
EVIDENCE FOR DISTINCT MECHANISMS OF … FOR DISTINCT MECHANISMS OF STARCH GRANULE BREAKDOWN IN...
24
1 EVIDENCE FOR DISTINCT MECHANISMS OF STARCH GRANULE BREAKDOWN IN PLANTS. Thierry Delatte, Martin Umhang, Martine Trevisan 1 , Simona Eicke, David Thorneycroft 2 , Steven M. Smith 3 and Samuel C. Zeeman * From the Institute of Plant Sciences, ETH Zurich, CH-8092 Zurich , Switzerland. Running Title : Mechanisms of starch degradation. 1 Institute of Plant Sciences, University of Bern, CH-3013 Bern, Switzerland. 2 Institute of Cell and Molecular Biology, University of Edinburgh, Edinburgh EH9 3JH, UK. 3 ARC Centre of Excellence in Plant Energy Biology, University of Western Australia, Crawley WA 6009, Australia. Address correspondence to: Samuel C. Zeeman, Institute of Plant Sciences, ETH Zurich, CH-8092 Zurich , Switzerland; Tel: +41 (44) 632 8275; Fax: +41 (44) 632 1044; E-mail [email protected] . The aim of this work was to understand the initial steps of starch breakdown inside chloroplasts. In the non-living endosperm of germinating cereal grains, starch breakdown is initiated by α- amylase secreted from surrounding cells. However, loss of α-amylase from Arabidopsis does not prevent chloroplastic starch breakdown [Yu et al, 2005, J. Biol. Chem. 250, 9773], implying that other enzymes must attack the starch granule. Here, we present evidence that the debranching enzyme isoamylase3 (ISA3) acts at the surface of the starch granule. Atisa3 mutants have more leaf starch and a slower rate of starch breakdown than wild-type plants. The amylopectin of Atisa3 contains many very short branches and ISA3-GFP localizes to granule-like structures inside chloroplasts. We suggest that ISA3 removes short branches from the granule surface. To understand how some starch is still degraded in Atisa3 mutants we eliminated a second debranching enzyme, limit dextrinase (LDA; pullulanase-type). Atlda mutants are indistinguishable from the wild type. However, the Atisa3/Atlda double mutant has a more severe starch- excess phenotype and a slower rate of starch breakdown than Atisa3 single mutants. The double mutant accumulates soluble branched oligosaccharides (limit dextrins) which are undetectable in the wild type and the single mutants. Together these results suggest that glucan debranching occurs primarily at the granule surface via ISA3, but in its absence soluble branched glucans are debranched in the stroma via LDA. Consistent with this model, chloroplastic α-amylase AtAMY3, which could release soluble branched glucans, is induced in Atisa3 and in the Atisa3/Atlda double mutant. Starch is the most abundant storage carbohydrate in plants. It is composed primarily of amylopectin, a branched polymer of glucose in which α-1,4-linked glucan chains are connected by α-1,6-bonds (branch points). In this way, amylopectin resembles glycogen, the soluble storage carbohydrate synthesized in prokaryotes, fungi and animals. However, unlike glycogen, amylopectin molecules can form a semi-crystalline granule. This capacity stems from the branching pattern of amylopectin, which produces clusters of linear chains that pack together in regular http://www.jbc.org/cgi/doi/10.1074/jbc.M513661200 The latest version is at JBC Papers in Press. Published on February 22, 2006 as Manuscript M513661200 Copyright 2006 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on May 22, 2018 http://www.jbc.org/ Downloaded from
Transcript of EVIDENCE FOR DISTINCT MECHANISMS OF … FOR DISTINCT MECHANISMS OF STARCH GRANULE BREAKDOWN IN...

1
EVIDENCE FOR DISTINCT MECHANISMS OF STARCH GRANULE BREAKDOWN IN PLANTS.
Thierry Delatte, Martin Umhang, Martine Trevisan1, Simona Eicke, David
Thorneycroft2, Steven M. Smith3 and Samuel C. Zeeman* From the Institute of Plant Sciences, ETH Zurich, CH-8092 Zurich , Switzerland.
Running Title : Mechanisms of starch degradation. 1Institute of Plant Sciences, University of Bern, CH-3013 Bern, Switzerland. 2Institute of Cell and Molecular Biology, University of Edinburgh, Edinburgh EH9 3JH, UK. 3ARC Centre of Excellence in Plant Energy Biology, University of Western Australia, Crawley WA 6009, Australia. Address correspondence to: Samuel C. Zeeman, Institute of Plant Sciences, ETH Zurich, CH-8092 Zurich , Switzerland; Tel: +41 (44) 632 8275; Fax: +41 (44) 632 1044; E-mail [email protected]. The aim of this work was to understand the initial steps of starch breakdown inside chloroplasts. In the non-living endosperm of germinating cereal grains, starch breakdown is initiated by α-amylase secreted from surrounding cells. However, loss of α-amylase from Arabidopsis does not prevent chloroplastic starch breakdown [Yu et al, 2005, J. Biol. Chem. 250, 9773], implying that other enzymes must attack the starch granule. Here, we present evidence that the debranching enzyme isoamylase3 (ISA3) acts at the surface of the starch granule. Atisa3 mutants have more leaf starch and a slower rate of starch breakdown than wild-type plants. The amylopectin of Atisa3 contains many very short branches and ISA3-GFP localizes to granule-like structures inside chloroplasts. We suggest that ISA3 removes short branches from the granule surface. To understand how some starch is still degraded in Atisa3 mutants we eliminated a second debranching enzyme, limit dextrinase (LDA; pullulanase-type). Atlda mutants are indistinguishable from the wild type. However, the Atisa3/Atlda double mutant has a more severe starch-excess phenotype and a slower rate of
starch breakdown than Atisa3 single mutants. The double mutant accumulates soluble branched oligosaccharides (limit dextrins) which are undetectable in the wild type and the single mutants. Together these results suggest that glucan debranching occurs primarily at the granule surface via ISA3, but in its absence soluble branched glucans are debranched in the stroma via LDA. Consistent with this model, chloroplastic α-amylase AtAMY3, which could release soluble branched glucans, is induced in Atisa3 and in the Atisa3/Atlda double mutant.
Starch is the most abundant storage carbohydrate in plants. It is composed primarily of amylopectin, a branched polymer of glucose in which α-1,4-linked glucan chains are connected by α-1,6-bonds (branch points). In this way, amylopectin resembles glycogen, the soluble storage carbohydrate synthesized in prokaryotes, fungi and animals. However, unlike glycogen, amylopectin molecules can form a semi-crystalline granule. This capacity stems from the branching pattern of amylopectin, which produces clusters of linear chains that pack together in regular
http://www.jbc.org/cgi/doi/10.1074/jbc.M513661200The latest version is at JBC Papers in Press. Published on February 22, 2006 as Manuscript M513661200
Copyright 2006 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
semi-crystalline arrays. This gives rise to a macromolecular granule structure in which semi-crystalline lamellae alternate with amorphous lamellae, where branch points are located (1, 2).
The way in which the insoluble starch granule is degraded to release stored glucose is understood only in the endosperm of germinating cereals. In this tissue, the process of starch breakdown is initiated by the action of α-amylase, secreted into the non-living starchy endosperm (3). This enzyme has an endoamylolytic action and releases a mixture of branched and linear oligosaccharides that serve as substrates for other hydrolytic enzymes including debranching enzymes (which hydrolyze the α-1,6-bonds) and exoamylases (β-amylases). There is mounting evidence that a different mechanism operates in living cells of other plant tissues, where the starch is degraded inside the plastid compartment. First, Arabidopsis mutants lacking all three α-amylases encoded in the genome have normal rates of starch breakdown under standard growth-room conditions, showing that the activity is not essential to initiate starch breakdown (4). Second, normal rates of starch breakdown in several plant organs require one or more enzymes belonging to the glucan water dikinase class (5-7), which phosphorylate glucosyl residues within amylopectin molecules (8). There is no evidence for a requirement for these enzymes in cereal endosperm starch degradation.
Evidence that chloroplastic enzymes other than α-amylase can attack starch granules is limited. Scheidig et al. (9) reported that a recombinant chloroplastic β-amylase from potato can liberate small amounts of maltose from isolated starch granules in vitro. Similarly, Hussain et al, (10) reported that a recombinant debranching enzyme from potato (StISA3) has a limited activity on isolated granules in
vitro. In both cases, antisense repression of these enzymes resulted in starch accumulation in potato leaves (9, 11). Similarly mutation of the orthologous genes in Arabidopsis leaves causes starch accumulation (12, 13).
A model can be envisaged in which β-amylase degrades long linear chains at the granule surface to within a few glucosyl residues of the branch points. Debranching enzyme may then remove the short branches to reveal the next layer of linear chains for further exoamylolytic attack (14). To test this hypothesis we investigated in detail the phenotypes of mutant plants lacking debranching enzyme activity. If debranching enzymes attack the granule surface, we would expect to see distinctive and predictable changes in the structure of starch polymers in their absence.
Debranching enzymes fall into two classes in plants; isoamylase (ISA) and limit dextrinase (LDA; also known as pullulanase or PUL). The Arabidopsis genome encodes three proteins of the isoamylase-type (ISA1, ISA2 and ISA3) and one of the limit dextrinase-type. Although related, the two types are readily distinguishable at the level of their amino acid sequence and also by the ability of the limit dextrinase-type, but not of the isoamylase-type, to hydrolyze the α-1,6-bonds of the yeast polysaccharide pullulan.
ISA1 and ISA2 contribute to a single isoamylase enzyme which is required for the normal biosynthesis of starch (15-17). Mutation of either gene in Arabidopsis leads to the accumulation of glucans with a different branching pattern from wild-type amylopectin, most of which accumulates as soluble phytoglycogen (17). All the branched glucans accumulated by these mutants are efficiently degraded during the dark (18), suggesting that ISA1/ISA2 may not be required for the hydrolysis of branch points during the remobilisation of starch.
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
In addition to the preliminary evidence reported for the role of ISA3 in starch breakdown (10, 12), research on maize suggests that limit dextrinase plays a role in starch breakdown. A mutation in the ZPU1 gene that encodes limit dextrinase causes a reduction in the rate of starch breakdown in leaves and in germinating seeds (19). However, in Arabidopsis, lda mutants (also called pul) are reported to have normal starch contents (12). Here we present a detailed characterization of Arabidopsis mutants lacking one or both of the debranching enzymes ISA3 and LDA. We show that both enzymes can participate in starch breakdown, present in vivo evidence that ISA3 acts at the surface of the granule and conclude that there are distinct α-amylase-dependent and α-amylase-independent mechanisms for starch granule degradation.
EXPERIMENTAL PROCEDURES Plants and growth conditions - Transposon insertion mutant lines from the Cold Spring Harbour Laboratory (Cold Spring Harbour, NY, USA), T-DNA insertion mutant lines from the Salk Institute (San Diego, CA, USA) and TILLING lines from the Arabidopsis Tilling Programme (Seattle, WA, USA) were obtained via the Nottingham Arabidopsis Stock Centre (Nottingham, UK). T-DNA insertion mutant lines from the GABI-Kat project were obtained from the Max Plank Institute for Plant Breeding Research (Cologne, Germany). Plants were grown in a controlled environment chamber. Unless otherwise stated the conditions were as follows; constant 20°C, 75% relative humidity and a 12-h/12-h light/dark cycle, with a uniform illumination of 175 µmoles photons m-2 s-1. Sown seeds were covered with a clear plastic lid and stratified at 4°C for 2 days. Lids were removed after the
cotyledons were fully emerged (approx. 10 days after sowing). Seeds were sown either directly onto a peat-based potting compost, or germinated first on fine-grade seed compost then transplanted into potting compost after 2-3 weeks. Native PAGE - Native PAGE of crude extracts of leaves for the detection of starch debranching enzyme activities was performed essentially as described previously (20). Briefly, gels contained 6 % (w/v) polyacrylamide and either 1% (w/v) red pullulan (Megazyme), 0.2% (w/v) β-limit dextrin or 0.2% (w/v) amylopectin. After electrophoresis, gels were incubated for 1-2h at 37°C in a medium containing 100 mM Tris, pH 7.2, 1 mM MgCl2, 1 mM CaCl2, 5 mM DTT. Clear bands developed in the red pullulan gel. β-Limit dextrin- and amylopectin-containing gels were stained with Lugol solution. Improved staining was achieved by incubating stained gels for 24h at 4°C and partially destaining by rinsing in water. Native gels for the detection of starch synthase (21), branching enzymes (22) and starch phosphorylase (23) were performed as described previously. Design of AtISA3 antigen and immunoblot analysis - Antibodies to the AtAMY3 protein were a gift from Dr. Jychian Chen (Academia Sinica, Taipei, Taiwan). Antibodies specific to the AtISA3 protein were purchased from Sigma Genosys (Haverhill, Suffolk, UK). A 16-amino acid peptide specific to AtISA3 (CFDELELQRRSNPRDH) was synthesized, conjugated to keyhole limpet hemacyanin and used for rabbit immunization. Anti-AtISA3 antibodies were purified by the manufacturer via immunoaffinity purification, using a column containing the covalently bound peptide antigen. The antibodies were used to probe protein gel blots of crude extracts and developed using Sigma-Fast™ (Sigma) or
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
Immun-Star™ (Bio-Rad) alkaline phosphatase reagents. Enzyme Assays - A method was developed for the quantitative assays of debranching enzyme using β-limit dextrin substrate. The β-limit dextrin was prepared as described previously (20), except that amylopectin (Sigma) was initially dissolved by boiling in water for 15 minutes. After digestion, the β-limit dextrin was precipitated by the addition 75% (v/v) methanol, 1% (w/v) KCl. Precipitate was collected by centrifugation, washed with 75% (v/v) methanol, dissolved by boiling in water for 15 minutes (final concentration of 4 mg mL-1) and stored at -20°C until used. Before use in the assay, β-limit dextrin was re-treated with 50 units mL-1 β-amylase (Megazyme, Bray, Ireland) in 10 mM NaAcetate, pH 6.0. Residual maltose was removed by passage though a NAP-25 column pre-equilibrated with assay medium (50 mM MOPS, pH 7.2, 1 mM EDTA, 1 mM DTT, 1 mM CaCl2). The β-limit dextrin and β-amylase were eluted in assay medium and residual maltose was retained in the column. Crude extracts were prepared by homogenizing leaf samples (200-300 mg), each from an individual plant, in 6 volumes of ice-cold extraction medium (50 mM MOPS, pH 7.2, 1 mM EDTA, 1 mM DTT) using an all-glass homogenizer. After centrifugation, the supernatants were desalted by passage through NAP-10 columns pre-equilibrated with assay medium. Duplicate assays (100 µL total volume) were started by the addition of the desalted extracts, incubated at 30°C for 1 h and stopped by boiling for 5 min. The assay contents were passed through sequential Dowex 50 and Dowex 1 mini-columns (24) to remove contaminating proteins and charged compounds, and neutral compounds were eluted with water. Maltose and maltotriose contents were determined by HPAEC-PAD, as described previously (25). No maltose was detected
when the plant extract was omitted. Control reactions (in which the β-limit dextrin was omitted) were used for each extract and gave very low values, which were subtracted from the assay values.
For the assay of α-amylase the CrealphaTM kit (Megazyme) was used (18). This employs a linear substrate and is not liable to interference from debranching enzymes. Carbohydrate measurements and structural analyses - Samples were harvested into liquid N2 and extracted using perchloric acid (17). Total glucans in soluble and insoluble fractions were determined by measuring the amount of glucose released by treatment with α-amylase and amyloglucosidase, as described previously (26) Chain length profiles of amylopectin were determined by HPAEC-PAD (17). Molecular methods - The full length ISA3 cDNA was cloned into the binary vector pMDC83 (27) by recombinant cloning. This construct produced a full-length ISA3 protein with GFP fused to its carboxyl terminus, under the transcriptional control of a double 35S CaMV promoter. Transient expression was carried out in protoplasts from the wild type and the starchless mutant pgm (28). Protoplasts were prepared as described by Fitzpatrick and Keegstra (29). PEG-mediated transformation was performed as described by Jin et al (30). Localization was carried out using a Leica DM IRBE and a Leica TCS SP laser confocal scanning laser microscope (Leica, Unterentfelden, Switzerland). Fluorophores were excited using a laser with 488nm wavelength. The GFP emission signal was collected in a wavelength window between 495-525nm. Chlorophyll autofluorescence was collected in a window of 625-690nm.
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
RESULTS Identification of ISA3 and LDA mutants - We identified mutant alleles of LDA and ISA3 from publicly available populations. Mutants lacking LDA were obtained from the SALK collection (http://signal.salk.edu; line SALK_060765)(31) and from the Cold Spring Harbor Gene Trap Collection (http://genetrap.cshl.org, line GT7150) and designated Atlda-2 and Atlda-3 respectively. The positions of the T-DNA insertion in Atlda-2 and of the transposon insertion in Atlda-3 were confirmed by PCR and DNA sequencing (Figure 1A) and plants homozygous for the insertions isolated. The effect of the insertions on LDA expression was determined using native PAGE of crude extracts in gels containing red pullulan (a substrate specific for LDA-class debranching enzymes). A single band of activity was detected in the two wild-type lines. In both Atlda-2 and Atlda-3 no activity was detectable (Figure 1B). Mutants lacking ISA3 were identified from the GABI-KAT collection (http://www.gabi-kat.de)(32) and via the Arabidopsis Tilling Program (http://tilling.fhcrc.org:9366)(33) and designated Atisa3-2 and Atisa3-3, respectively. The T-DNA insertion site in GABI_KAT_280G10 and the point mutation in CS88929 (TAIR polymorphism At4g09_120E3) were confirmed by PCR and DNA sequencing (Fasteris, Geneva, Switzerland)(Figure 1A). In CS88929 the EMS-induced point mutation lies on a splice junction. Plants homozygous for each of the mutations were isolated from segregating populations obtained from the source laboratories. To confirm that these mutations abolished expression of the ISA3 gene we acquired an antiserum raised to an AtISA3-specific peptide and used it to probe protein gel blots of crude extracts from the wild type and the Atisa3 mutants. A single 80 kD
protein was detected in the wild type, which was missing in both mutants (Figure 1C).
Both alleles of ISA3 had similar phenotypes. Likewise, both LDA mutants were alike. Unless specified, further work was carried out with Atisa3-2 and Atlda-2, both of which are mutants of the Columbia ecotype. We generated a double mutant by crossing Atlda-2 and Atisa3-2. The double mutant was selected from the segregating F2 population by screening for the T-DNA insertions in the respective genes using PCR.
Determination of debranching enzyme activity - Debranching enzyme activity is frequently determined using semi-quantitative native PAGE (zymogram) methods that rely on the debranching of a glucan substrate (e.g. amylopectin) incorporated into the gel. Activity is visualized by staining the gel with an iodine solution and the ISA1/ISA2 isoamylase and LDA activity are readily detected in Arabidopsis leaf extracts as clear or pale blue bands (12, 17, 18). No band of debranching enzyme activity has previously been attributed to ISA3 activity, leading to the suggestion that the protein is unstable (12). We tested a range of native gel conditions and substrates and observed that in gels containing β-limit dextrin (amylopectin treated with β-amylase to digest the external chains to within 2 or 3 glucosyl residues of a branch point), a dark blue band of activity was visible in wild-type extracts but missing in Atisa3 extracts (Figure 1D). This is consistent with the observation that the ISA3 orthologue from potato has high activity on β-limit dextrin in vitro (10).
We developed an assay to quantify β-limit dextrin debranching activity in extracts of Arabidopsis leaves (see Methods). The assay uses β-limit dextrin in a saturating background of β-amylase. The
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
external branches of β-limit dextrin are a mixture of ‘stubs’ with degree of polymerization (dp) of 2 and 3. Branches removed by debranching enzyme activity expose linear chains which are immediately degraded to maltose by β-amylase, thereby regenerating the β-limit dextrin substrate. The product of the assay is predominantly maltose (the dp2 stubs and the maltose produced by β-amylolysis of the newly-exposed linear chains), together with smaller amounts of maltotriose (the dp3 stubs). We quantified both using HPAEC-PAD (Figure 2A). Using wild-type extracts, we confirmed that this assay yields linear activity rates for both maltose and maltotriose production with respect to time and the volume of extract added in the ranges used in our experiments (data not shown).
Application of the debranching enzyme assay to extracts of wild-type and mutant plants showed that Atisa3 and Atlda both have reduced β-limit dextrin debranching activity. The production of maltose was reduced by 35% and 23% relative to the wild type in Atlda and Atisa3 respectively, and by 72% in extracts of the double mutant (Figure 2B). Interestingly, the changes in maltotriose production did not follow the changes in maltose production. Atlda extracts released slightly less maltotriose than wild-type extracts (22% reduction), whereas a large reduction (62%) was seen with Atisa3 extracts. In the double mutant extracts, almost no maltotriose was produced (96% reduction). A second experiment with a separate batch of plants yielded very similar data. Thus, both LDA and ISA3 have appreciable activity on β-limit dextrin, but ISA3 may have a greater affinity than LDA for dp3 stubs.
We also used this assay to measure the activity in the Atisa1/Atisa2 double mutant (17). No reduction in activity was seen in terms of maltose or maltotriose production (Figure 2B). This is intriguing, as
the activity is visible on a native gel containing the same substrate. One explanation is that the ISA1/ISA2 does not remove the dp2 and dp3 stubs, but rather the longer branches which carry stubs (i.e. B-chains). This would result in very little dp2 and dp3 release.
It should be noted that α-amylase may interfere with this assay as endoamylolysis will also expose chains for β-amylase. This may account for the residual maltose production in the Atlda/Atisa3 double mutant (39% of the wild-type activity). The absence of maltotriose produced in this line is also consistent with this explanation as β-amylase will release maltose from a branched substrate, but not maltotriose.
Phenotypes of Atisa3, Atlda and the double mutant - In a controlled environment chamber with a 12-h photoperiod, wild-type and Atlda mutant plants grew at the same rate whereas, Atisa3 mutants grew more slowly. The growth of the double mutant was even slower (Figure 3A and data not shown).
Based on measurements at a single time point it has been reported that an ISA3 mutant had a higher starch content than the wild type but that an LDA mutant (Atpul) did not (12). We confirmed these phenotypes and extended the analysis to examine the diurnal turnover of starch. Wild-type and Atlda plants had similar starch contents throughout the diurnal cycle (Figure 3B). This result was reproducible in three separate experiments and true for both Atlda mutant lines (data not shown). This result contrasts with the situation in maize leaves, in which the starch content was elevated in a mutant that lacks LDA (zpu1)(19). The starch content of Atisa3 leaves was higher than that of wild-type leaves throughout the diurnal cycle (Figure 3) supporting earlier suggestions of a role in
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
starch degradation (10, 12, 34). Old leaves contained more starch than young leaves (data not shown). However, appreciable amounts of starch were still degraded during the night (67.5% of that degraded by the wild type; the mean of three separate experiments including the one shown in Figure 3B), and extending the night for a further 48 h led to almost all the starch being remobilized (data not shown). These data suggest that the loss of ISA3 impairs but does not prevent the process of starch breakdown and that, in a day/night growth regime, starch accumulates in small increments due to an imbalance between daytime synthesis and night-time degradation.
To investigate how the starch is still degraded in the absence of ISA3 we analysed the Atisa3/Atlda double mutant. The phenotype of the double mutant was much more severe. Starch accumulated to higher levels than in Atisa3, and the extent of starch breakdown was further reduced (to 48.3% of the amount degraded by the wild type; the mean of two separate experiments including the one shown in Figure 3B) and, even after an extended night of 260 h, all but the youngest leaves still stained darkly for starch with iodine solution (data not shown). This suggests that in the absence of ISA3, LDA plays an important role in debranching glucans.
Glucans present in the soluble fraction of the leaf extracts were also determined. The soluble glucan content in the wild type and the two single mutant plants was very low, whereas significant amounts were detectable in the double mutant throughout the diurnal cycle. Soluble glucans in the double mutant increased during the night and decreased during the day (Figure 3B). These glucans could not be precipitated using 75% (v/v) methanol, 1% (w/v) KCl, indicating low molecular weight oligosaccharides rather than glycogen-type
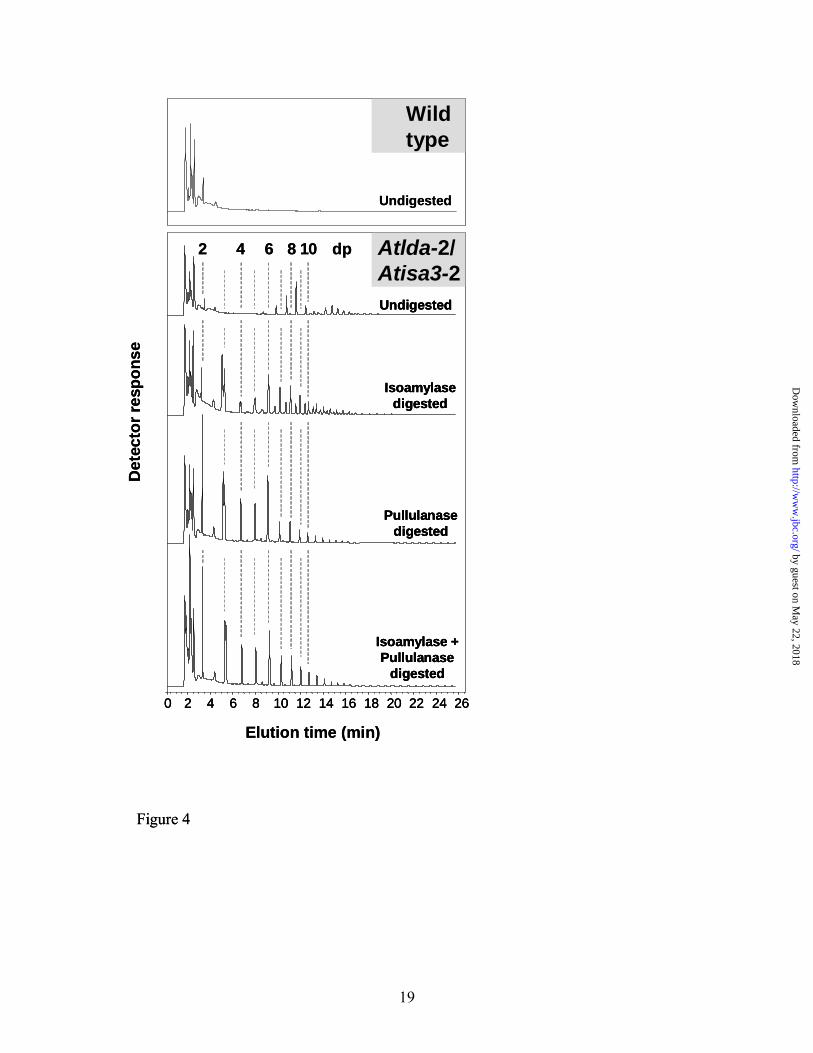
structures (data not shown). To gain more information about the nature of these oligosaccharides, we isolated the neutral compounds from the soluble extracts and analyzed them by HPAEC-PAD. The major oligosaccharide peaks in the chromatogram indicated a size range of between dp6 and dp20, but the peaks did not co-elute with linear malto-oligosaccharide standards (Figure 4). This is typical of branched oligosaccharides which elute slightly earlier than linear molecules of the same size. To evaluate this further, we treated the neutral fraction with a bacterial isoamylase, a bacterial pullulanase, or both. These treatments changed the appearance of the chromatogram. In all cases, linear malto-oligosaccharides were liberated including many chains as short as dp2. The overall size range of the oligosaccharides was reduced. Using isoamylase alone resulted in a mixture of linear and branched molecules, whereas pullulanase or both enzymes together effectively converted all the oligosaccharides to linear molecules (note that the bacterial isoamylase and pullulanase enzymes may not have exactly the same substrate specificity as the Arabidopsis proteins ISA3 and LDA, respectively). These results indicate that, in the absence of both ISA3 and LDA, branched oligosaccharides are released from the granule surface and metabolized to limit dextrins which are then inefficiently debranched. Increased chloroplastic α-amylase in Atisa3 and the double mutant - Branched oligosaccharides could be released from the granule by the action of α-amylase. Although elimination of the chloroplastic α-amylase does not result in elevated starch content under our growth conditions, it may still contribute to breakdown (4). Our native gels suggested an increased activity of an endoamylase, migrating in the same location
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
as the chloroplastic α-amylase AMY3 (Figure 1D). Therefore, we performed quantitative α-amylase measurements on crude extracts of leaves (Figure 5A). This revealed a significant increase in α-amylase activity in Atisa3 and an even greater increase in the Atisa3/Atlda double mutant. Protein gel blots probed with an antibody raised to the AMY3 protein confirmed that the amount of AMY3 protein was increased in both lines (Figure 5B). This suggests that the loss of ISA3 results in a pleiotropic increase in AMY3, which is increased further in the double mutant line. We performed further native gels to search for additional pleiotropic effects on starch biosynthetic enzymes (starch synthase and starch branching enzymes) and degradative enzymes (α-glucan phosphorylase and starch hydrolyzing enzymes), but no other differences were observed (data not shown). Changes in amylopectin structure in Atisa3 and the Atisa3/Atlda double mutant - Starch was extracted from leaves harvested at the end of the day and the chain-length distribution of the amylopectin was determined by HPAEC-PAD. The distributions of wild-type and Atlda mutant amylopectins were indistinguishable (Figure 6). In contrast, the chain length profile of amylopectin from Atisa3 differed markedly from that of the wild type, with an increase in the relative frequency of chains of dp3 and, to a lesser degree, dp4 and dp5 (Figure 6). Chains between dp9 and dp16 were decreased in relative abundance. The chain length profile of the amylopectin from the Atlda/Atisa3 double mutant was very similar to that of Atisa3. In each case, the amylopectin chain length profiles were consistent between at least three different batches of plants.
Chains of amylopectin shorter than dp6 are not thought to be produced during normal starch biosynthesis in Arabidopsis
leaves (35). Consequently, the short chains in Atisa3 and the Atisa3/Atlda double mutant most likely result from the action of degradative enzymes. Ritte et al (36) showed that starch granules isolated from potato leaves at night have more short chains on their surface (notably dp3 – dp5) compared to granules isolated from illuminated leaves. The inefficient removal of short chains from the granule surface, coupled with the repetitive diurnal cycles of synthesis and (partial) degradation could result in the build up of short chains within the granule to the levels that we observe. ISA3 localizes to the surface of the starch granule - The change in amylopectin structure in Atisa3 suggests that ISA3 could directly attack the starch granule in vivo as well as in vitro (10). To investigate this further we examined the sub-cellular localization of ISA3 by expressing an ISA3-GFP fusion protein in protoplasts. Confocal microscopy of transformed wild-type cells revealed GFP fluorescence from within the chloroplasts (Figure 7A-G). However, the signal was not uniformly distributed, but was strongest from oval structures within the chloroplasts that were similar in size, number and distribution to starch granules. This pattern of fluorescence was observed in several independent transformation experiments. In most cases the GFP fluorescence surrounded the oval structures, from the middle of which there was no chlorophyll fluorescence (Figure 7B-D). This is consistent with the presence of a starch granule, which would displace the chlorophyll-containing thylakoid membranes. We expressed the same construct in protoplasts isolated from the starchless mutant pgm (Figure 7H-N). In this case the GFP fluorescence signal was also exclusively from the chloroplast but the signal was uniform. These results provide
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
evidence that ISA3 is present at the surface of the starch granule.
DISCUSSION The work described here reveals the important role that two debranching enzymes play in starch breakdown inside chloroplasts and allows us to propose that there are distinct mechanisms of hydrolytic starch breakdown. ISA3 and LDA are both active on β-limit dextrins - Our results show that both Atisa3 and Atlda have lost single isoforms of debranching enzyme activity, detectable using β-limit dextrin-containing native gels. Both mutants also have a reduction in total β-limit dextrin debranching activity and the double mutant has a greater reduction. Furthermore, the results from the assay imply that ISA3 and LDA may have different specificities. Maltotriose production is reduced only slightly in Atlda, but markedly reduced in Atisa3, whereas the reduction in maltose production is similar in both lines. This could indicate that ISA3 preferentially removes dp3 stubs from the β-limit dextrin. However, an alternative explanation is that the increase in α-amylase detected in Atisa3 could interfere with the assay by enhancing maltose production resulting in an overestimated activity in Atisa3. Further experiments are required to confirm a preference of ISA3 for dp3 stubs. However, it is worth noting that the stubs which are over-represented in Atisa3 amylopectin are primarily dp3. Interestingly, the stubs in the soluble limit dextrins which accumulate when both Atisa3 and Atlda are missing are predominantly dp2. Therefore, we speculate that the results of our assay indicate a preference of ISA3 and LDA for stubs of dp3 and dp2 respectively.
If the rate of maltotriose production is assumed to represent half the rate of β-
limit dextrin debranching activity, it is possible to calculate an activity of 7.3 nmoles α-1,6-linkages min-1 g-1 FW for the wild type (the average wild type rate from figure 2B and a replicate experiment not shown). This is more than sufficient to catalyze the observed rate of branch point hydrolysis during starch breakdown (4 nmoles α-1,6-linkages min-1 g-1 FW, assuming that 4% of the linkages in starch are branch points). We emphasize that this value should be treated with caution as the assays have not been optimised and the activities of the enzymes may be different when acting on substrates in vivo. The role of ISA3 - Earlier results have implied that ISA3 is involved in starch degradation. A starch-excess phenotype is caused by antisense repression or mutation in potato and Arabidopsis, respectively (11, 12). In addition, ISA3 expression correlates with periods of starch breakdown in ripening banana fruit and Arabidopsis leaves (34, 37). Our results are consistent with these earlier reports and provide additional insight into the role that the enzyme plays in starch breakdown.
Firstly, the starch structure of the amylopectin in Atisa3 is altered with respect to wild-type amylopectin with increases in very short chains (especially dp3). The variability of our data is low and the finding was consistent in numerous batches of plants grown in a range of different conditions. This result contrasts to the conclusions of Wattebled et al, (12) where no changes in amylopectin structure were found. However, chains shorter than dp5 were not determined in this study, explaining this discrepancy. The increase in short chains implies that ISA3 would normally remove them and is consistent with the observation that ISA3 from potato has a particularly high activity on β-limit dextrin relative to other substrates (10).
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
Secondly, our results show that the ISA3-GFP fusion protein localises to the periphery of structures that are similar in size, shape and number to starch granules within wild-type chloroplasts. The uniform GFP signal inside starchless chloroplasts is consistent with the idea that these structures are indeed starch granules. If so, these results suggest that ISA3 acts on the surface of the granule, most likely during starch breakdown. If exoamylases (i.e. β-amylase) attack the surface of the granules, as has been suggested (9), short branches would be produced and further β-amylolysis would not be possible until these branches had been removed. Thus, ISA3 could act in conjunction with β-amylase to progressively degrade the starch granule. According to this hypothesis, some of the short chains that are exposed during breakdown are not removed in Atisa3, limiting the rate of degradation. Many of these short chains could be elongated again upon subsequent starch biosynthesis in the light, but some may become buried in the granule leading to the changes in chain length distribution that we observe. Although such short branches are not thought to be produced during normal amylopectin synthesis (35), we cannot exclude the possibility that ISA3 is also active during starch biosynthesis.
Thirdly, although the starch content of the Atisa3 leaves is significantly increased, there is still appreciable night-time degradation. It is possible that this occurs via the activity of the chloroplastic α-amylase AMY3. α-Amylase could release branched glucans from the granule which would serve as substrates for other stromal enzymes. This would simultaneously expose linear chains on the granule surface for β-amylolysis. The increase in chloroplastic α-amylase activity in Atisa3 provides some support for this hypothesis.
The role of LDA - Arabidopsis mutants lacking LDA do not display a high starch phenotype (this study)(12), although in maize, the equivalent mutant does (19). However, our results show that when the LDA and ISA3 are both missing, the double mutant line has a much higher starch content and a slower rate of starch breakdown. This suggests that LDA is involved in starch breakdown, at least in the Atisa3 mutant background. Importantly, this double mutant accumulates small, soluble branched glucans during the dark. These have the characteristics of limit-dextrins and are not observed in the single mutants or in the wild type. Furthermore, the chloroplastic α-amylase is increased appreciably in this line. Together, these data suggest that soluble branched glucans are produced by α-amylase give rise to limit-dextrins in the stroma, which can then be metabolised by limit dextrinase into linear glucans. Low levels of limit-dextrins were also observed in the LDA mutant of maize (19).
It remains possible that the ISA1/ISA2 isoamylase also contributes to debranching during starch breakdown. However, the slow rate of degradation observed, the accumulation of branched oligosaccharides in the Atisa3/Atlda double mutant, and the unaltered debranching enzyme activity on β-limit dextrin all suggest that its activity is either not sufficient or not suited to this role. Two pathways for starch granule attack in chloroplasts - Our data are consistent with the idea that there are several enzymes which can attack starch granules in vivo. We propose that in Arabidopsis, the major pathway involves β-amylase and ISA3, which progressively degrade the starch granule surface, releasing maltose for export from the chloroplast (38, 39) and short linear oligosaccharides for further metabolism (25) (Figure 8A). The predominance of this
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
pathway explains the absence of a starch-excess phenotype in Arabidopsis mutants lacking either AMY3 or LDA (4, 12).
We suggest that α-amylase can also attack the starch granule to release branched oligosaccharides that are degraded in the stroma by a combination of β-amylase and LDA, but that this pathway is, under normal circumstances not required in Arabidopsis leaves. In the absence of ISA3, however, the amount of α-amylase is increased and the flux through this pathway accounts for the observed rate of degradation (Figure 8B). The simultaneous removal of ISA3 and LDA would affect both of the proposed pathways, but would still allow some starch to be broken down. α-Amylase, further up-regulated in this line, can attack the granule exposing linear chains and the soluble glucans released can be degraded as far as branched limit dextrins (Figure 8C). Therefore, a slow rate of degradation is still achieved.
This model requires further testing to confirm the distinct roles of each enzyme and to establish whether the data for Arabidopsis are generally applicable to the leaves of other plants. Under some conditions, or in other species, the relative importance of these pathways may be different. For instance, in maize leaves a greater fraction of the flux might proceed via α-amylase and LDA, explaining the reduced starch breakdown in the maize LDA mutant (19). Similarly, in rice, repression of α-amylase results in elevated levels of leaf starch (40). On the other hand, the importance of ISA3 and β-amylase are not yet known for these species. It seems likely that enzymes attacking the granule are dependent on its prior phosphorylation by enzymes of the glucan, water dikinase family. However, the nature of this dependency is not understood and also needs further investigation.
REFERENCES
1. French, D. (1984) Starch: chemistry and technology. Academic Press, Orlando, USA 2. Buléon, A., Colonna, P., Planchot, V., and Ball, S. (1998) Internat. J. Biol.
Macromolecules 23, 85-112 3. Fincher, G. B. (1989). Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 305-346 4. Yu, T.-S., Zeeman, S. C., Thorneycroft, D., Fulton, D. C., Dunstan, H., Lue, W.-
L., Hegemann, B., Tung, S.-Y., Umemoto, T., Chapple, A., Tsai, D.-L., Wang, S.-M, Smith, A. M., Chen, J., and Smith, S. M. (2005) J. Biol. Chem. 280, 9773-9779
5. Lorberth, R., Ritte, G., Willmitzer, L., and Kossmann, J. (1998) Nature Biotech. 16, 473-477
6. Yu, T.-S., Kofler, H., Häusler, R. E., Hille, D., Flügge, U.-I., Zeeman, S. C., Smith, A. M., Kossmann, J., Lloyd, J., Ritte, G., Steup, M., Lue, W.-L., Chen, J., and Weber, A. (2001) Plant Cell 13, 1907-1918
7. Kötting, O., Pusch, K., Tiessen, A., Geigenberger, P., Steup, M., and Ritte, G. (2005) Plant Physiol. 137, 242-252
8. Ritte, G., Lloyd, J. R., Eckermann, N., Rottmann, A., Kossmann, J., and Steup, M. (2002) Proc. Natl. Acad. Sci. USA 99, 7166–7171
9. Scheidig, A., Frohlich, A., Schulze, S., Lloyd, J. R., and Kossmann, J. (2002) Plant J. 30, 581-591
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
10. Hussain H., Mant, A., Seale, R., Zeeman S. C., Hinchliffe, E., Edwards, A., Hylton, C., Borneman, S., Smith A. M., Martin, C., and Bustos R. (2003) Plant Cell 15, 133-149
11. Hussain M. H. (2002) PhD thesis, University of East Anglia, UK 12. Wattebled, F., Dond, Y., Dumez, S., Delvalle, D., Planchot, R., Berbezy, P., Vyas,
D., Colonna, P., Chatterjee, M., Ball, S., and D'Hulst, C. (2005) Plant Physiol. 138, 184-195
13. Kaplan, F., and Guy C. L. (2005) Plant J. 44, 730-743 14. Smith, A. M., Zeeman, S. C., and Smith S. M. (2005) Annu. Rev. Plant Biol. 56,
73-97 15. Myers, A. M., Morell, M. K., James, M. G. and Ball, S. G. (2000) Plant Physiol.
122, 989-997 16. Bustos, R., Fahy, B., Hylton, C. M., Seale, R., Nebane, N. M., Edwards, A.,
Martin, C., and Smith, A. M. (2004) Proc. Natl. Acad. Sci. USA 101, 2215-2220 17. Delatte, T., Trevisan, M., Parker, M. L., and Zeeman S. C. (2005) Plant J. 41,
815-830 18. Zeeman, S. C., Umemoto, T., Lue, W. L., Au-Yeung, P., Martin, C., Smith, A.
M., and Chen, J. (1998) Plant Cell 10, 1699-1711 19. Dinges, J. R., Colleoni, C., James, M. G. and Myers, A. M. (2003) Plant Cell 15,
666-680 20. Zeeman, S. C., Northrop, F., Smith, A. M., and ap Rees, T. (1998) Plant J. 15,
357-365 21. Edwards, A., Marshall, J., Sidebottom, C., Visser, R. G. F., Smith, A. M., and
Martin, C. (1995) Plant J. 8, 283-294 22. Yamanouchi, H., and Nakamura, Y. (1992) Plant Cell Physiol. 33, 985-991 23. Steup, M. (1990) Methods Plant Biochem. 3, 103-128 24. Harley, J. L., and Beevers, H. (1963) Plant Physiol. 38, 117-123 25. Critchley, J. H., Zeeman, S. C., Takaha, T., Smith, A. M., and Smith, S. M.
(2001) Plant J. 26, 89–100 26. Hargreaves, J. A., and ap Rees, T. (1988) Phytochem. 27, 1627-1629 27. Curtis, M. D., and Grossniklaus, U. (2003) Plant Physiol. 133, 462-469 28. Caspar, T., Huber, S. C., and Sommerville, C. R. (1985) Plant Physiol. 79, 11-17 29. Fitzpatrick, L. M. and Keegstra, K. (2001) Plant J 27, 59-65 30. Jin B. J., Kim, Y. A., Kim, S. J., Lee, S. H., Kim, D. H., Cheong, G. W., Hwang,
I. (2001) Plant Cell 13, 1511-1525 31. Alonso, J. M., Stepanova, A. N., Leisse, T. J., Kim, C. J., Chen, H., Shinn, P.,
Stevenson, D. K., Zimmerman, J., Barajas, P., Cheuk, R., Gadrinab, C., Heller, C., Jeske, A., Koesema, E., Meyers, C. C., Parker, H., Prednis, L., Ansari, Y., Choy, N., Deen, H., Geralt, M., Hazari, N., Hom, E., Karnes, M., Mulholland, C., Ndubaku, R., Schmidt, I., Guzman, P., Aguilar-Henonin, L., Schmid, M., Weigel, D., Carter, D. E., Marchand, T., Risseeuw, E., Brogden, D., Zeko, A., Crosby, W. L., Berry, C. C., and Ecker, J. R. (2003) Science 301, 653-657
32. Rosso, M. G., Li, Y., Strizhov, N., Reiss, B., Dekker, K., and Weisshaar, B. (2003) Plant Mol. Biol. 53, 247-259
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
33. Till, B. J., Reynolds, S. H., Greene, E. A., Codomo, C. A., Enns, L. C., Johnson, J. E., Burtner, C., Odden, A. R., Young, K., Taylor, N. E., Henikoff, J. G., Comai, L., and Henikoff, S. (2003) Genome Res. 13, 524-530
34. Bierhals, J. D., Lajolo, F. M., Cordenunsi, B. R., and Do Nascimento, J. R. O. (2004). J. Agric. Food Chem. 52, 7412-7418
35. Nielsen, T. H., Baunsgaard, L., and Blennow, A. (2002) J. Biol. Chem. 277, 20249-20255
36. Ritte, G., Scharf, A., Eckermann, N., Haebel, S., and Steup, M. (2004) Plant Physiol. 135, 2068–2077
37. Smith, S. M., Fulton, D. C., Chia, T., Thorneycroft, D., Chapple, A., Dunstan, H., Hylton, C., Zeeman, S. C., and Smith A. M. (2004) Plant Physiol. 136, 2687–2699
38. Niittylä, T., Messerli, G., Trevisan, M., Chen, J., Smith, A. M., and Zeeman, S. C. (2004) Science 303, 87-89
39. Weise, S., Weber, A.P.M., and Sharkey, T.D. (2004) Planta 218, 474–482 40. Asatsuma, S., Sawada1, C., Itoh1, K., Okito, M., Kitajima. A., and Mitsui T.
(2005) Plant Cell Physiol. 46, 858-869
FOOTNOTES We thank Cris Kuhlemeier for his support, Alison Smith and Cathie Martin for valuable discussions, Christopher Ball and Rebecca Alder for assistance in growing the plants. We also thank the Salk Institute Genomic Analysis Laboratory, the GABI-Kat project and the Cold Spring Harbour Laboratory for providing the sequence-indexed Arabidopsis T-DNA insertion mutants. We thank the Arabidopsis Tilling Programme for identifying point mutations in Atg09020 and the European Arabidopsis Stock Centre for distributing seed lines. The work was funded by the Swiss National Science Foundation (Grant 3100-067312.01/1).
FIGURE LEGENDS Fig 1. Identification and analysis of mutants at the AtLDA and AtISA3 loci. (A) Structure of the AtLDA and AtISA3 genes; exons are depicted as black boxes. Grey boxes depict 5’ and 3’ untranslated regions. T-DNA left border (LB) and transposon sequences are shown on a hatched background. Sequence lacking similarity to either the target gene or the T-DNA insert is underlined. T-DNA and transposon insertions in Atlda-2 and Atlda-3 occurred in exons 7 and 12, respectively. In Atisa3-2, a T-DNA insertion occurred in exon 21 and in Atisa3-3, a point mutation is present at the splice junction between intron 2 and exon 3. (B) Proteins in crude extracts of leaves from the Columbia wild type (WT Col), Atlda-2, Landsberg erecta wild type (WT L.er) and Atlda-3 were separated by native PAGE in gels containing red pullulan. LDA activity was detected as a clear band against the background. (C) Proteins in crude extracts of leaves from the wild type (WT), Atisa3-2, Atisa3-3 were separated by SDS-PAGE, blotted onto PVDF membranes and probed with anti-AtISA3 antibodies. Arrow indicates the migration position of a 75 kD molecular weight marker.
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
(D) Proteins in crude extracts of leaves from the wild type (WT), Atlda-2, Atisa3-2 and the Atlda/Atisa3 double mutant were separated by native PAGE in gels containing β-limit dextrin. After incubation of the gel, debranching enzyme activities were detected as blue-staining bands by iodine staining. D1 is the ISA1/ISA2 isoamylase. D2 is absent in Atlda-2 and D3 is absent in Atisa3-2. The colourless band A2 is the chloroplastic α–amylase AMY3. Fig 2. Determination of debranching enzyme activity in extracts of leaves. (A) Crude extracts of wild-type (WT), Atisa3-2 and Atlda-2 and Atisa3/Atlda double mutant leaves were desalted and incubated with β-limit dextrin substrate in the presence of saturating β-amylase (see Experimental procedures). Maltose (released by debranching and β-amylolysis of linear chains) and maltotriose (released by debranching) were detected by HPAEC-PAD. Representative chromatograms are shown. (B) Quantitative comparison of debranching enzyme activity measured as described in (A) in terms of maltose (top) and maltotriose production (bottom).Values are the means ± standard error from four replicate extracts, each made from an individual plant. Fig 3. Growth and starch-accumulating phenotypes of the wild-type, Atisa3, Atlda and the Atisa3/Atlda double mutant. (A) Wild-type (WT), Atlda-2, Atisa3-2 and Atisa3/Atlda double mutant plants after 5 weeks growth (top) and leaves harvested at the end of a 12-h night, decolorized with hot ethanol and stained for starch with iodine solution (bottom). (B) Samples comprising all the leaves of individual wild-type (open circles), Atlda-2 (closed triangles), Atisa3-2 (open squares) and Atlda/Atisa3 (closed diamonds) double mutant plants were harvested and immediately frozen in liquid N2. Insoluble (upper graph) and soluble (lower graph) glucans were extracted using perchloric acid and measured as described in Experimental procedures. Each point is the mean ± standard error from four replicate samples. Fig 4. Analysis of soluble glucans extracted from the Atisa3/Atlda double mutant at the end of the night. Neutral soluble compounds from perchloric acid-extracted leaves of the wild type (top panel) and the Atlda/Atisa3 double mutant lower panel were analysed by HPAEC-PAD. Samples from Figure 3B (24-h time point) were used and representative chromatograms are shown in each case. Samples were digested with Pseudomonas isoamylase, Klebsiella pullulanase or both. The chromatograms of the digested wild-type samples did not differ from the undigested sample and are not shown. The dotted lines indicate the retention times of linear malto-oligosaccharide standards with degrees of polymerization (dp) indicated. Fig 5. Increased chloroplastic α-amylase activity in Atisa3-2 and Atisa3/Atlda double mutant leaves. (A) α-Amylase activity in crude extracts of wild-type (WT), Atisa3-2, Atlda-2 and Atisa3/Atlda leaves. Measurements are the means ± standard errors of 4 extracts, each made from an individual plant.
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15
(B) Protein gel blots of the crude extracts measured in (A) probed with an antibody raised to the AMY3 chloroplastic α-amylase. The left-hand lane contains protein molecular weight markers, sizes given in kD. Replicate blots were performed with all 4 extracts, yielding similar results to those shown. Fig 6. Comparisons of the chain length distributions of amylopectin extracted from leaves of wild-type, Atisa3-2, Atlda-2 and the Atisa3/Atlda double mutant plants. Glucans were extracted from 4-6 individual plants at the end of the day. Equal quantities of starch from each plant were pooled, debranched with Pseudomonas isoamylase, and analyzed by HPAEC-PAD. Peak areas were summed and the areas of individual peaks expressed as a percentage of the total. The means ± standard errors of 3 independent isoamylase digests are shown. For difference plots, standard errors of the compared data sets were added together. Fig 7. AtISA3 localises to granule-like structures inside chloroplasts. Protoplasts from wild type (A-G) and the starchless mutant pgm (H-N) were transformed with a construct encoding ISA3 with GFP fused to its C-terminus. A and H: bright-field images of protoplasts. B and I; native chlorophyll fluorescence (the red objects are individual chloroplasts). C and J; GFP fluorescence. D, E, F and G; merged GFP- and chlorophyll-fluorescence images of wild-type protoplasts. G is an enlargement of D. E and F show different protoplasts. Note the non-uniform distribution of the GFP signal obtained in starch-containing wild-type chloroplasts. The GFP signal is distributed around structures that do not show chlorophyll fluorescence (marked with arrows). K, L, M and N; merged GFP- and chlorophyll-fluorescence images of pgm protoplasts. N is an enlargement of K. L and M show different protoplasts. Note the uniform distribution of the GFP signal in the starchless pgm chloroplasts. Several independent transformation experiments were performed with each line, yielding the same results. Scale bars = 8 µm. Fig 8. Distinct mechanisms for starch granule breakdown in Arabidopsis leaves. This model proposes that the starch granule surface is subject to the attack of multiple enzymes. In wild-type plants (A) exoamylases (β-amylases) release maltosyl residues from the surface of the granule to expose a β-limit dextrin structure with short external chains or ‘stubs’ (with dp3 or larger). The stubs are removed by the action of ISA3 and any linear chains longer than dp3 are degraded further by β-amylases to yield maltose and maltotriose. In the absence of ISA3 (B), endoamylases (α-amylases) release branched oligosaccharides into the stroma, which are degraded by a combination of exoamylases and LDA. If LDA is also absent, branched limit dextrins accumulate in the stroma (C).
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

16
Figure 1
ATG
T-DNALB
Atlda-2 Atlda-3
TransposonCATCCCTGATTGTACT
At5g04360; AtLDA
ATGAtisa3-3
TGATA (G A) GATTT
T-DNA LB
Atisa3-2
At4g09020; AtISA3
CTTTTCTG (30bp) AACAAATT
ATATCCTGTAGATTTCACACC
A
WT Atisa3-2 Atisa3-3
75 kD
C
WTCol Atlda-2 Atlda-3
WTL.er
B
D1
D2
D3
WT Atlda-2 Atisa3-2Atisa3-2/Atlda-2
D
A2
Figure 1
ATG
T-DNALB
Atlda-2 Atlda-3
TransposonCATCCCTGATTGTACT
At5g04360; AtLDA
ATGAtisa3-3
TGATA (G A) GATTT
T-DNA LB
Atisa3-2
At4g09020; AtISA3
CTTTTCTG (30bp) AACAAATT
ATATCCTGTAGATTTCACACC
A
WT Atisa3-2 Atisa3-3
75 kD
C
WTCol Atlda-2 Atlda-3
WTL.er
B
D1
D2
D3
WT Atlda-2 Atisa3-2Atisa3-2/Atlda-2
D
A2
ATG
T-DNALB
Atlda-2 Atlda-3
TransposonCATCCCTGATTGTACT
At5g04360; AtLDA
ATGAtisa3-3
TGATA (G A) GATTT
T-DNA LB
Atisa3-2
At4g09020; AtISA3
CTTTTCTG (30bp) AACAAATT
ATATCCTGTAGATTTCACACC
A
ATG
T-DNALB
Atlda-2 Atlda-3
TransposonCATCCCTGATTGTACT
At5g04360; AtLDA
ATGAtisa3-3
TGATA (G A) GATTT
T-DNA LB
Atisa3-2
At4g09020; AtISA3
CTTTTCTG (30bp) AACAAATT
ATATCCTGTAGATTTCACACC
A
WT Atisa3-2 Atisa3-3
75 kD
C
WTCol Atlda-2 Atlda-3
WTL.er
B
D1
D2
D3
WT Atlda-2 Atisa3-2Atisa3-2/Atlda-2
D
A2
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17
0.0 2.0 4.0 6.0 8.0 10.0
WT
Atlda-2
Atisa3-2
Atlda-2/Atisa3-2
dp2 dp3
Elution time (min)
Det
ecto
r res
pons
e
WT
Atld
a-2
Atisa
3-2
Atld
a-2/A
tisa3
-2
Mal
to-o
ligos
acch
arid
e pr
oduc
tion
(nm
oles
min
-1g
FW-1
)
BA
0
4
8
12
16
0
1
2
3
4
dp2
dp3
Atisa
1-1/A
tisa2
-1
Figure 2
0.0 2.0 4.0 6.0 8.0 10.0
WT
Atlda-2
Atisa3-2
Atlda-2/Atisa3-2
dp2 dp3dp2 dp3
Elution time (min)
Det
ecto
r res
pons
e
WT
Atld
a-2
Atisa
3-2
Atld
a-2/A
tisa3
-2
Mal
to-o
ligos
acch
arid
e pr
oduc
tion
(nm
oles
min
-1g
FW-1
)
BA
0
4
8
12
16
0
1
2
3
4
dp2
dp3
Atisa
1-1/A
tisa2
-1
Figure 2
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

18
0
10
20
30
40
Star
ch c
onte
nt(m
g/g
FW)
0
0.4
0.8
1.2
0 4 8 12 16 20 24
Time (h)
Solu
ble
gluc
an c
onte
nt
(mg/
g FW
)
Light DarkB
A
Wild type Atlda-2 Atisa3-2 Atlda-2/Atisa3-2
Figure 3
0
10
20
30
40
Star
ch c
onte
nt(m
g/g
FW)
0
0.4
0.8
1.2
0 4 8 12 16 20 24
Time (h)
Solu
ble
gluc
an c
onte
nt
(mg/
g FW
)
Light DarkB
A
Wild type Atlda-2 Atisa3-2 Atlda-2/Atisa3-2
0
10
20
30
40
Star
ch c
onte
nt(m
g/g
FW)
0
0.4
0.8
1.2
0 4 8 12 16 20 24
Time (h)
Solu
ble
gluc
an c
onte
nt
(mg/
g FW
)
Light DarkB
0
10
20
30
40
Star
ch c
onte
nt(m
g/g
FW)
0
0.4
0.8
1.2
0 4 8 12 16 20 24
Time (h)
Solu
ble
gluc
an c
onte
nt
(mg/
g FW
)
Light DarkB
A
Wild type Atlda-2 Atisa3-2 Atlda-2/Atisa3-2
A
Wild type Atlda-2 Atisa3-2 Atlda-2/Atisa3-2
Figure 3
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from
19
6 8 12 4
0 2 4 6 8 10 12 14 16 18 20 22 24 26
0 dp
Det
ecto
r res
pons
e
Elution time (min)
Undigested
Isoamylasedigested
Pullulanasedigested
Isoamylase +Pullulanase
digested
Undigested
Atlda-2/Atisa3-2
Wildtype
Figure 4
6 8 12 4
0 2 4 6 8 10 12 14 16 18 20 22 24 260 2 4 6 8 10 12 14 16 18 20 22 24 26
0 dp
Det
ecto
r res
pons
e
Elution time (min)
UndigestedUndigested
Isoamylasedigested
Isoamylasedigested
Pullulanasedigested
Pullulanasedigested
Isoamylase +Pullulanase
digested
Isoamylase +Pullulanase
digested
Undigested
Atlda-2/Atisa3-2
Wildtype
Figure 4
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

20
Figure 5
0
0.04
0.08
0.12
0.16
0.2
B
A
WT Atlda-2 Atisa3-2 Atlda-2/Atisa3-2
WT Atlda-2 Atisa3-2 Atlda-2/Atisa3-2
Activ
ity (a
rbitr
ary
units
)
100
75
Figure 5
0
0.04
0.08
0.12
0.16
0.2
B
A
WT Atlda-2 Atisa3-2 Atlda-2/Atisa3-2
WT Atlda-2 Atisa3-2 Atlda-2/Atisa3-2
Activ
ity (a
rbitr
ary
units
)
100
75
100
75
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

21
R
elat
ive
perc
enta
geWT
0
1
2
3
4
5
6
7
8
Atlda-2
0
1
2
3
4
5
6
7
Rel
ativ
e pe
rcen
tage
Atisa3-2
0
1
2
3
4
5
6
7
Rel
ativ
e pe
rcen
tage
Chain length
Atlda-2/Atisa3-2
0
1
2
3
4
5
6
7
3 13 23 33 43 53 63
Rel
ativ
e pe
rcen
tage
Diff
eren
ce
Atlda-2 - WT
-3
-2
-1
0
1
2
3
Atisa3-2 - WT
-3
-2
-1
0
1
2
3
Diff
eren
ce
Chain length
Atlda-2/Atisa3-2 - WT
-3
-2
-1
0
1
2
3
3 13 23 33 43 53 63
Diff
eren
ce
Figure 6
Rel
ativ
e pe
rcen
tage
WT
0
1
2
3
4
5
6
7
8
Atlda-2
0
1
2
3
4
5
6
7
Rel
ativ
e pe
rcen
tage
Atisa3-2
0
1
2
3
4
5
6
7
Rel
ativ
e pe
rcen
tage
Chain length
Atlda-2/Atisa3-2
0
1
2
3
4
5
6
7
3 13 23 33 43 53 63
Rel
ativ
e pe
rcen
tage
Rel
ativ
e pe
rcen
tage
WT
0
1
2
3
4
5
6
7
8
Atlda-2
0
1
2
3
4
5
6
7
Rel
ativ
e pe
rcen
tage
Atisa3-2
0
1
2
3
4
5
6
7
Rel
ativ
e pe
rcen
tage
Chain length
Atlda-2/Atisa3-2
0
1
2
3
4
5
6
7
3 13 23 33 43 53 63
Rel
ativ
e pe
rcen
tage
Diff
eren
ce
Atlda-2 - WT
-3
-2
-1
0
1
2
3
Atisa3-2 - WT
-3
-2
-1
0
1
2
3
Diff
eren
ce
Chain length
Atlda-2/Atisa3-2 - WT
-3
-2
-1
0
1
2
3
3 13 23 33 43 53 63
Diff
eren
ceD
iffer
ence
Atlda-2 - WT
-3
-2
-1
0
1
2
3
Atisa3-2 - WT
-3
-2
-1
0
1
2
3
Diff
eren
ce
Chain length
Atlda-2/Atisa3-2 - WT
-3
-2
-1
0
1
2
3
3 13 23 33 43 53 63
Diff
eren
ce
Figure 6
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

22
A B C D
Figure 7
I J KH
E F G
L M N
A B C D
Figure 7
I J KH
E F G
L M N by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

23
Figure 8
ISA3
β-amylase
α-amylaseLDA ISA3
β-amylase
Linear oligosaccharides
Maltose
Maltotriose
Branchedoligosaccharides
β-amylase
α-amylaseLDA
β-amylase
Linear oligosaccharides
Maltose
Maltotriose
Branchedoligosaccharides
β-amylase
α-amylase
β-amylase
Limitdextrins
Maltose
Branchedoligosaccharides
A: Wild Type
B: Atisa3
C: Atisa3/Atlda
Figure 8
ISA3
β-amylase
α-amylaseLDA ISA3
β-amylase
Linear oligosaccharides
Maltose
Maltotriose
Branchedoligosaccharides
β-amylase
α-amylaseLDA
β-amylase
Linear oligosaccharides
Maltose
Maltotriose
Branchedoligosaccharides
β-amylase
α-amylase
β-amylase
Limitdextrins
Maltose
Branchedoligosaccharides
A: Wild Type
B: Atisa3
C: Atisa3/Atlda
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Steven M. Smith and Samuel C. ZeemanThierry Delatte, Martin Umhang, Martine Trevisan, Simona Eicke, David Thorneycroft,
Evidence for distinct mechanisms of starch granule breakdown in plants
published online February 22, 2006J. Biol. Chem.
10.1074/jbc.M513661200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on May 22, 2018
http://ww
w.jbc.org/
Dow
nloaded from


















