
Experimental dmna induced hepatic necrosis: Early course of haemostatic disorders in the rat
8
Camp. Eiochem. Physiol. Vol. XOC. No. 2.pp.277-284, 1985 Printedin Great Britain 0306-4492/85 S3.00 +O.OO C 1985PergamonPress Ltd EXPERIMENTAL DMNA INDUCED HEPATIC NECROSIS: EARLY COURSE OF HAEMOSTATIC DISORDERS IN THE RAT* BERNARD CORNILLON, JACQUELINE PAUL, JEAN BELLEVILLE, ANNE MARIE AUROUSSEAU, GUY CLENDINNEN and ROSY ELOY Unit 37, Cardiovascular Surgery and Organ Transplantation Laboratory, Institut National de la Santt et de la Recherche Mtdicale, 18 avenue du Doyen L&pine, 69500 Bron, France (Received 8 Augusf 1984) Abstract-l. The course of haemostasis defects was investigated in dimethylnitrosamine (DMNA) acute liver necrosis. 2. Before 18 hr there was no evidence of disseminated intravascular coagulation (DIC) nor of abnormal fibrinolysis. 3. At 12 hr the level of the vitamin-K-dependent factors (factors II, VII, IX and X) was reduced to 25-63x of control. 4. Factors V and VIII : C levels decreased to about 10 and 20% by 12 hr. Factor V was the only molecule which decreased significantly by 6 hr. 5. The rapid decrease of these proteins might be related to an early parenchymal functional impairment attested by early structural lesions observed in the endoplasmic reticulum and nucleus. 6. The isolated decrease of factor V in the absence of any significant change in serum transaminase @GOT and SGPT) levels is proposed, at least in the rat, as an early criterion of hepatic failure. INTRODUCTION Major defects in coagulation were found as a compli- cation of acute hepatic necrosis in more than half the adults and 70% of the children studied by Saunders et al. (1979). It is generally considered that there is good correlation between coagulation abnormalities and the degree of liver failure (Chisholm, 1979) and thus the prothrombin time (PT) and partial throm- boplastine time (PTT) are used routinely to screen patients with liver disease. These tests may be consid- ered as liver function tests and may occasionally be abnormal while other biochemical tests are within normal limits. It is valuable, therefore, to evaluate the relative importance of criteria commonly used to evaluate acute hepatic failure and the specific assays of the different blood coagulation proteins which are now available. It is believed that serum amino- transferases are sensitive manifestations of hepatic injury but their level is not useful in predicting the onset of fulminant hepatic failure (Rosoff and Rosoff, 1977). A good correlation has, however, been shown between factor VII levels, prothrombin time and serum albumin in patients with liver disease (Green ef al., 1976) but factor VII assay is mostly considered as a prognostic criterion (Dimock et al., 1975). Rake ef al. (1973) showed that abnormalities in one stage coagulation tests and increased catabolism of fibrinogen in experimental hepatic necrosis in rats were similar to those found in acute viral and drug induced hepatic necrosis in man. The present study to assay blood coagulation factors in experimental he- patic necrosis has been set up to answer the following: *This work was supported by D.G.R.S.T. grant No. 80.7.6017. (1) what might be the earliest criterion of liver dysfunction and (2) what is the relative importance of the three pathogenic features that can lead to clotting abnormalities in the course of hepatic failure, viz (a) decreased protein factor synthesis (b) increased utilization of consumable factors or (c) excessive proteolysis and fibrinolysis (Roberts and Cederbaum, 1972). MATERIALS AND METHODS Dimethylnitrosamine (DMNA) was purchased from Merck Schuchardt. A solution of DMNA 1.6x, glucose 5%, ethanol 1% at a dose of 80mg DMNAjkg body wt was injected i.v. in Wistar rats (200 g wt) under ether ana- esthesia. Control animals were injected with the same solution minus DMNA. Animals were sequentially sacrificed in groups of eight to 17 animals in both experi- mental and control groups under ether anaesthesia at the 6th, l2th, 18th, and 24th hr following injection. Blood was collected by puncture of the infrarenal aorta directly into (1) glass tubes (1 ml) for determination of serum enzyme levels, (2) plastic tubes containing l/l0 volume of citrate solution for platelet count, coagulation assays and lysis time determination and (3) tubes containing 3600 NFU of soya bean trypsin inhibitors and 20NIH U of bovine thrombin per 2 ml of blood for fibrin degradation product (FDP) assays. Platelet counts of the whole blood were evaluated on a Coulter Counter (Model S). Fibrino- gen was assayed by the method of Clauss (1957) using bovine thrombin. Factor II, factor V, factor VII, factor VIII:C, factor IX and factor X were tested by classic one-stage assays using depleted (STAGO) or deficient (DADE) plasma (Alexander et al., 1956; Owren, 1947; Hardisty and McPherson, 1962; Bachman et al., 1958). The antithrombin III (AT III) level was determined spec- trophotometrically using chromogenic substrate (S 2238) according to Odegard (1975), fibrin degradation products (FDP) were measured according to Merskey ef al. (1969) 277
Transcript of Experimental dmna induced hepatic necrosis: Early course of haemostatic disorders in the rat

Camp. Eiochem. Physiol. Vol. XOC. No. 2. pp. 277-284, 1985 Printed in Great Britain
0306-4492/85 S3.00 +O.OO C 1985 Pergamon Press Ltd
EXPERIMENTAL DMNA INDUCED HEPATIC
NECROSIS: EARLY COURSE OF HAEMOSTATIC
DISORDERS IN THE RAT*
BERNARD CORNILLON, JACQUELINE PAUL, JEAN BELLEVILLE,
ANNE MARIE AUROUSSEAU, GUY CLENDINNEN and ROSY ELOY
Unit 37, Cardiovascular Surgery and Organ Transplantation Laboratory, Institut National de la Santt et de la Recherche Mtdicale, 18 avenue du Doyen L&pine, 69500 Bron, France
(Received 8 Augusf 1984)
Abstract-l. The course of haemostasis defects was investigated in dimethylnitrosamine (DMNA) acute
liver necrosis. 2. Before 18 hr there was no evidence of disseminated intravascular coagulation (DIC) nor of abnormal
fibrinolysis. 3. At 12 hr the level of the vitamin-K-dependent factors (factors II, VII, IX and X) was reduced to
25-63x of control. 4. Factors V and VIII : C levels decreased to about 10 and 20% by 12 hr. Factor V was the only molecule
which decreased significantly by 6 hr. 5. The rapid decrease of these proteins might be related to an early parenchymal functional impairment
attested by early structural lesions observed in the endoplasmic reticulum and nucleus. 6. The isolated decrease of factor V in the absence of any significant change in serum transaminase
@GOT and SGPT) levels is proposed, at least in the rat, as an early criterion of hepatic failure.
INTRODUCTION
Major defects in coagulation were found as a compli- cation of acute hepatic necrosis in more than half the adults and 70% of the children studied by Saunders et al. (1979). It is generally considered that there is good correlation between coagulation abnormalities and the degree of liver failure (Chisholm, 1979) and thus the prothrombin time (PT) and partial throm- boplastine time (PTT) are used routinely to screen patients with liver disease. These tests may be consid- ered as liver function tests and may occasionally be abnormal while other biochemical tests are within normal limits. It is valuable, therefore, to evaluate the relative importance of criteria commonly used to evaluate acute hepatic failure and the specific assays of the different blood coagulation proteins which are now available. It is believed that serum amino- transferases are sensitive manifestations of hepatic injury but their level is not useful in predicting the onset of fulminant hepatic failure (Rosoff and Rosoff, 1977). A good correlation has, however, been shown between factor VII levels, prothrombin time and serum albumin in patients with liver disease (Green ef al., 1976) but factor VII assay is mostly considered as a prognostic criterion (Dimock et al., 1975). Rake ef al. (1973) showed that abnormalities in one stage coagulation tests and increased catabolism of fibrinogen in experimental hepatic necrosis in rats
were similar to those found in acute viral and drug induced hepatic necrosis in man. The present study to assay blood coagulation factors in experimental he- patic necrosis has been set up to answer the following:
*This work was supported by D.G.R.S.T. grant No. 80.7.6017.
(1) what might be the earliest criterion of liver dysfunction and (2) what is the relative importance of the three pathogenic features that can lead to clotting abnormalities in the course of hepatic failure, viz (a) decreased protein factor synthesis (b) increased utilization of consumable factors or (c) excessive proteolysis and fibrinolysis (Roberts and Cederbaum, 1972).
MATERIALS AND METHODS
Dimethylnitrosamine (DMNA) was purchased from Merck Schuchardt. A solution of DMNA 1.6x, glucose 5%, ethanol 1% at a dose of 80mg DMNAjkg body wt was injected i.v. in Wistar rats (200 g wt) under ether ana- esthesia. Control animals were injected with the same solution minus DMNA. Animals were sequentially sacrificed in groups of eight to 17 animals in both experi- mental and control groups under ether anaesthesia at the 6th, l2th, 18th, and 24th hr following injection.
Blood was collected by puncture of the infrarenal aorta directly into (1) glass tubes (1 ml) for determination of serum enzyme levels, (2) plastic tubes containing l/l0 volume of citrate solution for platelet count, coagulation assays and lysis time determination and (3) tubes containing 3600 NFU of soya bean trypsin inhibitors and 20NIH U of bovine thrombin per 2 ml of blood for fibrin degradation product (FDP) assays. Platelet counts of the whole blood were evaluated on a Coulter Counter (Model S). Fibrino- gen was assayed by the method of Clauss (1957) using bovine thrombin. Factor II, factor V, factor VII, factor VIII:C, factor IX and factor X were tested by classic one-stage assays using depleted (STAGO) or deficient (DADE) plasma (Alexander et al., 1956; Owren, 1947; Hardisty and McPherson, 1962; Bachman et al., 1958). The antithrombin III (AT III) level was determined spec- trophotometrically using chromogenic substrate (S 2238) according to Odegard (1975), fibrin degradation products (FDP) were measured according to Merskey ef al. (1969)
277

278 BERNARD CORNILLON et al.
POST INJECTION TIME (hours)
Fig. 1. Course of vitamin-K-dependent coagulation factor II, factor VII, factor X and factor IX after DMNA administration. Results are expressed as a percentage of control values. Values are statistically
different from controls with a P value of less than 0.05 (*) or less than 0.01 (**).
and the euglobulin lysis time (ELT) was determined accord- ing to Dechaud et al. (1978). Human plasma was used as coagulation standard. The presence of factor VIII : WW was detected using Dodds’ procedure (1978) by agglutination of human washed platelets to ristocetin sulphate. Briefly, human platelets were washed twice in Tris 15 mM, pH 7.4, NaCl 137mM, KC1 5mM and EDTA 27mM. They were resuspended in the same solution minus EDTA. The agglu- tination medium included 200 ~1 of human washed platelet rich plasma (around 350 x lb9/l platelets), 50~1 of human washed plasma, 150~1 of Michaelis’ buffer pH 7.3 and 0.6 mg of ristocetin sulphate for induction. Aggregation was performed on a Dual 440 Chronologaggregometer (Coultronics) connected to a 2 pen Houston Omniscribe chart recorder. The evaluation. was based on the aggre- gation velocity expressed in mm per min. The serum levels of transaminase (SGOT and SGPT) were measured by classic methods.
Hepatocellular lesions were investigated by optic and by transmission electronic microscopy (TEM). For histology, liver biopsies were fixed in Bouin’s solution, dehydrated through graded ethanol solutions and embedded in paraffin. Slices 5 pm thick were stained with Hemalum-Phloxine- Safran solution. For TEM, biopsies were immediately trans- ferred to fresh 4% glutaraldehyde, then post-fixed in 1% osmium tetroxyde, dehydrated in graded ethanol and em- bedded in Epon. After thin section, they were mounted on copper grids and examined on a Phillips EM 300 electron microscope.
Student’s t test was used for the statistical analysis of differences between experimental and control groups.
RESULTS
There was no significant variation between the measurements in the different sham groups, so these results were pooled to form the control group. Changes in the vitamin-K-dependent factors (factor II, factor VII, factor IX and factor X) are shown in Fig. 1 and expressed as a percentage of control levels. There was no change at 6 hr but levels decreased between the 6th and the 12th hr. The most drastic drops were in factor X and factor IX which were reduced to 25 and 44%, respectively, at the 12th hr compared with 50 and 63% for factor VII and factor II. At the 24th hr they were 6, 18, 14 and 10% of control levels respectively.
Factor V and factor VIII:C levels (Fig. 2) de- creased rapidly to about IO and 20% of the control values by 12 hr and to 1 and 2%, respectively, at 24 hr. Factor V was the only substance which was significantly decreased by the 6th hr (67”/, of control value).
The development of consumption coagulopathy is illustrated in Figs 3 and 4. Until the 18th hr there was no significant variation of fibrinogen and platelet count but by 24 hr the levels were, respectively, 18 and 17% of control values. After an initial rise of 36% at 6 hr AT III level decreased significantly to reach 5% at the 24th hr. Even at 24 hr there was no significant change in euglobulin lysis time (representing systemic fibrinolytic activity), FDP ievels were constant at first but rose significantly by 24 hr to 575% of the con-
FACTOR V
POST INJECTION TIME (hours)
Fig. 2. Course of factor V and factor VIII:C levels after DMNA administration. Results are expressed as in Fig. 1.

DMNA induced hepatic necrosis 279
- I-‘~,1 50. PLAlELEY COUNT
\
mm 4 . .
*
m y+---‘1
so. FIBRINOGEN
2,
10 1 \r . .
Fig. 3. Course of criteria belonging to consumption coagu- lopathy syndrome: platelet count, fibrinogen and AT III.
Results are expressed as in Fig. 1.
trols. This increase in FDP and decrease in fibrinogen, platelet count and AT III suggest that disseminated intravascular coagulopathy appears between the 18th and 24th hr following DMNA injection.
Serum GPT levels (Fig. 5) were unchanged until the 18th hr. Then they increased rapidly so that by the 24th hr following administration of DMNA the mean level was 2560 IUjl(54 IUjl in control animals). Serum GOT level showed a slight but significant increase by 12 hr with a considerable further increase by 24 hr to 4230 IUjl (194 III/l in controls).
In rat plasma, ristocetin cofactor activity of factor VIII : VW was detected until the 18th hr (Table 1). At this time, aggregation velocity was slightly but significantly reduced. By 24 hr, the plasma proteins were precipitated by ristocetin.
Limited centrizonal necrosis characterized by an acidophilic degeneration was seen 6 hr after injection of DMNA. Cells in the mid zone of the lobule became vacuolated. Sinusoids were dilated around the centrilobular vein. These changes initially affected about 20% of the diameter of the lobule but had increased by 12 hr to affect 50% of the lobule. They were still maximal around the centrilobular vein. Hepatocytes showed loss of glycogen, increased acid- ophilic degeneration and ballooning. At this time, some free acidophilic bodies were seen in the mid- zone. At 18 hr less than 20% of the cells appeared
/ II --, -f
100 El1
x0- 0
POST INJECTION TIME (hours)
Fig. 4. Course of criteria belonging to consumption coagu- lopathy syndrome and/or excessive fibrinolysis: euglobulin lysis time (ELT) and fibrin degradation products (FDP).
Results are expressed as in Fig. 1.
normal. The central zone consisted only of blood filled spaces and debris of necrotic hepatocytes. The endothelial lining of the central veins and sublobular branches appeared swollen, or even detached. By 24 hr the necrosis of the centrilobular and midzone was general. The necrotic areas were confluent and haemorrhage into them was frequent. The toxic
tpop 0
I’..
I
5 GOT
/ _._.,/+--- **
SGPl
POST INJECTION TIME (hours)
Fig. 5. Course of serum transaminase (SCOT and SGPT) activities expressed in W/l after DMNA administration. SGOT and SGPT levels in control animals were 194 and
54 W/l, respectively.

280 BERNARD CORNILLON et al.
Table 1. Factor VIII:vW using ristocetin- induced agglutination of washed platelets in presence of rat plasma diluted four times, 6, 12, 18 hr after DMNA administration. The results are expressed in terms of aggregation velocity (mm/min) and compared to values obtained in the control group (‘P -C 0.05)
Sham group 46.7 & 4.2 Sixth hour 47.0 i: 10.2 Twelfth hour 43.5 i: 5.7 Eighteenth hour 36.0 rt 7.0’
injury was accompanied by an inflammatory response with aggregates of leucocytes, both polymorph and mononuclear.
Ultr~~tru~t~ral studies (Fig. 7)
Six hours. There was ~saggregation of the ribo- somes associated with swelling fragmentation and vesicle formation of the rough endoplasmic reticulum (RER). Lipid droplets had accumulated in the cyto- plasm in some areas. Little glycogen was observed in cells in the vicinity of central veins. The components of the nucleolus had segregated into fibrillar condens- ation surrounding granules. Autophagic vacuoles had developed in the centrilobular hepatocytes and contained fragments of endoplasmic reticulum, mito- chondrial debris and glycogen.
Twelue hours. In central hepatocytes, RER had largely disappeared and only scattered detached ribo- somes were seen. There were more autophagic vacu- oles and accumulation of lipids particularly in the cells of the peri-portal area. RER was increased in the bihary pole of the cells. Damage to mitochondria was rare. Microsegregation of the nucleolus was still more pronounced and there were small aggregated con- densations of the fibriltar component.
Eighteen hours. These lesions were now more ad- vanced and in addition there was evidence of damage affecting the mitochondria, lysosomes and plasma membrane. Numerous extracellular coated circular vesicles (60 pm) were seen apparently arising from the plasma membrane. There were also some similar vesicles within cells. Large cytolysosomes appeared where frank necrosis had taken place and there were concentric lamellar formations corresponding to both myelin and finger print configurations, the former being closely packed, the latter more loosely ar- ranged.
Microsegregation of the nucleolus was followed by depletion of the granular component leaving a clumped reticular pattern of fibrils. Several dense plaques developed at the periphery of the nucleolus. Erythrocytes were engulfed within sinusoids and cen- tral veins were occupied by cellular debris. The endothelial lining was often absent or damaged with interruption of plasma membranes or their invagi- nation. Fibrin meshes were observed at this time within the sinusoids.
Twenty four hours. There was further development of the cellular lesions with nuclear capping, con- centric lamellar proliferation and lesions of mitochondria. Complete cell necrosis was noted. Polymorphonuclear cells infiltrated enlarged inter- cellular spaces and microthrombi obstructed the cen- tral veins. Even at this time, no platelet aggregates were detected.
DISCUSSION
The potent hepatocellular toxic power of DMNA in different species is well known, In the presence of NADPH and molecular oxygen, a microsomal en- zymatic system present mainly in liver (Pegg, 1980) leads to the formation of a powerful methylating agent that reacts with everything within range. Atten- tion has been focused upon methylation of the gua- nine of ribonucleic and desoxyribonucleic acids (Pegg, 1980) and histidine and cysteine of proteins (Hemminki and Sovokinen, 1980). Inactivation of the ribosomes is probably responsible for the failure of protein synthesis in the liver although this would not necessarily lead to the frank necrosis of liver cells which appears 12-18 hr after poisoning with a DMNA dose of 50 mg/kg (McLean et al., 1965; Magee, 1966).
Liver cell necrosis might induce impairment of coagulation and haemostasis by at least three path- ways. First, by an increase of plasmin activity due to deficient clearance by liver cells, which might result in degradation of coagulation factors (Balkuv-Ulutin, 1978). Second, by disseminated intravascular coagu- lopathy in which debris form membranes (gly- coproteins and phospholipids) circulate in the blood- stream as the tissue factor able to activate factor VII and thereby the extrinsic coagulation pathway. Third, by decreased hepatic protein synthesis of factors II, VII, IX and X exclusively produced in the liver or fibrinogen, factor V, factor VIII : C and AT III partly synthesized by the liver. The aim of the present study, therefore, is to evaluate the contribution that each of these pathways makes in the severe coagu- lation defects associated with hepatic failure induced by DMNA.
Excessive fibrinolysis in liver disease is well estab- lished in patients with cirrhosis of the liver, whereas Ratnoff (1949) found little evidence for this in pa- tients with acute liver damage. The results obtained in the present study, however, suggest that for 18 hr after damage there is no increase of circulation piasmin activity since the euglobulin lysis time is unchanged and there are no fibrin degradation prod- ucts in the blood. Plasmin activity using chromogenic substrate has not been assayed directly in this study since, unlike human, rat plasminogen cannot be activated by urokinase or streptokinase.
Disseminated intravascular coagulation has been described both in liver necrosis (Rake et al., 1970) and cirrhosis (Horder, 1969; Titgat et al., 1971). In these cases coagulation defects were reversed by heparin. In human acute hepatic failure (Rake et ul., 1970) and in rat after acute liver necrosis induced with carbon tetrachioride (Rake et al., 1973) there was evidence of intravascular coagulation. The con- cept of DIC in liver disease has not, however, re- ceived universal support. The main features of DIC are (1) a drop or even absence of circulating anti- thrombin III (2) a decrease of all coagulation factors including fibrinogen (3) the appearance of circulating fibrin degradation products at a level of more than 30pg/ml in man while fibrinolysis remains un- modified (4) a factor VIII :C/factor VIII :vW ratio greater than 1 and (5) severe thrombopaenia (less than 100 x lo9 platelets/l).
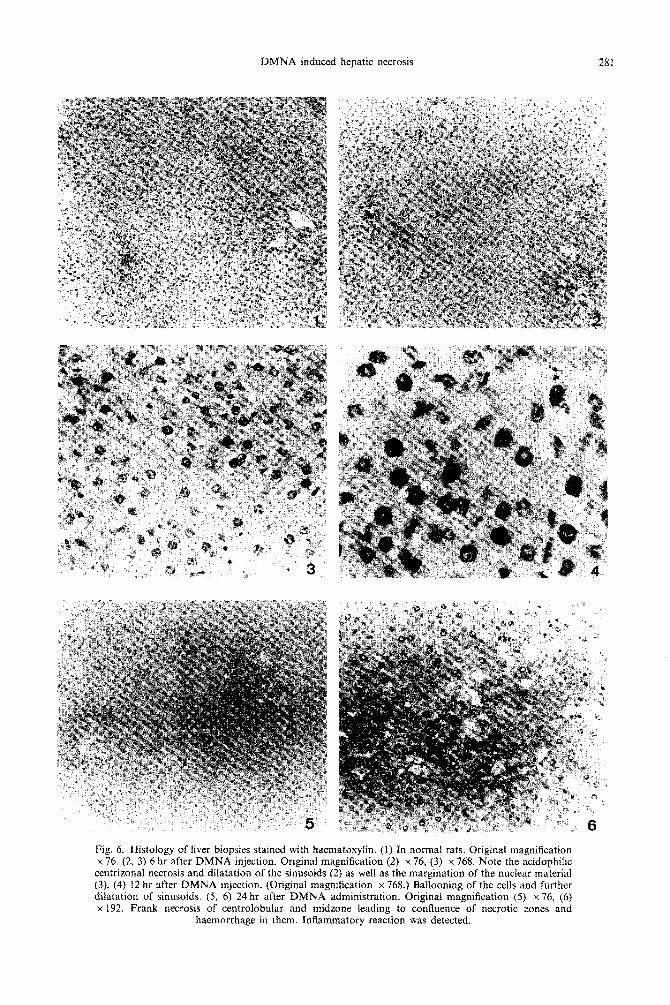
DMNA induced hepatic necrosis 281
Fig. 4. Histology of liver biopsies stained with haematoxylin. (1) In normal rats. Original magnification x 76. t2. 3) 6 hr after DMNA iniection. Original magnification t2) x 76, (31 x 768. Note the acidonhilic centrizonal necrosis and dilatation of the si&soids (2) as well ai the m&nation of the nuclear material (3). (4) 12 hr after DMNA injection. (Original magnification x 768.) Ballooning of the cells and further dilatation of sinusoids. (5, 6) 24 hr after DMNA administration. Original magnification (5) x 76. (6) x 192. Frank necrosis of centrolohular and midzone leading to confluence of necrotic zones and
haemorrhage in them. Inflammatory reaction was detected.

282 BERNAR~I CoiumLox ef al.

DMNA induced hepatic necrosis 283
All criteria of DIC were absent in the first 18 hr, but present between the 18th and 24th hr: low levels of coagulation proteins, a significant decrease in fibrinogen level, thrombocytopaenia, presence of cir- culating fibrin degradation products while fibrinolysis remains unmodified, no significant variation of factor VIII : VW level.
The histological and ultrastructural features of DIC were also present. They might be related to the early presence and further increase of membrane debris, pseudomyelinic bodies or circular vesicles detected within hepatic cells but also in surrounding sinusoids.
The third aetiopathogenic mechanism of impaired haemostasis during liver cell failure may involve decreased synthesis and/or transformation of clotting profactors.
The present study demonstrates that all vitamin-K- dependent factors, factor VIII:C and factor V had already decreased significantly in the first 12 hr of the experiment, a time at which no DIC criteria could be detected. All the vitamin-K-dependent factors (II, VII, IX and X) are synthesized and gam- macarboxylated in the rough endoplasmic reticulum of hepatocytes. Factor V is at least partly synthesized by the hepatocytes as shown by perfusion experi- ments (Workman and Lundblad, 1977) and hepa- tocellular cell culture (Wilson et al., 1984).
Despite many investigations, the site of factor VIII:C synthesis is not known. Clinical, immu- nological, transplantation and perfusion studies strongly suggest that factor VIII : C is released by the liver (Bloom, 1977; Shaw et al., 1979).
The important decrease of coagulation factors at 18 hr, i.e. factors II, V, VII, VIII:C and X, may be related to imnairment of orotein svnthesis in heoa- tocytes. Morihologic studies gave evidence of ulira- structural lesions of RER with detached ribosomes. But if decreased protein synthesis is exclusively in- volved then the rate of decrease of the different proteins should be of the same order as their biolog- ical half life. In this study using DMNA the half lives of the coagulation factors were considerably shorter than those found in man and in the control rats as shown in Table 2. This observation suggests that other mechanism may also be responsible for the rapid decline of coagulation factor levels.
The possibility that coagulation factors might be inactivated by DMNA methylation cannot be
Table 2. Comparative values of experimental rat half lives and human bioloeical half lives of coaeulations factors
Experimental calculated half Rat half
lives* lives7 Human half Factors (hr) (hr) lives
II 13.1 58 2-3 days V 2.6 IS-24 hr VII 5.4 6.0 4-6 hr VIII:C 5.2 12-18 hr IX 6.7 10.0 IS-30 hr X 3.1 11.4 48 hr
‘Prothrombin excepted, all decrease follow exponential functions between the 6th and 18th hr. Prothrombin value was calculated from a linear function.
tFrom data of Owen and Bowie (1978); values were calculated from levels obtained 8 and 24hr after i.v. injection of 0.15mg of sodium Warfarin.
excluded since DMNA is able to methylate histidine and cysteine of serum proteins (Hemminki and Sor- kinen, 1980). In this event, one might suggest the importance of histidine and cysteine on the active site (or in the ternary configuration) of the all in- vestigated coagulations factors, which seems unlikely. An alternative hypothesis is that the hepatic injury releases proteolytic enzymes into the blood; although our results demonstrate the absence of plasmin activ- ity until the 18th hr, other proteolytic activities have not been investigated at the present time. Never- theless this mechanism may explain why two high molecular weight proteins (factor V and factor VIII : C) which are particularly protease sensitive decrease so rapidly.
The rapid decrease of coagulation factors at 12 hr, i.e. factor II, factor VII, factor X, factor IX, factor V, factor VIII:C, and at the 6th hr for factor V may be related to the early histological and ultrastructural lesions of hepatocytes. Impaired protein synthesis by the hepatocytes together with possible release of hepatic proteases are likely to be early consequences of acute hepatic intoxication with DMNA. However whichever mechanism is involved, factor V is the coagulation protein to decrease significantly first following hepatocellular injury in this animal model.
REFERENCES
Alexander B., Meyer L., Kenny J., Goldstein R., Gurewich V. and Grinspoon L. (1956) Blood coagulation in preg- nancy: proconvertin and prothrombin, and the hyper- coagulable state. New Engl. J. Med. 254, 358-362.
Bachman F., Duckert F. and Keller F. (1958) The Stuart-Prower factor assay and its clinical significance. Thromb. Diath. Haem. 2, 24-38.
Balkuv-Ulutin S. (1978) Physiological response to enhanced fibrinolytic activity. In Fibrinolysis: Current Fundamental and Clinical Concepts (Edited by Gaffney P. J. and Balkuv-Ulutin S.), pp. 26-27.
Bloom A. L. (1977) Physiology of Factor VIII. In Recent Advances in Blood Coagulation (Edited by Poller L.). Churchill-Livingstone, London.
Chisholm M. (1979) Haematological disorders in liver dis- ease. In Liver and Biliary Disease (Edited by Wright R.), pp. 159-181. Saunders.
Clauss A. (1957) Gerinnungs physiologische schnell-method zur Bestimmung der Fibrinogen. Acta haemat. 17, 237-240.
Dechaud H., Belleville J., Benveniste E., Later J. and Quincv C. (1978) Determination du temos de lvse des euglobulines par une mkthode radioisotopique. Lqon Pharm. 29, 17-20.
Dodds W. (1978) Platelet functions in animals: species specificities. In Platelets: A Multidisciplinary Approach (Edited by de Gaetano G. and Garratini S.), pp. 49-50. Raven Press, New York.
Dimock I. W., Tucker J. S., Wollfil I. L. et al. (1975) Coagulation studies as a pronostic index in acute liver failure. Br. J. Haemat. 29; 385-395.
Green G., Poller L., Thomson J. M. et al. (1976) Factor VII as a marker of hepatocellular synthetic functions in liver disease. J. clin. Path. 29, 971-975.
Hardisty P. M. and McPherson J. C. (1962) A one stage factor VIII assay and its use in venous and capillary plasma. Thromb. Diath. Haem. 8, 215-229.
Hemminki K. and Sovokinen H. (1980) Alkylation of rat serum proteins by dimethylnitrosamine and acetylaminofluorene. Toxicol. Lett. 6, 433-437.

284 BERNARD C~RNILLON et al.
Horder M. H. (1969) Consumption coagulopathy in liver Ratnoff 0. D. (1949) Studies on a proteolytic enzyme cirrhosis. Thromb. Diaih. Haem. Suppl. 36, 313. in human plasma IV. The rate of lysis of plasma
McLean E., Bras G. and McLean A. E. M. (1965) Venous clots in normal and diseased individuals with particular occlusion in the liver following dimethylnitrosamine. Br. reference to hepatic disease. Bull. Johns Hopkins Hosp. 84, J. exp. Path. 46, 36. 29-42.
Magee P. N. (1966) Toxic liver necroses. Lab. Invest. 15, 111~116.
Merskey C., Lalezari P. and Johnson A. (1969) A rapid, simple, sensitive method for measuring ~b~nolytic split products in human serum. Proc. Sot. exp. Biol. Med. 281, 1023- 1026.
Roberts H. R. and Cederbaum A. I. (1972) The liver and blood coagulation physiology and pathology. Castro- enterology 63, 297-320.
RosoII’ L. and Rosoff L. (1977) Biochemical tests for hepatobihary disease. Surg. Cl&. North Am. 57, 257- 273.
@degard 0. (1975) Evaluation of an heparin cofactor method. Thromb. Res. 7, 351-360.
Owen C. A. and Bowie E. J. (1978) Rat coagulation factors V, VIII, XI and VII: vitamin-K-dependent. Huemosfasis 7, 189-201.
Saunders S. J., Seggie J. and Kirsch R. E. (I 979) Acute liver failure. In Liver and Biliuty Disease. (Edited by Wright R.), pp. 569-584. Saunders,
Owren P. (1947) The coagulation of blood. Investigation on a new# clotting tactor. Acta med. stand. 194, I.
Pegg A. E. (1980) Fo~ation and subsequent repair of alkilation lesions in tissues of rodents treated with nitro- samines. Archs Toxicol. Suppl. 3, 55-68.
Rake M. O., Flute P. T., Pannell G. and Williams R. (1970) Intravascular coagulation in acute hepatic necrosis. Lancer 1, 533-5371
Shaw E., Giddings J. C., Peake I. R. and Bloom A. L. (1979) Synthesis of procoagulant Factor VIII, Factor VIII related antigen and other coagulation factors by the isolated rat liver. Br. J. Haemat. 41, 585-596.
Titgat G. N., Cohen D. and Vestraete M. (1971) Metabo- lism of fibrinogen in cirrhosis of the liver. J. c&z. Inuesf. 50, 1690-1701:
Wilson D. B., Salem H. H., Mruk J. S., Maruyama I. and Maierus P. W. (1984) Biosvnthesis of coauulation factor V by a human hepatbcelluiar carcinoma c& line. J. c/in. Invest. 73, 654-658. Rake M. 0.. Flute P. T., Pannel G., Shilkin K. B. and
Williams R. 11973) Experimental hepatic necrosis: studies on coagulation abnormalities plasma clearance and organ distribution of 12-5-I-labelled fibrinogen. Gut l&574-580.
Workman E. F. and Lundblad R. L. (I 977) The role of liver in biosynthesis of the non vitamin-K-dependent clotting factors. Seminars Thromb. Haem. 4, 15-28.


















