
Evaluating a marine protected area in a developing country; Mafia Island Marine Park, Tanzania -...
17
Ocean & Coastal Management 47 (2004) 321–337 Evaluating a marine protected area in a developing country: Mafia Island Marine Park, Tanzania Albogast T. Kamukuru a, , Yunus D. Mgaya a , Marcus C. O ¨ hman b a Faculty of Aquatic Sciences and Technology, University of Dar es Salaam, P.O. Box 60091, Dar es Salaam, Tanzania b Department of Zoology, Stockholm University, 10691 Stockholm, Sweden Abstract The benefits of marine protected areas (MPAs) to fish productivity remain debated, and comprehensive research projects have been suggested to assess MPA function. This study evaluated MPA performance in a developing country in the context of local needs. We compared density and size of the blackspot snapper, Lutjanus fulviflamma (Forsska˚l 1775), in Mafia Island Marine Park (MIMP), Tanzania, with adjacent intensively fished areas (IFA) using underwater visual censuses (a total of 105, 50-m transects) as well as investigating the catches in the local fishery. The target species was over four times more numerous, its biomass six to ten times higher and individual sizes on an average 37% larger on reefs in MIMP compared to the IFA. Fish numbers and biomass were negatively correlated with fishing intensity and positively correlated with hard coral cover and structural complexity. This study supports predictions that MPAs can play a key role in the conservation of habitats and management of a fishery. It is suggested that for the purpose of management, it is possible to evaluate MPA performance with limited resources by focusing on key information. r 2004 Elsevier Ltd. All rights reserved. ARTICLE IN PRESS www.elsevier.com/locate/ocecoaman 0964-5691/$ - see front matter r 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.ocecoaman.2004.07.003 Corresponding author. Tel.: +468 164 022; fax: +468 167 715. E-mail address: [email protected] (M.C. O ¨ hman).
Transcript of Evaluating a marine protected area in a developing country; Mafia Island Marine Park, Tanzania -...

Ocean & Coastal Management 47 (2004) 321–337
Evaluating a marine protected area in adeveloping country: Mafia Island Marine Park,
Tanzania
Albogast T. Kamukurua,!, Yunus D. Mgayaa,Marcus C. Ohmanb
aFaculty of Aquatic Sciences and Technology, University of Dar es Salaam, P.O. Box 60091,Dar es Salaam, Tanzania
bDepartment of Zoology, Stockholm University, 10691 Stockholm, Sweden
Abstract
The benefits of marine protected areas (MPAs) to fish productivity remain debated, andcomprehensive research projects have been suggested to assess MPA function. This studyevaluated MPA performance in a developing country in the context of local needs. Wecompared density and size of the blackspot snapper, Lutjanus fulviflamma (Forsskal 1775), inMafia Island Marine Park (MIMP), Tanzania, with adjacent intensively fished areas (IFA)using underwater visual censuses (a total of 105, 50-m transects) as well as investigating thecatches in the local fishery. The target species was over four times more numerous, its biomasssix to ten times higher and individual sizes on an average 37% larger on reefs in MIMPcompared to the IFA. Fish numbers and biomass were negatively correlated with fishingintensity and positively correlated with hard coral cover and structural complexity. This studysupports predictions that MPAs can play a key role in the conservation of habitats andmanagement of a fishery. It is suggested that for the purpose of management, it is possible toevaluate MPA performance with limited resources by focusing on key information.r 2004 Elsevier Ltd. All rights reserved.
ARTICLE IN PRESS
www.elsevier.com/locate/ocecoaman
0964-5691/$ - see front matter r 2004 Elsevier Ltd. All rights reserved.doi:10.1016/j.ocecoaman.2004.07.003
!Corresponding author. Tel.: +468 164 022; fax: +468 167 715.E-mail address: [email protected] (M.C. Ohman).

1. Introduction
As coral reefs are important resources that provide humans with food and othergoods and services [1–4], the significance of protecting them is broadly recognised[5–7]. A widely accepted management strategy in the conservation of reef-fish speciesis the implementation of marine protected areas (MPAs) designated to protect fishstocks and habitats [8–20]. The expectation is that the spatial closure will act as arefuge for local reef fish communities, enhancing their densities and diversity. Thiscould then have a positive effect on fishery resources in the surrounding waters asadult fish may migrate (spillover effect) and/or fish larvae might disperse beyond thepark [8,21–25].
How MPAs perform in terms of protecting fish stocks has been the focus of manystudies [6,26–35]. A common conclusion is that there is an increase in fish numbers inMPAs compared to adjacent areas and/or compared to the situation before theMPAs were established [36]. However, evidence that MPAs enhance fisheries in theadjacent intensively fished areas is sparse [37]. In a paper assessing the effectivenessof MPAs as tools for fishery management, Russ [11] concludes that we need moreinformation on how well they work, because many studies have had problems intheir research design. He suggests that a study on the effects of MPAs on fishcommunities should include a before-after-control-impact-pairs design (BACIP)with replicate sampling units in replicate sites duplicated regionally, and that theinvestigation should be carried out over 5–20 years. In addition, habitat effectsshould be factored out, fishing mortality measured, a capture–recapture studyconducted and large transects that traverse both controls and reserves taken. Withsuch a design we would attain a broad scientific understanding on how successfulMPAs are and general principles on marine conservation could be achieved.
On the contrary, as most MPAs are situated in developing countries mainlyprotecting coral reefs, such large-scale studies that consider many aspects of marineprotection are not carried out due to limited resources. In addition, all thatinformation is usually not needed for the purpose of successful resource managementat a local scale in an area dominated by an artisanal fishery.
Along the coast of Africa lie some of the world’s poorest countries. In East Africa,where most coral reefs are situated, there is a population growth rate of 4–7% perannum [38]. In this area the information needed about MPA performance is lessacademic and more about the subsistence of local fishermen. Most studies on MPAfunction, so far, have mainly been carried out in Kenya [28,39–43]. Ecologicalstudies on the effects of MPAs in Tanzania and the other countries in the region arelimited (but see [6,44]).
The development of MPAs in Tanzania dates back to 1975 when eight areas weredesignated as no-take marine reserves, but were subjected to continuous fishingpressure, including blast fishing, and other abuses such as shell and coral collection,anchor damage, coral mining and human trampling [45]. Lack of financial, technical,and human resources caused the reserves to be left unmanaged. However, with theMarine Parks and Reserves Act of 1994, the first guide on the institutionalmechanisms for the management of MPAs in Tanzania was provided. To date two
ARTICLE IN PRESS
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337322

functional marine parks and one marine reserve have been established in Tanzania[46]. Unlike previous marine reserves, marine parks in Tanzania are now designedfor multiple usage applying a zoning scheme [47]. Like in adjacent areas wherefishing is usually carried out, extractive resource use is permitted within the park butin a more regulated manner.
In this study, we investigated the effects of an MPA on fishery resources in thecontext of local needs and resource limitations. This was done by focusing on aselected set of data instead of covering all aspects of MPA performance. Theabundance, biomass and size structure of a target fish species in Mafia Island MarinePark and adjacent intensively fished areas were compared. We also collectedinformation on catches as well as considered how the species may be influenced byhabitat structure.
2. Materials and methods
2.1. Study sites
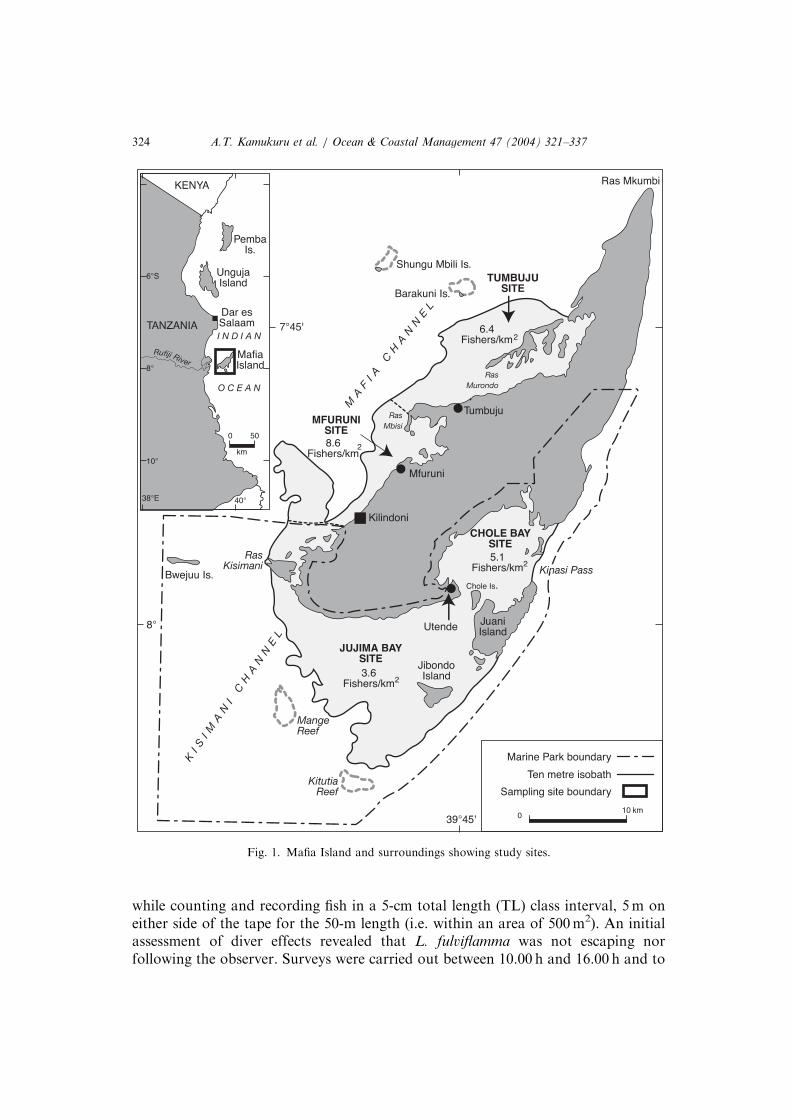
Mafia Island Marine Park (MIMP) is situated south-east of Dar es Salaam inTanzania, about 20 km east of the Rufiji delta (Fig. 1). MIMP contains a variety ofbiotopes that support a large number of species [48]. The park, which covers an areaof about 822 km2, was established in 1995. It is a large, multiple-use area beingoperated on the principles of integrated coastal management [46]. This ensures theneed to balance the protection of the natural resource base while maintaining thelocal communities’ right to the resources. Through boat patrols and localcommunity participation, coral mining and destructive fishing techniques such asbeach seining and blast fishing have been greatly controlled [47]. This surveillance isconducted on a regular basis and illegal fishers are either fined or have their gear andboats confiscated. Although violations still occur, the fishing pressure is considerablylower in MIMP than in intensively fished areas (IFA). This study was conducted onpatchy reefs at two sites in MIMP (Chole Bay and Jujima Bay) and at two sitesoutside the park (Mfuruni and Tumbuju) (see Fig. 1) during September/October2000.
2.2. Ecological data
The blackspot snapper, Lutjanus fulviflamma, is an important commercial reef-fishspecies in Tanzania [49] and elsewhere in the Indo-West Pacific region [50]. Althoughthe fishery at Mafia Island is multispecific, snappers (Lutjanidae) along withemperors (Lethrinidae) are preferred groups that contribute to about 42% of thetotal marine catch [44].
Numbers and size structure of L. fulviflamma were estimated by applying the 50mbelt-transect method in underwater visual censuses (UVC) [51]. Each transect wastaken by randomly laying a stretched fibreglass tape on the reef (2–15m depth). Theobserver swam along the tape at a constant speed (approximately 20m per minute)
ARTICLE IN PRESS
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337 323
while counting and recording fish in a 5-cm total length (TL) class interval, 5m oneither side of the tape for the 50-m length (i.e. within an area of 500m2). An initialassessment of diver effects revealed that L. fulviflamma was not escaping norfollowing the observer. Surveys were carried out between 10.00 h and 16.00 h and to
ARTICLE IN PRESS
KENYA
TANZANIA
PembaIs.
Shungu Mbili Is.
Chole Is.
JuaniIsland
JibondoIsland
Marine Park boundaryTen metre isobath
Sampling site boundary
0 10 km
Bwejuu Is.
Utende
Barakuni Is.
Ras Mkumbi
7°45'
8°
39°45'
RasMbisi
RasMurondo
RasKisimani
MangeReef
KitutiaReef
TUMBUJUSITE
Tumbuju
Mfuruni
Kilindoni
6.4Fishers/km2
MFURUNISITE
CHOLE BAYSITE
JUJIMA BAYSITE
K I S
I M A
N I
C H
A N
N E
L
M A
F I
A C H
A N
N E
L
8.6Fishers/km
5.1Fishers/km2
2
3.6Fishers/km2
MafiaIsland
UngujaIsland
Dar esSalaam
10°
0 50
km
38°E 40°
8°
6°S
I N D I A N
O C E A N
Rufiji River
Kinasi Pass
Fig. 1. Mafia Island and surroundings showing study sites.
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337324

avoid observer bias, all fish counts were conducted by the same person (A.T.K.).Water visibility was greater than 10m at all sites throughout the study period.Biomass was calculated as follows:
Biomass ðgÞ ¼ aLb; where L=mid class TL (cm) of fish with regression coefficientsobtained from Kamukuru [44] ða ¼ 1:165$ 10%5 and b=3.049).
The substrate characteristics were assessed using the line intercept transect (LIT)technique [52]. Substrate categories included live and dead coral, rock, rubble, sand,macroalgae, seagrass, soft coral and sponge. To measure structural complexity, a 50-m tape was laid on the bottom to conform to all irregularities and later pulledhorizontally tight to eliminate all the slack produced by the relief to obtain thehorizontal length. Structural complexity [R] was then calculated according to Grigg[53]
R ¼ 10% 10$H1
50;
where Hl is the horizontal length.
2.3. Fishery data
Fishery data (i.e. number of fishers, fishing boats and gear) were obtained from asurvey conducted by the Mafia Island District Fisheries Office in March 1999 (F.O.Hemile, pers. comm.). Fishing grounds were assumed to extend from the shoreline tothe 10-m isobath based on the type of fishing gear, boats and methods currentlyused. The area of each sampling site was estimated using a digital planimeter. Themeasurements for each sampling area were redigitised five times and the average areawas used in the calculation of the fishing intensity index given as number offishermen per unit area (Table 1).
2.4. Data analysis
The Kolmogorov–Smirnov goodness of fit test was used to test the normality ofdata collected from the underwater visual census. As the data did not conform to
ARTICLE IN PRESS
Table 1Estimation of fishing intensity index of the study sites in shallow waters of Mafia Island. Data is based onMafia District annual fisheries census conducted in March 1999
Study sites
Jujima Bay Chole Bay Tumbuju Mfuruni
Fishing intensity index (fishermen km%2) 3.63 5.13 6.38 8.62Fishing effort (fishermenvessel%1) 3.20 2.61 2.85 6.22Number of fishing vessels 209 142 201 99Number of fishermen 669 370 572 616Fished area by seine nets (km2) 184.28 72.18 89.70 71.44Seine nets stretched mesh size (mm) 50–64 50–64 19–38 19–38
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337 325

normal distribution the Kruskal–Wallis rank test, the Dunn’s multiple comparisonstest and the Mann–Whitney U-test were applied [54,55]. To test for the effects ofhabitat characteristics on fish abundance and biomass, the structural complexity andhard coral cover were regressed against the natural logarithm of fish abundance andbiomass to obtain Pearson correlation coefficients. Because of incidence of zerocounts, the data were transformed (x+1) prior to loge transformation [56].
3. Results
3.1. Fish counts
There were over four times more L. fulviflamma (H=46.359, po0.001, df=3) andtheir biomass was six to ten times higher (H=56.135, po0.001, df=3) within MafiaIsland Marine Park (MIMP) than in the intensively fished areas (IFA). There was anegative relationship between fish abundance/biomass and fishing intensity (Fig. 2).However, densities within a size range of 100–200mm total length (TL) did not differsignificantly between the study sites (H=2.76, p=0.430, df=3). The highestindividual count among the 105 transects was 72 fishes per 500 m2 at Jujima Baycompared to 2 fishes per 500m2 at more than 50% of the transects within the IFA(Fig. 3).
The mean (7 s.e.) visually assessed fish size was 229.071.5mm TL in MIMP incomparison to 167.272.6mm TL in IFA. The length–frequency distribution of fishrecorded on transects was compared with that of fish caught in the fishery within thecensus areas (Fig. 4). A similar trend of length–frequency distributions emerged witha modal class centred at 175 and 225mm TL for fish populations in IFA and MIMP,respectively. There was no significant difference in mean fish sizes assessed visually
ARTICLE IN PRESS
Fig. 2. Mean density and biomass of Lutjanus fulviflamma estimated using underwater visual census(UVC) in four study sites during September/October 2000. Values that are significantly different havedifferent letters for density and biomass following Dunn’s multiple comparisons test (error bars= 7 s.e.).Values in parentheses indicate no. of fishermen/km2 for each site.
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337326

and caught in the fishery (Jujima Bay: U=77058, Z=%0.772, p=0.429; Chole Bay:U=45718, Z=%1.016, p=0.307; Tumbuju: U=21066, Z=%1.497, p=0.134) exceptat Mfuruni (U=20881, Z=3.541, po0.001), with larger sizes recorded visually.
3.2. Habitat characteristics
All habitat characteristics, except sand, differed significantly among study sites(H=3.178, p=0.365, df=3) (See Table 2). Bare rock and rubble were the highestsubstrate categories and accounted for 30–36% and 22–31%, respectively within theIFA. Conversely, hard coral cover dominated in MIMP with a range of 26–28%(Table 2). In terms of structural complexity of reefs Chole Bay had the highestfigures (3.0) and Mfuruni the lowest (1.3) (Table 2). There was a significant increasein fish abundance and biomass with an increase in structural complexity and hardcoral cover (po0.001; Figs. 5 and 6).
4. Discussion
4.1. Fishery effects
This study examined fishery effects and MPA performance at Mafia Island,Tanzania. This was achieved by studying the presence in the field of a target species(blackspot snapper) and its catch composition in the fishery, and by collectingecological data on fish-habitat interactions. The results indicate significantdifferences in abundance, biomass and population structure of the species. Withinthe marine park there were four times more individuals of the species per unit area,the biomass was six to ten times greater and the fishes were larger than in theadjacent heavily fished areas. These observations conform to a number of studiesthat have assessed differences in reef fish communities between MPAs (or lessdisturbed areas) and more exploited locations, e.g. [28,30,33–35,57]. For example,Watson & Ormond [28] reported from Kenya that the abundance of Lutjanusfulviflamma and L. ehrenbergi (Peters, 1869) were 170 times higher in marine parks
ARTICLE IN PRESS
Fig. 3. Density–frequency distributions of Lutjanus fulviflamma visually assessed from shallow waters ofMafia Island during September/October 2000.
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337 327

than in adjacent fished areas. A similar pattern was observed from southernKenya and northern Tanzania where lutjanids were noted to have higherbiomass in protected areas [6]. In terms of size, e.g. Wantiez et al. [35] showedthat protection could increase the total length of the species by almost 30%.This is comparable with our study where the blackspot snappers were 37% larger inMIMP than in IFA. Consequently, our results indicate that there is an overall state
ARTICLE IN PRESS
(a)
(b)
(c)
(d)
Fig. 4. Comparison of length frequency distributions of Lutjanus fulviflamma caught in a fishery andvisually assessed from (a) Tumbujua, (b) Mfurunia, (c) Chole Bayb and (d) Jujima Bayb during September/October 2000. Note: Sites that are significantly different have different superscript letters following Dunn’smultiple comparisons test.
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337328

of overfishing, including growth overfishing [10], in the intensively fished areas ofMafia Island.
How MIMP affects the surrounding areas where more fish are removed is animportant question. MPAs aim to sustain a critical mass of fish and it is expectedthat this should have the secondary effect of boosting fish numbers in adjacent non-protected waters [37]. This perspective has been criticised as there are limits to thedispersal capabilities of reef fishes, and factors such as the hydrographic regime, thedistances separating reefs and the availability of suitable habitats, all have an impact[58]. Nevertheless, there is increasing quantitative evidence that enhanced recruit-ment and spillover (fish migration) are taking place [8,21,24]. Tupper and Rudd [25]noted that there was a linear relationship between the numbers of a target speciesand the distance to the centre of an MPA. In this study it was observed that Mfuruni,the most intensively fished location closest to Jujima Bay (where fishing intensity wasthe lowest) had a fairly higher fish abundance and biomass and larger mean size offish compared to Tumbuju, a site further away from MIMP. This could suggest thatfish spillover and larval settlement are occurring in the vicinity of the marineprotected area.
Habitat structure is an important factor in regulating fish numbers. The closerelationship between reef-fish communities and features of the reef habitat has beenwell documented [31,48,59–65]. In a quantitative ecological study examining reefstructure, benthos and fish at Mafia Island Marine Park, it was shown that 92% ofthe fish numbers were explained by habitat structure with live coral cover being theforemost predictor [48]. As L. fulviflamma is a mid-sized predator [66], the refugeavailability that is common on reefs with high rugosity would be expected toinfluence their survival. In addition, structurally complex reefs support preferredprey such as various invertebrates and smaller fish [67–68].
ARTICLE IN PRESS
Table 2Mean (7 s.e.) habitat characteristics at census sites listed in order of increasing fishing intensity index
Substrate Jujima Bay(n = 22)
Chole Bay(n = 14)
Tumbuju(n = 15)
Mfuruni(n = 14)
H p
Hard coral(%)
26.073.7a 28.074.0ab 13.272.5ac 18.272.8abc 9.427 *
Rock (%) 19.173.8a 12.572.7a 30.174.4ab 36.175.4b 16.224 ***Rubble (%) 9.072.2a 8.772.1a 31.174.0b 21.675.3ab 21.058 ***Sand (%) 17.273.5a 8.572.2a 13.473.5a 11.472.9a 3.178 NSAlgae &seagrass (%)
13.772.7a 15.372.4ab 4.971.7ac 5.171.8ac 15.699 ***
1Others (%) 15.073.6a 27.176.0ab 7.472.8ac 7.573.0ac 10.501 **Depth (m) 5.670.6a 9.270.8b 5.570.9a 4.470.6a 21.276 ***Rugosity 2.270.2a 3.070.2ab 1.370.2ac 1.570.2c 17.829 ***
1Others include dead coral, soft coral and sponge. Values that are significantly different have differentsuperscript letters in each row following Dunn’s multiple comparisons test.*** p o 0.001, ** p o 0.01, * po 0.05, NS = not significant p 4 0.05, df = 3.
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337 329
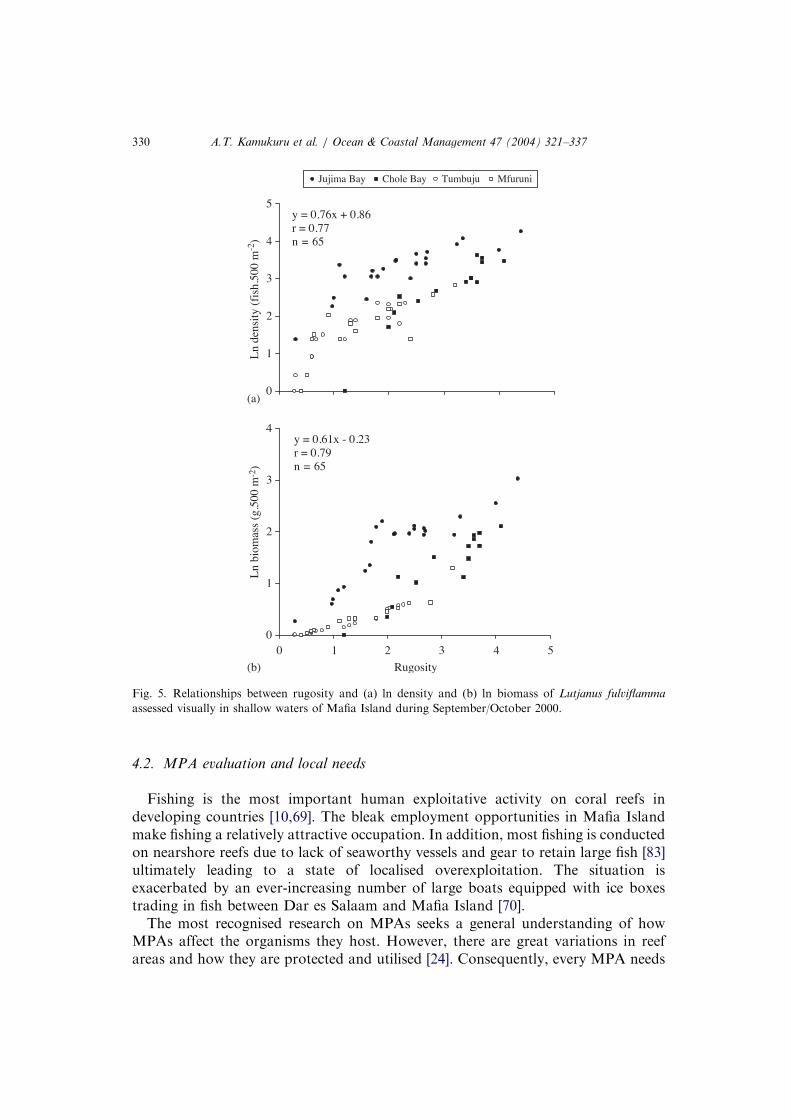
4.2. MPA evaluation and local needs
Fishing is the most important human exploitative activity on coral reefs indeveloping countries [10,69]. The bleak employment opportunities in Mafia Islandmake fishing a relatively attractive occupation. In addition, most fishing is conductedon nearshore reefs due to lack of seaworthy vessels and gear to retain large fish [83]ultimately leading to a state of localised overexploitation. The situation isexacerbated by an ever-increasing number of large boats equipped with ice boxestrading in fish between Dar es Salaam and Mafia Island [70].
The most recognised research on MPAs seeks a general understanding of howMPAs affect the organisms they host. However, there are great variations in reefareas and how they are protected and utilised [24]. Consequently, every MPA needs
ARTICLE IN PRESS
0
1
2
3
4
5
Ln d
ensit
y (fi
sh.5
00 m
-2)
Jujima Bay Chole Bay Tumbuju Mfuruni
y = 0.76x + 0.86 r = 0.77n = 65
(a)
0
1
2
3
4
0Rugosity
Ln b
iom
ass (
g.50
0 m
-2)
y = 0.61x - 0.23 r = 0.79n = 65
(b)1 2 3 4 5
Fig. 5. Relationships between rugosity and (a) ln density and (b) ln biomass of Lutjanus fulviflammaassessed visually in shallow waters of Mafia Island during September/October 2000.
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337330

its own evaluation as results from one reef area may only in part hold informationthat can be applied elsewhere. For example, McClanahan and co-workers [71–74] inKenya showed that the triggerfish–sea urchin interaction is an important factor inreef ecology which is influenced by the fishery. On the contrary, in many other reefareas in East Africa and elsewhere, sea urchins are not as common and consequentlythe organism is of less importance to MPA performance and fisheries management.
On comparing MPAs with adjacent areas, it is found that the more the number ofMPAs and comparable sites that are included in the study, the more reliable are thedata [11]. Nevertheless, if the aim is to generate data at low cost that will give anindication of MPA performance locally, a large-scale study may not be achievable or
ARTICLE IN PRESS
0
1
2
3
4
5
Ln d
ensit
y (fi
sh.5
00 m
-2)
Jujima Bay Chole Bay Tumbuju Mfuruni
(a)
y = 0.03x + 1.79 r = 0.39 n = 65
0
1
2
3
4
0 20 40 60 80Hard coral (%)
Ln b
iom
ass (
g.50
0 m
-2)
y = 0.02x + 0.50 r = 0.41 n = 65
(b)
Fig. 6. Relationships between percent hard coral cover and (a) ln density and (b) ln biomass of Lutjanusfulviflamma assessed visually in shallow waters of Mafia Island during September/October 2000.
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337 331

necessary. In addition, there is an upper limit to the number of sites that can beincluded because of limited numbers of comparable sites. In this study, we used twoplus two sites for comparing MIMP with adjacent IFAs. If other MPAs were to beincluded, the nearest well-functioning park would be Chumbe Island Coral Park inZanzibar Island (Unguja), whose composition differs from that of MIMP [75–77].
In this study we focused on a target species, which may act as a quick indicator ofhow well an MPA works. Depending on needs and available resources for additionalresearch, more exploited species can be examined. Moreover, certain species mayplay a key role in regulating functional groups within coral reefs [78]. Taxa such asButterfly fishes have been used as indicator species for disturbance [34] althoughthey, and other smaller species such as pomacentrids, are of less importance for thefishery in Mafia Island. Rare species may be more important to investigate than firstanticipated, as they could be the first to disappear. If time and resources permit, anear-complete coverage of all species may show how the entire reef community isaffected. However, as mentioned above, the project may then become more academicthan necessary for the immediate local needs. It should further be noted that theUVC-techniques used in this study have been criticised [79,80], one reason being thatit is difficult to count several species at the same time within a usually large area.However, with few, or as in our case, a single species, visual surveys are moreaccurate. It is further noticeable that the fishery-dependent data yielded the same fishsize structure with that of UVC. These observations are in agreement with Koslow etal. [81] who compared trap catches and UVC, and reported that the two estimatesprovided similar results of reef-fish sizes on Caribbean reefs.
In addition to fish data there are other factors that need to be considered whenexamining MPAs. Reef-fish communities are influenced by their habitat[31,34,48,60–65]. If this is not considered, differences between fished and non-fishedareas may be an effect of habitat only or a combination [47]. As there was a linearrelationship between fish and habitat structure in this study, it is difficult to separateone from the other. Reefs in the IFA had a lower percentage coral cover andstructural complexity than in the MIMP. Hence, the differences between the twoareas is probably not only an effect of fish removal, but also determined by habitat.This could, in part, also be the result of fishing practices that are caused by anchordamage, and/or destructive fishing techniques, which occur more frequently in theIFA.
5. Conclusions
In this study, we evaluated the performance of a marine protected area incomparison with surrounding intensively fished areas. For a rapid assessment at lowcost that would provide useful data on the local communities, we limited the study toone target species. The MPA had more individuals, higher biomass and larger fish.This seemed to be an effect of fishing, as fewer and smaller-sized fish were seen in theintensively fished areas. However, habitat was also an important aspect as fish datacorrelated positively with habitat features.
ARTICLE IN PRESS
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337332

There is much debate on how well MPAs work. The point made here is that thereis a difference between evaluating MPAs as a protecting measure in general termsand what the local needs are in a fishing village that depends on adjacent reefs forfood and income. For MPA evaluations a broad-scale quantitative study applyingvarious techniques including a BACIP design (Before-After-Control-Impact-Pairs)makes a strong case in fishery science but if fishers and managers only want to knowif there is a difference between their MPA and the surrounding reefs, such anendeavour becomes unnecessary for local needs.
This study gives information that should be considered in a management schemeregulating the fishery at Mafia Island. The MPA seemed to be a useful strategy forprotecting fish stocks and conserving habitats. Our results suggested that theintensively fished area was overfished, and of concern not only from its lowernumbers of fish but also their small size. The majority of the blackspot snappersassessed visually and caught in the fishery within the IFA had mean sizes far belowthe length of sexually mature fish [82]. This signifies that growth overfishing [10] is anadded measure of concern for the long-term sustainability of the fishery at thecurrent exploitation levels.
Acknowledgements
This study was funded by Sida/SAREC Bilateral Marine Science Programmebetween Sweden and Tanzania for research in marine zoology (Sida=SwedishInternational Development and Cooperation Agency). It was made possible throughthe co-operation of the Mafia Island Marine Park (MIMP) administration ingranting permission to conduct research within the marine park. We are mostgrateful to the Warden of MIMP Mr. George Msumi, the Technical Advisor of theWWF-MIMP project Mr. Jason Rubens, the Natural Resources Officer of MafiaDistrict Mr. F.O. Hemile and their staff for their help and assistance during thefieldwork. Thanks are also due to Mr. Nsajigwa Mbije at the Department ofZoology and Marine Biology, University of Dar es Salaam (currently at the SokoineUniversity of Agriculture, Morogoro, Tanzania), for his assistance during under-water reef surveys. Finally, the manuscript benefited from the comments andsuggestions of anonymous reviewers, to whom we are grateful.
References
[1] Anderson J, Ngazi Z. Marine resource use and the establishment of a marine park, Mafia Island,Tanzania. Ambio 1995;24:475–81.
[2] Berg HCE, Ohman MC, Troeng S, Linden O. Environmental economics of coral reef destruction inSri Lanka. Ambio 1998;27:627–34.
[3] Cesar HSJ, Lundin CG, Bettencourt S, Dixon J. Indonesian coral reefs—an economic analysis of aprecious but threatened resource. Ambio 1997;26:345–60.
[4] Cesar HSJ, Ohman MC, Espeut P, Honkanen M. Economic valuation of an integrated terrestrial andmarine protected area: Jamaica’s Portland Bight. In: Cesar HSJ, editor. Collected essays on theeconomics of coral reefs. Kalmar: Cordio; 2000. p. 203–14.
ARTICLE IN PRESS
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337 333

[5] Zann LP. The status of coral reef in south western Pacific islands. Marine Pollution Bulletin1994;29:52–61.
[6] McClanahan TR, Muthiga NA, Kamukuru AT, Machano H. Kiambo RW. The effects of marineparks and fishing on coral reefs of northern Tanzania. Biological Conservation 1999;89:161–82.
[7] Gladstone W. The ecological and social basis for management of a Red Sea marine protected area.Ocean & Coastal Management 2000;43:1015–32.
[8] Roberts CM, Polunin NVC. Are marine reserves effective in management of reef fisheries? Reviews inFish Biology and Fisheries 1991;1:65–91.
[9] Roberts CM, Polunin NVC. Marine reserves: simple solutions to managing complex fisheries? Ambio1993;22:363–8.
[10] Russ GR. Coral reef fisheries: effects and yields. In: Sale PF, editor. The ecology of fishes on coralreefs. San Diego: Academic Press; 1991. p. 601–35.
[11] Russ GR. Yet another review of marine reserves as reef fishery management tools. In: Sale PF, editor.Coral reef fishes; dynamics and diversity in a complex ecosystem. Amsterdam: Academic Press; 2002.p. 421–43.
[12] Bohnsack JA. Marine reserves: they enhance fisheries, reduce conflicts, and protect resources.Oceanus 1993;36:63–71.
[13] Bohnsack JA. Maintenance and recovery of reef fish productivity. In: Polunin NVC, Roberts CM,editors. Reef fisheries. London: Chapman & Hall; 1996. p. 283–313.
[14] Bohnsack JA. Application of marine reserves to reef fisheries management. Australian Journal ofEcology 1998;23:298–304.
[15] Clark CW. Marine reserves and the precautionary management of fisheries. Ecological Applications1996;6:369–70.
[16] Hall SJ. Closed areas for fisheries management—the case consolidates. Trends in Ecology andEvolution 1998;13:297–8.
[17] Lauck T, Clark CW, Mangel M, Munro GR. Implementing the precautionary principle in fisheriesmanagement through marine reserves. Ecological Applications 1998;8:72–8.
[18] Fogarty MJ. Essential habitat, marine reserves and fishery management. Trends in Ecology andEvolution 1999;14:133–4.
[19] Dayton PK, Sala E, Tegner MJ, Thrush SF. Marine protected areas: parks, baselines and fisheryenhancement. Bulletin of Marine Science 2000;66:617–34.
[20] Gell FR, Roberts CM. Benefits beyond boundaries: the fishery effect of marine reserves. Trends inEcology and Evolution 2003;18:448–55.
[21] Bohnsack JA. The potential of marine fishery reserves for reef fish management in the US southernAtlantic Miami. NOAA Technical Memo NMFS-SEFC, No. 261, 1990, 40pp.
[22] McClanahan TR, Kaunda-Arara B. Fishery recovery and its effect on the adjacent fishery.Conservation Biology 1996;10:1187–99.
[23] McClanahan TR, Mangi S. Spillover of exploitable fishes from a marine park and its effects on theadjacent fishery. Ecological Applications 2000;10:1792–805.
[24] Roberts CM, Bohnsack JA, Gell F, Hawkins JP, Goodridge R. Effects of marine reserves on adjacentfisheries. Science 2001;294:1920–3.
[25] Tupper M, Rudd MA. Species-specific impacts of a small marine reserve on reef fish production andfishing productivity in the Turks and Caicos Islands. Environmental Conservation 2002;29:484–92.
[26] Russ GR, Alcala AC. Effects of intensive fishing pressure on an assemblage of coral reef fishes.Marine Ecology Progress Series 1989;56:13–27.
[27] Russ GR, Alcala AC. Do marine reserves export adult fish biomass? Evidence from Apo Island,Central Philippines. Marine Ecology Progress Series 1996;132:1–9.
[28] Watson M, Ormond RFG. Effect of an artisanal fishery on the fish and sea urchin populations ofKenyan coral reef. Marine Ecology Progress Series 1994;109:115–29.
[29] Jennings S, Polunin NVC. Catch and effort relationships in Fijian multispecies reef fisheries subjectedto different levels of exploitation. Fisheries Management and Ecology 1995;2:98–101.
[30] Jennings S, Polunin NVC. Effects of fishing effort and catch rate upon the structure and biomass ofFijian reef fish communities. Journal of Applied Ecology 1996;33:400–12.
ARTICLE IN PRESS
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337334

[31] Jennings S, Boulle DP, Polunin NVC. Habitat correlates of the distribution and biomass ofSeychelles’ reef fishes. Environmental Biology of Fishes 1996;46:15–52.
[32] Jennings S, Marshall SS, Polunin NVC. Seychelles’ marine protected areas: comparative structureand status of reef fish communities. Biological Conservation 1996;75:201–9.
[33] Ohman MC, Rajasuriya A, Olafson E. Reef fish assemblages in northwestern Sri Lanka: distributionpatterns and influences of fishing practices. Environmental Biology of Fishes 1997;49:45–61.
[34] Ohman MC, Rajasuriya A, Svensson S. The use of butterflyfishes (Chaetodontidae) as bioindicatorsof habitat structure and human disturbance. Ambio 1998;27:708–16.
[35] Wantiez L, Thollot P, Kulbicki M. Effects of marine reserves on coral reef fish communities from fiveislands in New Caledonia. Coral Reefs 1997;16:215–24.
[36] Halpern BS. The impact of marine reserves: do reserves work and does reserve size matter? EcologicalApplications 2003;13:117–37.
[37] National Research Council. Marine Protected Areas: Tools for sustaining ocean systems. WashingtonDC: National Academy Press; 2001.
[38] Ngoile MAK, Francis J. Management of the coastal and marine environment. In: UNEP EasternAfrica Atlas of Coastal Resources (Tanzania). UNEP. 2001. p. 77–104.
[39] Samoilys MA. Abundance and species richness of coral reef fish on the Kenyan coast: the effects ofprotective management and fishing. Proceedings of sixth international coral reef symposium1988;2:261–6.
[40] McClanahan TR, Shafir SH. Causes and consequences of sea urchin abundance and diversity inKenyan coral reef lagoons. Oecologia 1990;83:362–70.
[41] Watson M, Dave D, Austin T, Ormond RFG. The effects of fishing on coral reef fish abundance anddiversity. Journal of Marine Biological Association UK 1996;76:229–33.
[42] McClanahan TR, Arthur R. The role of marine protected areas and habitat characteristics on EastAfrican coral reef fishes. Ecological Applications 2000;11:559–69.
[43] McClanahan TR, Uku JN, Machano H. Effect of macroalgae reduction on coral reef fish in theWatamuMarine National Park, Kenya. Journal of Marine and Freshwater Research 2002;53:223–31.
[44] Kamukuru AT. Effects of Fishing on growth and reproduction of Blackspot Snapper, Lutjanusfulviflamma (Pisces: Lutjanidae) in shallow waters of Mafia Island, Tanzania. Ph.D. Thesis,University of Dar es Salaam, Dar es Salaam; 2003 (274 pp).
[45] Horrill CJ, Darwall WRT, Ngoile MAK. Development of a marine protected area: Mafia Island,Tanzania. Ambio 1996;25:50–7.
[46] Francis J, Nilsson A, Waruinge D. Marine protected areas in the eastern African Region: howsuccessful are they? Ambio 2002;31:503–11.
[47] URT (United Republic of Tanzania). Mafia Island Marine Park: general management plan. Dar esSalaam: Board of Trustees, Marine Parks and Reserves Unit; 2000. 68pp.
[48] Garpe KC, Ohman MC. Coral and fish distribution patterns in Mafia Island Marine Park, Tanzania:fish-habitat interactions. Hydrobiologia 2003;498:191–211.
[49] Richmond MD, editor. A Guide to the seashores of Eastern Africa and the Western Indian Oceanislands. Stockholm: Swedish International Development Agency for the Swedish Agency forResearch Co-operation with Developing Countries (Sida/SAREC); 2002. 448pp.
[50] Druzhinin AD. The range and biology of snappers (Family Lutjanidae). Journal of Ichthyology1970;10:717–36.
[51] Samoilys MA. Underwater visual census surveys. In: Samoilys MA, editor. Manual for assessing fishstocks on Pacific coral reefs. Queensland Training Series QE 97009: Department of PrimaryIndustries; 1997. p. 16–29.
[52] English S, Wilkinson C, Baker V. Survey manual for tropical marine resources. ASEAN-AustraliaMarine Science Project. Townsville, Queensland: Australian Institute of Marine Sciences; 1994 358pp.
[53] Grigg RW. Effects of sewage discharge, fishing pressure and habitat complexity on coral ecosystemsand reef fishes in Hawaii. Marine Ecology Progress Series 1994;103:25–34.
[54] Conover WJ. Practical nonparametric statistics. NewYork: Wiley; 1980 493pp.[55] Sokal RR, Rohlf FJ. Biometry: The principles and practices of statistics in biological research, 3rd ed.
New York: WH Freeman & Company; 1995 887pp.
ARTICLE IN PRESS
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337 335

[56] Underwood AJ. Techniques of analysis of variance in experimental marine biology and ecology.Oceanography Marine Biology Annual Reviews 1981;19:513–603.
[57] Polunin NVC, Roberts CM. Greater biomass and value of target coral-reef fishes in two smallCaribbean marine reserves. Marine Ecology Progress Series 1993;100:167–76.
[58] Boehlert GW. Larval dispersal and survival in tropical reef fishes. In: Polunin NVC, Roberts CM,editors. Reef Fisheries. London: Chapman & Hall; 1996. p. 61–84.
[59] McClanahan TR. Kenyan coral reef lagoon fish: associations with reef management, complexity, andsea urchins. Coral Reefs 1994;13:231–41.
[60] Chabanet P, Ralambondrainy H, Amanieu M, Faure G, Galzin R. Relationships between coral reefsubstrata and fish. Coral Reefs 1997;16:93–102.
[61] Ohman MC, Rajasuriya A. Relationships between habitat structure and fish assemblages on coraland sandstone reefs. Environmental Biology of Fishes 1998;53:19–31.
[62] Ohman MC, Munday PL, Jones GP, Caley MJ. Settlement stratergies and distribution patterns ofcoral-reef fishes. Journal of Experimental Marine Biology and Ecology 1998;225:219–38.
[63] Bergman KC, Ohman MC, Svensson S. Influence of habitat structure on Pomacentrus sulfureus, awestern Indian Ocean reef fish. Environmental Biology of Fishes 2000;59:243–52.
[64] Lindahl U, Ohman MC, Schelten CK. The 1997/1998 mass mortality of corals: effects on fishcommunities on a Tanzanian coral reef. Marine Pollution Bulletin 2001;42:127–31.
[65] Rodwell LD, Barbier EB, Roberts CM, McClanahan TR. The importance of habitat quality formarine reserve—fishery linkages. Canadian Journal of Fisheries and Aquatic Sciences2003;60:171–81.
[66] Kamukuru AT, Mgaya YD. The food and feeding habits of blackspot snapper, Lutjanus fulviflamma
(Pisces: Lutjanidae) in shallow waters of Mafia Island, Tanzania. African Journal of Ecology2004;42:49–58.
[67] Kohn AJ, Leviten PJ. Effects of habitat complexity on population density and species richness intropical intertidal predatory gastropod assemblages. Oecologia 1976;25:199–210.
[68] Patton WK. Distribution and ecology of animals associated with branching corals (Acropora spp.)from the Great Barrier Reef, Australia. Bulletin of Marine Science 1994;55:193–211.
[69] McManus JW. Social and economic aspects of reef fisheries and their management. In: Polunin NVC,Roberts CM, editors. Reef fisheries. London: Chapman & Hall; 1996. p. 249–81.
[70] Mndeme YES. Mafia Island marine resources in peril. Naga, ICLARM Quarterly 1995;18:12–3.[71] McClanahan TR. Fish predators and scavengers of the sea urchin Echinometra mathaei in Kenyan
coral-reef marine parks. Environmental Biology of Fishes 1995;43:187–95.[72] McClanahan TR. Predation and the distribution and abundance of tropical sea urchin populations.
Journal of Experimental Marine Biology and Ecology 1998;221:231–55.[73] McClanahan TR, Kamukuru AT, Muthiga NA, Yebio MG, Obura D. Coral reef restoration: effect
of sea urchin reductions on algae, coral and fish assemblages. Conservation Biology 1996;10:136–54.
[74] Silva MC, McClanahan TR. Echinoid bioerosion and herbivory on Kenyan coral reefs: The role ofprotection from fishing. Journal of Experimental Marine Biology and Ecology 2001;262:133–53.
[75] Bergman KC, Ohman MC. Coral reef structure at Zanzibar Island. In: Richmond M, Francis J,editors. Marine Science development in Tanzania and Eastern Africa. Proceedings of 20th annualconference in Advances of Marine Sciences in Tanzania. 2001. p. 263–275.
[76] Muhando CA, Mohammed MS. Coral reef benthos and fisheries in Tanzania before and after the1998 bleaching and mortality event. Western Indian Ocean Journal of Marine Science 2002;1:43–52.
[77] Mbije NE, Wagner GM, Francis J, Ohman MC, Garpe KC. Patterns in the distribution andabundance of hard corals around Zanzibar Island. Ambio 2002;31:609–11.
[78] McClanahan TR. Recovery of the coral reef keystone predator, Balistapus undulatus, in East Africanmarine parks. Biological Conservation 2000;94:191–8.
[79] McCormick MI, Choat JH. Estimating total abundance of a large temperate-reef fish using visualstrip-transects. Marine Biology 1987;96:469–78.
[80] St John J, Russ GR, Gladstone W. Accuracy and bias of visual estimates of numbers, size structureand biomass of a coral reef fish. Marine Ecology Progress Series 1990;64:253–62.
ARTICLE IN PRESS
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337336

[81] Koslow JA, Hanley F, Wicklund R. Effects of fishing on reef fish communities at Pedro Bank andPort Royal Cays, Jamaica. Marine Ecology Progress Series 1988;43:201–12.
[82] Kamukuru AT, Mgaya YD. Effects of exploitation on reproductive capacity of blackspot snapper,Lutjanus fulviflamma (Pisces: Lutjanidae) in Mafia Island, Tanzania. African Journal of Ecology, inpress.
[83] Jiddawi NS, Ohman MC. Marine fisheries in Tanzania. Ambio 2002;31:518–27.
ARTICLE IN PRESS
A.T. Kamukuru et al. / Ocean & Coastal Management 47 (2004) 321–337 337


















