Ets1 blocks terminal differentiation of keratinocytes and...
10
3566 Research Article Introduction The stratified squamous epithelium of the skin is comprised of the inter-follicular epidermis (IFE), which covers most of the body surface, and interspersed hair follicles and sebaceous glands. Keratinocytes, the main cell type in the IFE, undergo a well- organized differentiation process to yield a water-impermeable, mechanically resistant cornified barrier that prevents both loss of internal fluids and entry of external substances and foreign bodies (Fuchs, 2007; Nagarajan et al., 2008; Proksch et al., 2008). During gestation, when a functional cornified layer has not yet developed, embryos are encased by periderm, a protective layer that acts as a transient barrier (M’Boneko and Merker, 1988). Stratified squamous epithelia of the hard palate and gingiva in the oral cavity undergo a differentiation process similar to that of IFE, leading to the production of a cornified epithelium (Presland and Jurevic, 2002; Squier and Kremer, 2001). In other areas of the oral mucosa, keratinocytes differentiate but do not cornify, hence producing a flexible, non-cornified layer capable of movements associated with speech, chewing and swallowing. The barrier function of the skin depends on the precise differentiation of the IFE keratinocytes, and disruptions to this sequence can cause serious, even life-threatening disease (Segre, 2006). Early stages of keratinocyte differentiation involve the expression of large amounts of intermediate filament proteins called keratins (Fuchs, 2007; Nagarajan et al., 2008; Proksch et al., 2008). In late stages of stratified squamous epithelial differentiation, the expression of genes encoding keratin proteins is downregulated and the expression of genes encoding cornified envelope proteins (such as involucrin, filaggrin, loricrin, small proline-rich proteins and late cornified envelope proteins) is upregulated (Fuchs, 2007; Nagarajan et al., 2008; Proksch et al., 2008). These cornified envelope constituents are crosslinked by the action of transglutaminase enzymes to form a tough sac enclosing the keratin filaments (Eckert et al., 2005; Hitomi, 2005). Granular layer keratinocytes synthesize and secrete a number of specialized lipids that contribute to the water-impermeable barrier function of the skin (Kalinin et al., 2002). The final differentiation step involves programmed cell death of the keratinocytes to yield anucleate squames that compose the cornified layer. Squamous cell carcinoma (SCC) is a malignant tumor that can arise in stratified squamous epithelia. SCC tumors exhibit aberrations in the normal differentiation pattern, which can include loss of cell polarity, hyperkeratosis, parakeratosis, acanthosis, increased mitotic figures, and crowded, pleomorphic and hyperchromatic nuclei (Arlette and Trotter, 2004; Bhawan, 2007; Cockerell, 2000). These changes to keratinocyte differentiation are caused by aberrant activation of oncogenes and inactivation of tumor suppressor genes. SCC lesions are also frequently associated with high-level expression of matrix metalloprotease (Mmp) enzymes, which are thought to contribute to tumor cell migration, invasion and proliferation (Ala-aho and Kahari, 2005; Kerkela and Saarialho-Kere, 2003; Westermarck and Kahari, 1999). Squamous cell tumors are often associated with a significant inflammatory response, which has been linked to tumor angiogenesis, Ets1 blocks terminal differentiation of keratinocytes and induces expression of matrix metalloproteases and innate immune mediators Priyadharsini Nagarajan 1, *, Shu Shien Chin 1 , Dan Wang 2 , Song Liu 2 , Satrajit Sinha 1,‡ and Lee Ann Garrett-Sinha 1,‡ 1 Department of Biochemistry, Developmental Genomics Focus Group, Center of Excellence in Bioinformatics and Life Sciences, State University of New York at Buffalo, Buffalo, NY 14203, USA 2 Department of Biostatistics, Roswell Park Cancer Institute, Buffalo, NY 14263, USA *Present address: Department of Pathology, Yale New Haven Hospital, New Haven, CT 06510, USA ‡ Authors for correspondence ([email protected], [email protected]) Accepted 9 July 2010 Journal of Cell Science 123, 3566-3575 © 2010. Published by The Company of Biologists Ltd doi:10.1242/jcs.062240 Summary The transcription factor Ets1 is normally expressed in the proliferative layer of stratified epithelium, but expression of Ets1 is significantly upregulated in squamous cell carcinomas. How elevated levels of Ets1 impact tumor initiation and progression is not well understood. To determine the biological consequences of overexpression of Ets1, we developed a transgenic mouse model that allows induction of Ets1 expression in keratinocytes of stratified epithelium in a regulatable fashion. Induction of Ets1 during embryonic development results in a dramatic alteration in epidermal structure and function by suppressing the expression of multiple stratum corneum constituents, while at the same time inducing expression of EGF ligands, AP1 transcription factors and matrix metalloproteases. Interestingly, expression of certain immune-related genes, including defensins, chemokines and cytokines was increased as well, suggesting a possible role for immune dysregulation in the promotion of squamous dysplasia. Experiments using cultured mouse keratinocytes indicate that Ets1 can induce expression of some of these mediators in a cell-intrinsic fashion. Collectively, our data reveal that elevated expression of Ets1 has a much broader array of pro-tumorigenic effects on epithelial cells than previously appreciated. Key words: Mmp, Chemokine, Differentiation, Keratinocyte, Transgenic, EGF ligand Journal of Cell Science
Transcript of Ets1 blocks terminal differentiation of keratinocytes and...

3566 Research Article
IntroductionThe stratified squamous epithelium of the skin is comprised of theinter-follicular epidermis (IFE), which covers most of the bodysurface, and interspersed hair follicles and sebaceous glands.Keratinocytes, the main cell type in the IFE, undergo a well-organized differentiation process to yield a water-impermeable,mechanically resistant cornified barrier that prevents both loss ofinternal fluids and entry of external substances and foreign bodies(Fuchs, 2007; Nagarajan et al., 2008; Proksch et al., 2008). Duringgestation, when a functional cornified layer has not yet developed,embryos are encased by periderm, a protective layer that acts as atransient barrier (M’Boneko and Merker, 1988). Stratified squamousepithelia of the hard palate and gingiva in the oral cavity undergoa differentiation process similar to that of IFE, leading to theproduction of a cornified epithelium (Presland and Jurevic, 2002;Squier and Kremer, 2001). In other areas of the oral mucosa,keratinocytes differentiate but do not cornify, hence producing aflexible, non-cornified layer capable of movements associated withspeech, chewing and swallowing.
The barrier function of the skin depends on the precisedifferentiation of the IFE keratinocytes, and disruptions to thissequence can cause serious, even life-threatening disease (Segre,2006). Early stages of keratinocyte differentiation involve theexpression of large amounts of intermediate filament proteinscalled keratins (Fuchs, 2007; Nagarajan et al., 2008; Proksch et al.,2008). In late stages of stratified squamous epithelial differentiation,the expression of genes encoding keratin proteins is downregulated
and the expression of genes encoding cornified envelope proteins(such as involucrin, filaggrin, loricrin, small proline-rich proteinsand late cornified envelope proteins) is upregulated (Fuchs, 2007;Nagarajan et al., 2008; Proksch et al., 2008). These cornifiedenvelope constituents are crosslinked by the action oftransglutaminase enzymes to form a tough sac enclosing the keratinfilaments (Eckert et al., 2005; Hitomi, 2005). Granular layerkeratinocytes synthesize and secrete a number of specialized lipidsthat contribute to the water-impermeable barrier function of theskin (Kalinin et al., 2002). The final differentiation step involvesprogrammed cell death of the keratinocytes to yield anucleatesquames that compose the cornified layer.
Squamous cell carcinoma (SCC) is a malignant tumor that canarise in stratified squamous epithelia. SCC tumors exhibitaberrations in the normal differentiation pattern, which can includeloss of cell polarity, hyperkeratosis, parakeratosis, acanthosis,increased mitotic figures, and crowded, pleomorphic andhyperchromatic nuclei (Arlette and Trotter, 2004; Bhawan, 2007;Cockerell, 2000). These changes to keratinocyte differentiation arecaused by aberrant activation of oncogenes and inactivation oftumor suppressor genes. SCC lesions are also frequently associatedwith high-level expression of matrix metalloprotease (Mmp)enzymes, which are thought to contribute to tumor cell migration,invasion and proliferation (Ala-aho and Kahari, 2005; Kerkela andSaarialho-Kere, 2003; Westermarck and Kahari, 1999). Squamouscell tumors are often associated with a significant inflammatoryresponse, which has been linked to tumor angiogenesis,
Ets1 blocks terminal differentiation of keratinocytesand induces expression of matrix metalloproteasesand innate immune mediatorsPriyadharsini Nagarajan1,*, Shu Shien Chin1, Dan Wang2, Song Liu2, Satrajit Sinha1,‡ andLee Ann Garrett-Sinha1,‡
1Department of Biochemistry, Developmental Genomics Focus Group, Center of Excellence in Bioinformatics and Life Sciences,State University of New York at Buffalo, Buffalo, NY 14203, USA2Department of Biostatistics, Roswell Park Cancer Institute, Buffalo, NY 14263, USA*Present address: Department of Pathology, Yale New Haven Hospital, New Haven, CT 06510, USA‡Authors for correspondence ([email protected], [email protected])
Accepted 9 July 2010Journal of Cell Science 123, 3566-3575 © 2010. Published by The Company of Biologists Ltddoi:10.1242/jcs.062240
SummaryThe transcription factor Ets1 is normally expressed in the proliferative layer of stratified epithelium, but expression of Ets1 issignificantly upregulated in squamous cell carcinomas. How elevated levels of Ets1 impact tumor initiation and progression is not wellunderstood. To determine the biological consequences of overexpression of Ets1, we developed a transgenic mouse model that allowsinduction of Ets1 expression in keratinocytes of stratified epithelium in a regulatable fashion. Induction of Ets1 during embryonicdevelopment results in a dramatic alteration in epidermal structure and function by suppressing the expression of multiple stratumcorneum constituents, while at the same time inducing expression of EGF ligands, AP1 transcription factors and matrix metalloproteases.Interestingly, expression of certain immune-related genes, including defensins, chemokines and cytokines was increased as well,suggesting a possible role for immune dysregulation in the promotion of squamous dysplasia. Experiments using cultured mousekeratinocytes indicate that Ets1 can induce expression of some of these mediators in a cell-intrinsic fashion. Collectively, our datareveal that elevated expression of Ets1 has a much broader array of pro-tumorigenic effects on epithelial cells than previouslyappreciated.
Key words: Mmp, Chemokine, Differentiation, Keratinocyte, Transgenic, EGF ligand
Jour
nal o
f Cel
l Sci
ence

proliferation and migration (Cataisson et al., 2009; Coppe et al.,2008; Coussens et al., 1999; Takahashi et al., 2010).
One of the oncogenes linked to the pathogenesis of SCC is thetranscription factor Ets1, a downstream effector of the Ras–MAPKpathway. In normal tissues, Ets1 is expressed only in the basallayer of stratified squamous epithelium (Nagarajan et al., 2009).However, in a large fraction of SCC tumors, Ets1 is highlyexpressed within the malignant keratinocytes, and its levels directlycorrelate with the invasive and metastatic potential of the tumor(Bai et al., 2009; Endo et al., 2006; Keehn et al., 2004; Pande etal., 1999; Saeki et al., 2000; Vairaktaris et al., 2007). Tumors thatexpress high levels of Ets1 typically also express high levels ofseveral matrix metalloprotease (Mmp) genes (Dittmer, 2003; Hahneet al., 2008; Lincoln and Bove, 2005) and Ets1 has been shown todirectly regulate expression of multiple Mmp genes (Baillat et al.,2006; Nagarajan et al., 2009; Ozaki et al., 2000; Reisdorff et al.,2002; Wei et al., 2009; Westermarck et al., 1997). Yet, it has beenunclear whether the sole oncogenic role of Ets1 is upregulation ofMmp expression or whether this protein also promotes squamouscell carcinoma development by affecting other pathways. To addressthis question, we developed an inducible transgenic mouse modelin which Ets1 can be induced in differentiating squamouskeratinocytes (Nagarajan et al., 2009). Our studies indicate thatinduction of Ets1 in adult mice leads to severe dysplastic lesionsaccompanied by high levels of several Mmp enzymes, most notablyMmp13. To further explore the role of Ets1 in squamous epithelialhomeostasis as well as carcinogenic transformation, we now reportthe immediate effects of Ets1 induction during embryonicdevelopment of mice. As described herein, our studies identify anumber of key regulatory programs that are influenced by Ets1 andthat might have causal implications in squamous carcinogenesis.
ResultsEctopic expression of Ets1 in differentiated layers of theepidermis leads to major epidermal defectsWe have previously described a bi-transgenic (designated BT)mouse model system in which we can inducibly express thetranscription factor Ets1 in a tightly regulated, tissue- anddifferentiation-specific pattern (Nagarajan et al., 2009). In thissystem, one transgenic line (the responder transgenic) contains anN-terminal hemagglutinin (HA)-tagged mouse Ets1 transgene,cloned downstream of the doxycycline-regulated pTight promoter.Another transgenic line (the driver transgenic), expresses thetetracycline-regulated transactivator protein (tTA) under the controlof the human involucrin promoter and enhancer elements (Jaubertet al., 2004). The involucrin promoter is active in the upperspinous and granular layers of stratified squamous epithelium,including skin epidermis. BT offspring derived from crossingthese two transgenic mouse lines express HA-tagged Ets1 in theupper spinous and granular layers of stratified squamous epithelialtissues in the absence of doxycycline (i.e. a Tet-off system).Induction of Ets1 in adult mice leads to severe dysplastic lesionsof the skin, reminiscent of squamous cell carcinoma-in-situ(Nagarajan et al., 2009).
To further explore the effects of Ets1 induction on the epidermis,we withheld doxycycline supplementation during gestation. Underthese conditions, the HA-Ets1 transgene is expected to be expressedwhen the involucrin promoter and enhancer elements becomeactive during embryonic development (i.e. beginning at E15.5, andupregulated at E16.5 and later time points) (Jaubert et al., 2004).Indeed, at embryonic day E18.5 and in newborn mice (NB), we
3567Oncogenic effects of Ets1 in skin
detected strong expression of the transgene using immunostaining(Fig. 1A). Overexpression of the Ets1 transgene was confirmedwith western blotting of skin extracts from E18.5 day embryos(supplementary material Fig. S1). BT mice were born in theexpected ratio and could be easily distinguished from their wild-type and single transgenic littermates because they were slightlysmaller and had a shiny, taut and translucent skin (Fig. 1B). Inaddition, some BT embryos also demonstrated an eyes-open-at-birth phenotype. The BT animals did not survive long after birth,probably because their severe skin phenotype led to rapiddehydration.
To analyze the epidermal defects caused by Ets1 induction, weexamined skin sections from E18.5 and newborn wild-type and BTmice (Fig. 1C,D). The BT epidermis was hyperplastic, as evidencedby the increased thickness. Cells in the epidermal basal layerappeared crowded, causing the cells to adopt a columnar appearanceand suggesting increased cellular proliferation in this compartment.The epidermis contained a compact cornified layer of reducedthickness, while the granular layer was largely absent(hypogranulosis) and, instead, the bulk of the suprabasal
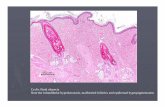
Fig. 1. Epidermal defects in mice overexpressing Ets1 in the suprabasallayers of epidermis. (A)Expression of the HA-Ets1 transgene in the skin ofBT mice. Arrow indicates transgene expression in the inner root sheath of thehair follicle. (B)BT newborn mice are smaller, with an eyes-open-at-birthphenotype (arrow) and have a shiny, translucent skin. (C,D)Hematoxylin andeosin staining to identify epidermal alterations in E18.5 (C) and newborn(NB) (D) BT mice as compared with the wild-type. The BT epidermis ishyperplastic, has a reduced granular layer (white brackets) and is characterizedby parakeratosis (arrows) and a compact stratum corneum. Arrowhead in Cindicates the presence of a superficial dermal capillary.
Jour
nal o
f Cel
l Sci
ence

compartment was occupied by spinous keratinocytes (acanthosis).In addition, there was retention of nuclei in the stratum corneum(parakeratosis) of the BT epidermis. These histological alterationsindicate that there was aberrant terminal differentiation of BTkeratinocytes. The dermal compartment appeared largely normal,except for the presence of increased numbers of superficial dermalcapillaries.
Induction of Ets1 results in epidermal barrier insufficiencyTo determine whether the epidermal alterations of the BT mice ledto an impairment in barrier function, we performed a whole mountskin permeability assay (Hardman et al., 1998) at various stages ofembryonic development (Fig. 2A). BT mouse embryos exhibitedan impaired epidermal barrier function at both E18.5 and at birth.To determine whether there were structural defects in the stratumcorneum, we compared the morphology of wild-type and BTcornified epidermocytes (Fig. 2B). Whereas the wild-type squameswere intact and polygonal, the BT cornified envelopes were smalland irregularly shaped. Less than 5% retained their polygonalshape, while the rest were grossly fragmented and disintegrating.This phenotype suggests that expression of Ets1 might interferewith the normal program of cornification in the epidermis, whichis the probable cause of the epidermal barrier defect and perinatallethality in these animals.
Expression of Ets1 in differentiating keratinocytesdisrupts the epidermal proliferation–differentiationequilibriumTo further examine the epidermal differentiation defects, wecompared the expression of keratinocyte marker proteins in newbornBT and wild-type skin. We have previously shown that in adultmice, induction of Ets1 leads to expansion of the domain thatexpresses early keratinocyte differentiation markers such as keratin14 (K14) and keratin 10 (K10), while there is a concomitant loss ofexpression of late differentiation markers, such as loricrin (Nagarajanet al., 2009). We observed a similar pattern of altered differentiation
3568 Journal of Cell Science 123 (20)
in BT embryos in that K10 and K14 expression was increased,whereas loricrin and filaggrin expression was decreased (Fig. 3;supplementary material Fig. S2). The expression of involucrin wasnormal in BT newborn mice (Fig. 3C), but was significantlydecreased in E18.5 embryos (supplementary material Fig. S2). Theresults of the immunostaining were confirmed with western blottingof skin protein extracts (supplementary material Fig. S1). Decreasedexpression of these late markers is consistent with the strikingreduction of granular layer keratinocytes, which contain keratohyalingranules composed of these proteins. It is also consistent with amajor defect in assembly of the cornified envelope. Defects in thedifferentiation of oral mucosa were also observed with increasedexpression of keratin 5 (K5), keratin (K6) and proliferating cellnuclear antigen (PCNA) and decreased expression of keratin 13(K13) and loricrin (supplementary material Figs S3 and S4).
Fig. 2. Epidermal permeability barrier deficiency in mice overexpressingEts1. (A)Dye exclusion assay on E16.5, E18.5 and newborn (NB) mice.(B)Morphology of newborn wild-type (top) and BT (bottom) cornifiedenvelopes; inset shows higher magnification of the boxed regions. The wild-type demonstrates intact, polygonal cornified envelopes, whereas the BTshows two intact and three fragmented cornified envelopes.
Fig. 3. Ets1 overexpression in the suprabasal layer blocks keratinocyteterminal differentiation. Immunofluorescent staining of epidermaldifferentiation markers on dorsal skin of newborn BT and wild-type micecounterstained with b4 integrin to mark the basement membrane. Thefollowing markers were used to assess the differentiation pattern of wild-typeand BT epidermis: (A) K14, (B) K10, (C) involucrin (INV), (D) filaggrin(FIL) and (E) loricrin (LOR).
Jour
nal o
f Cel
l Sci
ence

Similar to previous observations in adult mice, the skin of E18.5BT embryos expressed K6 (a marker normally absent from E18.5IFE) in the suprabasal layers (Fig. 4A). Expression of K6 by BTepidermis might reflect a stressed or hyperproliferative condition inthe late-stage gestation epidermis. Indeed, BT epidermis was highlyproliferative, as demonstrated by the expression of Ki67, PCNA andthe basal cell marker DNp63, all of which extended to three to fourlayers in the BT epidermis as compared with only the basal layer inthe controls (Fig. 4B–D). Together, our studies indicate that Ets1expression in differentiating keratinocytes inhibits terminaldifferentiation and cornification of the developing epidermis, whileat the same time promoting epidermal proliferation.
Ets1 induction does not affect periderm morphogenesisThe periderm is a transient intra-uterine barrier that protects thedeveloping embryo before the definitive epidermal barrier isgenerated (M’Boneko and Merker, 1988). When epidermalcornification is complete, the periderm is shed. Because Ets1expression blocks the cornification of stratifying keratinocytesresulting in a defective epidermal permeability barrier, the sheddingof periderm in BT embryos might be delayed. Moreover, because
3569Oncogenic effects of Ets1 in skin
periderm cells express involucrin (Akiyama et al., 1999), the Ets1transgene might be expressed in BT periderm, potentially leadingto alterations in its morphology, maturation or shedding. Toinvestigate these possibilities, we examined periderm organizationin BT embryos and their littermates by histological examination ofE16.5 embryonic skin (Fig. 5A). At this stage, the developingepidermis is typically two to three cell layers thick and overlaid bythe darkly staining periderm. The morphology of the periderm wassimilar in wild-type and BT embryos.
Because no morphologic differences could be identified betweenwild-type and BT periderm, we examined whether the expressionof the periderm marker protein K6 was altered in BT mice (Fig.5B). Both the wild-type and the BT sections stained similarly forthe expression of K6, indicating that the periderm is not affectedat this stage. We then examined E18.5 embryos, when peridermshedding is usually complete. Upon histological examination, noperiderm remnants were identified in either sample at this stage(Fig. 5C). Immunostaining for K6 revealed its localization in theinner root sheath (IRS) in the wild-type sample (Fig. 5D), whereasin the BT sample, K6 was expressed in the hyperplastic BTepidermal keratinocytes as well as in the IRS. Thus, peridermgeneration and shedding was not affected by ectopic expression ofEts1 in the suprabasal layers.
Ets1 induction does not affect the hair folliclemorphogenesis programAnother morphogenetic program that coincides with the later stagesof epidermal maturation is the development of the hair follicle. To
Fig. 4. Ets1 induction in the suprabasal layers results in keratinocyteactivation and enhanced proliferation. Immunostaining for keratinocyteactivation and proliferation markers on dorsal skin of BT newborn mice andlittermate wild-type mice. Samples were counterstained with DAPI to detectnuclei or b4 integrin to mark the basement membrane. The following markerswere used (A) K6, (B) DNp63, (C) Ki67 and (D) PCNA.
Fig. 5. Periderm shedding is not affected by suprabasal expression of Ets1.Skin sections of E16.5 embryos stained with (A) hematoxylin and eosin(dashed lines mark the periderm–epidermal junction) or (B) K6,counterstained with DAPI (white dashed line represents the location ofepidermal basement membrane, grey dashed line indicates the periderm–epidermal interface). (C,D)Same as above for E18.5 embryonic skin. Thearrowheads mark the surface of epidermis.
Jour
nal o
f Cel
l Sci
ence

investigate whether altered expression of Ets1 affects hair follicledevelopment, we examined sections of E18.5 embryo and newbornskin. Histological examination revealed that hair follicles of wild-type and BT embryos were at an equivalent developmental stage, i.e.the elongated bulbous peg stage (Fig. 6A). The maturation of dermalpapilla (dermal fibroblasts enclosed within the lower end of hairfollicle) as marked by alkaline phosphatase staining was normal inwild-type and BT skin (Fig. 6B). Similar results were obtained withsections from newborn mice (Fig. 6C,D). The expression of P-cadherin, a marker for the hair follicle matrix and regions of outerroot sheath and for the basal layer of inter-follicular epidermis, wassimilar in BT and control mice (Fig. 6E). We also examined earlierstages of hair follicle morphogenesis at E15.5 and E16.5. Theseearly morphogenetic events of hair follicle development were alsounaffected (supplementary material Fig. S5). Thus, the epidermo–mesenchymal interactions that induce and maintain hair follicles arenot affected by Ets1 induction, despite significant alterations in theoverall maturation of the epidermis.
3570 Journal of Cell Science 123 (20)
Induction of Ets1 alters programs of keratinocytedifferentiation at the mRNA levelTo understand the molecular mechanisms by which Ets1 functionsas a pro-proliferative and anti-differentiation signal in stratifiedsquamous epithelia, we chose to investigate global gene expressionprofiles using microarray technology. For these studies, weharvested skin from wild-type and BT epidermis at day 18.5 ofembryonic development. We chose to do these analyses at thisstage of differentiation for the following reasons: first, the uterineenvironment is sterile, which eliminates any possibility of secondarychanges in the skin owing to bacterial infection; and second, theshort duration of induction of the Ets1 transgene (2–3 days) meantthat the observed changes in gene expression were more likely tobe owing to primary or direct effects of Ets1 on keratinocytes,rather than on global changes in the physiology of the animal.
Gene expression microarray experiments identified a large numberof genes (~2200) that displayed statistically significant (P≤0.05)changes between wild-type and BT epidermis (supplementarymaterial Table S1). Of those, there were ~470 genes displaying atleast twofold upregulation and ~220 genes showing at least twofolddownregulation. Expression patterns of selected genes aresummarized in the heat map visualizations shown in Fig. 7. RT-PCRconfirmation of changes in the expression of some of these genes isshown in supplementary material Fig. S6.
Ets1 strikingly alters expression of genes encodingepidermal structural proteinsExamination of the data shows that induction of the Ets1 transgenedramatically inhibits the expression of many genes expressed in
Fig. 6. Early hair follicle morphogenesis is similar in wild-type and BTembryos. Skin sections of E18.5 wild-type and BT embryos stained with (A)hematoxylin and eosin or (B) for alkaline phosphatase activity (Alk. Ph.) tomark the dermal papilla (counterstained with hematoxylin). (C)Newborn skinstained with hematoxylin and eosin. (D)Newborn skin stained for alkalinephosphatase and counterstained with DAPI. (E)Newborn skin stained for P-cadherin and counterstained with b4 integrin to mark the basement membrane.
Fig. 7. Induction of Ets1 leads to major alterations in the differentiationprogram of keratinocytes. Heatmap visualizations of changes in expressionof genes involved in keratinocyte (A) structure, (B) proliferation, (C) motilityand (D) immune function. Red indicates increased expression, whereas greenindicates decreased expression.
Jour
nal o
f Cel
l Sci
ence

the cornified envelope (Fig. 7A) including Lor and Fil (encodingloricrin and filaggrin, respectively) as well as multiple group 1 latecornified envelope (Lce) genes (Lce1a1, Lce1a2, Lce1b, Lce1c,Lce1d, Lce1f, Lce1h and Lce1i). Interestingly, all of these genesare closely linked on mouse chromosome 3F1 and found withinthe epidermal differentiation complex (EDC) (Brown et al., 2007).However, the expression of certain other genes within the EDC,such as Ivl (encoding involucrin) and the S100 genes, was notchanged. Furthermore, the expression of some genes in the EDC,including many of the small proline-rich protein genes (Sprr genes)and some group 3 Lce genes (Lce3a and Lce3c), was dramaticallyupregulated. The Tgm1 gene (encoding transglutaminase 1) islocalized outside the EDC, but plays a major role in formation ofthe cornified envelope by crosslinking envelope proteins and lipidsto form the structurally resistant cornified envelope of thekeratinocyte (Eckert et al., 2005; Hitomi, 2005). Tgm1 demonstratedincreased expression, possibly as a response to impaired barrierformation. Decreased expression of loricrin, filaggrin and severalLce genes is highly consistent with the phenotypic abnormalitiesof the skin and corneocytes described above.
In keeping with our immunostaining results (Fig. 4), we detectedincreased expression of Krt6 (the gene encoding K6) and its partnerKrt16 (encoding keratin 16) in BT skin (Fig. 7A). There was alsoincreased expression of Krt17 (encoding keratin 17), another markerof keratinocyte activation. BT skin demonstrated reducedexpression of certain hair-associated genes including Tchh(trichohyalin) and Krt25, Krt27, Krt32 and Krt71, but increasedexpression of the hair-shaft-associated Tgm3 (transglutaminase 3)and Cdh3 (P-cadherin) genes.
Expression of several connexins (which are involved in gapjunction formation) was also affected, with Gjb2 and Gjb6(connexins 26 and 30) being upregulated in BT skin, while Gjb4,Gjb3 and Gjb5 (connexins 30.3, 31 and 31.1) were downregulated(Fig. 7A). Furthermore, the expression of the Prss8 gene [encodingchannel activating protease 1 (CAP1) or prostasin] was alsodownregulated, although expression of the epithelial sodiumchannel -subunit (which is activated by CAP1) was upregulated.Prss8 is required for normal skin barrier acquisition, and Prss8knockout epidermis demonstrates an absence of functional tightjunctions within the granular layer (Leyvraz et al., 2005). Thus, thestructural integrity of both gap junctions and tight junctions mightbe affected in BT skin.
Induction of Ets1 affects expression of growth-associatedgenes and matrix metalloproteasesWe detected upregulation in the expression of several epidermalgrowth factor receptor (EGF-R) ligands in BT skin, including Areg(encoding amphiregulin), Epgn (encoding epigen), Tgfa (encodingTGF) and Hbegf (encoding heparin-binding EGF) (Fig. 7B). Afifth EGF receptor ligand, Btc (encoding betacellulin), wasdownregulated in BT skin, whereas two others, EGF (Egf) andepiregulin (Ereg), were unchanged. EGF-R ligands are importantfor keratinocyte proliferation; hence, increased expression of certainEGF-R ligands by BT keratinocytes might be the cause of thehyperproliferative effects of Ets1 on the epidermis.
EGF signaling triggered by its ligands is known to stimulate theexpression of several AP1 proteins (Tomic-Canic et al., 1998) andwe detected increased expression of several AP1 family genes inBT skin, including Fos, Fosl1, Fosl2 and Junb (Fig. 7B). BothEts1 and AP1 proteins are known to be potent transactivators ofMmp genes. In keeping with this, we noted strong upregulation of
3571Oncogenic effects of Ets1 in skin
several members of the matrix metalloprotease gene family (Fig.7C), most prominently Mmp13. We have previously detected highMmp13 expression in adult BT mice, in which Ets1 transgeneexpression was induced at weaning age (Nagarajan et al., 2009).The induction of these Mmp enzymes might lead to degradation ofepidermal extracellular components such as adherens junctions ordesmosomes, or dermal components such as collagen, and mightcontribute to increased keratinocyte motility.
Induction of Ets1 causes a sterile inflammatory responseUnexpectedly, microarray analysis detected altered expression ofmany genes involved in the epidermal immune response, includingdefensins, cytokines, chemokines and chemokine receptors (Fig.7D). Because the skin was harvested from E18.5 embryos in a sterileuterine environment, it is highly unlikely that the upregulation ofimmune-response genes represents a secondary response to infection.Instead, it is likely that such genes are upregulated as a response tothe failure in barrier formation, perhaps as a physiological responseto potential infection at birth. Immunostaining confirmed that therewas an infiltration of CD11b+ myeloid cells in the dermis andepidermis of BT skin (supplementary material Fig. S7). In summary,our gene expression analyses indicate that expression of Ets1 in thedifferentiating keratinocytes of the suprabasal layers of the epidermisleads to a block in expression of cornified envelope proteins, whileat the same time promoting expression of genes involved inkeratinocyte proliferation, motility and immune response.
Ets1 induces inflammation in keratinocytes in a cell-intrinsic fashionThe expression of the immune-related genes might be owing totheir induction by Ets1 in keratinocytes. Alternatively, infiltrating
Fig. 8. Overexpression of Ets1 in cultured keratinocytes drives expressionof Mmps and chemokines. (A)Retroviral constructs used to infectkeratinocytes: MIGR1, control empty retrovirus; MIGR1-Ets1, retrovirusencoding mouse Ets1; MIGR1-R391D, retrovirus encoding the DNA-binding-defective mutant of Ets1. R391D represents a mutation in a key arginineresidue of Ets1 required for DNA binding. This mutant version of Ets1 fails tobind DNA or regulate target genes. (B)RT-PCR analysis of Mmp andchemokine gene expression in retrovirally infected mouse keratinocytes. Ccl2,Ccl20 and Cxcl5 are the three chemokines tested; Mmp9 and Mmp13 are thetwo Mmps tested; Hprt is an internal control for RNA quantity and integrity.
Jour
nal o
f Cel
l Sci
ence

inflammatory cells might be the major producers of these proteins.To determine whether Ets1 can directly induce expression ofinflammatory genes in keratinocytes, we infected cultured mousekeratinocytes with a retrovirus encoding mouse Ets1, with aretrovirus encoding a DNA-binding-defective mutant of Ets1 orwith a control empty retrovirus (Fig. 8A). Cells infected with thewild-type Ets1 retrovirus demonstrated strong upregulation of eachof three chemokine genes tested, Ccl2, Ccl20 and Cxcl5 (Fig. 8B).By contrast, both the empty retrovirus as well as the DNA-binding-deficient Ets1 retrovirus failed to induce these genes. These resultssuggest that Ets1 binds to the regulatory segments for thechemokine genes and directly induces their expression. Indeed,transduction of a wild-type Ets1 retrovirus, but not the mutant Ets1retrovirus, also induced the expression of two Mmp genes that arewell-validated Ets1 targets (Mmp9 and Mmp13) (Fig. 8B).
DiscussionThe Ets1 transcription factor is normally expressed within theproliferative basal layer of the epidermis (Nagarajan et al., 2009),but in many squamous cell cancers, Ets1 is overexpressed inmalignant keratinocytes, where its expression is correlated withinvasion and metastasis (Bai et al., 2009; Endo et al., 2006; Keehnet al., 2004; Pande et al., 1999; Saeki et al., 2000; Vairaktaris et al.,2007). In order to address the potential role of Ets1 in keratinocytebiology and in the induction of squamous cell tumors, we generateda mouse model in which we can inducibly express Ets1 inkeratinocytes that have already begun their differentiation process.When Ets1 is induced in the epidermis in adult mice, severedysplastic lesions develop, consistent with an important role forEts1 in driving the pathological process (Nagarajan et al., 2009).In this report, we further explore the effects of Ets1 on skindevelopment and investigate the mechanisms of Ets1 oncogenicaction.
Induction of Ets1 in differentiating keratinocytes leads to afailure of cornified envelope formationEts1 causes dramatic alterations to the normal differentiationprogram of the epidermis, leading to a failure in barrier formation(as demonstrated by impaired dye exclusion), resulting in earlypostnatal lethality. The striking effects induced by Ets1overexpression in differentiating keratinocytes appear to be limitedmainly to IFE keratinocytes of the skin. The refractoriness to theeffects of Ets1 in oral, hair follicle or periderm cells might reflectlower expression of the Ets1 transgene in these tissues.Alternatively, Ets1 might have different effects because a distinctset of developmental cues and signaling pathways mediatedevelopment and homeostasis in these tissues. The barrier defectin the skin is probably owing to aberrant formation of cornifiedenvelopes, because Ets1 prevents proper expression of filaggrin,loricrin and group 1 late cornified envelope (Lce) genes. Expressionof another major cornified envelope constituent, involucrin, wasdecreased in BT embryos at E18.5, but not at the newborn stage.This is most consistent with a delayed, but otherwise normal,involucrin expression.
Individual mutations in either loricrin (gene knockout) orfilaggrin (in the spontaneous flaky tail mouse mutant) do not leadto major defects in the assembly of the cornified envelope (Kochet al., 2000; Presland et al., 2000). However, the downregulationof loricrin, filaggrin and Lce proteins together, as seen in Ets1 BTmice, appears to result in major alterations to the cornified envelopeof the cell, as demonstrated by the irregular shapes and
3572 Journal of Cell Science 123 (20)
fragmentation of the squames extracted from newborn skin. It hasbeen proposed that alterations to the barrier function of skin mightmake the epidermal cells more susceptible to environmentalcarcinogens that can gain easier access to the deeper layers of theskin in the absence of a proper barrier (Demehri et al., 2009). Thusthe alteration to skin barrier function as seen in BT mice mightrepresent one important mechanism by which Ets1 acts as anoncogene.
The expression of some of the small proline-rich proteins (Sprr)is strongly upregulated in BT epidermis along with expression ofsome group 3 Lce genes. Interestingly, one study has identified anEts binding site in the promoter segment of the human SPRR1Agene, which was shown to bind the Ets factor ESE-1 (ELF3) (Sarket al., 1998). Potentially, other Ets factors, such as Ets1, could alsobind to the same site and result in induction of the Sprr1a gene inBT mice. Group 3 Lce genes are normally expressed in internalstratified squamous epithelia (such as oral epithelia), but not inepidermis. Dramatic upregulation of one group 3 Lce gene (LCE3E)has been reported in human epidermal keratinocytes in response tothe stress imposed by UV radiation (Jackson et al., 2005). Anotherstudy showed upregulation of human group 3 LCE genes inresponse to tape-stripping of the cornified layer, which temporarilyimpairs the local skin barrier function (de Cid et al., 2009). Wedetected significant upregulation of two group 3 Lce genes (Lce3aand Lce3c) in E18.5 BT skin, which might also reflect a responseto epidermal stress or to impaired barrier function. Alternatively,exposure to the aqueous environment in the uterus might sendsignals to the epidermis that results in upregulation of genesnormally expressed in internal epithelia.
In addition to the protein constituents of the cornified envelope,barrier function in the skin epidermis is also highly dependent onsecretion of specialized lipids and their assembly into lipid lamellae(Houben et al., 2007; Segre, 2003). Examination of microarraydata revealed that the expression of genes encoding enzymesinvolved in the synthesis and processing of lipids and thoseencoding the major protein substrates to which the lipids areattached was normal. The normal or elevated expression of thesegenes implies functional competence to assemble and crosslink thelipid envelope to the outer surface of the corneocytes.
Ets1 expression in differentiating keratinocytes inducesexpression of several genes involved in epidermalproliferation and motilityBT epidermis is thicker than wild-type epidermis and shows moreKi67 and PCNA staining. The increased proliferative response inthe basal layer is most probably secondary to changes in thesuprabasal layers, because the transgene is not expressed in thebasal compartment. One possible mechanism by which inductionof Ets1 might mediate this effect is through stimulation of theexpression of EGF-R ligands such as amphiregulin, epigen, heparin-binding EGF and TGF. These ligands are strongly mitogenic tokeratinocytes (Pastore et al., 2008; Schneider et al., 2008), and arecent report has implicated TGF as a major downstream mediatorof the oncogenic effects of Ets1 in several different human tumorcell lines (Holterman et al., 2010). Furthermore, Ets1 wasdemonstrated to bind to the human TGF gene promoter, indicatingthat TGF is a direct transcriptional target of Ets1 (Holterman etal., 2010).
Ets1 affects the expression of several genes involved in regulatingkeratinocyte homeostasis. Importantly, expression of several genesencoding AP1 family members Fos, Fosl1 (Fra-1), Fosl2 (Fra-2)
Jour
nal o
f Cel
l Sci
ence

and JunB is significantly upregulated in BT epidermis. Expressionof other AP1 genes such as Jun (c-jun), Jund (JunD) or Fosb (FosB)is not significantly changed. AP1 proteins form dimeric transcriptionfactor complexes consisting of Jun homodimers and Fos-Junheterodimers that regulate a variety of target genes. Interestingly,there are many examples of genes that are coordinately regulated byAP1 proteins and Ets family proteins. Indeed, adjacent AP1 and Etsmotifs in promoters can form so-called Ras-responsive elements thatmediate cellular responses to Ras–MAPK signaling (McCarthy etal., 1997; Westwick et al., 1994; Yang et al., 1996). Similar AP1 andEts motifs have been detected within the promoters of Mmp genes,where they play a major role in regulating expression of these genes(Kapila et al., 2009; Nelson et al., 2006; Watabe et al., 1998). SeveralMmp genes demonstrate dramatically increased expression in E18.5BT skin, and we have previously demonstrated upregulation ofmany Mmp genes in epidermis of BT mice induced at an adult age(Nagarajan et al., 2009). This appears to be a direct effect of Ets1 onthese genes because transduction of an Ets1-expressing retrovirusinto a keratinocyte cell line could induce expression of Mmp9 andMmp13, as shown in this study. We have previously shown bychromatin immunoprecipitation (ChIP) assay that Ets1 directly bindsto the promoter of the Mmp13 gene (Nagarajan et al., 2009). Otherlaboratories have demonstrated that Ets1 also directly regulates theMmp3 and Mmp9 genes (Baillat et al., 2006; Wei et al., 2009).Together, the Mmps induced by Ets1 expression function to degradea wide array of extracellular protein substrates, possibly leading todecreased epidermal keratinocyte cohesion and increased keratinocyteproliferation and motility.
Ets1 induction results in an innate immune response inthe skinOne of the most interesting findings of our study was theupregulation of many immunologically relevant target genes in BTepidermis. These include 16 different chemokines and sevendifferent cytokines with diverse biological activities. We haverecently reviewed the role of Ets1 in regulating cytokine andchemokine gene expression, and a great deal of evidence supportsa role for Ets1 in directly regulating expression of these genes bybinding promoter and/or enhancer elements (Russell and Garrett-Sinha, 2010). Expression of the chemokine molecules probablyhelps to recruit immune cells into the skin and, indeed, expressionof Itgam and Itgb2 (which encode the integrins that togetherconstitute the myeloid-specific Mac-1 receptor) is stronglyincreased in BT skin. We demonstrate that CD11b+ (Itgam+)myeloid cells are found in high numbers in BT skin. By contrast,we do not detect increased expression of B or T cell genes inmicroarray analysis and preliminary immunostaining experimentsdid not identify CD3+ T cells or CD19+ B cells in the skin of E18.5BT mice (data not shown). This contrasts with the situation foundin involucrin, envoplakin and periplakin triple-deficient mice,which also have a severe barrier defect, but exhibit recruitment ofCD4+ T cells into the skin (Sevilla et al., 2007).
As with the Mmp genes, Ets1 probably serves as a directregulator of chemokine gene expression. Transduction of an Ets1-expressing retrovirus into mouse keratinocytes induced strongexpression of the chemokines Ccl2, Ccl20 and Cxcl5 in a cell-intrinsic fashion. Previous ChIP studies have confirmed that theCcl2 gene (encoding monocyte chemoattractant protein 1, MCP-1)is a direct target of Etsl (Aoki et al., 2010; Zhan et al., 2005).Furthermore, the Ccl20 gene is a known target of another Etsfactor, ESE-1 (Elf3) (Kwon et al., 2003). Very interestingly, recent
3573Oncogenic effects of Ets1 in skin
studies have strongly linked inflammatory reactions to cancerinitiation and progression. Expression of chemokines and cytokinesby tumor cells is thought to recruit myeloid cells that secretefurther cytokines and chemokines to stimulate tumor cell growth,survival and invasion and to promote angiogenesis (Lazennec andRichmond, 2010).
In addition to chemokines and cytokines, the expression of b-defensins and lysozyme enzymes is strongly upregulated in BTskin. Increased expression of chemokines, cytokines, defensinsand lysozyme and the infiltration of myeloid cells cannot be owingto skin infection because we analyzed E18.5 embryos harvestedfrom the sterile environment of the uterus. However, several studiespoint to induction of an innate immune response in keratinocytesin response to wounding or to an impaired barrier formation (Aberget al., 2008; Zaja-Milatovic and Richmond, 2008). Thus, the mostlikely explanation for the induction of these inflammatory responsesis a keratinocyte response to the impaired barrier function of theskin in BT animals.
Potential involvement of Blimp1 in the Ets1 BT phenotypeWe have previously demonstrated that Ets1 binds to and inhibits thefunction of the transcription factor Blimp1 (John et al., 2008).Blimp1 is highly expressed within differentiating keratinocytes inthe granular layer of the epidermis. One study has demonstrated thatBlimp1 plays an important role in regulating terminal differentiationof epidermal keratinocytes (Magnusdottir et al., 2007), althoughanother study failed to detect alterations in the IFE of Blimp1conditional knockout mice (Horsley et al., 2006). Blimp1 is thoughtto regulate the differentiation of granular layer keratinocytes, and inits absence numerous genes involved in epidermal terminaldifferentiation are altered in their expression. Interestingly, some ofthese genes [such as late cornified envelope genes (Lce1d, Lce1f,Lce1h and Lce1i), small proline-rich protein genes (Sprr1a), EGFligands (Epgn, Areg and Btc) and inflammatory mediators (Il1a andCxcl16)] show similar patterns of upregulation or downregulation inBlimp1 conditional knockout epidermis and Ets1 BT epidermis.Thus, Ets1-dependent suppression of Blimp1 activity mightcontribute to its ability to block terminal differentiation ofkeratinocytes, a possibility that remains to be explored in the future.However, the effects of overexpressing Ets1 in the epidermis aremuch more striking than the effects of deleting Blimp1. Indeed, Ets1BT mice die at birth from severe barrier defects, whereas Blimp1conditional knockout mice do not. Together, these results imply thatthe effects of Ets1 on skin epidermis extend beyond its ability toblock Blimp1 activity and probably involve Ets1-dependent activationof target genes such as those encoding Mmps and chemokines.
ConclusionEts1 regulates a variety of transcriptional programs to alter thedifferentiation, proliferation, migratory and inflammatory propertiesof skin keratinocytes. Many of the effects of Ets1 are probablyowing to its direct binding to target sequences in the promoters orenhancers of genes such as those encoding Mmps and chemokines.It is also possible that Ets1 functions by additional mechanisms aswell. For instance, Ets1 might indirectly regulate the expression ofsome target genes by inducing other transcription factors that canturn these genes on or off.
Our results have major implications for how we view the role ofEts1 in squamous cell tumors. Previous research correlated Ets1expression with increased invasion and metastasis in squamous celltumors, which has been linked to the ability of Ets1 to upregulate
Jour
nal o
f Cel
l Sci
ence

3574 Journal of Cell Science 123 (20)
Mmp genes. Our results suggest that in addition to this importantfunction, Ets1 also serves to keep keratinocytes in an undifferentiatedor partially differentiated state and to promote their proliferation.Furthermore, Ets1 induces an innate immune response in the skin.Given the recent data linking immune activation with promotion ofsquamous cell carcinogenesis (Coussens and Werb, 2002; Vicari andCaux, 2002), this pathway might also be important in neoplastic orpre-neoplastic lesions expressing Ets1. On the whole, Ets1 exhibitsmultiple activities predicted to enhance the malignant properties ofkeratinocytes, suggesting that its function in tumor progression mightbe more complex than previously appreciated.
Materials and MethodsAntibodiesThe following primary antibodies were used for western blotting,immunofluorescence and/or immunohistochemistry: rat monoclonal anti-HA (clone3F10, Roche Applied Sciences), rabbit polyclonal anti-Ets1 (N-276 or C-20, SantaCruz Biotechnology), mouse monoclonal anti-GAPDH (clone 6C5, ChemiconInternational), rabbit polyclonal anti-Ki67 (NCL-Ki67p, Novocastra), rat monoclonalanti-b4 integrin (CD104, clone 346-11A, BD Biosciences), rat monoclonal anti-CD11b (M1/70, BD Biosciences), mouse monoclonal anti-E-cadherin (DECMA1,Santa Cruz Biotechnology), mouse monoclonal anti-P-cadherin (56/P-cadherin, BDBiosciences) and mouse monoclonal anti-PCNA (clone PC10, Dakocytomation). Arabbit polyclonal anti-DNp63 antibody (RR-14, developed in the laboratory of S.S.)was used for detecting p63 (Romano et al., 2006). In addition, several polyclonalrabbit antibodies specific for keratinocyte marker proteins (K5, K6, K10, K14,involucrin, filaggrin and loricrin) were generous gifts from Julie Segre (NationalHuman Genome Research Institute, NIH, Bethesda, MD).
AnimalsThe generation of BT mice carrying the involucrin-tTA transgene and the doxycycline-inducible Ets1 transgenes has previously been described (Nagarajan et al., 2009).Single transgenic mice on a diet lacking doxycycline were mated and checked forcopulatory plugs. Newborn mice and embryos at various gestational ages wereharvested for analysis.
HistologyFor histological analyses, skin samples from various regions of the body were fixedin 4% paraformaldehyde, embedded in paraffin and sectioned for staining withhematoxylin and eosin. For alkaline phosphatase staining, frozen sections of dorsalskin from BT and wild-type mice were air-dried and then stained with the VectorRed kit (Vector Labs) followed by counterstaining with DAPI or hematoxylin.
Whole mount dye penetration assay and extraction of cornified envelopesTo identify skin barrier defects, newborn mice and embryos obtained from timedpregnancies were dehydrated by incubating in methanol, rehydrated briefly in water,stained with 0.1% toluidine blue, rinsed quickly in water and then photographedimmediately. To isolate cornified envelopes from skin, the newborn epidermis wasseparated from the dermis by incubating in Dispase II or 0.1 M EDTA. The epidermiswas then heated in an extraction buffer containing 0.1 M Tris-HCl (pH 8.5), 2%SDS, 20 mM DTT and 5 mM EDTA for 10 minutes at 95°C, and the cornifiedenvelopes collected by centrifugation.
Western blotsWhole tissue lysates were prepared from BT and control mouse dorsal skins.Membranes from western blots were incubated with rat anti-HA, anti-Ets1, anti-GAPDH, anti-K6, anti-filaggrin or anti-loricrin and developed with horseradishperoxidase-conjugated anti-rat, anti-rabbit or anti-mouse secondary antibody. Specificbands were detected by chemiluminescence.
Indirect immunofluorescenceFor immunohistochemistry, 4-m thick skin sections were deparaffinized andsubjected to antigen retrieval by heating in sodium citrate buffer in a microwaveoven. The sections were then stained with specific primary antibodies (anti-HA, anti-Ets1 or anti-Ki67) using the Vectastain ABC kit (Vector Labs) using diaminobenzidineas the enzyme substrate. For immunofluorescence, 5-m thick OCT-embedded freshfrozen sections were fixed in methanol. After blocking, the sections were incubatedwith primary antibodies and then incubated with fluorescent (Alexa Fluor 488, AlexaFluor 568 or FITC) secondary antibodies (anti-rabbit IgG, anti-rat IgG or anti-mouseIgG). Sections were counterstained with DAPI, mounted in 80% glycerol andphotographed with an Axiophot Zeiss microscope.
Gene expression microarray analysisTotal RNA was extracted from E18.5 wild-type and BT embryos using TRIzol(Invitrogen, Carlsbad, CA) and then purified using the RNeasy Mini kit (Qiagen,Valencia, CA). Purified total RNA was analyzed on an Agilent Bioanalyzer 2100
(Agilent Technologies, Santa Clara, CA) to determine the quantity and RNA integrity.RNA was subjected to one round of amplification and labeling to obtain biotinylatedcRNA for hybridization to Affymetrix GeneChip Mouse Genome 430 2.0 Arrays.Two independent sets of biological replicates of each tested condition (wild-type andBT epidermis) were used for analysis.
Scanned microarray images were imported into GeneChip Operating Software(GCOS, Affymetrix, Santa Clara, CA) and checked for quality. Data was thennormalized using the MAS5.0 algorithm in the Affy package of Bioconductor in theR statistical computing environment. Probe sets, whose expression intensities wereclose to background noise across the majority of samples, were filtered out to yield~10,000 genes with significant expression. Comparison of BT and wild-typepopulations identified ~2200 of these genes whose expression was statisticallydifferent (P≤0.05) between samples.
Quantitative RT-PCR analysiscDNA was prepared from the same RNA samples that were used in microarrayanalysis using SuperScript II reverse transcriptase kit (Invitrogen). Primer sets for PCRamplification are given in supplementary material Table S2. PCR cycling was performedusing the following conditions: denaturation at 94°C for 4 minutes, 30 cycles ofamplification with denaturation at 94°C for 20 seconds, annealing at 55°C for 30seconds, extension at 72°C for 30 seconds, and final extension at 72°C for 10 minutes.
Retroviral transductions and RT-PCR assaysRetroviruses in the MIGR1 vector were produced as described before (John et al.,2008) using the Platinum-E packaging cell line. Retroviral supernatants were usedto infect a mouse keratinocyte cell line (mK cells) (Sinha and Fuchs, 2001). At least50% of infected cells were GFP-positive, indicating retroviral transduction. At 48hours post-infection, cells were harvested and RNA was prepared. RNA was reversetranscribed into cDNA and semi-quantitative PCR was performed to analyze theexpression of genes of interest.
These studies were supported by research grants from the AmericanCancer Society (Grant #0705201DDC) and the National Institutes ofDental and Craniofacial Research (Grant #DE016944). We thankKirsten Smalley and Irene Kulik for technical assistance, Steven Gillfor help with the microarray analyses and Julie Segre for providingtransgenic mice carrying the tetracycline-transactivator under thecontrol of the involucrin regulatory elements.
Supplementary material available online athttp://jcs.biologists.org/cgi/content/full/123/20/3566/DC1
ReferencesAberg, K. M., Man, M. Q., Gallo, R. L., Ganz, T., Crumrine, D., Brown, B. E., Choi,
E. H., Kim, D. K., Schroder, J. M., Feingold, K. R. et al. (2008). Co-regulation andinterdependence of the mammalian epidermal permeability and antimicrobial barriers.J. Invest. Dermatol. 128, 917-925.
Akiyama, M., Smith, L. T., Yoneda, K., Holbrook, K. A., Hohl, D. and Shimizu, H.(1999). Periderm cells form cornified cell envelope in their regression process duringhuman epidermal development. J. Invest. Dermatol. 112, 903-909.
Ala-aho, R. and Kahari, V. M. (2005). Collagenases in cancer. Biochimie 87, 273-286.Aoki, T., Kataoka, H., Nishimura, M., Ishibashi, R., Morishita, R. and Miyamoto, S.
(2010). Ets-1 promotes the progression of cerebral aneurysm by inducing the expressionof MCP-1 in vascular smooth muscle cells. Gene Ther. Epub ahead of print.
Arlette, J. P. and Trotter, M. J. (2004). Squamous cell carcinoma in situ of the skin:history, presentation, biology and treatment. Austr. J. Dermatol. 45, 1-9; quiz 10.
Bai, W., Wang, L., Ji, W. and Gao, H. (2009). Expression profiling of supraglotticcarcinoma: PTEN and thrombospondin 2 are associated with inhibition of lymphaticmetastasis. Acta Otolaryngol. 129, 569-574.
Baillat, D., Leprivier, G., Regnier, D., Vintonenko, N., Begue, A., Stehelin, D. andAumercier, M. (2006). Stromelysin-1 expression is activated in vivo by Ets-1 throughpalindromic head-to-head Ets binding sites present in the promoter. Oncogene 25, 5764-5776.
Bhawan, J. (2007). Squamous cell carcinoma in situ in skin: what does it mean? J. Cutan.Pathol. 34, 953-955.
Brown, S. J., Tilli, C. M., Jackson, B., Avilion, A. A., MacLeod, M. C., Maltais, L. J.,Lovering, R. C. and Byrne, C. (2007). Rodent Lce gene clusters; new nomenclature,gene organization, and divergence of human and rodent genes. J. Invest. Dermatol. 127,1782-1786.
Cataisson, C., Ohman, R., Patel, G., Pearson, A., Tsien, M., Jay, S., Wright, L.,Hennings, H. and Yuspa, S. H. (2009). Inducible cutaneous inflammation reveals aprotumorigenic role for keratinocyte CXCR2 in skin carcinogenesis. Cancer Res. 69,319-328.
Cockerell, C. J. (2000). Histopathology of incipient intraepidermal squamous cellcarcinoma (“actinic keratosis”). J. Am. Acad. Dermatol. 42, 11-17.
Coppe, J. P., Patil, C. K., Rodier, F., Sun, Y., Munoz, D. P., Goldstein, J., Nelson, P.S., Desprez, P. Y. and Campisi, J. (2008). Senescence-associated secretory phenotypesreveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor.PLoS Biol. 6, 2853-2868.
Jour
nal o
f Cel
l Sci
ence

Coussens, L. M. and Werb, Z. (2002). Inflammation and cancer. Nature 420, 860-867.Coussens, L. M., Raymond, W. W., Bergers, G., Laig-Webster, M., Behrendtsen, O.,
Werb, Z., Caughey, G. H. and Hanahan, D. (1999). Inflammatory mast cells up-regulate angiogenesis during squamous epithelial carcinogenesis. Genes Dev. 13, 1382-1397.
de Cid, R., Riveira-Munoz, E., Zeeuwen, P. L., Robarge, J., Liao, W., Dannhauser, E.N., Giardina, E., Stuart, P. E., Nair, R., Helms, C. et al. (2009). Deletion of the latecornified envelope LCE3B and LCE3C genes as a susceptibility factor for psoriasis.Nat. Genet. 41, 211-215.
Demehri, S., Turkoz, A. and Kopan, R. (2009). Epidermal Notch1 loss promotes skintumorigenesis by impacting the stromal microenvironment. Cancer Cell 16, 55-66.
Dittmer, J. (2003). The biology of the Ets1 proto-oncogene. Mol. Cancer 2, 29.Eckert, R. L., Sturniolo, M. T., Broome, A. M., Ruse, M. and Rorke, E. A. (2005).
Transglutaminase function in epidermis. J. Invest. Dermatol. 124, 481-492.Endo, K., Shirai, A., Furukawa, M. and Yoshizaki, T. (2006). Prognostic value of cell
motility activation factors in patients with tongue squamous cell carcinoma. Hum.Pathol. 37, 1111-1116.
Fuchs, E. (2007). Scratching the surface of skin development. Nature 445, 834-842.Hahne, J. C., Okuducu, A. F., Sahin, A., Fafeur, V., Kiriakidis, S. and Wernert, N.
(2008). The transcription factor ETS-1: its role in tumour development and strategiesfor its inhibition. Mini Rev. Med. Chem. 8, 1095-1105.
Hardman, M. J., Sisi, P., Banbury, D. N. and Byrne, C. (1998). Patterned acquisitionof skin barrier function during development. Development 125, 1541-1552.
Hitomi, K. (2005). Transglutaminases in skin epidermis. Eur. J. Dermatol. 15, 313-319.Holterman, C. E., Franovic, A., Payette, J. and Lee, S. (2010). ETS-1 oncogenic
activity mediated by transforming growth factor alpha. Cancer Res. 70, 730-740.Horsley, V., O’Carroll, D., Tooze, R., Ohinata, Y., Saitou, M., Obukhanych, T.,
Nussenzweig, M., Tarakhovsky, A. and Fuchs, E. (2006). Blimp1 defines a progenitorpopulation that governs cellular input to the sebaceous gland. Cell 126, 597-609.
Houben, E., De Paepe, K. and Rogiers, V. (2007). A keratinocyte’s course of life. SkinPharmacol. Physiol. 20, 122-132.
Jackson, B., Tilli, C. M., Hardman, M. J., Avilion, A. A., MacLeod, M. C., Ashcroft,G. S. and Byrne, C. (2005). Late cornified envelope family in differentiating epithelia-response to calcium and ultraviolet irradiation. J. Invest. Dermatol. 124, 1062-1070.
Jaubert, J., Patel, S., Cheng, J. and Segre, J. A. (2004). Tetracycline-regulatedtransactivators driven by the involucrin promoter to achieve epidermal conditional geneexpression. J. Invest. Dermatol. 123, 313-318.
John, S. A., Clements, J. L., Russell, L. M. and Garrett-Sinha, L. A. (2008). Ets-1regulates plasma cell differentiation by interfering with the activity of the transcriptionfactor Blimp-1. J. Biol. Chem. 283, 951-962.
Kalinin, A. E., Kajava, A. V. and Steinert, P. M. (2002). Epithelial barrier function:assembly and structural features of the cornified cell envelope. BioEssays 24, 789-800.
Kapila, S., Xie, Y. and Wang, W. (2009). Induction of MMP-1 (collagenase-1) by relaxinin fibrocartilaginous cells requires both the AP-1 and PEA-3 promoter sites. Orthod.Craniofac. Res. 12, 178-186.
Keehn, C. A., Smoller, B. R. and Morgan, M. B. (2004). Ets-1 immunohistochemicalexpression in non-melanoma skin carcinoma. J. Cutan. Pathol. 31, 8-13.
Kerkela, E. and Saarialho-Kere, U. (2003). Matrix metalloproteinases in tumorprogression: focus on basal and squamous cell skin cancer. Exp. Dermatol. 12, 109-125.
Koch, P. J., de Viragh, P. A., Scharer, E., Bundman, D., Longley, M. A., Bickenbach,J., Kawachi, Y., Suga, Y., Zhou, Z., Huber, M. et al. (2000). Lessons from loricrin-deficient mice: compensatory mechanisms maintaining skin barrier function in theabsence of a major cornified envelope protein. J. Cell Biol. 151, 389-400.
Kwon, J. H., Keates, S., Simeonidis, S., Grall, F., Libermann, T. A. and Keates, A. C.(2003). ESE-1, an enterocyte-specific Ets transcription factor, regulates MIP-3alphagene expression in Caco-2 human colonic epithelial cells. J. Biol. Chem. 278, 875-884.
Lazennec, G. and Richmond, A. (2010). Chemokines and chemokine receptors: newinsights into cancer-related inflammation. Trends Mol. Med. 16, 133-144.
Leyvraz, C., Charles, R. P., Rubera, I., Guitard, M., Rotman, S., Breiden, B., Sandhoff,K. and Hummler, E. (2005). The epidermal barrier function is dependent on the serineprotease CAP1/Prss8. J. Cell Biol. 170, 487-496.
Lincoln, D. W., 2nd and Bove, K. (2005). The transcription factor Ets-1 in breast cancer.Front. Biosci. 10, 506-511.
M’Boneko, V. and Merker, H. J. (1988). Development and morphology of the peridermof mouse embryos (days 9-12 of gestation). Acta Anat. (Basel) 133, 325-336.
Magnusdottir, E., Kalachikov, S., Mizukoshi, K., Savitsky, D., Ishida-Yamamoto, A.,Panteleyev, A. A. and Calame, K. (2007). Epidermal terminal differentiation dependson B lymphocyte-induced maturation protein-1. Proc. Natl. Acad. Sci. USA 104, 14988-14993.
McCarthy, S. A., Chen, D., Yang, B. S., Garcia Ramirez, J. J., Cherwinski, H., Chen,X. R., Klagsbrun, M., Hauser, C. A., Ostrowski, M. C. and McMahon, M. (1997).Rapid phosphorylation of Ets-2 accompanies mitogen-activated protein kinase activationand the induction of heparin-binding epidermal growth factor gene expression byoncogenic Raf-1. Mol. Cell. Biol. 17, 2401-2412.
Nagarajan, P., Romano, R. A. and Sinha, S. (2008). Transcriptional control of thedifferentiation program of interfollicular epidermal keratinocytes. Crit. Rev. Eukaryot.Gene Expr. 18, 57-79.
Nagarajan, P., Parikh, N., Garrett-Sinha, L. A. and Sinha, S. (2009). Ets1 inducesdysplastic changes when expressed in terminally-differentiating squamous epidermalcells. PLoS ONE 4, e4179.
Nelson, K. K., Subbaram, S., Connor, K. M., Dasgupta, J., Ha, X. F., Meng, T. C.,Tonks, N. K. and Melendez, J. A. (2006). Redox-dependent matrix metalloproteinase-1 expression is regulated by JNK through Ets and AP-1 promoter motifs. J. Biol. Chem.281, 14100-14110.
3575Oncogenic effects of Ets1 in skin
Ozaki, I., Mizuta, T., Zhao, G., Yotsumoto, H., Hara, T., Kajihara, S., Hisatomi, A.,Sakai, T. and Yamamoto, K. (2000). Involvement of the Ets-1 gene in overexpressionof matrilysin in human hepatocellular carcinoma. Cancer Res. 60, 6519-6525.
Pande, P., Mathur, M., Shukla, N. K. and Ralhan, R. (1999). Ets-1: a plausible markerof invasive potential and lymph node metastasis in human oral squamous cell carcinomas.J. Pathol. 189, 40-45.
Pastore, S., Mascia, F., Mariani, V. and Girolomoni, G. (2008). The epidermal growthfactor receptor system in skin repair and inflammation. J. Invest. Dermatol. 128, 1365-1374.
Presland, R. B. and Jurevic, R. J. (2002). Making sense of the epithelial barrier: whatmolecular biology and genetics tell us about the functions of oral mucosal and epidermaltissues. J. Dent. Educ. 66, 564-574.
Presland, R. B., Boggess, D., Lewis, S. P., Hull, C., Fleckman, P. and Sundberg, J. P.(2000). Loss of normal profilaggrin and filaggrin in flaky tail (ft/ft) mice: an animalmodel for the filaggrin-deficient skin disease ichthyosis vulgaris. J. Invest. Dermatol.115, 1072-1081.
Proksch, E., Brandner, J. M. and Jensen, J. M. (2008). The skin: an indispensablebarrier. Exp. Dermatol. 17, 1063-1072.
Reisdorff, J., En-Nia, A., Stefanidis, I., Floege, J., Lovett, D. H. and Mertens, P. R.(2002). Transcription factor Ets-1 regulates gelatinase a gene expression in mesangialcells. J. Am. Soc. Nephrol. 13, 1568-1578.
Romano, R. A., Birkaya, B. and Sinha, S. (2006). Defining the regulatory elements inthe proximal promoter of DeltaNp63 in keratinocytes: Potential roles for Sp1/Sp3, NF-Y, and p63. J. Invest. Dermatol. 126, 1469-1479.
Russell, L. M. and Garrett-Sinha, L. A. (2010). Transcription factor Ets-1 in cytokineand chemokine gene regulation. Cytokine 51, 217-226.
Saeki, H., Kuwano, H., Kawaguchi, H., Ohno, S. and Sugimachi, K. (2000). Expressionof ets-1 transcription factor is correlated with penetrating tumor progression in patientswith squamous cell carcinoma of the esophagus. Cancer 89, 1670-1676.
Sark, M. W., Fischer, D. F., de Meijer, E., van de Putte, P. and Backendorf, C. (1998).AP-1 and ets transcription factors regulate the expression of the human SPRR1Akeratinocyte terminal differentiation marker. J. Biol. Chem. 273, 24683-24692.
Schneider, M. R., Werner, S., Paus, R. and Wolf, E. (2008). Beyond wavy hairs: theepidermal growth factor receptor and its ligands in skin biology and pathology. Am. J.Pathol. 173, 14-24.
Segre, J. (2003). Complex redundancy to build a simple epidermal permeability barrier.Curr. Opin. Cell Biol. 15, 776-782.
Segre, J. A. (2006). Epidermal barrier formation and recovery in skin disorders. J. Clin.Invest. 116, 1150-1158.
Sevilla, L. M., Nachat, R., Groot, K. R., Klement, J. F., Uitto, J., Djian, P., Maatta,A. and Watt, F. M. (2007). Mice deficient in involucrin, envoplakin, and periplakinhave a defective epidermal barrier. J. Cell Biol. 179, 1599-1612.
Sinha, S. and Fuchs, E. (2001). Identification and dissection of an enhancer controllingepithelial gene expression in skin. Proc. Natl. Acad. Sci. USA 98, 2455-2460.
Squier, C. A. and Kremer, M. J. (2001). Biology of oral mucosa and esophagus. J. Natl.Cancer Inst. Monogr. 2001, 7-15.
Takahashi, H., Ogata, H., Nishigaki, R., Broide, D. H. and Karin, M. (2010). Tobaccosmoke promotes lung tumorigenesis by triggering IKKbeta- and JNK1-dependentinflammation. Cancer Cell 17, 89-97.
Tomic-Canic, M., Komine, M., Freedberg, I. M. and Blumenberg, M. (1998). Epidermalsignal transduction and transcription factor activation in activated keratinocytes. J.Dermatol. Sci. 17, 167-181.
Vairaktaris, E., Papageorgiou, G., Derka, S., Moulavassili, P., Nkenke, E., Kessler, P.,Vassiliou, S., Papakosta, V., Spyridonidou, S., Vylliotis, A. et al. (2007). Expressionof ets-1 is not affected by N-ras or H-ras during oral oncogenesis. J. Cancer Res. Clin.Oncol. 133, 227-233.
Vicari, A. P. and Caux, C. (2002). Chemokines in cancer. Cytokine Growth Factor Rev.13, 143-154.
Watabe, T., Yoshida, K., Shindoh, M., Kaya, M., Fujikawa, K., Sato, H., Seiki, M.,Ishii, S. and Fujinaga, K. (1998). The Ets-1 and Ets-2 transcription factors activate thepromoters for invasion-associated urokinase and collagenase genes in response toepidermal growth factor. Int. J. Cancer 77, 128-137.
Wei, G., Srinivasan, R., Cantemir-Stone, C. Z., Sharma, S. M., Santhanam, R.,Weinstein, M., Muthusamy, N., Man, A. K., Oshima, R. G., Leone, G. et al. (2009).Ets1 and Ets2 are required for endothelial cell survival during embryonic angiogenesis.Blood 114, 1123-1130.
Westermarck, J. and Kahari, V. M. (1999). Regulation of matrix metalloproteinaseexpression in tumor invasion. FASEB J. 13, 781-792.
Westermarck, J., Seth, A. and Kahari, V. M. (1997). Differential regulation of interstitialcollagenase (MMP-1) gene expression by ETS transcription factors. Oncogene 14,2651-2660.
Westwick, J. K., Cox, A. D., Der, C. J., Cobb, M. H., Hibi, M., Karin, M. and Brenner,D. A. (1994). Oncogenic Ras activates c-Jun via a separate pathway from the activationof extracellular signal-regulated kinases. Proc. Natl. Acad. Sci. USA 91, 6030-6034.
Yang, B. S., Hauser, C. A., Henkel, G., Colman, M. S., Van Beveren, C., Stacey, K. J.,Hume, D. A., Maki, R. A. and Ostrowski, M. C. (1996). Ras-mediated phosphorylationof a conserved threonine residue enhances the transactivation activities of c-Ets1 and c-Ets2. Mol. Cell. Biol. 16, 538-547.
Zaja-Milatovic, S. and Richmond, A. (2008). CXC chemokines and their receptors: acase for a significant biological role in cutaneous wound healing. Histol. Histopathol.23, 1399-1407.
Zhan, Y., Brown, C., Maynard, E., Anshelevich, A., Ni, W., Ho, I. C. and Oettgen, P.(2005). Ets-1 is a critical regulator of Ang II-mediated vascular inflammation andremodeling. J. Clin. Invest. 115, 2508-2516.
Jour
nal o
f Cel
l Sci
ence