![Evaluation of Sorghum [Sorghum bicolor (L.)] Reference ...oar.icrisat.org/9449/1/fpls-07-00529.pdf · Sudhakar Reddy et al. Reference Genes for qRT-PCR in. Sorghum bicolor. potential](https://static.fdocuments.us/doc/165x107/5f6067667afad96ee66a2cad/evaluation-of-sorghum-sorghum-bicolor-l-reference-oar-sudhakar-reddy-et.jpg)
Expression of cell wall invertase and several other genes of sugar metabolism in relation to seed...
14
Journal of Plant Physiology 165 (2008) 331—344 Expression of cell wall invertase and several other genes of sugar metabolism in relation to seed development in sorghum (Sorghum bicolor) Mukesh Jain a,b , Prem S. Chourey a,b,c, , Qin-Bao Li a,c , Daryl R. Pring a,c a Department of Plant Pathology, University of Florida, Gainesville, FL 32611-0680, USA b Department of Agronomy, University of Florida, Gainesville, FL 32611-0680, USA c US Department of Agriculture, Agricultural Research Service, Chemistry Research Unit, CMAVE, Gainesville, FL 32608-1069, USA Received 26 September 2006; received in revised form 2 December 2006; accepted 2 December 2006 KEYWORDS Caryopsis develop- ment; Cell wall invertase; Sugar-to-starch tran- sition; Sugar unloading Summary We report expression profiles of several genes of carbohydrate metabolism, cell wall invertase (CWI) in particular, to better understand sugar transport and its utilization in developing caryopses of grain sorghum [Sorghum bicolor (L.) Moench]. Gene expression analyses for CWI using RNA gel blot and real-time quantitative PCR approaches on developing caryopses, including the glumes (maternal tissue appended to the seeds), showed expression of SbIncw (ZmIncw2 ortholog) primarily in the basal sugar unloading zone of endosperm. The expression of ZmIncw1 ortholog was significantly less abundant and restricted to the glumes. The protein and enzyme activity data corroborated the temporal transcript expression profile that showed maximal CWI protein (INCW) expression preceding the starch-filling phase of endosperm development, i.e. 6–12 d-after-pollination (DAP). Protein gel blot analysis using polyclonal maize INCW1 antibodies showed a single polypeptide of 72 kDa. The highest level of enzyme activity was unique to the basal part of the endosperm, in particular the basal endosperm transfer cell (BETC) layer and the maternal pedicel region that were highly enriched for the INCW protein, as seen by immunolocalization. High hexose-to-sucrose ratio in 6–12 DAP seeds, and negligible starch deposition in glumes corroborated the CWI activity data. Additionally, we report transcription profiles of several other genes related to sugar-to-starch metabolism in developing sorghum endosperm. As in maize, the INCW-mediated apoplastic cleavage of sucrose in the BETC and pedicel during the early ARTICLE IN PRESS www.elsevier.de/jplph 0176-1617/$ - see front matter Published by Elsevier GmbH. doi:10.1016/j.jplph.2006.12.003 Abbreviations: AGPase, ADP-glucose pyrophosphorylase; BETC, basal endosperm transfer cells; CWI/Incw , cell wall invertase gene; DAP, days-after-pollination; INCW, cell wall invertase protein; Pgm, phosphoglucomutase; UGPase, UDP-glucose pyrophosphorylase Corresponding author. USDA ARS, C/O Department of Plant Pathology, University of Florida, Gainesville, FL 32611-0680, USA. Tel.: +1 352 392 3631 345; fax: +1 352 392 6532. E-mail address: [email protected]fl.edu (P.S. Chourey).
-
Upload
mukesh-jain -
Category
Documents
-
view
212 -
download
0
Transcript of Expression of cell wall invertase and several other genes of sugar metabolism in relation to seed...

ARTICLE IN PRESS
Journal of Plant Physiology 165 (2008) 331—344
0176-1617/$ - sdoi:10.1016/j.
AbbreviationDAP, days-after�Correspond
Tel.: +1 352 392E-mail addr
www.elsevier.de/jplph
Expression of cell wall invertase and several othergenes of sugar metabolism in relation to seeddevelopment in sorghum (Sorghum bicolor)
Mukesh Jaina,b, Prem S. Choureya,b,c,�, Qin-Bao Lia,c, Daryl R. Pringa,c
aDepartment of Plant Pathology, University of Florida, Gainesville, FL 32611-0680, USAbDepartment of Agronomy, University of Florida, Gainesville, FL 32611-0680, USAcUS Department of Agriculture, Agricultural Research Service, Chemistry Research Unit, CMAVE, Gainesville,FL 32608-1069, USA
Received 26 September 2006; received in revised form 2 December 2006; accepted 2 December 2006
KEYWORDSCaryopsis develop-ment;Cell wall invertase;Sugar-to-starch tran-sition;Sugar unloading
ee front matter Publisjplph.2006.12.003
s: AGPase, ADP-glucose-pollination; INCW, celling author. USDA ARS, C3631 345; fax: +1 352 3ess: [email protected]
SummaryWe report expression profiles of several genes of carbohydrate metabolism, cell wallinvertase (CWI) in particular, to better understand sugar transport and its utilizationin developing caryopses of grain sorghum [Sorghum bicolor (L.) Moench]. Geneexpression analyses for CWI using RNA gel blot and real-time quantitative PCRapproaches on developing caryopses, including the glumes (maternal tissueappended to the seeds), showed expression of SbIncw (ZmIncw2 ortholog) primarilyin the basal sugar unloading zone of endosperm. The expression of ZmIncw1 orthologwas significantly less abundant and restricted to the glumes. The protein and enzymeactivity data corroborated the temporal transcript expression profile that showedmaximal CWI protein (INCW) expression preceding the starch-filling phase ofendosperm development, i.e. 6–12 d-after-pollination (DAP). Protein gel blotanalysis using polyclonal maize INCW1 antibodies showed a single polypeptide of72 kDa. The highest level of enzyme activity was unique to the basal part of theendosperm, in particular the basal endosperm transfer cell (BETC) layer and thematernal pedicel region that were highly enriched for the INCW protein, as seen byimmunolocalization. High hexose-to-sucrose ratio in 6–12 DAP seeds, and negligiblestarch deposition in glumes corroborated the CWI activity data. Additionally, wereport transcription profiles of several other genes related to sugar-to-starchmetabolism in developing sorghum endosperm. As in maize, the INCW-mediatedapoplastic cleavage of sucrose in the BETC and pedicel during the early
hed by Elsevier GmbH.
pyrophosphorylase; BETC, basal endosperm transfer cells; CWI/Incw, cell wall invertase gene;wall invertase protein; Pgm, phosphoglucomutase; UGPase, UDP-glucose pyrophosphorylase/O Department of Plant Pathology, University of Florida, Gainesville, FL 32611-0680, USA.92 6532..edu (P.S. Chourey).

ARTICLE IN PRESS
M. Jain et al.332
developmental stages of caryopses is essential for the normal development of filialtissues. The unique cell-specificity of the INCW protein to both proximal and distalends of placental sac shown here for the first time is likely to greatly increaseuptakes of both hexose sugars and water through turgor sensing into developingseed. This trait is unique to sorghum among cereals and may facilitate its survival indrought environment.Published by Elsevier GmbH.
Introduction
Among grasses, sorghum is economically animportant cereal plant that is used both as a graincrop as well as forage. It is also a model cereal forgenomic studies due to a high level of molecularsimilarity and synteny to maize (Mullet et al.,2001), and it also has much smaller genome, one-fourth the size than maize, due to greatly reducedlevels of repetitive DNA (see review Kresowichet al., 2005). Despite these similarities, maize andsorghum are quite distinct in plant architecture andin developmental anatomy of seed. Developingsorghum seed, unlike maize, harbors a placentalsac as a result of post-fertilization loss of nucellarcells in the placental region adjacent to the basalpart of the seed. The placental sac is speculated toserve as an apoplastic reservoir for hexoses inbetween the maternal base and the basal transfercells of the endosperm (Maness and McBee, 1986).Sorghum, as compared to maize, is also highlyadaptable to adverse growth conditions, includinghot and dry environments, and is therefore referredto as a ‘dryland’ crop. Some of these character-istics are probably attributable to its unique originin Africa; unlike maize, Triticeae (wheat, barleyand oats) and rice that originated, in America,Middle East and Asia, respectively (Kresowichet al., 2005). Thus, a comparative study especiallybetween maize and sorghum may yield valuabledata on traits and genes that lead to unique aspectsof each of the two crop plants.
Sugar metabolism and its control are critical toall parts of the plant, including developing seeds –
a major site of sugar utilization. Sucrose is theprincipal and preferred from of photosynthate thatis transferred between cells and the long distancetransport to terminal storage tissues. Invertases(b-fructofuranosidase; EC 3.2.1.26) catalyze theirreversible hydrolysis of sucrose into its constitu-ent monosaccharides fructose and glucose, andhave been classified into three isoforms based onoptimal pH requirements and subcellular localiza-tion (reviewed by Roitsch and Gonzalez, 2004).Soluble acid invertases are present in vacuoles;
whereas the insoluble acid invertases are localizedto the apoplast, usually ionically bound to the cellwall. An apoplastic acid invertase, unbound tocell wall and characterized by short C-terminalextension and modified catalytic domain, hasbeen described in maize (Kim et al., 2000), rice(Hirose et al., 2002) and sugar beet (Gonzalezet al., 2005).
One of the earliest genetic demonstrations onthe role of cell wall invertase (CWI) in determiningsink strength in developing seeds was described inmaize. The loss-of-function, non-lethal mutation atthe Mn1 locus that encodes a major endosperm-specific CWI isoform, INCW2 (CWI protein; Millerand Chourey, 1992; Cheng et al., 1996), isresponsible for the miniature1 (mn1) seed pheno-type associated with a loss of 470% mature seedweight. Notably, Mn1 expression is restricted to thebasal endosperm transfer cells (BETC), the singleport of photosynthate unloading in the developingmaize kernel (Cheng et al., 1996), and coincideswith the rapid cell division phase of endospermgrowth. The reduced seed weight phenotype ofmn1 seed mutant is associated with reduced cellelongation and cell number in the endosperm(Vilhar et al., 2002). Of the four CWI genes inmaize, ZmIncw1–ZmIncw4, ZmIncw3 and ZmIncw4are putative CWI genes; their expression in varioustissues is detectable only by RT-PCR, and neither isexpressed in developing seed (Kim et al., 2000).The ZmIncw1 and ZmIncw2 (Mn1) genes areconsidered the major genes, and our recent datashow that the residual low-level CWI activity indeveloping mn1 seed mutation is due to theZmIncw1 locus that is expressed in many parts ofthe plant (Taliercio et al., 1999; Chourey et al.,2006).
A major role of CWIs in apoplastic cleavage ofsucrose, and establishment and maintenance ofsink metabolism has been illustrated across variousplant species, including tomato (Godt and Roitsch,1997), fava bean (Weber et al., 1996), barley(Weschke et al., 2003; Sreenivasulu et al, 2004),rice (Hirose et al., 2002; Cho et al., 2005; Ishimaruet al., 2005) and Arabidopsis (Sherson et al., 2003).

ARTICLE IN PRESS
Gene expression analyses of sugar metabolism in developing sorghum caryopses 333
A number of transgenic studies have also strength-ened a correlation between altered CWI activityand the modification of sink strength in transgenicplants. Transgenic over-expression of CWI in potato(Sonnewald et al., 1997) and its antisense repres-sion in carrot (Tang et al., 1999) have confirmedthat CWIs play crucial role in early differentiationand development of sink organs. Additionally inmaize, our previous in vitro kernel culture data alsosuggest a signaling role of the Mn1-encoded INCWenzyme in normal kernel development as themetabolic-released hexose, not an exogenoussource, is critical for normal seed development(Cheng and Chourey, 1999).
The present study was aimed towards under-standing the spatial and temporal variation inexpression of CWI genes, and their contribution inrelation to sucrose transport and metabolismduring grain filling in sorghum. Given the significantsyntenic similarities between maize and sorghum,an attempt was made to compare and contrast Incwexpression between developing seeds of the twocereal crops. Additionally, we report spatial andtemporal profiles of sugars and starch levels, andtranscript profiles of several genes of sugarmetabolism during caryopsis development.
Materials and methods
Plant material
Style emergence, pollen maturation and anthesisproceed basipetally in sorghum panicle, and are com-pleted in about 4 d. The field-grown male-sterile panicles(A3Tx7000; Tx7000 nucleus, IS1112C cytoplasm) werehand pollinated using male-fertile, normal cytoplasm lineB3Tx7000 (Tang et al., 1996) when flowering wascomplete at the base of panicle, in order to ensureuniform and synchronized seed set. The seeds wereharvested at the indicated developmental stages (0–30days-after-pollination (DAP)), flash frozen under liquidnitrogen and stored at �80 1C until further use.
RNA gel blot analyses
Total RNA was extracted from the frozen sorghumseeds following the method of Wadsworth et al. (1988).RNA was glyoxalated and resolved on a 1.2% (w/v)agarose gel essentially according to Ausubel et al.(1993), followed by transfer to Nytran membranes(Schleicher and Schull, Keene, NH) and UV cross-linking.Probes were prepared from the full-length/partial cDNAclones and labeled using the random priming method(Prime-Its RmT Random Primer Labeling Kit, Stratagene,LaJolla, CA). Pre-hybridization, hybridization and wash-ing conditions were same as described previously(Chourey et al., 2006). The blots were exposed to X-ray
film, in between two intensifying screens, for 1- to 4 ddepending on the transcript abundance. Transcript sizewas estimated based on the position of the rRNA speciesvisualized in the ethidium bromide-stained gel prior toblotting.
RT-PCR cloning and real-time quantitative PCR analyses
Five mg total RNA was reverse transcribed usingSuperScriptTM First-Strand Synthesis System (Invitrogen,Carlsbad, CA) essentially according to the manufacturer’sprotocol. Real-time quantitative PCR was performedusing the DyNAmoTM HS SYBRs Green qPCR Kit (Finn-zymes, Espoo, Finland) and Chromo 4TM CFD supported byOpticon MonitorTM Software version 2.03 (MJ Research,Alameda, CA). The PCR reactions were prepared accord-ing to the manufacturer’s instructions and contained400 nM of both the forward and reverse gene-specificprimers and 2 ml of the five-fold diluted RT reaction in afinal volume of 20 ml. The thermal cycling protocolentailed 50 1C incubation for 2min, followed by TbrDNA polymerase activation at 95 1C for 15min. The PCRamplification was carried out for 35 cycles with dena-turation at 94 1C for 10 s, and primer annealing andextension at 55 and 72 1C for 30 s each, respectively.Optical data were acquired following the extension step,and the PCR reactions were subject to melting curveanalysis beginning at 55 through 95 1C, at 0.1 1C s�1.Additionally, PCR products were also checked for purityby agarose gel electrophoresis. The data are presented asaverage7SD of three independently made RT prepara-tions used for PCR run, each having three replicates.
The various gene specific primers used for PCR cloning,real-time PCR analyses and cDNA probes are listed inTable 1. The amplification products were cloned inpCRs2.1 vector using the TA Clonings Kit (Invitrogen,Carlsbad, CA), and sequenced to verify the fidelity of thePCR amplification.
Preparation of protein extracts and invertase activityassays
Protein extracts enriched for CWI protein wereprepared using approximately the lower half-sections ofthe developing seeds as described in Carlson et al.(2002). The basal sections of the frozen seeds wereexcised, pulverized under liquid nitrogen and the powderwas resuspended in 5 volumes of 50mM Tris–maleate(pH 7.0) buffer, followed by centrifugation at 27,000g for20min. The pellets (containing the cell wall fraction)were resuspended in 2 volumes of extraction buffersupplemented with 1M NaCl, vigorously agitated for30min at 4 1C using an Eppendorf shaker, and centrifugedfor 20min at 27,000g. The resulting supernatantsenriched for the CWI fractions were desalted on Micro-Bio-Spin 6 chromatography columns (BioRad, Hercules,CA) as per the manufacturer’s recommendations. Acidinvertase activity was measured in a spectrophotometricassay as previously described (Tsai et al., 1970; Miller andChourey, 1992). Activity assays were performed in

ARTICLE IN PRESS
Table 1. Primers used for cDNA cloning and real-time quantitative PCR for genes investigated in the present study
Gene GenBank mRNA Forward primer (seq. 50-30) Reverse primer (seq. 50-30)
SbIncwa EF177465 TGTACTACAAGGGGTGGTACCA TCAAGCTCCATTCATGAGTGGCTTCTTCATZmIncw1c U17695 CCGTCTTCTTCAGGGTGTTC CGTAGAGGGTGAGCGTCCTTCSbIncwc EF177465 GCAAAGTCGGTCACTCTCAGGAA AAATCCTGCAAATGTCGGGCGSh1d X02400 ACTACAAGGGCACGACGATGATGT TTCCCGCGATTTCTTGGAATGTGCSus1d L29418 TTCCTCAACAGGCACCTGTCATCA TCCAGGTACGGCCAGACTTCAAATSus3d AY059416 AGGATGCTGCTGGTGAAATTGCTG AAGGTTTACCAGCTCCCTCAGCTTSus3c AY059416 GCCATGTCTGCGCCGAAGCTGGACCGC CGAACGGCGGCAGCACGATCGCCTCCSh2d DQ019881 TTGAAGGCGGGATCAACTTTGCTG TCTTGCACTTGTCCAATTGCGTCGPgm2d U89342 TCTGGTGATGGCCGCTATTTCTCA ATCAGCTGCAGCACCAAATTCAGGSbIvr2e NAb CTCACCAACTGGACCAAGTACGA CCATGTAGTCGTGGTTGTATGACGSbTua1e NAb CCTGAGGTTCGATGGTGCTCT CCATCATCACCTTCCTCACCC
aPrimers used for RT-PCR cloning of CWI genes from 6 to 12 DAP sorghum seed.bNot available.cPrimers used for real-time quantitative PCR.dPrimers used for making cDNA probes from 12 to 16 DAP maize endosperm.eThe soluble invertase-2 (SbIvr2) and a-tubulin 1 (SbTua1) cDNA clones were amplified from immature sorghum microspore RNA.
M. Jain et al.334
triplicate, using at least two separate extractions foreach sample. Protein concentrations were determinedusing the BioRad Protein Assay (BioRad, Hercules, CA),with BSA as the standard.
SDS-PAGE and protein gel blots
Samples prepared for SDS-PAGE electrophoresis andblotting were immediately denatured in 2� SDS-PAGEsample buffer [0.125M Tris–HCl, 4% (w/v) SDS, 20% (v/v)glycerol, 0.02% (w/v) BPB, pH 6.8] by boiling at 100 1C for5min. SDS-PAGE analyses were done using 7.5% (w/v)acrylamide Tris–HCl gels according to Laemmli (1970).Protein blotting to PVDF membrane (Immobilon-P, Milli-pore, Billerica, MA) was performed in the BioRad MiniTrans-Blot apparatus, following the manufacturer’s in-structions. As an additional blotting control, pre-stainedstandards were loaded along with the samples (Kaleido-scope Markers, BioRad). Protein gel blots were developedusing enhanced chemiluminescent substrate (PierceFemto Kit, Rockford, IL). The primary polyclonal anti-INCW1 (Carlson and Chourey, 1999), anti-UGPase (UDP-glucose pyrophosphorylase) (Carlson et al., 2002) andanti-SH2 (Datta et al., 2001) sera were used at 1:2000,1:6000 and 1:5000 dilutions, respectively. Anti-mouse(for INCW) and anti-rabbit (for UGPase and SH2) HRP-labeled secondary antisera were used at 1:5000 dilutions.
Sugar analyses
Frozen seeds and glumes were ground to a fine powderand the soluble sugars were extracted in hot ethanol toseparate soluble sugars from starch. After centrifugation,the pellet was used for starch analysis, and the super-natant was used for glucose, sucrose and fructoseanalyses in a microtiter plate assay (Kerr et al., 1985).Starch analysis was done following amyloglucosidasedigestion (EC 3.2.13, from Aspergillus niger, Roche
Diagnostics, Indianapolis, IN) as described by Rufty andHuber (1983).
Immunolocalization of the CWI protein
Developing caryopses at 10 and 16 DAP were harvestedand immediately fixed in cold formalin acetic alcohol(FAA; 3.7% formaldehyde, 5% acetic acid and 50% ethanol)for 24 h, followed by dehydration in a series of ethanoland tertiary butyl alcohol (TBA), and infiltration andembedding in Paraplast Plus paraffin (Sherwood MedicalCo., St. Louis, MO). Paraffin-embedded caryopses weresectioned with a rotary microtome (Microm 325, CarlZeiss, Germany). Polyclonal antibodies of maize CWI(Carlson and Chourey, 1999) were used for immunodetec-tion of INCW, and the immunogold signal was visualizedusing the ZYMED Kit (Zymed Laboratories Inc, SanFrancisco, CA).
Results
Cloning and sequence analysis of sorghumCWI cDNA clone
Figure 1 illustrates the deduced amino acidsequence alignment of the Incw-like partial cDNAclone of sorghum with ZmIncw1 and ZmIncw2. The1.6 kb SbIncw clone was PCR amplified fromreverse-transcribed mRNA isolated from 6 to 12DAP seeds, using the primer pair described inTable 1. The primers were designed following aBLAST search and alignment of maize Incw se-quences in the GSS data base (Genomic sequencesurveys), in order to consolidate relevant sequenceinformation available from sorghum. The SbIncwclone has the conserved cysteine catalytic motifMWECP, a hallmark feature of CWIs, as opposed to

ARTICLE IN PRESS
ZmINCW2 1 ----MRALVVVSFAS---ACLLLLLQLAGASHVVYNYKDLEAEAAAATDQVPPSIVNPLL
SbINCW 1 ------------------------------------------------------------
ZmINCW1 1 MGTRPRGVVLAPWAVVLVLVLVLALRLAGASHVIH--RSLEAEVPPS---VPASIVSPLL
consensus 1 ..... .. ... . ........ . .... .. ... ...
ZmINCW2 54 RTGYHFQPPKNWINDPNAPMYYKGWYHFFYQYNPKGAVWGNIVWAHSVSRDLINWVALEP
SbINCW 1 --------------------YYKGWYHFFYQYNPKGAVWGNIVWAHSVSRDLINWMALEP
ZmINCW1 56 RTGYHFQPPMNWINDPNAPLYYKGWYHLFYQYNPKGAVWGNIVWAHSVSRDLINWVALEA
consensus 61 ......... ..........*******.***************************.***.
ZmINCW2 114 ALRPSIPGDRYGCWSGSATVLPDGGGPVIMYTGVDHPDINYQVQNVAYPKNVSDPLLREW
SbINCW 41 AIKPSIPSDKYGCWSGSATTMPDG-TPVIMYTGIDRPNTNYQVQNVAYPRNKSDPLLREW
ZmINCW1 116 AIYPSIPSDKYGCWSGSATILEDG-TPAILYTGIDRPDINYQVQVLALPKNASDPLLREW
consensus 121 *..****.*.*********...** .*.*.***.*.*..*****..*.*.* ********
ZmINCW2 174 VKP-SHNPVIVPEGGINATQFRDPTTAWRG--PGPEQWRLLVG------SAAGSMPPRGV
SbINCW 100 VKP-SYNPVIVPEGGINATQFRDPTTAWRATAGGDGHWRLLIGSVRTTTTTGATAPPRGV
ZmINCW1 175 EKPEEYNPVATPAAGINATQFRDPTTAWRH--AGAGHWRMLVG------SVRG---ARGM
consensus 181 .** ..***..*..***************. .* ..**.*.* . ... ..**.
ZmINCW2 225 AYVYRSRDFRRWRRVRRPLHSAP-TGMWECPDFYPVSK---GGAPRAGLETSVPPGPRVK
SbINCW 159 AYVYRSRDFRRWTRVRRPLHSAA-TGMWECPDFYPVSSDEDGRRRRVGLETSVPSGARVK
ZmINCW1 224 ALVYRSRDFRTWTKAKHPLHSAALTGMWECPDFFPVSG----PGLQAGLDTSAPG---TK
consensus 241 *.********.*.....*****. *********.*** . . ..**.**.* . ..*
ZmINCW2 281 HVLKNSLDLRRYDYYTVGTYHPRAERYVPDDPAGD---EHRLRYDYGNFYASKTFYDPAK
SbINCW 218 HVLKNSLDLRRYDYYTVGTYDRDAERYVPDDPAGDDDGERRLRYDYGNFYASKTFYDPAK
ZmINCW1 277 YVLKSSLDLTRYDYYTIGSYDGGKDRYYPDDPAGD--YRRRLRYDYGNYYASKTFYDPVE
consensus 301 .***.****.******.*.*. ..**.******* ..********.*********..
ZmINCW2 338 RRRILWGWANESDTAADDVAKGWAGIQAIPRTVWLDPSGKQLLQWPIEEVEALRAKSVTL
SbINCW 278 RRRILWGWANESDTAADDVAKGWAGIQAIPRTVWLDPSGKQLLQWPVEEVEALRGKAVTL
ZmINCW1 335 RRRVLLGWANESDSVPDDKAKGWAGIHAIPRKIWLDPTGKQLLQWPIHEVEKLRGKAVSV
consensus 361 ***.*.*******...**.*******.****..****.********..***.**.*.*..
ZmINCW2 398 RNRVIKAGHHVEVTGIQTAQADVEVSFEVSP--AALAGAETLDPALAYDAERLCGVKRAD
SbINCW 338 KNRVIKPGQHVEVTGIQTAQADVEVSFEVSP--SALAGAERLDPALADDAERLCGVKRAD
ZmINCW1 395 DAKLVKPGDHFEVTGIATYQADVEVSFELEAGTSLLEKAEAFDPAYDDDAQKLCGVKGAD
consensus 421 .....*.* *.*****.*.*********... ..*..** .***...**..*****.**
ZmINCW2 456 VRGGVGPFGLWVLASANRKERTAVFFRVFKPAAG---DKPVVLMCTDPTKSSLNPNLYRP
SbINCW 396 VKGGVGPFGLWVLASANLKERTAVFFRVFKAAAGSSNNKPVVLMCTDPTKSSLNPNLYRP
ZmINCW1 455 ARGGVGPFGLWVLASADLQERTAVFFRVFRDGHG----KPKVLMCTDPTKSSLSPDLYKP
consensus 481 ..**************...**********. ..* **.************.*.**.*
ZmINCW2 513 TFAGFVDTDISNGKISLRSLIDRSVVESFGAGGKTCILSRVYPSLAIGKDARLYVFNNGR
SbINCW 456 TFAGFVDTDISNGKISLRSLIDRSVVESFGAGGKTCILSRVYPSLAIGNNARLYVFNNGK
ZmINCW1 511 TLAGLVDADISSGKITLRSLIDRSVVESFGAGGKTCILSRVYPSIAVGKDAHLYVFNNGE
consensus 541 *.**.**.***.***.****************************.*.*..*.*******.
ZmINCW2 573 AHVKVSRLTAWEMKKPVMNGA
SbINCW 516 ADVRVSRLTAWEMKKPLMNGA
ZmINCW1 571 VDVTVSGLTAWEMKKPLMNGA
consensus 601 ..*.**.*********.****
Figure 1. Comparison of deduced amino acid sequences for (partial) cell wall invertase (CWI) cDNA clone of sorghum(SbIncw, EF177465) and ZmIncw1 (U17695) and ZmIncw2 (AF165180). Conserved cysteine catalytic site (MWECPD) andthe three glycosylation motifs (NXS, NAT and NES) are shown in shaded regions. Gaps in the alignment are indicated bydashes and identical residues are marked as asterisks.
Gene expression analyses of sugar metabolism in developing sorghum caryopses 335
MWECV present in vacuolar invertases (Goetz andRoitsch, 1999). Additionally, there are three po-tential N-glycosylation sites (NXS/T), one of which(NES) is conserved in all plant CWIs identified so far.Deduced amino acid sequence of SbIncw gene wasalso aligned with those reported from variouspublished monocot and dicot plant species, and aphylogenetic tree (see Supplementary data) wasconstructed using the CLUSTAL W alignment (La-serGene 6, MegAlign 6.1). Phylogenetic and mole-cular evolutionary analyses placed the SbIncw genein the monocot-I group of CWIs, along with
ZmIncw1, ZmIncw2 and ZmIncw3, and was foundto be most similar to the maize ZmIncw2 and to alesser extent to ZmIncw1 or ZmIncw3. The SbIncwshared 86% homology with ZmIncw2 and 75% withZmIncw1 at DNA level, and 85% and 72% respec-tively, at the protein level.
Presently, the SbIncw cDNA clone lacks about200 bp sequences on the 50 end, since the ATG startcodon was not detectable following sequencealignments. The conserved b-fructosidase motif,NDPNG/A, and a potential cleavage site for target-ing extracellular localization for the translated

ARTICLE IN PRESS
M. Jain et al.336
protein are presumably included in the missingspan. Efforts to extend these sequences further,either by 50 RACE or sorghum database searches,have so far been unsuccessful. Attempts areunderway to screen a cDNA library of youngsorghum seeds for obtaining full-length clones.
Expression of soluble and cell wall invertasegenes during development of sorghumcaryopses
Figure 2 shows RNA gel blot analysis of SbIncwexpression in the developing sorghum caryopses.The RNA blot data showed temporal variability withmaximum transcript levels at 6 DAP (Fig. 2A).Significant transcript levels were seen both in theseed and glumes at 6 DAP, even though SbIncwtranscription was restricted only to lower half ofseed at 12 DAP (Fig. 2B). Given the close sequencesimilarity between the SbIncw clone and ZmIncw2(Fig. 1), the same RNA blots were stripped and re-hybridized with the ZmIncw2 probe. As expected,hybridization profiles were similar to those ob-tained with the SbIncw probe, with high transcript(2.0 kb) abundance in whole seed at 6 DAP andlower endosperm section at 12 DAP, with very lowexpression in 6 DAP glumes. Conversely, SbIncwclones did not hybridize to RNA extracted frommaize mn1-1 mutant lacking Incw2 (data notshown) thus confirming that the SbIncw genedescribed here is indeed ZmIncw2 ortholog.Furthermore, ZmIncw1 probe was also used to lookfor the expression of a respective ortholog insorghum seed. A 2.1 kb ZmIncw1 ortholog expres-
0 6 12 18 24 30 DAP
ZmIncw2
ZmIncw1
SbIncw
SbIvr2
Tua1
A
Figure 2. RNA gel blot showing temporal (A) and spatial (B) vainvertase genes during development of sorghum caryopses.unfertilized florets were used for RNA extraction at 0 DAP, and(A); and intact seeds were used for RNA extraction at 6 DAP,zone) and lower (sugar transport zone) half sections (B). Lo0.95 kb) (A), and sucrose synthase-3 (Sus3, 2.7 kb) transcript
sion was seen at high steady-state levels duringearly stages i.e. at 6 DAP, and reduced levels duringthe subsequent stages of seed development. TheZmIncw1 ortholog was readily detectable in 6 DAPglumes with limited expression in whole seed, andin lower endosperm and glumes at 12 DAP. Loadingcontrols for RNA levels are represented by a-tubulin(Tua1, 0.95 kb).
We also examined RNA expression levels of asoluble invertase gene (SbIvr2) using a partial cDNAclone previously isolated from young post-meioticsorghum microspores (Jain et al., unpublisheddata). The highest SbIvr2 (�2.2 kb) expressionlevels were detected prior to anthesis (0 DAP),declining post-6-DAP and was undetectable at 18DAP (Fig. 2A). At 6 DAP; SbIvr2 expression wassignificantly higher in seeds as compared to glumes(Fig. 2B). Steady-state levels of the SbIvr2 tran-scripts in upper and lower parts of 12 DAP seedwere comparable. Low-level expression of SbIvr2 inglumes at 6 DAP became undetectable at 12 DAP.
Real-time quantitative PCR analyses of cellwall invertase genes during development ofsorghum caryopses
Real-time PCR analyses (see Table 1) were usedfor absolute quantification of Incw transcriptabundance in sorghum (Fig. 3), and for corrobora-tion of the RNA blot data (Fig. 2). Maximumexpression of SbIncw gene was seen in the seedsat 6 DAP (Fig. 3C and D), whereas the expression ofZmIncw1 ortholog was restricted primarily to the 6DAP glumes (Fig. 3A and B). The expression of
6 DAP 12 DAP
glumes
glumes
upperlow
ersee
d
ZmIncw1
ZmIncw2
SbIncw
SbIvr2
Sus3
B
riability in the expression profiles of cell wall and solubleAll lanes were loaded with 15 mg total RNA. Completeseeds without glumes were sampled for 6–30 DAP stages
whereas the 12 DAP seeds were spliced in upper (storageading control for RNA is represented by a-tubulin (Tua1,profiles (B).

ARTICLE IN PRESS
b
0
4000
8000
12000
16000
0
1e+ 5
2e+ 5
3e+ 5
4e+ 5
5e+ 5
6e+ 5
0
5000
10000
15000
20000
25000
30000
seedglumes
upperlower
glumes
6 DAP 12 DAP
Zm
Incw
1 tr
ansc
ript
0
2000
4000
6000
8000
SbIn
cw tr
ansc
ript
0. 0
2.0 e+ 4
4.0 e+ 4
6.0 e+ 4
8.0 e+ 4
1.0 e+ 5
1.2 e+ 5
1.4 e+ 5
Sus3
tran
scri
pt
0
10000
20000
30000
40000
50000
0 6 12 18 24 30Developmental stage (DAP)
A B
C D
FE
Figure 3. Real-time quantitative PCR analyses of temporal and spatial variability in the expression profiles usingZmIncw1 (A, B) and SbIncw (C, D) primers, in glumes and endosperm during development of sorghum caryopses. Sucrosesynthase-3 (Sus3) represents the RNA quantification and reverse transcriptase (RT) reaction efficiency control (E, F).Complete unfertilized florets were used for RNA extraction at 0 DAP, and seeds without glumes were sampled for 6–30DAP stages (A, C and E); and intact seeds were used for RNA extraction at 6 DAP, whereas the 12 DAP seeds were cut inupper (storage zone) and lower (sugar transport zone) half sections (B, D and F). Five mg total RNA was used for first-strand cDNA synthesis reaction, diluted five-fold and used for quantitative PCR analyses as described in Materials andmethods. The transcript abundance was calculated by absolute estimation method. The amplified PCR products werecloned in pCRs2.1 (TOPO) vector, and used for generating the standard curve. Transcript abundance (10 ng�1 total RNA)is presented as average7SD of three independently made RT preparations used for PCR run, each having threereplicates.
Gene expression analyses of sugar metabolism in developing sorghum caryopses 337
SbIncw was many-fold higher than the expressionof ZmIncw1 ortholog at all the stages and intissues analyzed. The expression of Incw genes indeveloping sorghum caryopses was largely confinedto the lower sugar importing zone of the endo-sperm. Sucrose synthase isoform Sus3, anothergene involved in sucrose utilization in developingcereal endosperm, provided a good endogenousreference control for real-time transcript ana-lyses (Fig. 3E and F). Expression of Sus3 did notshow any significant temporal or spatial variabilityduring sorghum seed development, as was pre-
viously reported for maize endosperm (Carlsonet al., 2002).
Cell wall invertase activity and protein gelblot analyses in developing sorghum seeds
High CWI enzyme activity was seen in whole seedextracts at 6 and 12 DAP, and only a small fractionof it, 17% and 10%, persisted later at 18 and 24 DAPstages (Fig. 4A). The temporal reduction in CWIactivity was similar to that reported previously

ARTICLE IN PRESS
Cel
l wal
l inv
erta
se a
ctiv
ity(µ
mol
red
ucin
g su
gar
mg-
1 pr
otei
n m
in-1
)
0
2
4
6
8
10seedsglumes
6 DAP 12 DAP 18 DAP 24 DAP
A
6 DAP12 DAP
6 DAP12 DAP
seeds glumes
72 kDa
B
Figure 4. Cell wall invertase enzyme activity in 6–24 DAP(A) and protein gel blot analysis of CWI in 6 and 12 DAPsorghum seeds and glumes (B). The enzyme activity dataare presented as average7SD of three independentextractions with two assay replicates each. For proteingel blot analysis, 5mg protein extracted from seeds and10 mg protein from glumes were loaded per lane. Theprimary and secondary antibodies were used at 1:2000and 1:5000 dilutions, respectively.
M. Jain et al.338
during maize seed development (Cheng et al.,1996; Chourey et al., 2006). Notably, most of theCWI activity in 12 DAP seed was confined to thelower half section of the seed (7.38 mmol reducingsugarmg�1 proteinmin�1) and only a very smallfraction (0.92mmol reducing sugarmg�1proteinmin�1;12.5%) was extractable from the upper starchstorage region (data not shown). The decreasedCWI enzyme activity through endosperm develop-ment correlated well with the reduced transcriptabundance. However, a distinct lack of concor-dance between the transcript and protein contentmay be attributed to lower protein turnover in theglumes. A cell-specific intergenic cross-talk invol-ving post-transcriptional and post-translationalcontrol of ZmIncw1 and ZmIncw2 expression andINCW1 and INCW2 levels has been implicated duringendosperm maturation in maize (Chourey et al.2006). Figure 4B shows an immunoblot developedusing maize polyclonal INCW1 antiserum (Shankeret al., 1995) that does not discriminate the twoisoforms in maize (Carlson and Chourey, 1999;Chourey et al., 2006). Although a single polypep-tide of identical size (72 kDa) was seen in both seedand glumes, a possibility of more than one isoform
cannot be ruled out with in the detection limits ofthis assay since the SbIncw transcription is far moreabundant than the ZmIncw1 ortholog (Fig. 3).
Immunolocalization of INCW protein insorghum caryopses
Figure 5 depicts cell-specific localization of theCWIs in longitudinal sections of 10- and 16 DAPsorghum caryopses. As in maize (Cheng et al., 1996,Chourey et al., 2006), strong immunoreactivesignal was detectable in the BETC layer in 10 DAPkernels. Additionally, high level of INCW proteinwas also discernible in the maternal pedicel region,particularly in the basal vein that showed high levelof immunoreactivity at a larger dilution of theantibody (1:6000 as opposed to 1:2000). Also, therewas no detectable signal in upper endosperm regionof 10 or 16 DAP seeds, thus confirming, lowtranscript levels and CWI enzyme activity in upperparts of endosperm (Figs. 2B and 3).
RNA and protein gel blot analyses for genesinvolved in sugar-to-starch metabolictransition in developing sorghum caryopses
The transcript profiles of several other keyenzymes involved in sucrose utilization (sucrosesynthases; Sh1, Sus1 and Sus3) and sugar-to-starchmetabolism (Pgm2 (phosphoglucomutase), Sh2 andUGPase) in developing sorghum seeds were alsostudied in addition to cell wall and solubleinvertases. Sucrose synthases (Sh1, Sus1 andSus3), Pgm2 and Sh2 cDNA clones were amplifiedfrom 12 to 16 DAP maize endosperm and used asprobes for studying the respective ortholog expres-sion in developing sorghum seed (Fig. 6A and B).Shrunken1 (Sh1) mRNA was most abundant prior tofertilization and declined steadily through grainmaturation and starch filling phase. At 12 DAP, mostof the Sh1 expression was spatially restricted to theupper section of the seed, and the low-levelexpression in glumes evident at 6 DAP becameundetectable at 12 DAP. In contrast, Sus1 tran-scripts were readily detectable only in unfertilizedovules (0 DAP) and 6 DAP seeds (Fig. 2B), andlocalized to both lower and upper parts of theendosperm. RNA gel blot and real-time quantitativePCR analyses of Sus3 transcription (Fig. 3) did notshow any temporal or tissue specific variabilityduring sorghum seed development, as was pre-viously reported for maize endosperm (Carlson etal., 2002). During the development of sorghumseed, UGPase transcript levels increased from 0DAP to maximum at 24 DAP, declining later at 30

ARTICLE IN PRESS
A B C
FD E
em
en
pb ss
s
s
s
pp
p
p
p
b
en
en en
b
b
b
Figure 5. Immunolocalization of CWI protein in longitudinal sections of 10, 12 and 16 DAP sorghum caryopses usingmaize INCW1 antibodies. Primary antibody was used at 1:2000 dilution (B–E) and 1:6000 dilution (F). Pre-immunecontrol (1:2000 dilution), 10 DAP (A); 10 DAP showing INCW in filial BETC and maternal pedicel regions (B); 10 DAP (C);10 DAP showing pedicel (D); 16 DAP showing INCW in filial BETC and maternal pedicel regions (E) and 12 DAP showingINCW only in pedicel decorated with 1:6000 primary antibody dilution (F). Scale bars: 250 mm (A) and 200 mm (B–F). (b,BETC layer; em, embryo; en, endosperm; p, pedicel; s, placental sac).
Sh1
UGPase
Sus3
Sh2
Pgm2
Sus1
0 6 12 18 24 30 DAPA
Sus1
Sh1
Sus3
glumes
glumes
upperlow
ersee
d
6 DAP 12 DAPB
seeds glumes
6 12 18 24 6 12 18 24
UGPase
SH2
DAP
C
Figure 6. Temporal (A) and spatial (B) variability in the expression profiles of genes involved in sucrose cleavage andutilization in developing sorghum endosperm and glumes. All lanes were loaded with 15 mg total RNA. Completeunfertilized florets were used for RNA extraction at 0 DAP, and seeds without glumes were sampled for 6–30 DAP stages(A); and intact seeds were used for RNA extraction at 6 DAP, whereas the 12 DAP seeds were spliced in upper (storagezone) and lower (sugar transport zone) half sections (B). The RNA blot membranes used for hybridization in Fig. 3 werestripped and re-hybridized with respective probes. Protein gel blot analysis of UGPase and SH2 proteins in 6–24 DAPsorghum seeds and glumes (C). Ten mg total protein was resolved per lane. The primary antibodies for UGPase and SH2were used at 1:6000 and 1:5000 dilutions, respectively; and secondary antibodies were used at 1:5000 dilutions.
Gene expression analyses of sugar metabolism in developing sorghum caryopses 339
ARTICLE IN PRESS
M. Jain et al.340
DAP. Phosphoglucomutase (Pgm2) expression wasmost abundant at 6 DAP and declined at laterstages, whereas Sh2 expression coincided withstarch filling stages of seed development i.e. at12 and 18 DAP.
Protein gel blot analyses for UGPase and Sh2-encoded ADP-glucose pyrophosphorylase (AGPase)subunit in developing sorghum seeds and glumesare represented in Fig. 6C. Both UGPase and SH2proteins showed temporal expression patterns aswas predicted from the transcript level ana-lyses and maximum protein levels were associatedwith the starch filling phase of endosperm devel-opment, i.e from 12 to 24 DAP. The glumes atsimilar developmental stages showed very lowlevels of SH2, and UGPase levels were totallyundetectable.
µg
suga
rs /m
g fr
esh
wei
ght
0
2
4
6
8
10
12
14
16
18 glc
frc
suc
µg
star
ch/m
g fr
esh
wei
ght
0
50
100
150
200
250starch
seedglumes
seedglumes
see
6 DAP 12 DAP
Figure 7. Soluble sugar (A) and starch (B) content in glumes aalso analyzed for carbohydrate content in upper (storage zendosperm. The data presented are average7SD of threereplicates.
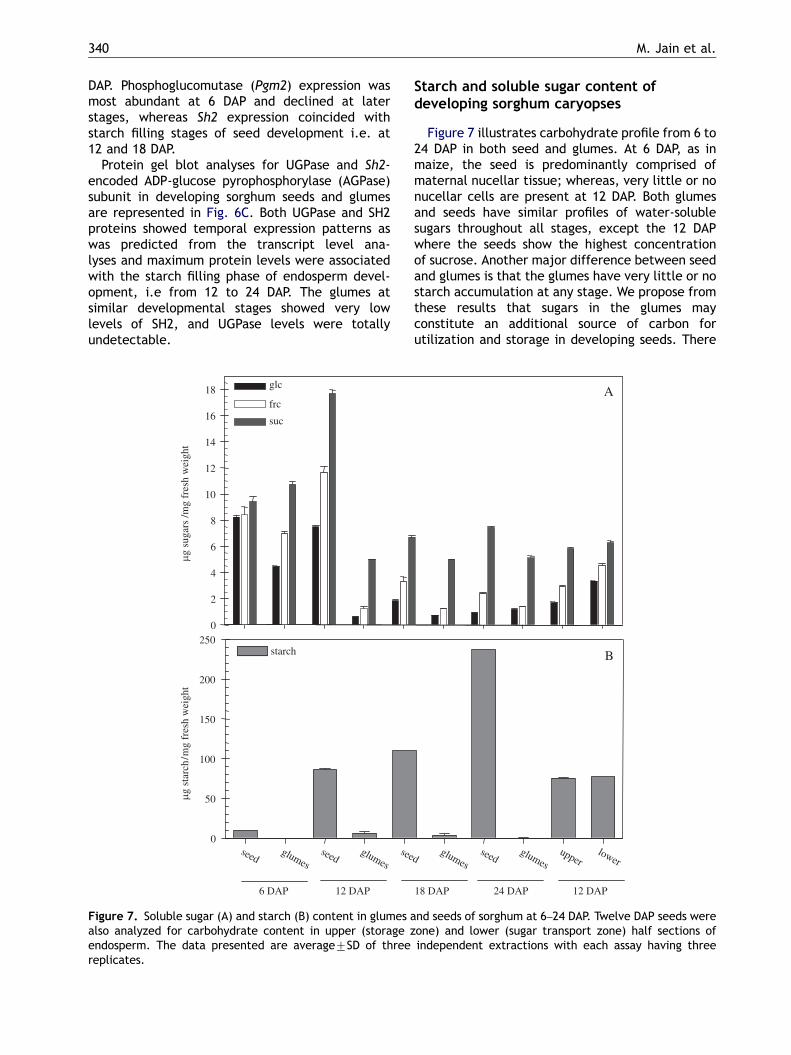
Starch and soluble sugar content ofdeveloping sorghum caryopses
Figure 7 illustrates carbohydrate profile from 6 to24 DAP in both seed and glumes. At 6 DAP, as inmaize, the seed is predominantly comprised ofmaternal nucellar tissue; whereas, very little or nonucellar cells are present at 12 DAP. Both glumesand seeds have similar profiles of water-solublesugars throughout all stages, except the 12 DAPwhere the seeds show the highest concentrationof sucrose. Another major difference between seedand glumes is that the glumes have very little or nostarch accumulation at any stage. We propose fromthese results that sugars in the glumes mayconstitute an additional source of carbon forutilization and storage in developing seeds. There
A
B
dglumes
seedglumes
upperlower
18 DAP 24 DAP 12 DAP
nd seeds of sorghum at 6–24 DAP. Twelve DAP seeds wereone) and lower (sugar transport zone) half sections ofindependent extractions with each assay having three

ARTICLE IN PRESS
Gene expression analyses of sugar metabolism in developing sorghum caryopses 341
was a steep increase in starch levels in seeds during18–24 DAP stage of development. High levels ofhexoses in the seeds at 6 and 12 DAP also coincidedwith the highest levels of CWI activity (Fig. 4A) ascompared to other stages. Likewise, high starchdeposition in seeds and almost negligible content inglumes correlated well with high steady-statelevels Sh2-encoded AGPase and UGPase RNA andprotein in 12–24 DAP seeds (Fig. 6A and C) and thelack of these two proteins in the glumes (Fig. 6C),respectively.
Discussion
Role of cell wall invertases duringdevelopment of sorghum caryopses
Temporal and spatial expression profiles ofSbIncw gene in developing sorghum seed (6–12DAP, in the filial BETC layer and maternal pedicelregion) correlate well with degeneration of nucel-lar tissue post-5-DAP and steep exponential growthof endosperm apparent during 8–12 DAP (Kladniket al., 2006). Thus, high transcript abundance,enzyme activity and immuno-localization data ofthe INCW protein primarily in the lower sugarimporting region of the developing sorghum seed,the single port of sugar-entry, are in close agree-ment with the proposed unique role of CWI in theestablishment and maintenance of developing sinkstrength in providing hexoses to the developingembryo and storage tissue in the cell division stageas reported previously in maize (Cheng andChourey, 1999), rice (Hirose et al., 2002) and favabean (Weber et al., 1996).
The SbIncw gene described here shows highhomology to ZmIncw2, and represents the predo-minant CWI expressed in the developing sorghumseed at 6 DAP. Despite the significant geneticrelatedness of sorghum and maize, the two plantspecies show strikingly dissimilar expression pro-files of Incw1 and Incw12 orthologs in the develop-ing seeds. The expression of SbIncw (ZmIncw2ortholog) in sorghum is confined to the basalsugar-importing region of the young 6–12 DAP seed,and the ZmIncw1 ortholog is expressed primarily inthe vegetative glumes of 6 DAP caryopses . Notably,the Incw transcript levels were below the detectionlimits in upper starchy storage region of 12 DAPendosperm, and the expression of SbIncw gene(ZmIncw2 ortholog) was several orders of magni-tude higher than ZmIncw1 ortholog throughoutsorghum seed development and glumes persis-tence. In sharp contrast, ZmIncw2 expression inmaize is restricted to the sugar-importing zone
during mitotic growth of the endosperm andembryo around 12 DAP and abundant expressionof ZmIncw1 is discernible in the upper starch-fillingpart of the endosperm later during the period ofkernel maturation and desiccation (Cheng et al.,1996; Chourey et al. 2006).
Low-level expression of ZmIncw1 ortholog wasdiscernible in glumes at 6 DAP, in addition to SbIvr2expression. It is conceivable that concerted cellwall and soluble invertase activities in photo-synthesizing glumes at 6 DAP in comparison tolower section of seed helps elevate the hexosecontent in glumes by sucrose inversion to supportenergy requirements for active growth, and anincrease in water potential aiding cellular expan-sion. The rapid increase in the size of glumes at thisstage keeps pace with the increase in seed size.High hexose-to-sucrose ratio in glumes at 6 DAP,and a dramatic reduction in the total soluble sugarpool in the glumes from 6 to 12 DAP supports thisargument. A similar role for vacuolar invertaseactivity has been proposed during rapid growthphase of juvenile petioles of sugar beet plants(Gonzalez et al., 2005).
Developing sorghum seed differs from othercereal seeds, including maize, in having a sac-likecavity, the placental sac, formed as a result of post-fertilization loss of nucellar cells in the placentalregion in the basal, abgerminal tissue. The placen-tal sac is speculated to serve as an apoplastic sinkfor hexoses in between the maternal base and thebasal transfer cells of the endosperm (Maness andMcBee, 1986). High CWI enzyme activity between 6and 12 DAP (present study) closely follows suit ofdifferentiation of BETC layer and appearance ofplacental sac (Maness and McBee, 1986) at 5 DAPand the beginning of rapid cell division phase ofendosperm growth (Kladnik et al., 2006). Interest-ingly, in sorghum the INCW protein is immunoloca-lized in the vein parenchyma of maternal pediceltissue and to a lesser extent in the filial BETCregion. The presence of INCW in the maternalpedicel region of sorghum caryopses is parallel tothat reported for in situ OsCIN1 mRNA localizationin vascular parenchyma of dorsal vein in young ricecaryopses (Hirose et al., 2002), whereas expressionof ZmIncw2 in maize is predominantly confined tofilial BETC region (Cheng et al., 1996). Maness andMcBee (1986) reported high concentrations ofglucose and fructose in the placental sac fluidsdeclining appreciably by 20 d post-anthesis,whereas the sucrose was always present at a lowconcentration remaining relatively constantthrough seed maturation. The detection of INCWprotein at both proximal and distal ends ofplacental sac is unique among cereal caryopses

ARTICLE IN PRESS
M. Jain et al.342
and is likely to be a beneficial trait for increasedsucrose unloading as well as water uptake/storageinto the developing seeds of sorghum.
Role of soluble invertases during earlydevelopment of sorghum seed
Soluble invertase SbIvr2 expression restricted to0–6 DAP in seeds consisting largely of maternalnucellar tissue (Kladnik et al., 2006) is in accor-dance with data published in maize (Carlson andChourey, 1999) and barley (Weschke et al., 2003),and thus rules out any participation of solubleinvertase activity in the establishment of filial sinkstrength. Infusion of sucrose into the maize stemsaround the pollination time resulted in droughtstress-induced transcriptional de-repression of Ivr2and Incw2 (in contrast to Incw1), and preventedstress-induced abortion of young ovaries (Anderson,2002; McLaughlin and Boyer, 2004). Sugar-respon-siveness and the distinct early temporal expressionof soluble invertase in maternal tissues of maizekernels indicate that Ivr2 maintains physiologicalreceptivity of ovary prior to fertilization. In sharpcontrast, association of Incw2 with the basalendosperm (sucrose-unloading zone) prior to thebeginning of assimilate import and storage phaseadjunct with development of filial sink tissuesensures reproductive success. The low-level, resi-dual soluble invertase activity (if any) may primar-ily be responsible only for providing hexoses for cellexpansion and growth in young developing seeds, asindicated by similar expression profiles of SbIvr2 inlower (sugar-unloading) as well as in upper (starch-storage) regions of 12 DAP seeds.
Sugar-to-starch transition in the developingsorghum seeds
Abundant steady-state levels of UGPase tran-script were observed throughout seed growth, witha significant increase in transcript abundanceevident during starch deposition stages of grainmaturation. In leaves, UGPase contributes tosucrose biosynthesis by providing UDP-glucose(UDPG) to sucrose-phosphate synthase, whereas insink tissues it participates in the breakdown ofsucrose using UDPG derived from sucrose synthasecatalysis. In cereal seed endosperm, the UDPGutilization is often coupled to the AGPase activityresulting in equimolar production of ADP-glucose(ADPG) from UDPG, ADPG being the sole immediateprecursor of starch biosynthesis (Kleczkowski et al.,2004). The formation of ADPG by AGPase is the firstcommitted step in the transfer of glucose moieties
into starch biosynthesis. In maize endosperm,AGPase is a heteromeric protein, encoded by twonon-allelic genes, Sh2 and Bt2 (Bhave et al., 1990).In young filial tissues, the requirement of UDPG forcell wall biosynthesis during active cell divisionphase may be fulfilled, directly by sucrose synthasereaction or partly through an invertase, andhexokinase- and /or PGM-dependent pathways forthe provision of glucose-1-P for UGPase-mediatedreaction. The role of Sh1-encoded SS1 isoform inproviding substrate moieties for cellulose biosynth-esis in maize (Chourey et al., 1998), and that ofRSs1-encoded RSuS2, a respective isoform in rice(Wang et al., 1999) is well documented. Macro-array-based transcript profiling also demonstratedan involvement of both sucrose synthase-3 (anortholog of maize Sh1) and invertase-driven path-ways for providing UDPG for early growth of barleyseeds (Sreenivasulu et al., 2004). High invertase,Sh1 and Pgm2 expression in young (6 and 12 DAP)and high Sh2 transcript and protein levels in latestages (12 and 24 DAP) prior to starch deposition insorghum seed (Figs. 3A and 7A, C, 8B) furthersupport and confirm the above argument.
Conclusions
In summary, we have described the expression ofCWI genes in developing sorghum caryopses. Ex-pression of CWI genes is spatially confined to BETCand pedicel tissue on either side of placental sac –
the sugar unloading region, and temporally prior tostarch deposition phase in the developing endo-sperm, as previously reported in several othercereal grains. The data in sorghum reaffirm thecrucial role of CWI-mediated cleavage of sucrose inthe early establishment of sink strength. Also, agradual shift from mitotic growth to the storagephase of growing seed is coincident with a shift inthe utilization of UDPG for cell wall biosynthesis inyoung seeds to starch accumulation upon maturity.Concerted UGPase and AGPase (Sh2 subunit)activities during storage phase corroborate thisobservation.
Acknowledgments
Mention of trade names or commercial productsin this publication is solely for the purpose ofproviding specific information and does not implyrecommendation or endorsement by the US Depart-ment of Agriculture. We thank J.R. Sovokinos,University of Minnesota and USDA-ARS for UGPasecDNA and antiserum, L.C. Hannah, University ofFlorida for the SH2 antibodies, Drs. Ed Exteberria,

ARTICLE IN PRESS
Gene expression analyses of sugar metabolism in developing sorghum caryopses 343
Sherry LeClere and Earl Taliercio for critical readingof the manuscript and Mr. A. Funk for excellenttechnical assistance. This was a cooperative in-vestigation of the US Department of Agriculture,Agricultural Research Service and the Instituteof Food and Agricultural Science, University ofFlorida.
Appendix A. Supplementary data
Supplementary data attached with this articlecan be found in the online version at doi:10.1016/j.jplph.2006.12.003.
References
Anderson MN, Asch F, Wu Y, Jensen CR, Naested H,Mogensen VO, et al. Soluble invertase expression is anearly target of drought stress during the critical,abortion-sensitive phase of young ovary developmentin maize. Plant Physiol 2002;130:591–604.
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG,Smith JA, et al. Current protocols in molecularbiology. New York: Wiley; 1993.
Bhave MR, Lawrence S, Barton C, Hannah LC. Identifica-tion and molecular characterization of Shrunken-2cDNA clones of maize. Plant Cell 1990;2:581–8.
Carlson SC, Chourey PS. A re-evaluation of the relativeroles of two invertases, INCW2 and IVR1, in developingmaize kernels and other tissues. Plant Physiol 1999;121:1025–35.
Carlson SJ, Chourey PS, Helentjaris T, Datta R. Geneexpression studies on developing kernels of maizesucrose synthase (SuSy) mutants show evidence for athird SuSy gene. Plant Mol Biol 2002;49:15–29.
Cheng WH, Chourey PS. Genetic evidence that invertasemediated release of hexoses is critical for appropriatecarbon partitioning and normal seed development inmaize. Theor Appl Genet 1999;98:485–95.
Cheng WH, Taliercio EW, Chourey PS. The Miniature1seed locus of maize encodes a cell wall invertaserequired for normal development of endosperm andmaternal cells in the pedicel. Plant Cell 1996;8:971–83.
Cho JI, Lee SK, Ko S, Kim HK, Jun SH, Lee YH, et al.Molecular cloning and expression analysis of the cellwall invertase gene family in rice (Oryza sativa L).Plant Cell Rep 2005;24:225–36.
Chourey PS, Taliercio EW, Carlson SJ, Ruan YL. Geneticevidence that the two isozymes of sucrose synthasepresent in developing maize endosperm are critical,one for cell wall integrity and the other for starchbiosynthesis. Mol Gen Genet 1998;259:88–96.
Chourey PS, Jain M, Li Q-B, Carlson SJ. Genetic control ofcell wall invertases in developing endosperm of maize.Planta 2006;223:159–67.
Datta R, Chourey PS, Pring DR, Tang HV. Gene expressionanalysis of sucrose-starch metabolism during pollenmaturation in cytoplasmic male-sterile and fertilelines of sorghum. Sexual Plant Reprod 2001;14:127–34.
Godt DE, Roitsch T. Regulation and tissue specificdistribution of mRNA for three extracellular invertaseisozymes in tomato suggests an important function inestablishing and maintaining sink metabolism. PlantPhysiol 1997;115:273–82.
Goetz M, Roitsch T. The different pH optima andsubstrate specificities of extracellular and vacuolarinvertases from plants are determined by a singleamino-acid substitution. Plant J 1999;20:707–11.
Gonzalez M-C, Roitsch T, Cejudo FJ. Circadian anddevelopmental regulation of vacuolar invertase ex-pression in petioles of sugar beet plants. Planta 2005;222:386–95.
Hirose T, Takano M, Terao T. Cell wall invertase indeveloping rice caryopsis: molecular cloning of OsCIN1and analysis of its expression in relation to its role ingrain filling. Plant Cell Physiol 2002;43:452–9.
Ishimaru T, Hirose T, Matsuda T, Goto A, Takahashi K,Sasaki H, et al. Expression patterns of genes en-coding carbohydrate-metabolizing enzymes andtheir relationship to grain filling in rice (Oryzasativa L.): comparison of caryopses located atdifferent positions in a panicle. Plant Cell Physiol2005;46:620–8.
Kerr PS, Rufty TW, Huber SC. Changes in non-structuralcarbohydrates in different parts of soybean (Glycinemax [L.] Merr.) plants during a light/dark cycle and inextended darkness. Plant Physiol 1985;78:576–81.
Kim JY, Mahe A, Guy S, Brangeon J, Roche O, Chourey PS,et al. Characterization of two members of the maizegene family, Incw3 and Incw4, encoding cell-wallinvertases. Gene 2000;245:89–102.
Kladnik A, Chourey PS, Pring DR, Dermastia M. Develop-ment of Sorghum bicolor caryopsis during the en-dosperm’s endoreduplication associated growthphase. J Cereal Sci 2006;43:209–15.
Kleczkowski LA, Geisler M, Ciereszko I, Johansson H. UDP-glucose pyrophosphorylase. An old protein with newtricks. Plant Physiol 2004;134:912–8.
Kresowich S, Barbazuk B, Bedell JA, et al. Towardssequencing the sorghum genome. A US NationalScience Foundation-sponsored workshop report. PlantPhysiol 2005;138:1898–902.
Laemmli UK. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature1970;227:680–5.
Maness NO, McBee GG. Role of placental sac inendosperm carbohydrate import in sorghum car-yopses. Crop Sci 1986;26:1201–7.
McLaughlin JE, Boyer JS. Sugar-responsive gene expres-sion, invertase activity, and senescence in abortingmaize ovaries at low water potentials. Ann Bot(London) 2004;94:675–89.
Miller ME, Chourey PS. The maize invertase-deficientminiature-1 seed mutation is associated with aberrant

ARTICLE IN PRESS
M. Jain et al.344
pedicel and endosperm development. Plant Cell1992;4:297–305.
Mullet JE, Klein RR, Klein PE. Sorghum bicolor – animportant species for comparative grass genomics anda source of beneficial genes for agriculture. Curr OpinPlant Biol 2001;5:118–21.
Roitsch T, Gonzalez M-C. Function and regulation of plantinvertases: sweet sensations. Trend Plant Sci 2004;9:606–13.
Rufty TW, Huber SC. Changes in starch formation andactivities of sucrose phosphate synthase and cytoplas-mic fructose-1,6-bisphosphatase in response tosource-sink alterations. Plant Physiol 1983;72:474–80.
Shanker S, Salazar RW, Taliercio EW, Chourey PS. Cloningand characterization of full-length cDNA encodingcell-wall invertase from maize. Plant Physiol 1995;108:873–4.
Sherson SM, Alford HL, Forbes SM, Wallace G, Smith SM.Roles of cell wall invertases and monosaccharidetransporters in the growth and development ofArabidopsis. J Exp Bot 2003;54:525–31.
Sonnewald U, Hajirezaei MR, Kossmann J, Heyer A,Trethewey RN, Willmitzer L. Increased potato tubersize resulting from apoplastic expression of a yeastinvertase. Nat Biotechnol 1997;15:794–7.
Sreenivasulu N, Altschmied L, Radchuk V, Gubatz S,Wobus U, Weschke W. Transcript profiles and deducedchanges of metabolic pathways in maternal and filialtissues of developing barley grains. Plant J 2004;37:539–53.
Taliercio EW, Kim JY, Mahe A, Shanker S, Choi J, ChengWH, et al. Isolation, characterization and expressionanalyses of two cell wall invertase genes in maize. JPlant Physiol 1999;155:197–204.
Tang GQ, Luscher M, Sturm A. Antisense repression ofvacuolar and cell wall invertase in transgenic carrotalters early plant development and sucrose partition-ing. Plant Cell 1999;11:177–89.
Tang HV, Pring DR, Shaw LC, Salazar FA, Muza FR, Yan B, etal. Transcript processing internal to a mitochondrialopen reading frame is correlated with fertility restora-tion in male-sterile sorghum. Plant J 1996;10:123–33.
Tsai CY, Salamini F, Nelson OE. Enzymes of carbohydratemetabolism in the developing endosperm of maize.Plant Physiol 1970;46:299–306.
Vilhar B, Kladnik A, Blejec A, Chourey PS, Dermastia M.Cytometrical evidence that the loss of seed weight inthe miniature1 seed mutant of maize is associatedwith reduced mitotic activity in the developingendosperm. Plant Physiol 2002;129:23–30.
Wadsworth GJ, Redinbaugh MG, Scandalios JG. A proce-dure for the small-scale isolation of plant RNA suitablefor RNA-blot analysis. Anal Biochem 1988;172:279–83.
Wang AY, Kao MH, Yang WH, Sayion Y, Liu LF, Lee PD, et al.Differentially and developmentally regulated expres-sion of three rice sucrose synthase genes. Plant CellPhysiol 1999;40:800–7.
Weber H, Buchner P, Borisjuk L, Wobus U. Sucrosemetabolism during cotyledon development of Viciafaba L. is controlled by the concerted action of bothsucrose-phosphate synthase and sucrose synthase:expression patterns, metabolic regulation and impli-cations for seed development. Plant J 1996;9:841–50.
Weschke W, Panitz R, Gubatz S, Wang Q, Radchuk R,Weber H, et al. The role of invertases and hexosetransporters in controlling sugar ratios in maternal andfilial tissues of barley caryopses during early develop-ment. Plant J 2003;33:395–411.






![THE POTENTIAL OF SWEET SORGHUM [Sorghum bicolor (L · The Potential of Sweet Sorghum [Sorghum Bicolor (L.) Moench] As A Bio- Resource for Syrup and Ethanol Production in Kenya EVANS](https://static.fdocuments.us/doc/165x107/5f0f52697e708231d44395b9/the-potential-of-sweet-sorghum-sorghum-bicolor-l-the-potential-of-sweet-sorghum.jpg)











