
Evidence for a C-terminal turn in PBAN An NMR and distance geometry study
8
Int. J. Peptide Protein Rex 47, 1996, 361-368 Printed in UK ~~ ull rights reserved INTERNATIONAL JOURNAL OF PEPTIDE & PROTEIN RESEARCH iSSN U367-6.377 Evidence for a C-terminal turn in PBAN An NMR and distance geometry study BARBARA A. CLARK and GLENN D. PRESTWICH Department of Chemistry, University at Stony Brook, Stony Brook, New York, USA Received 11 July, revised 27 September, accepted for publication 8 December 1995 The solution conformation of the pheromone biosynthesis activating neuropeptide (PBAN ) of the moth Helicoverpu zea has been determined using homonuclear two-dimensional nuclear magnetic resonance techniques and distance geometry-restrained energy minimization. The insect peptide hormone showed a random distribution of conformers in aqueous solution, whereas in a less polar medium of trifluoroethanol and water, a reordering process was observed. In particular, the C-terminal region (Phe-Ser-Pro-Arg- Leu-NH,) adopts a type I’ /I-turn conformation, residues 20-27 are in a helix conformation, and residues 1-19 exhibit a high degree of flexibility. Direct observation of the C-terminal /I-turn configuration of PBAN is consistent with a previous report that showed a rigid, cyclic analog of the C-terminal pentapeptide of PBAN retained pheromonotropic activity [ Nachman, R.J., Kuniyoshi, H., Roberts, V.A., Holman, G.M. & Suzuki, A. (1993). Biochem. Biophys. Res. Compnun. 661-6661, Furthermore, our results show no interaction between the C-terminal turn and the rest of the polypeptide chain, thus providing further evidence that the C-terminal turn is indeed the important conformation recognized by the PBAN receptor. 0 Munksgaard 1996. Key words: Helicovrrpu zeu; NMR spectroscopy; peptide hormone; sex pheromone The pyrokinins are a group of insect neuropeptides that share the C-terminal pentapeptide sequence, Phe- Xaa-Pro-Arg-Leu-NH,, where Xaa can be Thr, Ser, Gly or Val (1). The first member of the pyrokinin family to be isolated and characterized was leucopyro- kinin (Lem-PK) of the cockroach Leucophaea mad- erae, and the function of Lem-PK is to stimulate contractions in the hindgut and oviduct (2, 3). Like Lem-PK, the five pyrokinins of the locust Locusta migmtoria, labeled locustaniyotropins ( Lom-PK, Lorn-MT-I, -11, -111 and -1V) are involved in myotropic activities (4-7). Also sharing the C-ter- minal sequence of the pyrokinins are the diapause hormone of the silkworm moth Bombyx mori ( Born-DH) (S), and the pheromone biosynthesis activating neuropeptides (PBANs) of the corn ear- worm moth Helicoverpa :ea (Hez-PBAN) (9-1 l), Abbreviations: 2D, two-dimensional; COSY, homonuclear correla- tion spectroscopy; DG, distance geometry; DQF, double quantum filter; Hez-PBAN, pheromone biosynthesis activating neuropeptide of the moth Hrlicovrrpu zru; NMR, nuclear magnetic resonance spectroscopy; NOE, nuclear Overhauser enhancement; NOESY, NOE correlation spectroscopy; REM, restrained energy minimiza- tion; RMSD, root mean squared deviation; TFE, 2,2,2-trifluoro- ethanol; TOCSY, total correlation spectroscopy. the silkworm moth Bombyx mori (Bom-PBAN) (12), and the gypsy moth Lymantriu dispar (Lyd-PBAN) ( 13). Interestingly, there is a biological cross-activity among the pyrokinin family of peptides. Hez-PBAN demonstrates myotropic activity in the cockroach, displaying 25% of Lem-PK activity (1). In addition, Lem-PK, Lom-PK, Lom-MT-I and Lom-MT-11 all show pheromonotropic activity in the silkworm moth (14, 15). As shown in Table 1, the similarity in the C- terminal amino acid sequence is characteristic of the pyrokinin family. The importance of this region was determined when structure-activity studies revealed the minimum sequence required for myotropic and pheromonotropic activities was the C-terminal penta- peptide, Phe-Xaa-Pro-Arg-Leu-NH2 ( 16). The high amino acid sequence homology comprising the C- terminal residues of the pyrokinins may dictate the conformational preference of the peptides. For example, the solution structure of the cyclic pyrokinin analog, cyclo-(Asn-Thr-Ser-Phe-Thr-Pro-Arg-Leu-), indicates the presence of a p-turn encompassing the residues Thr-Pro-Arg-Leu (3). A recent NMR study by Wang et al. (17) identified the solution conforma- tion of the hexapeptide analog of the C-terminal 36 1
-
Upload
barbara-a-clark -
Category
Documents
-
view
218 -
download
5
Transcript of Evidence for a C-terminal turn in PBAN An NMR and distance geometry study

Int. J . Peptide Protein Rex 47, 1996, 361-368 Printed in UK ~~ ull rights reserved INTERNATIONAL JOURNAL OF
PEPTIDE & PROTEIN RESEARCH iSSN U367-6.377
Evidence for a C-terminal turn in PBAN An NMR and distance geometry study
BARBARA A. CLARK and GLENN D. PRESTWICH
Department of Chemistry, University at Stony Brook, Stony Brook, New York, USA
Received 11 July, revised 27 September, accepted for publication 8 December 1995
The solution conformation of the pheromone biosynthesis activating neuropeptide (PBAN ) of the moth Helicoverpu zea has been determined using homonuclear two-dimensional nuclear magnetic resonance techniques and distance geometry-restrained energy minimization. The insect peptide hormone showed a random distribution of conformers in aqueous solution, whereas in a less polar medium of trifluoroethanol and water, a reordering process was observed. In particular, the C-terminal region (Phe-Ser-Pro-Arg- Leu-NH,) adopts a type I’ /I-turn conformation, residues 20-27 are in a helix conformation, and residues 1-19 exhibit a high degree of flexibility. Direct observation of the C-terminal /I-turn configuration of PBAN is consistent with a previous report that showed a rigid, cyclic analog of the C-terminal pentapeptide of PBAN retained pheromonotropic activity [ Nachman, R.J., Kuniyoshi, H., Roberts, V.A., Holman, G.M. & Suzuki, A. (1993). Biochem. Biophys. Res. Compnun. 661-6661, Furthermore, our results show no interaction between the C-terminal turn and the rest of the polypeptide chain, thus providing further evidence that the C-terminal turn is indeed the important conformation recognized by the PBAN receptor. 0 Munksgaard 1996.
Key words: Helicovrrpu zeu; NMR spectroscopy; peptide hormone; sex pheromone
The pyrokinins are a group of insect neuropeptides that share the C-terminal pentapeptide sequence, Phe- Xaa-Pro-Arg-Leu-NH,, where Xaa can be Thr, Ser, Gly or Val (1). The first member of the pyrokinin family to be isolated and characterized was leucopyro- kinin (Lem-PK) of the cockroach Leucophaea mad- erae, and the function of Lem-PK is to stimulate contractions in the hindgut and oviduct (2, 3). Like Lem-PK, the five pyrokinins of the locust Locusta migmtoria, labeled locustaniyotropins ( Lom-PK, Lorn-MT-I, -11, -111 and -1V) are involved in myotropic activities (4-7). Also sharing the C-ter- minal sequence of the pyrokinins are the diapause hormone of the silkworm moth Bombyx mori ( Born-DH) (S), and the pheromone biosynthesis activating neuropeptides (PBANs) of the corn ear- worm moth Helicoverpa :ea (Hez-PBAN) (9-1 l),
Abbreviations: 2D, two-dimensional; COSY, homonuclear correla- tion spectroscopy; DG, distance geometry; DQF, double quantum filter; Hez-PBAN, pheromone biosynthesis activating neuropeptide of the moth Hrlicovrrpu zru; NMR, nuclear magnetic resonance spectroscopy; NOE, nuclear Overhauser enhancement; NOESY, NOE correlation spectroscopy; REM, restrained energy minimiza- tion; RMSD, root mean squared deviation; TFE, 2,2,2-trifluoro- ethanol; TOCSY, total correlation spectroscopy.
the silkworm moth Bombyx mori (Bom-PBAN) (12), and the gypsy moth Lymantriu dispar (Lyd-PBAN) ( 13). Interestingly, there is a biological cross-activity among the pyrokinin family of peptides. Hez-PBAN demonstrates myotropic activity in the cockroach, displaying 25% of Lem-PK activity (1). In addition, Lem-PK, Lom-PK, Lom-MT-I and Lom-MT-11 all show pheromonotropic activity in the silkworm moth (14, 15).
As shown in Table 1, the similarity in the C- terminal amino acid sequence is characteristic of the pyrokinin family. The importance of this region was determined when structure-activity studies revealed the minimum sequence required for myotropic and pheromonotropic activities was the C-terminal penta- peptide, Phe-Xaa-Pro-Arg-Leu-NH2 ( 16). The high amino acid sequence homology comprising the C- terminal residues of the pyrokinins may dictate the conformational preference of the peptides. For example, the solution structure of the cyclic pyrokinin analog, cyclo-(Asn-Thr-Ser-Phe-Thr-Pro-Arg-Leu-), indicates the presence of a p-turn encompassing the residues Thr-Pro-Arg-Leu (3). A recent NMR study by Wang et al. (17) identified the solution conforma- tion of the hexapeptide analog of the C-terminal
36 1

B.A. Clark and G.D. Prestwich
Peptide Amino-acid sequence
Hez-PBAN Born-PBAN Lyd-PBAN Born-DH Lorn-PK Lem-PK Lorn-MT-I Loin-MT-TI Lorn-MT-111 Lorn-MT-IV
L S D D M P A T P A D Q E M Y R Q D P E Q I D S R T K Y F S P R L N H , L S E D M P A r P A D Q E M Y Q P D P E E M Q S R T R Y F S P R L N H , L A D D M P A T M A D Q E V Y R P E P E Q I D S R N K Y F S P R L N H ,
T D M K D E S D R G A H S E R G A L C F G P R L - N H , E D S G D G W P Q Q P F V P RL-NHZ
E T S F T P RL-NH, G A V P A A Q F S P R L -NHz
E G D F T P R L -NH, R Q Q P F V P R L N H ,
R L H Q N G M P F S P R L N H ,
Abbreviations: Born. Boi i ih , . .~ i i i o r i : Hez. Helicowrptr ~ 1 ~ 1 : Lem. Lt~rri~o[~lrue~r r~rtrcfi~rrre: Loin. Locitstcr r?iigrtrtoriu, Lyd, Lyrnuiifriu dispur: DH. diapause hormone: MT. myotropin: PK. pyrokinin
sequence of PBAN. Tyr-D-Phe-Ser-Pro-Arg-Leu- NH,, as a type I1 [)-turn ( 17). Another example of a 0-turn conformation found in an insect peptide hormone, is the adipokinetic hormone red pigment- concentrating hormone ( AKHIRPCH ) family of pep- tides. This family of peptides is found in all orders of insects and is involved in influencing energy metabolism. The AKH from praying mantis is the octapeptide Glu-Val-Asn-Phe-Thr-Pro-Asn-Trp. and the C-terminal sequence, Thr-Pro-Asn-Trp. is in a /I- turn configuration ( 18). Although these reported structures of either analogs or fragments of insect peptide hormones have determined turn conforma- tions, the interaction between the C-terminal turn and the rest of the peptide molecule has not been established. Consider the three PBAN sequences: each has 33 amino acid residues, and each shares the identical C-terminal pentapeptide sequence, Phe-Ser- Pro-Arg-Leu-NH,, that structure-activity relation- ships reveal is required for pheromone production. Although the minimum sequence required to interact with both the binding and activation regions of the pheromonotropic receptor is the C-terminal pcnta- peptide ( 16), there may exist a cooperative interaction among the N-terminal and central regions of the peptide molecule and the active C-terminus.
We now report the first solution structure for the complete insect neuropeptide Hez-PBAN in a mixed solvent of 30% 2.2,2-trifluoroethanoI ( TFE ) and water. The conformational flexibility exhibited by small peptides in aqueous solution often hinders their analysis by NMR spectroscopy. The existence of compounds that promote secondary structure in poly- peptides has long been known. I n particular. tri- fluoroethanol (TFE) is a weakly polar alcohol that has been shown to promote secondary structures for peptides in solutions from 30 to 50% in water ( 19---22). The sequence specific proton resonance assignment for Hez-PBAN was determined. The interproton dis- tance constraints were estimated from the observed
362
nuclear Overhauser enhancements (NOES), and the experimental distance constraints were used as input for the distance geometry (DG) calculations. The peptide structures were further refined by restrained energy minimization (REM).
EXPERIMENTAL PROCEDURES Strriiplr pi.epuratioiz. Hez-PBAN was synthesized by standard solid-phase techniques at the Center for Analysis and Synthesis of Macromolecules (CASM, Stony Brook). The peptide was purified using a Vydak, 10 mni x 250 mm C-8 reversed-phase HPLC column, with a linear elution gradient of 20-60Y0 of a 95% acetonitrile 0.1% trifluoroacetic acid solution. This allowed removal of the methionine-oxidized forms that develop in PBAN samples upon storage. The NMR sample was prepared by dissolving ca. 5 mg of purified peptide in 0.45 mL of a 30% 2.2.2-triRuoroethanoI-d, 70% H,O (v/v) solution con- taining 20 mM phosphate. The sample pH was 5.7. An aliquot of the NMR sample was tested via HPLC prior to, and following, all NMR experiments to ensure no degradation of the sample had occurred. The NMR sample, stored at 4 "C, was stable for ca. two months.
Dtrru ucquisition und processing. All spectra were measured on a Bruker AMX-600 spectrometer equipped with a temperature control unit. Data pro- cessing was performed on a Silicon Graphics com- puter using the program FELIX (Hare Research Inc., Woodinville, WA).
Spin system identification, sequential resonance assignment, and experimental distance constraints were determined using the homonuclear two- dimensional (2D) NMR techniques; DQF-COSY, TOCSY and NOESY. Data acquisition wa5 in the phase-sensitive mode using the time-proportional phase increment (TPPI) method (23). The probe

NMR of PBAN
descents minimization was carried out within the Discover module to ensure a regular covalent geo- metry. The DG calculations were carried out using the program DGII (28, 29) within the NMR-refine module. Following triangle bounds smoothing, random distance matrices that satisfied the triangle inequality limits were generated using metrization. Coordinates satisfying the trial distances were obtained by embedding and majorization in four dimensions. The structures were optimized by simu- lated annealing in four dimensions and energy minim- ization in three dimensions.
temperature was set to 15 "C. The spectral width was 5434 Hz. Suppression of the water proton resonance was achieved by a low-power (55 dB) presaturation pulse. Each 2D-NMR experiment consisted of 4096 complex data points in t , and 450 increments in t,. The data were subsequently zero-filled to a final matrix size of 2048 x 2048 real data points. The DQF- COSY experiment (24) was collected with 32 transi- ents per free induction decay (FID). Prior to Fourier transformation (FT), a sine-bell apodization was applied in both dimensions. The TOCSY experiment (25) was collected with an MLEV-17 pulse sequence (26) with 32 transients per FID and a mixing time of 70 ms. The spin-lock field was 9.1 kHz. A sine- bell apodization in the first dimension, followed by a sine-squared apodization in the second dimension, was applied prior to FT. The NOESY experiments (27) were collected with 64 transients per FID at mixing times of 100, 200 and 300 ms. A sine-squared apodization was applied in both dimensions prior to FT.
Distance constraints. Spin diffusion effects were inspected by following the build-up of NOESY crosspeaks when mixing times were increased froin 100 to 300 ms. A mixing time of 200 ms was used to derive proton-proton distance constraints, a suitable compromise between spin diffusion effects and cros- speak intensity. The interproton distance constraints were estimated from the NOESY crosspeak volumes using the integration routine provided in FELIX. The distance constraint ri j between two protons i and j was estimated from the crosspeak volume vij of the NOESY spectrum acquired with a mixing time t , of 200 ms using the equation r i j = rkl(vkJvij)''6, where k and 1 are th,e /l and hydrogen atoms of Prol9, and r k l was 1.7 A. Since the method does not account for spin diffusion and internal motion, the calculated distances are approximate, and the constraints were classified into three categories corresppnding to upper distance limits of 2.7, 3.5 and 5.0A. Those peaks appearing only in the NOESY spectrum acquired at a mixing time of 300 ms were atsigned an upper bound distance constraint of 5.0 A. The following pseudoatom corrections wer? added to interproton distances where necessary; 1 A for methylpe, methyl and amino group hydrogen atoms; 2 A foro two equivalent aromatic hydrogen atoms; and 2.4 A for four equivalent aromatic hydrogen atoms. No dihed- ral angle constraints or stereospecific assignments were used in the structure calculations.
Distance geometry calculations. All DG calculations were carried out within the Insight 11 package (Biosym Technologies, Inc., San Diego, CA). A linear extended chain conformation of the peptide was generated within the Biopolymer module. Prior to the DG calculations, an initial unrestrained steepest
Structure rejinement calculations. REM calculations were carried out on the best DG structures (those with the lowest total constraint violations) within the Discover module. The all-atoem AMBER force field (30, 31) was used, with a 15 A cutoff for nonbonded interactions and a distance-dependent dielectric con- stant ( E = 4r) to compensate for lack of explicit solv- ent. The best DG structures were minimized using 1500 steps of steepest descents minimization, followed by a conjugate gradient minimization to a maximum gradient of 0.01 kcal mol-' A-' . The NOE con- straints were applied using a flat-bottomed square- well potential function with a force constant of 32 kcal A-' throughout.
RESULTS AND DISCUSSION
Data acquisition A preliminary 2D-NMR spectrum of the peptide was acquired in an aqueous solution at 25 "C. There were minimal NOESY crosspeaks observed, showing that the peptide exhibits conformational flexibility under these conditions. This conformational flexibility can be reduced by stabilizing a population of the peptide molecules that exhibit secondary structure. Literature reports have shown that solutions of up to 50% TFE in water promote secondary structure in peptides and proteins (19-22). Aliquots of TFE were added to the peptide solution and 1D-NMR spectra were recorded. At a 30% TFE concentration, no significant changes in linewidth were observed in the spectra. Preliminary CD studies of PBAN with 0, 15, 30 and 50% TFE further support this conclusion; however, at 50% TFE a significant increase in cc-helicity may be occur- ring (J.T. Elliott, unpublished results). At this point, a NOESY spectrum was recorded and secondary structural stabilization was apparent.
Spectral assignmens The DQF-COSY and TOCSY spectra were used to identify the spin systems of the amino acid side chains by their characteristic coupling patterns (32). In summary, the isoleucine and lysine residues were assigned from the spectra due to their uniqueness. The serines were distinguished from the other AMX
363

B.A. Clark and G.D. Prestwich
spin systems by the characteristic low field chemical shift of their 8-protons. The AMX spin systems from the aromatic residues were identified based on the NOESY coupling between the aromatic protons and the 3- or 8-protons. The remaining AMX spin systems were nonsequentially assigned to the aspartic acid residues. The alanine and threonine residues &ere readily distinguished from the two leucine residues. The three arginines, as well as the four prolines. were heavily overlapped making the assignments dificult. The methionine, glutamate and glutamine residues were identified in the sequential assignment. The amide proton resonances were assigned on the basis of the COSY connectivities between the C,H-NH proton resonances of each residue.
The sequential resonance assignment was deter- mined using the NOESY spectra (32) . The sequential assignment was based on the observed NOESY coup- ling between C,,H-N,, + ,H and C,,H-N,, I ,H. In the case of the proline residues where there is no amide proton, the crosspeak between the C,H of the preced- ing residue and the C,H of the proline residue was used to establish sequential connectivity. The finger- print region of the NOESY spectrum with the con- nectivities outlined is shown in Fig. 1 . The sequence-
FIGURE 1
specific chemical shift assignments for all of the proton resonances are given in Table 2.
E\~irk.ric.e ,for striic'turr The experimental distance constraints are derived mostly from the sequential and medium-range NOE connectivities. The observation of strong C,,H-N,, ,H NOE connectivities (Fig. 1 ) is indicat- ive of an extended chain or random coil conforma- tional ensemble of the peptide. As shown in Fig. 2, a number of NOE crosspeaks are observed between the amide proton resonances of sequentially adjacent amino acid residues. These N,H-N,+,H NOE con- nectivities, encompassing residues 2-8, 14-24 and 27-33. are indicative of folded structures in the peptide. Also shown in Fig. 2, are two N,H-Ni+,H NOE connectivities, between residues 22-25 and 24-27. which are characteristic of a helix conforma- tion. In addition, we observe three CBH-Ni+2H NOE crosspeaks between residues 1-3, 8-10 and 10-12, confirming that local structures with short-range order exist in the N-terminal region of the peptide.
There are four proline residues in Hez-PBAN. Categorizing a peptide bond in the cis configuration was based on the observed NOE connectivity between
0
s2
0 122
h M5 8 . 4 e. 2 e. o 7. e 7.6
D1 ( p p m )
Fingerprint region of the NOESY (100 ms) spectrum of Hez-PBAN in 30','t3 TFE water at 15 'C, pH 5.7. The sequential crosspeaks are connected to the intraresidue crosspeaks by solid lines.
364

N M R of PBAN
TABLE 2 Proton chemical shifrs of Hez-PBAN
Chemical shift (ppm)”
Residue N H CrH CBH Others
Leu- I 8.55 3.96 1.68*b yCH 1.61 6CH3 0.93* XCH, 0.89* Ser-2 8.47 4.43 3.78* Asp-3 8.37 4.61 2.79* Asp-4 8.17 4.61 2.77, 2.72 Met-5 7.82 4.69 1.98, 1.88 yCH, 2.51, 2.45 Pro-6 - 4.19 2.16, 1.85 ;CH, 1.91* K H , 3.69* Ala-7 8.06 4.32 1.30, Thr-8 7.79 4.58 4.30 yCH, 1.19* Pro-9 - 4.21 2.26, 1.89 yCH, 2.02* K H , 3.75* Ala-10 7.85 4.07 1.31* Asp- 1 1 7.84 4.46 2.96, 2.80 Gln-12 7.98 4.03 2.09* yCH, 2.32* 6NH, 7.16, 6.58 Glu- 13 8.00 4.07 2.11, 2.01 yCH, 2.32* Met-14 7.84 4.12 2.04, 1.99 yCH, 2.29* Tyr- 1 5 7.71 4.32 3.04, 2.90 Hz,6 6.96 H,,5 6.66 Arg- 16 7.63 4.08 1.81, 1.74 yCH, 1.55* 6CH, 3.06* NH 7.03 Gln-17 7.84 4.21 2.03, 1.92 yCH, 2.29* 6NH2 7.23, 6.55 Asp- 18 8.08 4.21 2.90, 2.75 Pro-19 - 4.30 2.14, 1.84 yCH, 1.89* 6CHz 3.69* Glu-20 7.98 4.07 2.03* yCH, 2.40* (3111-21 7.85 4.1 1 1.94, 1.89 jCH, 2.29* dNH, 7.21, 6.80
ASP-23 8.31 4.50 2.80* Ser-24 7.96 4.19 3.91, 3.84 Arg-25 7.96 4.21 1.93, 1.86 yCH, 1.72, 1.64 yCH, 3.09* NH 7.13 Thr-26 7.78 4.21 4.15 yCH3 1.15* Lys-27 7.78 4.05 1.61 yCH, 1.14* 6CH, 1.51* ECH, 2.81* NH, 7.44* Tyr-28 7.64 4.32 2.87, 2.73 H2,6 6.81 H3,5 6.62 Phe-29 7.70 4.46 3.02, 2.92 H2,6 7.11 H3,4,j 7.22 Ser-30 7.58 4.55 3.78, 3.70 Pro-3 1 - 4.27 2.20, 1.81 yCH, 1.90* 6CH2 3.57* Arg-32 7.88 4.18 1.78, 1.66 yCH, 1.54* 6CH2 3.09* NH 7.10 Leu-33 7.73 4.21 1.52* yCH 1.43 X H , 0.84* S’CH, 0.79*
a Chemical shifts are expressed relative to the water proton resonance at 4.80 ppm. Asterisks indicate degenerate proton chemical shifts.
Ile-22 7.70 3.84 I .80 yCH3 0.84* jCH2 1.45, 1.11 6CH, 0.76*
the C,H of the residue preceding the proline residue and the C6H of the proline residue. The peptide bonds for proline residues 6, 9 and 31 were deter- mined to be in the cis configuration (Fig. 3). The configuration of Pro19 is ambiguous, since neither an inter-residue H,-H, NOE nor an inter-residue He-H, NOE was observed.
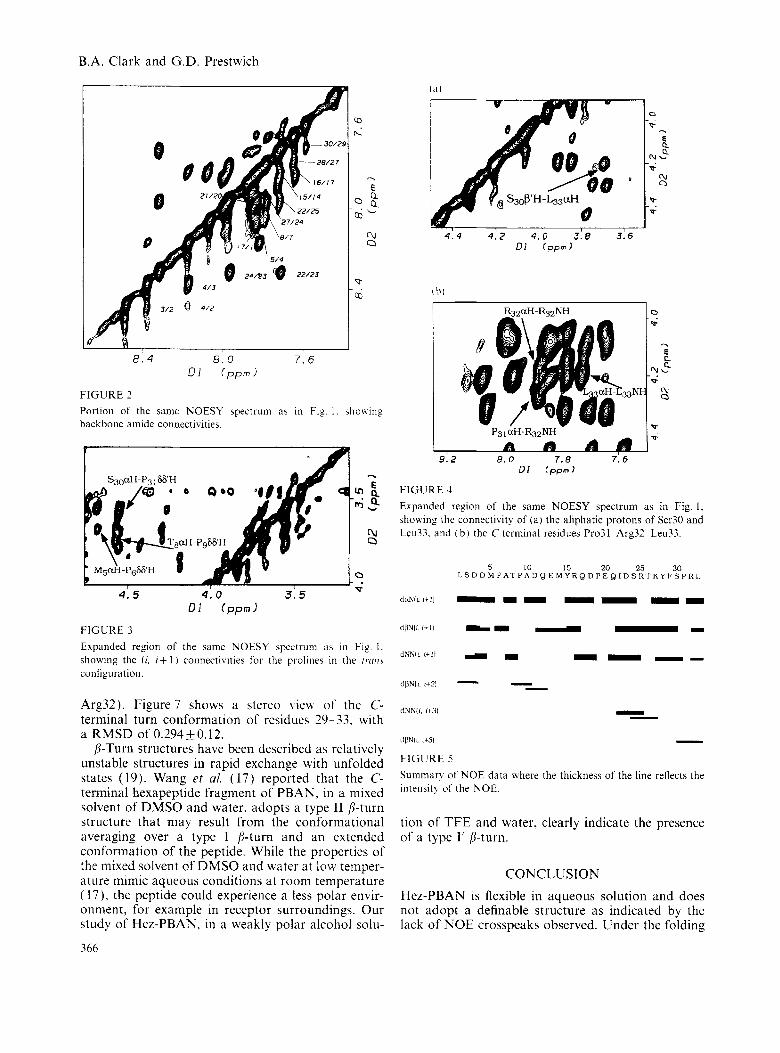
Evidence for a C-terminal turn is based on the observed C,H-C,, + 3H NOE connectivity between residues 30-33 (Fig. 4a), the C,H-C,,+,H NOE con- nectivity between residues 31-33 (data not shown), and the sequential C,H-N, + ,H NOE connectivities between residues 31-32-33 (Fig. 4b). A summary of the NOE data is given in Fig. 5, where the thickness of the line reflects the intensity of the NOE. A total of 223 NOE crosspeaks were assigned consisting of 137 intraresidue, 64 sequential and 22 medium-range connectivities.
Solution conformation DG calculations were used to generate 100 structures using a set of 223 NOE-derived distance constraints. From the 100 structures generated, the 25 structures with the lowest DGII error functions (<O.l) were chosen for further analysis. Figure 6 shows the super- position of the 25 best DGII/REM structures of Hez- PBAN aligned using residues 29-33. Although the N-terminal region shows several medium-range NOE connectivities, indicative of local structures with short range order, too few NOE-derived distance con- straints were determined to define any convergent set of coordinates. The C-terminal region exhibits a high degree of structural stabilization with the C-terminal pentapeptide sequence, Phe-Ser-Pro-Arg-Leu-NH,, adopting a type I’ p-turn conformation based on the dihedral angles of the central residues (4 = 55 k 4 and p=18+2forPro31, a n d 4 = 8 5 f 4 a n d p = l O f l for
365
B.A. Clark and G.D. Prestwich
‘15/14
22/25 24
17
5 / 4
2 4 0 3 8 22/23
FIGURE 2
lo r.
- E a ?,a
Q,
cu 4:4 4 : Z 4.0 3:8 3:6 Q DI Ippml
P ai cb1
Portion of the same NOESY spectrum as in Fig. I . bhotviiig backbone aniide connectivities.
I
4: 5 4: 0 3: 5 01 (pprnl
FIGURE 3 Expanded region of the same NOESY spectrum as in Fig. 1. showing the (i, i+ 1 ) connectivities for the prolines in the trmi.\ configuration.
Arg32). Figure 7 shows a stereo \iew of the C- terminal turn conformation of residues 29-33. with a RMSD of 0.294+0.12.
b-Turn structures have been described as relatively unstable structures in rapid exchange with unfolded states (19). Wang et ul. ( 1 7 ) reported that the C- terminal hexapeptide fragment of PBAN. in a mixed solvent of DMSO and water. adopts a type I1 p-turn structure that may result from the conformational averaging over a type I b-turn and an extended conformation of the peptide. While the properties of the mixed sohent of DMSO and water at IOK temper- ature mimic aqueous conditions at room temperature (17) . the peptide could experience a less polar envir- onment, for example in receptor surroundings. Our study of Hez-PBAN. in a weakly polar alcohol solu-
366
a 6 , A a 8.2 8.0 7.8 7. 6
D1 ( p p m )
FIGURE 4 Expanded region of the same NOESY spectrum as i n Fig. 1 , shoxing the connectivity of ( a ) the aliphatic protons of Ser30 and Leu33. and ( b ) the C-terminal residues Pro31-Arg32-Leu33.
5 10 15 L S D D M P A T P A D Q E M Y R Q
20 25 30 D P E Q I D S K T KY F S P R I ,
dBNli rc31 - FIGURE 5 Summary of NOE data where the thickness of the line reflects the intensit!, of the NOE.
tion of TFE and water, clearly indicate the presence of a type I ’ [j-turn.
CONCLUSION Hez-PBAN is flexible in aqueous solution and does not adopt a definable structure as indicated by the lack of NOE crosspeaks observed. Under the folding

NMR of PBAN
N
k -term
C-term 9 FIGURE 6 Superposition of the 25 best DGII/REM structures of Hez-PBAN aligned using residues 29-33.
conditions of the cosolvent TFE, the number of conformational states available to the peptide was reduced. Subsequent to the addition of TFE, the observation of several medium-range NOE connectiv- ities confirmed that local structures with short-range order exist in the peptide. Most notably, the small but measurable local structure of the N-terminal region of the peptide, the helical conformation of residues 20-27, and the type I’ p-turn conformation encompassing the C-terminal pentapeptide (Fig. 6). The /?-turn conformation of the C-terminal region of
Hez-PBAN is consistent with a previous study that showed a rigid, cyclic analog of the C-terminal hexa- peptide induced pheromonotropic activity (33). Also consistent with the study reported by Nachman rt ul. (33) is the existence of a /?-turn in the C-terminal hexapeptide fragment of PBAN that was essentially reported by Wang et ul. (17) The authors reported a type I1 p-turn structure, in a solution of DMSO and water, that may result from the coexistence of a type I p-turn and the extended conformation of the hexa- peptide. In the less polar environment of TFE and water, the stable structure is a type I’ /?-turn. In addition, there appears to be no interaction between the C-terminal turn and the rest of the peptide molecule, indicating the C-terminal turn is indeed the important conformation recognized by the PBAN receptor. Complete structural characterization of other insect neuropeptides, their carriers, and their receptors in target tissues are necessary prerequisites for the rational design of peptidomimetics as modi- fiers of insect behavior and development ( 1 ).
ACKNOWLEDGMENTS
We are grateful to Dr. G. Veglia (University of Rome) for his assistance with the distance geometry and energy minimization calculations, and to Mr. J. T. Elliott for permission to cite his CD results. We thank Dr. Veglia, Dr. J. F. Marecek, Mr. Elliott and Mr. M. R. Ziebell for helpful advice.
This work was supported by NIH grant A132498 to Dr. W.L. Roelofs (Cornell University) (through a subcontract to co-investigator G.D.P.) and by a Herman Frasch Foundation award to G.D.P. and Dr. S. McN. Sieburth (Stony Brook). The Bruker AMX-600 was acquired with instrument grants from NIH (RR05547A) and NSF (CHE8911350), with additional support from the University at Stony Brook and the Center for Biotechnology, a New York State Science and Technology Foundation Center for Advanced Technology.
REFERENCES
1. Nachman, R.J., Holman, G.M. & Haddon. W.F. ( 1993) Arch. Insect Biochem. Physiol. 22, 181-197
FIGLJRE 7 Stereo diagram of the backbone atoms of the C-terminal hexapeptide of Hez-PBAN
367

B.A. Clark and G.D. Prestwich
2. Nachman, R.J.. Holman. G.M. & Cook. B.J. ( 1986 1 Biocirent. BiophJx Res. Coniniun. 137. 936-942
3. Nachman. R.J.. Roberts. V.A.. Dysan. H.J.. Holman. G.M. & Tainer. J.A. (1991) Proc. ,Vtrrl. Act id Sci. 1-S.4 88. 4518-4522
4. Schoofs. L.. Holman. G.M.. Hayes. T.K.. Tips. A,. Nachman. R.J.. Vandesande. F. & DeLoof. A. (1990) Pepride.\ 11. 427-433
5. Schoofs. L., Holman. G.M.. Hayes. T.K.. Nachman. R.J. & DeLoof. A. (1990) fnsecr Biochent. 20. 479-484
6. Schoofs. L.. Holman. G.M. . Hayes. T.K.. Nachman. R J . & DeLoof. A. (1991) Gen. Cornp. Endocrinol. 81. 97-103
7. Schoofs. L., Holman, G.M.. Hayes. T.K.. Nachnian. R.J.. Kochansky, J.P. & DcLoof, A. ( 1992) hisect Biochrrn .\folec. Biol. 22. 447452
8. Imai. K.. Konno. T.. Nakazawa. Y.. Komiya. T.. Isobe. M.. Koga. K.. Goto. T.. Yaginuma. T.. Sakakibara. K.. Hasegatva. K. & Yamashita. 0. (1991 ) Proc. Jtrptrn A c , t r d 67B. 98-101
10. Raina. A.K.. Jaffe. H.. Kempe. T.G.. Keim. P.. Blacher. R.W.. Fales. H.M.. Riley. C.T.. Klun. J.A.. Ridgway. R.L. & H q e s . D. K. ( 1989) Science 244. 796-798
11. Raina. A.K. & Kempe. T.G. ( 1992) fnsecr Biochcwt. . tfo/cc. Biol. 22. 221--225
12. Kitamura. A,. Nagasawa. H.. Kataoka. H.. Inoue. T.. h d o . T. & Suzuki. A. ( 1989) Biochent. Biophi~s. Rcs. Connnirn. 163. 520-526
13. Masler. E.P.. Raina. A.K.. Wagner. R .M. & Kochansk?. J.P. ( 1994) Insecr Biocheitt. hfolcc. Biol. 24. 829 836
14. Kuniyoshi, H.. Nagasawa. H.. Ando. T.. Nachman. R.J & Holman, G.M. (1992) Biosci. Biorrch Biochern. 56. 167
15. Fonagy. A,. Schoofs. L., Matsumoto. S.. DeLoof. A . & Mitsui. T. (1992) J. Insect Phxsiol. 38. 651-657
16. Raina. A.K. & Kempe. T .G. (1990) I m w t Biochznr. 20. 849-85 1
17. Wang. Y.S.. Kempe. T.G.. Raina. A.K. & Mazzocchi. P.H. ( 1994) Iiit. J . Peptide Prorein Res. 43. 277-283
18. Zubrzycki. I.Z. & GSde. G . ( 1993) Biochent. Bioph~.,s, Rex Commun. 198. 228-235
19. Dyson. H.J.. Rance. M.. Houghton. R.A.. Wright. P.E. & Lerner. R.A. (1988) J. Mol. Biol. 201. 201 -217
9. Raina. A.K. & Klun. J.A. (1984) Scirrice 225. 531-533
20. Dyson. H.J.. Merutka. G.. Waltho, J.P., Lerner, R.A. &
21. Dyson. H.J.. Sayre. J.. Merutka, G. , Shin, H.-C.. Lerner, R.A.
22. Buck. M., Radford, S.E. & Dobson, C.M. (1993) Bioc.hemi.stry
23. Redfield. A.G. & Kunz. S.D. (1975) J. Mrrgn. Reson. 19. 250 254
24. Rance. M.. Snrensen, O.W., Bodenhausen, G., Wagner, G., Ernst. R.R. & Wiithrich, K. ( 1983) Bioche~?. Biophys. Res.
25. Braunschweiler. L. & Ernst, R.R. (1983) J. Mugn. Reson.
26. Bax. A. & Davis. D.G. (1985) J. Mugn. Reson. 65, 355-360 27. Jeener. J.. Meier. B.H., Bachmann, P. & Ernst, R.R. (1979)
J. Chetu. Phjx 71. 4546-4553 28. Havel. T.F. (1990) Biopo/jwiers 29, 1565-1585 29. Havel. T.F. (1991 ) Pmg. Biophys. M d Bid. 56, 43-78 30. Weiner. S.J.. Kollman, P.A., Case, D.A., Singh, U.C.. Ghio.
C. Alagona. G.. Profeta, S. & Weiner, S. (1984) J. Am. Cheni.
31. Weiner. S.J.. Kollman. P.A., Nguyen. D.T. & Case, D.A.
32. Wiithrich. K . ( 1986) N M R qfProteins and Nucleic Acids, John Wile?. New York
33. Nachman. R.J.. Kuniyoshi, H., Roberts. V.A., Holman, G.M. & Suzuki. A. ( 1993) Biochent. Biophys. RFS. Comnnn. 193,
Wright. P.E. (1992) J. Mol. Bid. 226, 795-817
& Wright. P.E. (1992) J. Mol. Biol. 226, 819-835
32. 669-678
C ~ t t i t t i ~ ~ i . 117.479-485
53. 521-528
SOC. 106. 765-784
( 1986) J. ConlpLrt. C/I<WI. 7. 230-252
66 I-66h
Address:
Professor Glenn D. Presiwich Department of Chemistry University at Stony Brook Stony Brook. New York 11794--3400 LISA
Phone: 5 I 6 632-7926 Fas: 5 16 632~- 7962 E-mail: [email protected]
368


















