
ENZYMATIC DETERMINATION OF TRANSMISSIBLE REPLICATING FACTORS ASSOCIATED WITH MOUSE TUMORS
29
ENZYMATIC DETERMINATION OF TRANSMISSIBLE REPLICATING FACTORS ASSOCIATED WITH MOUSE TUMORS* Vernon Riley Sloan-Kettering Institute for Cancer Research, Rye, N. Y. This report describes the characteristics of a viruslike agent (or agents) found associated with the Cloudman S91 melanoma, the Harding-Passey melanoma, the B-16 mouse melanoma, and about 30 other varieties of transplanted mouse tumors.l.*J Details of the enzyme methods employed in the original detection of these agents are described together with the related procedures for quantita- tive measurement of such agents. Although similar agents have been detected and studied in other tumor-bearing mice by the same techniques, the agent described in this report has been obtained mostly from a single source, namely Cloudman S91 tumor-bearing mice. While the association of these agents with mouse tumors is widespread, both geographically and by tumor type, their significance to the etiology of malig- nancy is uncertain. It would be as equally premature at this writing to attempt to disassociate them from the tumor process as it would be to identify the im- pressive ubiquitous agent-tumor association with tumor induction capability. Irrespective of the outcome of this important question, the ability of these agents to influence tumor and host metabolism and to affect the course of the malignant process as indicated by enhanced host anemia and other parameters will be illustrated. Although these agents are not uniquely associated with melanomas, they are, by perhaps unfortunate necessity, of interest to investigators of various dis- ciplines working with the pigment cell as well as other areas of biology. The following data will indicate a sufficient influence of these agents on fundamental biological properties of the cell to imply that their presence (or absence) in tumor or in host may alter, obscure, or enhance commonly studied metabolic or structural properties of the cell or its tissue whether in vitro, in tissue culture, embryo, egg, or adult host. Experimental Materials and Methods Mice. The experiments described here utilized Ha/ICR Swiss male mice, DBA/2 inbred, and DBA/2 X Swiss F1 hybrids of both sexes. Inbred DBA/2 mice were used for tumor induction with methylcholanthrene (MCA), and the resulting primary tumors were transplanted into DBA/2 X Swiss FI hybrids. In enzyme-agent studies such as reported here, it is essential to use “virus-free” mice that have uniformly normal plasma lactic dehydrogenase (LDH) values, preferably between 100 and 600 units. Reference to, or use of, the term virus- free is intended to imply only freedom from the LDH-elevating agents that can be readily detected by these techniques. Plasma samples. In most instances reported in these experiments, the LDH titer (lactic dehydrogenase activity) refers to that found in the peripheral blood * A portion of these studies was supported by Grant CY-3192 from the National Cancer Institute, Public Health Service, Bethesda, Md. 762
-
Upload
vernon-riley -
Category
Documents
-
view
212 -
download
0
Transcript of ENZYMATIC DETERMINATION OF TRANSMISSIBLE REPLICATING FACTORS ASSOCIATED WITH MOUSE TUMORS

ENZYMATIC DETERMINATION OF TRANSMISSIBLE REPLICATING FACTORS ASSOCIATED WITH
MOUSE TUMORS*
Vernon Riley Sloan-Kettering Institute for Cancer Research, Rye, N . Y .
This report describes the characteristics of a viruslike agent (or agents) found associated with the Cloudman S91 melanoma, the Harding-Passey melanoma, the B-16 mouse melanoma, and about 30 other varieties of transplanted mouse tumors.l.*J Details of the enzyme methods employed in the original detection of these agents are described together with the related procedures for quantita- tive measurement of such agents. Although similar agents have been detected and studied in other tumor-bearing mice by the same techniques, the agent described in this report has been obtained mostly from a single source, namely Cloudman S91 tumor-bearing mice.
While the association of these agents with mouse tumors is widespread, both geographically and by tumor type, their significance to the etiology of malig- nancy is uncertain. It would be as equally premature at this writing to attempt to disassociate them from the tumor process as it would be to identify the im- pressive ubiquitous agent-tumor association with tumor induction capability.
Irrespective of the outcome of this important question, the ability of these agents to influence tumor and host metabolism and to affect the course of the malignant process as indicated by enhanced host anemia and other parameters will be illustrated.
Although these agents are not uniquely associated with melanomas, they are, by perhaps unfortunate necessity, of interest to investigators of various dis- ciplines working with the pigment cell as well as other areas of biology. The following data will indicate a sufficient influence of these agents on fundamental biological properties of the cell to imply that their presence (or absence) in tumor or in host may alter, obscure, or enhance commonly studied metabolic or structural properties of the cell or its tissue whether in vitro, in tissue culture, embryo, egg, or adult host.
Experimental Materials and Methods Mice. The experiments described here utilized Ha/ICR Swiss male mice,
DBA/2 inbred, and DBA/2 X Swiss F1 hybrids of both sexes. Inbred DBA/2 mice were used for tumor induction with methylcholanthrene (MCA), and the resulting primary tumors were transplanted into DBA/2 X Swiss FI hybrids. In enzyme-agent studies such as reported here, it is essential to use “virus-free” mice that have uniformly normal plasma lactic dehydrogenase (LDH) values, preferably between 100 and 600 units. Reference to, or use of, the term virus- free is intended to imply only freedom from the LDH-elevating agents that can be readily detected by these techniques.
Plasma samples. In most instances reported in these experiments, the LDH titer (lactic dehydrogenase activity) refers to that found in the peripheral blood
* A portion of these studies was supported by Grant CY-3192 from the National Cancer Institute, Public Health Service, Bethesda, Md.
762

Riley : Transmissible Replicating Factors 7 63 plasma of the experimental and control mice. The blood was obtained serially during the course of a given experiment by the modified orbital bleeding tech- n i q ~ e . ~ This simple and rapid procedure permits LDH values to be determined daily on the same mice if experimental circumstances require it. The collected samples were centrifuged directly in heparinized, disposable collection tubes (2.8 mm. diam. X 147 mm. long*) and the plasma was transferred from the severed collection tube with a 25-microliter pipette for direct introduction into the enzyme setup tubes. The same procedures were employed for obtaining samples from the plasma of infected mice for agent passage or bioassay.
Enzyme assay. During the course of these experiments the lactic dehy- drogenase (LDH) enzyme procedure was adapted to an automatic recording spectrophotometer (Cary, Model 11). Also, in order to cope with the limited volume of mouse plasma available from a single animal on a daily or other serial basis, the determination was reduced to a semimicro level by reducing all volumes one fourth. The procedure is a modification of those previously
but since the altered details are useful for many DPN or TPN dependent determinations requiring semimicro volumes they are included here.
The enzyme determination is based upon the rate of change in optical density a t 340 mp as DPNH is oxidized to DPN by the plasma LDH in the presence of excess pyruvate. The following reagents are added in the order given to a 13 X 100 mm. test tube: phosphate buffer, 0.6 ml. (0.1 M, @H 7.4); DPNH, 0.05 ml. (Sigma, 2.5 mg./ml. water); and 0.025 ml. of plasma or other enzyme containing solution. The tubes are permitted to stand at room temperature for 10 to 20 min. to exhaust endogenous substrates and to become thermally equilibrated, following which 0.1 ml. of sodium pyruvate (0.5 mg./ml. water) is added to initiate the reaction. This total volume of 0.775 ml. (3.0 m1./4) is shaken immediately following the addition of the pyruvate and is quickly poured into a semimicro optical cuvette with inside dimensions of 3 X 10 X 40 mm.? The cuvette is placed in an appropriate spectrophotometer set at 340 mp, and readings are immediately begun to determine the rate of change in optical density (OD) which, of course, is a direct reflection of the LDH activity.
With the automatic instrument, recordings are made for slightly more than 1 min. which, with the proper adjustments, should give approximately a 4-inch straight line with the slope depending upon the activity of the preparation. The differences between the OD values obtained exactly 1 min. apart, with a linear reaction, are multiplied by 10,000 to give the units reported here. Under the altered conditions of this semimicro method the unit is still defined as that amount of LDH in 1.0 ml. (or 0.25 ml.) of plasma that reduces the optical density a t a rate of O.OOl/min., employing a wave length of 340 mp at room temperature (23"-25O C.). The interrelationship and calculation of the LDH units are made clear by the following definitions:
1 LDH unit = A 0.001 OD/min./ml. of plasma LDH units = A OD/min./l.O ml. plasma
= A OD/min./O.l ml. plasma X 1,000 (3.0 ml. macro procedure) X 10,000 (3.0 ml. macro procedure)
* Scientific Products Co., Flushing, N.Y. t Available through Pyrocell Co. or through Quaracell, Inc., both in New York, N.Y.

764 Annals New York Academy of Sciences = A OD/min./0.25 ml. plasma X 1,000 (0.775 ml. procedure) = A OD/min./0.025 ml. plasma X 10,000 (0.775 ml. procedure) = ODrmin, - OD:””min. X lo4
The semimicro method, employing a total volume of 0.775 ml. and a modified cuvette, gives essentially the same LDH values as the 3.0 ml macro method.1° It is desirable to use a light mask with the narrower cell to prevent the transmit- tance of light through the thick edges of the cuvette. This can be made from a thin strip of black spring steel into which a window measuring 3 X 16 mm. is cut and positioned appropriately by its attachment to the cuvette holder.
If desired, these units may be readily converted to the theoretically preferred basis of substrate utilization12 by multiplying them by an appropriate factor to give micromoles, or millimicromoles, of DPNH oxidized per minute per milli- liter of plasma.
If a manual spectrophotometer is employed, such as the Beckman DU, the same procedure is followed with the exception that it is usually necessary to dilute the plasma before adding i t to the setup tubes since it is difficult to read the instrument accurately every 15 sec. when the plasma enzyme activity is elevated. Such dilutions are also necessary with the automatic recording instrument when the LDH exceeds 20,000 U., since the substrate is exhausted so rapidly that linearity is not ob- tained over an adequate period. Additional dilution factors must, of course, be included in the final calculations.
The manual instrument is set to an initial OD of 0.400, and 4 to 9 readings are taken a t 15- or 30-sec. intervals to establish linearity of the reaction. A suitable 1 min. portion is then utilized for the calculation of LDH activity. I t might be mentioned that at the required wave length of 340 mp either a tung- sten or hydrogen lamp may be used as a light source with either instrument.
Agent assay. The LDH-elevating agent may be studied quantitatively by adaptation of standard infectivity end-point procedures in which the agent- containing material is diluted on an appropriate log basis through a series of decreasing concentrations in which some of the lower ones will fail to produce infectivity as indicated by LDH elevation failure. Tenfold or 100-fold log dilutions are satisfactory and permit the usual ID60 calculations by Reed and Muench or related methods.13J4
Epitomization of Procedures
Ordinarily, a 1 to 10 dilution is appropriate.
(1) Removal of blood sample from tumor-bearing donor (approximately 0.2 ml. by orbital bleeding) +
(2) Centrifuge whole blood in original micro collection tube and separate plasma4 (2,000 rpm, 20 min.).
(3) Injection of plasma into normal recipients for qualitative or quantitative agent assay (any route, usually ip, and any dilution from 0 to 10-lo).
(4) Removal of blood sample, as above, from recipients (any time after 48 hours).
(5) Determine LDH of plasma (spectrophotometric procedure, DPNH and pyruvate a t 340 mp, PH 7.4) for detection of the presence of LDH-elevating agents.
(6) Continue serial passage via plasma, organ, or tumor extracts as preferred.

Riley : Transmissible Replicating Factors 7 65 Are the “Agents” Viruses?
The agents found associated with tumor-bearing mice possess the following
(1) They pass readily through filters (Selas, 03 porosity and 10 mk Milli-
(2) They pass through Visking cellophane dialysis tubing that holds back
(3) They are partially sedimented in the centrifuge at 100,000 g for 2 hours. (4) They are inactivated by 100” C. for 10 min., or 60” C. for 40 min. ( 5 ) They replicate when injected into susceptible animals since they can be
detected in greater quantity than the amount introduced. Additional evi- dence for replication is their ability to pass through an indefinite series of mouse transplant passages in which the starting material is diluted beyond the theo- retical possibility of titer detection and yet the infectivity is found to be un- diminished on terminal testing.’ They canalso be passed for many generations in tissue culture and still infect mice when reinjected.16
(6) They are infectious by contact among mice in the same cage, and may be successfully injected by any standard route including dermal application to unbroken skin.
(7) While their potential disease characteristics are unknown they cause a host “biochemical lesion” by their induced plasma LDH elevation, and under some conditions, a moderate splenomegaly.
Preliminary quantita- tive experiments in which either DNase or RNase was incubated with the virus did not eliminate the infective capability of the agent.* This suggests a protective coating; or, possibly, the absence of vital components sensitive to the nucleases employed.
Although a number of the above characteristics are associated with classical viruses, the minute size and other unusual properties of these agents may place them eventually in a separate category.
LDH-Agent Mechanism The measurable enzyme alterations that occur in the agent infected hosts
manifest themselves by a substantial increase of a glycolytic enzyme, lactic dehydrogenase (LDH), in the peripheral blood of the infected animal. The present indications are that this is a secondary expression of agent replication, or other associated virus activity, and not a direct measurement of the agent itself. I t also appears reasonable to assume that in non-tumor bearing mice at least, some normal tissues or cellular components of the host are being at- tacked by the agent and, as a consequence, the enzymes of the insulted cells are somehow liberated, elicited, or dynamically secreted, and are thus free to accumulate in the blood plasma.
Plasma LDH Correlation with Tumor Growth With most transplanted mouse tumors there is good correlation between the
alterations in plasma LDH and the course of tumor growth. A typical re- sponse is shown in FIGURE 1. This illustrates the summary results of compiling
characteristics:
pore) that simultaneously retain bacteria.
molecules or particles in excess of about 20,000 molecular weight.
(8) Their nucleic acid character is undetermined.
* Deoxyribonuclease and ribonuclease from beef pancreas.

766 Annals New York Academy of Sciences the data of 20 individual experiments. The LDH curve, showing 5 separate phases, is the result of over 700 plasma enzyme determinations on 163 Swiss ICR mice bearing the solid Ehrlich carcinoma. Similar correlations have been obtained with the Cloudman S91 melanoma. I t is now known that irrespective of the impressive correlation of this enzyme parameter with tumor growth and regression, the entire curve is an expression of the activities of a previously unsuspected agent. The details will be discussed under subsequent headings.
TUMOR AGE-DAYS FIGURE 1. Typical five-phase plasma LDH curve associated with transplanted mouse-
tumor implantation and progressive growth. LDH elevation of phase 2 and 3 is due to agent only, while phase 4 represents a synergism between the agent and the tumor.
Extent of Association of LDH-Elevating Agents with Mouse Tumors If a cell-free tumor extract is prepared from any of some 30 different varieties
of transplanted mouse tumors, and this cell-free preparation is injected into normal healthy mice, either sucklings or adults, within 24 to 48 hours an in- crease may be observed in the plasma LDH activity of the recipients. The typical course of this response is shown in FIGURE 2. This plateau of enzyme activity is the same as phase 3 observed in the tumor implanted mice prior to tumor growth. It is also the activity level to which the chemotherapeutically or surgically treated mouse returns following the disappearance of its tumor.16 Once this LDH level is acquired it remains essentially permanent for the life

Riley : Transmissible Replicating Factors
i - 4 - -
- 3 - - L c 3
0 -
0
5 2 - -
m rn P l a s m a L D H A c t i v i t y - f
- Normal controls - "
' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ~ - 40
767 of the animal except, of course, for increases during any tumor growth which may occur.
The same result may be obtained by the injection of extracts of any organ from the melanoma or other tumor-bearing mouse, or more simply, by the in- jection of whole blood or plasma. Any standard inoculation route may be used including dermal application, although for simplicity we have usually employed intraperitoneal injections. Similar materials from a normal healthy mouse, or heated preparations (100' C./10 min.) from tumor mice do not induce such in- creases in plasma LDH activity. However, once the normal recipient has been injected with a preparation from a tumor-bearing mouse, its plasma is then infectious to other normal recipients.
FIGURE 2. Time-course response of plasma lactic dehydrogenase activity (LDH) follow- ing inoculation of normal mice with an agent obtained from any of the various mouse mel- anomas, or other transplanted mouse tumors such as Ehrlich carcinoma. The elevated plateau is the same as phase 3 in FIGURE 1.
TABLES 1, 2, and 3 illustrate the breadth of the association of LDH-elevating agents among various standard transplantable mouse tumors. An assortment of carcinomas is listed in TABLE 1, together with the plasma LDH response of normal mice injected with peripheral blood plasma from the tumor-bearing mice. The agents are not visibly sensitive to a change in mouse strain although they do not readily cross species barriers. Tumors such as the B-16 melanoma and the Cloudman S91 melanoma, which require C57 black and DBA mice for successful tumor transplantation, contain an agent that replicated readily in Swiss and other strains of mice. We have not been successful, however, in de- tecting replication of such agents in rats or in hamsters. I t is perhaps perti- nent to note in TABLE 1 that the various ascites cells that are frequently used for metabolic, chromosomal, and other critical cellular studies are inextricably associated with the LDH-elevating agents. TABLE 2 shows similarly the pres-

768 Annals New York Academy of Sciences ence of agents among experimental sarcomas, including the Crocker sarcoma, S-180, which is widely used in chemotherapy and compound screening studies.
An assortment of experimental leukemias, three mouse melanomas, and mis- cellaneous tumors are listed in TABLE 3, showing the presence of the same or
TABLE 1 TRANSPLANTABLE MOUSE CARCINOMAS ASSOCIATED WITH THE PRESENCE OF A TRANSMISSIBLE
AGENT AS DEMONSTRATED BY PASSAGE OF LDH ELEVATION TO NORMAL MICE
3400-5900 2200-4400 2000-3300 2500-4100 2500-4000
Tumor
4400 3600 2700 3000 3300
Carcinomas Ehrlich carcinoma (solid) Ehrlich carcinoma (ascitic) Nelson ascites Carcinoma 1025 Bashford carcinoma 63 Lewis bladder carcinoma Lewis lung carcinoma Adenocarcinoma EO-771 C3HBA mammary adenocarci-
Miyono adenocarcinoma Spontaneous mammary tumors$$ Carcinoma 755
(Normal controls) (Normal controls) (Normal controls)
noma$
2500-3100 3OOC-3400 2500-3700
260-510
Host'
2800 3200 3100
380
ICR (M) ICR (M) ICR (M) AKR (F) ICR (M) ICR (M) C57BL (F) C57BL (F)
ICR (M) ICR (M) ICR (M) C57BL (F)
ICR (M) C57BL (F) AKR (F)
Day testedt
2 2 4 2 3 5 2 2
6 3 3 3
- - -
Plasma LDH of recipient
Range
200l32400 2400-3300 3900-4900 1500-3000 1700-3000 4200-5400 3200-4200 58W6700
32004600 1600-2200 2300-2900 2350-3000
260-510 260-470 400-560
Average
2300 2800 4200 2200 2200 4500 3700 6400
4Ooo 1700 2600 2600
380 3 10 430
* Five mice per group. tumor-bearing donor strain.
t Number of days elapsing between plasma injection and LDH determination. 1 Known, or presumed virus-induced neoplasms. 8 Example of eight spontaneous tumors with agent.
Mouse strain indicated is normal recipient but not necessarily
These were primary mammary adeno- Other spontaneous tumors from carcinomas arising in Swiss O'Grady females, old breeders.
a different source failed to show the presence of LDH elevating agents.
TABLE 2
AGENT AS DEMONSTRATED BY PASSAGE OF LDH ELEVATION TO NORMAL MICE TRANSPLANTABLE MOUSE SARCOMAS ASSOCIATED WITH THE PRESENCE OF A TRANSMISSIBLE
Tumor
Sarcomas Sarcoma 180 (solid) Sarcoma 180 (ascitic) Sarcoma T-241 Ridgway osteogenic sarcoma Sarcoma MA387 Wagner osteogenic sarcoma Mecca lymphosarcoma Lymphosarcoma P1798
(Normal controls)
Host Day tested'
ICR (M) 4 ICR (M) ICR (M) ICR (M) ICR (MI ICR (Mj ICR (M) ICR (M)
ICR(M) I -
Plasma LDH of recipients
Range I Average
* Number of days elapsing between plasma injection and LDH determination.

Riley : Transmissible Replicating Factors 769 similar transmissible agents. I t may be noted that three of the leukemias listed have known oncogenic agents associated with them. It is uncertain a t this point whether the Friend1' and Moloneyl* agents are themselves capable of LDH elevation or whether this represents the presence of an additional inde- pendent factor. Indirect evidence suggests, but does not establish, the latter possibility.
In an effort to rule out a localized but general contamination of the animals and tumors of our institute we solicited standard tumors from various outside laboratories. The animals were tested immediately upon arrival to avoid con-
TABLE 3 TRANSPLANTABLE MOUSE LEUKEMIAS, MELANOMAS, AND MISCELLANEOUS TUMORS
ASSOCIATED WITH THE PRESENCE OF A TRANSMISSIBLE AGENT AS DEMONSTRATED BY PASSAGE OF LDH ELEVATION TO NORMAL MICE
Tumor
Leukemias Leukemia L-1210 Leukemia L-4946 Friend virus leukemia1 Friend virus leukemia (solid) 1 Moloney leukemia1 EL-4 leukemia (ascites)
Cloudman S91 melanoma Harding-Passey melanoma B-16 melanomalT
Melanomas
Miscellanv ~~
Glioma26 Adrenotropic atT-20 Plasma cell neoplasm 564711
(Normal controls) (Normal controls) (Normal controls)
Host'
ICR (M) ICR (M) ICR (M) ICR (M) ICR (M) ICR (M)
ICR (M) ICR (M) C57BL (M)
ICR (M) ICR (M) C3H (F)
ICR (M) C3H (F) C57BL (M)
Day tested
4 4 5 2 4 3
3 2 3
2 7
11
- - -
Plasma LDH of recipient
Range
3300-7800 27004600 2600-3800 2000-2900 2400-4000 2800-3100
2900-4400 1600-3300 2800-3000
1900-3200 1400-4500 2 100-34OO
260-510 150-500 200600
Average
4600 3400 3200 2400 3000 3000
3700 2400 2900
3100 3000 2800
380 300 350
* Recipient host for agent determination. 1 Known virus induced neoplasms. $ Received from Lloyd Old, Sloan-Kettering Institute for Cancer Research, Rye, N. Y. 7 Received from Frank Moyer, Johns Hopkins University, Baltimore, Md. 11 Received from Ira Pilgrim, University of Utah, Salt Lake City, Utah.
tamination. TABLE 4 illustrates typical results and indicates that the associa- tion of unsuspected LDH-elevating agents with transplantable mouse tumors is widespread and is independent of any single laboratory. A recent publica- tion by British workers also confirms the presence of similar agents in some of the tumor-bearing mice at the Imperial Cancer Research Fund Laboratory in London, England.19
Ejective Routes of Vi'irus Administration In an effort to obtain preliminary information concerning the various routes
by which the S91 associated agent could be effectively introduced into normal mice, the following experiment was performed.
Five S91 melanoma-bearing mice were orbitally bled? The plasma was

770 Annals New York Academy of Sciences separated and diluted 1 to 10 with 0.9 per cent NaCl, and this agentpreparation was then introduced by 7 different routes into 35 healthy mice with pretested normal LDH values.
Injections of 0.1 ml. of virus solution into each of 5 mice per group were made intraperitoneally (ip), intramuscularly (im) in the left thigh, and subcutaneously (sc) under skin of back. The same dose was given orally (PO) by forced intubation, 1 drop was instilled in the nostril (in), 1 drop was placed in the eye (io) without trauma, and 1 or 2 drops were rubbed into the unbroken skin with a wooden tongue depressor. The results are graphically shown in FIGURE 3 .
Three days following the various administrations, all groups had plasma LDH elevated mice with the exception of the intranasal and the intraocular groups. When the mice were retested on day 86, all groups showed a 100 per
TABLE 4 TRANSMISSIBLE AGENT IN VARIOUS SUBLINES OF TUNORS
CARRIED IN SEPARATE LABORATQRIES
Tumor I Plasma LDH’ I Day tested!
Carcinoma 755 Schabel subline Battelle subline Gellhorn subline
Sugiura subline Columbia subline
Burchenal subline Law-Hirschberg subline
Normal C57BL Controls SKI subline, Walker Lab. CRI subline, Columbia
Glioma-26
Leukemia LIZ10
2640 2550 2940
2500 3050
4650 2700
370 350
3 3 3
2 3
4 3
2 3
* Average plasma LDH in recipient mice following injection with plasma from tumor-
t Number of days elapsing between plasma injection and LDH determination. bearing mouse.
cent positive response except for the i.0. group, which had been accidentally destroyed before the second reading was obtained.
From this single qualitative experiment it would appear that there is no striking difference between ip, im, and sc routes of injection as far as early systemic infection is concerned. Infection is possible, but slower, by the oral and nasal routes, and perhaps of greatest interest was the infectivity by simple dermal application. This fact, paired with the observations that the agent is sometimes present in the urine and feces, points up the hazard of inadvertent laboratory contamination of “virus-free” mice through handling or other means of contact transmission.
Migration of Agent from Peritoneal Cavity to Peripheral Blood We had noticed that it was possible to transmit the LDH-elevating “disease”
to normal mice rather quickly following tumor implantation. Since many virus diseases have latent periods of the order of days or weeks during which
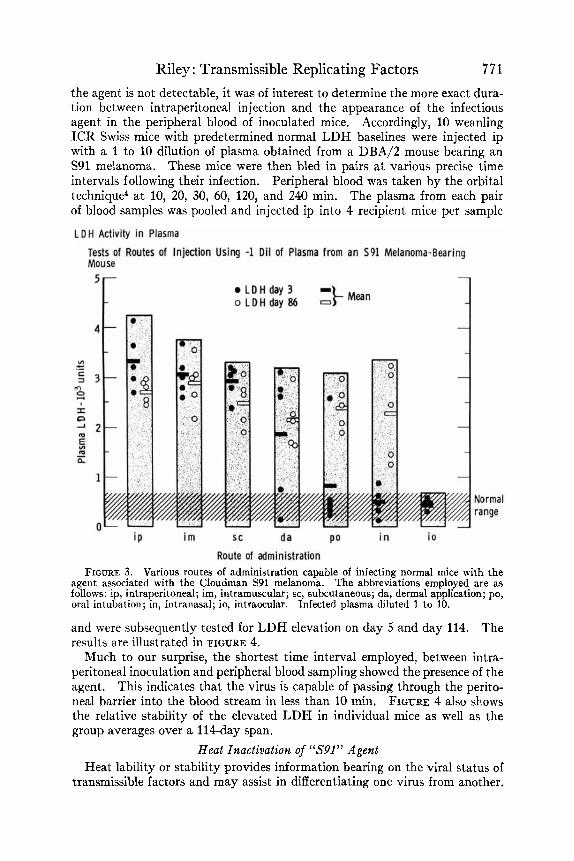
Riley : Transmissible Replicating Factors 771 the agent is not detectable, it was of interest to determine the more exact dura- tion between intraperitoneal injection and the appearance of the infectious agent in the peripheral blood of inoculated mice. Accordingly, 10 weanling ICR Swiss mice with predetermined normal LDH baselines were injected ip with a 1 to 10 dilution of plasma obtained from a DBA/2 mouse bearing an S91 melanoma. These mice were then bled in pairs a t various precise time intervals following their infection. Peripheral blood was taken by the orbital technique4 at 10, 20, 30, 60, 120, and 240 min. The plasma from each pair of blood samples was pooled and injected ip into 4 recipient mice per sample
LDH Activity in Plasma
Route of administration FIGURE 3. Various routes of administration capable of infecting normal mice with the
agent associated with the Cloudman S91 melanoma. The abbreviations employed are as follows: ip, intraperitoneal; im, intramuscular; sc, subcutaneous; da, dermal application; PO, oral intubation; in, intranasal; io, intraocular. Infected plasma diluted 1 to 10.
and were subsequently tested for LDH elevation on day 5 and day 114. The results are illustrated in FIGURE 4.
Much to our surprise, the shortest time interval employed, between intra- peritoneal inoculation and peripheral blood sampling showed the presence of the agent. This indicates that the virus is capable of passing through the perito- neal barrier into the blood stream in less than 10 min. FIGURE 4 also shows the relative stability of the elevated LDH in individual mice as well as the group averages over a 114-day span.
Heat Inactivation of “S91” Agent Heat lability or stability provides information bearing on the viral status of
transmissible factors and may assist in differentiating one virus from another.

7 72 Annals New York Academy of Sciences For example, polyoma is known to be relatively heat stable,20 and thus the inactivation of the agent associated with S91 melanoma in 30 min. at 60" C. tends to remove suspicion of its identity with polyoma. FIGURE 5 illustrates the qualitative inactivation of this agent with time following heating for various periods in a water bath at 60" C. and thermostatically controlled to within 0.5" c.
The virus was obtained by bleeding 6 mice bearing medium size intact S91 melanomas implanted im 42 days previously. The blood was pooled and the
Rapid Transmission of "agent" from Peritoneal Cavity to Peripheral Blood
Donors Interval between i p injection of agent and bleeding FIGURE 4. Demonstration of the ra idity with which the LDH elevating agent from S91
melanoma migrates from the peritonea! cavity to the peripheral blood Agent injected at time zero ip, and tested for by sampling at the retro-orbital plexus a t the various times indi- cated.
agent-containing plasma was separated in a refrigerated centrifuge at 2000 rpm (1000 g) for 20 min. The LDH activity of this plasma was 45,200 U., as ex- pected in a host bearing a tumor associated with an LDH-elevating agent.
The plasma was diluted 1 to 10 with 0.9 per cent NaCl and 1.0 ml. was carefully added to each of 5 thin-walled, wide-mouth vials of approximately 15 ml. capacity. Care was taken not to contaminate the upper part of the vial that was not exposed to submersion in the water bath. Sterile stoppers were added and a 26-gauge needle was inserted through the diaphram to relieve pressure but to prevent evaporation. Four of the vials were then placed in the water bath and held submerged by spring clips.
The zero-time control vial was placed in an ice bath and then removed to the refrigerator until the contents were injected into mice. The same proce-

Riley : Transmissible Replicating Factors 773 dure was followed for each vial as it was removed from the 60" C. bath after exposures of 5, 10, 15, and 30 min.
Five ICR Swiss mice, with pretested normal plasma LDH, were injected ip with 0.1 ml. of each sample, After 4 days the LDH level of their plasma was tested; the results are shown in FIGURE 5. Each point represents one mouse and the heavy crossbars represent the group averages. There was no significant qualitative difference between exposures of 0, 5 , and 10 min., but
Time Required to Inactivate Agent Associated with S91 Melanoma
Heating time - minutes FIGURE 5. Agent survival or inactivation following exposure to a temperature of 60" C .
for various periods. LDH values are those of the normal recipient test mice on day four following injection with the heated preparations.
the results of 30 min. at 60" C. indicate that most of the agent was inactivated. When such data are compared with quantitative results using known dilutions, we would estimate that more than 99.9 per cent of the original agent was de- stroyed under these conditions after 30 min. at 60" C.
Plasma LDH and S91 Melanoma Growth Response to Various Genetic Environments
The data of TABLE 5 illustrate the plasma LDH response of inbred and various F1 hybrid mouse strain combinations following their challenge with an

7 74 Annals New York Academy of Sciences intramuscular injection of 0.1 ml. of a 10 per cent suspension of Cloudman S91 melanoma. The tumor originated in a DBA strain mouse30 and has been carried by most laboratories in this inbred line or in a suitable F1 hybrid. We have employed Swiss X DBA/2 F1 mice for our S91 passage and for most experimental purposes.
This experiment indicates that the growth rate and per cent of tumor takes is approximately the same between DBA/2 inbred stock and the DBA/2 F1 hybrid with Swiss females, whereas the F1 combination employing C57BL/6
TABLE 5 PLASMA LACTIC DEHMROGENASE (LDH) RESPONSE IN VARIOUS MOUSE STRAIN
COMBINATIONS FOLLOWING INTRAMUSCULAR IMPLANTATION OF THE CLOUDMAN S91 MELANOMA
Swiss (random-bred)
DBA/2 (inbred)
DBA/2 (inbred)
Mice
F
M
F
1 Sex
C57BL/6 X DBA/2S
C57BL/6 X DBA/2
SWISS X DBA/2
SWISS X DBA/2
SWISS X C57BL/6
SWISS X C57BL/6
M
F
M I
F
M
F
Days: Post-tumor Implantation
0 1 18 1 35 1 41 1 60
- 550 o / w 500
0/10 500
0/10
0/10
0/10
0/10
0/10
360
300
400
600
2$!g 2,9009 0/10
Plasma Lactic Dehydrogenase' -- 4,200 0/10
6,500 6/10
6,000 5/10
4,200
3,900 0/10
0/10
5,500 8/10 5,300 7/10
4,100 3/10
4,600 0/10
- 0/10
25,000 7/10
22,000 8/9 -
3/10 -
1/10
18,200 7/10
30,100 9/9 -
5/10
O/ lO -
- 0/10
28,000 4/4
35,000 6/7
5/10
1/10
21,500 6/6
28,200 9/9
-
-
- 5/10 -
2/10
4,000 0/8
(dead) O/O
(dead) o/o
10,200 7/9 9,300 3/10
47,500 2/2
6/6 41,900
16,400 9/10 6,200 4/10
* Average LDH of all mice with and without tumors as indicated by denominator. t Number of tumors/number of surviving mice. $ F, mice of strains indicated. # Recipient mice contaminated with an unknown LDH elevating agent.
The female is listed first.
X DBA/2 yielded a longer latent period, a lower incidence, and slower growing tumors. This is reflected in the host plasma LDH as well as by the other parameters.
The unexpected growth of the tumor in F1 mice in which the parent DBA strain was not present indicates that this tumor is no longer completely strain specific. Although there were no tumor takes in Swiss female mice and no elevation of plasma LDH beyond that imposed by the agent present in this tumor, 90 per cent tumors were obtained in male F1 (Swiss X C57BL/6) and 40 per cent in similar FI females. The slower growth rate in this F1 combina- tion was also reflected by the lower terminal LDH.

Riley : Transmissible Replicating Factors 775 Throughout this experiment it was noted that the S91 melanoma grew faster,
had a greater percentage of takes, and induced higher plasma LDH in the male groups as compared with the analogous females.
The high baseline LDH values of the normal F1 Swiss X C57BL/6 groups are probably not a function of the genetic combination, but illustrate a contami- nation of unknown origin sometimes observed in mice received from various breeders. Such an infection of normal mice can also be acquired in the labora- tory mouse room through contact infection, This is discussed in greater de- tail with the experiments demonstrating the infectious character of the agents.
“Natz4ral” Transmission of Agent from 5’91 Melanoma by Contact
The plasma LDH activity of 10 HA/ICR Swiss male mice of approximately 5 weeks of age was determined and found to be within normal limits. In our experience, this establishes their freedom from infection with LDH-elevating viruses. The mice were numbered 1 through 10 by a standard dye-numbering code in distinction from the contaminating hazard of ear punching or toe clip- ping.
Five of the mice, those bearing the odd numbers, were injected ip with a 0.1-ml. dose of a 1 to 10 dilution of plasma obtained from a mouse bearing an S91 melanoma. All 10 mice were housed in a single stainless steel mouse cage suitable for long term observation of this number of animals. The maintenance of these mice followed the standard procedures for our laboratory. The bed- ding consisted of clean, nonsterilized shavings and was changed 3 times a week. The food pellets were placed in the bottom of the cage, and the unused portion was discarded with the soiled bedding. A common water bottle, fitted with a stainless steel drinking tube, was changed once a week, and the tap water was replenished daily. Ventilation was obtained through the quarter-inch wire mesh of the cage top. Room temperature was maintained at 72” F., and the humidity was controlled and maintained at approximately 40 per cent.
The plasma LDH was determined periodically as indicated in FIGURE 6. The LDH of the inoculated mice was elevated tenfold within 48 hours as ex- pected, while the normal inhabitants maintained a normal level for several weeks. The first noninjected mouse to show an LDH elevation was observed on day 26 of contact. By day 47 all cage inhabitants had approximately the same tenfold increase ix LDH indicating the acquirement of infection by the 5 noninjected cage mates. The average plasma LDH values of the 2 groups of mice and the course of transmission of the infection are shown in FIGURE 6.
Plasma LDH Response to Various Concentrations of Agent
Injection of normal mice with plasma from tumor-bearing or other animals carrying the LDH elevating agents will induce the same apparent qualitative enzyme effect irrespective of the dose, ranging from zero dilution to more than
However, by using adequate numbers of recipient mice, ap- propriate dilutions, and proper LDH determinations timewise, i t is not difficult to demonstrate a quantitative dose-response curve to an agent such as that found in the S91 melanoma and other mouse neoplasms. This dose response is expressed by the relative LDH activities a t a given time period following virus
dilution.

776 inoculation. FIGURE 7 illustrates the quantitative LDH host response to the S91 associated agent on postinjection day 0, 1, 2, and 3.
Various concentrations of the virus were prepared by diluting plasma ob- tained from a tumor-bearing mouse with 0.9 per cent NaCl solution. Five tenfold log dilutions were injected ip, as a 0.1-ml. dose, into 10 ICR Swiss recipients for each virus dilution. These groups of 10 were then divided into two groups of 5 mice each for bleeding on alternate days in order to have a daily LDH record. Thus each experimental point in FIGURE 7 represents the average plasma LDH of 5 mice, and the odd and even days plotted are separate but analogous groups. Since this experiment was done, we have found that
"Natural"7ransmission of Agent from infected to Normal Mice in Same Cage. (Injected Agent from Plasma of .s91 Melanoma-Bearing Mouse)
Annals New York Academy of Sciences
1 ' ' ' ' 1 ' ' ' ' 1 ' ' 1 8 1 ' " '
Days
FIGURE 6. Infection of normal mice through natural contact with agents-bearing ani mals in the same cage.
the LDH level is not altered by daily bleeding, at least for 5 successive days, so that the disadvantage of dividing experimental groups is not necessary.
This experiment demonstrates the importance of the time element in detect- ing a quantitative plasmaLDH response to agent dose. It is seen in FIGURE 7 that the 42-hour response is more reflective of the original injected agent dose than either the earlier or later response period. Agent preparations from dif- ferent tumor sources, or with variable virus titers, may yield their optimum dose-response curves at various postinjection periods. I t may also be noted that a 100,000-fold dilution is not sufficient to reduce the agent to its negative end point. Under appropriate conditions, which will be detailed elsewhere, other agent preparations have been found to be still infectious after being diluted 1,000-billionfold (10l2 ID60doses per 0.1 ml. of plasma). I t has also been observed in such experiments that the persisting elevated plasma titer of this

Riley : Transmissible Replicating Factors 777 giycolytic enzyme may be partially dependent upon the initial agent dose in- jected.
For quantitative virus assays it is usually simpler to employ end-point titra- tion followed by standard IDG0 ca lc~ la t ions ,~~J~ rather than to use the above quantitative plasma LDH response.
Spleen Enlargement with Tumors or with Associated Agent The phenomenon of splenomegaly associated with mouse cancer is suffi-
ciently well known to be generally ignored as a common and mysterious con-
P l a s m a L a c t i c D e h y d r o g e n a s e A c t i v i t y o n V a r i o u s D a y s F o l l o w i n g I P I n j e c t i o n o f Log D o s e s of a n A g e n t O b t a i n e d f r o m S 9 1 M o u s e M e l a n o m a
3
2 v)
c 3
c .-
m E:
n
, I
fu 5
" 1 E
m - n a,
a, > a
0
'2 1 1 I I I
Day2 (42 hrs)
Dav 1 (18 hrs)
Day0 (-1 hr)
-4 -3 -2 -1 I 1
Vi rus Dose - log dilution FIGURE 7. Quantitative LDH response to log dose of agent on day 2 in contrast to ab-
sence of a differential response at an earlier or later period.

7 78 Annals New York Academy of Sciences comitant of the malignant process. When we noted that large spleens occurred under some circumstances following the injection of cell-free preparations from tumors, or other tissue extracts from tumor-bearing mice, our interest in spleen enlargement and its possible reflection of virus activity was enhanced. FIGURE 8 illustrates the enlargement of spleens of mice infected with an agent obtained from spleen extracts of S91-bearing mice. Also shown is the absence of en- largement in similar untreated control mice or those injected with analogous normal spleen extract. Heated spleen extract (100' C./10 min.) from tumor mice also failed to induce splenomegaly.
FIGURE 8. Moderate spleen enlargement found in mice infected with agent from S91 melanoma-bearing mice, and comparison with the plasma LDH elevation. The mice were injected with spleen extract from normal and tumor-bearing animals but the same results are obtained with analogous plasmas.
TABLE 6 provides the experimental details of various conditions in the groups tested in this experiment, and indicates that agent-induced splenomegaly is essentially insensitive to mouse strain, sex, age, and duration of injection within the limits shown. Additional experiments have shown that agents from other tumors, such as the Ehrlich carcinoma, have much less ability to induce spleen- enlarging effects. This is suggestive evidence that the agents may differ in character depending upon the tumor source. Correlation between the agent- induced plasma LDH elevation and splenomegaly may also be noted in both FIGURE 8 and TABLE 6. Additive influence of Bartonella is also under study. FIGURE 9 illustrates a dose-response experiment in which the agent from an
S91 melanoma host was injected a t various log dilutions and by two separate

Riley : Transmissible Replicating Factors 779
TA
BL
E
6 C
OM
PAR
ISO
N
OF
PLA
SMA
LD
H A
ND
SP
LEEN
WEI
GH
TS OF
V
AR
IOU
S STR
AIN
S, SE
X AN
D A
GE OF M
ICE
FOL
LO
WIN
G
INJE
CT
ION
OF
SPLE
EN E
XT
RA
CT
FR
OM
N
OR
MA
L OR
S9
1 M
ELA
NO
MA
-BEA
RIN
G
AN
IMA
LS
Stra
in*
Sex
Age
whe
n in
ject
ed
Dur
atio
n (d
ays)
j N
umbe
r of
mic
e A
v. p
lasm
a L
DH
G
roup
av.
(L
DH
)
I N
orm
al c
ontr
ols
Inje
cted
wit
h no
rmal
spl
een
I D
BA
/FI
Swis
s D
BA
/F1
Swiss
Sw
iss
Swiss
F
M
F
M
M
/F
M/F
9
wks
5
wks
9
wks
5
wks
10
day
s 1 da
y 68
13
8 68
13
8 68
68
5
4 5
5 4
11
700
400
300
350
700
550
(550
) (4
75)
DB
A/F
1 F
9
wks
68
10
3 1
00
~~
~~
0.12
4 I
0.14
8 I
0.10
0 I
0.090
(0.1
32)
(0.1
16)
Av.
spl
een
wt. (gm.)
Gro
up a
v. (
sple
en w
t.)
Swiss
sw
iss
M
M/F
5
wks
10
day
s 13
8 68
10
4
3300
35
00
Inje
cted
wit
h sp
leen
from
S91
mou
se
' (3
300)
'
0.24
4 I
0.33
7 I
0.28
0 (0
.287
)
* DB
A/F
l ar
e F1
hyb
red
of Sw
iss X
DB
A/2
. Sw
iss a
re H
A/I
CR
Sw
iss m
ice.
+ T
ime
from
inoc
ulat
ion
with
spl
een
extr
act
until
spl
eens
wei
ghed
at s
acrif
ice.
T
he c
ontro
ls w
ere
anal
ogou
s bu
t with
no
inje
ctio
n.
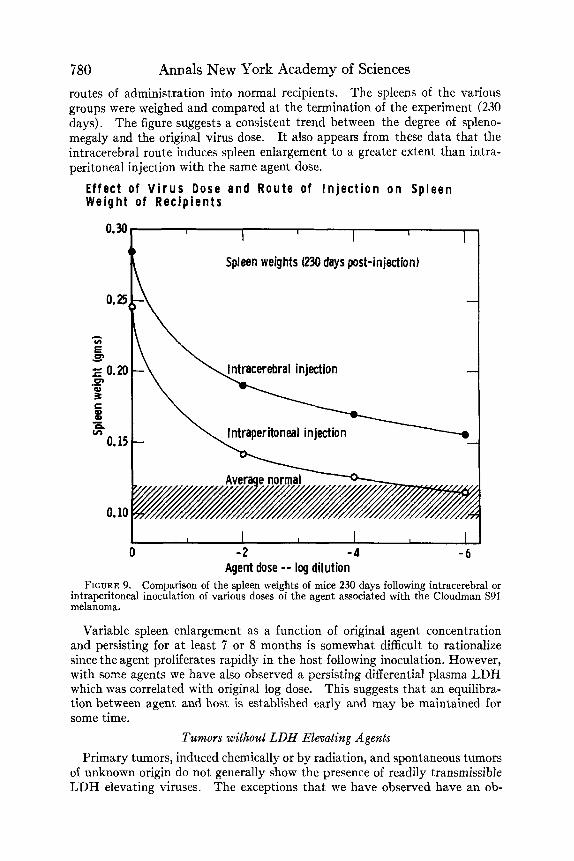
780 Annals New York Academy of Sciences routes of administration into normal recipients. The spleens of the various groups were weighed and compared at the termination of the experiment (230 days). The figure suggests a consistent trend between the degree of spleno- megaly and the original virus dose. It also appears from these data that the intracerebral route induces spleen enlargement to a greater extent than intra- peritoneal injection with the same agent dose.
Effect of V i r u s Dose and Route of I n j e c t i o n o n Spleen Weight of Recip ients
Spleen weights (230 days post-injection)
-
-
I ntraper itoneal injection n
0 - 2 -4 Agent dose -- log dilution
FIGURE 9. Comparison of the spleen weights of mice 230 days following intracerebral or intraperitoneal inoculation of various doses of the agent associated with the Cloudman S91 melanoma.
Variable spleen enlargement as a function of original agent concentration and persisting for a t least 7 or 8 months is somewhat difficult to rationalize since the agent proliferates rapidly in the host following inoculation. However, with some agents we have also observed a persisting differential plasma LDH which was correlated with original log dose. This suggests that an equilibra- tion between agent and host is established early and may be maintained for some time.
Tumors without LDH EEevating Agents Primary tumors, induced chemically or by radiation, and spontaneous tumors
of unknown origin do not generally show the presence of readily transmissible LDH elevating viruses. The exceptions that we have observed have an ob-
Riley : Transmissible Replicating Factors 78 1 scure significance and will not be discussed at this time. In addition to the above primary mouse tumors, we have been unable to detect agents by these techniques in long transplanted tumors of rats or hamsters. At present it is an open question of whether there are actually no such viral agents associated with their tumors, or whether a different enzyme parameter may be required to detect comparable agents in these species.
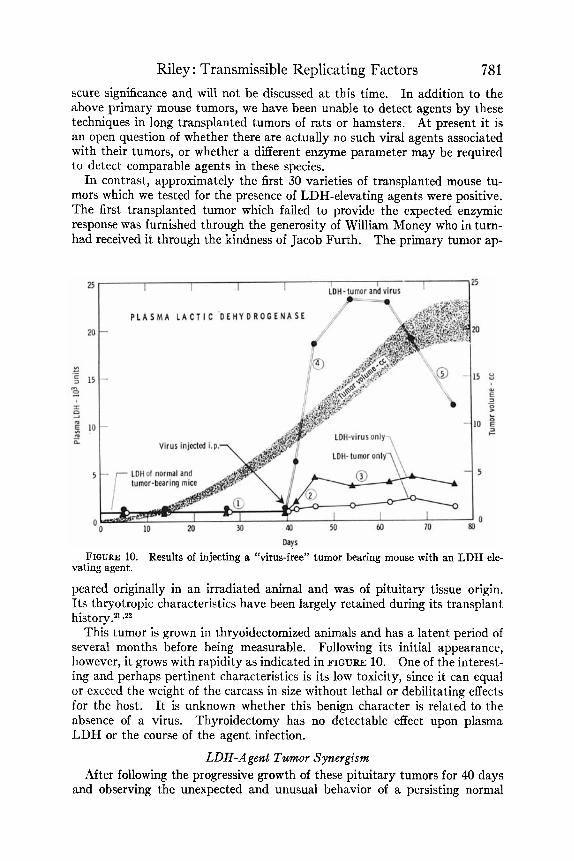
In contrast, approximately the first 30 varieties of transplanted mouse tu- mors which we tested for the presence of LDH-elevating agents were positive. The first transplanted tumor which failed to provide the expected enzymic response was furnished through the generosity of William Money who in turn- had received it through the kindness of Jacob Furth. The primary tumor ap-
Days FIGURE 10. Results of injecting a “virus-free” tumor bearing mouse with an LDH ele-
vating agent.
peared originally in an irradiated animal and was of pituitary tissue origin. Its thryotropic characteristics have been largely retained during its transplant history.2l zz2
This tumor is grown in thryoidectomized animals and has a latent period of several months before being measurable. Following its initial appearance, however, it grows with rapidity as indicated in FIGURE 10. One of the interest- ing and perhaps pertinent characteristics is its low toxicity, since it can equal or exceed the weight of the carcass in size without lethal or debilitating effects for the host. I t is unknown whether this benign character is related to the absence of a virus. Thyroidectomy has no detectable effect upon plasma LDH or the course of the agent infection.
LDB-Agent Tzcmor Synergism After following the progressive growth of these pituitary tumors for 40 days
and observing the unexpected and unusual behavior of a persisting normal

782 Annals New York Academy of Sciences plasma LDH, we intervened by adding an extraneous agent from an Ehrlich carcinoma-bearing mouse to this nonconforming neoplastic system. The re- sults are shown in FIGURE 10. Following the intraperitoneal injection of the agent there was approximately a 100-fold elevation in plasma LDH activity during the succeeding 10 days. However, during the same period there was no appreciable LDH rise in the control, “virus-free,” tumor-bearing mice although their tumors were continuing to enlarge a t the same rate. Only the usual moderate “phase 3” elevation was seen in comparable normal animals which had received the agent only.
As the experiment proceeded, it was interesting to observe the familiar 5-phases of plasma LDH activity assert themselves and thus further delineate the influence that these agents were undoubtedly exerting in the other “stand- ard” transplanted mouse tumors. This synergistic behavior involving tu- mor, agent, and host, as expressed by an elevated enzyme, has been confirmed in other similar experiments and also by employing somewhat different experi- mental designs wherein the agent was added during the chemical induction of tumors. See, for example, FIGURE 11.
Chemically Induced Tumors Since virus-free tumors seemed to be a necessary prerequisite for the study
of the direct influence of these agents on tumors, and on the tumor-bearing host, experiments were undertaken as follows. MCA (20-methylcholanthrene) was injected intramuscularly in DBA/2 inbred mice. Some of the groups received in addition an intraperitoneal inoculation of the LDH-elevating agent associated with the Cloudman S91 melanoma. Under the conditions of these experiments, no virus effect was observed on the latent period of the primary tumors or on their growth rate. However, the plasma LDH of the primary tumor-bearing hosts harboring the added agent showed the same synergistic elevation that had been found with transplanted tumors associated with the agent.
Upon subsequent parallel transplantation of the MCA induced tumors, with and without the added virus, a number of differences was observed. FIGURES 11 and 12 and TABLE 7 epitomize the data obtained. In FIGURE 11, there are shown three different host parameters with and without the agent. These tumors were in their first transplant generation and had been implanted in DBA/2 X Swiss FI hybrids. The comparative tumor volumes, plasma LDH, and blood hematocrit (packed RBC volume) are shown. The biological sig- nificance of the difference in tumor volume is uncertain but similar differences were obtained during the 6 subsequent passages of the 2 tumor lines. The plasma LDH was strikingly different between the 2 tumors and reached a maximum differential of about 100-fold, that is, from 250 to 25,000 LDH units. The usual terminal LDH decrease, which has been designated as “phase 5” of the tumor-virus LDH curve, may also be seen in FIGURE 11. This is thought to be a reflection of decrease in viable elements of the tumor that is not ac- curately reflected in 3-dimensional external caliper measurements.
Tumor-Agent Effect on Host Anemia In these studies,
the packed red cell volume (hematocrit), RBC count, and hemoglobin were Anemia associated with cancer is a common phenomenon.
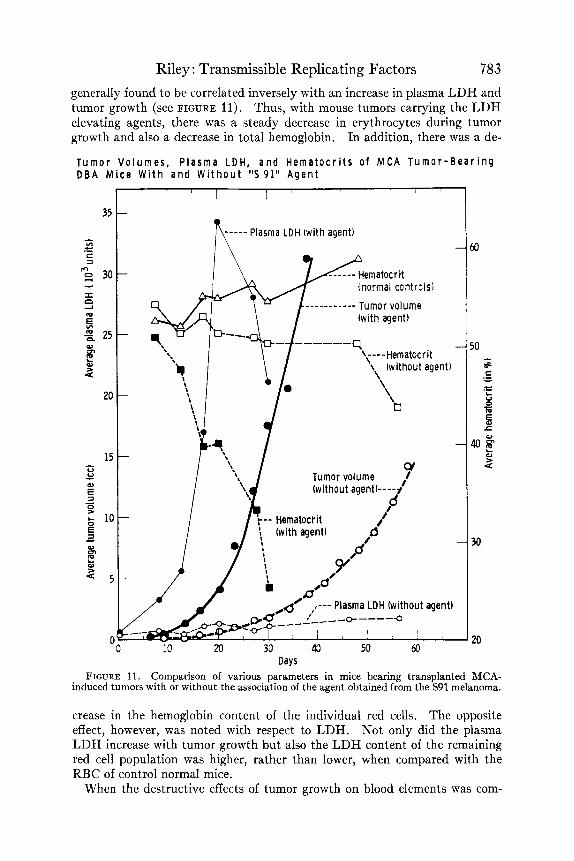
Riley : Transmissible Replicating Factors 783 generally found to be correlated inversely with an increase in plasma LDH and tumor growth (see FIGURE 11). Thus, with mouse tumors carrying the LDH elevating agents, there was a steady decrease in erythrocytes during tumor growth and also a decrease in total hemoglobin. In addition, there was a de-
T u m o r V o l u m e s , P l a s m a LDH, a n d H e m a t o c r i t s o f MCA T u m o r - B e a r i n g D B A M i c e W i t h a n d W i t h o u t ' 5 9 1 " A g e n t
---_- Plasma LDH (with agent) n d--- Hematocrit
__-_------- Tumor volume
(normal controls)
(wi th agent)
\ ----Hematocrit '\, (without agent)
Days FIGURE 11. Comparison of various parameters in mice bearing transplanted MCA-
induced tumors with or without the association of the agent obtained from the S91 melanoma.
crease in the hemoglobin content of the individual red cells. The opposite effect, however, was noted with respect to LDH. Not only did the plasma LDH increase with tumor growth but also the LDH content of the remaining red cell population was higher, rather than lower, when compared with the RBC of control normal mice.
When the destructive effects of tumor growth on blood elements was com-

784 Annals New York Academy of Sciences pared with tumors which were similar except for agent addition as described above, it was found that the presence of the LDH-elevating agents signifi- cantly increased the anemia inducing process. Details of this aspect of the studies will be reported elsewhere but the data listed in TABLE 7 are illus-
TABLE 7
COMPARED WITH SIMILAR HOSTS BEARING ANALOGOUS TUMORS WITHOUT AGENT* INCREASED ANEMIA OF HOSTS BEARING TUMORS INFECTED WITH AN LDH ELEVATING AGENT
1 9 2 3 I 1;;
Average 1
Tumor only
Tumor volume (cc)
0.1 1.2 2 .o 2.5 3.1 6 .9 8.7
3.5
Hematocri t (%)
53 .O 51 .o 50.0 48.8 42 .O 49.0 40.0
47.6
Tumor with virus
rumor volume (cc)
0.1 1.7 2.5 2.4 3.7 6.8 8.9
3.7
Hernatocrit (%)
51 .o 40.0 45 .O 40.0 21 .o 39.0 26.0
37.4
* Individual mice were selected for similarity of tumor volume in the two groups to elimi- nate possibility of tumor size effect on the hematocrit. Tumor pairs were of the same age. Agent obtained from an S91 melanoma-bearing mouse and was introduced during 20-methyl- cholanthrene tumor induction. Data obtained from primary tumors and various subsequent passages through four transplant generations.
FIGURE 12. Comparison of the average survival time of tumor-bearing mice, with and without the agent, in five separate experiments.

Riley : Transmissible Replicating Factors 785
Glucose Only Q k QE
Q::
Glucose f DPNH f Pyruvate Q:&
trative of the findings. Also, as might be expected, the absence of detectable viruses with their synergistic destructive effects, provided host benefits through an increase in average survival time of mice carrying the agent-free line of tu- mors. Among various experiments, as shown in FIGURE 12, there was a range from 6 to 128 per cent increase in longevity of such tumor-bearing mice.
1 6 500 2 10 400
5 13 160 6 18 200
Tumor Glycolysis and LDH-Elevating Agents The competence of these agents to flood the host plasma with a glycolytic
enzyme to more than 100 times the normal titer suggested that the influence of the agents on the glycolysis of the tumor tissue itself should be examined.
Tumors from mice bearing third generation transplants of chemically in- duced neoplasms with and without the agent were prepared in a standard manner except that special care was taken to perfuse the host with saline prior to removal of the tumor to eliminate blood contamination. Since the tumors were too soft to provide satisfactory slices, a conventional mince was used. Approximately 100 mg. of such tissue was added to each Warburg vessel which
TABLE 8
WITHOUT AN LDH-ELEVATING VIRUS COMPARATIVE GLYCOLYSIS BETWEEN TUMORS WITH AND
Per cent increase (with agent) I 1 Tumor (“agent-free”) Substrate

786 Annals New York Academy of Sciences These preliminary data raise obvious questions concerning the possible influ-
ence of virus-like agents, or other forms of mobile nucleic acid, upon tumor and tissue metabolism. The most important one, perhaps, is to what extent are they responsible for producing the Warburg effect of enhanced glycolysis in tumors?
Questions and Problems The observation that virus-like agents are associated with most transplanted
mouse tumors produces an obvious need for answers to several fundamental questions: Are these agents etiologically related to the cancer process? Do they arise de novo with tumor induction, or are they unrelated casual contami- nants?
Based on simplicity, the most tempting explanation of the significance of these ubiquitous agents in transplanted mouse tumors would undoubtedly be that they are contaminants, merely passenger viruses picked up from an en- demic infection in mice stock and perpetuated in the tumor and host through transplant passage. In support of this hypothesis, we have found LDH elevat- ing agents in “normal” mice from various breeders. Depending upon the source and the strain, the number of those infected have varied from 20 per cent to less than one per cent.
If, for statistical purposes, one assumed a 5 per cent over-all infection of mouse breeding colonies, there would be one chance in 20 of a given tumor passage acquiring a permanent virus contaminant. There is probably little doubt but that such a mechanism is constantly functioning with these and possibly other virus types. However, this still does not clearly answer the question of their etiological relationship to the neoplastic process, since the same situation may also apply to polyoma virus. In connection with possible modes of transmission in nature, we have demonstrated that tumor-bearing animals can infect normal adult mice by direct contact, or that contamination can take place from mother to offspring or vice versa.
On the surface, it would appear relatively simple to inject such agents into mice and record the ultimate presence or absence of tumors and settle the matter. However, some early investigators were victims of this attractive and reasonable appearing experimental simplification and thus overlooked such important neoplastic phenomena as the Gross leukemia-inducing agent,25 the Bittner mammary tumor agent,26 S. E. Polyoma ~ i r u s , ) ~ the Moloney leukemia agent,’* the Friend leukemia virus,17 etc. It is now recognized that successful experimental viral tumor induction is not necessarily subject to simple direct injection processes. The classical paradox is the Shope rabbit papilloma circumstance where tumors induced in domestic rabbits by a known tumor virus from wild cottontails fail to induce additional tumors by their cell-free filtrates. An analogous appearing situation presently holds for the S. E. polyoma virus-induced tumors in the hamster.28 The Gross leukemia agent also failed to induce disease experimentally, until the Dalldorf technique of employing new born mice was utilized.29 Another simple example of special requirement is that the Gross agent, as well as Moloney’s, fails to induce leukemia unless the appropriate mouse strain is utilized. Additional complica- tions could be cited to establish the present state of uncertainty on the me-

Riley : Transmissible Replicating Factors 78 7 chanics and requirements of tumor induction even when we are dealing with known biological carcinogens.
Respect for these lessons and the newer information available, includes avoidance of premature judgment concerning the etiological nature and sig- nificance, or the disease producing potential of these, or any agents, found associated with transplanted tumors. We are, however, approaching these problems experimentally from two somewhat separate viewpoints :
(1) What effect do viruses have on tumors, and on the tumor-bearing host, irrespective of whether these viruses induce tumors or not?
(2) Since under normal “blind” laboratory passage of mouse tumors an LDH-elevating agent is eventually demonstrable, can their “de novo” ap- pearance be observed, charted, and elucidated by following virus-free, radiation or chemically induced, primary tumors through serial passages in pretested LDH-elevating agent-free recipient mice?
Perhaps the most encouraging and useful factor in obtaining the necessary experimental information required for a firm solution to these problems is the acquisition of an enzymic method for the detection and quantitative measure- ment of virus-like agents which have heretofore been unsuspected of being associated with our classical experimental tumors.
Summary Techniques for the detection and measurement of previously unsuspected
transmissible agents in mouse melanomas and other transplanted mouse tu- mors are described. The various influences which these agents exert, both by themselves and in concert with their associated tumors, are illustrated.
Recently obtained information casts further doubt on the proper terminology for these infectious agents. While they seem to have many properties in common with classical viruses, there is accumulating evidence that they may represent a separate category at a yet more primitive level. This is based on data indicating that they are smaller by an order of magnitude than any known viruses, and by the apparent absence of sensitivity to either RNase or DNase. Their high i n vivo titer characteristics are also unknown for viruses since over 10l8 infectious units per ml of freshly drawn blood can be detected by appro- priate titration in normal mice. Until clarification of the ultimate nature of these replicating entities is obtained, use of a non-specific term such as “agent” is indicated.
Acknowledgments I acknowledge with gratitude the participation of laboratory colleagues
in these studies. Assistance in the experimental work was competently pro- vided by Editha Huerto, Mary Anne Fitzmaurice, David Bardell, Alba Co- zenza, Frank Lilly, and James Loveless. We are specially indebted to Eleanor Hevesi for volunteer secretarial services and assistance in the preparation of the manuscript. Winifred T. Powers and her volunteer staff have made many valuable contributions.
Tumors for agent testing in these studies were kindly provided by colleagues both within our institute and from other laboratories. We are indebted to the following investigators and their institutes: Ira Pilgrim, University of Utah, Salt Lake City, Utah; Frank Moyer, The Johns Hopkins University, Balti-

788 Annals New York Academy of Sciences more, Md.; Paul Hochstein, Columbia University, New York, N.Y.; John B. Nelson, The Rockefeller Institute, New York, N.Y.; Eric Hirschberg, Columbia University; J. M. Lampkin-Hibbard, University of Miami, Coral Gables, Fla. ; John Moloney, National Cancer Institute; Jacob Furth, Roswell Park Mem- orial Institute, Rye, N.Y.; and Ludwig Gross, Veteran’s Administration Hospi- tal, New York, N.Y. The following contributors are from our own institute: K. Sugiura, P. C. Merker, L. J. Old, G. W. Woolley, H. Balner, G. S. Tarnow- ski, M. N. Teller, D. J. Hutchison, and J. A. Jacquez. Chester Southam pro- vided miscellaneous virus samples and Gilbert Dalldorf provided similar mate- rials as well as friendly counsel and encouragement.
C. Chester Stock has provided essential support and valuable counsel for which appreciation is expressed.
References 1. RILEY, V., F. LILLY, E. HUERTO & D. BARDELL. 1960. Transmissible agent associated
with 26 types of experimental mouse neoplasms. 2. RILEY, V., E. HUERTO, F. LILLY, D. BARDELL, J. I). LOVELESS & M. FITZHAURICE.
1961. Some characteristics of virus-like entities associated with thirty varieties of experimental tumors.
3. RILEY, V. 1961. Virus-tumor synergism. Science. 134: 666-668. 4. RILEY, V. 1960. Ada tation of orbital bleeding technic to rapid serial blood studies.
Proc. SOC. Exptl. Biof Med. 104: 751-754 5. WARBURG, 0. 1938. Chemische Konstituiion von Fermenten. Ergeb. Enzymforch.
7: 210-245. 6. KUBOWITZ, F. & P. Om. 1943. Isolierung und Rristallization eines Garungsferments
aus Tumoren. 7. MEISTER, A. 1950. Lactic dehydrogenase activity of certain tumors and normal
tissues. 8. HILL, B. R. & C. LEVI. Elevation of a serum component in neoplastic disease.
Can. Research. 14: 513-515. 9. HILL, B. R. 1956. Some properties of serum lactic dehydrogenase. Can. Research.
16: 460466. 10. WR6BLEWSKI, F. & J. S. LADUE. 1955. Lactic dehydrogenase activity in blood.
Proc. SOC. Exptl. Biol. Med. 90: 210-213. 11. WR~BLEWSKI, F. Personal communication. 12. COOPER, J., P. A. SRERE, M. TABACHNICK & E. RACKER. 1958. The oxidative pentose
phosphate cycle. 11. Quantitative determination of intermediates and enzymes. Arch. Biochem. Biophys. 74: 306-314.
13. REED, L. J. & H. MUENCR. 1938. A simple method of estimating fifty per cent end- points.
14. IRWIN, J. 0. & E. A. CHEESEMAN. 1939. On an approximate method of determining the median effective dose and its error, in the case of a quanta1 response. Am. J. Hyg. 39: 574-580.
15. RILEY, V. & A. E. MOORE. Unpublished. 16. RILEY, V. & F. WR6BLEWSKI. Serial lactic dehydrogenase activity in plasma of
mice with growing or regressing tumors. 17. FRIEND, C. 1957. Leukemia of adult mice caused by a transmissible agent. Ann. N.Y.
Acad. Sci. 68( ): 522-532. 18. MOLONEY, J. B. Biological studies on a lymphoid-leukemia virus extracted from
sarcoma 37. J. Natl. Cancer Inst. 24: 933.
19. MUNDY, J. & P. C. WILLIAMS. 1961. Transmissible agent associated with some mouse neoplasms. Science. 134: 834-835.
20. BRODSKY, I., W. P. ROWE, J. W. HARTLEY & W. T. LANE. 1959. Studies of mouse polyoma virus infection.
21. FURTH, J., W. T. BURNETT & E. L. GADSDEN. 1953. Quantitative relationship between thyroid function and growth of pituitary tumors secreting TSH. Cancer Research. 13: 298-307.
22. KUMAOKA, S., W. L. MONEY & R. W. RAWSON. 1960. The effect of thyroxine analogues on a transplantable mouse pituitary tumor. Endocrinology. 86: 32-38.
Science. 132: 545-547.
Proc. Am. Assoc. Can. Research. 3: 261.
Biochem. Z. 314: 94-117.
J. Natl. Cancer Inst. lO(6): 1263-1271. 1954.
Am. J. Hyg. 27: 493-497.
1960. Science. 132: 151-152.
1960. I. Origin and introductory investigations.
J. Exptl. Med. 109: 439447.

Riley : Transmissible Replicating Factors 789 23. UMBREIT, W. W., R. H. BURRIS & J. F. STAUFFER. 1957. In Manometric Techniques.
3rd ed. Burgess Publ. Co. Minneapolis, Minn. 24. BARKER, S. B. & W. H. SUMMERSON. 1941. The colorimetric determination of lactic
acid in biological material. J. Biol. Chem. 138: 535-554. 25. GROSS, L. 1951. Spontaneous leukemia developing in C3H mice following inoculation,
in infancy, with AK-leukemic extracts, or AK-embryos. Proc. SOC. Exptl. Biol. 76: 27.
26. BITTNER, J. J. Some possible effects of nursing on the mammary gland tumor incidence in mice. Science. 84: 162.
27. STEWART, S. E., B. E. EDDY, N. M. GOCHENOUR, N. G. BORCESE & G. E. GRUBBS. 1957. The induction of neoplasms with a substance released from mouse tumors by tissue culture. Virology. 3: 380-400.
28. HABEL, K. 1961. Resistance of polyoma virus immune animals to transplanted polyoma tumors.
29. GROSS, L. 1961. In Oncogenic Viruses. : 280-281. Pergamon Press. New York, N.Y. 30. CLOUDMAN, A. M. 1941. The effect of an extra chromosomal influence upon trans-
1936.
Proc. SOC. Exptl. Biol. Med. 106: 722-725.
planted spontaneous tumors in mice. Science. 93: 380-381.
Discussion of the Paper
M. FOSTER (University of Michigan, Ann Arbor, Mich.) : There is one question concerning your table of serial dilutions in mice that puzzles me. Some rather quick calculations suggest that when your dilution is compared with Avogadro’s number of 6 X loza molecules per gram molecular weight, or say loz4 mole- cules, that a dilution of of whatever agent it is that you started with, would only leave you with =&a of some molecule-that is, loz4 divided by loz7.
This does not in any way cast doubt on the data as such, but I wonder whether it might not lead to a visualization of a mechanism that does not in- volve progressive dilution of a viruslike particle but rather something in the dilution medium itself that acts in some fashion so as to evoke the effect you observe.
V. RILEY (Sloan-Kettering Institute for Cancer Research, Rye, N . Y.) : I am pleased that you have raised this question so vividly.
The experiment was done for the purpose of showing that, since we continued to get an LDH response after nine serial passages in which the original mate- rial was theoretically diluted to 10-27, i t ruled out the possibility that we were simply carrying along some nonreplicating factor that was inducing the enzyme changes. This experiment is one way of showing that the agent must be replicating.
We can also do it more directly by quantitative methods in which we intro- duce a small amount of the viruslike factor into a normal mouse and get back a larger quantity. Evidence for agent multiplication was really all the experi- ment intended to show.
F. H. MOYER, JR. (Johns Hopkins University Medical School, Baltimore, Md.) : In view of the existence of five or six molecular forms of LDH in the mouse, I wonder if you have done any electrophoresis of the serum LDH in an attempt to find out whether it reflects any tissue-specific pattern of LDH.
RILEY: We have initiated such studies in collaboration with Louis Kaplan. We were counting on such experiments to tell us something about the nature and significance of the various plasma LDH phases. This was before we learned that the distinct phases all represent host reflections of the presence of these previously unsuspected agents.

790 Annals New York Academy of Sciences When we used starch gel electrophoresis and compared LDH isoenzymes in
the plasma from mice infected with the virus only and mice implanted with tumors, we found that the new LDH zones, appearing when the agent only was present, were also present with the tumor. These zones appeared similar to those found associated with the LDH of the mouse red cell.
S. ROTHMAN (University of Chicago, Argonne Cancer Research Hospital, Chicago, Ill.) : I think it would be of extreme significance to know whether all these very different tumors liberate the same molecular species of LDH, and whether it comes from one or more origins.
R. E. BILLINGHAM (Wistar Institzlte, Philadelphia, Pa.) : Have you examined any established lines of mouse cells maintained in vitro, and known not to be malignant, to see if the agent is present?
No agent was detectable in the normal mouse embryo cultures that she was using. However, our real purpose in the studies with her was to add the agent to such cultures to see if it could be propagated. We were able to maintain this agent for several months in tissue culture and then reinfect normal animals with such cultures with no difficulty.
We have also recently tested Earle’s L cells, maintained by John Jacquez, and two malignant cell lines (mouse and rat) maintained by Chester M. South- ham.
RILEY: We have done some work with Alice Moore in this connection.
These have all been negative.


















