
engrailed gene dosage determines whether certain recessive cubitus interruptus alleles exhibit...
10
DEVELOPMENTAL GENETICS 1934W49 (1996) engrailed Gene Dosage Determines Whether Certain Recessive cubitus interruptus Alleles Exhibit Dominance of the Adult Wing Phenotype in Drosophila JOHN LOCKE* AND SCOTT HANNA Department of Biological Sciences, University of Alberta, Edmonton, Alberta, Canacla ABSTRACT The cubitus interruptus (ci) lo- cus of Drosophila melanogaster is needed for nor- mal development. Some mutants of this gene result in embryonic lethality, while others just disrupt adult wing veins. While undertaking a genetic screen for additional ci mutations that affect the wing veins, we recovered a modifier mutation on chromosome two that produced a ci phenotype in recessive ci heterozygotes (cTecess’ve/ + ). We identified the modifier mutation as an allele of engrailed and have called it engrailed-enhancer of cubitus inter- ruptus (enEnc’). As a double heterozygote (en-/+; ci-/+) this new en allele dominantly generates a ci wing vein phenotype. As a double heterozygote, it also enhances the ci wing vein phenotype of the dominant alleles ciw and ace’, but not ciD. Other loss-of-function en alleles also enhance the ci phe- notype, with the en lethal alleles (and deletions) showing the strongest effect, while the homozy- gous viable en alleles show weaker enhancement. Strong en- alleles failed to induce a ti phenotype with heterozygotes of ci recessive lethal alleles 1/41 13, 1/41 17, or ciDrev, which are loss-of-function mutations. This supports a previous proposal that the ci wing vein phenotype is not due to loss of ci+ function, as would be expected for most recessive alleles. Instead, the adult wing vein abnormality is due to ectopic expression (or de-repression) of the ci transcript in the posterior compartment of the wing disc. We also observed that en-/+ heterozy- gotes could induce a ci phenotype in situations where the ci+ locus is either unpaired or hemizy- gous. Since loss of one en+ gene dose enhanced the ci phenotype, three doses of en+ were tested and found to suppress expression of the ci pheno- type in ci’ homozygotes and ciw heterozygotes. These observations show that correct regulation of the ci gene involves more than the simple interac- tion of upstream regulatory elements. Key words: Drosophila melanogaster, segment polarity genes, engrailed, cubitus interruptus, syn- thetic enhancement, genetic interactions, chromo- some pairing, pairing dependent gene repression, position effects. Dev Genet 19:340-349, 1996. 0 1996 Wiley-Liss, Inc. INTRODUCTION The cubitus interruptus (ci) locus in Drosophila mel- anogaster is a complex locus. The recessive viable wing vein mutants have interruptions in veins L4 and L5 of the adult wing giving a cubitus interruptus, or ci, phe- notype [Lindsley and Zimm, 19931. The dominant alle- les, ciD and cicd, also show ci wing vein interruptions and are recessive lethals. They do not complement Z(4)17 [Hochman, 19761; ciR also fails to complement another recessive lethal mutation, Z(4)13. The reces- sive lethal alleles 1(4)17 and 1(4)13 complement each other and each, as homozygotes, produce embryonic cu- ticle phenotypes typical of segment polarity genes [Orenic et al., 1987; Slusarski et al., 19951. Molecular cloning of the ci locus has identified a transcript that encodes a putative Zn-finger DNA bind- ing protein [Orenic et al., 19901. Sequence comparisons with other genes have shown similarity to the human GZi gene, which encodes a protein amplified in a human glioma [Ruppert et al., 19901, and the tra-l gene, which is involved in sex determination of C. elegans [Zar- kower and Hodgkin, 19921. The molecular lesions of some ci mutations have been described [Locke and Tartof, 1994; Slusarski et al., 1995; Schwartz et al., 19951. These studies indicate that there is a large reg- ulatory region upstream of the ci transcript and two separate elements, a proximal and a distal component, have been defined. The proximal fragment mediates embryonic expression, while the distal component me- diates anterior compartment expression in both em- bryos and discs [Schwartz et al., 19951. The molecular Received for publication 25 August 1995; accepted 27 June 1996. *Correspondence to Dr. John Locke, Department of Biological Sci- ences, University of Alberta, Edmonton, Alberta, Canada T6G 2E9. 0 1996 WILEY-LISS, INC.
-
Upload
john-locke -
Category
Documents
-
view
215 -
download
3
Transcript of engrailed gene dosage determines whether certain recessive cubitus interruptus alleles exhibit...

DEVELOPMENTAL GENETICS 1934W49 (1996)
engrailed Gene Dosage Determines Whether Certain Recessive cubitus interruptus Alleles Exhibit Dominance of the Adult Wing Phenotype in Drosophila JOHN LOCKE* AND SCOTT H A N N A Department of Biological Sciences, University of Alberta, Edmonton, Alberta, Canacla
ABSTRACT The cubitus interruptus (ci) lo- cus of Drosophila melanogaster is needed for nor- mal development. Some mutants of this gene result in embryonic lethality, while others just disrupt adult wing veins. While undertaking a genetic screen for additional ci mutations that affect the wing veins, we recovered a modifier mutation on chromosome two that produced a ci phenotype in recessive ci heterozygotes (cTecess’ve/ + ). We identified the modifier mutation as an allele of engrailed and have called it engrailed-enhancer of cubitus inter- ruptus (enEnc’). As a double heterozygote (en-/+; c i - /+) this new en allele dominantly generates a ci wing vein phenotype. As a double heterozygote, it also enhances the ci wing vein phenotype of the dominant alleles ciw and ace’, but not ciD. Other loss-of-function en alleles also enhance the ci phe- notype, with the en lethal alleles (and deletions) showing the strongest effect, while the homozy- gous viable en alleles show weaker enhancement. Strong en- alleles failed to induce a t i phenotype with heterozygotes of ci recessive lethal alleles 1/41 13, 1/41 17, or ciDrev, which are loss-of-function mutations. This supports a previous proposal that the ci wing vein phenotype is not due to loss of ci+ function, as would be expected for most recessive alleles. Instead, the adult wing vein abnormality is due to ectopic expression (or de-repression) of the ci transcript in the posterior compartment of the wing disc. We also observed that en-/+ heterozy- gotes could induce a ci phenotype in situations where the ci+ locus is either unpaired or hemizy- gous. Since loss of one en+ gene dose enhanced the ci phenotype, three doses of en+ were tested and found to suppress expression of the ci pheno- type in ci’ homozygotes and ciw heterozygotes. These observations show that correct regulation of the ci gene involves more than the simple interac- tion of upstream regulatory elements.
Key words: Drosophila melanogaster, segment polarity genes, engrailed, cubitus interruptus, syn- thetic enhancement, genetic interactions, chromo-
some pairing, pairing dependent gene repression, position effects. Dev Genet 19:340-349, 1996. 0 1996 Wiley-Liss, Inc.
INTRODUCTION The cubitus interruptus (ci) locus in Drosophila mel-
anogaster is a complex locus. The recessive viable wing vein mutants have interruptions in veins L4 and L5 of the adult wing giving a cubitus interruptus, or ci, phe- notype [Lindsley and Zimm, 19931. The dominant alle- les, ciD and cicd, also show ci wing vein interruptions and are recessive lethals. They do not complement Z(4)17 [Hochman, 19761; ciR also fails to complement another recessive lethal mutation, Z(4)13. The reces- sive lethal alleles 1(4)17 and 1(4)13 complement each other and each, as homozygotes, produce embryonic cu- ticle phenotypes typical of segment polarity genes [Orenic et al., 1987; Slusarski et al., 19951.
Molecular cloning of the ci locus has identified a transcript that encodes a putative Zn-finger DNA bind- ing protein [Orenic et al., 19901. Sequence comparisons with other genes have shown similarity to the human GZi gene, which encodes a protein amplified in a human glioma [Ruppert et al., 19901, and the tra-l gene, which is involved in sex determination of C. elegans [Zar- kower and Hodgkin, 19921. The molecular lesions of some ci mutations have been described [Locke and Tartof, 1994; Slusarski et al., 1995; Schwartz et al., 19951. These studies indicate that there is a large reg- ulatory region upstream of the ci transcript and two separate elements, a proximal and a distal component, have been defined. The proximal fragment mediates embryonic expression, while the distal component me- diates anterior compartment expression in both em- bryos and discs [Schwartz et al., 19951. The molecular
Received for publication 25 August 1995; accepted 27 June 1996.
*Correspondence to Dr. John Locke, Department of Biological Sci- ences, University of Alberta, Edmonton, Alberta, Canada T6G 2E9.
0 1996 WILEY-LISS, INC.

en ALLELES ENHANCE THE WING PHENOTYPE OF ci ALLELES 341 aberrations of the recessive wing vein ci alleles are consistent with the reduction or loss in the activity of a negative regulatory element within the distal regula- tory fragment [Locke and Tartof, 1994; Schwartz et al., 19951. Loss of this negative regulatory element leads to ectopic expression of the ci- transcript in the posterior compartment of the wing disc and is thought to be re- sponsible for the loss of wing veins seen in the ci phe- notype [Slusarski et al., 1995; Schwartz et al., 19951.
The recessive wing vein mutations of ci also show unusual position effects when heterozygous with cer- tain chromosome rearrangements (e.g., translocations or inversions) that move chromosome 4, the location of c i + , to distal regions on other chromosomes [Dubinin and Sidorov, 19341, Such chromosome rearrangements, called R(ci+), exhibit a ci phenotype when heterozy- gous with ci' or other recessive ci alleles. This ci effect is unusual in that if R(ci+) is heterozygous with a de- letion of the ci region or if it is homozygous (R(ci+)l R(ci+)), no ci wing vein phenotype is observed. This is unlike typical position effect variegation rearrange- ments, which express a mutant phenotype (variega- tion) in such situations. These position effects suggest that interruptions in the physical pairing at the ci lo- cus on homologous chromosomes are needed for the mutant ci phenotype [Ephrussi and Sutton, 1944; Locke and Tartof, 19941. Consequently, the ci effect has been regarded as a transvection-like phenomenon, in which correct gene expression may be dependent on homologous chromosome pairing [Tartof and Henikoff, 1991; Locke and Tartof, 19941. The DNA sequences and proteins involved in this transvection, or chromosome pairing, are unknown but may be obtained from an analysis of the ci locus and its mutants.
When screening for new ci alleles, we recovered a second site dominant enhancer of ci, which generated a strong ci wing vein phenotype in ci'l i heterozygotes which normally have wild type wings. In Drosophila and other organisms, such genetic interactions among mutations at different loci have been used to identify genes that regulate the same biological pathways [re- viewed by Guarante, 19931. We characterized this mod- ifier mutation further and found it to be an allele of engrailed (en). Interactions between ci' and en' alleles have been reported previously [House, 1953a,bl; how- ever, a pseudo-dominant effect on ci heterozygotes was not seen. We have re-examined the interactions of ci and en in the context of a wider spectrum of mutant alleles that have been described since the original ob- servations (see under Materials and Methods) [Linds- ley and Zimm, 19921. The interactions described here are stronger and more extensive than those previously reported [House, 1953a,bl and indicate that reduced en+ gene dose produced dominant expression of the ci wing vein phenotype in heterozygotes for recessive ci wing vein alleles. Furthermore, en-/ + heterozygotes also showed a ci phenotype with normal ci+ alleles that are unpaired, indicating that chromosome pairing may
facilitate the correct regulation (repression) of ci in the posterior compartment of the adult wing.
MATERIALS AND METHODS Drosophila Cultures
Drosophila melunogaster mutations used in this study, unless otherwise cited, are described by Lindsley and Grell [19681 or Lindsley and Zimm [19921. Cul- tures were grown at 21°C (unless otherwise stated) on standard yeastlsucrose medium (each liter contains 10 g agar, 100 g sucrose, 100 g brewer's yeast, 100 ml of 1 glL chloramphenicol, 10 ml proprionic acid, 4.3 g NaH2P0,, and 2.7 g Na2HP04).
Mutagenesis and Drosophila Crosses The mutagenesis for ci alleles involved irradiation
(4,000 rad of y-rays) of Oregon-R males, which were then crossed to dp; e; ci', e y virgin females. The het- erozygous progeny were screened for interruptions in wing vein L4, the fourth-longitudinal or cubital vein (the ci wing phenotype). Putative mutations were back- crossed to dp; e; ci', e y to confirm mutant transmis- sion and monitor for the co-segregation of other chro- mosomes. Co-segregation would indicate the mutation was associated with a translocation and would there- fore most likely be a ci-position effect.
Mutations Involving engruiled and cubitua interruptus
engrailed alleles have been divided into four classes that differ in their phenotypic interactions [Lindsley and Zimm, 19921. The original en1 mutation [Eker, 19291, which constitutes the first class, is homozygous viable and, among other defects, displays a variety of vein alterations in the posterior compartment of the wing. Its phenotype is not due to simple hypomorphic expression, but to a altered pattern and level of expres- sion in the wing disc [Brower, 1986; Condie and Brower, 19891. It is associated with an insert of 7-kb middle repetitive DNA element located 5' to the en- grailed transcript. The second en class consists of the recessive lethal alleles that have no cytologically visi- ble chromosome aberration (e.g., en4, en1034). The third class includes those en alleles that have cytologically visible aberrations and are recessive lethal (e.g., enpsL, Dff2R)ensFx). A new allele described in this study, In(2R)enEnCC, falls into the third class. The fourth class contains chromosome aberrations that are hemizygous viable (at the engrailed locus), yet do not complement en' (e.g., Dfl2R)edO and EnEs).
Tp(2;3)en+ is a complex rearrangement in which the engrailed locus and adjacent regulatory elements have been deleted from chromosome 2, to generate DfT2R)en28, and inserted into chromosome 3, to make Dp(2;3)en28 [Eberlein and Russell, 19831. This trans- position was used to generate progeny with 1, 2, OF 3 doses of en+.

342 LOCKE AND HANNA
There is also a variety of ci alleles. The dominant wing vein alleles (ciD, ciw, and cice2), recessive wing vein alleles (ci', ci36z, ci57g), and recessive lethal alleles (1(4)17 and 1(4)13), are now considered part of one com- plex locus on chromosome 4 [Hochman, 1976; Orenic et al., 1987; Locke and Tartof, 1994; Schwartz et al., 1995; Slusarski et al., 19951. The two recessive lethal alleles define two independent recessive lethal complementa- tion groups; both appear cytologically normal. No mo- lecular aberrations were identified for either lethal al- lele by Southern analysis using cloned fragments in the ci region [Locke and Tartof, 19941. These observa- tions are consistent with these being point mutant, loss-of-function, recessive lethal alleles. The revertants of ciD, ciDmGA, and ciDreugA, have reverted the dominant ci wing vein phenotype but retained the recessive le- thals and have not acquired a recessive ci wing vein phenotype. Furthermore, molecular analysis shows that they have. altered restriction fragments that are consistent with a small deletion in or near the ci- tran- script [Locke and Tartof, 19941.
Df4)M""f is deleted for the cytogenetic region 101E; 102B10-17, which includes the ci gene and an adjacent Minute locus. Df4)f13a is deleted for the cytogenetic region 101F2-102A1; 102A3-5, which includes the Minute locus, but not the ci gene; it nevertheless acts as a recessive ci allele through a second site mutation or a position effect [Locke and Tartof, 19941.
The s i m 4 chromosome is aDrosophila simulans chro- mosome 4 that has been introgressed into Drosophila melanogaster. Its genetic behavior is characteristic of a ci position effect allele [Muller and Pontecorvo, 1940a,bl, which could result from an inversion (relative to chromosome 4 of D. melanogaster) that involves a substantial number of the bands in the euchromatic portion of chromosome 4 [Horton, 19391. Furthermore, observations indicate s i m 4 remains unpaired in the polytene chromosomes of s im4lmel4 hybrids [Slizyn- ski, 19411.
T(1;4)wd, wd (3C2;101F1-2) is a reciprocal trans- location between chromosomes X and 4. It shows a var- iegated eye phenotype for white (on the X chromosome) and wing phenotype for ei (on chromosome 4) due to position effects.
The la& enhancer-trap insertion pciD-lacZlw +I [Eaton and Kornberg, 19901 has a P-element inserted upstream from the ci transcript described by Orenic et al. [1991]. pciD-lacZ[w +I homozygotes or heterozy- gotes with recessive ci wing vein alleles normally show no ci wing vein phenotype.
Scoring the cubitus interruptus Wing Vein Phenotype
The extent of ci mutant wing vein phenotype varies in penetrance and expressivity. The range of expres- sivity extends from weak vein thinning to almost com- plete loss of the L4 vein. We found the 0-4 numerical system for ci mutant phenotype used by Stern and
Fig, 1. Photographs of adult wings showing cubitus inter- ruptus phenotypes. a: An adult en+/en+; ci+lci+ wing shows a wild-type wing phenotype. b An example of a weak ci wing phenotype used in the scoring of ci phenotypes shows the ex- tra vein formation in the re- gion near or between the cross- veins. c: The mutant ci'lci' wing has an interruption of L4, typical for the ci wing vein phe- notype. d: The double heterozy- gote enEncL/ + ; c i 6 8 + shows a ci wing phenotype.
Kodani [1955] quantifies only the mutant aspects of distal L4 vein structure but fails to communicate the diversity of vein abnormalities observed in the ci phe- notype or the subtleties of a weak ci phenotype that is seen, especially in the proximal region of L4. Further- more, the 0-4 numerical system can not be considered a linear quantitative estimate of the extent of ci mutant phenotype and therefore is not amenable to simple sta- tistical analysis. Consequently we have scored the ci phenotypes as wild type, weak, or strong in our crosses for examining genetic interactions. Wild type showed no vein abnormalities and the wing was completely wild type (Fig. 1A). A weak ci phenotype included L4 vein thinning or the appearance of extra vein material in the region of L4 near the cross-veins (Fig. 1B). A strong ci phenotype included any break in L4, proximal or distal (Fig. 1C). Vein aberrations described in the weak ci class were also seen in those wings classified as strong ci (note L4 near the proximal cross vein in Fig. 1C).
Genetic Mapping Recombination mapping of the engrailed mutant,
, described in this study used these chromosome 2 markers: S (2-1.2; 21E1-21, Sp (2-22.0; 28Al-B4), Tfz (2-53.6; 37A3-6), nfl(2-79.6; 54A-55A), and Pin (2- 107.3; SOCS-DZ). en is located at map position 2-62.0; 48A2. The centromere is at approximately 55. The presence of the enEnci mutation was identified in the progeny as a ci wing vein phenotype in a cill+ back- ground. Crosses were grown at room temperature (-2 1°C).
Cytology Flies with the enEnci allele were crossed to wild type
(Oregon-R). The salivary gland polytene chromosomes
enEnci

en ALLELES ENHANCE THE WING PHENOTYPE OF ci ALLELES 343
Ag. 2, Photographs showing the enE"' mutation and phenotype. a: Salivary gland polytene chromosome of an enEmi/ + heterozygote showing a region of stretched chromosome (arrow) that connects re- gion 48A (the location of ergmiled) with the chromocenter through an inversion. Chromosome 4 (4) appears normal. b Adult wings of enEmci/ en' showing vein defects in the posterior compartment that include the complete disruption of L4 and L5 and the presence of abnormal anterior-like bristles on the posterior wing margin (c). d: The legs of en""'/en' males show duplications of the sex comb on the prothoracic leg.
of heterozygous third instar were examined for chro- mosome aberrations and photographed with phase-con- trast illumination on a Zeiss Axiophot microscope.
RESULTS Recovery of enE"' Mutant
During a small-scale mutagenesis to induce new ci alleles (see under Materials and Methods), we recov- ered seven putative ci mutants among approxi- mately 8,850 progeny. Upon further examination six were nontransmitting, lost, or translocations (presum- ably ci position effects). However one mutant, called In(2R)enE"' (hereafter enEnci), segregated with chro- mosome 2, but was not linked to chromosome 4, as would be expected for a T12;4) translocation. Further crosses showed this second chromosome mutation pro- duced a ci phenotype in ci' e f / + + heterozygotes, which usually have wild type wing veins (Fig. 1D). Expression of ep, another chromosome 4 recessive mu- tation, was not similarly modified, indicating that the modifer's affect was specific to ci.
Preliminary genetic mapping positioned the ci en- hancing mutant to the right of Tft and left of n d . Further mapping proved difficult for two reasons. First, the expression of a ci phenotype was only about 85% penetrant and therefore some mutants could not be classified correctly. Second, recombination was sup- pressed along chromosome 2, particularly on 2R. Cyto- logical examination of polytene chromosomes showed an inversion with breakpoints a t 48A (Fig. 2A) and the chromocenter (142R)40;48A). Chromosome 4 polytene banding was cytologically normal.
A breakpoint a t 48A suggested that this mutation might be an engrailed allele, so we tested for com- plementation with engrailed mutants. Crosses with various engraded alleles showed that this mutation
does not complement engrailed recessive lethal alleles en4, , Df12R)enSFx3' , en4sL, DflZR)en28, or enrPry+Xho251. Furthermore, enEnCilenl heterozygote adults have a posterior specific wing phenotype (Fig. 2B) typical of previously described enLethazlenl heterozy- gotes [Condie and Brower, 19891. This includes bristles characteristic of the anterior wing margin on the pos- terior margin (Fig. 2C). Furthermore, males have a duplication of sex combs on the prothoracic legs, an- other characteristic of enkthazlen1 heterozygotes (Fig. 2D). Heterozygotes between enEnci and Dfl2R)en30 or enEs have reduced adult viability (to -10%) and wings with vein defects in the posterior compartments only. The anterior compartment is wild type. Consequently we have called this mutation engrailed-Enhancer of cubitus interruptus (In(2R)enEmi 40;48A).
We have not identified the location of the breakpoint at the molecular level but, based on the interactions with other en alleles, enEnci is a class I11 allele (see under Materials and Methods); therefore, we would ex- pect a break in the en lethal region between -35 kb to + 10 kb in the map of Kuner et al. [19851. Since enEnci may have repositioned the engrailed locus to the prox- imal portion of chromosome 2 near the chromocenter, enEnci itself could be a position effect. We consider this unlikely, as enEnci is completely lethal over all enzethaz alleles tested and enEnci has similar effects to all other en alleles on the phenotype of ci alleles (see below).
enEmi Enhances the Wing Vein Phenotype of All Heterozygous Recessive ei Wing Vein Mutations
Since enEnci/ + enhanced the ci phenotype in cill+ , we tested other homozygous viable ci alleles (ci67g, ci""', and ciw) as double heterozygotes with enExi. In these enEnci I + ; ci-/ -i- double heterozygotes, the appearance of a ci phenotype is temperature dependent (Table 1). At 18"C, all three recessive ci alleles (cil, ci361, and ei57g), as double heterozygotes with enEnci show 100% penetrance of the ci phenotype, while at 25"C, pene- trance is only about 50%. We attribute this difference to variation in ci, rather than en, as expression of ho- mozygous recessive ci alleles is temperature sensitive and the phenotype is typical ci, not en. In these and other crosses, the mutant ci phenotype was qualita- tively typical of ci, in that usually only L4 is affected. It was distinct from the en wing phenotype of en' ho- mozygotes or enl/enzethal heterozygotes, as reported by Condie and Brower [ 19891, where much more than just L4 is affected.
The ciw allele, itself, is dominant and its phenotype temperature sensitive; a t 18"C, 56% of the ciwl + have a strong ci phenotype, but at 25°C all are wild type (Table 1). Here, enEnci/ t ; ciw/ + individuals have 100% ci penetrance at 25°C.
The allele ci' responds least to enEnci enhancement. The strength of the ci alleles in showing a ci phenotype in heterozygotes under these conditions was ci' <ci36z <ci57g << ciw, an order that is repeated with other en

344 LOCKE AND HA"A
TABLE 1. Effect of Temperature on the ci Wing Vein Pbenotwe of enE"'/ + xi/+ Double Heterozsgotes
Temperatureb Genotype Phenotype" 18°C (%) 21°C (%) 25°C (%) enEn~i I+ ; Strong ci 76 47
Wild type ci 0 22
CyOl+ Strong ci 0 0 ci'l+ Weak ci 0 0
Wild type ci 100 100
enEnci I + ; Strong ci 100 81 ci3"'/+ Weak ci 0 19
Wild type ci 0 0
C 0/+ ; Strong ci 0 0
Wild type ci 100 100
cil/ + Weak ci 23 31
(n = 114) (n = 106)
(n = 94) (n = 86)
(n = 63) (n = 164)
c&/+ Weak ci 0 0
(n = 52) (n = 163) I+ ; Strong ci 100 87
~ i ~ ~ g 1 - t Weak ci 0 13 enEnci
Wild type ci 0 0
Cy0l-t; Strong ci 0 0 cP7gl + Weak ci 0 0
Wild type ci 100 100
EnEncil+ ; Strong ci 92 100 ciw/ + Weak ci 8 0
Wild type ci 0 0
ci I + Weak ci 17 10 Wild type ci 26 62
(n = 31) (n= 157)
(n = 49) (n = 183)
(n=26) (n=117) CGOI + ; Strong ci 56 28
(n = 23) (n = 112)
T h e phenotype of + I + ; + /ci is wild type. bRearing temperature.
13 8
79 (n = 72)
0 0
100 (n = 78)
12 14 74
(n = 77) 0 0
100 (n = 68)
37 27 37
(n = 79) 0 0
100 (n = 67)
100 0 0
0 0
100 (n = 6)
(n = 10)
alleles (see below). This is consistent with ci' being the weakest of the three recessive ci alleles in its ability to produce a ci phenotype as a homozygote.
In these double heterozygotes with enEnci and ci, the 21°C crosses gave incomplete penetrance and variable expressivity, suggesting that the ci phenotype is at a balance point for detection and that 21°C should prove a good temperature for assaying genetic interactions. Hence the remaining crosses investigating ci and en interactions were grown at 21°C.
Other en Alleles Also Enhance the Wing Vein Phenotype of ci
To see whether the enhancement of ci heterozygotes was a general feature of en alleles, we tested represen- tatives of each of the four en classes. Crosses were made between en alleles balanced by a Cy marked chromo- some 2 (SM5 or CyO) and ci alleles (as homozygotes or balanced with a marked chromosome 4; ey" or M57g). Progeny were scored for ci phenotype and the results of various e n / + ; cil+ double heterozygotes is shown in Table 2. Since a ci phenotype was not seen in CyO/+ ;
c i /+ control progeny (except with the dominant ci al- leles), their values are not shown in Table 2.
All en alleles can enhance the ci phenotype of c i l i - heterozygotes, but not all are equally effective. en', which has previously been reported to enhance ci' ho- mozygotes [House, 1953a1, only weakly enhances the homozygous viable recessive ci alleles ci', ci3"', and ci"'", with most progeny (>70%) retaining a wild-type wing phenotype. However, the recessive lethal en alle- les (en4, enro, enSFX31 , and enEmi) have a strong affect on the ci heterozygotes, with most individuals showing a ci phenotype. The only exceptions to this strong interac- tion are among the ci l l+ individuals; ci' is the weakest allele. The fourth class of en allele, DfZR)en3O (viable as a heterozygote with enzethaz), is similar to en', in that it has a weak affect on ci heterozygotes. Like the ci alleles, the en alleles can be ranked with regard to their ability to induce a ci phenotype. This ranking is e n ' r D f 2 R ) e n 3 0 1 e n ~ ~ e n ~ ~ ' < ~ n ~ ~ ~ ~ ~ <enlo.
Interactions with the dominant ci alleles, ciD, ci", and cice2, were also tested. For ciD, the penetrance was complete and the expressivity consistent both with or without the en alleles; therefore, no alteration in the wing vein phenotype was seen. Penetrance of ciw/+ heterozygotes is normally incomplete (Table 1) but is enhanced to -100% in any en/+ heterozygote. Our strain of cice2 rarely expresses the dominant wing vein phenotype (<1%); therefore, we determined that ciCdI + heterozygotes behaved like the recessive ci al- leles, in that en' only weakly enhanced, while the
en Alleles Fail to Enhance the ci Phenotype of Cytologically Normal, Recessive Lethal ci Alleles
The two recessive lethal alleles, 1(4)17 and 1(4)13, are probably point mutant, loss-of-function, recessive lethal alleles that define two independent recessive le- thal complementation groups associated with ci. We tested the ability of en alleles to produce a ci phenotype in U4)13/+ and 1(4)17/+ heterozygotes, as well as with heterozygotes of two revertants of ciD, ciDmu3A , and
. None of these loss-of-function alleles, as het- erozygotes, showed any ci phenotype in the presence of any en- allele heterozygotes (Table 2).
en Alleles Also Generate a ci Wing Vein Phenotype With Chromosome 4 Rearrangements
That Unpair a Wild Type ci Allele Chromosome rearrangements that do not comple-
ment recessive ci wing vein alleles (e.g., cil) were also tested as heterozygotes for ci wing vein phenotype in en-/ + heterozygotes (Table 2). In these rearrange- ments, ci is wild type, and the lack of complementation has been attributed to a position effect that involves a lack of chromosome pairing of the two homologs at ci [Ephrussi and Sutton, 1944; Locke and Tartof, 19941.
Rearrangement T(1;4) wm5 represents a typical R(ci +), which gives a ci-position effect. T(1;4)wd
alleles showed much stronger enhancement. enlethal
ciDrev9A
en ALLELES ENHANCE THE WING PHENOTYPE OF ci ALLELES 345
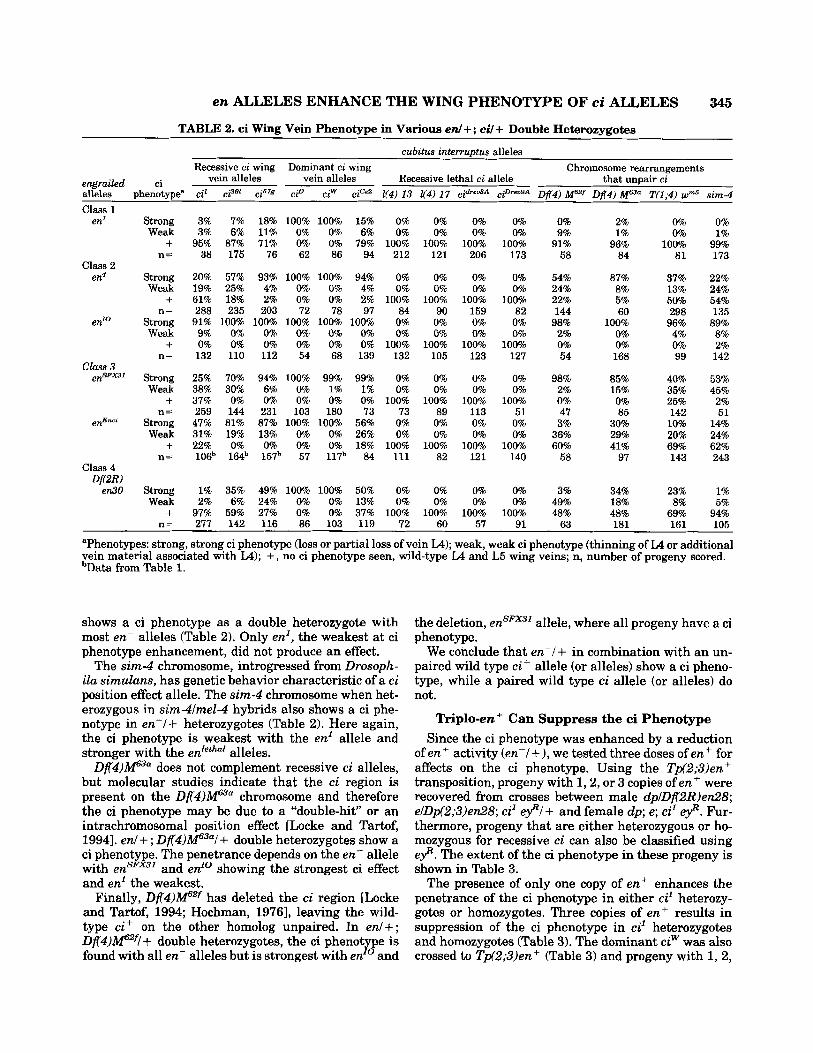
TABLE 2. ci Wing Vein Phenotype in Various en/+; ci/+ Double Heterozygotes
cubitus interruptus alleles
Class 1 en*
Class 2 en4
enlo
C h s s 3 enSFX31
enEIZC'
Class 4 Dff2R)
en30
Strong Weak
+ n =
Strong Weak
n = Strong Weak
n =
Strong Weak
+ n =
Strong Weak
n =
+
+
+
Strong Weak
3% 3% 95% 38
20% 19% 61% 288 91% 9% 0% 132
25% 38% 37% 259 47% 31% 22% 106b
1% 2%
7% 18% 100% 100% 15% 6% 11% 0% 0% 6% 87% 71% 0% 0% 79% 175 76 62 86 94
57% 25% 18% 235
100% 0% 0% 110
93% 4% 2% 203
100% 0% 0% 112
100% 0% 0% 72
100% 0% 0% 54
100% 0% 0% 78
100% 0% 0% 68
94% 4% 2% 97
100% 0% 0% 139
70% 30% 0% 144 81% 19% 0% 164b
94% 100% 99% 6% 0% 1% 0% 0% 0% 231 103 180 87% 100% 100% 13% 0% 0% 0% 0% 0% 157b 57 117b
99% 1% 0% 73
56% 26% 18% 84
35% 49% 100% 100% 50% 6% 24% 0% 0% 13%
0% 0%
100% 212
0% 0%
100% 84 0% 0%
100% 132
0% 0%
100% 73 0% 0%
100% 111
0% 0%
0% 0%
100% 121
0% 0%
100% 90 0% 0%
100% 105
0% 0%
100% 89 0% 0%
100% 82
0% 0%
0% 0%
100% 206
0% 0%
100% 159 0% 0%
100% 123
0% 0%
100% 113 0% 0%
100% 121
0% 0%
0% 0%
100% 173
0% 0%
100% 82 0% 0%
100% 127
0% 0%
100% 51 0% 0%
100% 140
0% 0%
0% 9% 91% 58
54% 24% 22% 144 98% 2% 0% 54
98% 2% 0% 47 3% 36% 60% 58
3% 49%
2% 1% 96% 84
87% 8% 5% 60
100% 0% 0% 168
85% 15% 0% 85
30% 29% 41% 97
34% 18%
0% 0%
100% 81
37% 13% 50% 298 96% 4% 0% 99
40% 35% 25% 142 10% 20% 69% 143
23% 8% ~~
+ 97% 59% 27% 0% 0% 37% 100% 100% 100% 100% 48% 48% 69% n= 277 142 116 86 103 119 72 60 57 91 63 181 161
aPhenotypes: strong, strong ci phenotype (loss or partial loss of vein LA); weak, weak ci phenotype (thinning of L4 or additional vein material associated with L4); + , no ci phenotype seen, wild-type L4 and L5 wing veins; n, number of progeny scored. bData from Table 1.
Recessive ci wing Dominant ci wing Chromosome rearranger
0% 1% 99% 173
22% 24% 54% 135 89% 8% 2% 142
53% 45% 2% 51
14% 24% 62% 243
1% 5% 94% 105
shows a ci phenotype as a double heterozygote with most en- alleles (Table 2). Only en', the weakest at ci phenotype enhancement, did not produce an effect.
The sim-4 chromosome, introgressed from Drosoph- ila simulans, has genetic behavior characteristic of a ci position effect allele. The sim4 chromosome when het- erozygous in sirn4lmel-4 hybrids also shows a ci phe- notype in en-/ + heterozygotes (Table 2). Here again, the ci phenotype is weakest with the en' allele and stronger with the enletha' alleles.
D f 4 ) P 3 " does not complement recessive ci alleles, but molecular studies indicate that the ci region is present on the Df('4)Pu chromosome and therefore the ci phenotype may be due to a "double-hit" or an intrachromosomal position effect [Locke and Tartof, 19941. en1 + ; Df4)f13al + double heterozygotes show a ci phenotype. The penetrance depends on the en- allele with ensFx3' and enro showing the strongest ci effect and enL the weakest.
Finally, D f 4 ) P f has deleted the ci region [Locke and Tartof, 1994; Hochman, 19761, leaving the wild- type ci+ on the other homolog unpaired. In en/+; Dfl)M62f! + double heterozygotes, the ci phenotype is found with all en- alleles but is strongest with en" and
the deletion, ensFx3' allele, where all progeny have a ci phenotype.
We conclude that en-/+ in combination with an un- paired wild type c i+ allele (or alleles) show a ci pheno- type, while a paired wild type ci allele (or alleles) do not.
Triplo-en+ Can Suppress the ci Phenotype Since the ci phenotype was enhanced by a reduction
of en+ activity (en-/ + ), we tested three doses of en+ for affects on the ci phenotype. Using the Tp(2;3)en+ transposition, progeny with 1,2, or 3 copies of enc were recovered from crosses between male dplDfT2RJen28; elDp(2;3)en28; ci' e y l + and female dp; e; ci' e y . Fur- thermore, progeny that are either heterozygous or ho- mozygous for recessive ci can also be classified using e y . The extent of the ci phenotype in these progeny is shown in Table 3.
The presence of only one copy of en+ enhances the penetrance of the ci phenotype in either ci' heterozy- gotes or homozygotes. Three copies of en+ results in suppression of the ci phenotype in ci' heterozygotes and homozygotes (Table 3). The dominant ciw was also crossed to Tp(2;3)en+ (Table 3) and progeny with 1,2,
346 LOCKE AND HANNA
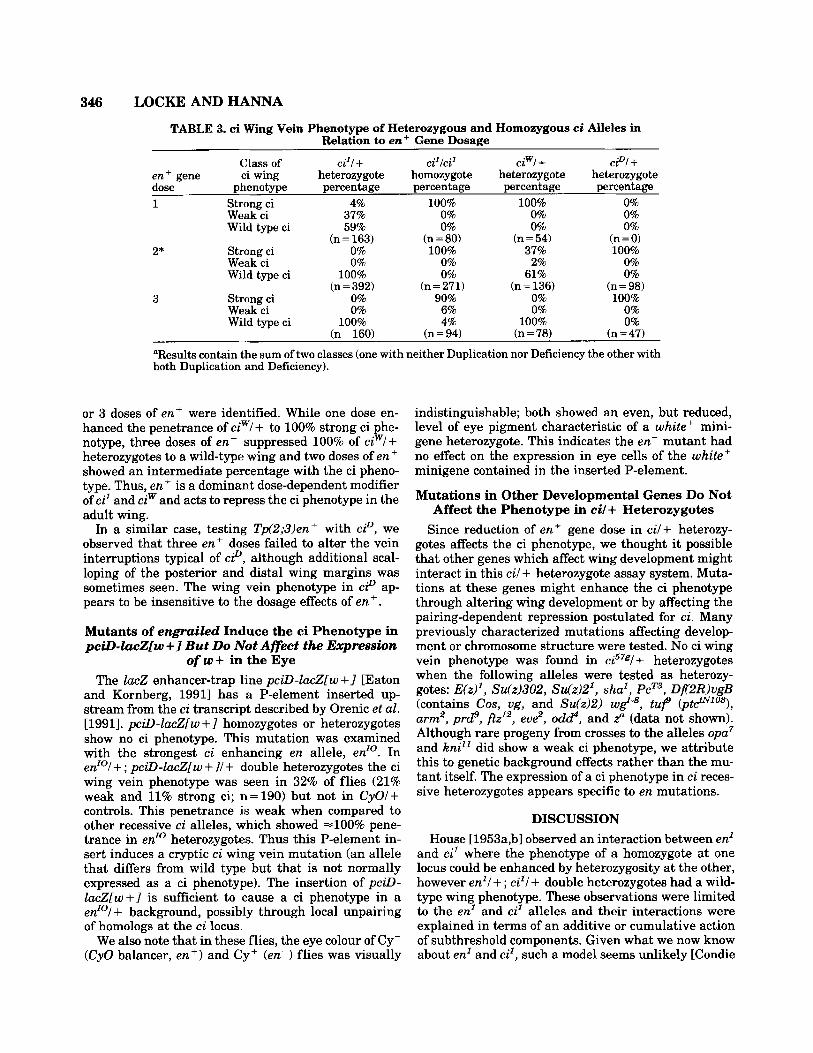
TABLE 3. ci Wing Vein Phenotype of Heterozygous and Homozygous ei Alleles in Relation to enz+ Gene Dosage
Class of ci'I + ci'Ici' ciw/ + ciDI +
1 Strong ci 4% 100% 100% 0% Weak ci 37% 0% 0% 0% Wild type ci 59% 0% 0% 0%
(n = 163) (n = 80) (n = 54) (n = 0) 2* Strong ci 0% 100% 37% 100%
Weak ci 0% 0% 2% 0% Wild type ci 100% 0% 61% 0%
3 Strong ci 0% 90% 0% 100% Weak ci 0% 6% 0% 0% Wild type ci 100% 4% 100% 0%
en+ gene ci wing heterozygote homozygote heterozygote heterozygote dose phenotype percentage percentage percentage percentage
(n = 392) (n = 271) (n = 136) (n = 98)
(n = 160) (n = 94) (n = 78) (n = 47)
"Results contain the sum of two classes (one with neither Duplication nor Deficiency the other with both Duplication and Deficiency).
or 3 doses of ent were identified. While one dose en- hanced the penetrance of ciwl + to 100% strong ci phe- notype, three doses of en' suppressed 100% of ciwl+ heterozygotes to a wild-type wing and two doses of e n f showed an intermediate percentage with the ci pheno- type. Thus, en+ is a dominant dose-dependent modifier of cz' and ciw and acts to repress the ci phenotype in the adult wing.
In a similar case, testing Tp(2;3)en+ with ciD, we observed that three enc doses failed to alter the vein interruptions typical of ciD, although additional scal- loping of the posterior and distal wing margins was sometimes seen. The wing vein phenotype in ciD ap- pears to be insensitive to the dosage effects of en+.
Mutants of engraikd Induce the ci Phenotype in pciLl-lmZ[w + I But Do Not Affect the Expression
of w + in the Eye The lacZ enhancer-trap line pciD-lacZ[w +I [Eaton
and Kornberg, 19911 has a P-element inserted up- stream from the ci transcript described by Orenic et al. [1991]. pciD-lacZlw +I homozygotes or heterozygotes show no ci phenotype. This mutation was examined with the strongest ci enhancing en allele, enro. In en'o! + ; pciD-lacZ[w +I! + double heterozygotes the ci wing vein phenotype was seen in 32% of flies (21% weak and 11% strong ci; n = 190) but not in CyO/+ controls. This penetrance is weak when compared to other recessive ci alleles, which showed ~ 1 0 0 % pene- trance in en'* heterozygotes. Thus this P-element in- sert induces a cryptic ci wing vein mutation (an allele that differs from wild type but that is not normally expressed as a ci phenotype). The insertion o f p c D - lacZ[w+l is sufficient to cause a ci phenotype in a enIol + background, possibly through local unpairing of homologs at the ci locus.
We also note that in these flies, the eye colour of Cy- (CyO balancer, en+) and Cy+ (en-) flies was visually
indistinguishable; both showed an even, but reduced, level of eye pigment characteristic of a whitef mini- gene heterozygote. This indicates the en- mutant had no effect on the expression in eye cells of the whitef minigene contained in the inserted P-element.
Mutations in Other Developmental Genes Do Not Affect the Phenotype in ci/+ Heterozygotes
Since reduction of en+ gene dose in cil+ heterozy- gotes affects the ci phenotype, we thought it possible that other genes which affect wing development might interact in this cil+ heterozygote assay system. Muta- tions at these genes might enhance the ci phenotype through altering wing development or by affecting the pairing-dependent repression postulated for ci. Many previously characterized mutations affecting develop- ment or chromosome structure were tested. No ci wing vein phenotype was found in ci57gl + heterozygotes when the following alleles were tested as heterozy- gotes: E(z)', Su(z)302, Su(z)2', sha', PcT3, Df2R)vgB (contains Cos, vg, and Su(z)2) w $ ' ~ , tuf (ptc"'oB), arm2, prd', fid2, e v 8 , od&, and z" (data not shown). Although rare progeny from crosses to the alleles opa7 and knill did show a weak ci phenotype, we attribute this to genetic background effects rather than the mu- tant itself. The expression of a ci phenotype in ci reces- sive heterozygotes appears specific to en mutations.
DISCUSSION House [1953a,bl observed an interaction between en'
and ci' where the phenotype of a homozygote a t one locus could be enhanced by heterozygosity a t the other, however en'l + ; till + double heterozygotes had a wild- type wing phenotype. These observations were limited to the en1 and cil alleles and their interactions were explained in terms of an additive or cumulative action of subthreshold components. Given what we now know about en' and ci', such a model seems unlikely [Condie

en ALLELES ENHANCE THE WING PHENOTYPE OF ci ALLELES 347
and Brower, 1989; Locke and Tartof, 1994; Schwartz et al., 1995; Slusarski et al., 19951.
The ci transcript is normally only expressed in the anterior compartment of the wing disc [Eaton and Korn- ber, 1990; Schwartz et al., 1995; Slusarski et al., 19951. The ci phenotype, which involves loss of wing vein struc- tures in the posterior compartment, is due to ectopic expression of the ci transcript in the posterior compart- ment cells [Eaton and Kornberg, 1990; Schwartz et al., 1995; Slusarski et al., 19951. This ectopic expression may be due to impairment of a cis-acting signal from an upstream silencing element or system normally func- tioning in the posterior compartment cells. Such a system could account for the overlap between the pleio- tropic en phenotype and the specific ci wing vein phe- notype. In both cases, the loss of vein L4 is due to the inability of en to prevent ectopic expression of ci, which produces the ci phenotype in the posterior wing com- partment.
In a search for cis-acting element upstream from the ci- transcript, Schwartz et al. (1995) assayed cloned fragments for regulatory elements with a lac2 reporter construct and for en protein binding using footprinting assays. Their results suggest en probably affects ci through direct transcriptional regulation. The reporter constructs exhibited proper en-dependent repression of ci in posterior compartment cells when two doses of en are present. However, in this report if only one func- tional copy of en is present (e.g., en-/ +), normal alleles of ci are unable to be repressed properly if they are chromosomally unpaired. In Table 2, the example of en''/ + ; p2? + double heterozygote (one ci' allele is chromosomally unpaired) shows a mutant ci wing phe- notype, while en1'/ f ; l(4)I 71 + (one ci+ allele paired with a ci- loss of function point mutant), has a normal wing vein. A simple cis-acting, en-dependent silencing element alone may not account for this regulation at ci in the posterior compartment of the wing disc.
An explanation of the unusual behavior of ci alleles and the wing vein phenotype (not the embryonic pat- terns), and the interactions between ci and en, could involve the trans-sensing [Tartof and Henikoff, 19901 of one ci allele by the other. Such a gene silencing system would have two components, a send function and a receive function, which may involve separate upstream regulatory elements at ci. This presupposes the gene is being expressed and needs silencing which appears to be the case with ci [Schwartz et al., 19951. If working together in cis, this system of two elements could silence an adjacent gene and would appear as a single silencing element, in that the sending of a si- lencing signal would be immediately received and acted on. Such a situation could explain the reporter constructs mentioned above. However, such a system would also be able to send a trans-silencing signal from one homolog to the other homolog. The receiving of this signal by a homolog would affect the expression in cis and repress or silence the expression of an adjacent
gene, in this case ci. The transmission of such a signal would only occur if the sequences were in close prox- imity, as would be the case for somatically paired chro- mosomes.
Mutations at the receive site, which transmits the silencing signal in cis to the adjacent ci gene, could explain the dominant alleles ciw and ciCd. The silenc- ing function would be unable or less effective in re- sponding to either cis- or truns-acting signals. This could be due to mutation of the receiving element or the silencer could be prevented from acting on the pro- moter as a consequence of mobile element insertion that are present in these alleles [Orenic et al., 1990; Locke and Tartof, 1994; Slusarski et al., 19951. Each is a partial loss of silencing function, as the ci phenotype is often incompletely dominant.
Loss of the send function could explain the recessive ci wing vein alleles. Such alleles would rely on trans- repression to silence ectopic expression of ci in the pos- terior compartment cells. Thus, ci expression from the recessive alleles can be repressed if paired with a nor- ma1 homologue (ci + /cirecessiue ), but not when homozy- gous as neither homologue has a send function. For example, ci57g, which is a recessive wing vein allele has a small deletion upstream from the promoter [Locke and Tartof, 1994; Schwartz et al., 19951. It is not dom- inant because even though the send function is inactive on that homologue and is unable to silence in cis, the receive component is still able to receive a silencing signal from the paired ci+ homologue. Somatic chro- mosome pairing provides an opportunity for physical interaction and sending a trans-acting silencing signal from the wild type homologue to the cirecessiue homolog. Receiving such a silencing signal prevents ci ectopic expression and the ci wing vein phenotype. However, if the ci .+ homologue is unpaired, as in R(ci+)lcimcesSive,
allele expresses ci ectopically, since the receive component would be unaffected by the inactive cis-send and out of contact with the functional send component on the ci+ homologue. Thus the recessive ci wing vein alleles become dominant when unpaired.
The recessive ci wing vein alleles also become dom- inant in the presence of reduced en+ protein (an en- allele). Reduced en+ produces a similar phenotypic ef- fect to that of an unpaired ci' allele. It may be that the en+ protein assists other proteins, through some type of cooperative action, in the pairing dependent trans- repression of cirecessiue alleles. It may be that the en+ product is involved in chromosome pairing directly or, alternatively, en + protein function may be facilitated by pairing at the ci locus. In either situation, en+ prod- uct should be associated with the ci gene sequences involved in the trans-silencing and such an association has been demonstrated by Schwartz et ul. (1995). An en+ binding site has been mapped to within the se- quences deleted in the ci5'g allele as would be expected for a recessive ci wing vein allele lacking the send com- ponent of the truns-silencing system. In cil and ci361,
the Cirecessiue

348 LOCKE AND HANNA
the presence of inserted mobile element sequences [Locke and Tartof, 19941 could preclude or interfere with the action of the send function.
The penetrance and expressivity of the ci phenotype in en-/+ Cirecesstve I + double heterozygotes correlates with enc allele strength (lethal en alleles enhance more than viable alleles) and gene dosage. Such dosage sensitivity further supports a direct effect of en+ pro- tein on ci transcription. Dose-dependent functioning of en+ has been observed previously, but in embryonic development [Bejsovec and Wieschaus, 19931. Here, changes in en+ gene dosage were observed to affect the embryonic cuticle pattern seen in wingless; naked dou- ble mutants. In both the imaginal and embryonic sit- uation the dosage of en+ seems critical only when cer- tain other mutations are also present. Such mutations may identify adjacent steps in pathways of gene regu- lation for normal development.
We find it significant that silencing of ci by en+ can be hampered or prohibited by unpairing ci. The en-/ + ; ci/+ double heterozygote assay differentiates between unpaired ci+ and paired ci’ . Cytologically visible de- ficiencies that remove the ci region, leaving the other homologue’s single copy of ci+ unpaired (e.g., Df (4)w) produce a ci phenotype with en-/ + . The un- pairing of two wild type ci+ alleles, as in chromosome rearrangements such as 2 ’ ( 1 ; 4 ) ~ ~ , sim-4, and possibly Df14)@3u, results in a ci phenotype in an en-/ + back- ground. These genotypes have two copies of ci+ un- paired. This suggests that a wild type ci+ allele, or alleles, in the presence of en-/ + , may not be properly silenced if ci+ is unpaired. Chromosome pairing at ci appears to be necessary for the posterior compartment silencing of ci in en-/ + heterozygotes.
The insensitivity of ciD heterozygotes to dosage ef- fects of en suggests the ciD wing vein phenotype has a different origin from that of the other ci alleles. Its molecular lesion is possibly within the transcription unit and an altered protein has been associated with this allele [Slusarski et al., 19951. The c?’ mutation is also unique in that it does not complement the reces- sive lethal 2(4)13 allele. The transcript from ciD may be derived from a different promoter with no receive ele- ments and thus regulated in a manner producing ec- topic expression and the dominant ci wing vein pheno- type [Slusarski et al., 19951. The ciD allele has retained a send component since the ciDre” alleles complement recessive ci wing vein mutations and do not produce an unpairing effect as en-/+; ciDreu/+ have wild type wings.
The situation at ci may be similar to that of en- graded, where upstream from the en transcription unit, pairing sensitive sites (PS sites) have been iden- tified through their ability to repress the expression of white+ gene in P-element constructs that have been transformed back into the Drosophila genome [Kassis etal., 19911. Three PS sites have been identified within 1.6 kb of the en+ promoter and are thought to influ-
ence transcription [Kassis, 19941. The pairing of these sites could provide a means for trans-regulation that may explain the ability of certain en alleles to comple- ment one another [Condie and Brower, 19891. Pairing between homologs may provide a mechanism that en- sures the coordinate repression of both alleles, possibly through an altered chromatin conformation that re- presses the expression of both genes.
Recent observations on the expression of genes rein- troduced into the genome have shown, that adjacent, tandem repeats on one homologue can repress genes within the repeats and that pairing of these similar DNA elements is part of, or a prerequisite for, the gene silencing in this situation [Dorer and Henikoff, 19941. This model proposes that physical pairing of adjacent tandem repeats is followed by a condensation of its chromatin and gene silencing. A critical mass of se- quences and proteins may be needed for chromatin con- densation and gene repression to occur. At ci reaching this critical mass may be facilitated by chromosome pairing and the physical juxtapositioning of ci pairing dependent sites on the two homologs. In the light of such a model for pairing dependent gene silencing pro- posed for tandem repeats, the dose dependent manner of en+ action is reminiscent of that proposed for genes subjected to position effect variegation and affected by dosage of second-site modifier loci [Locke et at., 19881. The interactions between en and ci described here offer a simple system for examining normal gene repression that involves trans-acting factors and chromosome pairing .
ACKNOWLEDGMENTS We thank P. Sekhon for technical assistance, the In-
diana University, Umea, and Bowling Green Drosoph- ila Stock Centers for providing stocks, as well as M. Russell for providing the diversity of developmental mutations tested. Financial support was provided by the Natural Sciences and Engineering Research Coun- cil of Canada and the University of Alberta Central Research Fund.
REFERENCES Ashburner M (1989): “Drosophila: A Laboratory Handbook.” Cold
Spring Harbor, NY: Cold Spring Harbor Laboratory Press. Bejsovec A, Wieschaus E (1993): Segment polarity gene interactions
modulate epidermal patterning in Drosophila embryos. Develop- ment 119501-517.
Brower DL (1986): engmiled gene expression in Drosophila imaginal discs. EMBO J 5:2649-2656.
Condie JM, Brower DL (1989): Allelic interactions a t the engmiled locus of Drusophila: engrailed protein expression in imaginal discs. Dev Biol 13531-42.
Dorer DR, Henikoff S (1994): Expansions of transgenic repeats cause heterochromatin formation and gene silencing in Dmsophilu. Cell 77:993-1002.
Dubinin NP, Sidorov BN (1934): Relation between the effect of a gene and its position in the system. Am Nat 68:377-381.
Eaton S, Kornberg TB (1990): Repression of c i D in posterior com- partments of Drosophila by engrailed. Genes Dev 4:1068-1077.

en ALLELES ENHANCE THE WING PHENOTYPE OF ci ALLELES 349
Eberlein S, Russell MA (1983): Effects of deficiencies in the engmiled region of Drosophila melanogaster. Dev Biol 100227-237.
Eker R (1929): The recessive mutant engruiled in Drosophila mela- nogaster. Hereditas 12:217-222.
Ephrussi B, Sutton E (1944): A reconsideration of the mechanism of position effect. Proc Natl Acad Sci USA 30:183-197.
Guarente L (1993): Synthetic enhancement in gene interaction: A genetic tool come of age. TIG 9:362-366.
Hochman B (1976): The fourth chromosome of Drosophila melanogas- ter. “The Genetics and Biology of Drosophila.” New York Academic Press, pp 903-928.
Horton IH (1939): A comparison of the salivary gland chromosomes of Drosophila melanogaster and D. simulans. Genetics 24234-243,
House VL (1953a): The interaction of mutants affecting venation of Drosophila melanogaster. I. Interaction of Hairless, engmiled, and cubitus interruptus. Genetics 38:199-215.
House VL (1953b): The interaction of mutant affecting venation of Drosophila melanogaster. 11. Additive and pattern effects of combi- nations of Hairless, engrailed, and cubitus interruptus. Genetics 38: 309-327.
Kassis JA (1994): Unusual properties of regulatory DNA from the Drosophilu engmiled gene: Three “pairing-sensitive’’ sites within a 1.6-kb region. Genetics 1361025-1038.
Kassis JA, Vansickle EP, Sensabaugh SM (1991): A fragment of en- gmiled regulatory DNA can mediate transvection of the white gene in Drosophila. Genetics 128:751-761.
Kinzler KW, Bigner SH, Bigner DD, Trent KM, Law ML (1987): Iden- tification of an amplified, highly expressed gene in a human glioma. Science 236:70-73.
Kuner JM, Nakanishi M, Ali Z, Drees B, Gustavson E, Theis J, Kau- var L, Kornberg T, OFarrell PH (1985): Molecular cloning of en- graded A gene involved in the development of pattern in Drosoph- ila melanogaster. Cell 42:309-316.
Lewis EB (1950): The phenomenon of position effect. Adv Genet 3:73- 115.
Lindsley DL, Grell EH (1968): “Genetic Variations of Drosophilu ml- anogaster.” Oak Ridge, TN: Carnegie Institution of Washington.
Lindsley DL, Zirnm GG (1992): “The Genome of Drosophila melano- gaster.” San Diego: Academic Press.
Locke J, Tartof KD (1994): Molecular analysis of cubitus interruptus (ci) mutations suggest an explanation for the unusual ci-position effects. Mol Gen Genet 243:234-243.
Locke J, Kotarski MA, Tartof KD (1988): Dosage-dependent modifiers of position effect varigation in Drosophila and a mass action model that explains their effect. Genetics 120:181-198.
Muller HJ, Pontecorvo G (1940a): Recombinants between Drosophila species the F1 hybrids of which are sterile. Nature 146:199-200.
Muller HJ, Pontecorvo G (1940b): The artificial mixing of incompat- ible germ plasms in Drosophila. Science 92:418(abst); Science 92: 476 correction.
Qrenic TV, Slusarski DC, Kroll KL, Holmgren RA (1990): Cloning and characterization of the segment polarity gene cubitus interrup- tus Dominant of Drosophila. Genes Dev 41053-1067.
Orenic T, Chidsey J , Holmgren R (1987): Cell and cubitus interruptus Dominant: T w o segment polarity genes on the fourth chromosome in Drosophila. Dev Biol 124:50-56.
Ruppert JM, Vogelstein B, Arheden K, Kinzler KW (1990): GLI3 en- codes a 190-kilodalton protein with multiple regions of GLI simi- larity. Mol Cell Biol 10:5404-5415.
Schwartz C, Locke J, Nishida C, Kornberg TB (1995): Analysis of cubitus interruptus regulation in Drosophilu embryos and imaginal disks. Development 121:1625-1635.
Slizynski BM (1941): The structural differentiation of the chromo- some IV of Drosophila simulans and its behavior in melanogaster genotype. Proc R Soc (Edin) 61:95-106.
Slusarski DC, Motzny CK, Holmgren R (1995): Mutations that alter the timing and pattern of cubitus interruptus gene expression in Drosophila melanoguster. Genetics 139229-240.
Spofford J B (1976): Position effect variegation in Drosophila. “Genet- ics and Biology of Drosophila.” Vol. 1C. London: Academic Press.
Stern C, Heidenthal G (1944): Materials for the study of the position effects of normal and mutant genes. Genetics 30:197-205.
Stern C, Kodani M (1955): Studies on the position effect at the cubitus interruptus locus of Drosophila melanogaster. Genetics 40:343-373.
Stern C, MacKnight RH, Kodani M (1946): The phenotypes of ho- mozygotes and hemizygotes of position alleles and heterozygotes between alleles in normal and translocated positions. Genetics 31:
Tartof KD, Henikoff S (1991): Trans-sensing effects from Drosophila to humans. Cell 65201-203.
Zarkower D, Hodgkin J (1992): Molecular analysis of the C. elegans sex-determining gene fra-1: A gene encoding two zinc finger pro- teins. Cell 70:237-249.
598-619.


















