
Essential Role for Pbx1 in Corneal...
9
Essential Role for Pbx1 in Corneal Morphogenesis Mark J. Murphy, 1,2 Bozena K. Polok, 2,3 Daniel F. Schorderet, 3,4,5 and Michael L. Cleary 1 PURPOSE. The Pbx TALE (three-amino-acid loop extension) ho- meodomain proteins interact with class 1 Hox proteins, which are master regulators of cell fate decisions. This study was performed to elucidate the role of the Pbx1 TALE protein in the corneal epithelium of mice. METHODS. Pbx1 f/f mice were crossed with mice containing Cre recombinase under the control of the K14 promoter. Subse- quently, the eyes of these mice were dissected and prepared for histologic or molecular analysis. RESULTS. Tissue-specific deletion of Pbx1 in the corneal epithe- lium of mice resulted in corneal dystrophy and clouding that was apparent in newborns and progressively worsened with age. Thickening of the cornea epithelium was accompanied by stromal infiltration with atypical basal cells, severe disorgani- zation of stromal collagen matrix, and loss of corneal barrier function. High epithelial cell turnover was associated with perturbed expression of developmental regulators and aber- rant differentiation, suggesting an important function for Pbx1 in determining corneal identity. CONCLUSIONS. These studies establish an essential role of the Pbx1 proto-oncogene in corneal morphogenesis. (Invest Oph- thalmol Vis Sci. 2010;51:795– 803) DOI:10.1167/iovs.08-3327 T he cornea covers the outer surface of the eye, and its refractive and transparent properties are essential for un- impaired vision. Loss of corneal integrity due to disease, injury, or surgical interventions can lead to loss of sight, which affects more than 1.5 million individuals annually, highlighting the importance of understanding corneal integrity. 1 The cornea is composed of three distinct layers. A single cell layer of endothelium separates the inner aqueous humor of the eye chamber from the corneal stroma, which constitutes up to 90% of the corneal thickness. The stroma contains inter- spersed neural crest– derived keratocytes that secrete the highly structured collagen-rich matrix essential for corneal transparency. An external, nonkeratinized, self-renewing epi- thelium of ectodermal origin acts as a barrier to the external environment. It consists of both basal and stratified squamous cells that are separated from the corneal stroma by a basement membrane. 2 Stratification of the cornea occurs in early devel- opment, and postnatally the corneal epithelium exhibits a dynamic homeostasis, turning over approximately every 10 days. 3,4 Defining the mechanisms that orchestrate differentia- tion and self-renewal of corneal epithelial cells will facilitate our understanding of corneal epithelial cell homeostasis. The TALE (three amino acid loop extension) class home- odomain transcription factor Pbx1 was initially described as a proto-oncogene (PBX1, pre-B cell leukemia transcription fac- tor 1 5,6 ) and has subsequently been characterized as a global developmental regulator in mice, zebrafish, 7,8 Caenorhabditis elegans, and Drosophila melanogaster. 9,10 The Pbx TALE pro- teins display unique abilities to interact with class 1 Hox proteins, which are master regulators of cell fate, and orphan homeodomain proteins, which are necessary for development of various tissues and organs. 11–16 Pbx interactions with these homeodomain transcription factors increase their DNA-binding affinity and specificity. Pbx1 also functionally interacts with the bHLH class of transcription factors, which regulate skeletal muscle development, differentiation, and regeneration, thus suggesting that Pbx1 serves an even broader and more impor- tant role as a general transcriptional cofactor integrating trans- duction signals in embryogenesis and organ development. 17–19 In this report, a conditional knockout mouse model was used to specifically inactivate Pbx1 in corneal epithelial cells. This method resulted in alteration of corneal cell fate, high epithelial cell turnover, and marked disruption of the corneal basement membrane with deficient epithelial barrier function. These studies demonstrate an essential role for Pbx1 in mor- phogenesis and maintenance of self-renewing adult tissues of the cornea. MATERIALS AND METHODS Mice K14Cre transgenic mice (Jackson Laboratory, Bar Harbor, ME) and Pbx1 / -knockout mice 20 were intercrossed to generate K14Cre .Pbx1 / mice. Subsequent breeding with Pbx1 f/f mice 16 produced K14Cre .Pbx1 /f or K14Cre .Pbx1 f/f mutants and their con- trol littermates. Mice were genotyped by PCR as previously described 16 and maintained in the Stanford animal facility. Mice aged 17 to 21 days were used for the experiments, except for those on neonatal (P0) mice. All experiments were performed with the approval of and in accordance with Stanford’s Administrative Panel on Laboratory Animal Care and the ARVO Statement for the Use of Animals in Ophthalmic and Visual Research. Anatomy and Histology Eye macroscopic morphology was photographed (model D100; Nikon, Tokyo, Japan). Enucleated whole eyes were washed in phosphate- buffered saline (PBS) and fixed in 4% paraformaldehyde in PBS for 45 minutes. Subsequently, the eyes were washed in PBS and stored at 4°C in 30% sucrose-buffered PBS. They were embedded in antifade Yazulla mounting medium and sectioned at 14-m thickness. From the 1 Department of Pathology, Stanford University School of Medicine, Stanford, California; 3 Institut de Recherche en Ophthalmolo- gie, Sion, Switzerland; the 4 Department of Ophthalmology, University of Lausanne, Lausanne, Switzerland; and 5 Ecole Polytechnique Fede- rale de Lausanne (EPFL), Lausanne, Switzerland. 2 Contributed equally to the work and therefore should be con- sidered equivalent authors. Supported by funds from National Cancer Institute Grant CA90735. Submitted for publication December 17, 2008; revised July 24 and August 25, 2009; accepted September 4, 2009. Disclosure: M.J. Murphy, None; B.K. Polok, None; D.F. Schor- deret, None; M.L. Cleary, None Corresponding author: Michael L. Cleary, Department of Pathol- ogy, Stanford University School of Medicine, Stanford, CA 94305; [email protected]. Cornea Investigative Ophthalmology & Visual Science, February 2010, Vol. 51, No. 2 Copyright © Association for Research in Vision and Ophthalmology 795 Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018
Transcript of Essential Role for Pbx1 in Corneal...

Essential Role for Pbx1 in Corneal Morphogenesis
Mark J. Murphy,1,2 Bozena K. Polok,2,3 Daniel F. Schorderet,3,4,5 and Michael L. Cleary1
PURPOSE. The Pbx TALE (three-amino-acid loop extension) ho-meodomain proteins interact with class 1 Hox proteins, whichare master regulators of cell fate decisions. This study wasperformed to elucidate the role of the Pbx1 TALE protein inthe corneal epithelium of mice.
METHODS. Pbx1f/f mice were crossed with mice containing Crerecombinase under the control of the K14 promoter. Subse-quently, the eyes of these mice were dissected and preparedfor histologic or molecular analysis.
RESULTS. Tissue-specific deletion of Pbx1 in the corneal epithe-lium of mice resulted in corneal dystrophy and clouding thatwas apparent in newborns and progressively worsened withage. Thickening of the cornea epithelium was accompanied bystromal infiltration with atypical basal cells, severe disorgani-zation of stromal collagen matrix, and loss of corneal barrierfunction. High epithelial cell turnover was associated withperturbed expression of developmental regulators and aber-rant differentiation, suggesting an important function for Pbx1in determining corneal identity.
CONCLUSIONS. These studies establish an essential role of thePbx1 proto-oncogene in corneal morphogenesis. (Invest Oph-thalmol Vis Sci. 2010;51:795–803) DOI:10.1167/iovs.08-3327
The cornea covers the outer surface of the eye, and itsrefractive and transparent properties are essential for un-
impaired vision. Loss of corneal integrity due to disease, injury,or surgical interventions can lead to loss of sight, which affectsmore than 1.5 million individuals annually, highlighting theimportance of understanding corneal integrity.1
The cornea is composed of three distinct layers. A singlecell layer of endothelium separates the inner aqueous humor ofthe eye chamber from the corneal stroma, which constitutesup to 90% of the corneal thickness. The stroma contains inter-spersed neural crest–derived keratocytes that secrete thehighly structured collagen-rich matrix essential for cornealtransparency. An external, nonkeratinized, self-renewing epi-thelium of ectodermal origin acts as a barrier to the externalenvironment. It consists of both basal and stratified squamouscells that are separated from the corneal stroma by a basement
membrane.2 Stratification of the cornea occurs in early devel-opment, and postnatally the corneal epithelium exhibits adynamic homeostasis, turning over approximately every 10days.3,4 Defining the mechanisms that orchestrate differentia-tion and self-renewal of corneal epithelial cells will facilitateour understanding of corneal epithelial cell homeostasis.
The TALE (three amino acid loop extension) class home-odomain transcription factor Pbx1 was initially described as aproto-oncogene (PBX1, pre-B cell leukemia transcription fac-tor 15,6) and has subsequently been characterized as a globaldevelopmental regulator in mice, zebrafish,7,8 Caenorhabditiselegans, and Drosophila melanogaster.9,10 The Pbx TALE pro-teins display unique abilities to interact with class 1 Hoxproteins, which are master regulators of cell fate, and orphanhomeodomain proteins, which are necessary for developmentof various tissues and organs.11–16 Pbx interactions with thesehomeodomain transcription factors increase their DNA-bindingaffinity and specificity. Pbx1 also functionally interacts withthe bHLH class of transcription factors, which regulate skeletalmuscle development, differentiation, and regeneration, thussuggesting that Pbx1 serves an even broader and more impor-tant role as a general transcriptional cofactor integrating trans-duction signals in embryogenesis and organ development.17–19
In this report, a conditional knockout mouse model wasused to specifically inactivate Pbx1 in corneal epithelial cells.This method resulted in alteration of corneal cell fate, highepithelial cell turnover, and marked disruption of the cornealbasement membrane with deficient epithelial barrier function.These studies demonstrate an essential role for Pbx1 in mor-phogenesis and maintenance of self-renewing adult tissues ofthe cornea.
MATERIALS AND METHODS
Mice
K14Cre transgenic mice (Jackson Laboratory, Bar Harbor, ME)and Pbx1�/�-knockout mice20 were intercrossed to generateK14Cre�.Pbx1�/� mice. Subsequent breeding with Pbx1f/f mice16
produced K14Cre�.Pbx1�/f or K14Cre�.Pbx1f/f mutants and their con-trol littermates. Mice were genotyped by PCR as previously described16
and maintained in the Stanford animal facility. Mice aged 17 to 21 dayswere used for the experiments, except for those on neonatal (P0)mice. All experiments were performed with the approval of and inaccordance with Stanford’s Administrative Panel on Laboratory AnimalCare and the ARVO Statement for the Use of Animals in Ophthalmicand Visual Research.
Anatomy and Histology
Eye macroscopic morphology was photographed (model D100; Nikon,Tokyo, Japan). Enucleated whole eyes were washed in phosphate-buffered saline (PBS) and fixed in 4% paraformaldehyde in PBS for 45minutes. Subsequently, the eyes were washed in PBS and stored at 4°Cin 30% sucrose-buffered PBS. They were embedded in antifade Yazullamounting medium and sectioned at 14-�m thickness.
From the 1Department of Pathology, Stanford University School ofMedicine, Stanford, California; 3Institut de Recherche en Ophthalmolo-gie, Sion, Switzerland; the 4Department of Ophthalmology, Universityof Lausanne, Lausanne, Switzerland; and 5Ecole Polytechnique Fede-rale de Lausanne (EPFL), Lausanne, Switzerland.
2 Contributed equally to the work and therefore should be con-sidered equivalent authors.
Supported by funds from National Cancer Institute GrantCA90735.
Submitted for publication December 17, 2008; revised July 24 andAugust 25, 2009; accepted September 4, 2009.
Disclosure: M.J. Murphy, None; B.K. Polok, None; D.F. Schor-deret, None; M.L. Cleary, None
Corresponding author: Michael L. Cleary, Department of Pathol-ogy, Stanford University School of Medicine, Stanford, CA 94305;[email protected].
Cornea
Investigative Ophthalmology & Visual Science, February 2010, Vol. 51, No. 2Copyright © Association for Research in Vision and Ophthalmology 795
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018

Immunohistochemistry
The presence of Pbx1b in tissue sections was detected with an anti-Pbx1b antibody21 using an ABC reagent (Vectastain), MOM, and DAB(Vector Laboratories, Burlingame, CA) according to the manufacturer’sinstructions.
For immunochemistry, the sections were blocked for 1 hour atroom temperature (RT) in PBS, 2% NGS, and 0.2% Triton-X and incu-bated overnight at 4°C with the primary antibodies. The sections werewashed three times for 5 minutes each at RT in PBS, and then incu-bated with secondary antibodies (1:2000 dilution) for 1 hour at RT.The slides were counterstained with DAPI, mounted with glycerol/PBS(AS1; Citifluor Ltd., London, UK), and photographed (BX51; Olympus,Tokyo, Japan). Primary antibodies consisted of anti-Notch1 (1:500;15580-100; Abcam, Cambridge, UK), anti-Ki67 (1:500; 49990-100; Ab-cam), anti-decorin (1:10; R&D Systems, Minneapolis, MN), anti-Pax6(1:100; Developmental Studies Hybridoma Bank, Iowa City, IA), anti-K12 (1:500; Santa Cruz Biotechnology, Santa Cruz, CA), anti-K10 (1:500; Santa Cruz), and anti-CD45 (1:300; Ebiosciences, San Diego, CA).Secondary antibodies consisted of anti-mouse (Alexa 488 and 494[Invitrogen-Molecular Probes, Eugene, OR], A21121 and A1105 [Sero-tec, Oxford, UK] and anti-rabbit (Alexa 488 and 494; A11008 andA11012) antibodies.
Cornea Fragility Assay
Corneal fragility was assessed postmortem, as previously described.22
One cornea was brushed in a circular fashion two times with a mi-crosponge soaked in PBS; the contralateral cornea was not disturbed.Subsequently, 0.5 mL of 2% fluorescein in PBS (Sigma, St Louis, MO)was applied for 1 minute to each eye, which was then copiouslywashed with PBS. The eyes were examined immediately afterward andphotographed (D100; Nikon) under a binocular microscope(1500SMZ; Nikon) with a GFP filter.
Electron Microscopy
Samples were fixed in Karnovsky’s fixative (2% glutaraldehyde and 4%paraformaldehyde in 0.1 M sodium cacodylate [pH 7.4]) for 1 hour atRT, then sectioned, postfixed in 1% osmium tetroxide for 1 hour at RT,washed three times with ultrafiltered water, and stained en bloc for 2hours at RT or held at 4°C overnight. Samples were then dehydratedthrough a series of ethanol washes for 15 minutes each at 4°C begin-ning at 50%, then 70%, then 95%, at which point the samples wereallowed to warm to RT, and the wash was changed to 100% ethanoland then to propylene oxide (PO) for 15 minutes. The samples wereinfiltrated with resin (EMbed-812; EMS cat. no. 14120) mixed 1:1 withPO for 2 hours followed by two parts resin and one part PO overnight.The samples were placed in resin for 2 to 4 hours, and then placed intomolds with labels and fresh resin, oriented, and held at 65°C overnight.
The sections (75–90 nm) were picked up on formvar/carbon-coated 75-mesh copper grids, stained for 20 seconds in 1:1 saturated�7.7% uracetate in acetone followed by staining in 0.2% lead citrate for3 to 4 minutes, observed by transmission microscopy (1230 TEM;JEOL. Tokyo, Japan) at 80 kV, and photographed with a digital camera(Multiscan 791; Gatan, Inc., Pleasanton, CA).
Quantitative Real-Time PCR
Eyes were enucleated postmortem, washed profusely in PBS, andenzymatically digested at 4°C for 8 hours in EMEM without calcium(Cambrex Biosciences, Cambridge, UK) containing 15 mg/mL dispaseII (Sigma). The ocular surface epithelial sheet was loosened by gentleshaking in PBS, transferred to RNA extraction reagent (TRIzol; Invitro-gen), and mechanically sheared. RNA was isolated and cDNA synthe-sized by using commercially prepared reagents according to the man-ufacturer’s instructions (Invitrogen), then subjected to real-time PCR(Prism 7700 Detector; Applied Biosystems, Inc. [ABI], Foster City, CA)using probes for Pbx1, cMyc, cyclin D1, p21, p27, p53, Notch1, Pax6,
and Decorin1 (TaqMan; ABI). Expression levels were standardized tothose of �-actin, which served as an internal control. Data wereanalyzed (using SDS ver. 1.9.1 software; SDS, Cary, NC; and Excel,Microsoft, Redmond, WA).
BrdU Labeling
Mice were injected with BrdU intraperitoneally (20 mg/mL in PBS pergram mouse weight) and killed 4 hours later. The eyes were removed,fixed in 4% paraformaldehyde in PBS at RT for 45 minutes, washed inPBS, and held in 30% sucrose-buffered PBS at 4°C before embedding.
Apoptosis Assay
Terminal dUTP nick end-labeling (TUNEL) assays to detect DNA strandbreaks in cell nuclei were performed on frozen sections (10 �m) usingcommercially prepared reagents (Roche, Indianapolis, IN) according tothe manufacturer’s instructions. The sections were counterstainedwith DAPI, and after three washes in PBS, were mounted (AF1; Citi-fluor Ltd) and viewed by fluorescence microscope (BX51; Olympus)equipped with appropriate filters. TUNEL-positive cells were countedin at least three corneas of wild-type (wt) and knockout (ko) mice (foreach cornea, three slides were obtained from the central portion of thecornea).
RESULTS
Efficient Deletion of Pbx1 in the Mouse Cornea
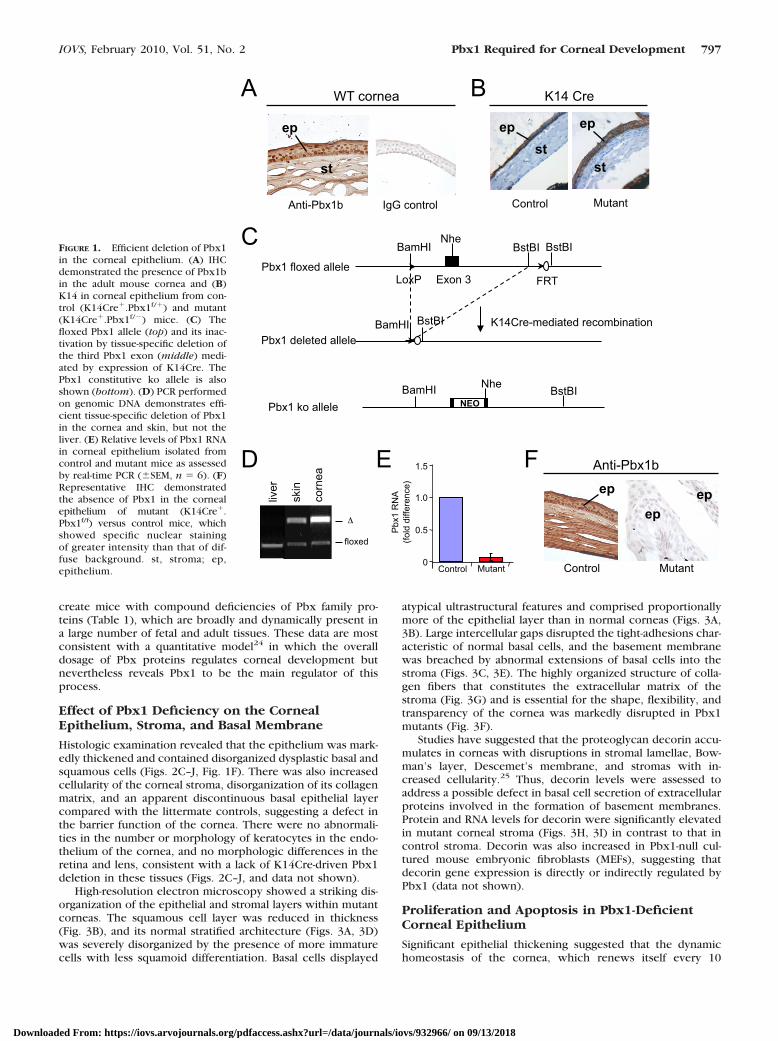
A genetic approach was used to conditionally delete Pbx1 inthe mouse corneal epithelium, which contains abundantPbx1b protein (Fig. 1A), the major isoform of Pbx1 present indeveloping and adult tissues. Cre recombinase was expressedunder control of the keratin 14 (K14) promoter, which isnormally active in basal and occasionally in suprabasal cells ofthe corneal epithelium23 as confirmed by immunohistochem-istry (IHC; Fig. 1B). Mice with a Pbx1 floxed allele (Pbx1f/f)16
were crossed with Pbx1�/� mice20 containing the K14Cretransgene to generate K14Cre:Pbx1f/� (mutant) and littermateK14Cre:Pbx1f/� (control) mice (Fig. 1C), which will hence-forth be referred to as mutant and control, respectively. Effi-cient and specific deletion of Pbx1 occurred in epithelialtissues of the cornea and skin, including the epithelial conjunc-tiva but not nonepithelial tissues (Fig. 1D), and was accompa-nied by a 90% reduction (�100% possibly due to inadvertentcontamination from cells not expressing K14Cre) in Pbx1transcript levels (Fig. 1E) and absence of Pbx1 protein (Fig. 1F)in the corneal epithelium of mutant mice, thus validating ourapproach to targeting Pbx1 in this cell type.
Corneal Clouding Due to Pbx Deficiency
The mutant mice were viable and fertile and opened their eyesat P13.5 to P14.5 similar to the control animals. However, themice displayed a conspicuous corneal clouding phenotype(Fig. 2A) that was apparent in the newborn mice and progres-sively worsened with age (data not shown). In contrast to theouter ocular surface of the control mice, the entire cornea ofthe mutant mice appeared opaque (Fig. 2B). The mutant miceexhibited normal development of skin epithelium and pelage;however, in aged mice (�6 months), hair loss and skin abra-sions developed around areas of stress (whiskers); these obser-vations will be described elsewhere.
The corneal clouding phenotype displayed 88% penetrancein K14Cre:Pbx1f/� mice (10% in K14Cre:Pbx1f/f mice raisedthe possibility that hemizygosity in stromal cells might com-promise mesenchymal/epithelial interaction for maintenanceof corneal transparency), but increased when Pbx1 conditionalmice were crossed with either Pbx2�/� or Pbx3�/� mice to
796 Murphy et al. IOVS, February 2010, Vol. 51, No. 2
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018
create mice with compound deficiencies of Pbx family pro-teins (Table 1), which are broadly and dynamically present ina large number of fetal and adult tissues. These data are mostconsistent with a quantitative model24 in which the overalldosage of Pbx proteins regulates corneal development butnevertheless reveals Pbx1 to be the main regulator of thisprocess.
Effect of Pbx1 Deficiency on the CornealEpithelium, Stroma, and Basal Membrane
Histologic examination revealed that the epithelium was mark-edly thickened and contained disorganized dysplastic basal andsquamous cells (Figs. 2C–J, Fig. 1F). There was also increasedcellularity of the corneal stroma, disorganization of its collagenmatrix, and an apparent discontinuous basal epithelial layercompared with the littermate controls, suggesting a defect inthe barrier function of the cornea. There were no abnormali-ties in the number or morphology of keratocytes in the endo-thelium of the cornea, and no morphologic differences in theretina and lens, consistent with a lack of K14Cre-driven Pbx1deletion in these tissues (Figs. 2C–J, and data not shown).
High-resolution electron microscopy showed a striking dis-organization of the epithelial and stromal layers within mutantcorneas. The squamous cell layer was reduced in thickness(Fig. 3B), and its normal stratified architecture (Figs. 3A, 3D)was severely disorganized by the presence of more immaturecells with less squamoid differentiation. Basal cells displayed
atypical ultrastructural features and comprised proportionallymore of the epithelial layer than in normal corneas (Figs. 3A,3B). Large intercellular gaps disrupted the tight-adhesions char-acteristic of normal basal cells, and the basement membranewas breached by abnormal extensions of basal cells into thestroma (Figs. 3C, 3E). The highly organized structure of colla-gen fibers that constitutes the extracellular matrix of thestroma (Fig. 3G) and is essential for the shape, flexibility, andtransparency of the cornea was markedly disrupted in Pbx1mutants (Fig. 3F).
Studies have suggested that the proteoglycan decorin accu-mulates in corneas with disruptions in stromal lamellae, Bow-man’s layer, Descemet’s membrane, and stromas with in-creased cellularity.25 Thus, decorin levels were assessed toaddress a possible defect in basal cell secretion of extracellularproteins involved in the formation of basement membranes.Protein and RNA levels for decorin were significantly elevatedin mutant corneal stroma (Figs. 3H, 3I) in contrast to that incontrol stroma. Decorin was also increased in Pbx1-null cul-tured mouse embryonic fibroblasts (MEFs), suggesting thatdecorin gene expression is directly or indirectly regulated byPbx1 (data not shown).
Proliferation and Apoptosis in Pbx1-DeficientCorneal Epithelium
Significant epithelial thickening suggested that the dynamichomeostasis of the cornea, which renews itself every 10
A
Pbx1 floxed allele Exon 3
Nhe
LoxP FRT
BstBI BstBI
BstBI
Pbx1 deleted allele
K14Cre-mediated recombination
Nhe BstBI
NEO Pbx1 ko allele
floxed
C
Anti-Pbx1b IgG control
0
0.5
1.0
1.5
Control Mutant
Pbx
1 R
NA
(f
old
diffe
renc
e)
F
Control Mutant
corn
ea
liver
skin
st st st
ep
ep
ep ep
ep ep
Control Mutant
B
Anti-Pbx1b
WT cornea K14 Cre
D E
BamHI
BamHI
BamHI FIGURE 1. Efficient deletion of Pbx1in the corneal epithelium. (A) IHCdemonstrated the presence of Pbx1bin the adult mouse cornea and (B)K14 in corneal epithelium from con-trol (K14Cre�.Pbx1f/�) and mutant(K14Cre�.Pbx1f/�) mice. (C) Thefloxed Pbx1 allele (top) and its inac-tivation by tissue-specific deletion ofthe third Pbx1 exon (middle) medi-ated by expression of K14Cre. ThePbx1 constitutive ko allele is alsoshown (bottom). (D) PCR performedon genomic DNA demonstrates effi-cient tissue-specific deletion of Pbx1in the cornea and skin, but not theliver. (E) Relative levels of Pbx1 RNAin corneal epithelium isolated fromcontrol and mutant mice as assessedby real-time PCR (�SEM, n � 6). (F)Representative IHC demonstratedthe absence of Pbx1 in the cornealepithelium of mutant (K14Cre�.Pbx1f/f) versus control mice, whichshowed specific nuclear stainingof greater intensity than that of dif-fuse background. st, stroma; ep,epithelium.
IOVS, February 2010, Vol. 51, No. 2 Pbx1 Required for Corneal Development 797
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018

days,3,4 was perturbed. Therefore, proliferation status was as-sessed by IHC with an anti-Ki67 antibody that stains cyclingcells. This assay revealed a large increase of basal cell prolifer-ation in mutant corneas (Fig. 4A) with 14% of cells in cyclecompared with 4% in control (Fig. 4B). Quantitative real-timePCR for transcripts of key cell cycle regulators showed in-creased levels (3.3-fold) for c-Myc, which is essential for drivingcells into the cycle,26 and cyclin D1 (3.1-fold), which is nec-essary for cell cycle progression, in corneal epithelium ofmutant mice (Fig. 4C). Conversely, RNA for p21, a cyclindependent kinase inhibitor (CDKI), was decreased 2.9-fold,whereas p27, another CDKI, was not significantly changed (Fig4C). These data collectively support a significant increase incell cycling activity due to loss of Pbx1 in corneal epithelialcells. Furthermore, the rate of apoptosis, which is typically
very low,27 was markedly increased (Fig. 4D) suggesting anabnormally high cell turnover in the mutant epithelium.
Defective Corneal Barrier Function Induced byLoss of Pbx1
The severe morphologic disorganization of the corneal epithe-lium and stroma raised the possibility of a barrier defect inPbx1-mutant corneas, which was addressed in corneal fragilityassays.22 When applied to the cornea, fluorescein dye wasunable to penetrate the epithelial barrier of control mice,either before or after slight mechanical brushing of the cornea(Figs. 2K, 2L). However, Pbx1-deficient corneas were readilypenetrated by the dye even before gentle brushing (Fig. 2M),
FIGURE 2. Loss of Pbx1 severelyperturbed corneal morphogenesis.(A, B) Representative macroscopicimages of in situ (A) and enucleated(B) eyes reveal corneal cloudiness inmutant but not control mice (3weeks old). (C–J) H&E-stained sec-tions show the histologic features ofcorneas visualized by bright-field mi-croscopy at low-power (C–F) andtheir respective high-power (G–J)magnifications. (K–N) Macroscopicimages from a corneal fragility assayof control (K, L) and mutant (M, N)eyes after application of fluoresceinto the corneas of anesthetized mice,with or without prior mechanicalbrushing. st: stroma, ep: epithelium.
TABLE 1. Corneal Phenotypes Caused by Pbx Deficiencies
Genotype Phenotype Viability Penetrance (%)
K14Cre Pbx1f/f Corneal cloudiness Normal 10%K14Cre Pbx1�/f Corneal cloudiness; skin problems in older mice Normal 88%K14Cre Pbx1f/f Pbx2�/� Corneal cloudiness; skin problems in older mice Normal 74%K14Cre Pbx1�/f Pbx2�/� Corneal cloudiness; skin problems in older mice Normal 100%K14Cre Pbx1f/f Pbx2�/� Corneal cloudiness; sunken eyes Decreased 100%K14Cre Pbx1�/f Pbx2�/� Pbx3�/� Corneal cloudiness; sunken eyes Decreased 91%K14Cre Pbx1f/f Pbx2�/� Pbx3�/� Corneal cloudiness when eyes open Decreased 94%K14Cre Pbx1�/f Pbx2�/� Pbx3�/� NA Embryonic lethality NAPbx2�/� Normal Normal NAPbx3�/� Normal Neonatal lethality NA
Mice with the indicated Pbx genotypes were assessed for corneal phenotypes, viability, and phenotype penetrance (n � 20 for all genotypes).NA, not applicable.
798 Murphy et al. IOVS, February 2010, Vol. 51, No. 2
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018

indicative of a barrier defect consistent with the observedmorphologic abnormalities of the corneal epithelium.
Disruption of the integrity of the corneal barrier can lead toan inflammatory response, which was assessed by staining forCD45, a marker for leukocytes and macrophages. Focal staining inthe stroma and epithelium was detected in mutants (Fig. 5A),suggestive of an inflammatory infiltrate in mature mice, but wasnot observed in neonatal mice (Supplementary Fig. S1, http://www.iovs.org/cgi/content/full/51/2/795/DC1). In addition, themutant stroma contained Bigh3, an extracellular keratoepithelinthat participates in cell adhesion and differentiation as a down-stream target of the TGF-� signaling pathway, possibly triggeredby secondary inflammatory processes in mature mice (Fig. 5B).
Effect of Loss of Pbx1 on the Expression ofDevelopmental Regulators in theCorneal Epithelium
Stratification of the cornea begins at postnatal day 4,28 andcorneal-type epithelial differentiation is marked by specificexpression of keratin12 (K12).29 IHC showed that K12 waspresent in the outermost epithelial cell layer in the controlcorneas, but was absent in mutant corneas (Fig. 5C), indicatinga defect in corneal epithelial cell differentiation. Loss of K12 inthe cornea is also a feature of mice deficient in Notch1, a broadregulator of embryonic and postnatal development in multipletissues including the cornea.23,30 After persistent inflammatoryinsult, Notch1 deficiency is associated with a change in their
fate from cornea to skin epithelium,23 which expresses keratin10 (K10); however, Pbx1-mutant corneas lacked K10 (Fig. 5D)as well as skin-associated appendages (Figs. 2C–J) indicatingthat corneal epithelium does not aberrantly differentiate intoskin in the absence of Pbx1. Nevertheless, Pbx1 deficiency wasassociated with perturbations of Notch1 as evidenced by sub-stantially elevated Notch1 protein and RNA levels in the mutantcorneal epithelium in contrast to control corneas, whichlacked Notch1 at this stage of development (Figs. 6A, 6C).Consistent with increased Notch1, mutant corneas also con-tained elevated levels of Pax6 (Figs. 6B, 6D), a transcriptionfactor that is a downstream target of Notch1 signaling31 and isimplicated in the maintenance of corneal epithelium.32
Corneal precursor cells have the capacity to differentiatealong the adipocyte and osteogenic lineages in culture.33 Theabsence of detectable lipid deposits by oil red O staining incorneas from mutant mice suggested a lack of aberrant differ-entiation toward an adipocyte fate (data not shown). However,alizarin red staining revealed large regions of calcification34 inthe stroma of mutant corneas (Fig. 5E), consistent with aber-rant differentiation toward an osteoblast fate in the absence ofPbx1.
DISCUSSION
Our study provided genetic evidence that the Pbx1 transcrip-tion factor plays an important role in controlling the integrity
FIGURE 3. Ultrastructural abnormal-ities of Pbx1-deficient corneas. (A–G)Electron photomicrographs show ul-trastructural features of corneal epi-thelium from control (A, D, G) andmutant (B, C, E, F) mice. (H) IHCdemonstrated increased decorin(green) in the mutant corneal epithe-lium compared with the control cor-nea (DAPI, blue). (I) Relative levelsof decorin RNA in corneal epithe-lium, as detected by quantitative real-time PCR (�SEM, n � 6; *P � 0.031)bc, basal cells; sc, stratified squa-mous cells; st, stroma; bm, bow-man’s membrane. Scale bars: (A, B,D) 10 �m; (C) 4 �m; (E) 2 �m; (G) 1�m; (F) 0.2 �m.
IOVS, February 2010, Vol. 51, No. 2 Pbx1 Required for Corneal Development 799
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018

of the cornea. Epithelia-specific deletion of Pbx1 by Cre recom-binase expressed under control of the keratin14 promoter ledto a progressive eye defect characterized by severe cornealopacity, thickening and degeneration of the corneal epithe-lium, disorganization of the stroma, and perturbation of cor-neal epithelial differentiation.
Corneal Morphology and Pathology
Corneal thickening in Pbx1-mutant mice is most likely due toincreased cellularity and proliferation within the stromal layer,partially caused by disruption of the basal membrane andinfiltration of dysplastic basal and squamous cells into thestroma. No abnormalities were detected in the number andmorphology of keratinocytes. Disruption of the basal barrier,suggested in some studies to be the mouse Bowman’s bar-rier35,36 is known to induce proliferation.37 In Pbx1-mutantcorneas, increased proliferation was also accompanied by ele-vated apoptosis.
Inflammatory changes were also present in mature but notin neonatal mice consistent with loss of corneal barrier func-tion. Although corneas usually lack resident lymphoreticularcells, they are capable of actively participating in an immune
response.38 Other authors suggest the presence of mesenchy-mal-derived cells, acting as antigen-presenting cells that canserve as immune sentinels. In the cornea these include macro-phages and dendritic cells,39 possible candidates in the inflam-matory process marked by the presence of CD45-expressingcells in mutant corneas.
Increased stromal deposition of keratoepithelin, a down-stream target of TGF-� signaling, suggests activation of thispathway in Pbx1-mutant corneas. Corneal opacities are in-duced by transgenic expression of TGF-�, whereas mice withthe mutation of TGF-�2 have thinner corneas.40 Activation ofthe TGF-� pathway may be secondary to inflammation or lossof Pbx1 in mutant animals may lead to de-repression of TGF-�signaling and subsequent corneal thickening. In support of thelatter, alterations of the TGF-� pathway recently reported inPbx1 deficient hematopoietic stem cells16 suggests a functionallink between these pathways.
Corneal Cell Fate
Loss of K12 expression, a specific corneal marker, indicatesaberrant differentiation of cells within Pbx1-deficient corneas.Cranial neural crest cells contribute to corneal development
FIGURE 4. Increased cycling and ap-optotic cells in Pbx1-deficient cor-neal epithelium. (A) IHC showed anincreased number of cells containingKi67 (green) in Pbx1-deficient versuscontrol corneal epithelium (DAPI,blue). (B) Quantification of Ki67-pos-itive cells in the corneal epithelium(�SEM, n � 4). (C) Relative levels ofRNA for the indicated cell cyclegenes in the corneal epithelium asdetected by quantitative real-timePCR (�SEM, n � 5; *P � 0.063, **P �0.041). (D) TUNEL staining showed anincreased number of apoptotic cells(green) in mutant compared with con-trol corneal epithelium (n � 5).
800 Murphy et al. IOVS, February 2010, Vol. 51, No. 2
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018

after their migration into the space between the superficialectoderm and the lens epithelium, lining the inner side of thecorneal epithelium.41
Notch signaling plays an important role in cranial neuralcrest formation and lateral inhibition within neural crest.42
Experiments in chick-quail chimeras demonstrate that neuralcrest grafts derived from different regions may contribute toocular and periocular structures.41 Thus, the loss of cornealepithelial differentiation may be due to misspecification orfailure in the processes related to the upregulation of Notchsignaling in mutant corneas. Since mesenchyme-derived cellsare present in the cornea, it is also possible that their differen-tiation occurs abnormally within keratinocytes and after ele-vated Notch signaling leads to the appearance of adipocytesand osteoblasts. The presence of focal calcified deposits inPbx1-mutant corneas supports the presence of osteoblast lin-eage cells.
Cross-talk between Notch1 and the TGF-� ligand BMP-2promotes osteogenic differentiation.43 Thus, activation of theTGF-� pathway may not only participate in the inflammatoryprocess of Pbx1�/� corneas but also, together with Notchsignaling, in the change in cell fate of the mesenchymal cellswithin the cornea. It remains to be clarified whether othermolecules known to play a role in the molecular control of
osteogenesis are also activated/induced by the absence ofPbx1.
Pax6 is a key developmental regulator capable of inducingectopic eye formation44–46 and is generally essential for mor-phogenesis of mesenchyme-derived tissues in the eye. Highand continuous expression of Pax6 is necessary for the expres-sion of several transcription factors, structural genes, and sig-naling molecules that are essential in the development of cellsthat derive from surface ectoderm and optic cup (e.g., lens andcorneal epithelium) and subsequently for the migration ofneural crest cells into the eye by inductive processes.47,48 Incontrast, low and transient expression of Pax6 serves a cellautonomous role in the differentiation of trabecular meshworkand in formation of corneal endothelium and keratino-cytes.47,49,50 Thus, upregulation of Pax6 expression in Pbx1 komice may drive differentiation of corneal cells to other celltypes.
In summary, our data indicate a role for Pbx1 in cornealmorphogenesis. The absence of Pbx1 leads to corneal opacifi-cation, which reflects high epithelial cell proliferation anddisorganization of the collagen matrix. Deficiency of Pbx1 isaccompanied by upregulation of Notch1 and components of itssignaling pathway. It remains to be demonstrated whetherPbx1 acts directly or indirectly to influence cell fate within the
A
Control
Mutant
CD45DAPI C
Control
Mutant
K12DAPI
Control
DAPI K10
Control-pos
Mutant
B
Control
Mutant
Bigh3DAPI
Mutant
Control Mutant E
D
FIGURE 5. Characterization of cells within corneas. (A) IHC demonstrated CD45 staining in Pbx1-deficient, but not wild-type, corneas. (B) TheBIGH3-encoding protein keratoepithelin was present within deposits in mutant corneas. (C) Pbx1-deficient cornea lacked the specific differenti-ation marker K12. (D) The K10 marker for skin epithelium was absent in wild-type corneas and weakly present in the stroma of Pbx1-deficientcorneas. (E) Alizarin staining within corneal deposits present in mutant mice provided evidence of calcification.
IOVS, February 2010, Vol. 51, No. 2 Pbx1 Required for Corneal Development 801
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018

cornea. In addition, compromised barrier function is associatedwith an acquired inflammatory process in mature but not inneonatal mice, most likely driven by the TGF-� pathway. Thepotential roles of other pathways remain to be elucidated.
Acknowledgments
The authors thank Maria Ambrus, Cita Nicolas, and Natacha Nanchenfor technical assistance.
References
1. Whitcher JP, Srinivasan M, Upadhyay MP. Corneal blindness: aglobal perspective. Bull World Health Org. 2001;79:214–221.
2. Daniels JT, Dart JK, Tuft SJ, Khaw PT. Corneal stem cells in review.Wound Repair Regen. 2001;9:483–494.
3. Haddad A. Renewal of the rabbit corneal epithelium as investigatedby autoradiography after intravitreal injection of 3H-thymidine.Cornea. 2000;19:378–383.
4. Hanna C, Bicknell DS, O’Brien JE. Cell turnover in the adult humaneye. Arch Ophthalmol. 1961;65:695–698.
5. Kamps MP, Murre C, Sun XH, Baltimore D. A new homeobox genecontributes the DNA binding domain of the t(1;19) translocationprotein in pre-B ALL. Cell. 1990;60:547–555.
6. Nourse J, Mellentin JD, Galili N, et al. Chromosomal translocationt(1;19) results in synthesis of a homeobox fusion mRNA that codesfor a potential chimeric transcription factor. Cell. 1990;60:535–545.
7. Popperl H, Rikhof H, Chang H, Haffter P, Kimmel CB, Moens CB.Lazarus is a novel pbx gene that globally mediates hox genefunction in zebrafish. Mol Cell. 2000;6:255–267.
8. Waskiewicz AJ, Rikhof HA, Hernandez RE, Moens CB. ZebrafishMeis functions to stabilize Pbx proteins and regulate hindbrainpatterning. Development. 2001;128:4139–4151.
9. Liu J, Fire A. Overlapping roles of two Hox genes and the exdortholog ceh-20 in diversification of the C. elegans postembryonicmesoderm. Development. 2000;127:5179–5190.
10. Mann RS, Affolter M. Hox proteins meet more partners. Curr OpinGenet Dev. 1998;8:423–429.
11. Capellini TD, Di Giacomo G, Salsi V, et al. Pbx1/Pbx2 requirementfor distal limb patterning is mediated by the hierarchical control ofHox gene spatial distribution and Shh expression. Development.2006;133:2263–2273.
12. Chang CP, Shen WF, Rozenfeld S, Lawrence HJ, Largman C, ClearyML. Pbx proteins display hexapeptide-dependent cooperativeDNA binding with a subset of Hox proteins. Genes Dev. 1995;9:663–674.
13. Peers B, Sharma S, Johnson T, Kamps M, Montminy M. The pan-creatic islet factor STF-1 binds cooperatively with Pbx to a regu-latory element in the somatostatin promoter: importance of theFPWMK motif and of the homeodomain. Mol Cell Biol. 1995;15:7091–7097.
14. Berkes CA, Bergstrom DA, Penn BH, Seaver KJ, Knoepfler PS,Tapscott SJ. Pbx marks genes for activation by MyoD indicating arole for a homeodomain protein in establishing myogenic poten-tial. Mol Cell. 2004;14:465–477.
15. Arata Y, Kouike H, Zhang Y, Herman MA, Okano H, Sawa H. Wntsignaling and a Hox protein cooperatively regulate psa-3/Meis todetermine daughter cell fate after asymmetric cell division in C.elegans. Dev Cell. 2006;11:105–115.
16. Ficara F, Murphy MJ, Lin M, Cleary ML. Pbx1 regulates self-renewalof long-term hematopoietic stem cells by maintaining their quies-cence. Cell Stem Cell. 2008;2:484–496.
17. Moens CB, Selleri L. Hox cofactors in vertebrate development. DevBiol. 2006;291:193–206.
18. Maves L, Waskiewicz AJ, Paul B, et al. Pbx homeodomain proteinsdirect Myod activity to promote fast-muscle differentiation. Devel-opment. 2007;134:3371–3382.
19. Laurent A, Bihan R, Omilli F, Deschamps S, Pellerin I. PBXproteins: much more than Hox cofactors. Int J Dev Biol. 2008;52:9–20.
20. Selleri L, Depew MJ, Jacobs Y, et al. Requirement for Pbx1 inskeletal patterning and programming chondrocyte proliferationand differentiation. Development. 2001;128:3543–3557.
Control
Mutant
Notch 1 DAPI A B
Control
Mutant
Pax 6 DAPI
C D
*
0
20
Control Mutant
Not
ch 1
RN
A
(fol
d di
ffere
nce)
16
12
8
4
**
0
2
Control Mutant
Pax
6 R
NA
(f
old
diffe
renc
e)
1
***FIGURE 6. Developmental regulatorsare abnormally expressed in Pbx1-deficient corneas. (A) IHC shows in-creased levels of Notch 1 (green) inPbx1-deficient corneal epithelium(DAPI, blue). (B) IHC shows in-creased levels of Pax6 in the outerlayer of Pbx1-deficient corneal epi-thelium. (C, D) Quantification ofRNA levels for Notch1 (C) and Pax6(D) RNA in corneal epithelium.(�SEM, n � 6 *P � 0.0017; **P �0.095).
802 Murphy et al. IOVS, February 2010, Vol. 51, No. 2
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018

21. Schnabel CA, Selleri L, Jacobs Y, Warnke R, Cleary ML. Expressionof Pbx1b during mammalian organogenesis. Mech Dev. 2001;100:131–135.
22. Davis J, Duncan M.K, Robison WG Jr, Piatigorsky J. Requirementfor Pax6 in corneal morphogenesis: a role in adhesion. J Cell Sci.2003;116:2157–2167.
23. Vauclair S, Majo F, Durham AD, Ghyselinck NB, Barrandon Y,Radtke F. Corneal epithelial cell fate is maintained during repair byNotch1 signaling via the regulation of vitamin A metabolism. DevCell. 2007;13:242–253.
24. Stankunas K, Shang C, Twu KY, et al. Pbx/Meis deficiencies dem-onstrate multigenetic origins of congenital heart disease. Circ Res.2008;103(7):702–709.
25. Funderburgh JL, Hevelone ND, Roth MR, et al. Decorin and bigly-can of normal and pathologic human corneas. Invest OphthalmolVis Sci. 1998;39:1957–1964.
26. Murphy MJ, Wilson A, Trumpp A. More than just proliferation: Mycfunction in stem cells. Trends Cell Biol. 2005;15:128–137.
27. Ren H, Wilson G. Apoptosis in the corneal epithelium. InvestOphthalmol Vis Sci. 1996;37:1017–1025.
28. Mukhopadhyay M, Gorivodsky M, Shtrom S, et al. Dkk2 plays anessential role in the corneal fate of the ocular surface epithelium.Development. 2006;133:2149–2154.
29. Moyer PD, Kaufman AH, Zhang Z, Kao CW, Spaulding AG, KaoWW. Conjunctival epithelial cells can resurface denuded cornea,but do not transdifferentiate to express cornea-specific keratin 12following removal of limbal epithelium in mouse. Differentiation.1996;60:31–38.
30. Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fatecontrol and signal integration in development. Science. 1999;284:770–776.
31. Koroma BM, Yang JM, Sundin OH. The Pax-6 homeobox gene isexpressed throughout the corneal and conjunctival epithelia. In-vest Ophthalmol Vis Sci. 1997;38:108–120.
32. Sivak JM, Mohan R, Rinehart WB, Xu PX, Maas RL, Fini ME. Pax-6expression and activity are induced in the reepithelializing corneaand control activity of the transcriptional promoter for matrixmetalloproteinase gelatinase B. Dev Biol. 2000;222:41–54.
33. Choong PF, Mok PL, Cheong SK, Then KY. Mesenchymal stromalcell-like characteristics of corneal keratocytes. Cytotherapy. 2007;9:252–258.
34. Pecorella I, McCartney AC, Lucas S, et al. Acquired immunodefi-ciency syndrome and ocular calcification. Cornea. 1996;15(3):305–311.
35. Hayashi S, Osawa T, Tohyama K. Comparative observations oncorneas, with special reference to Bowman’s layer and Descemet’s
membrane in mammals and amphibians. J Morphol. 2002;254:247–258.
36. Labbe A, Liang H, Martin C, Brignole-Baudouin F, Warnet JM,Baudouin C. Comparative anatomy of laboratory animal corneaswith a new-generation high-resolution in vivo confocal micro-scope. Curr Eye Res. 2006;31:501–509.
37. Zieske JD. Expression of cyclin-dependent kinase inhibitors duringcorneal wound repair. Prog Retin Eye Res. 2000;19:257–270.
38. Hamrah P, Dana MR. Corneal antigen-presenting cells. Chem Im-munol Allergy. 2007;92:58–70.
39. Gage PJ, Rhoades W, Prucka SK, Hjalt T. Fate maps of neural crestand mesoderm in the mammalian eye. Invest Ophthalmol Vis Sci.2005;46:4200–4208.
40. Flugel-Koch C, Ohlmann A, Piatigorsky J, Tamm ER. Disruption ofanterior segment development by TGF-beta1 overexpression inthe eyes of transgenic mice. Dev Dyn. 2002;225(2):111–125.
41. Creuzet S, Vincent C, Couly G. Neural crest derivatives in ocularand periocular structures. Int J Dev Biol. 2005;49:161–171.
42. Cornell RA, Eisen JS. Notch in the pathway: the roles of Notchsignaling in neural crest development. Semin Cell Dev Biol. 2005;16:663–672.
43. Nobta M, Tsukazaki T, Shibata Y, et al. Critical regulation of bonemorphogenetic protein-induced osteoblastic differentiation byDelta1/Jagged1-activated Notch1 signaling. J Biol Chem. 2005;280:15842–5848.
44. Ashery-Padan R, Gruss P. Pax6 lights-up the way for eye develop-ment. Curr Opin Cell Biol. 2001;13:706–714.
45. Chow RL, Lang RA. Early eye development in vertebrates. AnnuRev Cell Dev Biol. 2001;17:255–296.
46. Gehring WJ, Ikeo K. Pax 6: mastering eye morphogenesis and eyeevolution. Trends Genet. 1999;15:371–377.
47. Baulmann DC, Ohlmann A, Flugel-Koch C, Goswami S, Cvekl A,Tamm ER. Pax6 heterozygous eyes show defects in chamber angledifferentiation that are associated with a wide spectrum of otheranterior eye segment abnormalities. Mech Dev. 2002;118:3–17.
48. Cvekl A, Tamm ER. Anterior eye development and ocularmesenchyme: new insights from mouse models and human dis-eases. Bioessays. 2004;26:374–386.
49. Collinson JM, Quinn JC, Hill RE, West JD. The roles of Pax6 in thecornea, retina, and olfactory epithelium of the developing mouseembryo. Dev Biol. 2003;255:303–312.
50. Cvekl A, Yang Y, Chauhan BK, Cveklova K. Regulation of geneexpression by Pax6 in ocular cells: a case of tissue-preferredexpression of crystallins in lens. Int J Dev Biol. 2004;48:829–844.
IOVS, February 2010, Vol. 51, No. 2 Pbx1 Required for Corneal Development 803
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932966/ on 09/13/2018


















