
Exploring the extracellular electron transfer potential of ... · 1 Exploring the extracellular...
12
1 Exploring the extracellular electron transfer potential of biochar with soil microbes Samuel Barnett Department of Microbiology, Cornell University Microbial Diversity Course, Marine Biological Laboratory Summer 2017 Abstract Biochar is an agriculture soil amendment that improves crop yields and soil health. One function that biochar may have is as a mediator of extracellular electron transfer, coupling microbial metabolism with distant electron acceptor reduction. This study aimed to enrich for soil bacteria capable of transferring electrons to biochar and to examine difference in oxygen profiles in biochar amended soils. Using acetate as an electron source in a microbial fuel cell, I enriched for a currently unidentified bacterium, however no strong electrical current was generated by these enrichment cultures. Spherical structures of possible bacterial origin were also found highly enriched on biochar from a microbial fuel cell using succinate as electron source. No differences were measured in oxygen profiles due to biochar in amended soils with either succinate or acetate as added electron sources. While this study is a first step, more thorough experiments are needed to confirm and describe extracellular electron transfer capability of biochar with soil microbes. Introduction Biochar is a soil amendment that, while used by indigenous people of South America and Africa for hundreds of years, has only recently been introduced into modern agriculture. Biochar is produced by the slow anoxic pyrolysis of organic matter such as wood chips or agriculture waste. The resulting material is highly enriched in pure carbon. Application of biochar to fields increases yields of various crops especially when used in conjunction with fertilizer and nutrient amendments. The growth promoting properties of biochar are not fully understood, but evidence points to its immense water holding and cation exchange capacities, as well as its ability to adsorb compounds from the soil such as metals, organic matter, and nutrients. Biochar can also be used as a carbon sequestration mechanism as it cannot be easily broken down by organisms and remains in soils for hundreds of years [1]. The pure carbon throughout biochar also makes the material able to conduct electricity similar to graphite used in basic microbial fuel cells. Microbial fuel cells (MFC) harness microbial metabolism to generate an electric current. In a basic MFC, bacteria metabolize a given substrate using the anode, often graphite or carbon paper, as the electron acceptor. The electrons from this process then pass to a cathode where a final electron sink, such as oxygen, is reduced. The current produced by the electron movement between anode and cathode can be used for various applications such as powering environmental monitoring equipment. Electron transfer in an MFC requires either direct electron transfer between the bacteria and the electrode or by means of mediators such as humic acids, quinones, or Fe(II)/Fe(III) [2]. This extracellular electron transfer (EET) is also important in environmental systems where either electron donor and acceptor may not be easily accessible to organisms.
Transcript of Exploring the extracellular electron transfer potential of ... · 1 Exploring the extracellular...

1
Exploringtheextracellularelectrontransferpotentialofbiocharwithsoilmicrobes
SamuelBarnettDepartmentofMicrobiology,CornellUniversity
MicrobialDiversityCourse,MarineBiologicalLaboratorySummer2017
Abstract Biocharisanagriculturesoilamendmentthatimprovescropyieldsandsoilhealth.Onefunctionthatbiocharmayhaveisasamediatorofextracellularelectrontransfer,couplingmicrobialmetabolismwithdistantelectronacceptorreduction.Thisstudyaimedtoenrichforsoilbacteriacapableoftransferringelectronstobiocharandtoexaminedifferenceinoxygenprofilesinbiocharamendedsoils.Usingacetateasanelectronsourceinamicrobialfuelcell,Ienrichedforacurrentlyunidentifiedbacterium,howevernostrongelectricalcurrentwasgeneratedbytheseenrichmentcultures.Sphericalstructuresofpossiblebacterialoriginwerealsofoundhighlyenrichedonbiocharfromamicrobialfuelcellusingsuccinateaselectronsource.Nodifferencesweremeasuredinoxygenprofilesduetobiocharinamendedsoilswitheithersuccinateoracetateasaddedelectronsources.Whilethisstudyisafirststep,morethoroughexperimentsareneededtoconfirmanddescribeextracellularelectrontransfercapabilityofbiocharwithsoilmicrobes.Introduction
Biocharisasoilamendmentthat,whileusedbyindigenouspeopleofSouthAmericaandAfricaforhundredsofyears,hasonlyrecentlybeenintroducedintomodernagriculture.Biocharisproducedbytheslowanoxicpyrolysisoforganicmattersuchaswoodchipsoragriculturewaste.Theresultingmaterialishighlyenrichedinpurecarbon.Applicationofbiochartofieldsincreasesyieldsofvariouscropsespeciallywhenusedinconjunctionwithfertilizerandnutrientamendments.Thegrowthpromotingpropertiesofbiochararenotfullyunderstood,butevidencepointstoitsimmensewaterholdingandcationexchangecapacities,aswellasitsabilitytoadsorbcompoundsfromthesoilsuchasmetals,organicmatter,andnutrients.Biocharcanalsobeusedasacarbonsequestrationmechanismasitcannotbeeasilybrokendownbyorganismsandremainsinsoilsforhundredsofyears[1].Thepurecarbonthroughoutbiocharalsomakesthematerialabletoconductelectricitysimilartographiteusedinbasicmicrobialfuelcells. Microbialfuelcells(MFC)harnessmicrobialmetabolismtogenerateanelectriccurrent.InabasicMFC,bacteriametabolizeagivensubstrateusingtheanode,oftengraphiteorcarbonpaper,astheelectronacceptor.Theelectronsfromthisprocessthenpasstoacathodewhereafinalelectronsink,suchasoxygen,isreduced.Thecurrentproducedbytheelectronmovementbetweenanodeandcathodecanbeusedforvariousapplicationssuchaspoweringenvironmentalmonitoringequipment.ElectrontransferinanMFCrequireseitherdirectelectrontransferbetweenthebacteriaandtheelectrodeorbymeansofmediatorssuchashumicacids,quinones,orFe(II)/Fe(III)[2].Thisextracellularelectrontransfer(EET)isalsoimportantinenvironmentalsystemswhereeitherelectrondonorandacceptormaynotbeeasilyaccessibletoorganisms.

2
MicrobescapableofEETcanbefoundinmanyenvironments,allofwhicharegoodinoculumsourcesforMFCs.SoilhasbeenshowntoharbormanyEETcapablemicrobessuchasmembersofthegenresGeobacter[3]andRhizobiales[4]andClostridium[5].InsituEETisbelievedtoutilizeelectronacceptorssuchasironoxides[5]andhumicacids[6].Duetoitspurecarbonchemicalmakeup,biocharhasalsobeensuggestedasapossibleelectronconduitinamendedsoils[7].PreviousstudieshavefoundthatitcanbeusedeffectivelyasananodeinMFCsinoculatedwithwastewatersludge[8].OthershaveusedbiochartoconnectShewanellaoneidensismetabolism,aknownEETcapablemicrobe,withFe(III)reduction[9].
Thegoalofthisstudyistofurtherexaminetheroleofbiocharasanelectronconduitinsoils.TodothisItookbothanMFCandmicrocosmapproachtoenrichformicrobescapableofusingbiocharassoleelectronacceptorandmeasureprofilesofoxygen,themainenvironmentalelectronacceptor,inbiocharamendedsoils.MethodsSoilandbiochar: SoilwascollectedfromafieldatCoonamessettFarminEastFalmouth,MA(41.6762˚N,70.5767˚W).ThefieldsitehadnotbeenplantedsincetheFallofthepreviousyearandonlycontainedasparsecoveringofvariousweeds.Soilwastakenfromthetop10cmandconsistedprimarilyoftheorganichorizon.Soilwaspassedthrougha2mmsieveandincubatedatroomtemperaturewithadepthof3cmfor5dayspriortosuccinatefuelcells,6dayspriortosuccinatemicrocosms,8dayspriortoacetatefuelcells,and11dayspriortoacetatemicrocosms. Twotypesofbiochar,CCandSC,wereusedforthisexperiment,bothofwhichwereobtainedfromBrianvonHerzenoftheClimateFoundation.CCbiocharisofcellulosicoriginwhichunderwentpyrolysisataround500˚C.SCbiocharisfromsawdustfeedstockpyrolyzedatamaximumof480˚CandoriginallyfromZHAWinWadenswil,Switzerland.MicrobialFuelCells: Biocharanodesweremadebyfillinga1mlpipettetipwithgranulatedbiochar(Figure1a).Holesweremadeinthesideofthetiptoallowbetterentryofanolytemedia.Abarecopperwirewasinsertedintowideendofthepipettetipintobiocharandthensealedwithsiliconerubber. SuccinateMFCswerebuiltaspicturedinFigure1b.Theanaerobicandaerobicvesselsconsistedofa250mLand500mLPyrexbottlerespectively.Eachchamberwassealedwitharubberstoppercontainingholesforelectrodesandsaltbridge.Succinateanolytemediacontainedperlitersolution:2.06gsodiumsuccinatedibasichexahydrate,0.31gammoniumchloride,0.13gpotassiumchloride,2.93gsodiumphosphatemonobasicmonohydrate,4.09gsodiumphosphatedibasicanhydrous,1mLtraceelementsolution,5mLMESbufferatpH6.15,and1mLvitaminsolution[8].A1%agarosein3Msodiumchloridesaltbridgespanningtheanaerobicandaerobicchamberswasmadebyinjectingthemoltensolutionintoa3mmtubeandsolidifying.Thecounterelectrodewasagraphiterod(AlphaAesar#14738)insertedintotheaerobicchamber.TheAg/AgClreferenceelectrode(BASiMF-2052)wasinsertedintotheanaerobicchamber.TheexperimentalMFCusedabiocharanodeastheworkingelectrodewhilethecontrolMFCusedanindiumtinoxide(ITO)coatedslide(SigmaAldrich#703184)

3
attachedtoanalligatorclipthatwasnotsubmergedintheanolytemedia.Aconstantnitrogengasflowthroughtheanaerobicchambersmaintainedananoxicenvironment.BothMFCswereinoculatedwith5gsoil.Chamberswerefilledwithenoughanolytemediatopartiallycoverelectrodesbutmaintainasmallheadspaceforthenitrogengas. Acetatebasedfuelcellsweresinglechambersconsistingof125mLPyrexbottlesasshowninFigure1c.Chambersweresealedwithrubberstopperswithholesforworking,counterandreferenceelectrodes.Acetateanolytemediacontainedperlitersolution2.5gsodiumacetateanhydrous,0.31gammoniumchloride,0.13gpotassiumchloride,2.93gsodiumphosphatemonobasicmonohydrate,4.09gsodiumphosphatedibasicanhydrous,1mLtraceelementsolution,5mLMESbufferatpH6.15,and1mLvitaminsolution[8].BothMFCsusedagraphitecounterelectrodeandAg/AgClreferenceelectrode.Aconstantnitrogengasflowthroughtheanaerobicchambersmaintainedananoxicenvironment.TheexperimentalMFCusedabiocharanodeastheworkingelectrodewhilethecontrolMFCusedasecondgraphiterod.BothMFCswereinoculatedwith5gsoil.Chamberswerefilledwithenoughanolytemediatopartiallycoverelectrodesbutmaintainasmallheadspaceforthenitrogengas.Acetateanolytemediawasremovedandreplacedbyfreshmediaafter4days.
Figure1:a)biocharanodes.b)succinateMFCwithbiocharanodeusingbothanaerobic(small)andaerobic(big)bottles.c)AcetateMFCwithbiocharanodeinoneanaerobicbottle.
MFCswerehookeduptoseparateGamryInstrumentInterface1000potentiostatsandcontrolledusingGamryInstrumentFrameworkversion6.20software.Duringrun,MFCsweremaintainedusingcontrolledpotentialcoulometryatapotentialof200mV,samplingtimeof1min,andmaximumcurrentof1mA.Priortorunandattheendofeachrunweperformedcyclicvoltammetrywithapotentialrangefrom-500to400mV,scanrateof100mV/s,stepsizeof2mV,maxcurrentof10mA,andpowerfactorcorrectionof50ohm.AcetatedMFCswererestarted6timesinthefirst4daysduetocomputercutoffs.CyclicvoltammetrywasperformedatthebeginningandendofeachcontrolledpotentialcoulometryrunofallMFCsandbeforeandafterreplacementofanolytemediaintheacetateMFCs.
a b c

4
Microcosmsetup: Allmicrocosmsweresetupin20x150mmHungatetubesandsealedwithParafilm.Therewereeightmicrocosmstomeasuresuccinateusebysoilmicroorganisms.Eachsuccinatemicrocosmcontained10mLlowersoilslurrytoppedby20mLuppersoilslurry.Succinatemicrocosmsetupisdescribedintable1.Succinatesolutioncontained1.02gsodiumsuccinatedibasichexahydratein500mLwater.Fourmicrocosmsweresetuptomeasureacetateuse.Acetatemicrocosmsetupisdescribedintable2.Acetatesolutioncontained1.25gsodiumacetateanhydrousin250mLwater.Forbothmicrocosms,after24hoursofincubation,excessliquidonsurfaceofsoilwasremovedpriortomicroelectrodemeasurements.
Table1:Succinatemicrocosmdesigns.
Table2:Acetatemicrocosmdesigns.
ScanningElectronMicroscopy: Biocharanode,copperwirefrombiocharanode,andITOslidefromsuccinateMFCswereimagedusingaZeissSUPRA40VPscanningelectronmicroscope.Sampleswerefixedin4%paraformaldehydeatroomtemperaturefor4hoursfollowedby4˚Cfor2hours.Theywerethenwashedthreetimesin1xPBS.Allsamplesweredehydratedby25minutesubmersionsof25%,50%,75%and100%ethanol.Sampleswerestoredin100%ethanoluntilcriticalpointdryingandplatinumcoating.HPLC:
LevelsofacetateinsamplesofanolytemediafromacetateMFCsandextractsfromtheacetatemicrocosmsweremeasuredusingUV/visdetectiononanHPLC(ShimadzuLC-2010C-HTwithBio-RadAminexHPX-87Hcolumn).Initialanolytesolutionwassampledafter3daysandagainfromreplenishedanolytesolutionafter7daysrunningintotal.Freshunusedanolytesolutionwasmeasuredforbaselineacetateconcentration.Soilextractsweremadeattheconclusionofthemicrocosmexperiment,day3.25mLpurewaterwasaddedtoeachmicrocosmandtheslurrywashomogenizedbyshakingandvortexingtubes.10mLofslurrywasthencentrifugedat4100Gfor10minutes.Soilextractwaspurifiedbyfiltering3mLofslurrysupernatantwasfilteredthrough0.2µmmembranefilter.Fluorescenceinsituhybridization: AnolytesolutionwithsuspendedcellaggregateswassampledfrominitialacetateMFCsafter4days.Samplewasfixedwith2%paraformaldehydeat4˚Cfor24hours.Afterfixing,

5
samplewaswashed3timesin1xPBSandpassedthrough0.2µmfiltertocapturecells.Twosubsampleswerehybridizedfor2hoursat46˚Cin30%formamidewith2setsofprobes,onesettargetingGammaproteobacteria(mixofDelta495a-CY3,Delta495b-CY3,andDelta495c-CY3)andtheothertargetingDeltaproteobacteria(GAM42a-CY3).Bothprobesetsalsoincludedagenericeubacterialprobe(EUB338-FITC).Afterhybridization,sampleswerewashedinstandardwashbuffer,PBSandmilliQwaterfollowedbydehydrationin96%ethanolandstainingwithDAPI.SampleswereimagedusingaZeissAxio-A2fluorescentmicroscope.Microelectrodeoxygenmeasurements: Oxygenprofilesweremeasuredinallmicrocosmsusinga1.1x40mmneedlesensoroxygenprobe(UnisenseOX-N)coupledwithaUnisensefieldmicrosensormultimeter.SensorswerecontrolledanddataanalyzedusingUnisenseSensorTracesoftwareversion2.8.200.21688.Probesmeasuredtoadepthof1cmwithastepsizeof100µm.Oncealldatawascollected,gradientsfromasinglesamplingdaywerenormalizedtoaccountforvaryingstartingdepths.ResultsMFCbasedenrichments: NopeakincurrentwasobservedinanyMFCovertheentiretimecourse.Therewasaconstantcurrentat6.25µAinthesuccinatebiocharMFC,5.08µAinthesuccinateITOMFC,4.71µAintheacetatebiocharMFC,and4.53µAintheacetategraphiteMFC.CyclicvoltammogramsforbothsuccinateMFCsdidnotchangebetweenthestartandendofrun.PlotsforthebiocharanodeinthesuccinateMFCandfirstmeasurementoftheacetateMFCweremostlylinearwithnodistinguishingpeaks.TheplotforthebiocharanodeintheacetateMFCchangedbetweenstartofrunandthepre-andpost-replacementofanolytemediawithpeaksappearingataround175mVagainsttheAg/AgClreference(Figure2a).Thispeakwentawayinthefinalcyclicvoltammogramwhenexperimentwasfinishedandtheplotbecamerelativelylinear.ThecyclicvoltammogramforthegraphiteelectrodeintheacetateMFCalsochangedbetweentimepointswithapeakat260mVinitially,-200mVpre-replacementofanolytemedia,and50mVinbothpost-replacementofanolytemediaandfinalreading,allagainstAg/AgClreference(Figure2b).SamplesoftheanolytemediaweretakenfromtheacetateMFCspriortoreplacementofmediaandattheendofthestudyformeasurementbyHPLCbutnostrongdecreaseinacetateconcentrationwasobservedwhencomparedtofreshanolytemedia.
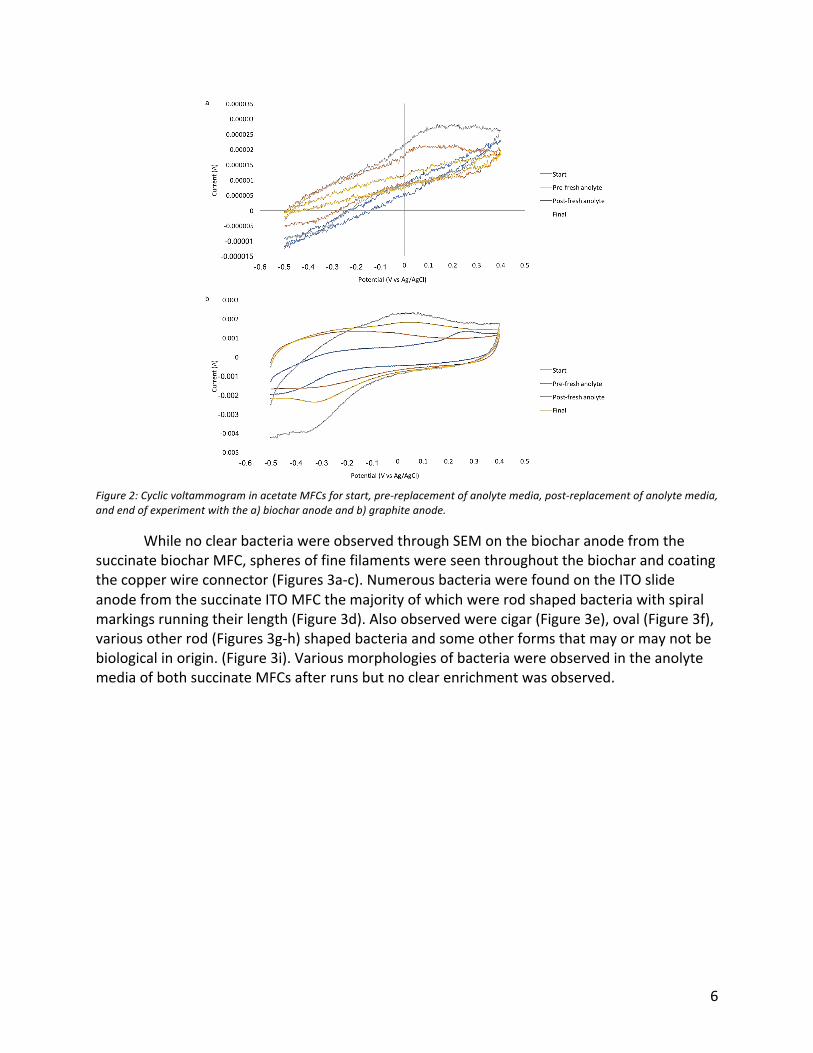
6
Figure2:CyclicvoltammograminacetateMFCsforstart,pre-replacementofanolytemedia,post-replacementofanolytemedia,andendofexperimentwiththea)biocharanodeandb)graphiteanode.
WhilenoclearbacteriawereobservedthroughSEMonthebiocharanodefromthesuccinatebiocharMFC,spheresoffinefilamentswereseenthroughoutthebiocharandcoatingthecopperwireconnector(Figures3a-c).NumerousbacteriawerefoundontheITOslideanodefromthesuccinateITOMFCthemajorityofwhichwererodshapedbacteriawithspiralmarkingsrunningtheirlength(Figure3d).Alsoobservedwerecigar(Figure3e),oval(Figure3f),variousotherrod(Figures3g-h)shapedbacteriaandsomeotherformsthatmayormaynotbebiologicalinorigin.(Figure3i).VariousmorphologiesofbacteriawereobservedintheanolytemediaofbothsuccinateMFCsafterrunsbutnoclearenrichmentwasobserved.

7
Figure3:SEMimagesofa)fibroussphereonbiocharanodefragmentfromasuccinateMFC,b)manyfibrousspheresonbiocharanodefragmentfromasuccinateMFC,c)fibrousspheresoncopperwirefrombiocharanodeinasuccinateMFCandbacteriaofvariousmorphologiesfoundontheITOslideanodefromasuccinateMFCincludingd)arodwithspiralpattern,e)cigar,f)oval,g)longerrod,h)roundedirregularrod,aswellasi)otherpossiblebiologicalstructures.

8
WhitewispyaggregatesofbacteriawerefoundfloatingandattachedtosurfacesintheacetatebiocharMFCpriortorefreshingofanolytemedia(Figure4a).Theaggregateswereformedbythickrodshapedbacteriacontainingnumerousphasebrightinclusionswithinthecells.Cellswerearrangedinclumpsandtwistedcords(Figure4b).NofluorescencewasobservedwithhybridizationofeitherDeltaproteobacteria(Figure5a)orGammaproteobacteria(Figure5b)FISHprobes,howeverfluorescencewasobservedwiththeeubacteriaprobeandDAPI.Nootherbacteriashowedclearenrichment.Whileaggregatescontinuedtobeseen,theenrichedbacteriawerenotseenaswidelyinanolytesamplesfromtherefreshedmediabiocharMFC.Aggregatesimagedunderphasecontrastmicroscopyappearedtobemadeupofcellulardebrisfromthepreviouslyenrichedbacteria.
Figure4:EnrichedbacteriainthesuccinateMFCwithbiocharanodeobservedasa)whitewispyaggregatesfloatingandattachedtoMFCsurfacesandasb)rodshapedbacteriaformingtwistedcordlikestructuresinphasecontrastmicroscopy
Figure5:FISHimageenrichedbacteriafromsuccinateMFCwithbiocharanodehybridizedwithprobesfora)Deltaproteobacteriaandb)GammaproteobacteriaalongwithEubacteriaandstainedwithDAPI.NotenofluorescencewitheithertheDeltaproteobacteriaorGammaproteobacteriaprobes.

9
Microcosmbasedcarbonutilization: Nodifferencewasobservedintheoxygenprofilesofanysuccinatemicrocosmsinanytimepoint(Figure6a-b).Nodifferencewasobservedintheoxygenprofilesoftheacetatemicrocosmsonday1(Figure7a),butthemicrocosmscontainingacetateonday3showedasteeperdecreaseinoxygenwithdepthascomparedtothenoacetatecontrols.Biocharshowednoinfluenceonthemicrocosmscontainingacetatebutseemedtoincreasetheslopeoftheoxygengradientinthenoacetatecontrols(Figure7b).AcetateconcentrationinsoilextractsweredeterminedattheconclusionoftheexperimentusingHPLCresultingin0.077mMand0.733mMinthe-biochar-acetateand+biochar-acetatemicrocosmsrespectivelyand7.758mMand8.084mMacetateinthe-biochar+acetateand+biochar+acetatemicrocosmsrespectively.
Figure6:Oxygenprofilesthroughfirst1cmofsuccinatemicrocosmsoils,a)measuredonday1andb)measuredonday3.
Figure7:Oxygenprofilesthroughfirst1cmofacetatemicrocosmsoils,a)measuredonday1andb)Measuredonday3.

10
Discussion Thisstudyaimedtoexaminetheabilityofbiochartoactasanelectronconduitforsoilmicrobes.WhilenoincreaseincurrentwasobservedintheMFCs,theincreasedsignalinthecyclicvoltammogramoverMFCruntimesuggestachangeinelectrontransferkineticsofthebiocharelectrode.Twopossiblecausesforthiscouldbechangeinanolytesolutionorchangetoanodecomposition.Replacementoftheanolytesolutionresultedinnochangetothecyclicvoltammogramwhencomparedtoplotcreatedjustpriortoreplacement.Thissuggeststhatitisamodificationtotheanodeitselfthatresultsintheelectrontransferkinetics.Twopossibleanodemodificationsincludecompletesaturationofanodebyanolytemediaorbuildupofbacterialbiofilmonthesurfacesoftheanodes.Thehighsurfaceareaandporespaceofbiocharcouldallowformoreanolytemediatocomeintocontactwiththeanodesurfaceovertime,increasingtheinteractionbetweenanolyteionsandtheanode[9].ThiscouldalsoexplainthehigherconstantcurrentobservedinthebiocharanodescomparedtographiteorITOslides.WhenreplacingtheanolytemediaintheacetateMFCs,theoldmediawithintheanodewouldlargelytransferwiththeanodeitself,keepingtheincreasedconductivitywhenthepotentialwasrestarted.However,thisshiftincyclicvoltammetrywasnotseeninthesuccinatebiocharMFC,wherethissaturationeffectshouldalsooccur.Biofilmdevelopmentontheanodeisanotherpossibilityfortheshiftinelectrontransferkineticsasenrichedbacteriawasobservedintheanodeduringtherun.Previousstudieshaveshownbiofilmdevelopmentincreasingcyclicvoltammetrysignalwithcarbonpaperanodes[10].Thelossofthepeaksinthecyclicvoltammogramattheendoftheexperimentcouldthenbeduetotheapparentlossoftheenrichedbacteria.Furtherstudiesshouldexaminetheextentofbiofilmdevelopmentonorbacterialinteractionwithbiocharanodestoseeifbacteriaareindeedthecauseoftheshiftcyclicvoltammograms. ThisstudywasabletoenrichforasinglebacterialtypeintheacetatebiocharMFCs.IwasunabletoidentifythebacteriabutnegativeresultsfromtheFISHexperimentsuggestthatitisnotamemberofeitherGammaproteobacteriaorDeltaproteobacteria,whichcontainknownEETcapablebacteriaShewanellaandGeobacterrespectively.Moreexperimentationincludingisolationandsequencingwillbeneededtoidentifythisbacterium.ItisalsounknownifthisbacteriumdirectlyinteractswiththebiocharanodeorconductsEET.Whileclustersofthebacteriawerefoundattachedtotheanode,theywerealsofoundfloatingintheanolytemediaaswellasattachedtothecounterandreferenceelectrodes.Thetwistedcordlikestructureofsomeofthebacterialaggregatesisalsointriguingandshouldbeexaminedinmoredetail.Thepoorhealthofthisenrichmentafterreplacementoftheanolytemediasuggestsacompoundinthesoilitselfwasrequiredforbacterialgrowth.UnderstandingthemicronutrientsinsoilswillbeimportantforfurtherenrichmentofsoilmicroorganismsandindevelopmentofmoreefficientMFCs.SEMimagesofbiocharandthecopperwirefromthesuccinatebiocharMFC,whilenotproducingclearsignsofbacteria,doshowinterestingstructuresthatmaybeofbacterialorigin.Thesefibrousspheresareinhighconcentrationonsomeofthebiocharfragmentsaswellasthecopperwire,suggestingacloserelationshiptotheelectronpotentialofthesesurfacesintheworkingMFC.Properidentificationandassessmentofthesestructuresisneeded. AccordingtoSEMimages,rod-shapedbacteriaalsoseemedtobeenrichedontheITOslideanodeofthesuccinateMFC.Thesebacteriahadaspiralgroovedstructurerunningthe

11
lengthofthecells.Itispossiblethatthesestructureswerecausedbythedehydrationandfixingprocesseshoweverfurtherimagingisneededtoexaminethenatureofthiscellmorphology.ContinuedculturingandlongerrunswiththisMFCarealsoneededtoconfirmtheconnectionbetweenthebacteriaandtheanode. Thisstudydidnotfindanymajordifferenceinoxygen-depthprofilesduetobiochar,exceptpossiblyanincreaseinslopeoftheoxygengradientinthebiocharamendedsoiloftheno-acetatemicrocosmascomparedtothenon-amendedsoil.Additionofacetatedoesshowasharpincreaseinoxygengradientsloperegardlessofbiocharaddition.Thisismostlikelyduetoincreasedmicrobialmetabolismfromexcesscarbonsource.Inordertofullytesttheelectrontransfercapabilityofbiocharinsoilitisimportanttomeasureconductivitythroughoutasoilprofilewithbiocharamendmentsandtoremoveorreduceanyothersourceofelectronflowotherthanbiochar.Useofbiocharembeddedmembranesmadeofnon-conductivematerialsuchasNafioncouldaccomplishthis.ExperimentswithopenairMFCssuchasthoseusedinWolińskaetal.[11]orJiangetal.[5]orinsituMFCscouldalsobeuseful.Conclusions Biocharhasthepotentialforbeinganelectronconduitformicroorganismsinanagriculturalsoilenvironment.ItsuseasananodeforMFCssuggeststhatmicrobescanuseitasanelectronacceptorandcoupletheirmetabolismswithreductionreactionsawayfromthecell.Moreworkneedstobedonetoshowthatthisisoccurringinsituanduncoverthemechanismsbehindtheprocess.IfEETisoccurringwithbiochar,thisprocesscouldexplainonebeneficialfunctionthisamendmenthasintreatedsoil.Acknowledgements IwouldliketothankDianneNewmanandJaredLeadbetterfortheirfantasticdirectionofthiscourseaswellasKurtHanselmann,ScottSaunders,BingranCheng,GeorgiaSquyres,andViolaKrukenbergforcloseassistanceandadviceonthisproject.Iwouldalsoliketothankalltheotherfaculty,TAsandlecturerswhohavecontributedtotherunningofthiscourseandthelaboratory.ThisprojectwouldnotbepossiblewithoutassistancefromthestaffatCoonamessettFarmforallowingmetosampletheirsoil,BrianvonHerzenfromtheClimateFoundationforhisdonationofbiochar,Unisenseforlendingusthemicroelectrodesetup,theGirguisLabatHarvardUniversityforlendingustheirpotentiostats,Zeissforlendingusfluorescentmicroscopes,andtheMBLCentralMicroscopyFacilityfortheirhelpwithSEMprepandimaging.ThefundingallowingmetoattendthiscoursecamefromtheSimonsMDScholarshipaswellascontributionfromtheBuckleyLabatCornellUniversity.References
1. Sohi,S.P.,Krull,E.,Lopez-Capel,E.,Bol,R.(2010)AReviewofBiocharandItsUseandFunctioninSoil,AdvancesinAgronomy105:47-82.
2. Du,Z.,Li,H.,Gu,T.(2007)Astateoftheartreviewonmicrobialfuelcells:Apromisingtechnologyforwastewatertreatmentandbioenergy,BiotechnologyAdvances,25:464-482.

12
3. Yates,M.D.,Kiely,P.D.,Call,D.F.,Rismani-Yazdi,H.,Bibby,K.,Peccia,J.,ReganJ.M.,Logan,B.E.(2012).Convergentdevelopmentofanodicbacterialcommunitiesinmicrobialfuelcells.TheISMEJournal,6:2002-2013.
4. Ishii,S.,Shimoyama,T.,Hotta,Y.,Watanabe,K.(2008)Characterizationofafilamentousbiofilmcommunityestablishedinacellulose-fedmicrobialfuelcell.BMCMicrobiology,8:6.
5. Jiang,Y.B.,Zhong,W.H.,Han,C.,Deng,H.(2016).CharacterizationofElectricityGeneratedbySoilinMicrobialFuelCellsandtheIsolationofSoilSourceExoelectrogenicBacteria.FrontiersinMicrobiology,7:1776.
6. Lovley,D.R.,CoatesJ.D.,Blunt-Harris,E.L.,Phillips,E.J.P.,WoodwardJ.C.(1996)HumicSubstancesasElectronAcceptorsforMicrobialRespiration.Nature382:445-48.
7. Sun,T.,Levin,B.D.A.,Guzman,J.J.L.,Enders,A.,Muller,D.A.,Angenent,L.T.,Lehmann,J.(2017).Rapidelectrontransferbythecarbonmatrixinnaturalpyrogeniccarbon.NatureCommunications,8:14873.
8. Huggins,T.,Wang,H.,Kearns,J.,Jenkins,P.,Ren,Z.J.(2014)Biocharasasustainableelectrodematerialforelectricityproductioninmicrobialfuelcells.BioresourceTechnology,157:114-119.
9. Friedl,J.,Stimming,U.(2017)DeterminingElectronTransferKineticsatPorousElectrodes.ElectrochimicaActa,227:235-245.
10. Liu,H.,Cheng,S.,Logan,B.E.(2005)ProductionofElectricityfromAcetateorButyrateUsingaSingle-ChamberMicrobialFuelCell.EnvironmentalScience&Technology39:658-662.
11. Woliaska,A.,Stapniewska,Z.,Bielecka,A.,Ciepielski,J.(2014)BioelectricityProductionfromSoilUsingMicrobialFuelCells.AppliedBiochemistryandBiotechnology173:2287-296.


















