
Evidence for fibromuscular pulleys of the recti extraocular muscles.
12
Evidence for Fibromuscular Pulleys of the Recti Extraocular Muscles Joseph L. Demer,*^ Joel M. Miller^ Vadims Poukens,*§ Harry V. Vinters,§ and Ben J. Glasgow* § Purpose. Magnetic resonance imaging (MRI) shows that the paths of recti extraocular muscle (EOM) bellies remain fixed in the orbit during large ocular rotations and across large surgical transpositions of their insertions. These findings imply that recti EOMs pass through pulleys coupled to the orbit and anterior to the muscle bellies, because the insertions must move with the globe. The present study was conducted to locate anatomically and to characterize histologically the pulley tissues. Methods. High-resolution MRI images were collected from volunteers, using multiple gaze directions to infer the locations of, and occasionally to visualize, recti EOM pulleys. Fresh cadaver orbits were exenterated and dissected to evaluate mechanical and structural properties of the orbital connective tissues. Lipid was cleared from whole specimens to reveal tissue relationships. Other specimens were selectively step- and serial-sectioned for histochemical and immunohistochemical staining. Results. Magnetic resonance imaging demonstrated dense connective tissue structures within posterior Tenon's fascia near the equator of the globe adjacent to the recti EOMs. Histochem- istry showed these structures to be pulleys—fibroelastic EOM sleeves consisting of dense bands of collagen and elastin, suspended from the orbit and adjacent EOM sleeves by bands of similar composition. A monoclonal antibody to human smooth muscle a-actin demonstrated substantial smooth muscle in the pulley suspensions and in posterior Tenon's fascia. Tenon's fascia itself was seen to be suspended at its periphery from the orbital walls like a drumhead. Conclusions. The human orbit contains specialized musculofibroelastic tissues in and just poste- rior to Tenon's fascia that serve as compliant pulleys and determine the pulling directions of recti EOMs. In this sense, the pulleys are the functional origins of the recti EOMs and are determinants of ocular motility. Invest Ophthalmol Vis Sci. 1995;36:1125-1136. .Before the development of computational models late binocular alignment from first principles then L of extraocular biomechanics, 1 " 5 it was acceptable to made it obvious that the paths of EOMs, gaze depen- r describe extraocular anatomy in terms of origins, in- dence of these paths, and the mechanical role of sup- i sertions, and cross-sections of recti extraocular mus- porting orbital tissues, were of critical importance in cles (EOMs) measured in cadavers. Attempts to calcu- determining EOM action. Surprisingly, nothing quan- , titative was known about these factors. For example, Pv" From the Departments of* Ophthalmology and-\ Neurology, Jules Stein Eye Institute, Computational simulations of muscle transpo ' University of California at Los Angeles; the ISmUh-Keltlewell Eye Research c •• j i .• 11 , , T^.-,»« Institute, San FrLasco, California; and the^epartment of Pathology and Surgery failed because the models assumed that EOM laboratory Medicine, University of California, J T JX Angeles. bellies COllId sideslip in the Orbit tO follow transposed k Presented in part at the. annual meeting of the Association for Research in Vision • . • 6 T> i J • I • i • • i 7 k and ophtJmoio®, Sarasota.Fiorida^ay 1994. insertions. 6 Early radiographic studies in monkeys 7 Supported by these grants from the National Eye Institute: consortium grant EY- and radiographic computed tomography in hu 08313 (IU), [MM); core grant EY-00331 to the Department olOphthalmology, , • . 8 1 t .. j ..u .. .. u iT n- r *u t University of CalifLa atUs Angeles; and core glnt EY-06883to.WA- Subjects later Suggested that the bellies of the reCtl V- Kettlewell Eye Research Institute. JID was a Research to Prevent Blindness William EOMs have paths that are Stable relative tO the t^ZTor^^S^TmXr^sedDe^,, m4; accepted despite changes in gaze. Miller used magnetic reso- December 6,1994. nance imaging (MRI) with three-dimensional image R^TriTjo^DeL, Jules stein Eye institute, wo stein plaza, uciA, reconstruction to demonstrate the extreme stability of IMS Angeles, CA 90095-7002. the recti EOM belly paths throughout the normal Investigative Ophthalmology & Visual Science, May 1995, Vol. 36, No. 6 Copyright © Association for Research in Vision and Ophthalmology 1125 Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018
Transcript of Evidence for fibromuscular pulleys of the recti extraocular muscles.

Evidence for Fibromuscular Pulleys of the RectiExtraocular Muscles
Joseph L. Demer,*^ Joel M. Miller^ Vadims Poukens,*§ Harry V. Vinters,§and Ben J. Glasgow* §
Purpose. Magnetic resonance imaging (MRI) shows that the paths of recti extraocular muscle(EOM) bellies remain fixed in the orbit during large ocular rotations and across large surgicaltranspositions of their insertions. These findings imply that recti EOMs pass through pulleyscoupled to the orbit and anterior to the muscle bellies, because the insertions must movewith the globe. The present study was conducted to locate anatomically and to characterizehistologically the pulley tissues.
Methods. High-resolution MRI images were collected from volunteers, using multiple gazedirections to infer the locations of, and occasionally to visualize, recti EOM pulleys. Freshcadaver orbits were exenterated and dissected to evaluate mechanical and structural propertiesof the orbital connective tissues. Lipid was cleared from whole specimens to reveal tissuerelationships. Other specimens were selectively step- and serial-sectioned for histochemicaland immunohistochemical staining.
Results. Magnetic resonance imaging demonstrated dense connective tissue structures withinposterior Tenon's fascia near the equator of the globe adjacent to the recti EOMs. Histochem-istry showed these structures to be pulleys—fibroelastic EOM sleeves consisting of densebands of collagen and elastin, suspended from the orbit and adjacent EOM sleeves by bands ofsimilar composition. A monoclonal antibody to human smooth muscle a-actin demonstratedsubstantial smooth muscle in the pulley suspensions and in posterior Tenon's fascia. Tenon'sfascia itself was seen to be suspended at its periphery from the orbital walls like a drumhead.
Conclusions. The human orbit contains specialized musculofibroelastic tissues in and just poste-rior to Tenon's fascia that serve as compliant pulleys and determine the pulling directionsof recti EOMs. In this sense, the pulleys are the functional origins of the recti EOMs and aredeterminants of ocular motility. Invest Ophthalmol Vis Sci. 1995;36:1125-1136.
.Before the development of computational models late binocular alignment from first principles thenL of extraocular biomechanics,1"5 it was acceptable to made it obvious that the paths of EOMs, gaze depen-r describe extraocular anatomy in terms of origins, in- dence of these paths, and the mechanical role of sup-i sertions, and cross-sections of recti extraocular mus- porting orbital tissues, were of critical importance in
cles (EOMs) measured in cadavers. Attempts to calcu- determining EOM action. Surprisingly, nothing quan-, titative was known about these factors. For example,Pv" From the Departments of* Ophthalmology and-\ Neurology, Jules Stein Eye Institute, Computational simulations of muscle transposition' University of California at Los Angeles; the ISmUh-Keltlewell Eye Research c •• j i .• 1 1 , , T^ . - , »«
Institute, San FrLasco, California; and the^epartment of Pathology and Surgery failed because the models assumed that EOMlaboratory Medicine, University of California, JTJX Angeles. bellies COllId sideslip in the Orbit tO follow transposed
k Presented in part at the. annual meeting of the Association for Research in Vision • . • 6 T> i J • I • i • • i 7k and ophtJmoio®, Sarasota.Fiorida^ay 1994. insertions.6 Early radiographic studies in monkeys7
Supported by these grants from the National Eye Institute: consortium grant EY- and radiographic computed tomography in human08313 (IU), [MM); core grant EY-00331 to the Department olOphthalmology, , • . 8 1 t .. j ..u .. ..u iT n- r *u
t University of CalifLa atUs Angeles; and core glnt EY-06883 to .WA- Subjects later Suggested that the bellies of the reCtlV- Kettlewell Eye Research Institute. JID was a Research to Prevent Blindness William EOMs have paths that are Stable relative tO the Orbit
t^ZTor^^S^TmXr^sedDe^,, m4; accepted despite changes in gaze. Miller used magnetic reso-December 6,1994. nance imaging (MRI) with three-dimensional imageR^TriTjo^DeL, Jules stein Eye institute, wo stein plaza, uciA, reconstruction to demonstrate the extreme stability ofIMS Angeles, CA 90095-7002. the recti EOM belly paths throughout the normal
Investigative Ophthalmology & Visual Science, May 1995, Vol. 36, No. 6Copyright © Association for Research in Vision and Ophthalmology 1125
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018

1126 Investigative Ophthalmology & Visual Science, May 1995, Vol. 36, No. 6
range of ocular rotations.1' Only the anterior portionsof the recti tendons move relative to the orbit, as theymust, because they are attached to the globe.
Two distinct (though not mutually exclusive)mechanisms were hypothesized to explain the ob-served path stability of recti EOMs: One involved con-nective tissue coupling of an anterior extent of eachmuscle to the globe, and the other involved each mus-cle's passage through some sort of pulley coupled tothe orbital wall.3-4910 Strong evidence for the pulleymodel was provided by MRI images demonstratingthat muscle belly paths are little affected by large surgi-cal transpositions of recti tendons.4 Because the surgi-cal dissections divided all connective tissue attach-ments of the EOM tendons to the globe to a level atleast 10 mm posterior to the insertions, it was con-cluded that EOM path stability was due to musculo-orbital coupling. At that time, however, neither rectiEOM tendons anterior to the equator, nor the putativepulleys themselves could be resolved in the early MRIimages.
Orbital connective tissues have aroused sporadicscientific interest. Since the time of Lockwood," inves-tigators have described networks of connective tissuesin the orbit and even the occurrence of smooth mus-cle cells.12"1'1 More recently, in investigations aimedat explaining the mechanical effects of orbital wallfractures,15 Koornneef14'16"19 performed systematichistologic studies of the orbital tissues, describing thestereotypic occurrence of connective tissue septaewithin the orbit. Although Koornneef recognized thatconnective tissues might have significance for normalEOM actions, his pioneering work did not have thebenefit of specific model-derived predictions and, con-sequently, it does not provide adequate basis for de-termining which, if any, connective tissue septae mightfunction as pulleys. The present study was conductedto determine if the recti EOM pulleys can be visualizedby MRI during gross dissection and microscopicallyand to determine the composition of these mechani-cally important structures.
METHODS
High-resolution MRI images were collected from vol-unteers who gave written informed consent to a proto-col conforming to the Declaration of Helsinki andapproved by the Human Subject Protection Commit-tee at the University of California, Los Angeles. Im-aging was performed using superconducting 1.5 TGeneral Electric Signa (Milwaukee, WI) or PickerVista (Cleveland, OH) scanners. Crucial aspects of thistechnique, described in detail elsewhere,20'21 involveuse of a 3-inch (General Electric) or 5-inch (Picker)surface coil over the orbit to be imaged and the main-tenance of steady ocular fixation on targets placed on
the inside of the bore of the MRI scanner. Coronal(Fig. 1A) or axial (Fig. IB) images of 2.5- to 3-mmthickness were obtained using T| weighting (repeti-tion time ~ 500 msec; echo time 15 to 20 msec), anda 10-cm square field of view with 256 X 192 or 256 X256 pixel matrices, giving a best pixel resolution of390 X 390 /xm. Digital image data were transferred toMacintosh Ilfx, Quadra 950, or PowerBook computers(Apple Computer, Cupertino, CA) for translation totagged image file format using locally developed soft-ware. Quantitative analysis was performed using thepublic domain National Institutes of Health Imageprogram (written by Wayne Rasband at the NationalInstitutes of Health and available from the Internetby anonymous ftp from zippy.nimh.nih.gov or onfloppy disk from NTIS, 5285 Port Royal Road, Spring-field, VA 22161, part number PB93-504868).
Fresh orbital specimens were obtained at autopsyfrom adult cadavers. Gross dissection was performedin situ, as well as after orbital exenteration. After enu-cleation of the globe, one specimen of posterior Ten-on's fascia was removed en bloc and fixed in 10%neutral buffered formalin while flatmounted to stiffcardboard. Another orbit was exenterated en blocand, after formalin fixation, placed in acetone for 5days to dissolve the orbital fat and disclose nonlipidtissues. Specimens of EOMs and connective tissueswere dissected selectively from other orbits. Tissue ori-entation was marked using colored dyes. One entireorbit was removed en bloc with periorbita intactthrough an intracranial approach after removal of thebrain. The orbital walls were removed with rongeursunder a dissecting microscope, and the orbit was fixedin 10% neutral buffered formalin for 4 days. Briefdecalcification in formalin-HCl solution (20 hours)was performed to soften residual bone particles andthe calcific trochlea. Processing of control specimensverified that this period of decalcification did not in-terfere with any of the staining procedures used here.
All specimens were fixed in 10% neutral bufferedformalin, dehydrated using graded solutions of alco-hol and chloroform, and embedded in paraffin. Thewhole orbital specimen was placed in a vacuum cham-ber to remove air bubbles before embedding. Stepand serial 7- to 10-//m sections were mounted on 50 X75 mm gelatin-coated glass slides. Masson's trichromestain was used to define muscle and collagen, VanGieson's stain to indicate elastin, and Alcian blue toindicate acid mucopolysaccharides consistent with car-tilage.22 Smooth muscle was immunohistochemicallyconfirmed using monoclonal mouse antibody to hu-man smooth muscle a-actin (Dako, Copenhagen,Denmark) applied at 25°C overnight at dilutions of1:50 to 1:500. This was followed by the addition ofa biotinylated anti-mouse IgG (Vector Laboratories,Burlingame, CA) that serves as a "bridge" for the
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018

Pulleys of Recti Extraocular Muscles
Superior
1127
Anterior
LacrimalGland
PulleyBands
Globe
PulleyBands
\Lateral °Ptic MedialRectus Nerve Rectus
B Posterior
A Inferior
FIGURE l. Magnetic resonance images of a representative right orbit illustrating evidence forrecti muscle pulleys. ON = optic nerve. LPS = levator palpebrae superioris. SO = superioroblique muscle. (A) (left) Coronal images at level of dashed line in B, obtained in primary,up, and down gaze. Arrowheads indicate change in position of globe-optic nerve junction.(right) Sagittally reconstructed images in 20° up gaze, primary gaze, and 20° down gaze.Note transverse bowing of anterior vertical recti muscle tendons in relaxation at location ofpulleys (arrows). (B) Axial image in primary gaze illustrating pulley tissues around medialrectus muscle.
primary antibody and the avidin-peroxidase complex(ABC Kit, Vector Laboratories). Diaminobenzidine(Sigma Chemical, St., Louis, MO) was used as thechromogen.23 Most sections contained vascularsmooth muscle and striated EOMs that provided in-trinsic positive and negative controls, respectively.
RESULTS
Magnetic Resonance Imaging
High-resolution surface coil MRI of some 70 orbitsuniformly confirmed that paths of the bellies of therecti EOMs remain stable relative to the orbit duringlarge (~20°) ocular rotations from a primary position(Fig. 1). Coronal images in Figure 1A were obtainedin primary gaze, as well as in 20° of elevation and 20°depression, at the level indicated by the dotted linein Figure IB. Stability of the horizontal recti duringlarge changes in vertical gaze was a consistent findingin all subjects. At the level in the orbit illustrated inFigure 1, the most obvious indication of gaze directionis the change in position of the globe-optic nerve
junction, indicated by arrowheads in Figure 1A (left).In addition to the indirect evidence for musculo-or-bital pulleys provided by muscle belly stability, carefulinspection of the highest quality images providedsome direct evidence. A full set of 12 continuous, 3-mm thick coronal images spanning the same orbit wasused to reconstruct a sagittal image set in a planecentered along the long axes of the vertical recti mus-cles (Fig. 1A, right). Note that the paths of the relaxingsuperior rectus muscle in down gaze, as well as therelaxing inferior rectus muscle in up gaze, are nottangent to the globe. Instead, the vertical recti musclesare bowed away from the globe at and just posteriorto the equator. Posterior to this point, they changedirection to run straight toward the orbital apex. Thisanterior departure from a straight-line path suggeststhe presence of some structures, demonstrated histo-logically to be the pulleys, pulling muscle paths towardthe orbital wall. Figure IB is a 2.5-mm thick axial im-age providing the best available MRI demonstrationof bands of connective tissue suspending the medialrectus pulley to the orbital wall. Similar suspensory
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018
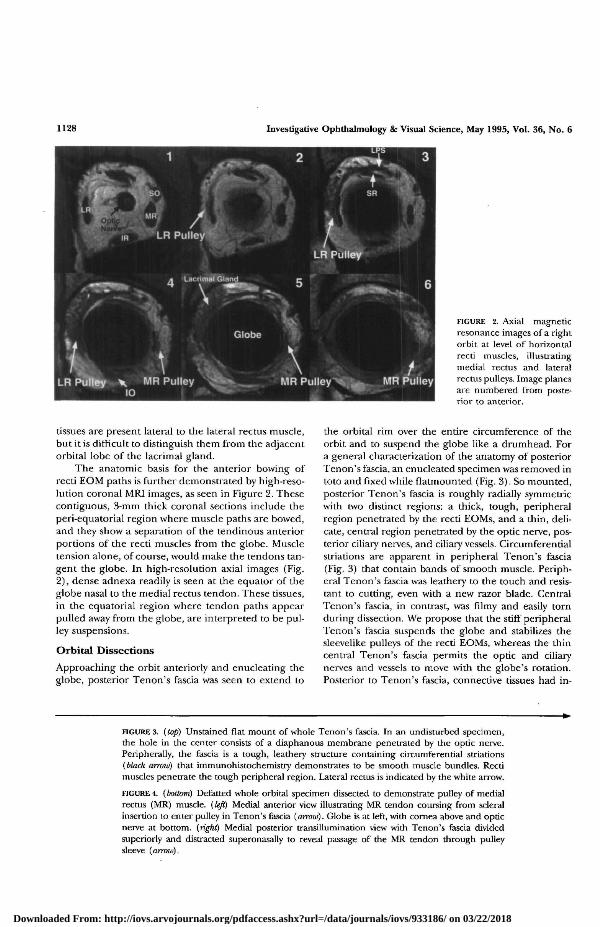
Investigative Ophthalmology & Visual Science, May 1995, Vol. 36, No. 6
FIGURE 2. Axial magneticresonance images of a rightorbit at level of horizontalreed muscles, illustratingmedial rectus and lateralrectus pulleys. Image planesare numbered from poste-rior to anterior.
tissues are present lateral to the lateral rectus muscle,but it is difficult to distinguish them from the adjacentorbital lobe of the lacrimal gland.
The anatomic basis for the anterior bowing ofrecti EOM paths is further demonstrated by high-reso-lution coronal MRI images, as seen in Figure 2. Thesecontiguous, 3-mm thick coronal sections include theperi-equatorial region where muscle paths are bowed,and they show a separation of the tendinous anteriorportions of the recti muscles from the globe. Muscletension alone, of course, would make the tendons tan-gent the globe. In high-resolution axial images (Fig.2), dense adnexa readily is seen at the equator of theglobe nasal to the medial rectus tendon. These tissues,in the equatorial region where tendon paths appearpulled away from the globe, are interpreted to be pul-ley suspensions.
Orbital Dissections
Approaching the orbit anteriorly and enucleating theglobe, posterior Tenon's fascia was seen to extend to
the orbital rim over die entire circumference of theorbit and to suspend the globe like a drumhead. Fora general characterization of the anatomy of posteriorTenon's fascia, an enucleated specimen was removed intoto and fixed while flatmounted (Fig. 3). So mounted,posterior Tenon's fascia is roughly radially symmetricwith two distinct regions: a thick, tough, peripheralregion penetrated by the recti EOMs, and a thin, deli-cate, central region penetrated by the optic nerve, pos-terior ciliary nerves, and ciliary vessels. Circumferentialstriations are apparent in peripheral Tenon's fascia(Fig. 3) that contain bands of smooth muscle. Periph-eral Tenon's fascia was leathery to the touch and resis-tant to cutting, even with a new razor blade. CentralTenon's fascia, in contrast, was filmy and easily tornduring dissection. We propose that the stiff peripheralTenon's fascia suspends the globe and stabilizes thesleevelike pulleys of the recti EOMs, whereas the thincentral Tenon's fascia permits the optic and ciliarynerves and vessels to move with the globe's rotation.Posterior to Tenon's fascia, connective tissues had in-
FIGURE 3. {top) Unstained flat mount of whole Tenon's fascia. In an undisturbed specimen,the hole in the center consists of a diaphanous membrane penetrated by the optic nerve.Peripherally, the fascia is a tough, leadiery structure containing circumferential striations{black arrow) that immunohistochemistry demonstrates to be smooth muscle bundles. Rectimuscles penetrate the tough peripheral region. Lateral rectus is indicated by the white arrow.
FIGURE 4. {bottom) Defatted whole orbital specimen dissected to demonstrate pulley of medialrectus (MR) muscle, {left) Medial anterior view illustrating MR tendon coursing from scleralinsertion to enter pulley in Tenon's fascia {arrow). Globe is at left, with cornea above and opticnerve at bottom, {right) Medial posterior transillumination view with Tenon's fascia dividedsuperiorly and distracted superonasally to reveal passage of the MR tendon through pulleysleeve {arrow).Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018

Pulleys of Recti Extraocular Muscles
sufficient stiffness to maintain spatial relationshipsamong orbital structures after subtotal exenteration.
Another orbital exenteration specimen wastreated with acetone to dissolve the orbital fat to revealmuscles and connective tissues better. The defattedspecimen was dissected (Fig. 4). Figure 4A depicts theanteromedial view seen by a strabismus surgeon, and
it illustrates the medial rectus tendon entering thepulley in Tenon's fascia. The complete course of themedial rectus tendon through the pulley sleeve inTenon's fascia is demonstrated using partial transillu-mination in Figure 4B. Note that the medial rectus istendinous within and anterior to the pulley sleeve,becoming muscular only posterior to Tenon's fascia.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018
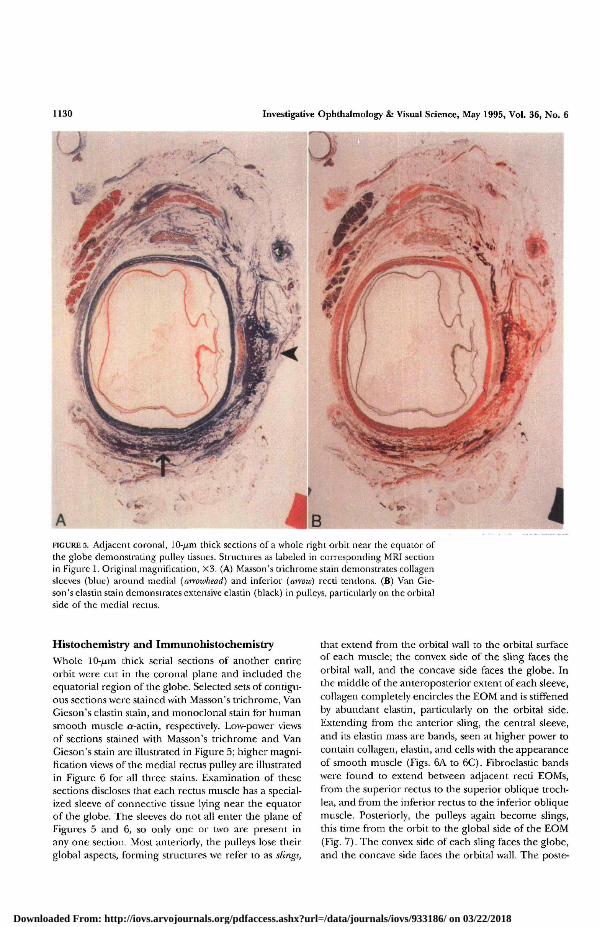
1130 Investigative Ophthalmology & Visual Science, May 1995, Vol. 36, No. 6
FIGURE 5. Adjacent coronal, 10-/̂ m thick sections of a whole right orbit near the equator ofthe globe demonstrating pulley tissues. Structures as labeled in corresponding MRI sectionin Figure 1. Original magnification, X3. (A) Masson's trichrome stain demonstrates collagensleeves (blue) around medial (arrowhead) and inferior (atroio) recti tendons. (B) Van Gie-son's elastin stain demonstrates extensive elastin (black) in pulleys, particularly on the orbitalside of the medial rectus.
Histochemistry and ImmunohistochemistryWhole 10-//m thick serial sections of another entireorbit were cut in the coronal plane and included theequatorial region of the globe. Selected sets of contigu-ous sections were stained with Masson's trichrome, VanGieson's elastin stain, and monoclonal stain for humansmooth muscle cr-actin, respectively. Low-power viewsof sections stained with Masson's trichrome and VanGieson's stain are illustrated in Figure 5; higher magni-fication views of the medial rectus pulley are illustratedin Figure 6 for all three stains. Examination of thesesections discloses that each rectus muscle has a special-ized sleeve of connective tissue lying near the equatorof the globe. The sleeves do not all enter the plane ofFigures 5 and 6, so only one or two are present inany one section. Most anteriorly, the pulleys lose theirglobal aspects, forming structures we refer to as slings,
that extend from the orbital wall to the orbital surfaceof each muscle; the convex side of the sling faces theorbital wall, and the concave side faces the globe. Inthe middle of the anteroposterior extent of each sleeve,collagen completely encircles the EOM and is stiffenedby abundant elastin, particularly on the orbital side.Extending from the anterior sling, the central sleeve,and its elastin mass are bands, seen at higher power tocontain collagen, elastin, and cells with the appearanceof smooth muscle (Figs. 6A to 6C). Fibroelastic bandswere found to extend between adjacent recti EOMs,from the superior rectus to the superior oblique troch-lea, and from the inferior rectus to the inferior obliquemuscle. Posteriorly, the pulleys again become slings,this time from the orbit to the global side of the EOM(Fig. 7). The convex side of each sling faces the globe,and the concave side faces the orbital wall. The poste-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018

Pulleys of Recti Extraocular Muscles 1131
1
TABLE i. Sagittal Distance From CornealSurface of Recti ExtraocularMuscle Pulleys*
MuscleAnterior PulleyExtent (mm)
Posterior PulleyExtent (mm)
Medial rectusInferior rectusLateral rectusSuperior rectus
6.014.116.6
5.8
25.627.031.319.9
* From serial 10-//m coronal sections starring at the anteriorcornea] surface.
FIGURE6. (left column) Adjacent coronal, 10-)um thick orbitalsections taken near the equator at medial rectus (MR) pulley(original magnification, Xl5). (A) Masson's trichrome staindemonstrates collagen sleeves (blue) around MR tendon. S= sclera;T = musculotendonous junction of MR; E = elastinmass on orbital side of pulley; M = smooth muscle bands.(B) Van Gieson's elastin stain demonstrates extensive elastin(black) in MR pulley and adjacent connective tissue bands.(C) lmmunohistochemical stain for human smooth musclea-actin demonstrates smooth muscle bands (brown reactionproduct) medial to MR pulley.
FIGURE 7. (right column) Coronal, 10-/xm thick section ofwhole right orbit near globe-optic nerve junction stainedwith Van Gieson's elastin stain. Posterior sling of lateralrectus extends From periorbita around global surface of lat-eral rectus (arroxo). Structures as labeled in correspondingmagnetic resonance imaging section in Figure 1. Originalmagnification, X3.
rior pulley slings were not found to contain smoothmuscle. The anteroposterior extent, including slings, ofeach of the pulleys is given in Table 1. Anteroposteriorlength ranged from 13 mm for inferior rectus to 19mm for medial rectus. The medial rectus pulley con-tained the greatest amount of fibroelastic tisstie, andthe superior rectus pulley contained the least.
The presence of smooth muscle was confirmedDownloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018

1132 Investigative Ophthalmology 8c Visual Science, May 1995, Vol. 36, No. 6
!B - "
FIGURE 8. (top) High-power view of 7-fim thick section showing smooth and striated musclein the region of the medial rectus (MR) pulley. Original magnification, X64. (A) Masson'strichrome stains striated muscle bright red and smooth muscle purple. (B) Immunohisto-chemical stain for human smooth muscle a-actin demonstrates brown reaction product inpulley smooth muscle and in intrinsic blood vessel walls (arrow) but not striated fibers ofmedial rectus muscle.
FIGURE 9. (bottovi) Low-power micrographs of coronal sections of mid-orbital region, poste-rior to pulleys. Structures as labeled in corresponding magnetic resonance image in Figure1. Original magnification, X3. (A) Masson's trichrome. Note that recti muscles (red) arelarge, essentially devoid of collagen sheaths (blue). Note also the fine collagenous septaeseparating orbital fat lobules. This area lacks smooth muscle. (B) Van Gieson's elastin stain.Note the scant elastin staining (black) within recti and superior oblique muscles themselvesand the absence of elastin in muscle sheaths and septae.
-Jf
using a specific immunohistochemical stain for hu-man smooth muscle a-actin (Figs. 6C, 8B). This anti-body-based stain does not bind to striated muscle fi-bers, although it does bind to the smooth muscle in-trinsic to blood vessel walls within the striated muscles
and elsewhere within the orbit (Fig. 8). Abundantbands of smooth muscle also were confirmed bysmoodi muscle a-actin immunohistochemistry in theperipheral region of Tenon's fascia near the equatorof the globe. The presence of smooth muscle in the
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018

Pulleys of Recti Extraocular Muscles 1133
pulleys and in Tenon's fascia was confirmed withsmaller specimens from other orbits. Smooth musclewas most abundant in the medial rectus pulley, lessabundant in the inferior rectus and lateral rectus pul-leys, and almost absent in the superior rectus pulley.Throughout the periorbita, occasional cells stainedpositive for smooth muscle a-actin. Alcian blue did notstain any rectus pulley tissue, indicating the absenceof acid mucopolysaccharides typical of cartilage. Themedial aspect of the trochlea was the only orbital struc-ture to stain positive with Alcian blue.
Whole coronal orbital sections posterior to Ten-on's fascia were examined at intervals approachingthe annulus of Zinn. Representative contiguous, 10-lim thick sections from approximately 6 mm posteriorto the globe-optic nerve junction are illustrated inFigure 9, stained with Masson's trichrome (Fig. 9A)and Van Gieson's elastin stain (Fig. 9B). The connec-tive tissues were similar throughout the orbit posteriorto the globe: Striated EOMs were surrounded by thin,or no, capsules containing little, if any, collagen andno elastin or smooth muscle. The insubstantial struc-ture of the posterior orbital connective tissues is con-sistent with palpation at dissection, which encoun-tered essentially no mechanical resistance from thesestructures. Fat lobules in the posterior orbit were sur-rounded by thin septae composed of collagen, but noelastin. Except for the media of blood vessels, the onlysmooth muscle in the posterior orbit was found inthe orbital muscle of Muller, which spans the inferiororbital fissure, and as scattered cells within the perior-bita itself. The orbital muscle was found to be com-posed of a mixture of collagen and smooth muscleand reached a surprisingly great maximum thicknessof more than 6 mm at a point approximately 36 mmposterior to the corneal surface.
Structure of Orbital Connective Tissues ^
From these data, we infer the architecture of the or-bital connective tissues schematized in Figure 10. Theglobe itself is supported by posterior Tenon's fasciain a drumhead arrangement supported throughoutits periphery by sturdy attachments to the periorbita,particularly at the orbital rim. The peripheral portionof posterior Tenon's fascia is a tough, fibromuscularstructure composed of dense collagen, elastin, circum-ferentially arranged smooth muscle bundles and someradially arranged smooth muscle bundles. Most of thesmooth muscle in Tenon's fascia is at and anterior tothe equator of the globe. The central, most posteriorportion of Tenon's fascia is very thin and flexible,permitting free motion of the optic nerve, ciliary ves-sels, and nerves during globe rotation.
The recti muscle pulley sleeves are continuouswith the tough peripheral portion of posterior Ten-on's fascia and become incomplete sleeves, or slings,
anterior and posterior to the sleeves. The sleeves arestabilized by fibromuscular septae extending from thepulleys to the orbital walls and to adjacent pulleys byposterior Tenon's fascia itself, which is fibromuscularas well. The pulleys are centered at and just posteriorto the equator of the globe, with effective anteropost-erior extents of approximately 13 to 19 mm. The pul-leys are composed of dense collagen, elastin, andsmooth muscle, but they do not contain acid muco-polysaccharides typical of cartilage. The recti tendonstravel within the pulley sleeves by the sliding or tele-scoping of membranous sheaths composed primarilyof collagen.
DISCUSSION
Structure of the Pulleys
Previously, the existence of pulleys constraining thepaths of the recti EOMs could only be inferred indi-rectly from magnetic resonance images of musclepaths themselves. With refined MRI techniques, wevisualized pulleys in vivo (Figs. 1, 2). Gross dissec-tions and histologic studies showed fibroelastic (col-lagen and elastin) sleeves in posterior Tenon's fas-cia at approximately the equator of the globe, ap-proximately 10 mm posterior to the EOMinsertions. The sleeves were coupled to the orbitsby musculofibroelastic (smooth muscle, collagen,and elastin) septae, extending to the periorbita andto adjacent EOM sleeves, and by the periphery ofTenon's fascia. Tenon's fascia suspended the globeand became thin and pliable posteriorly, permittingfree rotation of structures attached to the posteriorpole of the globe. Posterior to the pulleys, no struc-tures were found that would stabilize recti musclebellies, which there appear to follow straight-linepaths to the annulus of Zinn.
Smooth muscle cells near the pulleys have beendescribed, though not immunohistochemically con-firmed, by other workers.121*'24'25 The smooth musclea-actin immunohistochemical stain we used is specificand sensitive, revealing more smooth muscle than pre-viously seen with histochemical stains, includingsmooth muscle in pulley suspensions, in the periph-eral region of posterior Tenon's fascia, and evensparsely within the dense collagen of the periorbita.These muscle cells are distinct from the more anteriorsmooth muscles of the lids and the posterior orbitalmuscle of Muller. The dense smooth muscle bundlesassociated with the medial rectus muscle can be dis-cerned by MRI.
Function of the Pulleys
A notable aspect of this study is that it was motivated bywork on a computational biomechanical model,15'26
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018

1.134 Investigative Ophthalmology & Visual Science, May 1995, Vol. 36, No. 6
Horizontal Section Anterior Slings
SRtndn
Posterior SlingsSleeves tndn
Tendon Striated Smooth Elastin Collagen CartilageMuscle Muscle
FIGURE 10. Diagrammatic representation of strucLure of orbital connective tissues. IR = infe-rior rectus; LPS = levaior palpebrae superioris; LR = lateral rectus; MR = medial rectus;SO = superior oblique; SR. = superior rectus. The three coronal views are represented atthe levels indicated bv arrows in horizontal section.
which showed that known extraoctilar structures couldnot account for the stability of muscle paths observedin magnetic resonance images.IA(> This suggested thatthere were unidentified, functionally important me-chanical structures where one might reasonably haveexpected anatomic study to yield nothing new. Themusculo-fibroelastic structures here described are lo-cated just where theoretical considerations requiredthem to be to stabilize recti muscle paths relative tothe orbit, preventing them from slipping sideways(that is, perpendicular to their long axes) over theglobe. Most effective stabilization would occur in thepulley sleeves, but the adjacent slings would be ex-pected to influence EOM paths. Thus, the muscles aredeflected from the shortest "great-circle" paths they
would assume if acted on only by their intrinsic ten-sions. Pulleys also explain why magnetic resonanceimages show that conventional strabismus surgicaltransposition of recti muscle insertions leave the pathsof the EOM bellies essentially unchanged.'
Recti tendons travel through their thick pulleys bysliding inside thin collagenous sheaths that probablytelescope within the pulley sleeves to permit passageof the entire tendon. This mechanism of passage isdistinct from that reported for the superior obliquetendon in the trochlea, where fibrous layers of thetendon itself reportedly telescope relative to one an-other so that full longitudinal sliding of the tendon ispresent only in the central core.'7
The present findings conflict with several conven-
x
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018

Pulleys of Recti Extraocular Muscles 1135
tional notions of EOM function. Whereas the direc-tion of pull (or axis of rotation) of a rectus muscle isusually thought to be determined by its functionalinsertion (or point of tangency with the globe) andits anatomic origin in the annulus of Zinn, the presentfindings imply a functional origin at the muscle's pulley.The length of a muscle's path, which determines itsdegree of stretch and affects its tension, is increasedby deflection through the pulley. Although stiffenedby elastin and in the awake state by smooth muscle,we expect that the pulleys are somewhat compliant.Thus, we would expect functional origins to vary some-what as a function of gaze. Finally, we might need toconsider disorders of the pulleys. For example, underthe chronically high muscle forces of co-contractivesyndromes, pulley tissues might become distendedand weakened.
Regardless of how a gaze position is actuallyreached, the ocular torsion normally associated withit is always the same (Donder's law), and it is as thoughthe eye rotated from a primary position around a sin-gle axis lying in the equatorial plane (Listing's law).We propose that the roughly symmetrical arrange-ment of the four recti pulleys, and the drumhead-likesuspension of the globe provided by Tenon's fascia,provide a "passive" mechanism for Listing's law. Ithad been supposed that there were no orbital struc-tures that could passively determine torsion accordingto Listing's law, which was assumed to have an activeneural basis. It was taken as evidence for this view thatListing's law is violated in sleep28 by convergence2930
and by the vestibulo-ocular reflex.Sl However, theseviolations are small and could be produced by neuralnoise in sleep, in vergence, and by vestibulo-ocularcommands stretching connective tissues. That is, wepropose that Listing's law is "overdetermined" firstby orbital connective tissue and smooth muscle andsecond by EOM coordination that tends to be consis-tent with passive constraints.
The complete role of smooth muscle in the pul-leys and in Tenon's fascia is not known. Possible func-tions include maintenance of appropriate stiffness ofposterior Tenon's fascia and pulleys during aging and,perhaps, as suggested by Bergen,12 a dynamic role infine tuning the position of the pulleys. The smoothmuscle in pulley suspensions is well located to modifymuscle-pulling directions. Dynamic contraction ofsmooth muscle around the pulley sleeves could playa role in vergence eye movements. Such a role inhorizontal vergence is suggested by the particularlywell-developed smooth muscle struts extending medi-ally from the medial rectus pulley in an arrangementthat could displace the pulley to stretch the medialrectus and augment convergence. An analogous func-tion is served by Muller's smooth muscle, which helpsthe striated levator palpebrae superioris elevate the
upper eyelid. Neural control of smooth muscle in thepulleys and Tenon's fascia is poorly understood. How-ever, unmyelinated nerves and motor end plates seenwithin the smooth muscle have histochemical and im-munohistochemical characteristics suggesting thatthese nerves are nitroxidergic and catecholaminer-
Surgical Implications
Strabismus surgeons normally do not encounter therecti pulleys. The usual technique of engaging EOMtendons on blunt hooks probably pulls the tendonsanteriorly through the sleeves, leaving the pulleys be-hind. The anterior sling portion of each pulley is onthe orbital side of the muscle and is simply avoidedor retracted out of the field during surgery. Surgeonsare taught not to violate posterior Tenon's capsuleto avoid releasing orbital fat and producing the fatadherence syndrome.33 Thus, the pulleys tend to re-main outside the surgical field, hindering their recog-nition. The posterior pulley slings are too far back inthe orbit to be encountered at all during conventionalstrabismus surgery.
As we have argued, conventional rectus muscletransposition surgery leaves the pulleys intact and, so,causes little or no change in recti muscle belly paths.'1
In fact, the only situation in which these paths areknown to change is as a function of gaze in somepatients with Duane's syndrome exhibiting up ordown shoots.10 Here, perhaps, pulleys are weakenedby unusually high muscle forces produced by chronicco-contraction of misinnervated horizontal recti. How-ever, all available post-transposition MRI data werecollected within days of surgery; therefore, we cannoteliminate the possibility that, given sufficient time, thepulley of a surgically transposed muscle would notweaken or displace. Perhaps supplementing inser-tional transposition with transposition of recti pulleyswould enhance the effects of surgery for EOM paraly-ses. Markedly exaggerated effects have been observedin patients with recti muscle transposition in whomTenon's fascia has been divided (Rosenbaum AL, un-published data, 1983). We expect that knowledge—particularly as embodied in quantitative models—oforbital connective tissue anatomy will lead to rationaldesign of novel surgical approaches to the pulleys.
Key Words
connective tissue, extraocular muscle, magnetic resonanceimaging, orbit, smooth muscle
Acknowledgments
The authors thank Galit Katsir for technical assistance withtissue processing, William O'Day For assistance with photo-micrography, and Igor Legkholts and Karl Beykirch for com-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018

1136 Investigative Ophthalmology & Visual Science, May 1995, Vol. 36, No. 6
puter programming for the magnetic resonance image anal-ysis.
References
1. Miller JM, Robinson DA. A model of the mechanicsof binocular alignment. Comput Biomed Res.1984; 17:436-70.
2. Miller JM. Applications of the SQUINT computer stra-bismus model. ARVO Abstracts. Invest Ophthalmol VisSd. 1985;26:253.
3. Miller JM, DemerJL. Biomechanical analysis of strabis-mus. Binocular Vision Eye Muscle Surg Q. 1992; 7:233-248.
4. Miller JM, DemerJL, Rosenbaum AL. Effect of trans-position surgery on rectus muscle paths by magneticresonance imaging. Ophthalmology. 1993; 100:475-487.
5. Robinson DA. A quantitative analysis of extraocularmuscle cooperation and squint. Invest Ophthalmol.1975; 14:801-25.
6. Miller JM. Functional anatomy of normal human rec-tus muscles. Vision Res. 1989;29:223-240.
7. Miller JM, Robins D. Extraocular muscle sideslip andorbital geometry in monkeys. Vision Res. 1987; 27:381-392.
8. Simonsz HJ, Harting F, de Waal BJ, Verbeeten BWJM.Sideways displacement and curved path of recti eyemuscles. Arch Ophthalmol. 1985; 103:124-128.
9. Miller JM, Robinson DA, Scott AB, Robins D. Side-slipand the action of extraocular muscles. ARVO Ab-
i stracts. Invest Ophthalmol Vis Sd. 1984;25:182.10. Miller JM, Demer JL, Rosenbaum AL. Two mecha-
nisms of up-shoots and down-shoots in Duane's syn-drome revealed by a new magnetic resonance imaging(MRI) technique. In: Campos EC, eds. Strabismus andOcular Motility Disorders. London: Macmillan; 1990:229-234.
11. Lockwood CB. The anatomy of the muscles, liga-ments, and fasciae, etc. JAnat Physiol. 1886;20:l-25.
12. Bergen MP. Some histological aspects of the structureof the connective tissue system and its relationshipswith the blood vessels in the human orbit. Ada MorpholNeerlScand. 1982; 20:293-308.
13. Jones LT. A new concept of the orbital fascia andrectus muscle sheaths and its surgical implications.Trans Am Acad Ophthalmol Otolaryngol. 1968; 72:755-64.
14. Koornneef L. Spatial Aspects of Orbital Musculo-fibrousTissue in Man. Amsterdam: Swets & Zeitlinger; 1977.
15. Koornneef L. Current concepts on the managementof orbital blow-out fractures. Ann Plast Surg.1982;9:185-200.
16. Koornneef L. Orbital septa: Anatomy and function.Ophthalmology. 1979;86:876-80.
17. Koornneef L. New insights in the human orbital con-nective tissue: Result of a new anatomical approach.Arch Ophthalmol. 1977;95:1269-73.
18. Koornneef L. Details of the orbital connective tissuesystem in the adult. Acta Morphol Neerl Scand.1977; 15:1-34.
19. Koornneef L. The architecture of the musculo-fibrousapparatus in the human orbit. Acta Morphol NeerlScand. 1977; 15:35-64.
20. DemerJL, Miller JM, Rosenbaum AL. Clinical correla-tions of models of orbital statics. In: Scott AB, eds.Mechanics of Strabismus Symposium. San Francisco:Smith-Kettlewell Eye Research Institute; 1992:141-161.
21. DemerJL, Miller JM. Magnetic resonance imaging ofthe functional anatomy of the superior oblique mus-cle. Invest Ophthalmol Vis Sd. 1995; 36:906-913.
22. Sheehan DC and Hrapchak BB. Theory and Practice ofHistotechnology. St. Louis: CV Mosby; 1973:95-116.
23. Sternberger LA. Immunocytochemistry. New York: Wiley;1979.
24. Koornneef L. The first results of a new anatomicalmethod of approach to the human orbit following aclinical enquiry. Acta Morphol Neerl Scand. 1974;12:259-82.
25. Koornneef L. Sectional Anatomy of the Orbit. Amster-dam: Aolus Press; 1981:1-33.
26. Miller JM, Shamaeva I. Orbif® 1.0 Gaze Mechanics Simu-lation User's Manual. San Francisco: Eidactics; 1993.
27. Helveston EM, Merriam WW, Ellis FD, Shellhamer RHand Gosling CG. The trochlea: A study of the anatomyand physiology. Ophthalmology. 1992;80:124-133.
28. Nakayama K, Balliet R. Listing's law, eye positionsense, and perception of the vertical. Vision Res.1977; 17:453-457.
29. Allen MJ. The dependence of cydophoria on conver-gence elevation and the system of axes. Am J Optom.1954;31:297-307.
30. Hering E. Der Raumsinn und die Bewegungen desAuges. In: Hermann L, eds. Handbuch der Physiologie.Leipzig: FCW Vogel; 1879:343-601. In German.
31. Crawford JD, Vilis T. Axes of eye rotation and Listing'slaw during rotations of the head. J NeurophysioL1991;65:407-423.
32. Demer JL, Poukens V, Micevych PE. Nitroxideregicand catecholaminergic innervation of the smoothmuscle of the medial rectus pulley in humans. ARVOAbstracts. Invest Ophthalmol Vis Sd. 1995; 36: in press.
33. Parks MM. Ocular Motility and Strabismus. Hagerstown,PA: Harper & Row; 1975.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933186/ on 03/22/2018


















