
Enigmol: A Novel Sphingolipid Analogue with Anticancer ... · cancer (20, 21), and then its...
11
2011;10:648-657. Published OnlineFirst March 11, 2011. Mol Cancer Ther Holly Symolon, Anatoliy Bushnev, Qiong Peng, et al. Intestinal and Prostate Cancer Models for In vivo Activity against Cancer Cell Lines and Enigmol: A Novel Sphingolipid Analogue with Anticancer Updated version 10.1158/1535-7163.MCT-10-0754 doi: Access the most recent version of this article at: Material Supplementary http://mct.aacrjournals.org/content/suppl/2011/03/13/1535-7163.MCT-10-0754.DC1.html Access the most recent supplemental material at: Cited Articles http://mct.aacrjournals.org/content/10/4/648.full.html#ref-list-1 This article cites by 48 articles, 22 of which you can access for free at: Citing articles http://mct.aacrjournals.org/content/10/4/648.full.html#related-urls This article has been cited by 1 HighWire-hosted articles. Access the articles at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications on August 4, 2014. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754 on August 4, 2014. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754
Transcript of Enigmol: A Novel Sphingolipid Analogue with Anticancer ... · cancer (20, 21), and then its...

2011;10:648-657. Published OnlineFirst March 11, 2011.Mol Cancer Ther Holly Symolon, Anatoliy Bushnev, Qiong Peng, et al. Intestinal and Prostate Cancer
Models forIn vivoActivity against Cancer Cell Lines and Enigmol: A Novel Sphingolipid Analogue with Anticancer
Updated version
10.1158/1535-7163.MCT-10-0754doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2011/03/13/1535-7163.MCT-10-0754.DC1.html
Access the most recent supplemental material at:
Cited Articles
http://mct.aacrjournals.org/content/10/4/648.full.html#ref-list-1
This article cites by 48 articles, 22 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/10/4/648.full.html#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754

Therapeutic Discovery
Enigmol: A Novel Sphingolipid Analogue with AnticancerActivity against Cancer Cell Lines and In vivo Models forIntestinal and Prostate Cancer
Holly Symolon1,2, Anatoliy Bushnev3, Qiong Peng1, Harsha Ramaraju1,4, Suzanne G. Mays4,Jeremy C. Allegood1, Sarah T. Pruett3, M. Cameron Sullards1, Dirck L. Dillehay5,Dennis C. Liotta3, and Alfred H. Merrill, Jr.1
AbstractSphingoid bases are cytotoxic for many cancer cell lines and are thought to contribute to suppression of
intestinal tumorigenesis in vivo by ingested sphingolipids. This study explored the behavior of a sphingoid
base analogue, (2S,3S,5S)-2-amino-3,5-dihydroxyoctadecane (Enigmol), that cannot be phosphorylated by
sphingosine kinases and is slowly N-acylated and therefore is more persistent than natural sphingoid bases.
Enigmol had potential anticancer activity in a National Cancer Institute (NCI-60) cell line screen and was
confirmed to be more cytotoxic and persistent than naturally occurring sphingoid bases using HT29 cells, a
colon cancer cell line. Although the molecular targets of sphingoid bases are not well delineated, Enigmol
shared one of the mechanisms that has been found for naturally occurring sphingoid bases: normalization of
the aberrant accumulation of b-catenin in the nucleus and cytoplasm of colon cancer cells due to defect(s)
in the adenomatous polyposis coli (APC)/b-catenin regulatory system. Enigmol also had antitumor
efficacy when administered orally to Min mice, a mouse model with a truncated APC gene product
(C57Bl/6JMin/þ mice), decreasing the number of intestinal tumors by half at 0.025% of the diet (w/w), with
no evidence of host toxicity until higher dosages. Enigmol was also tested against the prostate cancer cell lines
DU145 and PC-3 in nudemouse xenografts and suppressed tumor growth in both. Thus, Enigmol represents a
novel category of sphingoid base analogue that is orally bioavailable and has the potential to be effective
against multiple types of cancer. Mol Cancer Ther; 10(4); 648–57. �2011 AACR.
Introduction
Sphingolipids are highly bioactive compounds thatmodulate many cell signaling pathways that are relevantto tumor biology and cancer control (1, 2). Much attentionhas been given to ceramide (Cer) and Cer analogues (3, 4)as inhibitors of cell growth and inducers of apoptosis(5), and to sphingosine 1-phosphate (S1P), which caninhibit apoptosis, induce cell migration and other pro-carcinogenic behaviors (1, 2). However, sphingosine,sphinganine, and other sphingoid bases (such as safingol)
also have the potential to be useful for cancer control,because they inhibit transformation of normal cellsinduced by irradiation (6) and chemical carcinogens(7), induce differentiation of transformed cells (8), andare growth inhibitory and cytotoxic for many cancer celltypes (9–13). The mechanism(s) for these effects isunclear; however, sphingoid bases and analogues affectmultiple signaling pathways (14) and important pro-cesses such as autophagy (15).
Because dietary sphingolipids are hydrolyzed to thesphingoid base backbones that are taken up by intestinalcells (16, 17), many of the studies of cancer suppression invivo have focused on colon cancer. Orally administeredsphingolipids decrease precancerous lesions and tumorsin models of chemically induced and inherited coloncancer (18, 19), and sphingoid bases normalize one ofthe regulatory defects in colon cancer the accumulationof b-catenin in the nucleus and cytosol of intestinal cells(20, 21).
The effectiveness of sphingoid bases seems to belimited by their phosphorylation by sphingosinekinase—an enzyme that has been called an oncogene(22); indeed, studies with sphingosine kinase 1 knock-out mice have found that this enzyme is required for
Authors' Affiliations: 1Schools of Biology, Chemistry, and Biochemistry,and the Petit Institute for Bioengineering and Bioscience, Georgia Instituteof Technology; and the 2Program in Nutrition and Health Science, Depart-ments of 3Chemistry, 4Urology, and 5Pathology and Division of AnimalResources, Emory University, Atlanta, Georgia
Note: Supplementary material for this article is available at MolecularCancer Therapeutics Online (http://mct.aacrjournals.org/).
Corresponding Author: Alfred H. Merrill, School of Biology, 310 FerstDrive, Georgia Institute of Technology, Atlanta, GA 30332. Phone: (404)385-2842; Fax: (404) 385-2917. E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-10-0754
�2011 American Association for Cancer Research.
MolecularCancer
Therapeutics
Mol Cancer Ther; 10(4) April 2011648
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754

small intestinal tumor cell proliferation in multipleintestinal neoplasia mice (Min; ref. 23). On the basisof this rationale, one would predict that compounds thatcannot be phosphorylated would be more effective incancer suppression than the naturally occurring sphin-goid base sphingosine.This study describes the findings with a sphingoid
base analogue, (2S,3S,5S)-2-amino-3,5-dihydroxyoctade-cane (named Enigmol), that cannot be phosphorylated bysphingosine kinase, and is also poorly N-acylated (24).Enigmol was compared with natural sphingoid bases forcytotoxicity for cancer cell lines, cellular uptake andmetabolism, and reduction of nuclear b-catenin, whichhas previously been associated with suppression of coloncancer (20, 21), and then its efficacy was evaluated usingmouse models for colon and prostate cancer. Enigmoldisplayed anticancer activity in all these systems andhence represents a novel category of sphingoid baseanalogue that is worthy of additional consideration incancer control.
Materials and Methods
Additional information about the materials and meth-ods has been provided in Supplementary files that areavailable online.
Sphingolipids and analoguesEnigmol [(2S,3S,5S)-2-amino-3,5-dihydroxyoctadecane;
Fig. 1B] was prepared using a highly diastereoselectivechemical methodology developed in the Liotta laboratory(25) and references cited in Supplementary Materials andMethods), except for the studies using mouse xenograftswith PC-3 cells, where itwas obtained from theNCI underthe RANDprogram. Other compounds and reagentswereobtained commercially.
Cell culture and treatmentsThe cell lines referred to in this article and Supplemen-
tary Data were obtained from the American Type CultureCollection (ATCC) and grown inmedia standard for eachcell type as described in the Supplementary files. Theidentity of the HT29, DU145, and PC-3 had been estab-lished by the ATCC by its criteria for authentication,which are described on their Web site (http://www.atcc.org/Science/CollectionsResearchandDevelopment/CellBiology/tabid/205/Default.aspx) and the lines hadnot been passaged in culture for greater than 6 monthsfrom their receipt or resuscitation.For treatment of the cells in culture, the sphingoid bases
were prepared as the 1:1 molar complex with fatty acid–free bovine serum albumin (BSA; Calbiochem; ref. 26).Unless otherwise noted, viability was measured usingthe WST-1 reagent (Roche). The NCI-60 Human TumorCell Line Screen was conducted by the NCI’s Develop-mental Therapeutics Program (NCI DTP; http://www.dtp.nci.nih.gov/branches/btb/ivclsp.html) using theBSA complex of Enigmol provided by our laboratory.
Analysis of sphingolipids by liquid chromatography,electrospray ionization tandem mass spectrometry
Lipid analysis was conducted by extracting the sphin-golipids from the cells followed by liquid chromatogra-phy, electrospray ionization tandem mass spectrometry(LC ESI-MS/MS) in positive ion mode as described pre-viously (27), with minor modifications described in Sup-plementary Materials and Methods.
Other assaysThe localization of b-catenin in HT29 cells was deter-
mined using a primary anti–b-catenin antibody (Trans-duction Labs; 1:500 dilution) and Alexa Fluor 488 goatanti-mouse IgG (Molecular Probes) as the second anti-body (20, 21).
Animals and treatmentsProtocols involving animals were approved by the
Institutional Animal Care and Use Committee and con-ducted according to National Research Council Guide-lines. MaleMinmice (C57Bl/6JMin/þmice; 32 days of age;Jackson Laboratory) were fed a semipurified AIN 76Adiet (Dyets) that is essentially sphingolipid free (28)supplemented with 0%, 0.025%, or 0.1% (w/w) Enigmol.At 39 days of age, themicewere changed to these diets for45 days and then killed for analysis of the tissues.
In the first mouse xenograft study, athymic nu/nuBalb/c mice (Charles River Laboratories) of both sexesat 32 days of age were given subcutaneous injectionsof DU145 cells (5 � 106/100 mL PBS) in the right andleft flanks, then the groups received Enigmol i.p. (0 or8 mg/kg mouse body weight in 200 mL olive oil) dailyfor 5 days. Tumors were measured with calipers (widthand length) 2 times a week and the tumor volume wascalculated as 0.5 � (length � width2). In the second,male, athymic (nu/nu) mice (from the NCI) received5 � 106 PC-3 cells, and when tumors became palpable,mice were assigned to groups (n ¼ 15) that received 0 or10 mg/kg Enigmol orally by daily gavage until termina-tion of the experiment. For these studies, Enigmol wasdissolved in a small volume of 100% ethanol and thendiluted into olive oil; the daily dosage was delivered in200 mL per mouse.
Results
The hypotheses of this study were that Enigmol willdisplay potential anticancer activity with human cancercell lines, have greater potency than sphingosine andsphinganine because it will undergo slower metabolism,modulate at least some of the regulatory processes thathave been seen previously for sphingoid bases, andsuppress tumorigenesis in vivo in animals models.
Results from an NCI-60 human tumor cell linescreen
When Enigmol was tested by the NCI (http://dtp.nci.nih.gov/branches/btb/ivclsp.html; ref. 29), it displayed
Anticancer Activities of Enigmol
www.aacrjournals.org Mol Cancer Ther; 10(4) April 2011 649
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754
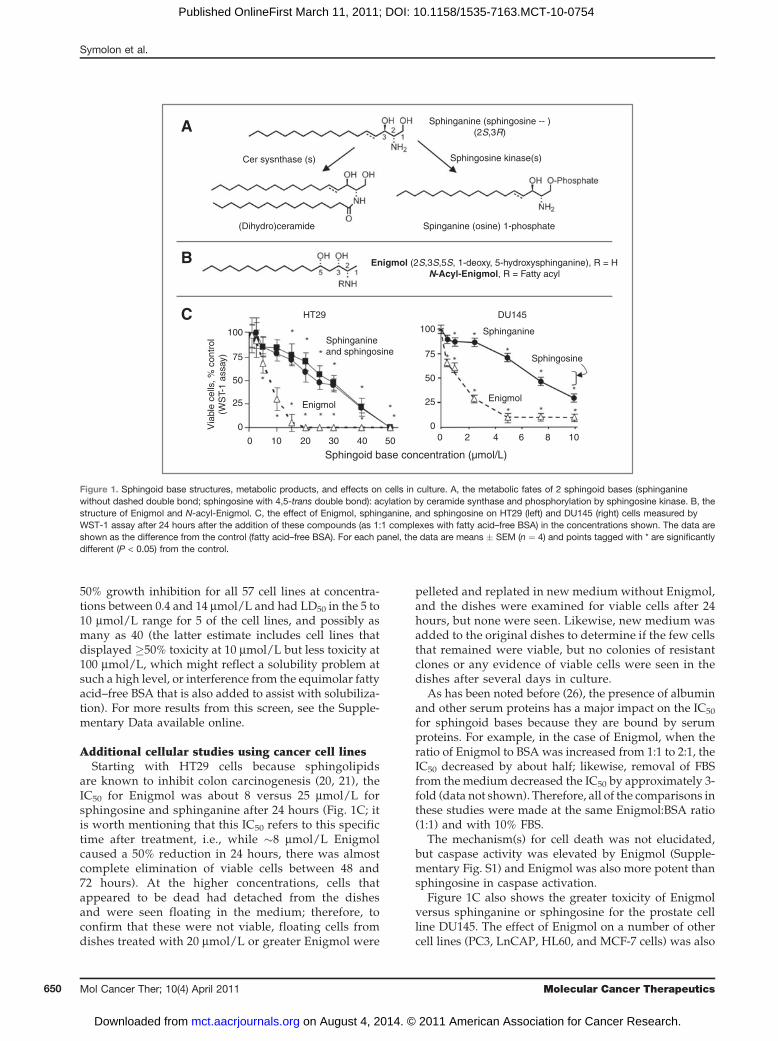
50% growth inhibition for all 57 cell lines at concentra-tions between 0.4 and 14 mmol/L and had LD50 in the 5 to10 mmol/L range for 5 of the cell lines, and possibly asmany as 40 (the latter estimate includes cell lines thatdisplayed �50% toxicity at 10 mmol/L but less toxicity at100 mmol/L, which might reflect a solubility problem atsuch a high level, or interference from the equimolar fattyacid–free BSA that is also added to assist with solubiliza-tion). For more results from this screen, see the Supple-mentary Data available online.
Additional cellular studies using cancer cell linesStarting with HT29 cells because sphingolipids
are known to inhibit colon carcinogenesis (20, 21), theIC50 for Enigmol was about 8 versus 25 mmol/L forsphingosine and sphinganine after 24 hours (Fig. 1C; itis worth mentioning that this IC50 refers to this specifictime after treatment, i.e., while �8 mmol/L Enigmolcaused a 50% reduction in 24 hours, there was almostcomplete elimination of viable cells between 48 and72 hours). At the higher concentrations, cells thatappeared to be dead had detached from the dishesand were seen floating in the medium; therefore, toconfirm that these were not viable, floating cells fromdishes treated with 20 mmol/L or greater Enigmol were
pelleted and replated in new medium without Enigmol,and the dishes were examined for viable cells after 24hours, but none were seen. Likewise, new medium wasadded to the original dishes to determine if the few cellsthat remained were viable, but no colonies of resistantclones or any evidence of viable cells were seen in thedishes after several days in culture.
As has been noted before (26), the presence of albuminand other serum proteins has a major impact on the IC50
for sphingoid bases because they are bound by serumproteins. For example, in the case of Enigmol, when theratio of Enigmol to BSA was increased from 1:1 to 2:1, theIC50 decreased by about half; likewise, removal of FBSfrom the medium decreased the IC50 by approximately 3-fold (data not shown). Therefore, all of the comparisons inthese studies were made at the same Enigmol:BSA ratio(1:1) and with 10% FBS.
The mechanism(s) for cell death was not elucidated,but caspase activity was elevated by Enigmol (Supple-mentary Fig. S1) and Enigmol was also more potent thansphingosine in caspase activation.
Figure 1C also shows the greater toxicity of Enigmolversus sphinganine or sphingosine for the prostate cellline DU145. The effect of Enigmol on a number of othercell lines (PC3, LnCAP, HL60, and MCF-7 cells) was also
A
Cer sysnthase (s) Sphingosine kinase(s)
Sphinganine (sphingosine -- )(2S,3R)
Spinganine (osine) 1-phosphate
Enigmol (2S,3S,5S, 1-deoxy, 5-hydroxysphinganine), R = HN-Acyl-Enigmol, R = Fatty acyl
(Dihydro)ceramide
100
Via
ble
cells
, % c
ontr
ol(W
ST-
1 as
say) 75
50
25
0
100
DU145HT29
Sphinganine
Sphingosine75
50
25
0
0 0 2 4 6 8 1010 20 30 40
Enigmol
Sphingoid base concentration (µmol/L)
Enigmol
Sphinganineand sphingosine
50
B
C
Figure 1. Sphingoid base structures, metabolic products, and effects on cells in culture. A, the metabolic fates of 2 sphingoid bases (sphinganinewithout dashed double bond; sphingosine with 4,5-trans double bond): acylation by ceramide synthase and phosphorylation by sphingosine kinase. B, thestructure of Enigmol and N-acyl-Enigmol. C, the effect of Enigmol, sphinganine, and sphingosine on HT29 (left) and DU145 (right) cells measured byWST-1 assay after 24 hours after the addition of these compounds (as 1:1 complexes with fatty acid–free BSA) in the concentrations shown. The data areshown as the difference from the control (fatty acid–free BSA). For each panel, the data are means � SEM (n ¼ 4) and points tagged with * are significantlydifferent (P < 0.05) from the control.
Symolon et al.
Mol Cancer Ther; 10(4) April 2011 Molecular Cancer Therapeutics650
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754

examined using the WST-1 assay (as in Fig. 1C) and allhad IC50 values in the range of 8 to 12 mmol/L (data notshown); therefore, this sphingoid base analogue affects awide variety of cancer cell lines, as was indicated by theNCI-60 Human Tumor Cell Line Screen.
Enigmol uptake and metabolismThe greater toxicity of Enigmol is not due to increased
uptake by cells (Fig. 2A); however, Enigmol persistedlonger, as seen in the pulse-chase portion of the experi-ment (right graph of Fig. 2A)—that is, where the cellswere treated with sphingosine or Enigmol for 1 hour andthen the medium was removed and replaced with newmedium minus these compounds and incubated forvarying times and finally analyzed by LC ESI-MS/MS.While very little free sphingosine was found in the cells(only 10% at 3 hours and <2% at 6 hours), more than 50%of the Enigmol was present after 3 hours and about 25%after 12 hours. Under these conditions, Enigmol killed98% � 2% of the cells versus about 50% for sphingosine(Supplementary Fig. S2).A substantial portion (�5 nmol/106 cells) of the
Enigmol that disappeared during the chase period(�6 nmol/106 cells) could be accounted for as N-acyl-
Enigmols (Fig. 2B), which is consistent with this com-pound being a modest substrate for Cer synthase(s) (ref.24; the finding of multiple fatty acyl derivatives sug-gests that it is acylated by several Cer synthases,because each is relatively selective with respect to thefatty acyl coenzyme A that it uses; ref. 30). HT29 cellsalso produced small amounts (�0.1 � 0.02 nmol/106
cells at the 12-hour time point) of N,N,N-trimethyl-Enigmol, a type of sphingoid base metabolite that hasrecently been found with safingol (31).
As a weak Cer synthase inhibitor (12, 24, 32), Enigmolelevated sphinganine and sphinganine 1-phosphate(Sa1P) by several fold (Fig. 2C and D), but had little orno effect on endogenous cellular sphingosine (Fig. 2C)and S1P (Fig. 2D). The latter is somewhat surprisingbecause Enigmol inhibited sphingosine kinase in vitro(see Supplementary Fig. S3). Enigmol had little or noeffect on the amounts of Cer, sphingomyelin, and mono-hexosylCer (Supplementary Figs. S4 and S5).
The sphingosine that was added to the cells resulted inextremely large increases in S1P (Fig. 2D), from 0.07 �0.01 to 7 � 1 nmol S1P/106 cells. There was also a largeincrease in Cer (from �1 to 15 nmol/106 cells at 12 hours;Supplementary Fig. S2) and, interestingly, the fatty acid
15 3
2
1
016:0 18:0 20:0
N-Acyl chain length
22:0 24:1 24:0 01 h3 h chase
6 h chase12 h chase
10S
phin
goid
(nm
ol/1
06 c
ells
)
5
0
0.3
10
1
0.1
0.01
LOD Enigmol-P
Sa1P
S1P
0.2
Cel
lula
r sp
hing
oid
base
nmol
/106
cel
ls
Cel
lula
r sp
hing
oid
base
-1-P
(nm
ol/1
06 c
ells
)0.1
00 1 2 3
01 2
Enigmol
Sa in Enigmol-treatedcells
Sa in Enigmol-treated cells
Sa in So-treated cellsSo-treated
Enigmol-treated
Enigmol-treated
So-treated
Treatment duration (h) Treatment duration (h)
So
So
A
C D
B
Treatment duration (h) Duration after 1 h pulse (h)
Enigmol
N-A
cyl-E
nigm
ol(n
mol
/106
cel
ls)
Termination of pulseand change to
new medium
3 3 6 12
Figure 2. Cellular amounts of Enigmol versus sphingosine in HT29 cells and their effects on other lipids. A, left graph, HT29 cells were administered30 mmol/L sphingosine (So) or Enigmol and incubated for the shown times before analysis of the amounts of these compounds in the cells by LC ESI-MS/MS;right graph, amounts of sphingosine and Enigmol associated with the cells in a pulse-chase treatment, that is, the medium from some of the dishes used forthe left graph was removed after 1 hour, new medium without sphingoid base was added, and the lipids were analyzed at the times shown. The dataare expressed in nmol/mg protein as the mean� SD, n¼ 3. B, the amounts and subspecies (i.e., N-acyl-chain length variants) of Enigmol associated with theHT29 cells after the first hour and at each time point during the incubation in new medium. C and D, the effects of the added Enigmol or sphingosineon other sphingolipids of interest: sphinganine (Sa), sphinganine 1-phosphate (Sa1P), and sphingosine 1-phosphate (S1P). The lower limit of detection (LOD)for Enigmol 1-phosphate by LC ESI-MS/MS is shown by hashed lines (D). The points tagged with * and S1P in the sphingosine-treated cells are significantlydifferent (P < 0.05) from untreated (time zero) cells.
Anticancer Activities of Enigmol
www.aacrjournals.org Mol Cancer Ther; 10(4) April 2011 651
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754
compositions of the Cer in sphingosine-treated cells dif-fered considerably from theN-acyl-Enigmols in Enigmol-treated cells (Supplementary Fig. S6), therefore, it ispossible that different CerS are involved. Sphingomyelinsand monohexosylCers also increased when the cells weretreated with sphingosine, but by much lower amounts(�1 nmol/106 cells each) than was seen with Cer (Sup-plementary Fig. S5). These results show that the additionof sphingosine alters many sphingolipid subspecies, withprobably themost noteworthy being the 100-fold increasein S1P and 15-fold increase in Cer.
Other effects of Enigmol on HT29 cellsBecause a large number of signaling pathways are
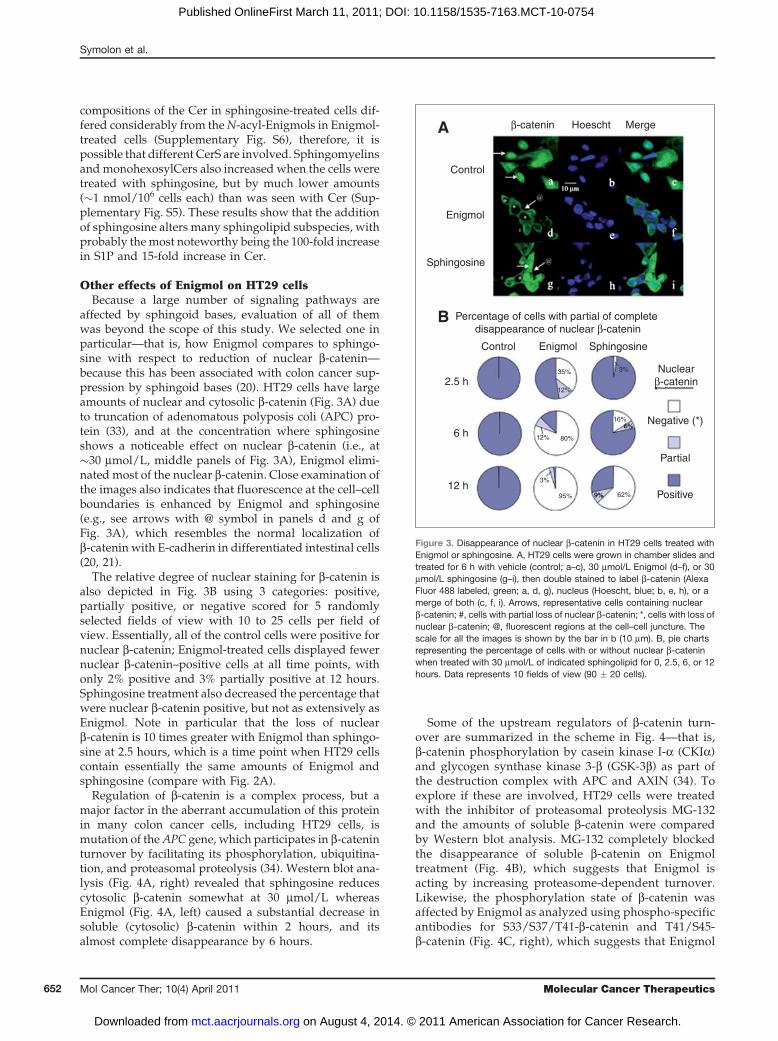
affected by sphingoid bases, evaluation of all of themwas beyond the scope of this study. We selected one inparticular—that is, how Enigmol compares to sphingo-sine with respect to reduction of nuclear b-catenin—because this has been associated with colon cancer sup-pression by sphingoid bases (20). HT29 cells have largeamounts of nuclear and cytosolic b-catenin (Fig. 3A) dueto truncation of adenomatous polyposis coli (APC) pro-tein (33), and at the concentration where sphingosineshows a noticeable effect on nuclear b-catenin (i.e., at�30 mmol/L, middle panels of Fig. 3A), Enigmol elimi-nated most of the nuclear b-catenin. Close examination ofthe images also indicates that fluorescence at the cell–cellboundaries is enhanced by Enigmol and sphingosine(e.g., see arrows with @ symbol in panels d and g ofFig. 3A), which resembles the normal localization ofb-catenin with E-cadherin in differentiated intestinal cells(20, 21).
The relative degree of nuclear staining for b-catenin isalso depicted in Fig. 3B using 3 categories: positive,partially positive, or negative scored for 5 randomlyselected fields of view with 10 to 25 cells per field ofview. Essentially, all of the control cells were positive fornuclear b-catenin; Enigmol-treated cells displayed fewernuclear b-catenin–positive cells at all time points, withonly 2% positive and 3% partially positive at 12 hours.Sphingosine treatment also decreased the percentage thatwere nuclear b-catenin positive, but not as extensively asEnigmol. Note in particular that the loss of nuclearb-catenin is 10 times greater with Enigmol than sphingo-sine at 2.5 hours, which is a time point when HT29 cellscontain essentially the same amounts of Enigmol andsphingosine (compare with Fig. 2A).
Regulation of b-catenin is a complex process, but amajor factor in the aberrant accumulation of this proteinin many colon cancer cells, including HT29 cells, ismutation of theAPC gene, which participates in b-cateninturnover by facilitating its phosphorylation, ubiquitina-tion, and proteasomal proteolysis (34). Western blot ana-lysis (Fig. 4A, right) revealed that sphingosine reducescytosolic b-catenin somewhat at 30 mmol/L whereasEnigmol (Fig. 4A, left) caused a substantial decrease insoluble (cytosolic) b-catenin within 2 hours, and itsalmost complete disappearance by 6 hours.
Some of the upstream regulators of b-catenin turn-over are summarized in the scheme in Fig. 4—that is,b-catenin phosphorylation by casein kinase I-a (CKIa)and glycogen synthase kinase 3-b (GSK-3b) as part ofthe destruction complex with APC and AXIN (34). Toexplore if these are involved, HT29 cells were treatedwith the inhibitor of proteasomal proteolysis MG-132and the amounts of soluble b-catenin were comparedby Western blot analysis. MG-132 completely blockedthe disappearance of soluble b-catenin on Enigmoltreatment (Fig. 4B), which suggests that Enigmol isacting by increasing proteasome-dependent turnover.Likewise, the phosphorylation state of b-catenin wasaffected by Enigmol as analyzed using phospho-specificantibodies for S33/S37/T41-b-catenin and T41/S45-b-catenin (Fig. 4C, right), which suggests that Enigmol
β-catenin
Controla
d
b10 μm
c
e
h ig
f
A
B
Enigmol
Sphingosine
2.5 h
6 h
12 h
Control Enigmol
35%
12%
3%
16%6%
80%12%
3%
95% 62%9%
Sphingosine
Nuclearβ-catenin
Negative (*)
Partial
Positive
Percentage of cells with partial of complete disappearance of nuclear β-catenin
Hoescht Merge
Figure 3. Disappearance of nuclear b-catenin in HT29 cells treated withEnigmol or sphingosine. A, HT29 cells were grown in chamber slides andtreated for 6 h with vehicle (control; a–c), 30 mmol/L Enigmol (d–f), or 30mmol/L sphingosine (g–i), then double stained to label b-catenin (AlexaFluor 488 labeled, green; a, d, g), nucleus (Hoescht, blue; b, e, h), or amerge of both (c, f, i). Arrows, representative cells containing nuclearb-catenin; #, cells with partial loss of nuclear b-catenin; *, cells with loss ofnuclear b-catenin; @, fluorescent regions at the cell–cell juncture. Thescale for all the images is shown by the bar in b (10 mm). B, pie chartsrepresenting the percentage of cells with or without nuclear b-cateninwhen treated with 30 mmol/L of indicated sphingolipid for 0, 2.5, 6, or 12hours. Data represents 10 fields of view (90 � 20 cells).
Symolon et al.
Mol Cancer Ther; 10(4) April 2011 Molecular Cancer Therapeutics652
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754

enhances the phosphorylation of b-catenin by CKI-aand/or GSK-3b, which occurs at these sites (34). GSK-3b is also phosphorylated on S9 by protein kinase C (35),which might also be affected because protein kinaseC (PKC) is inhibited by sphingoid bases (36), andindeed, there was a decrease in phospho-S9-GSK-3bin cells treated with Enigmol (Fig. 4C, left).These might not be the only mechanisms whereby
Enigmol induces turnover of b-catenin in these cellsbecause other proteases are also activated by sphingoidbases (as shown in Supplementary Fig. S1 for caspaseactivation by sphingosine and Enigmol). All in all, theseresults establish that Enigmol has effects similar to, butmore potent than, sphingosine with respect to decreasingthe amount of b-catenin in the nucleus and cytosol.
Inhibition of adenoma formation in Min miceTo determine if Enigmol has antitumor activity
in vivo, Min mice (19, 20) were fed an essentiallysphingolipid-free diet with no supplement (control;n ¼ 9), 0.025% (w/w) Enigmol (n ¼ 8), or 0.1%
(w/w) Enigmol (n ¼ 8). Mice fed the control diet had75.0 � 13.6 tumors per mouse, whereas those fed Enig-mol had 52% and 37% fewer tumors per mouse for the0.025% and 0.1% fed groups, respectively (P < 0.05 vs.control; Fig. 5A). Suppression by Enigmol was seenthroughout the proximal, mid, and distal portions ofthe intestine (Fig. 5A). Because only a few tumorsdevelop in the colon of Min mice, Enigmol did not havea statistically significant effect, although the averagesseem lower (i.e., 0.63 � 0.43 for 0.025% and 0.88 � 0.44for 0.1% vs. 1.0 � 0.4 for the control, with P ¼ 0.2 and0.4, respectively).
Enigmol had no adverse effect on body weight (20.2 �0.9 g for the control mice and 20.2 � 1.3 g) nor on any ofthe biomarkers for liver and kidney function (Fig. 5C) at0.025% of the diet. However, 0.1% Enigmol reducedweight gain to 4.3 � 0.7 g during the course of the 5.5-week study (vs. 6.0 � 0.5 g for the control mice and 6.2 �0.4 g for the 0.025% Enigmol group, both of which weresignificantly different from the 0.1% group with P < 0.05)for a final weight of 18.0 � 0.8 g (P < 0.05). We do not
Enigmol 30 µmol/L
β-Catenin
A
B
C
EnigmolMG-132
DestructionComplex
(APC, AXIN,CK, GSK)
β-Catenin
β-Catenin-P
β-Catenin-Ubn
Proteasomaldegradation
Enigmol
+– –
–
+
+
+– +
+––
β-Catenin
GSK-3β
P-(S9)GSK-3β
β-Catenin
P(S33/S37/T41)
P(T41/S45)
-β-catenin
-β-catenin
Actin
Actin
0 h 2 h 4 h 6 h Con
trol
Sphi
ngos
ine
Enig
mol
Actin
Actin
Figure 4. Effects of sphingoid bases on the amount of soluble b-catenin and regulators of b-catenin turnover in HT29 cells. A, the induction of b-cateninturnover by Enigmol. For the left gel, HT29 cells were treated with 30 mmol/L Enigmol for varying time points; for the right gel, the cells were treatedwith vehicle control or 30 mmol/L Enigmol or sphingosine for 6 hours; in each case, soluble proteins were separated by SDS-PAGE and immunoblotted using amonoclonal anti–b-catenin antibody. B, the effect of a proteasomal inhibitor (MG-132) on b-catenin degradation and attenuated of Enigmol-induced turnoverof b-catenin by this proteasomal inhibitor. HT29 cells were pretreated with 50 mmol/L MG-132 for 1 hour, followed by the addition of 30 mmol/L Enigmol for6 hours. C, HT29 cells were treated with or without 30 mmol/L Enigmol for 6 hours and analyzed using phospho-specific antibodies for GSK-3b and b-catenin,and cytosolic proteins (i.e., after centrifugation to remove nuclei and membranes) were analyzed by Western blotting.
Anticancer Activities of Enigmol
www.aacrjournals.org Mol Cancer Ther; 10(4) April 2011 653
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754

know whether this is due to an effect of 0.1% Enigmol onfood consumption, which was not recorded; however, wedid not notice differences in the amounts of food left inthe food containers. In addition, although the biomarkersfor liver and kidney function were in the normal range,the average blood urea nitrogen level was significantlyhigher (P < 0.05) for mice given 0.1% Enigmol versus thecontrol. Total serum protein and albumin levels weresomewhat higher for the Enigmol-treated mice comparedwith the control, which is generally not thought to be anindication of toxicity, but may indicate dehydration(Fig. 5C, right panels). All together, these results establishthat Enigmol can reduce intestinal tumorigenesis in thismouse model with no evidence of host toxicity at 0.025%of the diet, but the higher level of 0.1% may be moreproblematic.
In vivo antitumor activity and toxicity of Enigmol inmouse xenografts for prostate cancer
Although the focus of our studies has been on coloncancer due to the relatively well-established link forthis cancer (18–21), the screening of Enigmol against othercancer cell lines indicates that it might have broader effi-cacy (Fig. 1C and Supplementary Data). To test thispossibility, Enigmolwas tested against the prostate cancerline DU145 in a mouse xenograft model (nu/nu Balb/cmice), because it was toxic for these cells in culture(Fig. 1C). As shown in Fig. 6A, Enigmol significantly
suppressed the growth (i.e., a �50% reduction in tumorsize comparedwith the control) of tumors from these cellswhen injected i.p. at 8 mg/kg body weight for 5 days. Atthis dosage, there was no sign of host toxicity by histo-pathology or by clinical pathology for either study; themean � SD for 3 randomly selected male mice givenEnigmol at this dosage were as follows: blood urea nitro-gen (25� 2 mg/dL), creatinine (0.65� 0.49 mg/dL), ALT(alanine aminotransferase; 33 � 9 U/L), albumin (2.6 �0.4 g/dL), and total serum protein (4.8 � 0.3 g/dL), allwithin normal values.
In a second study (Fig. 6B), Balb/c nu/nu miceimplanted with PC-3 cells were assigned to 3 groups(n ¼ 15) that received a daily oral gavage with vehiclealone (200 mL of olive oil) or 10 mg/kg Enigmol in200 mL of olive oil until termination of the experiment.This also resulted in an approximately 50% reductionin tumor size for the Enigmol group versus the vehiclecontrol. The animals in these groups displayed similarchanges in weight during the treatment period; thatis, the starting/ending weights were as follows:22.8 � 2.4 g/19.5 � 2.4 g (loss of 3.3 g) for the controlsversus 23.5 � 3.5 g/20.8 � 2.9 g (loss of 2.7 g) for theanimals administered Enigmol at 10 mg/kg.
Therefore, these studies show that Enigmol has antic-ancer activity against 2 human prostate cancer cell lines ina mouse xenograft model and the latter study shows thatan effect can be seen when Enigmol is administeredorally.
100Control0.025% Enigmol)0.1% Enigmol
8
6
4
2
0
Control 0.025%Enigmol as %diet
0.1%
80
Ave
rage
no.
of t
umor
spe
r m
ouse
Wei
ght g
ain
durin
g fe
edin
g (g
)
60
40
20
0
50
A
C
B
0.12(*) (*) (*) (*)50 10 6
4
2
8
6
4
2
40
30
20
10
0 0.1.025 0 0 0 00.1 0.1 0.1 0.1.025 .025
Enigmol as %diet
.025 .025
0.08
Cre
atin
ine
(mg/
dL)
BU
N (
mg/
dL)
ALT
(U
/L)
Tota
l pro
tein
(g/
dL)
Alb
umin
(g/
dL)
0.04
40
30
20
10
Proxim
alM
idDist
al
colon To
tal
Figure 5. Effect of Enigmol on tumorigenesis and host toxicity in Min mice. A, the effect of Enigmol on tumor number in APCMin/þ mice after 6 weeksfeeding starting at 32 days of age. Tumor number was assessed by light microscopy and adenomas throughout the entire intestinal tract werecounted. Data bars represent mean � SEM (n ¼ 9, control; n ¼ 8, Enigmol-treated). *, P < 0.05, significant difference from untreated control (the groupsgiven 0.025% and 0.1% Enigmol were not significantly different from each other, P < 0.1). B, the effect of Enigmol administered in the diet on thetotal weight gained at the end of 45 days feeding. Data represent mean � SD. C, the blood levels of common hepato- and renal toxicity markers.Shaded area represents normal diagnostic range for values. Data represented as mean (—) and individual values (¤). *, P < 0.05, significantdifference from untreated control.
Symolon et al.
Mol Cancer Ther; 10(4) April 2011 Molecular Cancer Therapeutics654
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754

Blood and tissue levels of EnigmolAn analysis of the pharmacokinetics of Enigmol uptake
and elimination will be published separately (37); how-ever, it is worth mentioning here that the amounts ofEnigmol in blood on the fourth day of administration of4 mg Enigmol/kg i.p. was 0.41 � 0.20 mmol/L (n ¼ 4),following the protocol used for Fig. 6A. In the study usingPC-3 cells to prepare the xenografts (Fig. 6B), analysis of 3of the tumors at the end of the study showed that theycontained 0.18 � 0.07 nmol of Enigmol/g of tissue. Theseresults confirm that Enigmol appears in both blood andthe tumors.
Discussion
These studies have established that Enigmol is toxic fornumerous human cancer cell lines in the NCI-60 screenand suppressed tumor growth inmousemodels for colonand prostate cancer. The toxicity for so many cell lines inthe NCI-60 screen raised concern that Enigmol might be
toxic for host and cancer cells; however, the in vivo studiessuggested that tumors are most affected.
It is usually difficult to predict if compounds thatdisplay potential anticancer activity in studies of cellsin culture will have efficacy in vivo, and this can beespecially problematic when the agent is hydrophobicand its delivery to the target site might be limiting.Therefore, on a practical level, the most remarkablefindings of these studies were that Enigmol suppressestumor growth not only in a colon cancer model (Minmice), as has been seen before for dietary sphingolipids(20), but also in mouse xenografts with 2 prostate cancercell lines, thereby extending the types of cancer thatmight be affected by this category of compounds, andthat oral administration of Enigmol could affect the PC3cell xenografts. These properties, plus the apparently lowhost toxicity of Enigmol when fed to animals at levels thatdecrease tumor growth, suggest that this compound haspromise for cancer control.
The rationale behind the selection of this type of ana-logue was that sphingoid bases lacking the 1-hydroxygroup cannot be metabolized by sphingosine kinase(s)and the 2S,3S stereochemistry (compared to 2S,3R forsphingosine) also makes Enigmol a poorer substrate forN-acylation (24, 38); therefore, Enigmol would be pre-dicted to be more persistent as the free sphingoid base,which was borne out by the cell culture studies withHT29 cells and the appearance of free Enigmol in tumorsin the mouse xenograft studies. A longer persistence ofthe free sphingoid base might also contribute to thecytotoxicity of the synthetic analogues safingol (39), N,N-dimethylsphingosine (40) and naturally occurringsphingoid base variants (41, 42), including one recentlyevaluated in a phase I trial (43).
The inability of Enigmol to undergo phosphorylationseems to be especially important because sphingosinekinase and S1P clearly play important roles in colontumorigenesis (44, 45). Nonetheless, it does not seem thatEnigmol acts via elimination of endogenous S1P(although it can inhibit sphingosine kinases in vitro),based on no measurable reduction in S1P in the cells(Fig. 2). This warrants further study, however, becausethe pertinent S1P might be localized in a specific sub-compartment, secreted from the cells, and/or involved inreceptor cross-talk (46), that would not have beendetected in our analysis. It was striking that the additionof sphingosine to HT29 cells elevated S1P levels by morethan 100-fold, which might be a major factor in its lowertoxicity, because S1P can sometimes protect cells againstapoptosis. The addition of sphingosine also elevated Cerbymore than 10-fold and sphingomyelin and glycosphin-golipids by smaller but detectable amounts (Supplemen-tal Figs. S2 and S3), which illustrates the complexity ofstudies of naturally occurring sphingolipids, because theadded compound undergoes extensive metabolism tomultiple bioactive species.
The mechanism(s) for the tumor suppression by Enig-mol has been only partially defined by these experiments;
200
300A
B
100
1,600
1,200
800
400
0
Ave
rage
tum
or v
olum
e (m
m3 )
Ave
rage
tum
or v
olum
e (m
m3 )
0 5 10
Period of dailyadministration
Control
Control
Enigmol8 mg/kg daily
Enigmol (10 mg/kg)
PC-3
DU145
to day 40
15Days after xenograft
Days after xenograft
20 25 30 35
0 5 10 15 20 25 30 35 40
Figure 6. Evaluation of Enigmol in a nude mouse xenograft models forhuman prostate cancer. A, DU145 cells were injected in both flanks ofathymic nu/nu Balb/c mice (female) at 32 days of age and after tumorswere palpable (63 � 8 and 58 � 7 mm3 for the control and treated mice,respectively; mean � SE for n ¼ 10/group), the control and treatmentgroups were given daily intraperitoneal injections of 200 mL olive oil orEnigmol at 8 mg/kg mouse body weight in 200 mL olive oil for 5 days(designated by the bar). Twice per week, the tumors were measuredwith calipers and tumor volume was calculated as follows: (greatestdiameter � smallest diameter2)/2. B, PC-3 cells were injected in male,athymic (nu/nu) mice and when tumors became palpable (day 10), micewere assigned to groups (33 � 5 and 30 � 7 mm3 for the control andtreated mice, respectively; mean � SE for n ¼ 15) that received dailygavage with the vehicle (200 mL of olive oil) or vehicle containing 10 mg ofEnigmol/kg body weight until termination of the experiment. Tumorvolumes were measured as described above. *, for both graphs, timepoints where the differences between the tumor volumes for the treatedmice were statistically different from the control by P � 0.05 by a 2-tailedunpaired t test.
Anticancer Activities of Enigmol
www.aacrjournals.org Mol Cancer Ther; 10(4) April 2011 655
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754

nonetheless, Enigmol can normalize one of the defectsthat is important in colon cancer—the aberrant appear-ance of b-catenin in the nucleus (20, 21) where it plays amajor role in regulating proliferation via the T-cell factor/lymphoid enhancer transcription factor (47). Further-more, these studies have established that Enigmol and,to a lesser extent sphingosine, increases proteasomalturnover of b-catenin, apparently by increasing its phos-phorylation by GSK-3b (and possibly CKI-a, althoughthis was not tested), which precedes the polyubiquitina-tion and degradation of b-catenin. The immediate targetof Enigmol is not known but might include PKC, which iselevated in colon tumors (48) and is known to be inhibitedby sphingoid bases (36); however, other protein kinases,such as PKB (protein kinase B)/Akt (49–51), might alsoplay a role because they are also known to be affected bysphingoid bases. It is noteworthy that interference withthese mitogenic signaling pathways might also provide alink between Enigmol and cell death, because Akt andother signaling pathways influence caspase activationand apoptotic cell death.
All in all, these studies suggest that Enigmol representsa novel category of sphingoid base analogue that is orally
bioavailable and has the potential to be effective againstmultiple types of cancer.
Disclosure of Potential Conflicts of Interest
A.H. Merrill, Jr. has patents pertaining to sphingolipids, but nonelicensed to companies.
Acknowledgments
The authors thank David Menaldino, Kena Desai, Trey Perkins, andCarrie Pack for conducting some of the initial experiments of this studyand Selwyn Hurwitz, David Pallas, and FrankMcDonald for many usefuldiscussions.
Grant Support
This work was supported by NIH grant U19-CA87525 (to allauthors) and funds from the Smithgall Institute Chair in Molecularand Cell Biology at Georgia Tech (to A. Merrill).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received August 11, 2010; revised January 20, 2011; accepted February12, 2011; published OnlineFirst March 11, 2011.
References1. Saddoughi SA, Song P, Ogretmen B. Roles of bioactive sphingo-
lipids in cancer biology and therapeutics. Subcell Biochem2008;49:413–40.
2. Takabe K, Paugh SW, Milstien S, Spiegel S. "Inside-out" signaling ofsphingosine-1-phosphate: therapeutic targets. Pharmacol Rev 2008;60:181–95.
3. Dindo D, Dahm F, Szulc Z, Bielawska A, Obeid LM, Hannun YA,et al. Cationic long-chain ceramide LCL-30 induces cell death bymitochondrial targeting in SW403 cells. Mol Cancer Ther 2006;5:1520–9.
4. Heakal Y, Kester M. Nanoliposomal short-chain ceramide inhibitsagonist-dependent translocation of neurotensin receptor 1 to struc-tured membrane microdomains in breast cancer cells. Mol CancerRes 2009;7:724–34.
5. Ogretmen B, Hannun YA. Biologically active sphingolipids in cancerpathogenesis and treatment. Nat Rev Cancer 2004;4:604–16.
6. Borek C, Ong A, Stevens VL, Wang E, Merrill AH Jr. Long-chain(sphingoid) bases inhibit multistage carcinogenesis in mouse C3H/10T1/2 cells treated with radiation and phorbol 12-myristate 13-acetate. Proc Natl Acad Sci U S A 1991;88:1953–7.
7. Borek C, Merrill AH Jr. Sphingolipids inhibit multistage carcinogenesisand protein kinase C. Basic Life Sci 1993;61:367–71.
8. Stevens VL, Owens NE, Winton EF, Kinkade JM Jr, Merrill AH Jr.Modulation of retinoic acid-induced differentiation of human leukemia(HL-60) cells by serum factors and sphinganine. Cancer Res 1990;50:222–6.
9. Jarvis WD, Fornari FA, Traylor RS, Martin HA, Kramer LB, Erukulla RK,et al. Induction of apoptosis and potentiation of ceramide-mediatedcytotoxicity by sphingoid bases in human myeloid leukemia cells. JBiol Chem 1996;271:8275–84.
10. Auzenne E, Leroux ME, Hu M, Pollock RE, Feig B, Klostergaard J.Cytotoxic effects of sphingolipids as single or multi-modality agentson human melanoma and soft tissue sarcoma in vitro. Melanoma Res1998;8:227–39.
11. Schmelz EM, Dombrink-Kurtzman MA, Roberts PC, Kozutsumi Y,Kawasaki T, Merrill AH Jr. Induction of apoptosis by fumonisin B1 inHT29 cells is mediated by the accumulation of endogenous freesphingoid bases. Toxicol Appl Pharmacol 1998;148:252–60.
12. Desai K, Sullards MC, Allegood J, Wang E, Schmelz EM, Hartl M, et al.Fumonisins and fumonisin analogs as inhibitors of ceramide synthaseand inducers of apoptosis. Biochim Biophys Acta 2002;1585:188–92.
13. Sweeney EA, Sakakura C, Shirahama T, Masamune A, Ohta H,Hakomori S, et al. Sphingosine and its methylated derivative N,N-dimethylsphingosine (DMS) induce apoptosis in a variety of humancancer cell lines. Int J Cancer 1996;66:358–66.
14. Zheng W, Kollmeyer J, Symolon H, Momin A, Munter E, Wang E, et al.Ceramides and other bioactive sphingolipid backbones in health anddisease: lipidomic analysis, metabolism and roles in membrane struc-ture, dynamics, signaling and autophagy. Biochim Biophys Acta2006;1758:1864–84.
15. Coward J, Ambrosini G, Musi E, Truman JP, Haimovitz-Friedman A,Allegood JC, et al. Safingol (l-threo-sphinganine) induces autophagyin solid tumor cells through inhibition of PKC and the PI3-kinasepathway. Autophagy 2009;5:184–93.
16. Schmelz EM, Crall KJ, Larocque R, Dillehay DL, Merrill AH Jr. Uptakeand metabolism of sphingolipids in isolated intestinal loops of mice. JNutr 1994;124:702–12.
17. Duan RD, Nilsson A. Metabolism of sphingolipids in the gut and itsrelation to inflammation and cancer development. Prog Lipid Res2009;48:62–72.
18. Schmelz EM, Dillehay DL, Webb SK, Reiter A, Adams J, Merrill AH Jr.Sphingomyelin consumption suppresses aberrant colonic crypt fociand increases the proportion of adenomas versus adenocarcinomasin CF1 mice treated with 1,2-dimethylhydrazine: implications for diet-ary sphingolipids and colon carcinogenesis. Cancer Res 1996;56:4936–41.
19. Symolon H, Schmelz EM, Dillehay DL, Merrill AH Jr. Dietary soysphingolipids suppress tumorigenesis and gene expression in 1,2-dimethylhydrazine-treated CF1 mice and ApcMin/þ mice. J Nutr2004;134:1157–61.
20. Schmelz EM, Roberts PC, Kustin EM, Lemonnier LA, Sullards MC,Dillehay DL, et al. Modulation of intracellular beta-catenin localizationand intestinal tumorigenesis in vivo and in vitro by sphingolipids.Cancer Res 2001;61:6723–9.
21. Simon KW, Roberts PC, Vespremi MJ, Manchen S, Schmelz EM.Regulation of beta-catenin and connexin-43 expression: targets for
Symolon et al.
Mol Cancer Ther; 10(4) April 2011 Molecular Cancer Therapeutics656
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754

sphingolipids in colon cancer prevention. Mol Nutr Food Res2009;53:332–40.
22. Bergelin N, Blom T, Heikkila J, Lof C, Alam C, Balthasar S, et al.Sphingosine kinase as an oncogene: autocrine sphingosine 1-phos-phate modulates ML-1 thyroid carcinoma cell migration by a mechan-ism dependent on protein kinase C-alpha and ERK1/2. Endocrinology2009;150:2055–63.
23. Kohno M, Momoi M, Oo ML, Paik JH, Lee YM, Venkataraman K, et al.Intracellular role for sphingosine kinase 1 in intestinal adenoma cellproliferation. Mol Cell Biol 2006;26:7211–23.
24. Humpf HU, Schmelz EM, Meredith FI, Vesper H, Vales TR, Wang E,et al. Acylation of naturally occurring and synthetic 1-deoxysphinga-nines by ceramide synthase. Formation of N-palmitoyl-aminopentolproduces a toxic metabolite of hydrolyzed fumonisin, AP1, and a newcategory of ceramide synthase inhibitor. J Biol Chem 1998;273:19060–4.
25. Bushnev AS, Baillie MT, Holt JJ, Menaldino DS, Merrill AH Jr, LiottaDC. An efficient asymmetric synthesis of Enigmols (1-deoxy-5-hydro-xysphingoid bases), an important class of bioactive lipid modulators.ARKIVOC 2010; 2010:263–77.
26. Hannun YA, Merrill AH Jr, Bell RM. Use of sphingosine as inhibitor ofprotein kinase C. Methods Enzymol 1991;201:316–28.
27. Merrill AH Jr, Sullards MC, Allegood JC, Kelly S, Wang E. Sphingo-lipidomics: high-throughput, structure-specific, and quantitative ana-lysis of sphingolipids by liquid chromatography tandem massspectrometry. Methods 2005;36:207–24.
28. Dillehay DL, Webb SK, Schmelz EM, Merrill AH Jr. Dietary sphingo-myelin inhibits 1,2-dimethylhydrazine-induced colon cancer in CF1mice. J Nutr 1994;124:615–20.
29. Shoemaker RH. The NCI60 human tumour cell line anticancer drugscreen. Nat Rev Cancer 2006;6:813–23.
30. Pewzner-Jung Y, Ben-Dor S, Futerman AH. When do Lasses (long-evity assurance genes) become CerS (ceramide synthases)? Insightsinto the regulation of ceramide synthesis. J Biol Chem 2006;281:25001–5.
31. Morales PR, Dillehay DL, Moody SJ, Pallas DC, Pruett S, AllgoodJC, et al. Safingol toxicology after oral administration to TRAMPmice: demonstration of safingol uptake and metabolism by N-acylation and N-methylation. Drug Chem Toxicol 2007;30:197–216.
32. Sullards MC, Merrill AH Jr. Analysis of sphingosine 1-phosphate,ceramides, and other bioactive sphingolipids by high-performanceliquid chromatography-tandem mass spectrometry. Sci STKE 2001;2001:PL1.
33. Yang J, Zhang W, Evans PM, Chen X, He X, Liu C. Adenomatouspolyposis coli (APC) differentially regulates beta-catenin phosphor-ylation and ubiquitination in colon cancer cells. J Biol Chem2006;281:17751–7.
34. Wu D, Pan W. GSK3: a multifaceted kinase in Wnt signaling. TrendsBiochem Sci 2010;35:161–8.
35. Ali AS, Ali S, El-Rayes BF, Philip PA, Sarkar FH. Exploitation of proteinkinase C: a useful target for cancer therapy. Cancer Treat Rev2009;35:1–8.
36. Smith ER, Merrill AH, Obeid LM, Hannun YA. Effects of sphingosineand other sphingolipids on protein kinase C. Methods Enzymol2000;312:361–73.
37. Garnier-Amblard EC, Mays SG, Arrendale RF, Baillie MT, Bushnev AS,Culver DG, et al. A Novel Synthesis and Biological Evaluationof Enigmols (2-Amino-3,5-Dihydroxyoctadecanes) as TherapeuticAgents for Treating Prostate Cancer. ACS Med Chem Letters2011 (in press).
38. Seiferlein M, Humpf HU, Voss KA, Sullards MC, Allegood JC, Wang E,et al. Hydrolyzed fumonisins HFB1 and HFB2 are acylated in vitro andin vivo by ceramide synthase to form cytotoxic N-acyl-metabolites.Mol Nutr Food Res 2007;51:1120–30.
39. Ling LU, Lin H, Tan KB, Chiu GN. The role of protein kinase C in thesynergistic interaction of safingol and irinotecan in colon cancer cells.Int J Oncol 2009;35:1463–71.
40. Kim HL, Im DS. N,N-Dimethyl-D-erythro-sphingosine increases intra-cellular Ca2þ concentration via Naþ-Ca2þ-exchanger in HCT116human colon cancer cells. Arch Pharm Res 2008;31:54–9.
41. Padron JM, Peters GJ. Cytotoxicity of sphingoid marine compoundanalogs in mono- and multilayered solid tumor cell cultures. InvestNew Drugs 2006;24:195–202.
42. Sugawara T, Zaima N, Yamamoto A, Sakai S, Noguchi R, Hirata T.Isolation of sphingoid bases of sea cucumber cerebrosides and theircytotoxicity against human colon cancer cells. Biosci BiotechnolBiochem 2006;70:2906–12.
43. Baird RD, Kitzen J, Clarke PA, Planting A, Reade S, Reid A, et al.Phase I safety, pharmacokinetic, and pharmacogenomic trial of ES-285, a novel marine cytotoxic agent, administered to adult patientswith advanced solid tumors. Mol Cancer Ther 2009;8:1430–7.
44. Oskouian B, Saba J. Sphingosine-1-phosphate metabolism and intest-inal tumorigenesis: lipid signaling strikesagain.CellCycle 2007;6:522–7.
45. Kawamori T, Kaneshiro T, Okumura M, Maalouf S, Uflacker A, Bie-lawski J, et al. Role for sphingosine kinase 1 in colon carcinogenesis.FASEB J 2009;23:405–14.
46. Shida D, Fang X, Kordula T, Takabe K, Lepine S, Alvarez SE, et al.Cross-talk between LPA1 and epidermal growth factor receptorsmediates up-regulation of sphingosine kinase 1 to promote gastriccancer cell motility and invasion. Cancer Res 2008;68:6569–77.
47. Morin PJ, Sparks AB, Korinek V, Barker N, Clevers H, Vogelstein B,et al. Activation of beta-catenin-Tcf signaling in colon cancer bymutations in beta-catenin or APC. Science 1997;275:1787–90.
48. Davidson LA, Jiang YH, Derr JN, Aukema HM, Lupton JR, ChapkinRS. Protein kinase C isoforms in human and rat colonic mucosa. ArchBiochem Biophys 1994;312:547–53.
49. Ahn EH, Schroeder JJ. Sphinganine causes early activation of JNKand p38 MAPK and inhibition of AKT activation in HT-29 human coloncancer cells. Anticancer Res 2006;26:121–7.
50. King CC, Zenke FT, Dawson PE, Dutil EM, Newton AC, Hemmings BA,et al. Sphingosine is a novel activator of 3-phosphoinositide-depen-dent kinase 1. J Biol Chem 2000;275:18108–13.
51. Fyrst H, Oskouian B, Bandhuvula P, Gong Y, Byun HS, Bittman R,et al. Natural sphingadienes inhibit Akt-dependent signaling andprevent intestinal tumorigenesis. Cancer Res 2009;69:9457–64.
Anticancer Activities of Enigmol
www.aacrjournals.org Mol Cancer Ther; 10(4) April 2011 657
on August 4, 2014. © 2011 American Association for Cancer Research.mct.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2011; DOI: 10.1158/1535-7163.MCT-10-0754


















