
comme marqueur qui correspond à Leishmania infantum zymodème MON-1 (MON=
Montpellier).
2.1.6.2. Morphologie
Les leishmanies sont des eucaryotes unicellulaires caractérisés par la présence d’un
kinétoplaste (organite propre aux kinétoplastidés) qui contient deux types de molécules
d’ADN, les maxi-cercles et les mini-cercles. Elles apparaissent sous deux formes
morphologiques différentes au cours de leur cycle biologique:
-Forme promastigote: organisme allongé, d’environ 5 à 2 µm de long et 1 à 4 µm de large
avec un noyau approximativement central, prolongé au pôle antérieur par un flagelle libre
pouvant atteindre jusqu’à 20 µm de long. Le kinétoplaste se situe entre le noyau et le flagelle
(Killick-Kendrick,1990; Dedet, 1999) (Fig.11 A).
- Forme amastigote: organisme ovalaire ou arrondi de 2 à 6 μm de diamètre, immobile,
enveloppé d’une membrane bien définie, présentant un noyau, et muni d’un flagelle
intracytoplasmique très réduit. Le kinétoplaste est juxtanucléaire (Killick-Kendrick,1990,
Dedet, 1999) (Fig.11 B).
Fig.11. Formes de Leishmania1
A. Formes amastigotes intracellulaires et libres (MGG, x 1 000)
B. Formes promastigotes (MGG, x 200)
2.1.6.3. Cycle biologique
La réalisation du cycle nécessite deux hôtes avec deux stades évolutifs distincts: le stade
promastigote mobile dans le tube digestif de l’hôte invertébré, le phlébotome femelle et le
stade amastigote intracellulaire chez l’hôte vertébré, le mammifère (Homme, chien, rongeurs
sauvages) (Fig.12).
1http://www.uvp5.univ-paris5.fr/campus-parasitologie/cycle2/poly (consulté le 10/08/2018)

Chapitre I Synthèse bibliographique
28
Le phlébotome infecté inocule les formes promastigotes métacycliques au mammifère lors de
son repas sanguin. Avant d’être phagocytés, les parasites adhèrentaux macrophages suite à la
reconnaissance de molécules de liaison présentes à la surface des parasites, la glycoprotéine
63 et les lipophosphoglycanes (GP63 et LPG), par des récepteurs spécifiques de la membrane
du macrophage. Ils sont ensuite internalisés dans la vacuole parasitophore (ou phagosome) par
le biais de pseudopodes émis par le macrophage (Rittig et Bogdan, 2000).Une fois
internalisés, les promastigotes métacycliques perdent la longueur de leurs flagelles (Basset et
al., 2005) et se transforment en amastigotes qui vont survivre (grâce à des stratégies variées)
et se diviser par scissiparité, provoquant l’éclatement du macrophage. Les amastigotes libérés
vont être alors phagocytés par d’autres macrophages. Ils peuvent se disséminer par voie
sanguine et lymphatique et atteindre le système réticulo-endothélial (rate, foie et moelle
osseuse) dans le cas de la LV ou les cellules lymphoïdes de la peau dans le cas de la LC.
Les macrophages contenant les formes amastigotes sont ingérés par le phlébotome au cours
d’un repas sanguin sur un mammifère infecté. Les cellules sont digérées mais les parasites
échappent à la digestion grâce à une membrane péritrophique protectrice élaborée par
l’insecte. Les amastigotes se transforment en promastigotes procycliques qui prolifèrent
activement puis deviennent des formes plus allongées et plus mobiles mais non prolifératives,
ce sont des promastigotes nectomonades. Ces derniers sécrètent une enzyme qui dégrade la
chitine composant la membrane péritrophique puis migrent vers la région antérieure du
mésentéron. Pour éviter d’être éliminés avec les déchets du repas sanguin, ils s’attachent aux
cellules intestinales par le biais d’une liaison entre les lectines présentes à la surface de ces
cellules et les LPG (Sacks et al., 2000). Les nectomonades se transforment en leptomonades
qui se multiplient avant de devenir des promastigotes métacycliques infectants. Le processus
de la métacyclogenèse correspond à un épaississement de la membrane cellulaire par
élongation des LPG, diminuant ainsi leur adhérence aux lectines des cellules intestinales. Ce
phénomène permet aux parasites de migrer vers la valve stomodéale (jonction entre l’intestin
moyen et l’intestin antérieur) et d’être transmis lors d’un prochain repas sanguin, à un nouvel
hôte mammifère.

Chapitre I Synthèse bibliographique
29
Fig.12. Cycle biologique de Leishmania2
2.1.7. Leishmanies et réactions immunitaires
Les Leishmania, parasites des cellules du système des phagocytes mononucléés, survivent et
se multiplient dans ces cellules grâce à des molécules de surfaces telles que les LPG et la
GP63 (63KDa) à rôle protecteur vis à vis de la réponse immunitaire des mammifères (Mosser
et Rosenthal, 1993).
Les formes promastigotes inoculées au mammifère doivent faire face aux protéines du
complément. L’activation de celles-ci par la voie classique entraine la fixation de C3 sur la
membrane plasmique des parasites et son clivage en C3a et C3b. Dès lors, le C3b peut se fixer
au LPG ou à la GP63 du parasite et provoquer sa destruction suite à la formation du complexe
lytique C5b-9 (Puentes et al., 1990). Par ailleurs, les promastigotes utilisent des stratégies qui
leur permettent de résister à la lyse par le complément, telle la protéolyse de C3b et sa
conversion en molécule inactive C3bi par la GP63 (Mosser et Rosenthal, 1993).
2 http://www.parasitologie.univ-montp1.fr/images/cycle.jpg

Chapitre I Synthèse bibliographique
30
Même si ces mécanismes ne sont pas complètement efficaces pour empêcher la formation du
complexe C5b-9, les promastigotes sont partiellement protégés de la lyse grâce au LPG qui
bloque l’accès des complexes C5b-9 à la membrane plasmique (Flippi et al., 2001).
L’allongement des LPG chez les formes métacycliques expliquent la résistance de celles-ci à
la lyse par le complément contrairement aux formes procycliques (Sacks et al., 1995).
Les promastigotes ayant échappé à la lyse par le complément peuvent être phagocytés par les
macrophages. En plus des macrophages; les parasites sont phagocytés par les neutrophiles, les
cellules dendritiques et les mastocytes.
Dans les conditions normales, les macrophages détruisent les pathogènes par la production et
la libération intracellulaire d’anions superoxide O2-, du peroxyde d’hydrogène H2O2, du
radical hydroxyle HO et de l’oxyde nitrique (Bogdan, 1990).
En plus de leur échappement à la lyse par le complément, les leishmanies exploitent le C3
pour une invasion silencieuse dans le macrophage (Bogdan et Röllinghoff, 1998). Ce dernier
présente à sa surface des récepteurs CR1 et CR3 qui se lient aux C3b et C3bi fixés à la surface
du promastigote métacyclique. Cette liaison est suivie par l’internalisation du parasite dans
une vacuole parasitophore, facilitée par lafibronectine, molécule de surface du macrophage
(Mosser et Rosenthal, 1993).
Les LPG présents en abondance chez le promastigote métacyclique empêchent
temporairement la formation du phagolysosome et la libération des anions O2- et du HO après
l’activation du NADPH durant la phagocytose. La fusion tardive des lysosomes au
phagosome permet à la forme promastigote de se transformer en forme amastigote (Bogdan
et Röllinghoff, 1998). Cette forme résiste au milieu hostile (hydrolytique et acide contenant
des protéases et des hydrolases) de la vacuole parasitophore par la présence à sa surface d’une
couche dense de glycocalyx composée de glycol-inositol-phospholipides (GIPL) et des
glycophospholipides (GPL) à rôle protecteur (Burchmore et Barett, 2001).
Les amastigotes se multiplient dans le macrophage devenu quiescent. Une activation tardive
des macrophages peut se produire grâce à l’interféron γ (INF) (Gant et al., 2001).
Les cellules dendritiques infectées produisent l’interleukine 12 (IL-12) qui active les Natural
Killer (NK) lesquels produisent à leur tour l’IFN-ɣ. Ce dernier agit sur les lymphocytes
Thelper 1 CD4+ (Th1 CD4+) qui libèrent les IL-2 et TNF- α (Scharton-Kersten et al., 1995).
Il agit aussi sur les lymphocytes T cytotoxiques CD8+ (LTcCD8+) à réponse immunitaire à
médiation cellulaire. Les LTc libèrent les perforines qui détruisent la cellule infectée (Fig.13).

Chapitre I Synthèse bibliographique
31
Les cellules dendritiques activent aussi les CD4+Th2 (qui peuvent être activé aussi par IL-4
provenant des mastocytes) qui produisent des anticorps à rôle inconnu et assurent le maintien
du parasite (Mutiso et al., 2013; Khadem et Uzonna, 2014; Rodrigues et al., 2016).
Fig. 13. Illustration de l’interaction entre les deux types de réponse Th1 et Th2 (Baneth et al.,
2008)
Les leishmanies sont particulièrement connues par les immunologistes parce que ce sont des
outils très utiles pour comprendre le fonctionnement du système immunitaire. C’est en
étudiant des souris de laboratoire infectées par des leishmanies que ces scientifiques ont réussi
à élucider une partie des mécanismes qui gouvernent la différenciation des différents
lymphocytes au cours d’une réponse immunitaire physiologique.
2.2. Arbovirus
2.2.1. Généralités
Les arboviroses (Arthropod-Borne Virosis en anglais») sont des maladies vectorielles
transmises par différents groupes d’arthropodes hématophages (tiques, moustiques,
phlébotomes et culicoïdes) à des vertébrés réceptifs (mammifères, oiseaux, reptiles et
batraciens). Ce sont des anthropozoonoses dues à un ensemble hétérogène de plus de 500
virus à ARN, de familles différentes (mais dont l'épidémiologie est commune). Ces virus
appartiennent aux Togaviridae, Flaviviridae, Reoviridae, Rhabdoviridae et Bunyaviridae. Ces
familles renferment également des virus autres que les arbovirus.
Les arbovirus, dont une centaine est pathogène pour l’Homme et une quarantaine pour les
animaux, sont rencontrés dans les différentes régions du monde avec une répartition plus ou
moins étendue, selon la présence des vecteurs. Ceux qui affectent l’Homme sont présents en
zones intertropicales et en zones tempérées (tels le virus West-Nil et le virus de l’encéphalite

Chapitre I Synthèse bibliographique
32
à tique). Ils se transmettent par voie sanguine de vecteur à vertébré et de vertébré à vecteur.
L’Homme constitue un hôte accidentel dans le cycle de transmission de plusieurs virus mais
peut en être lui-même le réservoir naturel comme c’est le cas pour les virus de la dengue.
La transmission virale peut être verticale de vecteur à vecteur, le virus persiste dans les œufs
pondus par l’arthropode (après transmission transovarienne) et infecte la génération suivante,
permettant une persistance virale même sans infection de l’hôte vertébré habituel. Ce
phénomène s’observe particulièrement pendant les périodes où le vecteur est saisonnièrement
absent, par exemple pendant l’hiver en zones tempérées.
La réplication virale se produit dans les cellules cibles aussi bien chez le vecteur que chez le
vertébré. L’insecte s’infecte en aspirant le sang de vertébrés infectés; le virus se multiplie
dans son tube digestif puis diffuse et gagne les glandes salivaires. Le vecteur qui ne souffre
nullement de l’infection, transmettra le virus par sa salive au vertébré lors d’une prochaine
piqûre.
Dans le cas des arboviroses humaines, le virus inoculé dans le sang par le vecteur, se multiplie
dans les monocytes-macrophages (système réticulo-endothélial) et l’invasion de l’organisme
donne une virémie qui correspond à la phase systémique de la maladie. Dans la plupart des
cas, l’infection n’évolue pas et reste asymptomatique. Cependant, suite à une incubation
silencieuse de 1à10 jours, elle peut évoluer en deux phases séparées par une courte phase de
rémission. La phase d’invasion se manifeste brutalement, elle est peu caractéristique, dure 2 à
3 jours, avec une fièvre élevée, un malaise général associé à des douleurs diffuses (céphalées,
nausées, myalgies et arthralgies).Une courte rémission de 12 à 36 heures lui succède avec une
température de 37-38°C et la disparition de presque tous les signes généraux. Survient ensuite
une phase d’état caractérisée par un retour de la fièvre en plateau (autour de 40 °C) et des
mêmes signes que ceux de la première phase. A ces signes peuvent s’ajouter d’autres
symptômes permettant de distinguer, selon les virus en cause, trois grands tableaux cliniques:
- Syndromes aigus fébriles: qui sont souvent spontanément résolutifs en 1à 2 semaines, sans
séquelles, avec une asthénie persistance pendant quelques jours à quelques semaines: virus
Chikungunya (Togaviridae), virus Ziga, virus de la dengue et virus West Nil (Flaviviridae),
phlébovirus (virus Toscana, virus de Sicile, virus de Naples) (Bunyaviridae) …
-syndromes encéphalitiques : l’évolution se fait soit vers une guérison sans séquelles, soit vers
l’installation de séquelles neurologiques graves, soit vers un coma fatal : virus des
encéphalites à tiques (Flaviviridae), virus de Californie (Bunyaviridae) etc…

Chapitre I Synthèse bibliographique
33
-syndromes hémorragiques: qui peuvent évoluer rapidement de manière fatale ou vers une
guérison sans séquelles : virus de la fièvrejaune (Flaviviridae), phlébovirus (virus de la vallée
du Rift, Nairovirus) (Bunyaviridae )…
Les arbovirus véhiculés par les phlébotomes se répartissent en trois genres: Phlebovirus
(Bunyaviridae), Orbivirus (Reoviridae) et Vesiculovirus (Rhabdoviridae), présents
respectivement dans l’Ancien Monde et le Nouveau monde, dans le Nouveau Monde et dans
l’ancien Monde et le Nouveau Monde. Dans la région méditerranéenne, seuls les phlébovirus
ont un impact en santé humaine (Izri et al., 2006).
2.2.2. Bunyaviridae
2.2.2.1. Généralités
Les Bunyaviridae (isolés à Bunyamwera, une ville de l’Ouest de l'Ouganda) est l’une des plus
grandes familles des virus à ARN, formant un groupe très diversifié et largement distribué à
l’échelle mondiale et dont les membres sont connus comme des Bunyavirus (Calisher, 1986;
Nichol et al., 2005). Les 350 virus inventoriés se répartissent dans cinq genres
Orthobunyavirus, Hantavirus, Nairovirus, Phlebovirus et Tospovirus. Ces virus infectent les
animaux, invertébrés et vertébrés à l’exception des Tospovirus qui sont des pathogènes de
plantes.
La transmission virale est assurée par des arthropodes à l’exception des Hantavirus. Ces
derniers sont uniques parmi les Bunyaviridae car ils sont transmis par exposition à des
sécrétions ou excrétions contaminées de leurs hôtes (rongeurs, musaraignes ou taupes)
(Elliott, 1997; Kang et al., 2011).
De nouveaux membres de cette famille sont régulièrement identifiés dont quatre au cours du
1er
semestre de l’année 2011(Kang et al., 2011; Marklewitz et al., 2011; Savji et al, 2011; Yu
et al., 2011). La classification a été à l'origine basée sur la réactivité sérologique croisée puis a
inclus la morphologie du virion, l'organisation du génome, les cycles de réplication et les
relations phylogénétiques (Porterfield, 1975; Schmaljohn, 1996; Nichol et al., 2005).
Récemment, il a été proposé que le virus de Gouléako, isolé des moustiques de la Côte
d'Ivoire, soit classé comme un sixième genre de cette famille en raison de sa distance
phylogénétique et du manque de réactivité sérologique croisée avec d'autres Bunyaviridae. Il
se caractérise par un génome de plus petite taille, une absence de protéines non-structurales et
une restriction de réplication dans les cellules chez l’insecte (Marklewitz et al., 2011).
Comme tous les virus à ARN (à l’exception des myxovirus), les Bunyaviridae se répliquent
dans le cytoplasme de la cellule-hôte et la maturation des virions (assemblage des acides
nucléiques et des protéines) se produit au niveau de l’appareil de Golgi. Du fait de la

Chapitre I Synthèse bibliographique
34
segmentation de leur génome, de nombreux membres subissent un réassortiment génétique ce
qui augmente à la fois la diversité et le potentiel évolutif de cette famille et fournit des outils
génétiques pour déterminer le rôle de la pathogenèse de chaque segment (Janssen et al.,
1984).
Les membres de cette famille qui existent dans le monde entier, se propagent pour la plupart
par des cycles de transmission selvatique entre vertébrés ou plantes sensibles et arthropodes
vecteurs. Les facteurs mondiaux qui favorisent l'apparition des infections à Bunyaviridae
comprennent la migration, le changement climatique, l'urbanisation, le volume croissant du
commerce international et des transports, les pressions sur les systèmes de santé et les
changements dans les écosystèmes (déforestation, irrigation et perte de la biodiversité)
(Soldan et Gonzalez-Scarano, 2005; Elliott, 2009; Hollidge et al., 2010).
Les Bunyaviridae ont un impact sur la santé humaine et animale et sur les cultures végétales
et par conséquent sur le développement économique (Nichol et al., 2000; Soldan et Gonzalez-
Scarano, 2005; Elliott, 2009). Généralement, les infections humaines à Bunyaviridae vont
d'une infection bénigne et asymptomatique à une maladie grave, notamment une méningite ou
une fièvre hémorragique.
2.2.2.2. Structure
Les Bunyaviridae sont des virions enveloppés et sphériques, à génome segmenté, constitué de
trois molécules d’acide nucléique simple brin, de polarité négative. Les complexes
ribonucléoprotéiques correspondant aux segments génomiques recouverts par les
nucléoprotéines présentent une symétrie hélicoïdale. L’ensemble est enveloppé d’une double
couche lipidique avec à sa surface des projections de nature glycoprotéique (Gc et Gn) de 5-
10 nanomètre (Schmaljohn, 1996; Gonzalez-Scarano et Soldan, 2011) (Fig.14).
Les segments génomiques sont de tailles différentes : S (Small), M (medium) et L (Large). Ils
codent pour quatre protéines de structure (Gc, Gn, S, and L) et un nombre variable de
protéines non-structurales.
Le segment L code pour la polymérase virale, qui est responsable de la transcription et de la
réplication du génome. Le segment M code pour un seul cadre de lecture ouvert (Open
Reading Frame : ORF) qui est traduit en glycoprotéines virales (Gc, et Gn) et chez le genre
Orthobunyavirus et quelques membres des genres Phlebovirus et Nairovirus, en protéines non
structurales (NSm ou NSm1 et NSm2). Ces dernières ne sont pas essentielles à la réplication
du virus mais pourraient jouer un rôle dans l'assemblage du virus et la suppression de
l’apoptose induite par le virus (Fuller et Bishop, 1982; Pollitt et al., 2006; Shi et al., 2006;
Won et al., 2007).

Chapitre I Synthèse bibliographique
35
Le segment M des Tospovirus code également pour une protéine non structurelle NSm en
générant deux messages subgénomiques et en utilisant le codage ambisens pour exprimer les
protéines Gn et Gc à partir d'un ORF dans l'ARNc et une protéine non structurale NSm à
partir d’un ORF dans l'ARNv (Schmaljohn, 1996).
Le segment S code pour la protéine N qui, en plus d’encapsider l'ARN génomique, fonctionne
comme un chaperon d'ARN (Mir et Panganiban, 2006). De plus, le segment S des
Orthobunyavirus, Tospovirus et Phlebovirus code pour une autre protéine non structurale
(NSs) par un cadre de lecture chevauchant (Orthobunyavirus) ou par un mécanisme de codage
ambisens (Phlebovirus, Tospovirus) (Schmaljohn,1996).
Les études moléculaires ciblant les NSs ont montré que cette protéine joue un rôle important
dans l’échappement à la réponse immunitaire innée, servant de suppresseur d'ARN silencieux
et inhibant la synthèse protéique de la cellule hôte (Bridgen et al., 2001; Weber et al., 2001;
Takeda et al., 2002; Soldan et al., 2005).
Fig. 14. Structure du génome et stratégie de codage des Bunyaviridae (Soldan et Gonza Lez-
Scarano, 2014)
2.2.2.3. Dissémination virale et réponse immunitaire
L’injection de salive virulente par l’arthropode est suivie d’une réplication à proximité du
point d’inoculation et dans les ganglions lymphatiques régionaux, puis d’une phase de virémie
de quelques jours, jusqu’à l’apparition des premiers anticorps. Cette virémie assure la

Chapitre I Synthèse bibliographique
36
contamination de nouveaux arthropodes et une dissémination systémique du virus vers le
foie, les reins, les glandes endocrines et le système nerveux central. La réponse immune
humorale aboutit rapidement à la production d’anticorps neutralisants qui protègent
durablement contre une infection ultérieure par le même virus. L’immunité, à médiation
cellulaire, est à l’origine de réactions inflammatoires périvasculaires parfois intenses, en
particulier au niveau du système nerveux central, pouvant être à l’origine de lésions (Rodhain,
2001).
2.2.3. Phlébovirus
2.2.3.1. Généralités
Le genre Phlebovirus, un des arbovirus émergeants, s’est progressivement enrichi de virus
apparentés (Fig.15). Il est actuellement composé deplus de 68 membres antigéniquement
distincts dont les virus des fièvres à phlébotomes (transmis par les phlébotomes) et les virus
Uukuniemi (transmis par les tiques) (Nunes-Neto et al., 2017).
Les virus des fièvres à phlébotomes représentent un problème de santé publique majeure dans
les différentes régions du monde et notamment dans le bassin méditerranéen (Izri et al., 2006;
Depaquit et al., 2010).
Sur le plan taxonomique et selon la relation antigénique, 9 espèces, correspondant à des
sérocomplexes, sont reconnues comme appartenant au genre Phlebovirus: virus Sandfly Fever
Naples (SFNV:virus des fièvres à phlébotomes de Naples incluant le virus Toscana),virus
Punta Toro, virus Salehaba, virus Rift Valleyfever (RVFV: virus de la fièvre de la vallée du
Rift), virus Bujaru, virus Candiru, virus Chilibre, virus Frijoles et virus Uukuniemi. Selon le
9ème
rapport du Comité International de Taxonomie des Virus (ICTV), ces virus comprennent
37 sérotypes distincts et 33 sérotypes non classés. Plusieurs nouveaux virus ont été identifiés
mais non encore classés (Plyusnin et al., 2011; Charrel, 2014) (Tableau VI).
Une minorité des phlébovirus connus, incluant le SFNV (dont le virus Toscana), virus Punta
Toro et Sandfly Fever Sicilian (SFSV: virus siciliens des fièvres à phlébotomes qui incluent
lesvirus Corfou, Utique, de Chypre et de Turquie) ont été impliqués dans l’apparition des
fièvres à phlébotomes chez l’Homme (Perrone et al., 2007).
Les virus SFN et SFS existent dans toutes les régions du monde en raison de la grande
répartition géographique de leurs vecteurs. Ces virus ont été isolés de phlébotomes en Asie
centrale, en Amérique, en Europe, en Afrique et notamment en Afrique du Nord (Maroc,
Algérie, Tunisie et Egypte) (Cusi et al., 2010) (Fig.16).

Chapitre I Synthèse bibliographique
37
Fig. 15. Arbre phylogénétique des phlébovirus transmis par les phlébotomes dans l’Ancien
Monde (Neighbour-Joining) (Alkan et al., 2010)

Chapitre I Synthèse bibliographique
38
Tableau VI. Liste des virus du genre Phlebovirus (Charrel, 2014 )
Espèce virale Nom du virus Vecteur
Bujaru virus Bujaru virus
Munguba
ND
Phlébotomes
Candiru virus Candiru virus
Alenquer virus
Itaituba virus
Nique virus
Oriximina virus
Turuna virus
ND
ND
ND
Phlébotomes
Phlébotomes
Phlébotomes
Chilibre virus Chilibre virus
Cacao virus
Phlébotomes
Phlébotomes
Frijoles virus Frijoles virus
Joa virus
Phlébotomes
ND
PuntaToro virus PuntaToro virus
Buenaventura virus
Phlébotomes
Phlébotomes
Rift Valley Fever virus Rift Valley Fever virus
Belterra virus
Icoaraci virus
Moustiques
ND
Phlébotomes
Moustiques
Salehabad virus Salehabad virus
Arbia virus
Adria virus*
Phlébotomes
Phlébotomes
Phlébotomes
Sandfy Fever Naples virus Toscana virus
Sabin virus
Tehran virus
Massilia virus
Granada virus*
Punique virus*
Phlébotomes
Phlébotomes
Phlébotomes
Phlébotomes
Phlébotomes
Phlébotomes
Uukuniemi Uukuniemi virus
EgAN 1825-61 virus
Fin V707 virus
Grand Arbaud virus
Manawa virus
Murre virus
Oceanside virus
Ponteves virus
Precarious Point virus
RML 105355 virus
Saint Abb’s Head virus
Tunis virus
ZalivTerpeniya virus
Tiques
ND
ND
Tiques
Tiques
ND
Tiques
Tiques
Tiques
Tiques
Tiques
ND
Tiques

Chapitre I Synthèse bibliographique
39
Virus non classé Aguacate virus
Ambe virus
Anhanga virus
Arboledas virus
Ariquemes virus
Armero virus
Arumowot virus
Calmito virus
Chagres virus
Corfou virus
Durania virus
Escharte virus
Gabek Forest virus
Gordil virus
Hualyangshan virus
(Severe fever with
thrombocytopenia syndrome
virus [SFTSV])
Itaporanga virus
Ixcanal virus
Jacunda virus
Leticia virus
Mariquita virus
Morolillo virus
Morumbi virus
Mucura virus
Odrenisrou virus
Pacui virus
Rio Grande virus
Salobo virus
Sandfly Fever Sicilian virus
Saint-Floris virus
Serra Norte virus
Tapara virus
Uriurana virus
Urucuri virus
Phlébotomes
Phlébotomes
ND
Phlébotomes
Phlébotomes
Phlébotomes
Moustiques
Phlébotomes
Phlébotomes
Moustiques
Phlébotomes
ND
ND
ND
Tiques
Moustiques
Phlébotomes
ND
Phlébotomes
Phlébotomes
ND
ND
Moustiques
Moustiques
Phlébotomes
ND
ND
Phlébotomes
ND
ND
Phlébotomes
Phlébotomes
ND
ND : Non Défini
* Non officiellement reconnu par l’ICTV, car de découverte récente.

Chapitre I Synthèse bibliographique
40
Fig. 16. Distribution des phlébovirus transmis par les phlébotomes en Afrique (Alkan et al.,
2013)
2.2.3.2.. Découverte des phlébovirus en Algérie
En 1976, des tests de neutralisation dont les résultats se sont révélés négatifs, ont été effectués
parmi la population du sud-est de l’Algérie (Tamanrasset et Djanet) pour la détection du virus
sicilien, SFSV (Tesh et al., 1976).
En 2006, une étude moléculaire et de séroprévalence a été menée en Kabylie, dans la région
de Larbaa Nath Irathen pour la mise en évidence de la circulation du même virus SFS (Izri et
al. 2008). Des anticorps anti-SFSV ont été détectés dans 3 sérums humains sur les 60 testés.
Une séquence d’ARN viral apparentée à celle du virus SFS a été isolée chez un phlébotome
Phlebotomus ariasi sur les 460 constituant l’échantillon examiné. L’analyse phylogénétique a
indiqué que ce virus algérien forme un cluster avec la souche Sabin du SFSV et le virus
sicilien de Chypre (Fig. 17).

Chapitre I Synthèse bibliographique
41
Fig.17. Analyse phylogénique du virus SFS, Algérie, basée sur la séquence 207-nt (A) ou 67
aa(B) du gène de la polymérase (Izri et al., 2008)
Les phlébovirus ont fait l’objet d’une autre étude, du même type que la précédente, dans le
Nord de l’Algérie. Deux tests de séroprévalence ont mis en évidence des IgG spécifiques du
virus SFN ou du virus SFN-like dans des échantillons sériques colligés à Bou Ismail (Alger)
et à Larbaa Nath Irathen avec des taux de 10.6% (11/104 sérums) et 21.6% (13/60 sérums)
respectivement. A partir de l’étude moléculaire de 785 phlébotomes capturés en 2007 dans
ces deux régions et identifiés, trois pools se sont révélés positifs. Deux séquences d’ARN
viral obtenues chez Phlebotomus longicuspis et qui étaient étroitement liées à la séquence de
la souche Poona du SFNV, appartiennent à l’espèce SFNV. Une séquence d’ARN viral isolée
de P.papatasi était étroitement apparentée au virus de Chypre. Ce nouveau virus SFS est
distinct du SFSV détecté dans l’étude précédente (Moureau et al., 2010) (Fig.18 ).
Selon l’analyse phylogénétique, ces séquences présentent un degré d’apparenté étroit avec la
séquence de la souche prototype SFSV de Sabin, formant un cluster (Moreau et al., 2010).

Chapitre I Synthèse bibliographique
42
Fig.18. Analyse phylogénétique de séquences de phlébovirus obtenues à partir de
phlébotomes algériens, basée sur la séquence 67-AA du gène de la polymérase (Moureau et
al. , 2010)
2.2.4. Virus Toscana (Toscana virus ou TOSV)
2.2.4.1. Introduction
TOSV est présent dans plusieurs pays du sud de l’Europe et au Maghreb (Tunisie et Maroc).
Il a été isolé ou détecté chez descas d’infections humaines ou dans des phlébotomes (Alkan et
al., 2013; Bichaud et al., 2013; Gabriel et al.,2010; Kay et al., 2009; Venturi et al.,
2007;Bahrietal., 2010; Es-Sette et al., 2016)
Les transmissions transovarienne et sexuelle de TOSV ont été démontrées du fait de son
isolement à partir de phlébotomes mâles et femelles en Italie, Espagne et France.
Il s’agit d’un sérotype de l’espèce Sandfly Fever Naples qui comprend actuellement les virus
Sabin, Tehran et Karimabad. De nouveaux phlébovirus (virus Granada, virus Punique),
étroitement liés mais distincts de TOSV, ont été découverts et pourraient appartenir à SFNV
mais ne sont pas encore classés par le Comité International de Taxonomie des Virus
(Plyusnin et al., 2011).
TOSV est la troisième cause de méningites estivales, infectant l’Homme avec la prévalence la
plus élevée en Europe et dans le bassin méditerranéen. L’apparition de ces affections est
limitée à la période d’activité du vecteur, durant les mois chauds, d’avril à octobre (Charrel et
al., 2014).

Chapitre I Synthèse bibliographique
43
Le schéma épidémiologique de l’infection au TOSV reste mal connu, le point d’ombre
concerne notamment l’existencedu réservoir mammifère primaire qui n’a pas encore été
identifié. Cependant, la récupération du virus du cerveau d'une chauve-souris capturée dans
une zone d'endémie peut suggérer le rôle possible de cet animal en tant qu’hôte amplificateur
(Verani et al., 1988).
2.2.4.2. Structure et propriétés biologiques
Comme tous les phlébovirus inclus dans la famille des Bunyaviridae, TOSV est un virus
enveloppé, qui apparait sous forme de particule sphérique de 80 à 120 nm de diamètre. Son
génome est constitué de trois segments d'ARN à simple brin de polarité négative ou ambisens,
de 1869 (segment S), 4215 (segment M) et 6404 (Segment L) nucléotides. Chaque segment
est entouré d’une nucléoprotéine (N), formant un complexe hélicoïdal de ribonucléoprotéines.
Ce virus possède :
-deux protéines structurales, les glycoprotéines externes, Gc et Gn (selon leur proximité avec
les terminaisons aminées et carbonées) enchâssées dans la bicouche lipidique de l’enveloppe,
de 5 nm d’épaisseur et codées par le segment M;
- une protéine de nucléocapside (N) codée par le segment S ;
- une protéine non structurale (Ns) exprimée à partir du segment S ;
- un ARN polymérase ARN-dépendante codée par le segment L.
Il se réplique alternativement chez le vecteur et chez le vertébré avec un effet cytopathique
pour les cellules. Le cycle de réplication se déroule dans le cytoplasme de la cellule-hôte
selon les étapes principales suivantes (Charrel, 2014):
- adhésion à la cellule par le biais d’une interaction entre les glycoprotéines de l’enveloppe et
des récepteurs cellulaires non identifiés;
- pénétration par endocytose puis décapsidation par fusion de la membrane virale avec les
membranes endosomales;
- transcription primaire : synthèse des ARNm complémentaires du génome par la polymérase
virale en utilisant des amorces coiffées cellulaires;
- traduction des ARNm correspondant aux trois segments et clivage cotraductionnel du
précurseur des glycoprotéines pour l’obtention des glycoprotéines Gc et Gn;
- synthèse et encapsidation de l’antigénome servant de matrice pour la synthèse de l’ARN
génomique;
- réplication du génome;
- transcription secondaire, synthèse des ARNm et transcription ambisens;

Chapitre I Synthèse bibliographique
44
-morphogénèse : accumulation des glycoprotéines Gn et Gc dans l’appareil de Golgi,
glycosylations et acquisition des membranes virales dans les citernes de l’appareil de Golgi et
bourgeonnement à la surface de la cellule;
- fusion des vésicules cytoplasmiques avec la membrane plasmique et libération des nouveaux
virions.
2.2.4.3. Découverte et pathogénicité de TOSV
TOSV a été isolé pour la première fois chez Phlebotomus perniciosus en Italie centrale en
1971 (Verani et al., 1980). La première preuve de sa pathogénicité chez l'Homme a été
démontrée suite à son implication dans l'infection du système nerveux central (SNC)
diagnostiquée chez des patients suédois et américains à leur retour d’un voyage
respectivement du Portugal et de l’Italie (Calisher et al., 1987; Ehrnst et al., 1985).
Par la suite, l'isolement de ce virus chez une femme atteinte de méningite aseptique a
confirmé qu’il est la cause majeure d'infections du SNV en Italie centrale (Nicoletti et al.,
1991). D'autres souches de TOSV ont été isolées en Italie à partir de P. perfiliewi (Verani et
al., 1988). Une étude rapporte la présence de ce virus chez Sergentomyia minuta (espèce
herpétophile) à partir de spécimens collectés à Marseille (Charrel et al., 2006). Cependant la
pertinence de l’implication du genre Sergentomyia dans le cycle de réplication du virus reste
inconnue.
Suite à sa découverte en Italie centrale, TOSV s'est avéré endémique dans plusieurs autres
régions d'Italie, où il provoque des infections neuro-invasives pendant l'été (Cusi et al., 2010;
Nicoletti et al., 1991; Valassina et al., 2000, 1998, 1996). Les régions endémiques à ce virus
se sont étendues à d’autres pays, dont le Portugal, l’Espagne, la France, la Croatie, la Grèce,
Chypreet la Turquie. Il a été associé aux SFNV et SFSV dans l’apparition des syndromes
fébriles isolés, observés au cours de la saison chaude. A ce jour, il est le seul phlébovirus
transmis par les phlébotomes à être associé aux manifestations du système nerveux central et
l’un des agents viraux les plus souvent impliqués dans les infections neuroméningées chez
l’Homme (Sanchez-Seco et Navarro, 2005; Dionisio et al., 2001; Bahri et al., 2010).
2.2.4.4. Syndromes cliniques
L'infection clinique par TOSV commence par une maladie fébrile légère, après une période
d'incubation de 3 à 7 jours, sans implication du SNC (Charrel et al., 2005). Les infections
neuroinvasives commencent généralement par des maux de tête, de la fièvre, des nausées, des
vomissements et des myalgies. L'examen physique peut révéler une rigidité de la nuque, signe
de Kernig et dans certains cas, une perte de conscience, des tremblements, une parésie et un

Chapitre I Synthèse bibliographique
45
nystagmus. Dans la plupart des cas, le liquide céphalo rachidien (LCR) contient plus de 5 à 10
cellules/ml, avec des taux de glucose et de protéines normaux. Une leucopénie ou une
leucocytose peut être observée.
Le résultat est généralement favorable sans séquelles. D'autres manifestations neurologiques
ont été rapportées telles que l'encéphalite (Dionisio et al., 2001), la méningo-encéphalite
sévère (Baldelli et al., 2004), la surdité (Martinez-Garcia et al., 2008; Pauli et al., 1995), une
perte de conscience durable avec des convulsions (Bartels et al.,2012). Les troubles de la
parole et la parésie peuvent persister pendant des mois après la phase aiguë (Sanbonmatsu-
Gamez et al., 2009).
2.2.4.5. Diagnostic
Le diagnostic spécifique direct repose sur l’isolement du virus sur culture cellulaire, une fois
détecté dans le LCR et dans le sang du patient pendant la phase fébrile. Parmi les techniques
utilisées, la RT-PCR en temps réel donne rapidement un résultat confirmant l’infection.
Plusieurs techniques sérologiques peuvent être utilisées pour un diagnostic spécifique indirect,
inhibition de l’hémagglutination, fixation du complément ou séroneutralisation,
immunofluorescence indirecte et ELISA.

CHAPITRE II
MATERIEL ET METHODES

Chapitre II Matériel et Méthodes
46
1. Caractéristiques de la zone d’étude
Situé au nord-est d’Algérie et à 110 km au sud-est d’Alger, Draa El Mizan est chef-lieu de
daïra qui relève de la wilaya de Tizi-Ouzou et regroupe 4 communes, Draa El Mizan, Aïn-
Zaouïa, Frika et Aït-Yahia Moussa (Fig.19). Celles-ci s’étendent sur une superficie de 80.84
km2
pour une population de 38.844 habitants (RGPH de 2008) qui se répartissent à raison de
51% dans le chef-lieu de commune, 39% dans 9 agglomérations secondaires et le reste dans
des zones éparses à 650 mètres d’altitude, 36°32'8"N, 3°50'3"E. La région de Draa El Mizan
est enclavée dans une cuvette entourée de collines, au pied ouest de la chaîne de montagne du
Djurdjura (Fig.20). Elle bénéficie d’un climat méditerranéen, tempéré, humide et riche en eau
grâce aux cours d’eau et à l’existence d’un barrage (classification de Koppen : Csa).
La pluviométrie annuelle est de 600 à 800mm; l’étage bioclimatique est subhumide, à hiver
doux, fréquemment entre 0 et 3°C. Le mois le plus froid de l’année se situe entre décembre et
janvier pour la température minimale et le plus chaud entre juillet et août avec une
température maximale allant de 36 à 40°C.
Fig.19.Situation géographique de Draa El Mizan (Données cartographiques, Google, 2018)
Le relief et la géologie sont constitués du sud au sud-ouest de Draa El Mizan, essentiellement
des massifs de Moula Yahia à Béni Khalfoun. Le relief est formé surtout de conglomérats de
gré jaunâtre friable. Au nord-ouest, la chaine Flissa M’Kira et les massifs montagneux de Sidi

Chapitre II Matériel et Méthodes
47
Ali Bounab sont composés de micaschistes, de gneiss des granulites et de grés et de terrains
cristallophylliens issus de formations schisteuses.
Fig.20. Commune de Draa El Mizan (Photo personnelle)
La formation végétale est celle des moyennes altitudes (400 à 800 m), caractérisant les forêts
typiques de chêne liège, un cortège floristique: arbousier (Arbutus unedo), filaire à large
feuille (Phillyrea latifolia), bruyère arborescente (Erica arborea), cistes à trois feuillets
(Cistus monspeliensis), cistes velus (Cistus villosus), daphné garou (Daphne gnidium),
lavande stéchade (Lavandula stoechas), chèvrefeuille (Lonicera implexa), myrte commune
(Myrtus communis), aubépine blanche (Crataegus oxyacanthoides), genévrier oxycèdre
(Juniperus oxycedrus), olivier sauvage (Oléastre), inule visqueuse (Inula viscosa) et lentisque
(Pistacia lentiscus).
2. Etude entomologique
2.1. Méthodes d’échantillonnage des phlébotomes adultes
Les phlébotomes ayant fait l’objet de cette étude ont été capturés dans la région de Draa El
Mizan (36° 320'146’’ N, 3° 500’850’’ E) à une altitude moyenne de 380 m, dans des sites
sélectionnés en fonction des cas de leishmaniose enregistrés (données épidémiologiques
fournies par les services de santé) et de la présence d’animaux domestiques, principalement
d’animaux d’élevage (Fig.21).

Chapitre II Matériel et Méthodes
48
Fig.21. Exemples de sites de capture des phlébotomes
Deux méthodes d’échantillonnage ont été utilisées, la capture au piège adhésif et la capture au
piège lumineux.
- Piège adhésif : connu depuis les importants travaux des épidémiologistes russes, Vlazov
(1932) et Petrischeva (1935), ce type de piège consiste en un support papier (blanc mat de
80g/m² découpé en carrés de 20cm de côté) imprégné d’huile de ricin et sur lequel viennent se
poser les insectes (Rioux et al., 1967; Croset et al., 1974).
De 2012 à 2013, des captures mensuelles ont été réalisées entre les mois de mai et septembre.
Les pièges ont été tendus en fin de journée à l’aide d’un support métallique, selon le dispositif
mis au point par Izri (1998) (Fig.22) et placés dans différents biotopes, tels les gîtes de repos
et lieux de ponte des phlébotomes (anfractuosités de murs, fenêtres ou autres ouvertures
d’habitations) et dans les abris d’animaux domestiques. Ils ont été récupérés le lendemain,
regroupés selon les lieux de capture et mis dans des sacs en plastique numérotés.
Fig.22. Lieux d’emplacement des pièges adhésifs

Chapitre II Matériel et Méthodes
49
-Piège lumineux: de type CDC (Center for Disease Control): consiste en un moteur surmonté
d’une lampe et alimenté par une batterie pour assurer le fonctionnement d’un ventilateur
(destiné à entretenir une aspiration continue de faible intensité). Les phlébotomes, attirés par
la lumière, sont aspirés par le ventilateur et piégés dans une nasse. Les pièges utilisés sont des
«Miniature Light Traps CDC (John WHock Company, Gaines ville, FL)» modifiés, la nasse
étant remplacée par une cage moustiquaire à maille très fine et insérée dans un cadre rigide,
cubique, de 20 cm de côté. Ce dispositif utilise une lumière jaune fournie par une ampoule à
incandescence de 0.3A et un petit aspirateur alimenté par une batterie de 6 volts.
Les pièges ont été déposés durant cinq nuits consécutives du mois d’août 2013, dans les abris
d’animaux domestiques, à 1ou 2 mètres au-dessus du sol et dans des coins ombragés comme
décrits par Rioux et al. (1986) et Izri et al. (1990) (Fig.23). Ils restent fonctionnels toute la
nuit pour être récupérés tôt le matin. Afin d’éviter toute fuite des phlébotomes, le manchon
des pièges est noué à son extrémité à l’aide d’un cordon juste avant le débranchement de
l’appareil. Chaque cage a été placée dans un sac en plastique noir dont l’atmosphère est
humidifiée par un morceau de coton imbibé d’eau pour empêcher la dessiccation des insectes.
Fig.23.Pièges lumineux CDC accrochés
Les différents pièges ont été acheminés vers le Laboratoire National Vétérinaire de Draa El
Mizan où s’est effectué le prélèvement des phlébotomes. Ces derniers ont été décollés des
pièges adhésifs, délicatement à l’aide d’un pinceau fin et plus facilement en déposant dessus
une goutte d’éthanol à 70% qui dissout l’huile. Ceux capturés par pièges lumineux ont été
récupérés au moyen d’un aspirateur d’entomologie (Fig.24) et anesthésiés en utilisant une
compresse imprégnée de chloroforme placée quelques secondes à l’ouverture de l’aspirateur.
Les spécimens récoltés morts dans les deux types de pièges ont été conservés dans des tubes
en verre à fermeture hermétique, contenant de l’éthanol à 70%, en vue de leur identification.
Les échantillons de phlébotomes obtenus vivants par pièges lumineux, ont été divisés en deux
lots, l’un examiné à frais pour une étude taxonomique et parasitologique et l’autre destiné à

Chapitre II Matériel et Méthodes
50
une étude moléculaire. Ce dernier a été organisé en 247 pools de 20 à 30 individus et
conservés par congélation à -80° C dans des cryotubes de 2 ml.
Fig.24.Capture des phlébotomes aux pièges lumineux CDC
A. Phlébotomes piégés, B. phlébotomes aspirés , C. phlébotomes collectés
La combinaison de ces deux méthodes de piégeages permet de maximaliser la possibilité de
capturer les différentes espèces de phlébotomes présentes dans la région d’étude.
Après échantillonnage, une approche écologique a été abordée avec l’analyse de la
composition de la population phlébotomienne étudiée, à l’aide de l’abondance relative,
comme indice écologique.
L’abondance relative (Ar) ou fréquence centésimale (Fc) d’une espèce donnée correspond au
pourcentage d'individus de cette espèce (ni) par rapport au total des individus recensés (N)
d’un peuplement (Dajoz, 1985).
ni: nombre d’individus d’une espèce i.
N: nombre total d’individus toutes espèces confondues
2.2. Identification des phlébotomes
Elle a été réalisée selon les clés d’Abonnenc (1972), de Léger et al., (1983) et de Killick-
Kendrick (1991), basées sur des critères morphologiques spécifiques après dissection des
individus.

Chapitre II Matériel et Méthodes
51
Les structures utilisées sont les spermathèques chez les femelles; le génitalia chez les mâles et
l’armature pharyngienne et/ou cibariale chez les phlébotomes des deux sexes.
2.2.1. Identification à frais
Les phlébotomes gardés vivants ont été disséqués sous la loupe binoculaire: chaque individu
est déposé sur une lame porte-objet dans une goutte d’eau physiologique stérile, disséqué à
l’aide de deux aiguilles stériles et jetables, puis selon le sexe :
- la spermathèque est extraite puis observée entre lame et lamelle au microscope optique;
- l’extrémité abdominale est sectionnée au niveau du 7ème
segment et placée entre lame et
lamelle pour l’examen au microscope optique des armatures génitales, valves péniennes,
coxites et styles.
En cas de difficulté de détermination, la tête de l’insecte est examinée, placée face ventrale
vers le haut pour une meilleure visualisation de l’armature pharyngienne et/ou cibariale.
Quelques spécimens ont été dessinés à l’aide d’un microscope optique muni d’un tube à
dessin.
2.2.2. Identification après éclaircissement et montage
Les phlébotomes conservés dans l’éthanol à 70% ont été transvasés dans un verre de montre
une fois l’alcool aspiré à l’aide d’une pipette Pasteur, puis immergés pendant 24 heures dans 6
ml de solution Marc-André (Abonnenc, 1972) pour éclaircissement. Chaque phlébotome est
ensuite prélevé à l’aide d’un pinceau fin, déposé sur une lame porte-objet dans une goutte de
Poly Vinyl Alcool (PVA) et disséqué pour effectuer les mêmes observations microscopiques
que celles décrites ci-dessus.
3. Etude parasitologique
3.1. Recherche des formes promastigotes chez le phlébotome
Un examen parasitologique direct a été réalisé pour la recherche de leishmanies dans
l’intestin de phlébotomes femelles capturés vivants. L’insecte est disposé sur une lame stérile
dans une grosse goutte d’eau physiologique stérile et une fois la tête coupée, il est disposé en
position latérale pour pratiquer deux incisions, une ventrale et l’autre dorsale entre les 7ème
et
8ème
segments abdominaux. La partie postérieure est bloquée à l’aide d’une aiguille montée
stérile et la partie antérieure retirée pour faire apparaitre progressivement l’intestin (Fig.25).
La tête et la partie postérieure de l’abdomen sont transférées dans une goutte de Marc-André
(déposée sur la même lame) et observées au microscope optique.
Avant de procéder à leur dissection, les phlébotomes ont été disposés sur un lit de glace pour
prolonger leur engourdissement et maintenir vivantes d’éventuelles formes promastigotes.

Chapitre II Matériel et Méthodes
52
Fig.25.Dissection de phlébotome femelle pour la détection de promastigotes
(D’après El-Hossary, 2006)
A. Enlèvements des pattes, B. Enlèvement de la tête, C. incision de l’abdomen
3.2. Détection de cas humains de leishmaniose viscérale par Western blot
Le Western blot (WB) est un test sérologique sensible et très spécifique. Il permet de détecter
de faibles taux d’anticorps et de distinguer les sujets malades des porteurs asymptomatiques.
Dans le but de détecter des cas de leishmaniose viscérale parmi la population de Draa El
Mizan, 370 échantillons sériques ont été testés par le WB, IgG : LES-WB24G. Ces sérums
ont été collectés à partir de prélèvements sanguins pratiqués chez des personnes d’âge et de
sexe différents, apparemment saines et choisies aléatoirement. Avant la réalisation du test,
chaque sérum est réparti en deux aliquotes.
Les sérums sont incubés séparément avec une membrane de nitrocellulose sensibilisée par les
protéines antigéniques. Les couples antigène-anticorps sont révélés par l’adjonction d’anti-
immunoglobuline marqué à la phosphatase alcaline. La révélation du ligand immuno-
enzymatique se fait par l’addition du substrat spécifique de l’enzyme. Les antigènes reconnus
par les anticorps anti-Leishmania sont révélés sous forme de bandes transversales violettes.
Il s’agit des bandes 14KDa et/ou 16KDa spécifiques à la leishmaniose, bien visibles en bas de
la bandelette.
Protocole :
- établir un plan de distribution des échantillons;
- distribuer 1.2 ml du Tampon de l’échantillon dans chacun des puits ;
- agiter doucement la cuve;
- placer la bandelette de nitrocellulose, face vers le haut, dans chacun des puits (Fig.26);
- attendre 1mn puis agiter doucement la cuve pour immerger la bandelette;
- distribuer 25µl de chaque sérum et 25µl du contrôle positif;
- agiter doucement la cuve après chaque dépôt;
- incuber 90 mn sur agitateur;
- laver 3 fois avec la solution de lavage diluée au 1/10;

Chapitre II Matériel et Méthodes
53
- distribuer 1.2 ml du conjugué anti IgG;
- agiter doucement la cuve;
- incuber 60 mn sur agitateur ;
- laver 3 fois avec la solution de lavage diluée au 1/10;
- distribuer 1.2 ml du substrat;
- agiter doucement la cuve;
- incuber 60 mn sur agitateur;
- arrêter plus tôt les bandelettes dont la coloration devient très forte ;
- arrêter la réaction par 2 lavages à l’eau distillée;
- transférer les bandelettes sur un papier filtre;
- laisser sécher à l’air 15 mn;
- comparer le profil de l’immunoblot de chaque échantillon avec celui du contrôle positif.
Fig.26. Incubation des sérums avec les bandelettes du Western blot
Analyse statistique
L'étude de la variation de la positivité du test sérologique (Western blot) entre les différentes
classes de taille a été réalisée par le test Khi 2.
L'analyse statistique a été réalisée à l'aide du logiciel STATISTICA (Version 10.0.228.8).
4. Recherche et caractérisation moléculaire de phlébovirus
Cette étude a été menée sur les 247 pools de phlébotomes conservés par congélation et les
techniques utilisées ont été réalisées au laboratoire des Virus Emergents, UMR 190, Faculté
de Médecine, La Timone (Marseille).
4.1. Broyage et homogénéisation des pools de phlébotomes
Les pools ont été placés dans des cryotubes renfermant chacun une bille de tungstène de 3mm
et 600µl d’Eagle Minimal Essential Medium (EMEM) (Charrel et al., 2009; Alkan et al.,
2015).

Chapitre II Matériel et Méthodes
54
Les tubes sont ensuite placés dans un broyeur « Tissu Lyser » pendant 3 à 5 minutes à une
fréquence de 30 tours/min selon leur homogénéisation. L’homogénat est centrifugé à 5800
tours/min et seul le surnageant est récupéré et réparti en aliquotes.
4.2. Extraction de l’ARN
L’extraction d’ARN est réalisée avec le robot virol RNA & DNA isolation (Machenery Nagel)
qui prend à la fois, 2 plaques de 96 échantillons chacune et complétée manuellement avec le
Kit Quiagen EZ1® pour les 55 échantillons restants.
.4.2.1. Extraction par le robot (Well Bloch, Eppendorf epMotion 5075)
- Prendre 140 µl du surnageant des 192 pools de phlébotomes avec une pipette multicanaux et
les disposer dans une plaque;
- rajouter 560 µl du tampon RAV 1 préalablement préparé;
- placer la plaque des échantillons dans le robot selon le plan indiqué par le logiciel.
4.2.2. Extraction manuelle à partir du Kit Quiagen EZ1®. Virus MiniKit V2.0
- Prendre 140 µl du surnageant des 55 échantillons avec une pipette multicanaux et les
disposer dans une plaque, ajouter ensuite 560 µl du tampon (RAV 1) préalablement préparé ;
- laisser incuber à température ambiante pendant 5 minutes, ajouter 400 µl d’éthanol (grade
HPLC);
- mélanger puis transférer dans les colonnes;
- centrifuger à 5600 tours/min le mélange pendant 4 minutes puis vider les colonnes une à une
et les remplacer par de nouvelles;
- faire le 1er
lavage du contenu avec 500 µl du tampon RAV1 et centrifuger à 5600 tours/min
pendant 4 minutes ; vider les colonnes pour la 2ème
fois et les changer par d’autres;
- faire le 2ème
lavage avec 700 µl du tampon RAV3 et centrifuger à 5600 tours/min pendant 4
minutes et remplacer les colonnes;
- le 3ème
lavage est effectué avec 700 µl du tampon RAV3, centrifuger à 5600 tours/min
pendant 15 minutes et remplacer les colonnes par des tubes Eppendorf;
- faire une élution (Round-Wall block low) en rajoutant 90 µl de H2O dans chaque échantillon;
- laisser incuber les échantillons à température ambiante pendant 5 minutes ensuite centrifuger
à 5600 tours/min pendant 4 minutes.

Chapitre II Matériel et Méthodes
55
4.3. Amplification génique
La PCR est une technique d’amplification d’ADN in-vitro qui permet d’obtenir un très grand
nombre de copies d’une séquence d’ADN cible. Chaque cycle de PCR est constitué de trois
étapes :
- une dénaturation de l'ADN par chauffage pour séparer les deux brins qui le composent,
- une hybridation des amorces aux extrémités de la séquence recherchée,
- une élongation grâce à l'action d'une ADN polymérase.
Ce cycle est répété un grand nombre de fois pour obtenir une multiplication exponentielle de
la séquence d'ADN cible. Il existe différentes variantes de la PCR. Les techniques de RT-PCR
et de PCR nichée ont été retenues dans cette étude.
La RT-PCR (Reverse transcriptase PCR) est une technique qui associe une transcription
inverse suivie d’une PCR standard. Elle permet de synthétiser le brin complémentaire d’un
ARN en utilisant une ADN polymérase ARN dépendante.
Pour ce qui du principe de la PCR nichée, l’amplicon obtenu après une première PCR est
amplifié à l’aide d’un second couple d’amorces qui s’hybride à une partie interne de la
séquence amplifiée (gain en sensibilité et spécificité). Cette technique est largement utilisée
pour la recherche et la détection des virus.
Les réactions d’amplification des gènes de phlébovirus sont réalisées avec des couples
d’amorces spécifiques (Eurogentec) dans un thermocycleur de type 2720 Thermal Cycler AB
Applied Biosystems en présence de l’enzyme polymérase en respectant les conditions
d’utilisation spécifiées par le fournisseur. Les amplicons sont ensuite purifiés et séquencés.
Les séquences sont comparées à celles déposées dans une banque de données
(www.ncbi.nlm.nih.gov).
4.3.1. Amorces
Plusieurs amorces ciblant des gènes différents sont utilisées dans les réactions indépendantes
de RT-PCR et de PCR nichée. Les amorces consensus SFN des phlébovirus (virus Sandfly
Fever Naples) ciblent les nucléoprotéines situées dans le segment S de l’ARN virale (Charrel
et al., 2007) et les amorces spécifiques N-Phlebo ciblent le gène de la polymérase qui est
localisé dans le segment L de l’ARN (Sanchez-Seco et al., 2003) (Tableaux VII et VIII

Chapitre II Matériel et Méthodes
56
Tableau VII : Séquences des amorces pour la RT-PCR
Gènes cibles Amorces Séquences Concentration TM Taille de
l’amplicon (pb)
Références
Polymérase,
Segment L
N-Phlebo1+ ATGGARGGITTTGTIWSICIICC 0.4 µM 48°C 553
(Sanchez-Seco et
al., 2003) N-Phlebo1- AARTTRCTIGWIGCYTTIARIGTIGC 0.4 µM
nucléoprotéines
Segment S
SFNV-NP-S1 CTTYTTRTCYTCYCTRGTGAAGAA 0.8 µM 48°C 437 (Charrel et al.,
2007) SFNV-NP-R1 ATGATGAAGAARATGTCAGAGAA 0.8 µM
Tableau VIII : Séquences des amorces pour la PCR nichée
Gènes cibles Amorces Séquences Concentration TM Taille de
l’amplicon (pb)
Références
Polymérase,
Segment L
N-Phlebo2+ WTICCIAAICCIYMSAARATG 0.4 µM 48°C 244 (Sanchez-Seco et
al., 2003) N-Phlebo2- TCYTCYTTRTTYTTRARRTARCC 0.4 µM
Nucléoprotéines,
Segment S
SFNV-NP-R2 CCTGGCAGRGACACYATCAC 0.8 µM 48°C 304 (Charrel et al.,
2007) SFNV-NP-S2 GCRGCCATRTTKGGYTTTTCAAA 0.8 µM

Chapitre II Matériel et Méthodes
57
4.3.2. Préparation des mix
Le mix est manipulé dans une pièce spéciale sous PSM avec le port de gants pour éviter toute
contamination, il est suite distribué dans des tubes Eppendorf numérotés.
La RT-PCR est réalisée avec le kit-Access RT-PCR (Promega, Charbonnières-Les Bains,
France), selon les recommandations du fabricant. Le volume final utilisé est de 50 µl dont 5
µl d’ARN (Tableau IX).
Tableau IX. Mix de la RT-PCR
La PCR nichée est réalisée avec une Taq polymérase standard (Promega, Charbonnières-Les
Bains, France) selon les recommandations du fabricant. Le volume final utilisé est de 50 µl,
47 µldu mix et 3 µl du produit de la RT-PCR. (Tableau X).
Tableau X. Mix de la PCR nichée
Composants Volumes pour un échantillon
Tampon 10X de la Taq polymérase 5 µl
dNTP 1 µl
Amorce Forward 2.5 µl
Amorce Reverse 2.5 µl
MgCl2 1.5 µl
Taq polymérase 0.25 µl
H2O 34.25 µl
Produit de RT-PCR 3 µl
Composants Volumes pour un échantillon
Tampon AMV 5X 10 µl
dNTP 1 µl
Amorce Forward 2 µl
Amorce Reverse 2 µl
MgSO4 2 µl
TFi DNA polymérase 1 µl
AMV Reverse Transcriptase 1µl
H2O 26 µl
ARN à analyser 5 µl

Chapitre II Matériel et Méthodes
58
Chaque constituant de ces mix est multiplié par le nombre d’échantillons étudiés, puis 45µl et
47 µl de ces mélanges sont distribués dans des barrettes pour la RT-PCR et la PCR nichée,
respectivement :
- Témoin négatif : 5 µl d’eau distillé stérile plus 45µl de mix pour la RT-PCR et 3µl d’eau
distillé stérile plus 47 µl de mix pour la PCR nichée;
- échantillons : 5 µl de notre échantillon (ARN) est ajouté à 45µl de mix pour la RT-PCR et
3µl du produit de la RT-PCR est ajouté à 47µl de mix pour la PCR nichée;
- volume total : 50 µl par échantillon pour le témoin négatif et les échantillons à analyser.
Le mix et les échantillons sont transférés dans des barrettes et sont introduits dans le
thermocycleur.
4.3.3. Programmation du thermocycleur
Cette réaction est cyclique et se réalise en trois étapes :
- dénaturation : consiste à chauffer avec une température élevée pour séparer l’ADN à double
brin en brisant les liaisons hydrogènes ;
- hybridation : une fois l’ADN séparé, on refroidit rapidement pour que les amorces se fixent
à chaque extrémité complémentaire de l’ADN à simple brin;
- polymérisation ou élongation: après la fixation des amorces, l’ajout de la Taq polymérase et
de nucléotide permet la synthèse des nouveaux brins d’ADN.
- Nouvelle amplification.
Une exception est faite pour la RT-PCR, il s’agit d’une étape de rétrotranscription avant le
début du cycle d’amplification afin de transformer l’ARN en ADN.
Les réactions d’amplification sont réalisées dans le thermocycleur selon les conditions
suivantes (Tableaux XI et XII).
Tableau XI. Cycle de la RT-PCR
Température Temps Nombre de cycle
Rétrotranscription 48°c 45 minutes 1 cycle
Dénaturation initiale 94°c 2 minutes 1 cycle
Dénaturation 94°c 30 secondes 40 cycles
Hybridation 48°c 45 secondes 40 cycles
Elongation 68°c 2 minutes / 7 minutes
30*
40 cycles
Extension 20°c 2 minutes 1 cycle
* 2 minutes correspondent au temps d’élongation en utilisant l’amorce N-Phlebo alors que les 7
minutes 30 correspondent au temps d’élongation en utilisant l’amorce SFN.

Chapitre II Matériel et Méthodes
59
Tableau XII. Cycle de la PCR nichée
Température Temps Nombre de cycle
Dénaturation initiale 94°c 5 minutes 1 cycle
Dénaturation 94°c 30 secondes 40 cycles
Hybridation 48°c 30 secondes 40 cycles
Elongation 72°c 2 minutes / 7
minutes 30*
40 cycles
Extension 20°c 2 minutes 1 cycle
* 2 minutes correspondent au temps d’élongation en utilisant l’amorce N-Phlebo alors que les 7
minutes 30 correspondent au temps d’élongation en utilisant l’amorce SFN.
4.4. Lecture des résultats de PCR par électrophorèse sur gel d’agarose
Les fragments d’ADN obtenus (ou amplicons) ont été visualisés par une électrophorèse sur
gel d’agarose.
Cette technique, sensible, rapide et simple à mettre en œuvre, est utilisée soit à des fins
analytiques (pour séparer des fragments d'ADN et pour déterminer leur taille), soit pour
obtenir un fragment d'ADN de taille connue en vue de sa purification.
La taille des fragments d’ADN extraits est comprise entre 0,2 et 50 kb. Ces derniers sont
facilement détectés sur le gel d’agarose grâce à un colorant fluorescent, le BET (Bromure
d'éthidium). Des quantités très faibles d'ADN peuvent être ainsi visualisées en lumière UV (de
l'ordre de 5-10 ng).
Préparation du gel d’agarose
- Diluer 4 g de poudre d’agarose dans 200 ml de tampon (Tris, Borate, EDTA) (90 mM Tris-
borate, 2mM EDTA pH 8,2) afin d’obtenir un gel concentré à 2% ;
- chauffer le mélange au micro-onde environ 2 minutes pour dissoudre l’agarose et obtenir un
liquide clair et limpide ;
- transvaser le gel dans un flacon approprié au BET (Bromure d’éthidium) ;
- refroidir le gel jusqu’à environ une T° entre 40-45°C;
- ajouter du BET à 0.5 µg/ ml;
- agiter et verser le gel dans le support déjà préparé avec des peignes pour former les puits de
dépôt des échantillons;
- placer le gel dans la cuve d’électrophorèse en ajoutant le tampon TBE 0.5X dans la cuve
préalablement remplie jusqu’à ce que le gel soit immergé (environ 1mm au-dessus);
- déposer au milieu des puits 4µl du marqueur de poids moléculaire (100 pb DNA ladder,
Invitrogen);

Chapitre II Matériel et Méthodes
60
- mélanger extemporanément 5 µl de tampon de charge (Bleu de Bromo-phénol 2X) avec
10µl de chaque échantillon d’ADN à tester afin d’assurer le maintien du dépôt en immersion
dans le puits et permettre la visualisation de la migration ;
- déposer les échantillons dans chaque puits en réservant un puits pour le témoin négatif ;
- régler l’électrophorèse à 120V pendant 30 minutes.
Visualisation par transilluminateur
La visualisation des bandes d’ADN du gel se fait sur la table UV du transilluminateur dans
une chambre noire grâce aux molécules du BET qui s'intercale entre les paires de base d’ADN
où sa fluorescence est exaltée. En fonction de la taille des fragments d’ADN étudiés, on
repèrera la bande et sa taille respectivement par rapport au marqueur de poids moléculaire.
4.5. Purification des produits de PCR
- Prélever les bandes d'intérêt;
- découper la bande désirée à l'aide d'un scalpel et transférer celle-ci dans un tube Eppendorf ;
- tarer le tube Eppendorf et peser les fragments d'agarose contenant l’ADN à purifier.
Les échantillons sont purifiés sur colonne (Amicon Ultra Centrifucal Filters, Millipore) selon
les recommandations du fournisseur :
- Insérer les dispositifs Amicon Ultra-0.5 dans les tubes Eppendorf ;
- ajouter jusqu'à 500 μl d'échantillon au dispositif de filtration Amicon Ultra 0.5 et le boucher
(en fonction du poids pesé);
- centrifuger à 14 000 tours /min pendant environ 10-30 minutes ;
- retirer le dispositif assemblé de la centrifugeuse et séparer le filtre Amicon Ultra 0.5 du tube
Eppendorf ;
- pour récupérer le soluté concentré, placer le filtre Amicon Ultra 0.5 à l'envers dans un tube
Eppendorf propre ;
- placer dans la centrifugeuse, en alignant le bouchon ouvert vers le centre du rotor ;
- centrifuger pendant 2 minutes à 1000 tours /min pour transférer l'échantillon concentré du
filtre Amicon Ultra 0.5 vers le tube Eppendorf où l'ADN peut être stocké.
4.6. Séquençage
Le séquençage (selon Sanger et al., 1977) des échantillons positifs, dans les deux directions,
est fait en utilisant l’ABI Prism®
BigDye®Terminator v1.1 Cycle Sequencing Kit (Applied
Biosystems, USA) selon les indications du fournisseur.

Chapitre II Matériel et Méthodes
61
4.6.1. Préparation de la réaction du séquençage
La manipulation du mix pour la réaction du séquençage se fait comme pour une PCR standard
dans une cuvette contenant de la glace pilée. Il est distribué dans des tubes Eppendorf
numérotés à raison de 2 cupules pour chaque échantillon contenant chacun une amorce
(Forward et reverse) (Tableaux XI et XII).
Tableau XIII. Produits du mix du séquençage
Produits Réaction pour un échantillon
BigDye® Terminator v1.1 3μl
SFNV-NP* Forward (S2 ) ou Reverse ( R2 ) 0,5μl
Eau distillée Ultra-pure stérile 4μl
ADN purifié 3μl
Volume total du mix 10,5μl
(*) Pour chaque échantillon, deux cupules contenant 7µl du mix plus 0,5µl de l’amorce sens ou 0,5µl
de l’amorce antisens.
Tableau XIV. Programme du thermocycleur
96°C
96°C
50°C
60°C
4°C
2 min
10 sec
5 sec
4 min
∞
4.6.2. Purification au Sephadex®
- Préparer une plaque de filtration de 96 puits de marque millipore A1=Cup1 ;
- remplir les puits de la plaque avec la poudre Sephadex à l’aide d’un moule spécial ;
- ajouter 300µl d’eau ultra-pure dans chaque puits; la plaque Sephadex est prête après 3h ;
- centrifuger à 1200 tours pendant 2 minutes à 4°C en mettant une plaque collective (poubelle)
au-dessous de la plaque de purification pour récupérer les 300µl d’eau; le Sephadex forme un
gel pour la purification des produits de PCR ;
- ajouter 10μl de l’eau distillée dans chaque cupule contenant l’ADN (produits du BigDye),
prendre une plaque de récupération propre en respectant l’ajustement des lettres et numéros
avec la plaque Sephadex ;
- transférer les 20µl dans la plaque du Sephadex et faire une deuxième centrifugation à 1200
tours pendant 2 minutes à 4°C;
25 cycles

Chapitre II Matériel et Méthodes
62
- récupérer l’ADN purifié au Sephadex dans la plaque de récupération et le déposer sur une
autre plaque appropriée au séquenceur.
5. Lecture des résultats
Le séquençage de l’ADN est réalisé à l’aide de l’appareil ABI 3730 automate de séquenceur
(Applied Biosystems, Foster City, California, United States). Les résultats obtenus sont
corrigés grâce au logiciel Codon Code Aligner 3.7.1.1 sequencing analysis software.
Les séquences obtenues sont analysées à l'aide du Blast N dans la base de données NCBI
(www.ncbi.nlm.nih.gov).
6. Mise en culture des phlébovirus
Afin d’isoler le virus, des cellules Vero (ATCC CCL81) (une lignée isolée en 1962 à partir
de cellules épithéliales de rein de singe vert d’Afrique) ont été inoculées par le surnageant du
pool positif de phlébotomes. La détection du virus est fondée sur la recherche d’un éventuel
effet cytopathogène sur les cellules.
6.1. Préparation des cellules
Préparer 3 flasques de 25 cm2 de cellules Vero, 2 ou 3 jours avant la date d’inoculation. Les
cellules doivent être à 70% de confluence le jour de l’infection.
6.2. Trypsinisation des cellules
La trypsinisation (ou trypsination qui permet le détachement des cellules confluentes adhérant
à la surface interne des flasques) est pratiquée pour maintenir les cultures cellulaires. Cette
opération se réalise en plusieurs étapes :
- laver le tapis cellulaire avec 10 ml de tampon HBSS déposé sur la face opposée au tapis
cellulaire et ramené progressivement sur celui-ci;
- éliminer le produit de lavage en vidant la plaque dans un récipient ou à l’aide d’une pipette ;
- ajouter 4 ml de trypsine dans la flasque toujours dans le sens opposé de la nappe cellulaire;
- vider la flasque partiellement en conservant environ 1/3 de son contenu (environ 1-2ml);
- incuber la flasque à 37°C pendant 10 minutes (jusqu’à ce que les cellules soient bien
détachées et dissociées);
- ajouter 8 ml de milieu EMEM et de sérum de veau fœtal (SVF) à la culture afin d’arrêter
l’effet de la trypsine ;
- dissocier le reste des cellules par aspiration/refoulements puis transférer dans un tube de
centrifugation de 50 ml ;
- centrifuger 5mn à 1500 tours/mn ;

Chapitre II Matériel et Méthodes
63
- vider le surnageant en faisant attention au culot ;
- suspendre avec du milieu (5 ml pour un flasque de 25 cm2) en homogénéisant ;
- réaliser un comptage de cellules (à l’aide d’un automate ou d’un protocole manuel) ;
- déposer une suspension cellulaire nécessaire pour l’obtention de 2.105 cellules/ml et ajouter
le volume d’EMEM et de SVF nécessaire.
6.3. Inoculation du virus
Les surnageants des pools de phlébotomes sont centrifugés à 13200 tours/min pendant 1
minute. La mise en place dans les flasques des cellules Vero a été réalisée sous hotte à flux
laminaire.
- distribution dans chaque flasque de 250 µl de la suspension cellulaire précédemment diluée ;
- addition de 200 µl du milieu SVF à 0% et 50 µl du surnageant dans chaque tube;
- inoculation de la suspension cellulaire de chaque tube avec 50 µl de surnageant de broyat de
phlébotomes;
- préparation d’un tube témoin dans lequel sont ajoutés 50 µl du milieu SFV à 0% et la
suspension cellulaire;
- incubation à température ambiante pendant 1 heure en agitant les tubes toutes les 5 minutes
pendant 10 minutes pour éviter que les cellules ne se déposent;
- addition de 2,5 ml de milieu SVF à 5% dans chaque tube ;
- incubation des tubes infectés à 37° C dans une ambiance de 5% de CO2;
- examen, dès le lendemain et jusqu’au 7ème
jour (7ème
passage) des cellules sous microscope
optique inversé pour rechercher un effet cytopathogène.
Cet effet est dit positif si une modification morphologique des cellules est notée avec
agrégation en amas et augmentation de la réfringence.
7. Séquençage du génome complet
La caractérisation complète du génome viral est réalisée par la technique de séquençage de
nouvelle génération (Next-Generation-Sequencing, NGS) à partir du passage 3 de la culture
Vero. Cette nouvelle technique fonctionne en 3 étapes. La première étape consiste en la
préparation et l'amplification des molécules d'ADN à analyser. La seconde étape permet
l'incorporation des bases complémentaires du brin à séquencer et la troisième comprend la
lecture de la séquence proprement dite.
Pour ce faire, 140 µl du surnageant de la culture cellulaire sont incubés à 37°C pendant 7 h
avec 30 U de Benzonase (Novagen, n° de catalogue 70664-3); puis l'ARN est extrait en
utilisant le minikit d’ARN Viral (Qiagen) sur le Bio Robot EZ1 Advanced (Qiagen). Des

Chapitre II Matériel et Méthodes
64
amorces aléatoires marquées spécifiquement pour une amplification aléatoire pour la RT-PCR
et la PCR standard sont utilisées. Les produits de PCR sont purifiés (Amicon ultracentrifugal
filtres, Millipore) puis 200 ng sont utilisés pour le séquençage du génome en utilisant le
séquenceur à ion PGM (Life Technologies SAS, Saint Aubin, France).
Les séquences brutes du virus obtenues ont été reconstruites à partir des « NGS reads » basées
sur les résultats du meilleur BLAST (Basic Local Alignment SearchTool) en fonction de la
similarité avec les bases de données de référence. Les « NGS reads », de longueur minimale
de 30 nucléotides, ont été coupés en utilisant le logiciel CLC Genomic Work bench 6.5, avec
un minimum de qualité de 99% par base et comparés avec la séquence de référence (TOSV,
souches tunisiennes, GenBank, N° d'adhésion. JX867534, JX867535 et JX867536 pour les
segments L, M et S, respectivement). Les paramètres ont été mis de telle sorte que chaque
lecture acceptée devrait correspondre à la séquence de référence avec au moins 50% de sa
longueur, et au moins 80% d’identité avec la séquence. Les trous dans le génome ont été
séquencés à partir des produits de PCR chevauchante amplifiés par des amorces conçues à
partir de la séquence de référence, en utilisant soit le séquençage direct par la technique de
Sanger soit la technique de NGS.
Les extrémités 5’et 3’de chaque segment ont été séquencées en utilisant une amorce
comprenant 8 nucléotides conservés des séquences communes aux phlébovirus précédemment
décrite (Palacios et al., 2013). Une ultime vérification du génome a été effectuée par la
technique de séquençage de Sanger depuis les produits de PCR chevauchante couvrant la
totalité du génome.
7.1. Distances génétiques et analyse phylogénétique
Les séquences complètes d'acides aminés des segments S (les gènes N et Ns), M (les gènes
Gn et Gc) et L (polymérase) de la souche virale isolée dans cette étude ont été alignées avec
des séquences proches de celles de souches de TOSV récupérées à partir de
la base de données GenBank jusqu'en Avril 2015.
Les acides nucléiques et les acides aminés ont été alignés en utilisant l'algorithme CLUSTAL
du logiciel MEGA 5 (Tamura et al.,2011). L’analyse, par Neighbor-joining en utilisant le
paramètre Kimura-2 et le modèle de p-distance, a été réalisée par le logiciel MEGA 5, avec
1000 pseudoréplications de bootstrap.
8. Séroneutralisation
Principe (Fig.27): Le test de neutralisation virale est une technique quantitative permettant de
mettre en évidence la présence d’anticorps neutralisants dans un sérum. Son principe repose

Chapitre II Matériel et Méthodes
65
sur l'interaction des anticorps contre les protéines virales externes. Ces anticorps empêchent la
pénétration et donc la multiplication du virus dans les cellules sensibles.
Pour mettre en œuvre la réaction, des quantités constantes de virus sont incubées avec des
dilutions en série du sérum testé et inoculées dans des microplaques de culture cellulaire et
incubées pendant 3 à 7 jours. L'absence d'effet cytopathogène reflète la présence d'anticorps
neutralisants dans le sérum testé.
Fig. 27. Principe de la réaction de le séroneutralisation 3
Un test de séroneutralisation a été réalisé sur le deuxième aliquote des échantillons de sérums
après consentement des prélevés suivant la réglementation algérienne pour de telles études. La
séroneutralisation a été adaptée suivant le protocole décrit par Alkan et al. (2013) et réalisée
en utilisant des microplaques de 96 puits avec des cellules Vero (ATCC CCL81).
Après inactivation des sérums à 56°C pendant 30 min (pour éviter la lyse cellulaire par le
complément présent dans le sérum), des dilutions de 1 :10 à 1 :160 ont été réalisées pour
chaque sérum: A partir de la dilution 1 :10, des dilutions de ½ en ½ ont été préparées. Dans
une microplaque 96 puits et à l’aide d’un automate de pipetage Eppendorf epMotion 5075
(afin d’avoir une bonne reproductibilité de l’essai), un volume de 50 µl de chaque dilution a
été déposé dans les puits correspondants.
___________________
3http://test10.vetion.de/fr/maison/diagnostic/methodes-diagnostiques/anticorps. Consulté le
25/09/2018 à 8:20

Chapitre II Matériel et Méthodes
66
50 µl de la dose challenge contenant 1000 DICP50 ou 100 DICP50 (deux essais indépendants)
du virus TOSV (souche MRS2010-4319501, préalablement déterminée par titrage: 106.42
DITC50/ml) ont été ajoutés dans chaque puits à l'exception des sérums contrôles (contenant
des sérums témoins dilués à 1:10 et des cellules Vero sans virus) et des contrôles cellulaires
négatifs (contenant des cellules Vero sans virus et sans sérum) dans lesquelles un volume de
50 µl du PBS a été ajouté (Tableau XV). Après homogénéisation par des mouvements
d’aspiration et refoulement, la plaque est scellée et incubée 1h à 37°C.
Tableau XV. Distribution des sérums et des contrôles sur une plaque de micro-titration
utilisée dans le test de séroneutralisation virale
Une suspension de cellules Vero (contenant environ 2 x 105cellules /ml dans du milieu
EMEM enrichi avec 5% de sérum fœtal bovin, 1% de Pénicilline streptomycine, 1% de L-
glutamine 200 mM, 1% de la kanamycine et 3% de Fungizone) a été déposée à raison de 100
µl par puits tout en veillant à homogénéiser fréquemment la suspension. La plaque est alors
scellée et incubée à 37 ° C en présence de 5% de C02.
Au bout de 5 jours d’inoculation, les microplaques sont observées à l’aide un microscope
inversé. La présence de l'effet cytopathique (séroneutralisation avec un titre de 10, 20, 40, 80
ou 160) ou l’absence de cet effet (pas de séroneutralisation) ont été enregistrées. Les seuils de
positivité (Cut off) pour la dose 100 DICP50 et la dose 1000 DICP50 sont respectivement fixés
à 1:20 et 1:10.
L'étude de la variation des taux de séroprévalence entre les différentes classes de taille a été
réalisée par le test Khi 2.
L'analyse statistique a été réalisée à l'aide du logiciel STATISTICA (Version 10.0.228.8).

CHAPITRE III
RESULTATS

Chapitre III Résultats
67
1. Etude entomologique
1.1. Résultats des piégeages
Un total de 1405 pièges huileux a été récupéré sur les 1750 déposés durant une période de 10
moisrépartis sur les années 2012 et 2013 et représentant une surface de 7 m².
Ces pièges ont permis la capture de 2783 phlébotomes adultes dont 2051 mâles (73,70%) et
732 femelles (26,30%) soit un sex-ratio de 2,80.
A partir de 33 pièges CDC, tendus pendant 5 nuits en août 2013, 22998 phlébotomes adultes
ont été récoltés, soit des moyennes de 4600 phlébotomes par jour et de 700 par piège et par
nuit (700 phlébotomes/nuit/piège).
L’échantillonnage était constitué de 13431 (58%) individus vivants et de 9567 (42%)
individus morts. Parmi ces derniers, 1500 ont été examinés pour la détermination du sexe, 869
(57,93%) étaient de sexe mâle et 627 (41,80%) de sexe femelle, soit un sex-ratio de 1,38, et 4
de sexe indéterminé.
Le sexe de l’ensemble de ces insectes a été déterminé en se basant essentiellement sur l’aspect
du génitalia, qui est allongé chez le mâle (Fig.28A) et simple chez la femelle (génitalia interne)
(Fig.28 B).
Fig. 28. Phlébotome adulte mâle (×4) (A). Phlébotome adulte femelle gorgée (×4) (B)
1.2. Inventaire taxonomique
La totalité des phlébotomes capturés par pièges huileux et les 1500 phlébotomes capturés
morts par pièges CDC ont été identifiés sur la base de critères morphologiques (d’après les
clés de diagnose établies par Abonnenc, 1972; Léger et al., 1983 et Killick-Kendrick, 1991).
Cinq espèces du genre Phlébotomes et une espèce du genre Sergentomyia ont été recensées à
partir de l’ensemble des captures: Phlebotomus perniciosus, P. perfiliewi, P. longicuspis,
P. papatasi, P. segenti et Sergentomyia minuta.
A B

Chapitre III Résultats
68
Les pièges adhésifs ont permis l’enregistrement de deux autres espèces du genre
Phlebotomus : Phlebotomus langeroni et P. chabaudi. L’espèce de quatre spécimens du genre
Sergentomyia n’a pu être déterminée en raison de leur état détérioré.
• Genre Phlebotomus Rondani, 1843
- Soies des tergites abdominaux, toutes dressées;
- armature cibariale absente ou rudimentaire;
- spermathèques complètement ou incomplètement segmentées;
- 4 à 5 épines sur le style des génitalia.
o Sous-genre Larroussius Nitzulescu, 1931
- Style avec 5 longues épines ; épine médiane plus rapprochée de l’épine basale que des
épines terminales (Fig. 29 A) ;
- touffe de 12 soies longues et bien différenciées à l’apex du coxite ;
- petit tubercule basal du coxite portant plus de 6 soies ;
- fourreau (valve) pénien long et de forme variable ;
- pharynx avec armature formée de nombreuses petites dents punctiformes (Fig. 29 B) ;
- spermathèques segmentées avec un long processus terminal.
Fig. 29. Critères d’identification du sous-genre Larroussius
Génitalia (×10) (A). Pharynx (×100) (B)
❖ Phlebotomus (Larroussius) perniciousus Newstead, 1911.
▪ Mâle
- Style avec 5 épines ;
- valve pénienne bifurquée à l’extrémité avec occasionnellement une petite dentsurnuméraire
(Fig.30 A, B) ;

Chapitre III Résultats
69
- fourche terminale très polymorphe à pointes égales ou inégales et pourvues ou non de
denticulations latérales ;
▪ Femelle
- Corps de la spermathèque constitué d’une dizaine d’anneaux ;
- processus terminal avec un long coudiminuant progressivement de diamètre de la base à
l’extrémité distale avec une petite tète arrondie (Fig.30 C) ;
- spermathèque (corps, processus terminal et conduit) de structure morphologique identique
à celle observée chez P. longicuspis; caractère différentiel représenté par l’aspect de la partie
terminale du conduit de la spermathèque (Leger et al., 1983).
Fig. 30. Critères d’identification morphologique de P. perniciosus
Valve pénienne (×40, ×10) (A et B). Cœcum (×40) (C). Dessin des spermathèquesau
microscope optique avec chambre claire (×10) (D)
❖ Phlebtomus (Larroussius) longicuspis Nitzulescu,1930
▪ Mâle
- Style avec 5épines (Fig. 31 A, C, D) ;
- valves péniennes terminées par une pointe unique, longue, plus ou moins recourbée vers le
bas.

Chapitre III Résultats
70
▪ Femelle
- Pharynx de morphologie similaire à celle de P.perniciosus ;
- spermathèques avec 9 à 10 anneaux (Fig. 31 B, E).
Fig. 31. Critères d’identification morphologique de P. longicuspus
Valves péniennes (×10) (A).Cœcum et spermathèque (×40) (B). Génitalia mâle (×4) (C). Dessin à la
chambre claire du génitalia mâle (×4) (D) et du génitalia femelle (x10) (E)
❖ Phlebotomus (Larroussius) perfiliewi Parrot, 1930
▪ Mâle
- Valves péniennes à pavillon large, à extrémité distale spatulée (Fig. 32 A, B) ;
- présence d’une série de denticulations sur le bord externe des valves.
▪ Femelle
- Armature pharyngienne occupant le ¼ de la longueur de l’organe ;
- plage de denticules longs et minces, disposés en rangées semi-circulaires ;
A B
D
C

Chapitre III Résultats
71
- spermathèque avec un nombre d’anneaux plus élevé (12-18) (Fig. 32 C).
Fig. 32. Critères d’identification morphologique de P. perfiliewi
Styles et valves péniennes (×4) (A).Valves péniennes (×40) (B). Spermathèque (×40) (C)
❖ Phlebotomus (Larroussius) langeroni Nitzulescu, 1930
- Bord dorsal de la valve pénienne avec 4 à 6 dents ;
- valve arquée et spatulée à l’apex.
o Sous-genre Phlebotomus Rondani, 1843
- Génitalia grand ;
- style long et mince avec 3 courtes épines spatulées à l’extrémité et 2 à proximité du milieu
du segment ;
- fourreau pénien court et conique ;
- spermathèques segmentées avec un bouton terminal.
❖ Phlebotomus (Phlebotomus) papatasi Scopoli, 1786
▪ Mâle
- Génitalia très développé (Fig.33 A, C) ;
- style long portant 5 épines courtes et spatulées (3 terminales et 2 échelonnées sur le tiers
distal) ;

Chapitre III Résultats
72
- lobe latéral terminé par 2 épines spatulées ;
- pénis conique et mousse à son extrémité ;
- paramères trilobés.
▪ Femelle
- Pharynx postérieur armé dans son tiers postérieur d’une plage de dents écailleuses
irrégulières dans leur forme et leur implantation ;
- spermathèques annelées (6-9 anneaux) et sans col (Fig. 33 B, D).
Fig. 33. Critères d’identifications morphologiques de P. papatasi
Styles et valves péniennes (×10) (A). Cœcum (×40) (B). Dessin au microscope optique avec
chambre claire, du génital mâle (×10) (C) et du génitalia femelle (×10)
o Sous-genre Paraphlebotomus Theodor, 1948
-Style à 5 épines ;
- Style long à 5 épines courtes et spatulées, dont trois terminales ;
- paramère trilobé, lobe latéral armé de deux épines terminales.

Chapitre III Résultats
73
❖ Phlebotomus (Paraphlebotomus) sergenti Parrot, 1917
▪ Mâle
- Génitalia court ;
- style court avec 4 longues épines dont 2 sont terminales ou subterminales et 2 à proximité de
la base du segment (Fig. 34 A) ;
- fourreau pénien court avec un crochet terminal (Fig. 34B).
▪ Femelle
- Pharynx avec une armature de fortes dents cunéiformes (Fig. 34 C) ;
- spermathèques segmentées avec un segment terminal élargi (Fig. 34 D).
Fig. 34. Critères d’identification morphologique de P. sergenti
Styles et valves péniennes (×10) (A).Valves péniennes (×40) (B). Pharynx (×100) (C).
Spermathèques (×100) (D)
❖ Phlebotomus (Paraphlebotomus) chabaudi Croset, Abonnenc et Rioux, 1970
- Valve pénienne recourbée en « bec de rapace » à l’apex ;
- une seule épine terminale sur le style ;
- lobe latéral plus court ou égal au coxite et à la base du coxite, présence d’un lobe court,
trapu et nettement dilaté à l’extrémité.
A
B
C
D
A

Chapitre III Résultats
74
• Genre Sergentomyia França et Parrot, 1921
- Soies abdominales couchées dans la plupart des espèces, quelques-unes dressées dans un
sous-genre ;
- armature cibariale bien développée ;
- spermathèques tubulaires, capsuliformes ou segmentées.
Génitalia des mâles : style avec 4 longues épines et 1 petite soie sur le côté interne ; 2 longues
épines et 1 petite soie dans un sous-genre.
❖ Sergentomyia (Sergentomyia) minuta Parroti, Adler et Theodor, 1927
▪ Mâle
- Style trapu, pourvu de 4 épines terminales groupées et d’une soie non caduque insérée à
l’union des deux tiers proximaux et du tiers apical (Fig. 35A) ;
- édéage court et robuste, se terminant en doigt de gant (Fig. 35 A).
▪ Femelle
- Cibarium armé avec une plage pigmentée plus ou moins développée (Fig. 35 B) ;
- spermathèques de formes variées (Fig. 35C) ;
- conduits lisses ou ondulés ou capsules lisses avec ou sans épines, mais non segmentées.
Fig. 35. Critères d’identification morphologique de S. minuta
Styles et valves péniennes (×10) (A). Cibarium(×4) (B). Spermathèques (×40) (C)
A
B
C

Chapitre III Résultats
75
1.3. Aspect écologique
Les résultats de l’étude morphologique et taxonomique ont été traités par l’abondance relative,
un indice écologique de composition de la faune phlébotomienne.
Ce paramètre a été évalué selon le genre, le sous-genre et l’espèce et par sexe. Les valeurs
obtenues et présentées séparément selon le type de piégeage sont exprimées sous formes de
tableaux et de figures.
1.3.1. Abondance relative selon le taxon des phlébotomes piégés sur papiers huileux
Il ressort des valeurs obtenues que :
- le genre Sergentomyia, représentant 73% des captures, est plus répandu que le genre
Phlebotomus (27%) (Fig.36).
- Dans le genre Phlebotomus, Larroussius est le sous-genre le plus abondant, 25,54%, contre
respectivement 1,29% et 0,57% pour Phlebotomus et Paraphlebotomus (Tableau XVI).
- Sergentomyia minuta est l’espèce la plus fréquente, totalisant 72,66% des individus piégés,
suivie de Phlebotomus perniciosus (19,4%) et de P. perfiliewi (4,6%). P. Langeroni est
l’espèce la moins fréquente (0,1% ), les autres espèces étaient rares (Tableau XVII et Fig.37).
Les résultats montrent par ailleurs que toutes les espèces étaient représentées par une
dominance des individus mâles.
Fig. 36.Abondance relative selon le genre des phlébotomes capturés par pièges huileux
Phlebotomus27%
Sergentomyia73%

Chapitre III Résultats
76
Tableau XVI.Abondance relative selon le sous-genredes phlébotomes capturés par pièges
huileux
Genre et sous genre Espèce Nb d’individus
(n=2783)
Fréquence
%
Phlebotomus (larroussius)
P. perfiliewi
(Parrot, 1930)
129
25,54
P. perniciosus
(Newstead 1911)
540
P. longicuspis
(Nitzulescu, 1930)
40
P. langeroni
(Nitzulescu, 1950)
02
Phlebotomus (Phlebotomus) P. papatasi
(Scopoli, 1786)
36 1,29
Phlebotomus (Paraphlebotomus)
P. sergenti
(Parrot, 1917)
15
0,57 P. chabaudi
(Croset, Abonnenc
et Rioux 1970)
01
Sergentomyia S. minuta
Parroti (Adler et
Theodor, 1927)
2020
72,58
Total 2783 100

Chapitre III Résultats
77
Tableau XVII. Abondance relative par sexe des espèces recensées
Espèces Nombre de
phlébotomes
mâles
A.R des
phlébotomes
mâles(%)
Nombre de
phlébotomes
femelles
A.R des
phlébotomes
femelles(%)
Nombre de
phlébotomes
mâles et
femelles
A.R des
phlébotomes
mâles et
femelles (%)
P. perniciosus 490 23,9 50 6,8 540 19,4
P. papatasi 24 1,2 12 1,6 36 1,3
P. longicuspis 37 1,8 3 0,4 40 1,44
P. perfiliewi 119 5,8 10 1,4 129 4,6
P. langeroni 2 0,1 0 0 2 0,1
P. sergenti 14 0,7 1 0,1 15 0,5
P. chabaudi 1 0,05 0 0 1 0,04
Sergentomyia
minuta
1364 66,45 656 89,7 2020 72,66
Total 2051 100 732 100 2783 100
Fig.37. Abondance relative par sexe des espèces recensées
1.2.3. Abondance relative selon le taxon des phlébotomes capturés par pièges lumineux
Les valeurs obtenues montrent :
- une forte abondance du genre Phlebotomus (97,4%) contrairement au genre Sergentomyia
(2,6%) (Fig.38) ;
- une abondance élevée des sous-genres Larroussius et Phlébotomes comparée à celle du
sous-genre Paraphlebotomus (Tableau XVIII) ;
0.00
20.00
40.00
60.00
80.00
100.00
72.66
19.40
4.60 1.40 1.30 0.50 0.10 0.04
Ab
on
da
nce
rela
tiv
e (%
)
Espèce
Pièges huileux Mâle Femelle Total

Chapitre III Résultats
78
- une large distribution de l’espèce P. perfiliewi (51,60%) suivie de celle de P. perniciosus
(36,7%) ;
- une présence quasiment rare de P. sergenti (0,13%) ;
- une présence faible de S. minuta (2,34%) (Tableau XIX et Fig.39).
Fig. 38. Abondance relative selon le genredes phlébotomes capturés par pièges lumineux
Tableau XVIII. Abondance relative selon le sous-genredes phlébotomes capturés par pièges
lumineux
Genre et sous genre Espèce Nb d’individus
(n=1.500)
Fréquence
(%)
Phlebotomus (larroussius)
P. perfiliewi
(Parrot, 1930)
772 90,8
P. perniciosus
(Newstead, 1911)
550
P. longicuspis
(Nitzulescu, 1930)
40
Phlebotomus (Phlebotomus)
P. papatasi
(Scopoli, 1786)
97 6,47
Phlebotomus (Paraphlebotomus)
P. sergenti
(Parrot, 1917)
02 0,13
Sergentomyia
S. minuta
(Parroti, Adler et
Theodor, 1927)
35 2,6
Sergentomyia sp 4
Total 1500 100
Phlebotomus97%
Sergentomyia3%

Chapitre III Résultats
79
Tableau XIX.Abondance relative par sexe des espèces recensées
Espèces Nombre de
phlébotomes
mâles
A.R des
phlébotomes
mâles(%)
Nombre de
phlébotomes
femelles
A.R des
phlébotomes
femelles(%)
Nombre de
phlébotomes
mâles et
femelles
A.R des
phlébotomes
mâles et
femelles (%)
P. perniciosus 422 48,56 128 20,41 550 36,76
P. papatasi 57 6,56 40 6,38 97 6,48
P. longicuspis 22 2,53 18 2,87 40 2,67
P. perfiliewi 348 40,05 424 67,62 772 51,60
P. langeroni 0 0 0 0 0 0
P. sergenti 0 0 2 0,32 2 0,13
P. chabaudi 0 0 0 0 0 0
Sergentomyia minuta 20 2,30 15 2,39 35 2,34
Total 869 100 627 100 1496 100
Fig. 39. Abondance relative des espèces recensées
Les individus mâles de toutes les espèces sont dominants (58.08% vs 41.91%) à l’exception
de P. Perfiliewi représenté majoritairement par des individus femelles (sex-ratio de 0,82).
2. Etude parasitologique
2.1. Recherche des leishmanies chez le vecteur
Les phlébotomes capturés vivants au CDC ont été examinés au Laboratoire d’Hygiène de
Draa El Mizan dans le but de détecter des formes promastigotes. Cette recherche s’est révélée
négative et par conséquent la culture du parasite n’a pu être envisagée pour son identification.
0.00
20.00
40.00
60.00
80.00
2.34
36.76
51.60
2.676.48
0.13 0.00 0.00
Ab
on
da
nce
rela
tiv
e (%
)
Espèce
Pièges CDC Mâle Femelle Total

Chapitre III Résultats
80
2.2. Diagnostic sérologique de cas humains de leishmaniose viscérale
Le test de western-blot effectué sur les 370 sérums collectés à partir des personnes habitant la
région de Draa El Mizan a révélé 284 cas négatifs et 86 cas positifs à Leishmania (Fig.40).
Les caractéristiques de la population testée sont représentées dans le tableau ci-dessous.
Tableau XX. Caractéristiques de la population étudiée
Fig.40.Résultats du test Western-Blot
Les résultats indiquent que la tranche d’âge de 21 à 40 ans est celle où le parasite a été le plus
souvent détecté, avec 36 sérums positifs (Fig.41). Cependant, l’analyse statistique, effectuée
par le test Khi2 (Annexe I) n’a pas montré de différence significative entre les tranches d’âge
(Khi2 de Pearson: 4.0323, p=0.2579).
Par ailleurs, ce portage asymptomatique de leishmanies s’est révélé plus fréquent chez le sexe
féminin avec 53 cas (53/248) que chez le sexe masculin avec 33 cas (33/122) (Fig.42).
0
50
100
150
200
250
300
Test N Test P
No
mb
re d
e c
as
Age Nombre % Féminin Féminin(%) Masculin Masculin(%)
< 20 24 6,48 13 54,16 11 45,83
21-40 155 41,89 115 74,19 40 25,80
41-60 97 26,21 69 71,13 28 28,86
> 60 94 25,40 51 54,25 43 45,74
Total 370 100 248
122

Chapitre III Résultats
81
Fig.41.Répartition selon les tranches d’âges des cas positifs à Leishmania
Fig.42.Répartition selon le sexe des cas positifs à Leishmania
0
20
40
60
80
100
120
140
0-20 21-40 41-60 > 60
No
mb
re d
e c
as
Tranches d'âges
Test N
Test P
0
50
100
150
200
250
Féminin Masculin
P N

Chapitre III Résultats
82
3. Etude virologique
3.1. Isolement et caractérisation du virus Toscana par analyse moléculaire
3.1.1. Recherche du virus
Sur les 22998 phlébotomes capturés par pièges lumineux CDC, 21498 non identifiés, ont été
répartis en 247 pools. L’ARN viral a été extraitde ces pools par deux techniques, extraction
par le robot et extraction manuelle. Les produits d’extraction ont fait objet de deux techniques
d’amplification afin de rechercher le virus Toscana, la RT-PCR, afin de transformer l’ARN en
ADN, et la PCR nichée, dans le but d’amplifier les produits de la première PCR.
Un des pools, le 189, s’est révélé positif par RT-PCR et PCR nichée en utilisant les amorces
SFNV-NP1 et SFNV-NP12, respectivement
Le produit de la RT-PCR correspondait à 437 pb et celui de la PCR nichée à 304 pb
(Fig.43,44).
Fig.43. Produit de la RT-PCR
Fig.44. Produit de la PCR nichée
Une partie de la séquence du gène N codant pour la nucléocapside a été identifiée avec une
similitude de 96% avec la souche tunisienne TOSV, T152 (numéro d’accession, GenBank.
JX867536) (Fig.45).

Chapitre III Résultats
83
Fig.45. Blast du gène N codant pour la nucléocapside du pool 189 comparé à celui de la
souche tunisienne TOSV T152 (JX867536)
3.1.2. Isolement du virus
Au jour 4 de passage 0, un effet cytopathogène a été observé sur la culture des cellules Vero,
inoculée avec le pool 189. Cette souche virale dépistée pour la première fois en Algérie a été
nommée TOSV-Algérie 189. L’absence du CPE a été notée dans le contrôle négatif.
L’échantillon a été lyophilisé pour une conservation à long terme, la souche virale a été
incluse dans la collection des Archives de Virus européenne (http: //www.europeanvirus-
archive.com/) où ils sont accessibles au public sous la référence "Toscana virus Algeria strain
189" pour les recherches universitaires.
3.1.3. Séquençage du génome complet
Le surnageant du troisième passage de la culture cellulaire a été utilisé pour le séquençage du
génome complet par la technique de NGS. Après analyses bio-informatiques utilisant les
logiciels BLAST et CLC Genomic Workbench, nous avons démontré que le génome complet
de TOSV-Algérie 189 se compose de 6404, 4215 et 1869 nucléotides correspondant aux
segments L, M et S, avec les numéros d’accession GenBank respectifs: KP694240, KP694241
et KP694242.
Dans le segment L, le gène de la polymérase est composé de 6285 nucléotides (Open Reading
Frame, ORF) et code pour une protéine de 2095 aa. Dans le segment M, le gène de la

Chapitre III Résultats
84
glycoprotéine est composé de 4017 nucléotides (ORF) et code pour une protéine de 1339 aa.
Le segment S est composé de 759 et 948 nt (ORF) codant respectivementpour la protéine de
la nucléocapside composée de 253 aa et d’une protéine non structurale de 316 aa (Fig.46).
Fig.46. Structure génomique du Virus TOSV-Algérie 189
Les séquences obtenues à partir de la technique de séquençage du génome complet de TOSV-
Algérie 189 ont été confirmées par la technique de séquençage de Sanger directement à partir
des produits de PCR.
3.1.4. Analyse phylogénétique
La souche TOSV-Algérie 189 a été groupée avec d'autres séquences de souches TOSV en
tenant compte des 3 segments utilisés dans l’analyse à savoir le segment S, L, et M. D’après
l’analyse des arbres phylogénétiques, la souche TOSV-Algérie 189 est étroitement liée à des
souches TOSV, originaires de la Tunisie et de l’Italie. Ceci implique que la souche algérienne
appartient à la lignée A à la différence des souches originaires de l’Espagne, de la France et
du Portugal qui font partie de la lignée B. La discrimination entre les lignées A et B était sans
ambiguïté pour les trois segments analysés (Fig.47).

Chapitre III Résultats
85

Chapitre III Résultats
86
Fig.47. Analyse phylogénétique des souches de phlébovirus en utilisant les séquences
d’acides nucléiques des segments L (1), M (2) et S (3).
3.2. Séroneutralisation
En utilisant une dose de virus à 1000 DICT50, 171 sérums humains sur les 370 testés (46,2%)
contenaient des anticorps neutralisants contre le TOSV considérant le titre de neutralisation de
10 comme limite. Cependant, en utilisant une dose de virus à 100 DICT50, 170 sérums
(45,95%) contenaient des anticorps neutralisants contre TOSV considérant le titre de
neutralisation de 20 comme limite. La corrélation entre les deux résultats observés à partir de
la dose virus 100 et 1000 TCID50 a été significative avec un p⩽ 0,05 (Coefficient: 0,427). La
moyenne géométrique des titres de TOSV MN selon le groupe d'âge est montrée dans les
figures 48 et 49. La variation des taux de séroprévalence chez les groupes d'âge a été calculée
et jugée statistiquement significative par le test de Khi2 (p⩽ 0,05) (Annexe II).

Chapitre III Résultats
87
Tableau XXI. Répartition des anticorps neutralisants du virus Toscana selon le sexe et l’âge
pour 1000 TCDI50
Age Nombre
de cas
Sexe Titre de neutralisation
F M Négatif ≥ 10 ≥ 20 ≥ 40 ≥ 80 ≥ 160 Total, n (%)
0-20 24 13 11 22 2 0 0 0 0 2(8,3%)
21-40 155 115 40 90 59 5 0 0 1 6 5(41,9%)
41-60 97 69 28 50 37 9 0 1 0 47 (48,5%)
>60 94 51 43 37 47 9 0 1 0 57 (60.6%)
Total 370 248 122 199 145 23 0 2 1 171 (46.2%) TCID50 :(Tissue Culture Infectieuse Dose)la dose infectieuse de culture tissulaire qui pourra infecter 50% si les
monocytes sont infectés par un inoculum défini.
Tableau XXII. Répartition des anticorps neutralisants du virus Toscana selon le sexe et l’âge
pour 100 TCDI50
Age Nombre de cas Sexe Titre de neutralisation
F M Négatif ≥ 10 ≥ 20 ≥ 40 ≥ 80 Total, n (%)
0-20 24 13 11 21 2 0 1 0 1 (4,17%)
21-40 155 115 40 80 13 20 25 17 62 (40,0%)
41-60 97 69 28 39 8 13 26 11 50 (51,55%)
>60 94 51 43 25 12 18 16 23 57 (60,64%)
Total 370 248 122 165 35 51 68 51 170(45,95%) TCID50 :(Tissue Culture Infectieuse Dose)la dose infectieuse de culture tissulaire qui pourra infecter 50% si les
monocytes sont infectés par un inoculum défini.
Fig.48. Moyenne géométrique des titres pour les anticorps neutralisants de TOSV selon les
groupes d’âges (1000 TCID50)
0
2
4
6
8
10
12
14
16
18
20
< 20 21-40 41-60 > 60
Mo
yen
ne
gé
om
étr
iqu
e d
u t
itre
po
ur
TO
SCV

Chapitre III Résultats
88
Fig.49. Moyenne géométrique des titres pour les anticorps neutralisants de TOSV selon les
groupes d’âges (100 TCID50)
4. Co-infections Leishmania-Toscana
Des anticorps anti-leishmania ont été détectés dans 86 sérums et les anticorps anti-TOSV
dans 171 sérums (positifs pour 1000 TCDI50), par respectivement les tests Western Blot et de
séroneutralisation.
11 sérums se sont révélés positifs aux deux tests, ce qui indique la présence de cas d’infection
virale concomitante à une leishmaniose viscérale parmi la population de Draa El Mizan
(Tableau XXIII). La répartition selon l’âge de ces cas co-infectés a montré que les adules de
plus de 60 ans sont les plus touchés avec une prédominance féminine (Tableau XXIV).
Tableau XXIII. Résultats du Western Blot et de la séroneutralisation
Nombre Western Blot Séroneutralisation
86 P N
171 N P
11 P P
102 N N
0
5
10
15
20
25
20> 21-40 41-60 60<
Mo
yen
ne
gé
om
étr
iqu
e d
u t
itre
po
ur
TO
SCV
5,8
12
15,2
20

Chapitre III Résultats
89
Tableau XXIV. Résultats du Western Blot et de la séroneutralisation selon l’âge et le sexe
Age Western Blot / Séroneutralisation
Total Féminin Masculin
< 20 0 0 0
21- 40 1 1 0
41-60 3 2 1
> 60 7 5 2

CHAPITRE IV
DISCUSSION

Chapitre IV Discussion
90
Les phlébotomes vivent, prolifèrent et se dispersent localement dans des environnements
chauds et humides de toutes les régions du monde. Des espèces du genre Phlebotomus sont
des vecteurs de plusieurs groupes de pathogènes dont les principaux sont représentés par les
leishmanies et des membres du genre phlébovirus.
Le présent travail avait pour but initialement de montrer le rôle du phlébotome avec
identification des espèces, dans la transmission à l’échelle nationale de Leishmania et de
Phlebovirus à partir de deux études, parasitologique et moléculaire. La détection d’infections
humaines dues à ces deux types de pathogènes a été ensuite envisagée.
Les leishmanioses constituent un problème de santé publique en Algérie, essentiellement
dans leur forme viscérale. Des enquêtes épidémiologique ont permis de mettre en évidence les
foyers à leishmanioses et de dresser l’inventaire des espèces de phlébotomes vectrices (Izri et
al., 2014). A ce jour, 24 espèces ont été recensées après l’enregistrement de l’espèce
Phlebotomus mascitti à Larba Nath Irathen en Kabylie (Berdjane-Brouk et al., 2011).
En raison de sa position géographique, de son cadre bioclimatique et de son activité basée sur
l’élevage, Draa El Mizan (Kabylie) offre des gites favorables à la prolifération de plusieurs
espèces de phlébotomes. Comparée aux autres sites de la Kabylie, c’est la région où la densité
des phlébotomes est la plus élevée (Mouloua et al. (2017). Ces caractéristiques naturelles
expliquent le choix de cette région comme zone d’étude pour y rechercher la présence de
phlébovirus.
Les phlébotomes étudiés ont été capturés par des pièges adhésifs et des pièges lumineux type
CDC installés dans différents sites de la zone périurbaine, pendant la période d’activité de ces
insectes.
Un total de 25781 phlébotomes a été récolté à partir de 33 pièges lumineux, déposés pendant
5 nuits du mois d’aout 2013 et de 1405 pièges huileux, déposés au cours des dix mois de
l’année 2012-2013. L’inventaire faunistique global de 4283 phlébotomes a mis en évidence la
présence de 8 espèces de phlébotomes réparties sur deux genres, Phlebotomus et
Sergentomyia. Il s’agit de Phlebotomus perniciosus, P. perfiliewi, P. longicuspis, P. papatasi,
P. sergenti, Phlebotomus langeroni, P. chabaudi et Sergentomyia minuta
Mouloua et al. (2017) ont recensé 6 espèces du genre Phlebotomus et une espèce du genre
Sergentomyia à partir de 4806 phlébotomes capturés dans la région de Draa El Mizan durant
la période de 2007 à 2010, à l’aide de pièges lumineux et de pièges adhésifs. Ces espèces sont
Phlebotomus perniciosus, P. perfiliewi, P. longicuspis, P. papatasi, P. sergenti, P. ariasi et
Sergentomyia minuta.

Chapitre IV Discussion
91
La composition de ces deux faunes phlébotomiennes indique l’absence de P. ariasi dans nos
captures et celle de P. langeroni et P.chabaudi dans l’inventaire dressé par ces auteurs. Ces
résultats sont probablement liés aux conditions météorologiques ou environnementales ou
encore à la méthode d’échantillonnage, notamment le type de biotopes choisi pour la capture
des phlébotomes.
Une étude antérieure a montré que parmi les 15 espèces rencontrées en Algérie, seules deux
dominent par leur abondance, Phlebotomus perniciosus et Sergentomyia minuta. P.
perniciosus représentait à lui seul, 64,8% des exemplaires du genre Phlebotomus (Dedet et al.,
1984). Ce genre regroupe la quasi-totalité des espèces anthropophiles, donc d’intérêt médical
(Abonnenc, 1972).
Dans notre étude, Sergentomyia minuta, l’espèce la plus répandue en Algérie, a représenté
48,07% des captures totales contre 25,45% pour Phlebotomus perniciosus.
Mouloua et al. (2017) ont rapporté une dominance de P. perniciosus avec un taux de présence
de 44,3%, à partir de l’ensemble des captures.
Dans l’Est algérien, Phlebotomus perniciosus est retrouvé dans les étages, bioclimatique
humide et semi-aride avec une fréquence respective de 38% et de 46,33% (Bounamous,
2010). Cet insecte occupe un vaste territoire géographique dans le bassin méditerranéen,
particulièrement dans la partie septentrionale, jusqu’à la lisière saharienne (Tassili et Hoggar)
(Rioux et al., 1967). Il est répandu dans les régions tempérées à une altitude inférieure à
1.000 mètres et est capable de vivre dans différentes zones notamment rurales, périurbaines
ou urbaines (Dedet et al., 1984; Izri, 1993; Bounamous, 2010).
Cette espèce, décrite dans l’ile de Malte par Newstead en 1911 et signalée pour la 1ère
fois en
Algérie en 1914 par Sergent, est considérée comme espèce importante du point de vue
épidémiologique (Dedet et al., 1984). Elle est incriminée dans la transmission de Leishmania
infantum dans le bassin méditerranéen (Harrat et al.,1986; Rioux et al.,1986) et c’est l’espèce
responsable de la forme viscérale et de la forme cutanée propre à Draa El Mizan (Dedet et al.,
1984; Boudrissa et al., 2012; Izri et al., 2014). Mise à part sa vaste distribution, elle présente
deux formes atypiques retrouvées surtout dans l’Ouest et à l’extrême Sud (Rioux et al., 1967;
Berchi et al.,2007). Ses variations morphologiques sont à l’origine de la description de
nombreuses sous-espèces ou variétés qui ont été par la suite élevées au rang d’espèces comme
P. tobbi, P. langeroni, P.orientalis et P.longicuspis.
La leishmaniose cutanée sporadique du Nord est transmise par P. Perfiliewi (Izri et Bellazoug,
1993) que nous avons recensé à Draa El Mizan avec une abondance de 21,4%. Cette
fréquence pourrait expliquer l’apparition depuis 1996 de la forme cutanée dans un foyer

Chapitre IV Discussion
92
connu initialement pour la forme viscérale (Dedet et al., 1984; Izri et al., 2006; Boudrissa et
al., 2012). Il s’agit d’une espèce décrite pour la première fois en Algérie par Parrot à Médéa
(Parrot, 1935) et qui se distribue dans les étages humide, subhumide avec une fréquence
élevée et semi-aride, mais elle est absente dans les étage arides et saharien (Dedet et al.,
1984). Elle est signalée avec une abondance de 34,9% dans l’étude de Mouloua et al. (2017);
selon ces auteurs, elle évolue dans le même biotope que l’espèce P. perniciosus.
P. papatasi, à répartition mondiale très vaste (Lewis, 1982) est le vecteur majeur de la forme
cutanée à L. major qui sévit dans les zones arides (Bellazoug, 1983; OMS, 1990; Harrat et
al., 1992). Sa densité est maximale aux étages péri-aride et aride mais elle devient rare dans
les étages subhumide, humide et semi-aride (Guernaoui et al., 2005). Les travaux de
Boudrissa et al. (2012) ont mis en évidence l’extension de L. major des zones arides vers les
zones semi-arides de la vallée de la Soummam, ce qui expliquerait la présence de cette espèce
à Draa El Mizan. Ce phénomène est probablement lié à l’impact des changements climatiques
et particulièrement du phénomène de la désertification de la région steppique au Nord du
Sahara. P. papatasi abonde dans les régions à basse altitude, entre 300 et 400 mètres; il est
retrouvé au Sahara plus souvent autour des terriers du rongeur Psammomys obesus (OMS,
1990; Boubidi et al., 2011). Il devient moins fréquent au fur et à mesure que l’on s’éloigne de
ces gites. C’est probablement la raison pour laquelle nous l’avons enregistré avec une faible
fréquence (3,11%.).
Depuis longtemps, la leishmaniose cutanée du Nord à Leishmania infantum et la leishmaniose
cutanée du Sud à Leishmania major ont été séparées par la chaîne montagneuse de l’Atlas
Tellien qui constitue une barrière naturelle. Un nouveau foyer de leishmaniose cutanée est
apparu dans la localité d’El M’Hir, située sur le versant Nord de cette chaîne. Les
prospections entomologiques ont montré la prédominance de P. papatasi et
P.perniciosus (Izri et al., 1992).
Suite à des investigations menées dans des foyers au Nord du Maroc, Rioux et al. (1997) ont
lié l’augmentation de l’incidence des leishmanioses, d’une part à l’augmentation des densités
des vecteurs, en particulier P. sergenti et P. longicuspis, et d’autre part à l’allongement de
leurs périodes d’activité.
P. longicuspis (1,87%) était faiblement représenté dans nos captures; P. sergenti et les
espèces P.langeroni et P.chabaudi étaient rares avec des taux de présence n’atteignant pas 1%.
L’ensemble de ces espèces a été recensé avec des taux de fréquence différents selon le mode
de piégeage. P. perniciosus a été retrouvé dans les captures aux pièges CDC avec une
abondance relative élevée (36.76%) par contre il n’a représenté que 19,4% des spécimens

Chapitre IV Discussion
93
récoltés dans les pièges adhésifs. C’est une espèce zoo-anthropophile et endophile, qui semble
bien adaptée à l’environnement humain (Mouloua et al., 2017). Elle peut être capturée à
l’intérieur des abris d’animaux domestiques ou de bâtiments d’élevage. Elle présente aussi un
tropisme pour l’Homme plus élevé que P. ariasi.
P. perfiliewi a été recensé avec une abondance de 51,60% et de 4,6% à partir respectivement
des pièges huileux et des pièges lumineux. C’est une espèce plus endophile que P.
perniciosus, elle pénètre dans divers abris d’animaux et dans les habitations humaines.
L’étude de Moulahem (1998) a montré que l’essentiel des captures réalisées dans un
appartement à Constantine était représenté par P. perfiliewi (75,5%) contre 23,3% pour P.
perniciosus.
Sergentomyia minuta dont l’abondance est très élevée (72,58%) à partir des captures par
pièges adhésifs, a été est très faiblement retrouvé (2.34%) dans les pièges lumineux. Ce
résultat peut s’expliquer par le fait qu’il s’agit d’une espèce herpétophile qui cherche son hôte
dans les anfractuosités où sont posés les papiers huileux. Boussaa et al. (2005) ont enregistré
Sergentomyia spp avec une fréquence de 36.5% dans la région de Marrakech mais avec une
différence de richesse en espèces entre un milieu rural et un milieu urbain. Des travaux menés
dans des foyers méditerranéens, indiquent une prévalence comprise entre 70 et 96.6%
(Haddadi et Dedet, 1974; Benallal et al., 2014).
Nos échantillonnages de phlébotomes ont été fructueux notamment avec le mode de capture
par les pièges lumineux type CDC. Ce type de piège nous a permis de récolter un nombre de
spécimens ayant donné lieu à un échantillon global représentatif pour un inventaire
taxonomique. Il a démontré sa qualité en termes de quantité et de qualité de spécimens
capturés, mais aussi de temps. Il permet de récolter des espèces rares à potentiel épidémique
voire pandémique. Aussi les spécimens capturés vivants permettent d’entreprendre des
examens de génomique et de protéomique nécessaires à l’identification de l’agent pathogène
en cause. De ce fait, ils restent un outil de choix pour la surveillance entomologique des
leishmanioses et autres maladies vectorielles émergentes ou réémergentes.
La recherche chez le phlébotome de l’ADN parasitaire n’a pu pas aboutir et celle des
leishmanies était négative et ce probablement en raison du taux d’infection qui selon certains
auteurs est généralement très faible (0,01 à 1%), même en zone d’endémie (Hashiguchi,
2003). Ces résultats ont été obtenus lors d’investigations menées à Taourirt Aden, dans la
région de Mekla (Kabylie) (Izri et al., 1988). Ces auteurs avaient trouvé une femelle P.
perniciosus infectée sur 400 disséquées et une femelle P. perfiliewi infectée sur 710

Chapitre IV Discussion
94
disséquées soit un taux de 0,16% de femelles contaminées. Seuls les promastigotes provenant
de P. perniciosus ont pu être cultivés et l’espèce identifiée était L. infantum MON-1.
Dans l’étude de Mouloua et al. (2017), l’emploi de la PCR pour la détection de l’ADN du
parasite, a permis d’obtenir un taux d’infection de 31,9%.
Les sérums de patients développant une leishmaniose-maladie reconnaissent en moyenne une
trentaine de fractions antigéniques. Parmi ces fractions protéiques, les antigènes de 14 et 16
kDa semblent les plus fréquents (Marty et al., 1995). Ainsi, un sérum est considéré comme
positif en WB lorsqu'il reconnaît une fraction antigénique de 14 et/ou 16 kDa. Lors de l’étude
prospective, 370 sérums ont été soumis à ce test. Parmi les 86 cas qui se sont révélés positifs,
40,7% reconnaissent les 2 antigènes de 14 et 16 kDa, 36% l'antigène de 16 kDa et 23,3%
celui de 14 kDa.
Ce test permet donc d'explorer, de façon indirecte, une partie de l'immunité à médiation
cellulaire spécifique anti-Leishmania infantum. Marty et al. (1992), signalent que 65% des
sérums de 20 individus asymptomatiques vivant en zone d’endémie marseillaise reconnaissent
par la technique de WB, des antigènes de 14 et 16 (18) kDa. Ils proposent d'utiliser cette
technique pour détecter des anticorps dirigés contre ces 2 protéines qui seraient des marqueurs
biologiques au contact d'un individu avec L. infantum et peut-être d'un portage
asymptomatique. De plus, l'absence de reconnaissance en WB des antigènes de 14 et/ou 16
kDa permet d’éliminer une suspicion de portage asymptomatique dans 90% des cas.
Le WB semble être un outil épidémiologique efficace, sa réalisation peut être différée et
seulement quelques gouttes de sang sont nécessaires puisque 25µl de sérum suffisent pour sa
réalisation. Cependant, la lecture tardive des bandelettes (48 heures après) entraînerait un
absentéisme à l'interprétation (environ 12% des sujets par rapport à la population initiale des
volontaires dans l’étude de Marty et al., 1992). L'étude de Dunan et al. (1990) souligne
l’équivalence de ce test avec l'Intra Dermo Réaction à la Leishmanine (test de Montenegro) in
vitro. Cependant, il ne permet pas de prédire l'évolution clinique d'un sujet séropositif vis à
vis des antigènes de Leishmania (Badro et al., 1996). C’est un test sérologique utile pour le
diagnostic des LV humaines à L. infantum (Alvar et al., 1997; Rolland et al., 1995), plus
sensible que l’'IFI car il est positif dans tous les cas et l'IFI dans moins d'un cas sur deux.
C’est la raison pour laquelle certains auteurs le proposent avant tout prélèvement invasif
(ponction de moelle osseuse).
Il faut souligner que la comparaison des résultats avec d'autres auteurs est difficile parce que
les souches de parasites, les antigènes utilisés et les modalités techniques sont variées.

Chapitre IV Discussion
95
Néanmoins, le WB reste une technique de sensibilité supérieure à quelques tests sérologiques
qui nécessitent un consensus pour la réalisation et l’interprétation de Leishmania spp. Il est
utile pour confirmer une suspicion clinique de leishmaniose ou une suspicion de portage
asymptomatique de cette affection en zone d'endémie.
Des études récentes ont démontré que TOSV, un membre du genre Phlebovirus, n’est pas
seulement confiné à l'Europe du Sud et au Moyen-Orient mais est également présent dans les
pays d'Afrique du Nord (Tunisie et Maroc).
Une étude recherchant la distribution mondiale des anticorps neutralisants dans les
populations humaines contre plusieurs phlébovirus a été réalisée mais aucun anticorps
neutralisants n’a été trouvé que ce soit contre le virus de Sicile (SFSV) ou le virus de Naples
(SFNV). La présence d’anticorps neutralisants contre TOSV n'a pas été recherchée (Tech et
al., 1976).
Au début de cette étude, peu de données étaient disponibles sur l'Algérie. Les deux seules
investigations, réalisées dans le Nord du pays, ont rapporté la présence d’Ig G spécifiques des
virus SFS et SFN, utilisant les tests Elisa et d’immunofluorescence. Ces résultats suggèrent
que ces virus ou des virus apparentés au point de vue antigénique circulent dans les régions
prospectées (Izri et al., 2008; Moreau et al., 2010).
Dans la première étude, l'ARN du SFS-like-virus a été détecté et partiellement séquencé chez
P. ariasi retrouvé dans les captures réalisées dans la région de Kabylie. Le virus n'a pas été
isolé car les échantillons ont été stockés dans du thiocyanate de guanidinium (Izri et al.,
2008). Dans la seconde, l'ARN du SFN-like-virus a été détecté chez P. longicuspis et celui du
SFS-like-virus chez P. papatasi (Moreau et al., 2010). Le virus n'a pas été isolé pour les
mêmes raisons décrites précédemment. Comme les séquences obtenues étaient courtes,
empêchant ainsi des études phylogénétiques détaillées et aussi du fait des réactions croisées
des tests employés, les souches n’ont pas été identifiées. Par conséquent, ces résultats n’ont
pas permis de conclure sur la présence ou l’absence de TOSV en Algérie.
Les résultats de notre étude mettent en évidence la détection, l’isolement et l’identification de
TOSV chez les phlébotomes, en Algérie. La séquence complète de son génome nous a permis
de classer cette souche comme appartenant à la lignée A; et il est nettement transmis aux
populations humaines locales, tel qu'évalué par le taux d’anticorps neutralisants proche de
50%.
À notre connaissance, cette étude est la première à isoler et à caractériser le génome d'un
membre du genre Phlebovirus en Algérie. La souche de TOSV, isolée à partir d'un pool de
phlébotomes collecté à Draa El Mizan, appartient à la lignée A, comme la souche isolée en

Chapitre IV Discussion
96
Tunisie (Bichaud et al., 2013). En revanche, la souche TOSV isolée au Maroc appartient à la
lignée B (Izri et al., 2008). Fait intéressant, il semble que la délimitation des lignées observées
au Sud de l’Europe existe également au Nord de l’Afrique autour du bassin méditerranéen. En
effet, la lignée B est présente au Portugal, en Espagne, en France et au Maroc, alors que la
lignée A est présente en France (en particulier en Corse), Italie, Tunisie et Algérie. Par contre
en Turquie, les deux lignées ont été décrites, bien que le virus n'ait jamais été isolé (Dincer et
al., 2015). En Croatie (Punda-Polic et al., 2012) et en Grèce (Papa et al., 2014), une troisième
lignée (C) a été identifiée sur la base des séquences partielles de polymérase. Cette lignée (C)
diffère des lignées A et B de 29% et 30%, respectivement (Papa et al., 2014).
À ce jour, le taux des phlébotomes infectés par TOSV dans les pays où l'infection a été
détectée sont comme suit; 0,22% en Italie (Verani et al., 1988), 0,05% en Espagne
(Sanbomatsu-Gamez et al., 2005), 0,29% en France (Charrel et al., 2007) et 0,03% en Tunisie
(Bichaud et al., 2013).
Dans cette étude, le taux de phlébotomes infectés est de 0,004%, supposant que seul un
phlébotome a été infecté dans le pool 188. Ces variations ne reviennent pas à des différences
dans les protocoles car ces derniers ont été réalisés dans les mêmes conditions (Bichaud et al.,
2013; Charrel et al., 2007).
La discordance entre ces résultats peut s’expliquer par des variations des conditions
environnementales lors des captures des phlébotomes sur le terrain. Des campagnes
répétitives de piégeage au cours des années successives sont nécessaires pour mieux
comprendre la dynamique des taux d'infection. Il n'a pas été possible d’identifier les espèces
de phlébotomes dans lesquelles la souche algérienne TOSV a été isolée car le nombre de
spécimens capturés par jour était relativement grand (700 phlébotomes/nuit/piège) et les
conditions de travail dans le Laboratoire National Vétérinaire de Draa El Mizan étaient
difficiles. L’échantillon de 1500 phlébotomes identifiés était représentatif et sa composition
phlébotomienne coïncide avec les données de la littérature qui rapportent que TOSV est
transmis par des phlébotomes appartenant au sous-genre Larroussius, le plus souvent par les
espèces P. perniciosus et P. perfiliewi (Verani et al ; 1980; Verani et al., 1984).
Bien que sa présence chez les phlébotomes soit fortement suggérée, la circulation de TOSV
parmi la population a été également recherchée en testant 370 échantillons de sérum collectés
auprès de personnes vivant au voisinage des sites de capture de ces insectes. Comme la
population locale ne se déplace pas souvent à l'étranger, les taux de séroprévalence sont
susceptibles de refléter le degré d’exposition locale à TOSV et de fournir des informations sur
sa circulation.

Chapitre IV Discussion
97
En fonction des critères retenus pour définir un cas positif, la séroprévalence varie largement.
Pour tenter de déterminer les critères qui sont adaptés de façon meilleure à une identification
positive, nous avons calculé la corrélation entre les résultats observés en utilisant une dose de
virus à 100 et 1000 DICT50.
Après comparaison, les mêmes résultats ont été observés avec la combinaison de 100 DICT50
(titre 20) et 1000 DICT50 (titre 10), qui sont des valeurs des cut-off. Dans les deux cas,
l’analyse de la moyenne géométrique des titres selon le groupe d'âge a révélé qu'il existe une
exposition continue pour un sujet pendant une durée de vie, bien que le résultat semble plus
clair avec la dose de virus à 100 DICT50.
Lorsqu’on retient la valeur la plus stricte du cut-off pour déterminer la positivité (1000
TCID50 au titre 20), 6,21% de la population avaient des anticorps neutralisants contre TOSV.
Cette valeur est en accord avec les taux observés dans les régions les plus exposées du Sud-
Est de la France où TOSV circule le plus. Toutefois, il semble plus réaliste d'utiliser la
corrélation de tous les titres. Les anticorps neutralisants ont été détectés chez 46,2% de la
population. Ces résultats sont en accord avec ceux rapportés au Nord de la Tunisie (Sakhria et
al., 2013).
Ces résultats suggèrent que TOSV affecte fortement les personnes exposées aux piqûres de
phlébotomes infectés dans le Nord d'Algérie. Les régions côtières sont les plus peuplées avec
un total d'au moins 34 millions d'habitants qui courent donc le risque de contracter ce virus.
En conclusion, nos résultats confirment que TOSV (lignée A) est
présent chez les phlébotomes appartenant au genre Larroussius dans le Nord de l'Algérie et
infecte la population à des taux comparables à ceux observés dans le Nord de la Tunisie
(Bichaud et al., 2013). Ces taux sont beaucoup plus élevés que ceux décrits dans la majorité
des pays du Sud d'Europe (Charrel et al., 2007; Dincer et al., 2015; Verani et al., 1988).
Par ailleurs, onze cas présentant une co-infection aux deux agents pathogènes ont été
retrouvés parmi la population de Draa El Mizan mais qui ne peuvent pas être confrontés à
cause des variations des techniques de leur dépistage. Néanmoins, ce travail a mis en évidence
la concomitance de ces deux types d’infections, transmises par le même diptère dans une
région d’endémie.

CONCLUSION ET PERSPECTIVES

Conclusion
98
Ces dernières années ont vu un nombre croissant de maladies vectorielles émergentes ou
réémergentes à l’échelle mondiale du fait de la prolifération et de la dissémination de leurs
vecteurs. Elles sont de plus en plus documentées en raison de leur impact sur la santé humaine
et animale. Ces maladies, qu’elles soient humaines ou zoonotiques, sont causées par une large
variété d’agents pathogènes, protozoaires (leishmanioses, paludisme, maladie de Chagas…)
métazoaires (loase, filariose de Bancroft..), bactéries (borréliose de Lyme, borréliose à tique,
peste…) et arbovirus (dengue, chikungunya, fièvre jaune, fièvres à phlébotomes…).
La transmission vectorielle dépend des modifications environnementales qui sont dues
essentiellement aux changements climatiques locaux ou généraux et à l’Homme lui-même et
ses activités qui directement ou indirectement transforment la surface de la Terre en
n’épargnant aucun écosystème (pratiques de l’élevage et de l’agriculture, assèchement des
zones humides, déforestation, aménagement hydro-agricoles…).
Ces changements entrainent l’introduction d’un pathogène dans une région où le vecteur est
présent et /ou l’introduction de vecteurs dans une région préalablement indemne.
Les arboviroses sont pour la plupart des infections émergentes et dont la problématique est
reflétée par les résultats de nombreux travaux. Leur émergence est en partie liée à l’expansion
de plusieurs espèces de vecteurs (moustiques, phlébotomes, tiques).
Des organismes de recherche tels les Instituts Pasteurs, ont établi un inventaire des arbovirus
dont la circulation se faisait selon un mode enzootique mais qui sont devenus transmissibles à
l’Homme. Ce sont des virus qui ont franchi la barrière d’espèce en passant de l’animal à
l’Homme.
Les phlébovirus sont responsables chez l’Homme de maladies qui sont décrites depuis
longtemps mais dont la situation épidémiologique demeure mal connue. Ces virus circulent
dans tous les pays d’Europe et plus fréquemment dans ceux du bassin méditerranéen comme
c’est le cas de TOSV. Celui-ci est la cause d’un syndrome fébrile mais souvent d’une forme
grave, une infection neuro-invasive.
Peu de données sont disponibles sur la circulation des phlébovirus en Algérie et notamment
sur celle de TOSV. Ce virus et Leishmania peuvent coexister dans un même écosystème du
fait de leur vecteur commun qui est principalement Phlebotomus perniciosus dans le sud de la
France. Des études statistiques ont montré que les personnes exposées aux infections dues à
TOSV présentent plus de risque d’être infectées par les leishmanies.

Conclusion
99
Notre contribution à travers ce travail a été de mettre en évidence la co-circulation de
phlébovirus et de leishmanies à Draa El Mizan, une zone d’endémie à partir de l’étude du
vecteur et de la population pour établir le lien épidémiologique entre ces agents pathogènes.
Les résultats de la première partie de ce travail, consacrée à une étude entomologique, ont
montré la présence d’une population phlébotomienne importante dans cette région et assez
riche en espèces. Parmi ces espèces, certaines assurent le maintien de ce foyer endémique de
leishmanioses
Les résultats de la recherche de leishmanies chez les phlébotomes ont été négatifs. Cependant
et à partir de tests sérologiques par le WB, des cas de leishmaniose viscérale sous sa forme
asymptomatique ont été détectés chez des personnes vivant à Draa El Mizan et choisies
aléatoirement.
Les résultats de l’étude virologique ont révélé et pour la première fois à l’échelle nationale, la
présence du virus Toscana. La souche isolée chez les phlébotomes capturés dans la région
d’étude, cultivée puis identifiée après séquençage complet du génome, a été nommée TOSV-
Algérie 189 (souche lignée A). L’espèce (ou les espèces) de phlébotomes responsable de la
transmission virale n’a pas pu être identifiée. Par ailleurs des tests de séroneutralisation ont
révélé des cas positifs à TOSV avec la détection d’anticorps dirigés contre ce virus.
L’ensemble de ces résultats montre le rôle des phlébotomes dans la transmission de
leishmanies et du virus Toscana dans un même foyer et le risque encouru par la population de
Draa El Mizan de contracter ces deux pathogènes. Des cas de co-infections ont été détectés.
Les moyens de lutte contre les infections à TOSV et les leishmanioses font appel à la lutte
contre les phlébotomes en utilisant les insecticides, la réduction des gîtes et la protection
individuelle contre les piqûres (répulsifs cutanés, vêtements et moustiquaires imprégnés). La
promotion des mesures de lutte antivectorielle et de protection individuelle est donc
d’actualité.
Au vu des résultats obtenus, des études supplémentaires plus poussées sont à prévoir pour une
meilleure investigation, il serait ainsi nécessaire de :
- poursuivre cette recherche sur une période plus longue pour compléter les connaissances sur
le cycle de transmission de TOSV vu que les phlébotomes jouent à la fois le rôle de vecteur et
de réservoir du virus,
- identifier les espèces de phlébotomes vectrices;
- déterminer la spécialisation vecteur-virus pour estimer la circulation et la propagation des
virus
- élargir ce type d’étude à d’autres régions du pays connues comme foyers de leishmanioses.

Conclusion
100
Il faudrait aussi que les structures sanitaires des zones d’endémie mettent en œuvre le
diagnostic de TOSV chez les personnes présentant des accès fébriles pendant la saison d’été
et surtout chez celles souffrant d’infections du système nerveux central.

REFERENCES BIBLIOGRAPHIQUES

Références bibliographiques
101
A
Abonnenc E. (1972). Les phlébotomes de la région éthiopienne (Diptera, Psychodidae),
Ed O.R.S.T.O.M (France), 55: 1-289.
Alkan C., Bichaud L., De Lamballerie X., Alten B., Gould E.A., Charrel R.N. (2013).
Sandfly-borne phleboviruses of Eurasia and Africa: Epidemiology, genetic diversity,
geographic range, control measures. Antiviral Res, 100:54-74.
Alvar J., Canavate C., Gutierrez-solar B., Jimenez M., Laguna F., Lopez-velez R.,
Molina R., Moreno J. (1997). Leishmania and Human Immunodeficiency Virus
Coinfection : the First 10 Years. Clin. Microbiol. Rev. 10: 298-319.
Alvar J., Desjeux (2003). Leishmania/HIV co-infections: epidemiology in Europe Pages
3 15 : Published online: 18 Jul 2013
Alvar, J., Veılez, I.D., Bern, C., Herrero, M., Desjeux, P., Cano, J., Jannin, J., den
Boer,M. (2012). Leishmaniasis worldwide and global estimates of its incidence. PLosone
7, e35671
Aransay A.M., Scoulica E., Tselentis Y., Ready P.D. (2000). Phylogenitic relationships
of phlebotomine sandfies inferred from small subunit nuclear ribosomal DNA. Isect.
Molecular. Biology, 9:157-168.
B
Badaro R., Benson D., Eulalio M.C., Freire M., Cunha S., Netto ETTO E.M., Pedral-
sampaio D., Madureira C., Burns J.M., Houghton R.L., David J.R., Reed S.G.
(1996). RK 39: A cloned antigen of Leishmania chagasi that predicts active visceral
leishmaniasis. J. Inf. Dis, 173: 758-761.
Bahri O., Dhifallah I., Ben Alaya-Bouafif N., Fekih H., Gargouri J., Triki H. (2010).
Sero epidemiological study of West Nile virus circulation in human in Tunisia. Bull Soc
Pathol Exot. Oct, 104 (4):272.6.
Bahri, O., Fazaa, O., Ben Alaya-Bouafif, N., Bouloy, M., Triki, H., Bouattour, A.,
(2011). Role of Toscana virus in meningo-encephalitis in Tunisia. Pathol. Biol. (Paris) 59,
e125-e127
Baneth G., Koutinas AF., Solano-Gallego L., Bourdeau P., Ferrer L. (2008). Canine
leishmaniosis new concepts and insights on an expanding zoonosis.
Managing canine vector-borne diseases of zoonotic concern : part one. Trends Parasitol.
Jul, 24(7):324-30.
Basset D., Faraut F., Marty P., Dereure J., Rosenthal E., Mary C., Pratlong F.,
Lachaud L., Bastien P., Dedet JP. (2005).Visceral leishmaniasis in organ transplant
recipients: 11 new cases and a review of the literature. Microbes Infect. Oct, 7(13):1370-5.

Références bibliographiques
102
Belazzoug S. (1983). Isolation of Leishmania major Yakimoff & Schokhor, 1914 from
Psammomys obesus Gretzschmar, 1828 (Rodentia: Gerbillidae) in Algeria. Trans RSoc.
Trop. Med. Hyg, 77(6) : 876.
Belazzoug S. (1983). Le nouveau foyer de leishmaniose cutanée de M'sila (Algérie).
Infestation naturelle de Psammomys obesus (rongeur, Gerbillidae). Bull. Soc. Pathol.
Exot.76: 146-149
Belazzoug S. (1986a). Découverte d'un Meriones shawi (Rongeur, Gerbillidae)
naturellement infesté par Leishmania dans le nouveau foyer de leishmaniose cutanée de
Ksar Chellala (Algérie). Bull Soc Path Exot, 79: 630-633
Bellazoug S. (1986b). Les leishmanioses en Algérie. A propos de l’identification
enzymatique de 32 souches d’origine humaine et animale. In: Leishmania . Taxonomie et
phylogénese (Coll.int. CNRS/INSERM, 1984). Ed by Rioux J.A. Montpelier: IMEEE,
1986, 397- 400.
Belazzoug S. (1991). The sandflies of Algeria. Parasitologia, 33 (Suppl), 85- 87.
Benallal K., Benikhlef R., Garni R., Gassen B., Dedet J.P., Harrat Z. (2014). Presence
of Phlebotomus perniciosus atypical form in Algeria. Journal of Arthropod-Borne
Diseases.
Berchi S., Rioux J.A., Belmonte A., Russo J. (1986). Un phlébotome nouveau pour
l'Algérie, Phlebotomus (Praphlebotomus) kazeruni Theodor et Mesghali, 1964. Ann.
Parasit. Hum. Comp. 61(4): 507-508.
Berchi S. (1990). Ecologie des phlébotomes de l’Est Algérien. Thèse Magister Univ.
Constantine, Algérie, 129.
Berchi S. (1993). Les phlébotomes (Insecta, Diptera, Psychodidae), vecteurs de
leishmanioses dans l’Est algérien. Bull. Soc. Zool. Fr, 118, 3, 341-349.
Berdjane-Brouk Z., Charrel R. N., Bitam I., Hamrioui B. & Izri A. (2011). Record of
Phlebotomus (Transphlebotomus) mascittii Grassi, 1908 and Phlebotomus (Larroussius)
chadlii. In Rioux, Juminer & Gibily (1966). Female in Algeria. Parasite, 18(4), 337-339.
Bichaud L., Dachraoui K., Piorkowski G., Chelbi I., Moureau G., Cherni S et al.
(2013). Toscana virus isolated from sandflies, Tunisia. Emerg. Infect. Dis., 19:322-4.
Bogdan C., Röllinghoff M. (1998). The immune response to Leishmania: mechanisms of
parasite control and evasion. Int J Parasitol. Jan, 28(1):121-34.
Bogdan C., Stosiek N., Fuchs H., Rollinghoff M., Solbach W. (1990). Detection of
potentially leishmanial antigens by Western blot analysis of sera from patients with kala-
azar or multilesional cutaneous leishmaniasis J. Inf. Dis., 1990, 162, 1417-1418.

Références bibliographiques
103
Boubidi S.C., Benallal K., Boudrissa A., Bouiba L., Bouchareb B., Garni R.,
Bouratbine A., Ravel C., Dvorak V., Votypka J., Volf P., Harrat Z. (2011).
Phlebotomus sergenti (Parrot, 1917) identified as Leishmania killicki host in Ghardaia,
south Algeria. Microbes and Infection 13, 691-696.
Boudghene-strambouli O., Merad-boudia A. (1991). L’extension de la leishmaniose
cutanée en Algérie: 25 cas observés dans la wilaya de Tlemcen (Ouest Algérie. Bull. Soc.
Path. EX: 63-69
Boudrissa A., Cherif K., Kherrachi I., Benbetka S., Bouiba L., Boubidi S.C.,
Benikhlef R, Arrar L., Hamrioui B., Harrat Z. (2012). Extension de Leishmania major
au nord de l’Algérie. Spread of Leishmania major to the north of Algeria. Bull. Soc. Path,
105:30-3.
Bounamous A. (2010). Biosystématique et caractérisation par la biologie moléculaire des
phlébotomes de l’Est algérien. Thèse de Doctorat. Univ. de Constantine, 304.
Bounamous A., Boudabous R., Jouet D., Augot D., Ferté H., Babba H., Berchi S.,
Depaquit J. (2008). Caractérisation moléculaire et morphologique de deux espèces
affines de Paraphlebotomus: phlebotomus chabaudi in Croset, Abonnenc & Rioux, 1970
et P.riouxi Depaqui, Killck-Kendrick & Leger, 1998 (Diptra, Psychodidae). Parasite, 15,
565-571.
Bourabtine A., Aoun K., Gharb J., Harrat Z., Ezzedini M.S. et Etlijani S. (2005).
Spread of Leishmania killicki to Central and South-West Tunisia. Parasite, 12, 59-63.
Boussaa S., Guernaoui S., Pesson B., Boumezzough A. (2005). Seasonal fluctuations of
phlebotomine sandfly populations (Diptera: Psychodidae) in the urban area of Marrakech
Moroco. Acta Tropica, 95.
Bridgen A., Weber F., Fazakerley J.K. et al., (2001). Bunyamwera bunyavirus
nonstructural protein NSs is a nonessential gene product that contributes to viral
pathogenesis. Proc Natl Acad Sci U S A 98: 664–669
Brunhes J., Hassain K., Rhaim A., Hervy J.P. (2000). Les espèces de l’Afrique
méditerranéenne: Espèces présentes et répartition: (Diptera: Nematocera). Bull. Ent.
France, Extrait: 105, 195-204
Burchmore R.J., Barrett M.P. (2001). Life in vacuoles nutrient acquisition by
Leishmania amastigotes. Int J Parasitol. Oct, 31(12):1311-20.
C
Carhan A., Uyar Y., Ozkaya E., Ertek M., Dobler G., Dilcher M., Wang Y., Spiegel
M., Hufert F., Weidmann M. (2010). Characterization of a sandfly fever Sicilian virus
isolated during a sandfly fever epidemic in Turkey. J. Clin. Virol. 48, 264- 269

Références bibliographiques
104
Calisher C.H. (1986). History, classification, and taxonomy of viruses in the family
Bunyaviridae. In: R Elliot (Ed.).The Bunyaviridae. Plenum Press, New York.
Calisher C.H., Weinberg A.N., Muth D.J., Lazuick J.S. (1987). Toscana virus infection
in United States citizen returning from Italy. Lancet. Jan 17, 1(8525) :165-6.
Centre National d’Expertise sur les Vecteurs (C.N.E.V.). (2014). Collections de
références d’arthropodes vecteurs en France.
Charrel R., Izri M.A., Temmam S., Delaunay P., Toga I., Dumon H., Marty P., De
Lamballerie X., Parola P. (2007). Cocirculation of 2 genotypes of Toscana Virus,
Southeastern France. Emerging Infectious Diseases.
Charrel R.N., Moureau G., Temmam S., Izri, A., Marty P., Parola P., da Rosa A.T.,
Tesh R.B., de Lamballerie, X. (2009). Massilia virus, a novel Phlebovirus (Bunyaviridae)
isolated from sandflies in the Mediterranean. Vector Borne Zoonotic Dis. 9, 519-530.
Charrel R. (2014). Infection par le virus Toscana. EMC. Biologie médicale, 90-55-0183.
Cusi M.G., Savellini G.G., Zanelli G. (2010). Toscana virus epidemiology: from Italy
to beyond. Open Virol J 4, 22-28.
D
Dajoz R. (1985). Précis d’écologie. Ed. Dunod, Paris; 631p.
Dedet J.P., Addadi K. (1977). Epidémiologie des leishmanioses en Algérie. 4. Les
phlébotomes (Diptera, Psychodidae). Des Aurès. Arch. Enst. Past. Algérie. 52, 85-94.
Dedet J. P. (1999). Les leishmanioses. Ed. Ellipses, 253.
Dedet J. P., Addadi, K., Belazzoug, S. (1984). Les phlébotomes (Diptera, Psychodidae)
d'Algérie. Cahiers-O.R.S.T.O.M. Entomologie médicale et parasitologie, 22(2), 99-127.
Dedet J.P. (2001). Répartition géographique des leishmanioses. Méd. Mal. Infect., 31
Suppl 2 : 178-183.
Dedet J. P. (2009). Leishmanies, leishmanioses : biologie, clinique et thérapeutique. EMC.
Ed. Elsevier Masson SAS, Paris, Maladies infectieuses, 8, 506-510.
Depaquit J. (1997). Révision du sous genre Paraphlebotomus (Phlebotomus :
Phlebotominae: Diptera). Approches morphologique et moléculaire. Thèse, Univ. Reims,
Pharm. N° 211.
Depaquit J. (2014). Molecular systematics applied to Phlebotomine sandflies: review and
perspectives. Infect. Genet. Evol. Dec., 28:744-56.
Depaquit J., Ferté H., Léger N. (2000a). Révision du sous-genre Paraphlebotomus
(Phlebotomus-Phlebotominae Psychodidae: Diptera). Approche morphologique et
moléculaire. Ann. Pharm. Fr. 58: 333-340.

Références bibliographiques
105
Depaquit J., Ferté H., Léger N., Killick-Kendrick R., Rioux J.A., Killick-Kendrick
M., Hanafi H.A., Gobert S. (2000b). Molecular systematics of the Phlebotomine
sandflies of the subgenus Paraphlebotomus (Diptera: Psychodidae) and Phlebotomus)
based on ITS 2r DNA sequences. Hypotheses of dispersion and speciation. Insect
Molecular Biology, 9, 293-300.
Depaquit J., Grandadam M., Fouque F., Andry P.E., Peyrefitte C. (2010). Arthropod-
borne viruses transmitted by Phlebotomine sandflies in Europe: a review. Euro Surveill;
15: 19507.
Dereure J. (1993). Place du chien dans les complexes pathogènes leishmaniens des pays
du pourtour méditerranéen et du Moyen-Orient. (Algérie, Egypte, France, Maroc, Syrie,
Yemen).Thèse de Sciences, 1993, Faculté de Médecine Montpellier.
Desjeux P. (1991). Information sur l’épidémiologie des leishmanioses et la lutte contre
ces maladies par pays ou territoires. O.M.S, 30.
Desjeux P. (1999). Les leishmanioses. Aspect de santé publique et lutte. Ed. Ellipses, 253.
Dincer E., Gargari S., Ozkul A., Ergunay K. (2015). Potential animal reservoirs of
Toscana virus and coinfections with Leishmania infantum in Turkey. Am.J. Trop. Med.
Hyg., 92:690-7.
Dionisio D., Valassina M., Ciufolini M.G., Vivarelli A., Esperti F., Cusi M.G.,
Marchi, A., Mazzoli F., Lupi C. (2001). Encephalitis without meningitis due to sandfly
fever virus serotype Toscana. Clin Infect Dis 32, 1241-1243.
Djerbouh A., Toudjine M., Djoudi M., Benikhlef R., Harrat Z. (2005). La
leishmaniose canine en Algérie : essai de traitement par l’allopurinol. Ann. Med. Vet, 149,
132-134.
Dolmatova A.V., Demina N.A. (1971). Les phlébotomes (Phlebotominae) et les maladies
qu’ils transmettent. Cah. O.R.S.T.O.M Documentation Tech, 55, 289.
Dvorak V., Halada P., Hlavackova K., Dokianakis E., Antoniou M., Volf P. (2014).
Identification of phlebotominae sand flies (Diptera: Psychodidae) by matrix-assisted laser
desorption/ionization time of flight mass spectrometry. Parasites &Vector, 7:21. Ed.
Ellipses, 253.
E
Eddaikra N., Kherachi Djenad I., Benbetka S., Benikhlef R., Ait-Oudhia Kh.,
Moulti-Mati F., Bruno O., Denis S. & Harrat Z. (2016). Development of a Murine
Infection Model with Leishmania killicki, responsible for cutaneous leishmaniosis in
Algeria: Application in Pharmacology Biology and Medical Research International. 8 p
Elliott R.M. (2009). Bunyaviruses and climate change. Clin. Microbiol. Infect. 15: 510-
517.

Références bibliographiques
106
Elliott R.M. (1997). Emerging viruses: the Bunyaviridae. Mol. Med. 3: 572577.
Encyclopedic Médico-chirnrgicale (Ed. Elsevier, Paris). Maladies infectieuses, 10 :506-
508.
Ergunay K., Saygan M.B., Aydogan S., Lo M.M., Weidmann M., Dilcher M., Sener
B., Hascelik G., Pinar A., Us D. (2011). Sandfly fever virus activity in central/ northern
Anatolia, Turkey: first report of Toscana virus infections. Clin. Microbiol. Infect 17, 575–
581
Ergunay K, Erisoz Kasap O, Kocak Tufan Z, Turan MH, Ozkul A, Alten B. (2012).
Molecular evidence indicates that Phlebotomus major sensu lato (Diptera: Psychodidae) is
the vector species of the recently-identified sandfly fever Sicilian virus variant: sandfly
fever turkey virus. Vector Borne Zoonotic Dis. 12: 690–698.
F
Flippi C., Malherbe L., Julia V., Glaichenhaus N. (2001). L'immunité contre les
leishmanies. Médecine/Science 17(11): 1120-1128.
Fuller F., Bishop D.H. (1982). Identification of virus-coded nonstructural polypeptides in
bunyavirus-infected cells. J Virol,41: 643-648.
G
Gonzalez-Scarano F., Soldan S.S. (2011). Orthobunyavirus, Bunyaviridae. In: Darai
G., Konie S., Simniok J.et al. Springer index of viruses. Springer, Heidelberg.
H
Harrat Z., Addadi K., Belkaid M., Tabet-Derraz O. (1992). La leishmaniose viscérale
en Algérie: recensement des cas de leishmaniose (période 1985-1990). Bull.Soc. Pathol.
Exot, 85, 296-301.
Harrat Z., Benkherrouf K., Taharboucht Z., Bendali I.S., Yah T., Hamrioui B.
(1995). La leishmaniose canine urbaine. Arch. Inst. Past.Alger, 60, 157-165.
Harrat Z., Hamrioui B., Belkaid M. & Tabet-Derraz O. (1995). Point actuel sur
l'épidémiologie des leishmanioses en Algérie. Bull. Soc. Patho. Exot, 88. 180 -184.
Harrat Z., Pratlong F., Belazzoug S., Dereure J., Deniau M., Rioux J.A., Belkaid M.,
Dedet J.P. (1996). Leishmania infantum and L. major in Algeria. Trans. Roy. Soc. Trop.
Med. Hyg. 90,625-29.
Harrat Z., Belkaid M. 2002. Co-infection Leishmaniose viscérale-Sida en Algérie.
Algérie Santé-7. 37-38.

Références bibliographiques
107
Harrat Z., Boubidi S.C., Pratlong F., Benikhlef R., Selt B., Dedet J.P., Ravel C.,
Belkaid M. (2009). Description of Leishmania close to L.killicki (Rioux, Lanotte et
Pratlong, 1986) in Algeria.Trans. R. Soc. Trop. Med. and Hyg, 103 716-720.
Hollidge B.S., Gonzalez-Scarano F., Soldan S.S. (2010). Arboviral encephalitides:
transmission, emergence, and pathogenesis. J Neuroimmune Pharmacol 5: 428-542.
I
Izri M.A., Belazzoug S., Boudjebla Y., Dereure J., Pratlong F., Delalbre-Belmonte A.,
Rioux J.A. (1990). Leishmania infantum MON-1 isole de Phlebotomus perniciosus, en
Kabylie (Algérie). Ann. Parasitol. Hum. Comp, 65: 151-152.
Izri M. A., Marty P., Rahal A., Lelievre A., Ozon C., Baldelli G., Presiozo J., HaasP.,
Le Fichoux Y. (1992). Phlebotomus perniciosus Newstead, 1911 naturellement infesté
par des promastigotes dans la région de Nice (France).Bul. Soc. Path. Exot, 85, 385-387.
Izri M.A., Belazzoug S., Pratlong F., Rioux J.A. (1992). Isolement de Leishmania
major chez Phlebotomus papatasi à Biskra (Algérie). Fin d’une épopée
écoépidémiologique. Ann. Parasitol. Hum. Comp. 67: 31-32.
Izri M. A., Belazzoug S. (1993). Phlebotomus (Larroussius) perjiliewi naturally infected
with dermotropic Leishmania infantum at Tenes, Algeria. Tran. Royal. Soc.Trop. Med.
Hyg, 87, 399.
Izri A., Marty P., Fauran P., Le Fichoux Y., Rousset J. (1994). Phlebotomus perfiliewi
Parrot, 1930 (Diptera : Psychodidae) dans le Sud-Est de la France. Parasite 1 : 286.
Izri A., Marty P., Fauran P et al. (1996). Presumed vectors of leishmaniosis in the
principality of Monaco. Trans R Soc Trop Med Hyg 90-114.
Izri A., Depaquit J., Parola P. (2006). Phlébotomes et transmission d’agents pathogènes
autour du bassin méditerranéen. Med. Trop, 66:429-35.
Izri A., Temmam S., Moureau G., Hamrioui B., de Lamballerie X., Charrel R.N.
(2008). Sandfly fever Sicilian virus, Algeria. Emerg Infect Dis, 14:795-7.
Izri M.A., Bendjaballah A., Andriantsoanirina V. and Durand R. (2014). Cutaneous
leishmaniasis caused by Leishmania killicki, Algeria. Emerg Infect Dis. Mar, 20(3): 502–
504.
J
Janssen R., Gonzalez-Scarano F., Nathanson N. (1984). Mechanisms of bunyavirus
virulence. Comparative pathogenesis of a virulent strain of La Crosse and an avirulent
strain of Tahyna virus. Lab Invest 50: 447-455.
K

Références bibliographiques
108
Kang H.J., Bennett S.N., Hope A.G. et al. (2011). Shared ancestry between a newfound
mole-borne Hantavirus and Hantaviruses harbored by cricetid rodents. J Virol 85: 7496-
7503.
Konstantinou G.N, Papa A., Antoniadis A. (2007). Sandfly-fever outbreak in Cyprus:
are phleboviruses still a health problem. Travel Med Infect Dis, 5:239-42.
Khadem F., Uzonna J.E. (2014). Immunity to visceral leishmaniasis: implications for
immunotherapy. Future Microbiol. 9(7):901-15.
Killick Kendrick R. (1990). Phlebotomine vectors of leishmaniasis: Review. Med. Vet.
Entomol, 4, 1-24.
Killick-Kendrick R. (1985). Some epidemiological consequences of the evolutionary fit
between Leishmania and their phlebotomine vectors. Bull. Soc. Path. Exot, 78, 747-755.
Killick-Kendrick R., Tang Y., Killick-Kendrick M. (1991). The identification of female
sandflies of the subgenus Larroussius by the morphology of the spermathecal ducts.
Kocak Tufan Z., Weidmann M., Bulut C., Kinikli S., Hufert FT., Dobler G., Demiroz
A.P. (2011). Clinical and laboratory findings of a sandfly fever Turkey Virus outbreak in
Ankara. J. Infect. 2011 Nov; 63(5):375-81. doi: 10.1016/j.jinf.2011.07.011.
L
Lachaud L., Marchergui-Hammami S., Chabbert E., Dereure J., Dedet J. et Bastien.
(2000). Optimized PCR using patient blood for diagnosis and follow-up of visceral
leishmaniasis, with special reference to AIDS patients. Journal of Clinical Microbiology;
38: 236-240.
Le Houerou H.N. (1977). Etudes bioclimatiques des steppes algériennes (avec une carte
bioclimatiques à 1/1.000.000ème). Bull. Soc. Hist. Nat. Afr. Nord Alger, 68, 33-74.
Léger N., Pesson B., Madulo-Leblond G., Abonnenc E. (1983). Sur la différentiation
des femelles du sous-genre Larroussius Nitzulescu, 1931 (Diptera: Phlebotomidae) de la
région méditerranéenne. Ann. Parasitol. Hum. Comp, 58, 611-623.
Léger N., Depaquit J. (1999). Les phlébotomes .In: J-P. Dedet (ed.), les leishmanioses,
Paris : Ellipses : 90-108.
Léger N., Depaquit J. (2001). Les phlébotomes et leur rôle dans la transmission des
leishmanioses. Rev. Fr. labo, 338, 41-48.
Léger N., Depaquit J. (2001). Les phlébotomes et leur rôle dans la transmission des
leishmanioses. Rev. Fr. labo., 338, 41-48.

Références bibliographiques
109
Léger N., Depaquit J. (2002). Systématique et Biogeographie des phlébotomes (Diptera:
Psychodidae). Ann. Soc. Entomol. Fr. (n.s.), 38 (1-2), 163-175. Comparison of six PCR
methods using peripheral blood for detection of canine visceral leishmaniasis.
Levine N.D., Corliss J.O., Cox F.F.G., Deroux G., Grain J., Honigberg B.M., Leedale
G.F., Loeblich A.R., Lom J., Lynn D., Merinfeld E.G., Page F.C., Poljansky G.,
Sprague V., Vavra J. & Wallace F. G. (1980). A newly revised classification of the
Protozoa. J. Protozool. 27. 37-58.
Lewis D. J. (1973). Phlebotomidae and Psychodidae (sand-flies and moth-flies in insects
and other arthropods of medical importance. Edited by Kenneth G. V. Smith, London,
155-179.
Lewis D.J. (1982). A taxomonic review of genus Phlebotomus (Diptera: Psychodidae).
Bull. Brit. Mus. Nat. Hist. (Ent), 45. 121-209.
M
Mansouri R., Pratlong F., Bachi F., Hamrioui B., Dedet J.P. (2012). The first isoen-
zymatic characterizations of the Leishmania strains responsible for cutaneous
leishmaniasis in the Area os Annaba (Eastern Algeria). Open Conf. Proc. J. 3,6-11.
Marklewitz M., Handrick S., Grasse W et al. (2011). Gouleako virus isolated from
West African mosquitoes constitutes a proposed novel genus in the family Bunyaviridae. J
Virol 85: 9227-9234.
Marty C., Lamouroux D., Dunan S., Quilici M. (1992). Western blot analysis of
antibodies to Leishmania infantum antigens. Potential of the 14-kD and 16-kD antigens
for diagnosis and epidemiologic purposes. Am. J. Trop. Med. Hyg., 47, 764-771.
Marty P., Leleivre A., Quaranta J.F., Suffia I., Eulalio M., Garitoussaint M., Le
fichoux Y., Kubar J. (1995). Detection by Western blot of four antigens characterizing
acute clinical leishmaniasis. Trans. R. Soc. Trop. Med. Hyg. 89, 690-691.
Mir M.A, Panganiban A.T. (2006). The bunyavirus nucleocapsid protein is an RNA
chaperone: possible roles in viral RNA panhandle formation and genome replication.
RNA 12: 272-282.
Moureau G., Bichaud, L., Salez N., Ninove L., Hamrioui B., Belazzoug S.,
deLamballerie X., Izri A., Charrel R.N. (2010). Molecular and serological evidence for
the presence of novel phleboviruses in sandflies from northern Algeria.Open Virol. J. 4,
15-21.
Moulahem T., Fendri A.H., Harrat Z., Benmezdad A., Aissaoui K. (1998).
Contribution à l’étude des phlébotomes de Constantine : espèces capturées dans un
appartement urbain. Bull Soc Pathol Exot. 91 (4) 344-345.

Références bibliographiques
110
Mouloua A., Boubidi S.C., Bouiba L., Mezai G., Madiou M., Harrat Z. (2017). Impact
environnemental sur la répartition des leishmanioses dans le foyer de Tizi-Ouzou
(Algérie). Revue Méd. Vét., 168, 10-12, 252-261.
Mutiso J.M., Macharia J.C., Kiio M.N., Ichagichu J.M., Rikoi H., Gicheru M.M.
(2013). Development of Leishmania vaccines: predicting the future from past and present
experience. J Biomed Res. Mar, 27(2):85-102.
N
Niang A.A., Geoffroy B., Angel G., Trouillet J., Killik-Kendrick R., Hervy J-
P.,Brunhes J. (2000). Les phlébotomes de l'Afrique de l'Ouest. Logiciel d'identification et
d'enseignement, IRD Edition.
Nichol S.T., Beaty B.J., Elliot R.M et al. (2005). Bunyaviridae. In: CM Fauquet, MA
Mayo, J Mailoff et al. (Eds).Virus taxonomy eight: Report of the International Committee
on Taxonomy of Viruses. Academic, Amsterdam.
O
O.M.S. (1990). World Health Organization. Control of the leishmaniases. World Health
Organ Tech Rep Ser, 793: 1-177.
O.M.S. (2010). Control of the Leishmaniases: Report of a Meeting of the WHO Expert
Commitee on the Control of Leishmaniases. World Health Organ Tech Rep Ser949
Geneva.
O.M.S. (2017). Leishmaniose. Aide-mémoire n°375.
P
Palacios G., Tesh R.B., Savji N., Travassos da Rosa A.P., Guzman H., Bussetti A.V.,
Desai A., Ladner J., Sanchez-Seco M., Lipkin W.I. (2014). Characterization of the
Sandfly fever Naples species complex and description of a new Karimabad species
complex (genus Phlebovirus, family Bunyaviridae). J. Gen. Virol. Feb.;95(Pt 2):292-300.
Papa A., Konstantinou G., Pavlidou V., Antoniadis A. (2006). Sandfly fever virus
outbreak in Cyprus. Clin. Microbiol. Infect., 12:192-4.
Papa A., Paraforou T., Papakonstantinou I., Pagdatoglou K., Kontana A.,
Koukoubani T. (2014). Severe encephalitis caused by Toscana virus, Greece.Emerg
Infect Dis ,20:1417-9.Parasitologia, 33: 335-347.
Pearson R.D., Sousa A.Q. (1996). Clinical spectrum of Leishmaniasis. Clin Infect Dis,
22(1),1-13.

Références bibliographiques
111
Perrone L.A., Narayanan K., Worthy M et al. (2007). The S segment of Punta Toro
virus (Bunyaviridae, Phlebovirus) is a major determinant of lethality in the Syrian hamster
and codes for a type I interferon antagonist. J. Virol. 81: 884-892.
Porterfield J.S .(1975). The basis of arbovirus classification. Med. Biol. 53: 400-405.
Puentes S.M., Da Silva R.P., Sacks D.L. Hammer C.H., Joiner K.A. (1990). Serum
resistance of metacyclic stage Leishmania major promastigotes is due to relaese of C5b-9.
J.Immunol.145 (12): 4311-4316.
Pratlong F., Dedet J.P, Marty P., Portús M., Deniau M., Dereure J., Abranches P.,
Reynes J., Martini A., Lefebvre M. et al. (1995). Leishmania-human immunodeficiency
virus coinfection in the Mediterranean basin: isoenzymatic characterization of 100 isolates
of the Leishmania infantum complex. ). J. Infect. Dis., Jul;172 (1):323-6. PMID: 7797943.
Punda-Polic V., Mohar B., Duh D., Bradaric N., Korva M., Fajs L., Saksida A.,
Avsic-Zupanc T. (2012). Evidence of an autochthonous Toscana virus strain in Croatia. J
Clin Virol; 55, 4-7.
R
Ready P.D. (2013). Biology of phlebotomine sand flies as vectors of disease agents.
Annu. Rev. Entomol. 58, 227-250.
Reithinger R., Dujardin J.C., Louzir H et al. (2007). Cutaneous leishmaniasis. Lancet
Infect Dis 7,581-96.
Rioux J. A., Croset H., Juminer B. (1969a). Présence en Tunisie de Phlebotomus
alexandri (Sinton, 1928) Sergentomyia clydei (Sinton, 1928) et Sergentomyia dryfussi
(Parrot, 1933). Ann. Parasitol. Hum. Comp. 44, 825-826.
Rioux J.A., Golvan Y.J., Croset H., Tour S., Houin R., Abonnenc C., Petitdidier M.,
Vollhardt Y., Dedet J.P., Albaret J.L., Lanotte G., Quilici M. (1969b). Epidémiologie
des leishmanioses dans le sud de la France. Monographie I.N.S.E.R.M. 37, 223.
Rioux J.A, Akalay O., Perieres J., Dereure J., MAhjour J., Lehouerou H.N., Leger N.,
Desjeux P., Gallego M., Saddiki A et al. (1997). L’évolution éco-épidémiologique du
risque leishmanien au Sahara atlantique marocain. Intérêt heuristique de la relation
phlébotomes bioclimats. Ecole Méditerranée, 23, 73-92.
Rioux J.A., Ashford R.W., Khiami A. (1992). Ecoepidemiology of leishmaniases in
Syria. 3. Leishmania major infection in Psammomys obesus provides clues to life history
of the rodent and possible control measures. Ann. Parasitol. Hum. Comp., 67(6):163-5.
Rioux J.A., Golvan Y.J., Croset H., Houin R., Juminer B., Bain O., Tour S. (1967).
Ecologie des leishmanioses dans le Sud de France. Echantillonnage. Ethologie. Ann.
Parasitol. Hum. Comp., 42: 561-603.

Références bibliographiques
112
Rioux J.A., Lanotte G. (1993). Apport de la cladistique à l’analyse du genre Leishmania
Ross, 1903 (Kinetoplastida: Trypanosomatidae). Corollaires épidémiologiques.
Biosystema 1993, 8 : 79-80.
Rioux J.A., Lanotte G., Petter F., Dereure J., Akalay O., Pratlong F., Velez G., Fikri
N.B., Maazoun R., Denial M. (1986). Les leishmanioses cutanées du bassin
méditerranéen occidental: de l’identification enzymatique à l’analyse éco-
épidémiologique, l’exemple de trois ‘foyers’, tunisien, marocain et français, 365-395 in:
Rioux J.A. Leishmania. Taxonomie et phylogénèse. Applications écoépidémiologiques.
Montpellier, France: Institut Méditerranéen d’Etudes Épidémiologiques et Ecologiques.
Rioux J.A., Lanotte G., Serres E., Pratlong F., Bastien P., Perières J. (1990).
Taxonomy of Leishmania. Use of isoenzymes. Suggestions for a new classification. Ann.
Parasitol. Hum. Comp;, 65, 111-115.
Rioux JA., Ashford RW., Khiami A. (1991). Ecoepidemiology of leishmaniases in Syria.
3. Leishmania major infection in Psammomys obesus provides clues to life history of the
rodent and possible control measures. Ann. Parasitol. Hum. Comp., 67(6):163-5
Rittig M.G., Bogdan C. (2000). Leishmania-host-cell interaction: complexities and
alternative views. Parasitol Today. Jul, 16(7):292-7.
Rodhain F. (2001). Fièvre jaune, dengue et autres arboviroses. EMC . Maladies
infectieuses ; 8-062-A-10, 19.
Rolland L., Belkaid M., Seye A., Schneider P., Gentilini M. (1995). Detection of serum
antibodies against Leishmania 94 kDa antigen in visceral and cutaneous leismaniosis due
to L. infantum. Parasite, 2, 13-21.
S
Sacks D.l., Modi G., Rowton E., Späth G., Epstein l., Turco S.J., Beverley S.M.
(2000). The role of phosphoglycans in Leishmania-sand fly interactions. Proc Natl Acad
Sci U S A., 97(1), 406-11.
Sacks D.L., Pimenta P.F., Mc Conville M.J., Schneider P., Turco S.J .(1995). Stage-
specific binding of Leishmania donovani to the sand fly vector mid gut is regulated by
conformational changes in the abundant surface lipophosphoglycan. J. Exp. Med. 181(2):
685-697.
Sakhria S., Bichaud L., Mensi M., Salez N., Dachraoui K., Thirion L. et al. (2013). In
Co-Savji N., Palacios G., Travassos da Rosa A et al. (2011). Genomic and phylogenetic
characterization of Leanyer virus, a novel Orthobunyavirus isolated in northern Australia.
J. Gen. Virol. 92: 1676-1687.

Références bibliographiques
113
Sánchez-Seco M. P., Echevarria J. M., Hermndez L., Estévez D., Navarro-Mari J.
M., Tenorio A. (2003). Detection and identification of Toscana and other phleboviruses
by RT-nested-PCR assays with degenerated primers. J. Med. Virol. 71, 140–149.
Sánchez-Seco M.P., Navarro J.M. (2005). Infections due to Toscana virus, West Nile
virus, and other arboviruses of interest in Europe. Enferm. Infec. Microbiol. Clin. Nov., 23
(9) :560-8.
Sanbonmatsu-Gamez S., Perez Ruiz M. et al., (2005). Toscana virus in Spain. Emerg
Infect Dis.; 11(11):1701-7.
Sanger F., Nicklen S., Coulson A.R. (1977). DNA sequencing with chain-terminating
inhibitors. Proc. Natl. Acad. Sci. U S A. Dec; 74(12):5463-7.
Savji N., Palacios G., Travassos da Rosa A. et al. (2011). Genomic and phylogenetic
characterization of Leanyer virus, a novel orthobunyavirus isolated in northern Australia.
J. Gen. Virol. 92: 1676–1687.
Schmaljohn C. (1996). The viruses and their replication. In: Fields B.N., Knipe D.M.,
Howley P.M (Eds.), Fields virology. Lippincott-Raven, Philadelphia.
Sergent E.D.M., Sergent E.T., Parrot L., Donatien A., Beguet M. (1921). Transmission
du Clou de Biskra par le phlébotome (Phlebotomus papatasi). C.R. Acad. Sci. Paris 173,
1030.
Shi X., Kohl A., Leonard V.H et al. (2006). Requirement of the N-terminal region of
0rthobunya virus non structural protein NSm for virus assembly and morphogenesis. J.
Virol. 80: 8089-8099.
Soldan S.S., Gonzalez-Scarano F. (2005). Emerging infectious diseases: the
Bunyaviridae. J. Neurovirol. 11: 412–423.
Soldan S.S., Plassmeyer M.L., Matukonis M.K et al. (2005). La Crosse virus
nonstructural protein NSs counteracts the effects of short interfering RNA. J. Virol. 79:
234-244.studies on the epidemiology of sandfly fever in the Old World. Bull World
Health Organ, 54:663-74.
Soldan. S., Gonza Lez-Scarano F., (2014). The Bunyaviridae. Handbook of Clinical
Neurology, Vol. 123 (3rd series). Neurovirology A.C. Tselis and J. Booss, Ed. © Elsevier
B.V.
T
Takeda A., Sugiyama K., Nagano H. et al. (2002). Identification of a novel RNA
silencing suppressor, NSs protein of Tomato spotted wilt virus. FEBS Lett 532: 75-79.

Références bibliographiques
114
Tamura K., Peterson D., Peterson N., Stecher G., Nei M, Kumar S. (2011). MEGA5:
molecular evolutionary genetics analysis using maximum likelihood, evolutionary
distance, and maximum parsimony methods. Mol. Biol. Evol. Oct. 28(10):2731-9.
Tesh R.B., Saidi S., Gajdamovic J.J., Rodhain F., Vesenjak Hirjan J. (1976).
Serological studies on the epidemiology of sandfly fever in the Old World. Bull World
Health Organ 54: 663-674.
V
Verani P., Nicoletti L. Ciufolini M.G., (1984). Antigenic and biological characterization
of Toscana virus, a new Phlebotomus fever group virus isolated in Italy. Acta Virol. 28,
39-47.
W
Weber F., Dunn E.F., Bridgen A. et al. (2001). The Bunyamwera virus nonstructural
protein NSs inhibits viral RNA synthesis in a minireplicon system. Virology 281: 67-74.
Who, 1990;
Won S., Ikegami T., Peters C.J. et al. (2007). NSm protein of Rift Valley fever virus
suppresses virus-induced apoptosis. J. Virol. 81: 13335-13345.
Y
Yu X.J., Liang M.F., Zhang S.Y et al. (2011). Fever with thrombocytopenia associated
with a novel Bunyavirus in China. N. Engl. J Med. 364: 1523-1532. Jan, 175(1):14-5.

ANNEXES

Classe Test (N) Test (P) Totaux (Ligne)
Effectif C1 20 4 24
%age Colonne
7,04% 4,65%
%age Ligne
83,33% 16,67%
%age Total 5,41% 1,08% 6,49%
Effectif C2 119 36 155
%age Colonne
41,90% 41,86%
%age Ligne
76,77% 23,23%
%age Total 32,16% 9,73% 41,89%
Effectif C3 79 18 97
%age Colonne
27,82% 20,93%
%age Ligne
81,44% 18,56%
%age Total 21,35% 4,86% 26,22%
Effectif C4 66 28 94
%age Colonne
23,24% 32,56%
%age Ligne
70,21% 29,79%
%age Total 17,84% 7,57% 25,41%
Effectif Ts Grpes 284 86 370
%age Total 76,76% 23,24%
Classe Test (N) Test (P) Totaux (Ligne)
C1 18,4216 5,57838 24,0000
C2 118,9730 36,02703 155,0000
C3 74,4541 22,54595 97,0000
C4 72,1514 21,84865 94,0000
Ts Grpes 284,0000 86,00000 370,0000
Annexe I
L’analyse statistique effectuée par le test Khi2 sur les résultats du test de Western-Blot
Khi2 de Pearson: 4.0323, dl=3, p=0.2579

Classe de taille
Test (N)
Test (P)
Totaux (Ligne)
Effectif C1 22 0 22
%age Colonne
6,43% 0,00%
%age Ligne
100,00% 0,00%
%age Total 5,99% 0,00% 5,99%
Effectif C2 148 6 154
%age Colonne
43,27% 24,00%
%age Ligne
96,10% 3,90%
%age Total 40,33% 1,63% 41,96%
Effectif C3 87 10 97
%age Colonne
25,44% 40,00%
%age Ligne
89,69% 10,31%
%age Total 23,71% 2,72% 26,43%
Effectif C4 85 9 94
%age Colonne
24,85% 36,00%
%age Ligne
,43%
9,57%
%age Total 23,16% 2,45% 25,61%
Effectif Ts Grpes 342 25 367
%age Total
93,19% 6,81%
Annexe II
L’analyse statistique effectuée par le test Khi2 sur les résultats de la séroneutralisation

Classe Test (N) Test (P) Totaux (Ligne)
Effectif C1 21 3 24
%age Colonne
12,73% 1,46%
%age Ligne
87,50% 12,50%
%age Total 5,68% 0,81% 6,49%
Effectif C2 80 75 155
%age Colonne
48,48% 36,59%
%age Ligne
51,61% 48,39%
%age Total 21,62% 20,27% 41,89%
Effectif C3 39 58 97
%age Colonne
23,64% 28,29%
%age Ligne
40,21% 59,79%
%age Total 10,54% 15,68% 26,22%
Effectif C4 25 69 94
%age Colonne
15,15% 33,66%
%age Ligne
26,60% 73,40%
%age Total 6,76% 18,65% 25,41%
Effectif Ts Grpes 165 205 370
%age Total
44,59% 55,41%
Classe Test (N) Test (P) Totaux (Ligne)
Effectif C1 22 2 24
%age Colonne
11,06% 1,17%
%age Ligne
91,67% 8,33%
%age Total 5,95% 0,54% 6,49%
Effectif C2 90 65 155
%age Colonne
45,23% 38,01%
%age Ligne
58,06% 41,94%
%age Total 24,32% 17,57% 41,89%
Effectif C3 50 47 97
%age Colonne
25,13% 27,49%
%age Ligne
51,55% 48,45%
%age Total 13,51% 12,70% 26,22%
Effectif C4 37 57 94
%age Colonne
18,59% 33,33%
%age Ligne
39,36% 60,64%
%age Total 10,00% 15,41% 25,41%
Effectif Ts Grpes 199 171 370
%age Total
53,78% 46,22%

ORIGINAL ARTICLE VIROLOGY
Virus isolation, genetic characterization and seroprevalence of Toscanavirus in Algeria
C. Alkan1,2, A. B. Allal-Ikhlef3, S. Alwassouf1,2, A. Baklouti1,2, G. Piorkowski1,2, X. de Lamballerie1,2, A. Izri4 and R. N. Charrel1,2
1) Aix Marseille Université, IRD French Institute of Research for Development, EHESP French School of Public Health, EPV UMR_D 190 “Emergence des
Pathologies Virales”, 2) IHU Mediterranee Infection, APHM Public Hospitals of Marseille, Marseille, France, 3) Département de Biologie, Faculté des Sciences de la
Nature et de la Vie, Université d’Oran 1, Ahmed Ben Bella, Algeria and 4) Service de Parasitologie-Mycologie, Hôpital Avicenne, Université Paris, Bobigny, Paris,
France
Abstract
Toscana virus (TOSV; Bunyaviridae, Phlebovirus) is transmitted by sandflies of the genus Phlebotomus in the Mediterranean area. One strain of
TOSV was isolated from a total of almost 23 000 sandflies collected in Kabylia, Algeria. The complete genome was sequenced, and
phylogenetic studies indicated that it was most closely related with TOSV strain from Tunisia within lineage A, which also includes Italian,
French and Turkish strains. A seroprevalence study performed on 370 sera collected from people living in the same area showed that
almost 50% possessed neutralizing antibodies against TOSV, a rate much higher than that observed in Southern Europe. Sandfly species
distribution in the study area suggests that the vector of TOSV in this region belongs to the subgenus Larroussius. These data support the
rapid implementation of the diagnosis of TOSV in clinical microbiology laboratories to estimate the burden in patients presenting with
neuroinvasive infections and febrile illness.
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All
rights reserved.
Keywords: Complete genome sequencing, microneutralization, Phlebovirus, seroprevalence, Toscana virus, vector-borne infection
Original Submission: 18 May 2015; Revised Submission: 22 June 2015; Accepted: 13 July 2015
Editor: T. Av�sic-Zupanc
Article published online: XXX
Corresponding author: R.N. Charrel, UMR 190 Unité des VirusÉmergents, Faculté de Médecine, 27 Bvd Jean Moulin, 13005 Marseille,FranceE-mail: [email protected]. Alkan and A.B. Allal-Ikhlef contributed equally to this article,and both should be considered first author. A. Izri and R.N.Charrel contributed equally to this article and both should beconsidered last author.
Introduction
Toscana virus (TOSV; Bunyaviridae, Phlebovirus) is an enveloped
virus, with three segments of single-stranded, negative-senseRNA which is transmitted by phlebotomine sandflies. It was
first isolated from Phlebotomus perniciosus in central Italy in 1971[1]. The first evidence that TOSV can cause disease in humans
was reported 12 years later when the virus was detected in thecerebrospinal fluid of two patients with meningitis after they
vacationed in Portugal and Italy [2,3]. Subsequent studiesshowed that TOSV was the main cause of meningitis in Central
Italy during summertime, before enteroviruses, mumps andherpesviruses [4–7]. TOSV was also identified as among the
three main causes of meningitis during summertime in Spain andFrance [8], along with enteroviruses and herpesviruses. After-wards, TOSV human cases were also diagnosed by direct
methods (such as virus isolation and/or molecular detection) inGreece, Turkey and Croatia [9–11]. Recently, direct and in-
direct evidence have indicated that TOSV is present in Malta,Cyprus, Bosnia-Herzegovina, Kosovo, Tunisia and Morocco
[12–16].At the outset of this study, there were no data from Algeria
except for seroprevalence studies and Phlebovirus sequencedetection (distinct from TOSV) [17–19]. Using an approach
Clin Microbiol Infect 2015; -: 1.e1–1.e9Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reserved
http://dx.doi.org/10.1016/j.cmi.2015.07.012

ORIGINAL ARTICLE VIROLOGY
Virus isolation, genetic characterization and seroprevalence of Toscanavirus in Algeria
C. Alkan1,2, A. B. Allal-Ikhlef3, S. Alwassouf1,2, A. Baklouti1,2, G. Piorkowski1,2, X. de Lamballerie1,2, A. Izri4 and R. N. Charrel1,2
1) Aix Marseille Université, IRD French Institute of Research for Development, EHESP French School of Public Health, EPV UMR_D 190 “Emergence des
Pathologies Virales”, 2) IHU Mediterranee Infection, APHM Public Hospitals of Marseille, Marseille, France, 3) Département de Biologie, Faculté des Sciences de la
Nature et de la Vie, Université d’Oran 1, Ahmed Ben Bella, Algeria and 4) Service de Parasitologie-Mycologie, Hôpital Avicenne, Université Paris, Bobigny, Paris,
France
Abstract
Toscana virus (TOSV; Bunyaviridae, Phlebovirus) is transmitted by sandflies of the genus Phlebotomus in the Mediterranean area. One strain of
TOSV was isolated from a total of almost 23 000 sandflies collected in Kabylia, Algeria. The complete genome was sequenced, and
phylogenetic studies indicated that it was most closely related with TOSV strain from Tunisia within lineage A, which also includes Italian,
French and Turkish strains. A seroprevalence study performed on 370 sera collected from people living in the same area showed that
almost 50% possessed neutralizing antibodies against TOSV, a rate much higher than that observed in Southern Europe. Sandfly species
distribution in the study area suggests that the vector of TOSV in this region belongs to the subgenus Larroussius. These data support the
rapid implementation of the diagnosis of TOSV in clinical microbiology laboratories to estimate the burden in patients presenting with
neuroinvasive infections and febrile illness.
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All
rights reserved.
Keywords: Complete genome sequencing, microneutralization, Phlebovirus, seroprevalence, Toscana virus, vector-borne infection
Original Submission: 18 May 2015; Revised Submission: 22 June 2015; Accepted: 13 July 2015
Editor: T. Av�sic-Zupanc
Article published online: 30 July 2015
Corresponding author: R.N. Charrel, UMR 190 Unité des VirusÉmergents, Faculté de Médecine, 27 Bvd Jean Moulin, 13005 Marseille,FranceE-mail: [email protected]. Alkan and A.B. Allal-Ikhlef contributed equally to this article,and both should be considered first author. A. Izri and R.N.Charrel contributed equally to this article and both should beconsidered last author.
Introduction
Toscana virus (TOSV; Bunyaviridae, Phlebovirus) is an enveloped
virus, with three segments of single-stranded, negative-senseRNA which is transmitted by phlebotomine sandflies. It was
first isolated from Phlebotomus perniciosus in central Italy in 1971[1]. The first evidence that TOSV can cause disease in humans
was reported 12 years later when the virus was detected in thecerebrospinal fluid of two patients with meningitis after they
vacationed in Portugal and Italy [2,3]. Subsequent studiesshowed that TOSV was the main cause of meningitis in Central
Italy during summertime, before enteroviruses, mumps andherpesviruses [4–7]. TOSV was also identified as among the
three main causes of meningitis during summertime in Spain andFrance [8], along with enteroviruses and herpesviruses. After-wards, TOSV human cases were also diagnosed by direct
methods (such as virus isolation and/or molecular detection) inGreece, Turkey and Croatia [9–11]. Recently, direct and in-
direct evidence have indicated that TOSV is present in Malta,Cyprus, Bosnia-Herzegovina, Kosovo, Tunisia and Morocco
[12–16].At the outset of this study, there were no data from Algeria
except for seroprevalence studies and Phlebovirus sequencedetection (distinct from TOSV) [17–19]. Using an approach
Clin Microbiol Infect 2015; 21: 1040.e1–1040.e9Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reservedhttp://dx.doi.org/10.1016/j.cmi.2015.07.012

combining (a) an entomologic campaign for the capture of
sandflies, (b) the direct detection of viruses relying on molec-ular techniques and cell culture and (c) a seroprevalence study
in local human populations using neutralization assay, weinvestigated the current situation in Algeria. Similar research
programs were previously implemented in other regions andprovided invaluable data [12,20,21]. We present here the re-sults obtained in northern Algeria in the region of Kabylia, a
well-known area for the circulation of sandflies and endemic forleishmaniasis [17].
Materials and methods
Study site, sandfly trapping and identificationSandfly trapping campaigns were conducted during 17–23August 2013 in Draa El Mizan (36° 320 1460’N, 3° 500 85000 E) inthe Kabylia region of Algeria at an average altitude of 380 musing CDC Miniature Light Traps as previously reported [17].
The collection sites were chosen on the basis of the epidemi-ologic data provided by the health services (358 recorded cases
of confirmed cutaneous leishmaniasis since 1990) and thepresence of domestic animals [22]. Draa El Mizan is situated at
the west side of the Djurdjura Mountains, is surrounded by hillsand experiences a Mediterranean climate with plenty of wa-terways and dams. The population was 38 844 according to the
2008 General Population and Housing Census. Vegetationconsists of oak, olive trees and fig trees. Domestic animals (cats,
dogs, goats, sheep, rabbits and chickens) are frequently shel-tered in basements or near human housings. In addition, there
are many government-subsidized beef cattle livestock lots, thesurroundings of which are the most productive for the captures
of sandflies. A total of 22 998 sandflies were trapped. Speciesidentification was only performed for a random subset of 1500captured sandflies using morphologic keys [23,24] because it is
a time-consuming process (>1 minute per sandfly) which de-creases the chances of virus detection and isolation. The
remaining 21 498 sandflies were pooled and stored at −80°Cfor virus detection and isolation.
Virus detectionPools of sandflies were homogenized in 600 μL of Eagleminimal essential medium (EMEM) as previously reported
[20,21]. A 200 μL volume of the homogenized pool was usedfor viral nucleic acid extraction with the BioRobot EZ1-XL
Advanced (Qiagen) using the Virus Extraction Mini Kit (Qia-gen); 5 μL of nucleic acid were used for RT-PCR and nested
PCR assays with primers targeting the polymerase gene andthe nucleoprotein gene using protocols previously described
[25,26]. PCR products of the expected size were column
purified (Amicon Ultra Centrifugal filters, Millipore) and
directly sequenced.
Virus isolationA 50 μL volume of homogenized sandfly pools was inoculated
onto a 12.5 cm2flask of Vero cells as previously reported [20].
The flasks were incubated at 37°C in a 5% CO2 atmosphere
and examined daily for cytopathic effects.
Complete genome sequencingTOSV passage 0 was used for complete genome characteriza-tion through next-generation sequencing (NGS), as previously
described [20]. Virus sequences were reconstructed from theNGS reads based on the best BLAST (Basic Local AlignmentSearch Tool) similarity against reference databases. NGS reads,
of minimum length 30 nucleotides, were trimmed using CLCGenomic Workbench 6.5, with a minimum of 99% quality per
base and mapped to reference sequences (TOSV, Tunisianstrain, GenBank accession nos. JX867534, JX867535 and
JX867536 for the L, M and S segments, respectively). Parame-ters were set such that each accepted read had to map to the
reference sequence for at least 50% of its length, with a mini-mum of 80% identity to the reference. Gaps in the genomewere sequenced from overlapping PCR products amplified by
sequence-based designed primers, using either Sanger directsequencing or NGS. The 50 and 30 extremities of each segment
were sequenced using a primer including the 8 nt conservedsequence common to sandfly-borne phleboviruses as previ-
ously described [27]. Ultimate verification of the sequence wasdone by Sanger sequencing of overlapping PCR products
covering the entire genome.
Genetic distances and phylogenetic analysisThe complete amino acid sequences of S (N and Ns genes), M
(Gn and Gc genes) and L (polymerase) segments of the TOSVstrain isolated in this study were aligned together with ho-
mologous sequences of other strains of TOSV retrieved fromthe GenBank database (until April 2015). Nucleotide and amino
acid sequences were aligned using the CLUSTAL algorithm ofMEGA 5 software [28]. Neighbor-joining analysis (Kimura 2-
parameter and p-distance models) was performed by MEGA5, with 1000 bootstrap pseudoreplications.
Microneutralization-based (MN) seroprevalence studyHuman sera were collected in Draa El Mizan and were used inagreement with the Algerian regulations for such studies. The
MN assay was adapted from a previously described protocol[20]. Briefly, twofold serial dilutions from 1:10 to 1:160 were
prepared for each serum, and a volume of 50 μL of eachdilution was transferred into 96-well plates. A volume of 5 0μL
CMI Alkan et al. Toscana virus in Algeria 1040.e2
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reserved, CMI, 21, 1040.e1–1040.e9

containing either 1000 tissue culture infectious dose that will
infect 50% if cell monolayers are challenged with definedinoculum (TCID50) or 100 TCID50 (two independent assays) of
TOSV (strain MRS2010-4319501) was added to each wellexcept for the controls, which contained PBS. The plates were
incubated at 37°C for 1 hour. Then a 100 μL suspension ofVero cells containing approximately 2 × 105 cells/mL of EMEMmedium enriched with 5% fetal bovine serum, 1% penicillin-
streptomycin, 1% L-glutamine 200 mM, 1% kanamycin and 3%Fungizone was added to each well and incubated at 37°C in the
presence of 5% CO2. The first row of each plate containedcontrol sera diluted 1:10 and Vero cells without virus. After 5
days, the microplates were read under an inverted microscope,and the presence (neutralization titre at 10, 20, 40, 80 and 160)
or absence (no neutralization) of cytopathic effect was noted.Cutoff values to consider a serum as positive were set asdilution 1:20 for the 100 TCID50 and dilution 1:10 for the 1000
TCID50.The geometric mean of the titres for TOSV MN ac-cording to age groups was calculated. To attempt to determine
which criteria were best, we calculated the Pearson correlationbetween results observed using 100 and 1000 TCID50 viral
inoculums using IBM SPSS Statistics 21.0 software. The variationof seroprevalence rates among age groups was also calculated
by the chi-square test.
Results
Sandfly trapping and virus detectionA total of 22 998 sandflies were collected during 17–23 August
2013 from Draa El Mizan. A total of 1500 flies were identifiedmorphologically. Five species were identified, as follows:
P. perfiliewi (n = 772, 51.4%), P. perniciosus (n = 550, 36.7%),P. longiscupis (n = 40, 2.6%), P. papatasi (n = 97, 6.5%) andS. minuta (n = 35, 2.3%). All the remaining 21,498 sandflies were
organised into 247 pools. Pool 189 was positive with primersSFN-1S and SFN-1R [25]. The corresponding 443 nt sequence
in the N gene was most closely related with the homologousregion of the TOSV Tunisian strain T152 (GenBank accession
no. JX867536) sequence (96% identity at the nt level).
Virus isolationVero cells that were inoculated with pool 189 showed a clear
cytopathic effect at day 4 of passage 0. This strain was namedTOSV–Algeria 189. A supernatant of passage 3 was used for
full-length genome sequencing and freeze-drying for long-term conservation; vials have been included in the collec-
tion of the European Virus Archive (http://www.european-virus-archive.com/), where they are publicly available under
the reference “Toscana virus Algeria strain 189” for academic
research.
Complete genome sequencingThe complete genome of TOSV–Algeria 189 consists of 6404,
4215 and 1869 nt for the L, M and S segments, respectively(GenBank accession nos. KP694240, KP694241 and
KP694242). The polymerase gene encodes a 6285 nt long openreading frame (ORF) (2095 aa), whereas the glycoprotein gene
encodes a 4017 nt long ORF (1339 aa). The small segmentencodes a 759 and 948 nt long ORF, which are translated to a
nucleocapsid protein (253 aa) and a nonstructural protein (316aa), respectively. Sequences obtained using NGS were
confirmed by direct sequencing performed on overlapping PCRproducts using Sanger sequencing.
Phylogenetic analysisTOSV–Algeria 189 was grouped together with other TOSVsequences regardless the gene used for analysis. Among other
TOSV strains, Algeria 189 was most closely related, with strainsoriginating from Tunisia, Italy, Spain, France and Portugal, thus
showing that this strain belongs to lineage A (Fig. 1). Thediscrimination between lineages A and B was unambiguousexcept for analysis done with the amino acid sequence of the N
gene.
MN-based seroprevalence studyA total of 370 human sera (from 248 female and 122 malesubjects) was collected from blood donors in Draa El Mizan.
Detailed results and the characteristics of the population seraobtained are presented in Tables 1 and 2. Using a virus dose at1000 TCID50, 171 sera (46.2%) contained neutralizing anti-
bodies against TOSV considering 10 as the cutoff titre. Using avirus dose at 100 TCID50, 170 sera (45.95%) contained
neutralizing antibodies against TOSV considering 20 as thecutoff titre. The correlation between results observed using
100 and 1000 TCID50 viral inoculums was significant (Pearsoncorrelation (2-tailed) coefficient: 0.427 was significant at the
0.01 level, p � 0.05). The geometric mean of the titres forTOSV MN according to age group is shown in Fig. 2. The
variation of seroprevalence rates among age groups wascalculated and found to be statistically significant by the chi-square test (two tailed) at the 0.001 level (p � 0.05).
Discussion
Recent studies have demonstrated that TOSV is not onlyconfined to Southern Europe and the Middle East but is also
present in Northern African countries such as Tunisia and
1040.e3 Clinical Microbiology and Infection, Volume 21 Number 11, November 2015 CMI
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reserved, CMI, 21, 1040.e1–1040.e9

(a)
(b)
(c)
TOSV-Algeria-KP694240
TOSV-Tunisia-JX867534
TOSV-Italy-NC_006319
TOSV-Italy-X68414
TOSV-Spain-FJ153280
TOSV-Spain-FJ153279
TOSV-France-FJ153281
TOSV-France-KC776216
TehranVirus-JF939846
SFNV-PoonaStrain-HM566176
SFNV-HM566172
SFNV-HM566183
PuniqueVirus-JF920133
GranadaVirus-GU135606
MassiliaVirus-EU725771
SFTV-IzmirStrain-GQ847513
100
100100
100
79
100
10098
99
100
88
100
84
0.05
TOSV-Algeria- KP694241
TOSV-Tunisia-JX867535
TOSV-ItalyEU003177
TOSV-Italy-NC_006320
TOSV-Italy-X89628
TOSV-Italy-EU003180
TOSV-Italy-JF330279
TOSV-Spain-FJ153282
TOSV-Spain-FJ153283
TOSV-France-HQ184337
TOSV-France-FJ153284
Tehran-JF939847
SFNV-HM566171
SFNV-HM566177
Punique-JF920134
Massilia-EU725772
Granada-GU135607
SFTV-NC_015411
100
100
81
96
96
100
100100
100
100
79100
0.05 TOSV-Algeria-KP694241
TOSV-Italy-X89628
TOSV-Italy-NC_006320
TOSV-ItalyEU003177
TOSV-Italy-EU003180
TOSV-Italy-JF330279
TOSV-Tunisia-JX867535
TOSV-Spain-FJ153282
TOSV-Spain-FJ153283
TOSV-France-HQ184337
TOSV-France-FJ153284
Punique-JF920134
Massilia-EU725772
Tehran-JF939847
SFNV-HM566171
SFNV-HM566177
Granada-GU135607
SFTV-NC_015411
100
100
99
99
99
100
99
98
77
99
0.05
FIG. 1. Phylogenetic analysis of selected
strains of Sandfly fever Naples virus using
complete amino acid sequences. (a) L pro-
tein, (b) Gn protein, (c) Gc protein, (d)
Nucleocapsid protein, (e) Non-structural
protein.
CMI Alkan et al. Toscana virus in Algeria 1040.e4
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reserved, CMI, 21, 1040.e1–1040.e9

Morocco. However, at the outset of this study, few data wereavailable for Algeria. In the founding study on the worldwide
distribution of neutralizing antibodies (NT Ab) in human pop-ulations against several phleboviruses, NT Ab were not foundagainst either Sicilian or Naples viruses; the presence of NT Ab
against TOSV had not been tested in this study [19]. Tworecent serologic studies reported the presence of IgG-specific
viruses belonging to the sandfly fever Naples and sandfly fe-ver Sicilian serocomplexes using enzyme-linked immunosor-bent assay and immunofluorescence assay tests, which are
(d) TOSV-France-KC776214
TOSV-Italy-JF330274
TOSV-Tunisia-JX867536
TOSV-Algeria-KP694242
TOSV-Italy-JF330275
TOSV-Italy-NC_006318
TOSV-Italy-EU327772
TOSV-Spain-FJ153286
TOSV-Spain-FJ153285
TOSV-Spain-EF120631
TOSV-Spain-EF120632
TOSV-Spain-EF120630
TOSV-Spain-EF120629
TOSV-Portugal-EF201833
TOSV-Spain-EF120628
SFNV-SabinStrain-EF201829
PuniqueVirus-FJ848987
MassiliaVirus-EU725773
GranadaVirus-GU135608
TehranVirus-JF939848
SFTV-NC_015413
100
93
83
73
100
0.05
(e) TOSV-Tunisia-JX867536
TOSV-Algeria-KP694242
TOSV-Italy-JF330275
TOSV-Italy-EU327772
TOSV-Italy-JF330274
TOSV-Italy-NC_006318
TOSV-Spain-EF120631
TOSV-Spain-FJ153285
TOSV-Portugal-EF201833
TOSV-Spain-FJ153286
TOSV-France-KC776214
TehranVirus-JF939848
SFNV-SabinStrain-EF201829
PuniqueVirus-FJ848987
MassiliaVirus-EU725773
GranadaVirus-GU135608
SFTV-NC_015413
100
100
88 87
94
98
100
100
10090
75
0.1 FIG. 1. (continued).
1040.e5 Clinical Microbiology and Infection, Volume 21 Number 11, November 2015 CMI
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reserved, CMI, 21, 1040.e1–1040.e9

notoriously prone to cross-reactivity, thus indicating that these
viruses or antigenically related viruses had to be present in thestudied regions of northern Algeria [17,18].
In addition, Sicilian-like virus RNA was detected and partiallysequenced in P. ariasi sandflies trapped in the Kabylia region
(northern Algeria); the virus was not isolated because thespecimens were stored in guanidinium thiocyanate [17]. Inanother study, Naples-like virus RNA was detected in
P. longicuspis, and Sicilian-like virus RNA was detected inP. papatasi [18]; again, virus isolation was not attempted, for the
same reasons as above. However, the sequences obtained wereshort, thus precluding detailed phylogenetic studies; in addition,
as a result of the cross-reactions of enzyme-linked immuno-sorbent assays and immunofluorescence assays, there was pre-
viously no definitive evidence that TOSV was present in Algeria.Although not unexpected, the results of this study provide
undisputable evidence of the following: TOSV is present insandflies in Algeria; its complete genome sequence allowed usto classify this strain as belonging to lineage A; and it is signifi-
cantly transmitted to local human populations, as assessed byrates of NT Ab approaching 50%.
To our knowledge, this study was the first to isolate andcharacterize the genome of a member of the Phlebovirus genus
in Algeria. This TOSV strain, isolated from a pool of sandfliescollected in Draa El Mizan, belongs to lineage A, as was the
strain isolated in Tunisia [12]. In contrast, the TOSV strainisolated in Morocco belongs to lineage B [17]. Interestingly, itappears that the delineation observed in Southern Europe in
the countries bordering the Mediterranean also exists in
northern Africa in the southern border of the Mediterranean.
Indeed, lineage B is present in Portugal, Spain, France andMorocco, whereas lineage A is present in France (particularly
Corsica Island), Italy, Tunisia and Algeria. Interestingly, inTurkey, both lineages were described [29], although the virus
was never isolated. In Croatia [11] and Greece [10], a thirdlineage (C) was identified on the basis of partial polymerasesequences. Whether this delineation is accurate merits future
investigation by lineage-specific molecular tests, as moleculartests are the only tests capable of distinguishing among TOSV
lineages in sandflies and in human specimens.To date, TOSV infection rates of sandflies in countries where
it was detected were as follows; 0.22% in Italy [30], 0.05% inSpain [31], 0.29% in France [25] and 0.03% in Tunisia [12]. In
this study, the infection rate of sandflies was 0.004%, assumingthat only one sandfly was infected in the TOSV-positive pool. It
is unlikely that these differences are due to technical differencesof the protocols because several of the above-mentionedstudies were done by the same group with identical tech-
niques [12,25]. These discrepant results can be explained byhighly changing environmental conditions that depend on the
period during which the fieldwork was performed. Repetitivetrapping campaigns during successive years are necessary to
better understand the dynamics of infection rates.A total of 22 species of sandflies, including 12 Phlebotomus and
10 Sergentomyia species, were identified in Algeria [32]. Amongsandfly fever vectors, P. perfiliewi, P. perniciosus and P. papatasihave been reported to be present in this country [33–35]. In the
region where this study was done, >47% of the sandfly
TABLE 2. Distribution of Toscana virus neutralizing antibodies according to sex and age for 100 TCID50
Age group (years) No of sera
Sex Neutralizing titre
Total, n (%)F M Ratio (M/F) Negative ‡ 10 ‡ 20 ‡ 40 ‡ 80
0–20 24 13 11 0.85 21 2 0 1 0 1 (4.17%)21–40 155 115 40 0.35 80 13 20 25 17 62 (40.0%)41–60 97 69 28 0.41 39 8 13 26 11 50 (51.55%)>60 94 51 43 0.84 25 12 18 16 23 57 (60.64%)Total 370 248 122 0.49 165 35 51 68 51 170 (45.95%)
TCID50, tissue culture infectious dose that will infect 50% if cell monolayers are challenged with defined inoculum.
TABLE 1. Distribution of Toscana virus neutralizing antibodies according to sex and age for 1000 TCID50
Age group (years) No. of sera
Sex Neutralizing titre
Total, n (%)F M Ratio (M/F) Negative ‡10 ‡20 ‡40 ‡80 ‡160
0–20 24 13 11 0.85 22 2 0 0 0 0 2 (8.3%)21–40 155 115 40 0.35 90 59 5 0 0 1 65 (41.9%)41–60 97 69 28 0.41 50 37 9 0 1 0 47 (48.5%)>60 94 51 43 0.84 37 47 9 0 1 0 57 (60.6%)Total 370 248 122 0.49 199 145 23 0 2 1 171 (46.2%)
TCID50, tissue culture infectious dose that will infect 50% if cell monolayers are challenged with defined inoculum.
CMI Alkan et al. Toscana virus in Algeria 1040.e6
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reserved, CMI, 21, 1040.e1–1040.e9

population consisted of P. perniciosus, whereas P. perfiliewi and
P. longicuspis occupied ranks 2 and 3, respectively. In this study,we identified 51.4% and 36.7% of the 1500 sandflies as
P. perfiliewi and P. perniciosus, respectively. As a result of thenature of this study, it was not possible to define the species of
sandflies in which the Algerian strain of TOSV was isolated.However, because, according to previous studies in the same
region [17,18,34] most sandflies belonged to P. perniciosus, andgiven the fact that TOSV is known to be transmitted by speciesbelonging to the Larroussius subgenus, most frequently by
P. perniciosus and P. perfiliewi, we can speculate that the situationin Algeria does not contradict the existing data.
Although the presence of TOSV in sandflies strongly sug-gested that the virus was causing human infections, its existence
and the level of exposure of humans to the virus was addressed
by testing 370 human serum samples collected from the pop-
ulation living in the vicinity of the sandfly capture sites, andtherefore exposed to sandfly bites and possibly infected with
the virus. Because the local population does not frequentlytravel abroad, seroprevalence rates are likely to reflect local
exposure to TOSV, thus providing information concerning thelocal presence and circulation of TOSV. Depending on the
criteria adopted to define a positive case, the seroprevalencevaries greatly. To attempt to determine which criteria are bestsuited to positive identification, we calculated the correlation
between results observed using 100 and 1000 TCID50 virusinoculums. Interestingly, when compared, the same results
were observed with the combination of 100 TCID50/titre 20and 1000 TCID50/titre 10 cutoff values. In both cases, the
analysis of the geometric mean of titres according to age group
(a)
(b)
1000 TCID50
100 TCID50
5.3
77.6
8.3
0
2
4
6
8
10
0 - 20 21 - 40 41 - 60 > 60
Geom
etric
Mea
nof
Tite
rsfo
rTO
SV
Age group
5.8
12
15.2
20
0
5
10
15
20
25
0 - 20 21 - 40 41 - 60 > 60
Geom
etric
Mea
nof
Tite
rsfo
rTO
SV
Age group
FIG. 2. Geometric mean of titres for
TOSV neutralisation antibodies accord-
ing to age groups.
1040.e7 Clinical Microbiology and Infection, Volume 21 Number 11, November 2015 CMI
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reserved, CMI, 21, 1040.e1–1040.e9

revealed that there is continuous exposure during a subject’s
lifetime, although it appears more clearly using the 100 TCID50
assay. As previously demonstrated [12], sandfly bites occur
mostly at home; the occupational risk seems to be limited [36].When the most stringent cutoff value for positivity is
retained (1000 TCID50 at titre 20), 7% of the population hadneutralizing antibodies against TOSV, which is in agreementwith the rates observed in the most exposed regions of
southeastern France—that is, the areas with the highest cir-culation of TOSV. However, it appears more realistic to use the
correlated combinations as described above. In this case, 46%of the population demonstrates the presence of neutralizing
antibodies, which is also in agreement with results recentlyreported in northern Tunisia [16].
Together, these results strongly suggest that TOSV is heavilyaffecting sandfly-exposed people in northern Algeria. Thesecoastal regions are the most highly populated, with a total of at
least 34 million Algerian inhabitants at risk of TOSV infection.Because TOSV causes various types of diseases, ranging from self-
resolving febrile illnesses to central and peripheral nervous sys-tem infections, it is now necessary to implement the diagnosis of
TOSV in regions where the circulation is high during the warmseason.
The microneutralization-based serologic data confirm thecirculation of TOSV in the region. However, TOSV and Naples
virus show a one-way relationship in neutralization assays:TOSV sera neutralize at almost the same titre for both TOSVand Naples virus, whereas the Naples virus sera do not
neutralize TOSV. Even if the Naples virus was not found in thearea, the low infectious rate of sandflies does not permit us to
exclude its presence, now or in the past. Finding sera capable ofneutralizing Naples virus only (not TOSV) would demonstrate
that Naples virus is still circulating in the area. In this study, wedid not perform this type of assay. However, during the last 15
years, tens of thousands of sandflies have been tested forphleboviruses using RT-PCR assays capable of amplifying Naplesvirus. Naples-like virus RNA was detected only once [18],
despite the fact that many other phleboviruses (including newones) were detected and/or isolated. In addition, the last report
of the isolation of Naples virus was in the 1990s in Cyprus andAfghanistan [37]. It is therefore plausible that the Naples virus
may be close to extinction. This hypothesis might be confirmedin the future using human sera, which may show during which
decade the Naples virus drastically decreased.In conclusion, our results confirm that TOSV (lineage A) is
present in northern Algeria in sandflies which are likely tobelong to the Larroussius subgenus, and that TOSV infectshumans at rates that are comparable to those observed in
northern Tunisia [12] and that are much higher than thosedescribed in the majority of Southern European countries
[25,29,30]. This fact should be taken into consideration to
rapidly implement the diagnosis of TOSV in the NationalReference Centre for arboviruses and in clinical microbiology
laboratories to create the capacity to detect TOSV infections inpatients with central nervous system infections and to estimate
the burden caused by this virus in Algeria.
Transparency declaration
All authors report no conflicts of interest relevant to this
article.
Acknowledgements
Fieldwork was supported by project 105738-001 of the CRASC(Centre de Recherche en Anthopologie Sociale et Culturelle).The authors thank A. Sellam and coworkers for the help during
the field trip, the team of the Draa El Mizan hygiene laboratory,and Dr Mezreg for providing the human sera. This work was
supported in part by the European Virus Archive (EuropeanUnion Seventh Framework Programme under grant agreement
no. 228292) (http://www.european-virus-archive.com/), and theEDENext FP7- n°261504 EU project. The work of RNC was
done under the frame of EurNegVec COST Action TD1303.
References
[1] Verani P, Lopes MC, Nicoletti L, Balducci M. Studies on Phlebotomus-transmitted viruses in Italy: I. Isolation and characterization of a sandflyfever Naples-like virus. In: Vesenjak-Hirjan J, Porterfield JS,Arslanagi�c E, editors. Arboviruses in the Mediterranean countries.Stuttgart-New York: Gustav Fischer on behalf of the Federation ofEuropean Microbiological Societies; 1980. p. 195–201.
[2] Calisher CH, Weinberg AN, Muth DJ, Lazuick JS. Toscana virusinfection in United States citizen returning from Italy. Lancet1987;1(8525):165–6.
[3] Ehrnst A, Peters CJ, Niklasson B, Svedmyr A, Holmgren B. Neuro-virulent Toscana virus (a sandfly fever virus) in Swedish man after visitto Portugal. Lancet 1985;1(8439):1212–3.
[4] Nicoletti L, Verani P, Caciolli S, Ciufolini MG, Renzi A, Bartolozzi D,et al. Central nervous system involvement during infection by Phlebovirustoscana of residents in natural foci in central Italy (1977–1988). Am JTrop Med Hyg 1991;45:429–34.
[5] Valassina M, Cuppone AM, Bianchi S, Santini L, Cusi MG. Evidence ofToscana virus variants circulating in Tuscany, Italy, during the summersof 1995 to 1997. J Clin Microbiol 1998;36:2103–4.
[6] Valassina M, Cusi MG, Valensin PE. Rapid identification of Toscanavirus by nested PCR during an outbreak in the Siena area of Italy. J ClinMicrobiol 1996;34:2500–2.
[7] Valassina M, Meacci F, Valensin PE, Cusi MG. Detection of neurotropicviruses circulating in Tuscany: the incisive role of Toscana virus. J MedVirol 2000;60:86–90.
CMI Alkan et al. Toscana virus in Algeria 1040.e8
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reserved, CMI, 21, 1040.e1–1040.e9

[8] Charrel RN, Gallian P, Navarro-Mari JM, Nicoletti L, Papa A, Sánchez-Seco MP, et al. Emergence of Toscana virus in Europe. Emerg Infect Dis2005;11:1657–63.
[9] Ergünay K, Saygan MB, Aydo�gan S, Lo MM, Weidmann M, Dilcher M,et al. Sandfly fever virus activity in central/northern Anatolia, Turkey:first report of Toscana virus infections. Clin Microbiol Infect 2011;17:575–81.
[10] Papa A, Paraforou T, Papakonstantinou I, Pagdatoglou K, Kontana A,Koukoubani T. Severe encephalitis caused by Toscana virus, Greece.Emerg Infect Dis 2014;20:1417–9.
[11] Punda-Poli�c V, Mohar B, Duh D, Bradari�c N, Korva M, Fajs L, et al.Evidence of an autochthonous Toscana virus strain in Croatia. J ClinVirol 2012;55:4–7.
[12] Bichaud L, Dachraoui K, Piorkowski G, Chelbi I, Moureau G, Cherni S,et al. Toscana virus isolated from sandflies, Tunisia. Emerg Infect Dis2013;19:322–4.
[13] Es-Sette N, Nourlil J, Hamdi S, Mellouki F, Lemrani M. First detectionof Toscana virus RNA from sand flies in the genus Phlebotomus(Diptera: Phlebotomidae) naturally infected in Morocco. J Med Entomol2012;49:1507–9.
[14] Fezaa O, Bahri O, Alaya Bouafif NB, Triki H, Bouattour A. Seropre-valence of Toscana virus infection in Tunisia. Int J Infect Dis 2013;17:e1172–5.
[15] Fezaa O, M’ghirbi Y, Savellini GG, Ammari L, Hogga N, Triki H, et al.Serological and molecular detection of Toscana and other phlebovi-ruses in patients and sandflies in Tunisia. BMC Infect Dis 2014;14:598.
[16] Sakhria S, Bichaud L, Mensi M, Salez N, Dachraoui K, Thirion L, et al.Co-circulation of Toscana virus and Punique virus in northern Tunisia:a microneutralisation-based seroprevalence study. PLoS Negl Trop Dis2013;7:e2429.
[17] Izri A, Temmam S, Moureau G, Hamrioui B, de Lamballerie X,Charrel RN. Sandfly fever Sicilian virus, Algeria. Emerg Infect Dis2008;14:795–7.
[18] Moureau G, Bichaud L, Salez N, Ninove L, Hamrioui B, Belazzoug S,et al. Molecular and serological evidence for the presence of novelphleboviruses in sandflies from Northern Algeria. Open Virol J 2010;4:15–21.
[19] Tesh RB, Saidi S, Gajdamovic SJ, Rodhain F, Vesenjak-Hirjan J. Sero-logical studies on the epidemiology of sandfly fever in the Old World.Bull World Health Organ 1976;54:663–74.
[20] Alkan C, Alwassouf S, Piorkowski G, Bichaud L, Tezcan S, Dincer E,et al. Isolation, genetic characterization and seroprevalence of Adanavirus, a novel phlebovirus belonging to the Salehabad virus complex inTurkey. J Virol 2015;89:4080–91.
[21] Charrel RN, Moureau G, Temmam S, Izri A, Marty P, Parola P, et al.Massilia virus, a novel Phlebovirus (Bunyaviridae) isolated from sandfliesin the Mediterranean. Vector Borne Zoonotic Dis 2009;9:519–30.
[22] Achour Barchiche N, Madiou M. [Outbreak of cutaneous leishmaniasis:about 213 cases in the province of Tizi-Ouzou]. Pathol Biol (Paris)2009;57:65–70.
[23] Abonnenc E. Les phlébotomes de la région éthiopienne (Diptera:Phlebotomidae). Mem Orstom 1972;55:1–289.
[24] Leger N, Pesson B, Madulo-Leblond G, Abonnenc E. Sur la différ-enciation des femelles du genre Laroussius Nitzulescu, 1931 (DipteraPhlebotomidae) de la région méditerranéenne. Ann Parasitol HumComp 1983;58:611–23.
[25] Charrel RN, Izri A, Temmam S, Delaunay P, Toga I, Dumon H,et al. Cocirculation of 2 genotypes of Toscana virus, southeasternFrance. Emerg Infect Dis 2007;13:465–8.
[26] Sánchez-Seco MP, Echevarría JM, Hernández L, Estévez D, Navarro-Marí JM, Tenorio A. Detection and identification of Toscana and otherphleboviruses by RT-nested-PCR assays with degenerated primers.J Med Virol 2003;71:140–9.
[27] Palacios G, Savji N, Travassos da Rosa A, Guzman H, Yu X, Desai A,et al. Characterization of the Uukuniemi virus group (Phlebovirus:Bunyaviridae): evidence for seven distinct species. J Virol 2013;87:3187–95.
[28] Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S.MEGA5: molecular evolutionary genetics analysis using maximumlikelihood, evolutionary distance, and maximum parsimony methods.Mol Biol Evol 2011;28:2731–9.
[29] Dincer E, Gargari S, Ozkul A, Ergunay K. Potential animal reservoirs ofToscana virus and coinfections with Leishmania infantum in Turkey. AmJ Trop Med Hyg 2015;92:690–7.
[30] Verani P, Ciufolini MG, Caciolli S, Renzi A, Nicoletti L, Sabatinelli G,et al. Ecology of viruses isolated from sand flies in Italy and charac-terized of a new Phlebovirus (Arabia virus). Am J Trop Med Hyg1988;38:433–9.
[31] Sanbonmatsu-Gámez S, Pérez-Ruiz M, Collao X, Sánchez-Seco MP,Morillas-Márquez F, de la Rosa-Fraile M, et al. Toscana virus in Spain.Emerg Infect Dis 2005;11:1701–7.
[32] Belazzoug S. The sandflies of Algeria. Parassitologia1991;33(Suppl.):85–7.
[33] Kabbout N, Merzoug D, Chenchouni H. Ecological status of phlebot-omine sandflies (Diptera: Psychodidae) in rural communities of North-eastern Algeria. J Arthropod Borne Dis 2016;10:24–38.
[34] Maroli M, Feliciangeli MD, Bichaud L, Charrel RN, Gradoni L. Phle-botomine sandflies and the spreading of leishmaniasis and other dis-eases of public health concern. Med Vet Entomol 2013;27:123–47.
[35] Izri MA, Belazzoug S, Boudjebla Y, Dereure J, Pratlong S, Delalbre-Belmonte A, et al. [Leishmania infantum MON-1 isolated from Phlebo-tomus perniciosus, in Kabylia (Algeria)]. Ann Parasitol Hum Comp1990;65:151–2.
[36] D’Ovidio MC, Venturi G, Fiorentini C, Barbati G, Di Renzi S,Vonesch N, et al. Occupational risk associated with Toscana virusinfection in Tuscany, Italy. Occup Med (Lond) 2008;58:540–4.
[37] Alkan C, Bichaud L, de Lamballerie X, Alten B, Gould EA, Charrel RN.Sandfly-borne phleboviruses of Eurasia and Africa: epidemiology, ge-netic diversity, geographic range, control measures. Antiviral Res2013;100:54–74.
1040.e9 Clinical Microbiology and Infection, Volume 21 Number 11, November 2015 CMI
Clinical Microbiology and Infection © 2015 European Society of Clinical Microbiology and Infectious Diseases. Published by Elsevier Ltd. All rights reserved, CMI, 21, 1040.e1–1040.e9

Inventaire des phlébotomes dans le foyer leishmanien de Draa El Mizan, Kabylie
37 JFMO : N°0, Mars 2017
Inventaire des phlébotomes dans le foyer leishmanien de Draa El Mizan, Kabylie
Sandflies’s inventory in a leishmaniasis home in Draa El Mizan, Kabylie
Amina Bouchra Allal -Ikhlef1, Karima Senouci1, Kamel Benallal2, Rachid Triki-Yamani3, Zoubir Harrat2, Leila Houti4
1 Faculté des Sciences de la Nature et de la Vie. Université Oran 12 Laboratoire d’Eco-Epidémiologie Parasitaire et Génétique des Populations, Institut Pasteur, Alger3 Laboratoire de Parasitologie- ISV-Blida-14 LABSIS. Faculté de Médecine. Université Oran 1
MOTS CLÉS
Phlébotomes, inventaire, piègeslumineuxtype CDC, Draa El-Mizan.
Résumé
Les leishmanioses, maladies à transmission vectorielle liée au genre Phlebotomus, constituent un problème de santé publique en Algérie, essentiellement dans leur forme cutanée. Dans la région de Draa El Mizan (Kabylie), coexistent depuis long-temps laleishmaniose viscérale, et la forme cutanée liée àLeishmania infantum.
Durant l’été 2013, une enquête entomologique a été menée à Draa El-Mizan, dans le but de réaliser un inventairefaunistique des phlébotomes à l’aide de pièges lumineux type CDC.Sur un total de 22.998 spécimens récoltés, un échantillon représentatif de 1500 individus a fait l’objet d’une identificationmorphologique par microscope optique.
Le genre Phlebotomus est prédominant avec 97,41% des individus représentant 5 taxons où prévalent Phlebotomus perfiliewi (4,63%) et Phlebotomus perniciosus (19,4%). Le sex-ratio M/F global est de 1,39, mais en faveur des individus femelles pour l’espèce Phlebotomus perfiliewi (sex-ratio M/F=0,82).A Draa El Mizan, les conditions climatiques associées aux activités humaines de type pastorales entretiennent la prolifération du genre Phlebotomus inféodé à L. Infan-tum. Par ailleurs, l’utilisation des pièges lumineux type CDC a montré son intérêt dans la surveillance entomologique des maladies vectorielles.
Auteur correspondant :[email protected]
Abstract
Leishmaniasis, vector-borne diseases related to the genus Phlebotomus, is a public health problem in Algeria, mainly in its cutaneous form. The region of Draa El Mizan (Kabylie), is known as home to the visceral form related to Leishmania infantum and
KEY WORDSDuchenne musculardys
Articles originauxJournal de la Faculté de Médecine d’Oran
Jfmo
Article original

A.B. Allal -Ikhlef, K. Senouci, K. Benallal, R. Triki-Yamani, Z. Harrat, L. Houti
38 JFMO : N°0, Mars 2017
Introduction
La leishmaniose est un complexe de maladies avec un large éventail de formes cliniques (cutanée, viscérale etcuta-néo-muqueuse). Elle est due à des protozoaires du genre Lei-shmania,parasite du système des phagocytes mononucléés d’un grand nombre de mammifères, incluant l’homme [1]. La maladie est cosmopolite, spécialement en zone tropicale et tempérée. Elle affecte 12 millions de personnesdans 88 pays,avec 2 millions de nouveaux cas et 60.000 décèschaque année [2, 3]. Ainsi, dans le monde350 millions de personnes sontà risque [4,5]. Approximativement 20 espèces de Leish-mania sont reconnues pathogènes pour l’homme [5, 6].
Cette maladie qui a un impact important sur la santé des populations, est de nature anthopo-zoonotique [7]. La crois-sance démographique dans les zones rurales et périurbaines, associée aux activités humaines qui altèrent les écosystèmes naturels [2] sont responsables de la prolifération du réservoir, le chien ou le rongeur [8, 9, 10]et du vecteur [10, 11].
En Algérie, les leishmanioses constituent un vrai problème de santé publique [7, 12, 13] et bénéficient de programmes spé-cifiques de prévention. Elles sévissent sous plusieurs formes clinico-épidémiologiques :
- la forme cutanée zoonotique du Sud, la plus fréquente, sévissant dans les régions steppiques du Nord du Sahara et s’étendant aux régions cotières [12, 14].
- la forme viscérale, mortelle en l’absence de traitement, sous forme épidémique dans le Nord du pays, particulière-ment en Kabylie [15] et sporadique dans l’extrême Sud[16]. Pour la forme viscérale humaine dont le chien est le principal réservoir, Phlebotomus perniciosus en est le vecteur confirmé [17];
- la forme cutanée chronique, récemment décrite dans la ré-
gion de Ghardaïacausée par le parasite Leishmania killicki [18] récemment rapportée à Tipaza [19] et dont le vecteur est Phlebotomus sergenti [20].
Dans le pourtour méditerranéen,l’espèce de parasite incri-minée est Leishmania infantum, avec le chien comme hôte réservoir principal [17, 21]. Sa transmission est saisonnière en relation avec la densité et l’activité des populations de phlébotomes. Phlebotomus perniciosus est leprincipal vec-teur de Leishmania infantum [17] et il assure également la transmission du virus Toscana [22, 23].
En Kabylie, les données épidémiologiques mettent en évi-dence l’existence d’un foyer endémique de leishmaniose cu-tanéedu Nord et viscérale,dans la région de Draa El Mizan liée à Leishmania Infantum [15, 17].
L’objectif de cette étude est de réaliser un inventaire des-phlébotomes qui sévissentdans cette région particulière.
Matériel et méthodes
1. Site d’étude
La région de Draa El-Mizan en Kabylie (36°32’8»N, 3°50’3»E) est situéeà 110 km au Sud-Est d’Alger, en zone montagneuse culminant jusqu’à650 mètres d’altitude. Elle bénéficie d’un climat méditerranéen, tempéré et humide, à l’étage bioclimatique subhumide. La pluviométrie annuelle varie entre 600 et 800 mm. L’hiver est doux, oscillant souvent entre 0 et 3°C. Le mois le plus froid de l’année se situe entre dé-cembre et janvier, et le plus chaud entre juillet et août avec une température maximale de 36° à 40°C et une humidité accrue, surtout après la construction du barrage Taksebt.2.
trophy, Dystrophin, Mul-tiplex PCR, deletions, X chromosome
became since 20 years an area related to cutaneous leishmaniasis.
During summer 2013, an entomological survey was conducted at Draa El-Mizan in order to make a fauna inventory of sandflies, using light traps type CDC. Out of 22998 har-vested specimensa representative sample of 1500 individuals underwent morphological identification by optical microscope.
The genus Phlebotomus is predominant with 97.41% of individuals representing 5 taxa where prevail Phlebotomus perfiliewi (4.63%) and Phlebotomus cease (19.4%). The glo-bal M/F sex ratio is 1.39. Conversely, individuals females haematophagous are more frequent in species Phlebotomus perfiliewi (sex ratio M/F = 0, 82).
At Draa El Mizan, the climatic conditions associated with the human pastoral activities maintain the proliferation of the genus Phlebotomus subservient to Leishmania infan-tum. Furthermore, the use of light traps type CDC showed his interest in Entomological surveillance of vector diseases.

Inventaire des phlébotomes dans le foyer leishmanien de Draa El Mizan, Kabylie
39 JFMO : N°0, Mars 2017
Echantillonnage et traitement des phlébotomes
La collecte des phlébotomes a été réalisée durant le mois d’Août 2013. Durant 5 nuitées consécutives, 8 pièges type CDC «mini light-trap CDC » (John WHock Company, Gainse-ville, FL) (figure 1) modifiés,numérotés, ont été placés en fin de journée, dans différents abris d’animaux d’élevage des ovins, bovins et volailles,et récupérés tôt le matin.Ils sont placés à 12 mètres au-dessus du sol dans un coin ombragé, selon la technique de Rioux et Izri [19, 21]. Au total, 33 pié-geages ont été réalisés dans différents sites géo-localisés, à des altitudes variant entre 379 et 386 mètres.
Les phlébotomes sont anesthésiés à l’aide d’une compresse imprégnée de chloroforme. Les insectes sont ensuite déver-sés dans un plateau blanc émaillé, puis délicatement récupé-rés à l’aide d’une pince douce et placés dans des cryotubesde 2ml,numérotés et portant les références du piège.
Un échantillon représentatif de spécimens tiré à partir de chacun des sites a été retenu pour identification microsco-pique. Le reste des phlébotomes sont conservés à -80°C pour des analyses virologiques ultérieures.
3. Dissection et identification des phlébotomesLes phlé-botomes ont été identifiés morphologiquement suivant les clésd’identification d’Abonnenc et de Léger [24, 25], en te-nant compte quelques fois du nombre des soies médianes des coxites.
Les phlébotomes ont subi un éclaircissement dans du NaOH 20% durant 2 heures, suivi de deux à trois lavages consécutifs à l’eau puis laissés toute la nuit dans la solution de Marc Andrée. Dans la même solution, l’insecte (mâle et femelle) a été monté entre lame et lamelle et examinés au microscope optique.
Les résultats sont présentés sous forme de fréquences rela-tives.
Résultats
Au total, 22.998 phlébotomes ont été récoltés, soit en moyenne 4600 spécimens par jour et 700 par piège. Parmi les spécimens capturés, l’étude morphologique a concerné un échantillon de 1.500 phlébotomes, soit 6,52% des individus récoltés.
Dans la région de Draa El Mizzan, le genre Phlebotomus est prédominant (97,41%) suivi par le genre Sergentomyia (2.6%). L’espèce Phlebotomus perfiliewiest prédominante (51,4 %), suivie de Phlebotomus perniciosus (36.7 %) et de Phleboto-mus papatasi (6,4%) (Tableau 1).
Cette étude a montré par ailleurs une prédominance des indi-vidus mâles qui sont deux fois plus fréquents que les individus femelles (68,3% vs 31,7%) (Tableau 2). Cette prédominance masculine est particulièrement observée chez Phlebotomus perniciosus (sex-ratio M/F = 3,29). Par ailleurs, nous avons relevé une prédominance féminine dans la population de
Phlebotomus perfiliewi (sex-ratio M/F=0,82).
Discussion
En Algérie, les enquêtes des foyers ont permis de mettre en évidenceles vecteurs des leishmanioses et la prolifération des phlébotomes est liée à la présence des hôtes [10, 26].A ce jour, l’Algérie compte 24 espèces de phlébotome après le report de la présence de l’espèce Phlebotomus mascitti en Kabylie [27].
Ainsi en 1984, Dedet a montré que parmi les 15 espèces ren-contrées en Algérie, seules deux espèces dominent par leur abondance et leur fréquence. Il s’agit de Sergentomyia mi-nuta (78.9%) et de Phlebotomus. perniciosus (10.4%) [28].
Genre et sous genre
Espèce Nb d’in-dividus
(n=1500)
Fréquence (%)
Phlebotomus ( larroussus)
P. perfiliewi 772 51,47
P. pernicio-sus 550 36,67
P. longicuspis 40 2,67
Phlebotomus (Phlebotomus)
P. papatasi 97 6,47
Phlebotomus (Paraphleboto-mus)
P. sergenti02 0,13
Sergentomyia S. minuta 35 2,33
Sergen-tomyia sp 4 0,27
Tableau 1 : Identificationdes phlébotomes capturés par les pièges DC.
Espèce Mâles Fe-melles Sex-ratio M/F
P. perniciosus 422 128 3,35
P. papatasi 57 40 1,42
P. longicuspis 22 18 1,22
P. perfiliewi 348 424 0,82
P. sergenti 0 2 0
S. minuta 20 15 1,33
Sergentomya sp Indéterminé
Tableau 2 : Répartition des individus par sexe.

A.B. Allal -Ikhlef, K. Senouci, K. Benallal, R. Triki-Yamani, Z. Harrat, L. Houti
40 JFMO : N°0, Mars 2017
Dans notre étude, la présence de Phlebotomus perniciosus dans les captures aux pièges CDC a été observée avec une fréquence relativement élevée (36,67%). Ces phlébotomes abondent dans les régions tempérées à une altitude infé-rieure à 1000 mètres. Selon Rioux et al. (1967), cette espèce occupe un très vaste territoire géographique dans tout le bassin méditerranéen, particulièrement dans la partie sep-tentrionale, jusqu’à la lisière saharienne (Tassili et Hoggar) [29].
Dans l’Est algérien, cette espèce se retrouve dans tous les étages bioclimatiques avec cependant une fréquence plus élevée dans les zones humides [29, 30]. Phlebotomus per-niciosus est considéré depuis longtemps comme vecteur de Leishmania infantum dans le bassin méditerranéen [11, 21]. D’un point de vue épidémiologique, il est responsable de la forme viscérale et cutanée propre à Draa El Mizan [17, 21, 31, 32]. Son aire de distribution coïncide avec celle de la maladie humaine et animale [2, 10, 17]. Cette espèce, mise à part sa vaste distribution, présente deux formes atypiques surtout retrouvées dans l’Ouest et l’extrême Sud [33].
La forme cutanée du Nord, ayant également pour réservoir le chien [11, 34],est transmise par Phlebotomus perfiliewi, que nous avons recensé avec une fréquence de 51.47% à Draa El Mizan. Cette fréquence explique l’apparition depuis 1996, selon les services de santé, de la forme cutanée dans un foyer connu initialement pour la forme viscérale [15, 16, 31, 32].
L’espèce Phlebotomus papatasi a été peu retrouvée à Draa El Mizan (6.47%). Phlebotomus papatasi est le vecteur ma-jeur de la forme cutanée à L. major sévissant dans les zones arides [8, 9, 35] dont les principaux réservoirs sont les ron-geurs Gerbillidés ; Merione shawi et Psammomys obesus. On
considère que Phlebotomus papatasi présente sa densité maximale aux étages péri-aride et aride et devient rare dans les étages subhumide, humide et semi-aride. Guernaoui et al. (2006) corrèlent négativement son abondance avec l’al-titude et trouvent qu’il est plus abondant dans les stations à basse altitude, entre 300 et 400 mètres [36]. Phlebotomus papatasi est retrouvé plus souvent autour des terriers du rongeur Psammomys obesus au Sahara [8, 9, 10]. Il devient moins abondant au fur et à mesure que l’on s’éloigne de ces gites. C’est un insecte des régions arides, plus abondant au Sud qu’au Nord [9, 10]. A proximité, les matières organiques constituent un lieu de pullulation, soit une nouvelle niche écologique. Cette niche secondaire ou niche anthropique, prend le relais de la niche primaire sauvage et permet l’in-troduction et la prolifération des phlébotomes dans le milieu péri-domestique ou domestique [10].
Depuis longtemps, la leishmaniose cutanée du Nord à Lei-shmania infantum et la leishmaniose cutanée du Sud à Lei-shmania major ont été nettement séparées par la chaîne montagneuse de l’Atlas Tellien qui constitue une barrière naturelle. Un nouveau foyer de leishmaniose cutanée est ap-paru dans la localité d’El M’hir, située sur le versant Nord de cette chaîne. Les prospections entomologiques ont montré la prédominance de Phlebotomus papatasi et Phlebotomus perniciosus. Le premier est reconnu comme vecteur de L. major dans le Sahara algérien. L’étude de Boudrissa et almet en évidence l’extension de L. major des zones arides vers les zones semi-arides de la vallée de la Soummam [9,10]. Ce phénomène pourrait être lié à l’impact des changements climatiques et particulièrement du phénomène de la déser-tification de la région steppique au Nord Sahara [9]. Rioux et al. (1986) signalent que, sur le pourtour du bassin méditer-ranéen, le développement des leishmanioses dépend du vo-lume des populations de phlébotomes [21]. Dans des foyers similaires au Nord du Maroc, les auteurs, Rioux et al. (1997), ont lié l’augmentation de l’incidence de la leishmaniose, d’une part à l’augmentation des densités des vecteurs, en particulier Phlebotomus sergenti et Phlebotomus longicus-pis, et d’autre part à l’allongement de leurs périodes d’ac-tivité [37]. Ces auteurs expliquent que la prolifération des vecteurs est plutôt liée à une détérioration des conditions d’hygiène et plus particulièrement à l’installation d’abris animaux de plus en plus au voisinage des habitations hu-maines [37].
Le genre Sergentomyia est très faiblement retrouvé (2,6%). Cette très faible représentation est liée au mode de pié-geage. En effet, on connait le comportement herpétophile de ces espècesqui cherchent leurs hôtes, essentiellement des reptiles, dans les anfractuosités où sont posés les papiers huilés destinés à capturer les moucherons. Les observations de Bousaaet al en 2005 ont signalé la présence de 36.5% de genre Sergentomyia dans la région de Marrakech avec une
Figure 1 : Piège lumineux type CDC

Inventaire des phlébotomes dans le foyer leishmanien de Draa El Mizan, Kabylie
41 JFMO : N°0, Mars 2017
différence de richesse des espèces de Sergentomiya entre un biotope rural et urbain. Les travaux de ces auteurs menés dans des foyers méditerranéens, indiquent une prévalence comprise entre 70 et 96,6% [38].
Plusieurs facteurs ont ainsi concouru à l’obtention de ces résultats quantitatifs et qualitatifs : le mode de capture par les pièges lumineux type CDC qui ont permis de récolter un grand nombre de spécimens ayant donné lieu à un échantil-lon représentatif pour l’identification microscopique, l’alti-tude inférieure à 1000 mètres avec une zone bioclimatique favorable ainsi que la présence de matières organiques pré-sentes dans le fumier au niveau des multiples élevages dans la région.
Précédemment, une campagne répétée de piégeages réali-sée dans le même site, au cours des périodes d’été s’étalant entre 2010 et 2013, par la technique du papier huilé, n’a permis de récolter qu’un nombre réduit de spécimens en 3 années (n=2783), tous morts. Ainsi, les pièges lumineux ont démontré leur qualité en termes de quantité et de qualité de spécimens capturés, mais aussi de temps. S’ils restent un outil de choix pour la surveillance entomologique des lei-shmanioses et autres maladies vectorielles émergentes, les spécimens capturés vivants permettent d’engager ultérieu-rement tous les examens de génomique et de protéomique nécessaires afin d’identifier le parasite en cause [39]. Même s’ils nécessitent un appareillage complexe de coût élevé, ils peuvent être rapidement amortis dans le cadre des pro-grammes de prévention, comme ils permettent de récolter des espèces rares à potentiel épidémique voire pandémique [40].
Conclusion
Les résultats de ce travail ont montré une diversité en es-pèces des phlébotomes capturés. Le genre Phlebotomus prédomine avec plus de la moitié des spécimens, représen-tant 7 taxons où prévalent Phlebotomus perfiliewet Phle-botomusperniciosus.Cette prolifération est en faveur de la circulation du vecteur, inféodé aux leishmanioses du type Leishmania infantum, dans ses deux formes cutanée du Nord et viscérale. Elle entretient le cycle naturel de la maladie, au sein d’un écosystème favorable, constitué par un étage bioclimatique subhumide associé à une activité économique basée sur l’élevage.
Par ailleurs, la capture des phlébotomes par les pièges type CDC montre l’intérêt de ce type de piège qui a permis, en un temps limité, de récolterune quantité importante de spéci-mens de bonne qualité. A ce titre, il apparait intéressant de mettre en place un programme de surveillance du vecteur des leishmanioses et autres maladies vectorielles, utilisant
les pièges lumineux type CDC.
Conflit d’intérêt
Les auteurs ne signalent aucun conflit d’intérêt.
Remerciements
Ce travail a été soutenu par le Centre de Recherche en An-thropologie Sociale et Culturelle-CRASC (Projet d’Etablisse-ment: 2013-2016). Les auteurs remercient pour leur aide à Draa El Mizan,l’ensemble du staff de l’APC sous la direction de A. Sellami ainsi que l’équipe du Laboratoire d’Hygiène.
Références bibliographiques
[1] Gramiccia M, Gradoni L. The current status of zoonotic leishma-
niases and approaches to disease control. Int J Parasitol 2005 35:1169-
1180.
[2] Desjeux P. The increase of risk factors for leishmaniasis worldwide.
Trans. R. Soc. Trop. Med. Hyg., 95, 2001, 239-243.
[3] Who - World Health Organization. Control of the leishmaniasis:
report of a meeting of the WHO Expert Committee on the Control of
Leishmaniases. WHO Press, Geneva 2012. 2010:186.
[4] Dedet J.P. Leishmanioses en Afrique du Nord. Bulletin de l’Institut
Pasteur d’Algérie 1979,77 (1): 49-82.
[5] Desjeux P. Leishmaniasis: current situation and new perspectives.
Comp. Immunol. Microbiol. Infect. Dis 2004, 27, 305-318.
[6] Choi C.M.; Lerner E.A. Leishmaniasis as an emerging infection. J.
Invest. Dermatol. Symp. 2001, Proc 6, 175-182.
[7] WHO - World Health Organization. Control of the leishmaniases
1990. World Health Organ Tech Rep Ser 1990, 793: 1-177.
[8] Belazzoug S. Isolation of Leishmania major from Psammomys
obesus (Rodentia: Gerbillidae) in Algeria. Trans R Soc Med Hyg 1983,
77:876.
[9] Boudrissa A, Cherif K, Kherrachi I et al. Extension de Leishmania
major au Nord de l’Algérie. Spread of Leishmania major to the north
of Algeria Bull. Soc. Path. Exot 2012, 105:30-35.
[10] Izri AM, Depaquit J, Parola P. Phlébotomes et transmission
d’agents pathogènes autour du bassin méditerranéen Med Trop 2006;
66 : 429-435.
[11] Izri MA, Belazzoug S, Benhabyles N et al. Le clou de Biskra (Al-
gérie). Résultats d’une étude éco-entomologique. Bulletin de la so-
ciété française de parasitologie, 1998, tome 16, numéro1.
[12] Institut National de la Santé publique. REM: Relevé Epidémi-

A.B. Allal -Ikhlef, K. Senouci, K. Benallal, R. Triki-Yamani, Z. Harrat, L. Houti
42 JFMO : N°0, Mars 2017
ologique Mensuel 2000-2014.Available from: http://www.ands.dz/
insp/rem. html.
[13] Harrat Z, Hamrioui B, Belkaid M et al. Point actuel sur l’épidé-
miologie des leishmanioses en Algérie Bulletin de la Société de Pathol-
ogie Exotique.1995. T: 88. 180 -184.
[14] Boudrissa A, Cherif K, Kherrachi I et al. Spread of Leishmania
major to the north of Algeria. Bull Soc Pathol Exot.2012 Feb ;105
(1):30-5.
[15] Achour N, Madiou M. Recrudescence des leishmanioses cutanées
: à propos de 213 cas dans la wilaya de Tizi-Ouzou.Pathologie Biologie
2008, doi:10.1016/j.
[16] Benallal K, Gassen B, Bouiba L et al.Entomological investigation
following the resurgence of human visceral leishmaniasis in southern
Algeria.Acta Trop. 2013, 128(3):518-21.
[17] Izri MA, Belazzoug S, Boudjebla Y et al. Leishmania infantum
Mon-1 isolé de Phlebotomus perniciosus en Kabylie (Algérie). Ann.
Parastitol Hum Comp 1990, 65(3), 151–152.
[18] Harrat Z, Boubidi SC, Pratlong F et al.Trans R. Description of a
dermatropic Leishmania close to L. killicki (Rioux, Lanotte & Pratlong
1986) in Algeria.Soc Trop Med Hyg. 2009 Jul;103(7):716-20.
[19] Izri MA, Bendjaballah A, Andriantsoanirina V et al. Cutaneous
Leishmaniasis Caused by Leishmania killicki, Algeria. Emerg Infect Dis.
2014 Mar; 20(3): 502–504.
[20] Boubidi ASC, Benallal K, Boudrissa A et al. Phlebotoms sergenti
(Parrot, 1917) identified as Leishmania killicki host in Ghardaïa, south
21 February Elsevier Masson SAS 2011.
[21] Rioux JA, Lanotte G, Petter F et al. Les leishmanioses cutanées
du bassin méditerranéen occidental:de l’identification enzymatique à
l’analyse éco-épidemiologique, l’exemple de trois ‘foyers’, tunisien,
marocain et français 1986, p. 365–395 in: Rioux JA. Leishmania. Tax-
onomie et phylogenése. Applications écoépidémiologiques. Montpel-
lier, France: Institut Méditerranéen d’Etudes Épidémiologiques et
Ecologiques.
[22] Charrel R, Izri MA, Temmam S et al. Cocirculation of 2 genotypes
of Toscana Virus, Southeastern France. Emerging Infectious Diseases
2007. WWW.cdc.gov/eid. vol. 13, N°03.
[23] Sanbonmatsu-Gamez S, Perez-Ruiz M, Collao X et al. Toscana virus
in Spain. EmerInfect Dis 2005, 11: 1701-1707.
[24] Abonnenc E. Les phlébotomes de la région éthiopienne (Diptera:
Phlebotomidae). Mémoire de l’ORSTOM 1972, 55, 1–289.
[25] Leger N, Pesson B, Madulo-Leblond Get al. Sur la différenciation
des femelles du genre LaroussiusNitzulescu, 1931 (DipteraPhleboto-
midae) de la région méditerranéenne. — Ann. Parasitol. Hum. Comp.,
1983, 58, 611-623.
[26] Berchi S. Les phlébotomes (Insecta, Diptera, Phlebotomidae),
vecteurs de leishmanioses dans l’est algérien. Bull de la Société
Zoologique de France 1993, 118: 341-349.
[27] Berdjane-Brouk Z, Charrel RN, BitamI et al. Record of Phlebot-
omus (Transphlebotomus) mascittii Grassi, 1908 and Phlebotomus
(Larroussius) chadlii Rioux, Juminer & Gibily, 1966 female in Algeria.
Research note 337 Parasite, 2011, 18, 337-339.
[28] Dedet JP, Addadi K, Belazzoug S. Cahiers O.R.S : Les phléboto-
mes (Diptera, Psychodidae) d’Algérie. Cah O.R.S.T.O,M:, abr. sér. Ent.
méd. et Paradol 1984, vol. XXI-I, n°2: -99-127.
[29] Rioux JA, Golvan YJ, Croset H et al. Ecologie des leishmanioses
dans le Sud de la France. I. les Phlébotomes. Echantillonnage. Ethol-
ogie. Annales de ParasitologieHumaine Comparée, 1967,42: 561-603.
[30] Rispail P, Dereure J, JARRY D. Risk zones of human leishmania-
sis in the western mediterranean basin. Correlations between vector
sandflies, bioclimatology and phytosociology. Memorias do Instituto
Oswaldo Cruz 2002, 97(4):477.
[31] Harrat, Z, Addadi, K, Belkaid M et al. La leishmaniose viscéraleen
Algérie : recensement des cas de leishmaniose (période 1985-1990).
Bull.Soc. Pathol Exot, 1992, 85, 296-301.
[32] Haddadi K, Dedet J.P. Des leishmanioses en Algérie. 6. recensem-
ent des cas de leishmanioses viscérales infantiles entre 1965 et 1974.
Bulletin de la Société de Pathologie Exotique 69: 68 – 75.
[33] Benallal K, Benikhlef R, Garni R et al. Presence of Phlebotomus
perniciosus atypical form in Algeria. (sous presse Journal of Arthro-
pod-Borne Diseases).
[34] Benikhlef R, Harrat Z, Toudjine M et al. Detection of Leishmania
infantum MON-24 in the dog. Med Trop (Mars). 2004;64(4):381-3.
[35] Belazzoug S. Découverte d’un Meriones shawï (Rongeur, Gerbill-
idé) naturellement infesté par Leishmania dans le nouveau foyer de
leishmaniose cutanée de Ksar Chellala (Algérie). Bull Soc Pathol Exo
1986;79:630-3.
[36] Guernaoui S, Boussaa S, Pesson B et al. Nocturnal activity of phle-
botomine sandflies
(Diptera: Psychodidae) in a cutaneous leishmaniasis focus in
Chichaoua, Morocco. Parasitol Res (2006) 98: 184–188.
[37] Rioux JA, Akalay O, Perieres J et al. L’évolution éco-épidémi-
ologique du risque leishmanien au Sahara atlantique marocain. Intérêt
heuristique de la relation phlébotomes bioclimats. Ecole Méditer-
ranée 1997, 23, 73–92.
[38] Boussaa S, Guernaoui S, Pesson B et al. Seasonal fluctuations of
phlebotomine sand fly populations (Diptera: Psychodidae) in the urban
area of Marrakech Moroco. ActaTrpica 2005, 95 2°: 8691.
[39] Alkan C, Allal-Ikhlef AB, Alwassouf S et al. Virus isolation, genetic
characterization and seroprevalence of Toscana virus in Algeria. Clin-
ical Microbiology and Infection © 2015 European Society of Clinical
Microbiology and Infectious Diseases. Published by Elsevier Ltd. All
rights reserved, CMI, 21, 1040.e1–1040.

Inventaire des phlébotomes dans le foyer leishmanien de Draa El Mizan, Kabylie
43 JFMO : N°0, Mars 2017
[40] Benallal KE, Allal-Ikhlef A, Benhamouda K et al. First report of
Aedes (Stegomyia) albopictus (Diptera: Culicidae) in Oran, West of
Algeria.Acta Trop. 2016 Dec;164:411-413.


Résumé
Les leishmanioses, maladies parasitaires observées chez les mammifères dont l’Homme, sont causées
par un protozoaire du genre Leishmania, transmis par un insecte piqueur, le phlébotome femelle.
Deux formes cliniques de leishmanioses sévissent en Algérie, la leishmaniose viscérale et les
leishmanioses cutanées. Récemment, des arbovirus du genre Phlebovirus, les virus Sicilien et de
Naples ayant le phlébotome comme vecteur commun avec Leishmania, ont été signalés dans le Nord
de l’Algérie. Notre travail a été consacré à la recherche en Algérie, du virus Toscana et à la mise en
évidence de l’existence de son lien épidémiologique avec Leishmania, durant la période de 2012 à 2014.
L’étude a été menée dans la région de Draa El Mizan (Kabylie), un foyer endémique à leishmanioses. Un
total de 25781 phlébotomes a été capturé dont 4283 ont fait l’objet d’une étude taxonomique pour
déterminer la composition de la population phlébotomienne. Huit espèces du genre Phlebotomus et
Sergentomyia ont été recensées avec la prédominance de P. perniciosus. La recherche de Leishmania
chez les phlébotomes femelles s'est révélée négative. Une analyse moléculaire, basée sur la technique
de RT-PCR suivie de celle de la PCR nichée, a été utilisée pour la détection des phlébovirus chez
ces insectes. Ces techniques ont permis d’identifier et de caractériser pour la première fois en Algérie,
le virus Toscana (TOSV). Après la culture du virus sur des cellules Vero (ATCC CCL81), le séquençage
complet du génome viral a été réalisé par la technique de séquençage de nouvelle génération
(Next-Generation- Sequencing, NGS). La souche obtenue, nommée «Toscana Virus Algérie, souche
189», et classée phylogénétiquement dans la lignée A, est proche de la souche TOSV isolée en
Tunisie. L’étude a été complétée par deux tests sérologiques, Western Blot et séroneutralisation,
effectués sur 370 sérums provenant de personnes choisies aléatoirement parmi la population de
Draa El Mizan. Ces tests ont détecté respectivement 86 cas positifs à Leishmania et des anticorps
neutralisants contre TOSV dans 171 sérums en utilisant une dose de virus à 1000 DICT50. Ces résultats,
qui indiquent que TOSV circule à Draa El Mizan, sont en adéquation avec la présence du virus chez
les phlébotomes sévissant dans cette région. Par ailleurs, la séropositivité, aux deux tests, de 11
sérums ndique l’existence d’infections virales à TOSV concomitantes à une leishmaniose viscérale parmi la
population de Draa El Mizan.
Mots clés :
Leishmania; Phlebovirus; Virus Toscana; Phlébotome; PCR; Whole genome sequencing (WGS) ;
Séroneutralisation; Western Blot; Draa El Mizan; Algérie.


















