
ELECTROPHYSIOLOGICAL RESPONSES OF THE TERMINAL...
22
J. Exp. Biol. (i974), 60, 275-293 275 With 2 plates and 11 text-figures Printed in Great Britain ELECTROPHYSIOLOGICAL RESPONSES OF THE TERMINAL SENSILLA ON THE MAXILLARY PALPS OF LOCUSTA MIGRATORIA (L.) TO SOME ELECTROLYTES AND NON-ELECTROLYTES BY W. M. BLANEY Zoology Department, Birkbeck College, Malet Street, London, WCiE 7HX (Received 27 July 1973) INTRODUCTION It has been shown that the maxillary palps of Locusta migratoria (L.) play an import- ant part in food selection when the insect has not been deprived of food for a long period (Blaney & Chapman, 1970; Blaney, Chapman & Wilson, 1973). The receptors primarily concerned are those on the flexible dome at the distal tip of the palp, and in the fifth instar larvae of Locusta there are about 370 of these sensilla on the dome of each maxillary palp. The fine structure of these sensilla (Blaney, Chapman & Cook, 1971) corresponds to that generally ascribed to contact chemoreceptors (Lewis, 1970; Slifer, 1970) and a chemoreceptive function has been confirmed by Haskell & Schoonhoven (1969) using electrophysiological techniques. This paper describes part of an electrophysiological study to determine the role of these receptors in food selection, and in this preliminary investigation an examination is made of the responses of the receptors to some electrolytes and non-electrolytes. MATERIALS AND METHODS Insects All the experiments were carried out with male fifth-instar larvae of Locusta migratoria (L.) obtained from the normal laboratory stock at the Centre for Overseas Pest Research (Hunter-Jones, 1961) and kept subsequently in 12 1 cylindrical Perspex cages at 26-30 °C with about 12 insects in each cage. The insects were marked individually within 12 h of ecdysis and used in the period 2-5 days after ecdysis so that the results were not influenced by the proximity of the moult. Recording technique Electrophysiological recordings were obtained using a modified version of the technique of Hodgson, Lettvin & Roeder (1955); silver/silver chloride electrodes were used to detect, extracellularly, electrical changes within the sensilla. The indifferent electrode was connected via a back-off d.c. source and calibration source to earth. The back-off source was used to compensate for the potential difference between the
Transcript of ELECTROPHYSIOLOGICAL RESPONSES OF THE TERMINAL...

J. Exp. Biol. (i974), 60, 275-293 275With 2 plates and 11 text-figures
Printed in Great Britain
ELECTROPHYSIOLOGICAL RESPONSESOF THE TERMINAL SENSILLA ON THE MAXILLARY
PALPS OF LOCUSTA MIGRATORIA (L.) TO SOMEELECTROLYTES AND NON-ELECTROLYTES
BY W. M. BLANEYZoology Department, Birkbeck College, Malet Street,
London, WCiE 7HX
(Received 27 July 1973)
INTRODUCTION
It has been shown that the maxillary palps of Locusta migratoria (L.) play an import-ant part in food selection when the insect has not been deprived of food for a longperiod (Blaney & Chapman, 1970; Blaney, Chapman & Wilson, 1973). The receptorsprimarily concerned are those on the flexible dome at the distal tip of the palp, andin the fifth instar larvae of Locusta there are about 370 of these sensilla on the dome ofeach maxillary palp.
The fine structure of these sensilla (Blaney, Chapman & Cook, 1971) correspondsto that generally ascribed to contact chemoreceptors (Lewis, 1970; Slifer, 1970) anda chemoreceptive function has been confirmed by Haskell & Schoonhoven (1969)using electrophysiological techniques.
This paper describes part of an electrophysiological study to determine the role ofthese receptors in food selection, and in this preliminary investigation an examinationis made of the responses of the receptors to some electrolytes and non-electrolytes.
MATERIALS AND METHODS
Insects
All the experiments were carried out with male fifth-instar larvae of Locustamigratoria (L.) obtained from the normal laboratory stock at the Centre for OverseasPest Research (Hunter-Jones, 1961) and kept subsequently in 12 1 cylindrical Perspexcages at 26-30 °C with about 12 insects in each cage. The insects were markedindividually within 12 h of ecdysis and used in the period 2-5 days after ecdysis sothat the results were not influenced by the proximity of the moult.
Recording technique
Electrophysiological recordings were obtained using a modified version of thetechnique of Hodgson, Lettvin & Roeder (1955); silver/silver chloride electrodes wereused to detect, extracellularly, electrical changes within the sensilla. The indifferentelectrode was connected via a back-off d.c. source and calibration source to earth. Theback-off source was used to compensate for the potential difference between the

276 W. M. BLANEY
Fig. 1. Two examples of the 'unstimulated' response of terminal sensilla recorded by the side-wall technique. Notice the different levels of 'spontaneous' activity. Horizontal bar shows
sensillum tip and the tip of the capillary containing the stimulating solutions and therecording electrode. Compensation, which was approximated on adjacent hairs notused for recording, prevented blocking of the amplifier and allowed recording of theearliest impulses elicited by the stimulating solution (see van der Starre (1972) fordiscussion). The calibration source provided a square wave of 50 mV peak-to-peakwith a frequency of 1 kHz.
Electrical signals from the sensilla were fed from the recording electrode through alaboratory built a.c.-coupled pre-amplifier with a cathode-follower input stage givingan input grid current of < io~12 A (Rees, 1968). Amplified signals were displayed ona C.R.O. (Tektronix 502 A) and recorded on a tape recorder (Thermionic T3000).For subsequent analysis the electrical phenomena were printed out using a u.v.recorder (S.E. 3006/DL) or photographed on oscillograph paper.
This method of tip recording prevented the testing of compounds which were notelectrolytes except by the addition of an electrolytic substance to the stimulatingsolution. In practice, all substances other than salts were presented in a solution of0-05 M sodium chloride. This constraint could have been circumvented by using thesidewall recording technique of Morita (1959); but, besides the difficulty of applyingthis technique to very small sensilla, when it was successfully applied the level of'spontaneous' firing indicated was very variable (Fig. 1) and was often much higherthan that reported in other insects (Rees, 1968, in Phormia; 1969, in Chrysolina;McCutchan, 1969, in Phormia). It seems likely that this represents an injury effect,perhaps arising from interference with the conduction of receptor potentials along thedendrites (Rees, 1967). For these reasons it was decided not to use this technique.
Solutions and pipettes
The stimulating solutions were made using Analar or biochemical grade compounds(B. D. H., Hopkins & Williams) in glass-distilled de-ionized water. They were storedwhen not in use at — 25 °C. The test solution, with the recording electrode, wasapplied to the sensillum in a glass capillary drawn and cut to give a tip diameter ofapproximately 10/tm. There was some tendency for water to evaporate from the tip,so increasing the concentration of the test solution, but this was offset by smoothingthe cut end of the capillary by heating and by maintaining the experimental area at

Electrophysiological responses of terminal sensilla of maxillary palps 277
Cathodefollower
7?c=<10-'2APre-amplifier
Recordingelectrode
Preparation
28 "C70-90% R.H.
Indifferentelectrode
C.R.O. Camera
Audio/ visualmonitors
Tape recorder
Back-ofTd.c. source Calibration source
U.v. recorder
Fig. 2. Diagram of the recording arrangements.
high humidity (R.H. 70-90 %). In addition, to ensure that the concentration appliedwas typical of the solution, fluid was drawn from the tip of the capillary by means oflens tissue immediately before each stimulation.
Method of preparation
Before being used in an experiment, normally feeding locusts were taken individu-ally in 350 ml glass jars and observed until they had fed to repletion on fresh grass(Blaney & Chapman, 1970V They were then kept without food for a precisely knownperiod, commonly 2 h, before preparation for electrophysiology. This ensured thata good proportion of the sensilla had regained the readily functional state after feeding(Bernays, Blaney & Chapman, 1972) and avoided any abnormalities arising fromunusually prolonged periods without food.
Consistent results were only obtained by using intact but restrained insects;isolated palps or heads frequently gave no results due to the effects of injury (see alsoHaskell & Schoonhoven, 1969).
Insects were anaesthetized briefly (< 2 min) with carbon dioxide, the legs wererestrained and the head and thorax were immobilized as shown in Plate 1, using CotrellSticky Wax, care being taken to avoid damage by overheating. The trussed insect wasattached with wax to a Perspex cradle so that the left maxillary palp was exposed forrecording. The indifferent electrode was inserted into the haemolymph adjacent tothe right antennal socket. The Perspex cradle, bearing the connecting terminal forthe indifferent electrode, was clipped onto the movable stage of a Leitz microscope,and manipulation of the preparation and recording electrode was accomplished by

278 W. M. BLANEY
4 IFig. 3. Recordings of attempted movements by the insect (between arrows) superimposed on
the response by terminal sensilla to o-i M sodium chloride. Horizontal bar shows o-i sec.
combined use of the microscope stage controls and micromanipulators (Narishige).A schematic diagram of the recording situation is given in Fig. 2.
Attempted movements by the insect during recording were always accompanied byvolleys of impulses due to motor neurones firing. This produced a recognizable soundon the audio monitor and visually appeared as a burst of low-amplitude impulses(Fig. 3). When this occurred the record was not used for analysis and any insecthaving a tendency to be active was discarded. Further, because of the proximity ofadjacent sensilla on the palp tip, the possibility was investigated of inadvertentlyrecording spontaneous activity from sensilla other than being tested. With therecording electrode in place over an adapted and fatigued sensillum, mechanical andchemical stimuli were applied to adjacent sensilla. At no time was activity from othersensilla recorded in this way (see also van der Starre, 1972). I am therefore confidentthat the results obtained from the terminal sensilla are not influenced by nervousactivity from elsewhere in the body.
RESULTS
General considerations
Three types of sensilla are distinguishable on the basis of their fine structure(Blaney et al. 1971; Blaney, in preparation) on the tip of the maxillary palp of Locustamigratoria, and the distinction is confirmed by electrophysiological recordings. About90 % of the sensilla are approximately 25 /an high with a structure as described byBlaney et al. (1971). Of the remainder about half, i.e. 5 % of the total, are less than20 /im high and lack a distal crest (PI. 2). Le Berre, Sinoir & Boulay (1967) suggestthat these are mechanoreceptors, and this is in accord with the lack of response fromthese sensilla on stimulation by the tip-recording method. A further 4-5 % also lackthe distal crest but are about 30 fim high and are readily distinguished on the basis oftheir electrophysiological responses. A preliminary study suggests that these sensillamay mediate the common chemical sense (Blaney, in preparation).
This paper is concerned only with the responses of the first group of sensilla.
Responses to c i M sodium chloride
Representative responses by the terminal sensilla to stimulation with o-i M sodiumchloride are shown in Fig. 4. The phasic-tonic response typical of insect chemo-receptors is seen here. Although there is much variation in the absolute and relative

Electrophysiological responses of terminal sensilla of maxillary palps 279
' ' ' * T
(A)
Fig. 4. Responses of terminal sensilla to stimulation with o-i M sodium chloride. In (a) and(6), impulses of one amplitude category are present; (c) and (d) show impulses of two amplitudecategories, while in (e) and (/) there are impulses of three amplitude categories. Horizontal barshows o-i sec.
duration and vigour of the phasic and tonic elements, it is always possible to recognizeboth (Fig. 7).
The traces obtained are, however, very variable with respect to the occurrence ofimpulses of different amplitudes. Very rarely (in 4 cases out of 192) only one amplitudecategory appears to exist (Figs. \a, b; 5 a, b). Commonly two (Figs. 4c, d; 5 c, d) or threeFigs. 4 c,/, $e,f) discrete amplitude categories are recognizable, but sometimes the

280 W. M. BLANEY
CO
20 25 30
15 20 25
Amplitude classes (mm)
30
Fig. 5. Frequency histograms of impulse amplitudes during the first second of theresponses shown in Fig. 4.
variation in amplitudes is such that it is not possible to assign them to discretecategories.
Fig. 6 shows the adaptation curves for the records shown in Fig. 4, and Fig. 7expresses the rate of adaptation as derived from Fig. 6. It is apparent that initial, rapidadaptation followed by a tonic level of discharge occurs in all recognizable amplitudeclasses.
There is great variability in the total number of impulses occurring during the firstsecond of stimulation if the same sensillum is repeatedly stimulated with a givensolution at intervals long enough (10 min) to allow complete recovery from theprevious stimulation (Table 1). Similarly Fig. 8 shows the variation occurring in thesecircumstances between successive responses in respect of absolute and relative ampli-tude classes, and in the relative rate of firing between amplitude classes (see also denOtter & van der Starre, 1967, and van der Starre, 1972). The number of amplitudeclasses is, however, consistent in any one sensillum for a given stimulus solution.
Table 1 also shows the variation between hairs on initial and subsequent stimulations.

Electrophysiological responses of terminal sensilla of maxillary palps 281
30
20
10
30
20
10
-
-
1
v (*)
1 1 1 1
60
50
40
30
20
10
Time (sec)
Fig. 6. Adaptation curves for the first five seconds of responses shown in Fig. 4.A-A, large impulses; • — • , medium impulses; y—Ti small impulses.
Table i. Variability of response in the first second of successive stimulations
of five sensilla by o-i M sodium chloride solution
Number of stimulation
Sensillum
1
2
3
4
5
MSMSMSMSMS
1
1817
5417
49294216
2626
2
3511
4129
47245413
2518
3
27J33738
4730
3718
3930
4
2812
263i
3925
7338
2421
5
173i5622
39435142463°
6
29382721
593666373619
M = number of medium-amplitude impulses. S = number of small-amplitude impulses.

282 W. M. BLANEY
% 100
90
80
70
60
50
Time (sec)
Fig. 7. Curves derived from Fig. 6 showing rate of adaptation of each amplitude category.Each point, except the first, is expressed as a percentage of the preceding one.
Table 2. The number of impulses in the first second of stimulation offour sensilla with different concentrations of sodium chloride
Sensillum
olarity
o-oi0-050-075o-i0-25o-5
1
371 0 0
989692
1 1 7
2
3961
46738196
3
64525512
2 1
34
4
1 2
24336427
6
Responses to a range of concentrations of sodium chloride
Individual sensilla were tested with solutions of sodium chloride at six differentconcentrations (o-oi M, O 05 M, 0-075 M> o ' x M> o-25 M a nd 0-5 M). The order of appli-cation of the stimuli was randomized and at least 10 min were allowed between suc-cessive applications of stimulus to any one sensillum.
As before, individual sensilla responded with impulses having one or more ampli-tude categories. In about half of the sensilla tested there was a tendency for the firingrate to increase as the concentration of salt increased (Table 2, sensilla 1 and 2). Ofthe remainder, some responded to an increase in salt concentration with a decline infiring rate (Table 2, sensillum 3), but in most the highest firing rate occurred atintermediate concentrations of salt with a decline at either end of the range (Table 2,sensillum 4). Considerable variability was noted, but in general, individual sensillaappeared to have a similar distribution of impulse amplitude categories at allconcentrations.

Electrophysiological responses of terminal sensilla of maxillary palps 283
15 20 25 5 10
Amplitude classes (mm)
15 20
Fig. 8. (a) Frequency histogram of impulse amplitudes during the first second of ten successivestimulations with o-i M sodium chloride of a sensillum having impulses of two amplitudecategories. (6) The same from a sensillum having impulses of three amplitude categories.
Responses to a range of different compounds
The responses of the terminal sensilla to a number of compounds other than sodiumchloride were obtained. The compounds tested were not necessarily at the concentra-tions at which they occur in plants because that information is not generally available.However, in most cases the concentration chosen was that which had been shown togive an unequivocal response in behavioural experiments (A. G. Cook, personal
IQ E X B 60

284 W. M. BLANEY
Table 3. Responses often sensilla to various organic and inorganic compounds
Sensillum
Stimulus solution
o-i M sodium chlorideo-i M potassium
chlorideo-i M ammoniumchloride
0-025 M glucose0-025 M fructose0025 M xylose0-025 M maltose0-025 M sucrose0-0125 M serine0-0125 M alanineo-1 M citric acid0-1 M oxalic acid1-5% hydrochloricacid
Quinine hydrochlorideNeemAnthroquinone
1
+ +0
+
0
+0
+——0
0
0
0
0
2
+ + •+ +
++ +++0
0
0—
0
+
0
+—
3
+ ++ +
+
0
+ +0
+++ ++
+ ++ +
++0
4
+ + •+ +
+
++ ++ ++ ++0
0
0
+ ++ +
+0—
S
+• 0
0
H0
0
0
0
H+ 0
0
0
+ + o+ + + o+ + o— 0 0
o — —0 0 —
0 0 0
o — +0 — 0
00 +
+ O O O O
o — — o o— O —
Responses during the first second of stimulation are considered. Those which do not differ from thecontrol level are indicated as o, those showing a marked increase in firing rate are indicated as + + + ,those showing a marked decrease as , and the other categories are intermediate.
communication). Otherwise concentrations were chosen so as to avoid being eithersubthreshold or damagingly high.
To ensure adequate electrical conductivity of the stimulating solution, all thecompounds tested were presented in 0-05 M sodium chloride solution and the re-sponses were compared with the average of a number of responses to stimulation with0-05 M sodium chloride alone.
The results, shown in Table 3, are coded according to the number of impulsesoccurring in the first second; and, because of the variability inherent in the system,only responses which differed by 20 % from the control level were regarded as indi-cating an effect of the additional stimulant (see also Dethier & Kuch, 1971).
No single sensillum would seem capable of differentiating between compounds noreven classes of compounds, yet it is clear that many sensilla are capable of respondingin some way to a larger number of compounds than there are neurones in the sensilla.The full extent of this effect may be masked by the conservative interpretation ofresponse level; some compounds recorded as ineffective may in fact be weakly stimu-lating or inhibiting.
Mechanoreception
In addition to the effects of chemical stimulation, those of mechanical manipulationwere investigated. Sensilla which were already being stimulated by a o-i M solutionof sodium chloride were displaced laterally by the stimulating capillary.
The response, when it occurred, was always the same and consisted of a burst ofimpulses whose amplitude was considerably greater than that of impulses elicited bychemical stimulation (Bernays, Blaney & Chapman (1972), fig. 4). The response was

Electrophysiological responses of terminal sensilla of maxillary palps 285
strongly phasic as reported by Haskell & Schoonhoven (1969); but, whereas theseauthors found it to occur in all the sensilla which they tested, in the present investiga-tion only 47% of 390 sensilla (on 13 insects) responded.
INTERPRETATION OF RESULTS
The uncertainty in identifying the responses of individual neurones by inspectionof the amplitude and temporal pattern of impulses on traces is compounded with thevariability of response between different sensilla, between subsequent stimulations ofthe same sensillum with the same solution and between stimulations of the samesensillum with different concentrations of the same stimulant.
It is generally assumed that impulses of characteristically different and constantamplitudes derive from different neurones (Dethier & Hanson, 1965; Dethier & Han-son, 1968; Hodgson, 1968), but it does not necessarily follow that impulses of the samesize derive from the same neurone. Unusually large impulses occurring infrequentlyappear to result from a temporal coincidence and consequent summation of two ormore smaller impulses. This diagnosis may be confirmed by inspection of traces whenpairs of different-sized impulses may be seen to occur successively closer together,finally to be replaced by one or more larger impulses before reappearing as separate,smaller impulses again. Significantly, this phenomenon may occur with pairs of similar-sized impulses which, but for the coincidence, would otherwise be regarded as de-riving from one neurone.
Clearly this event occurs most frequently when the overall rate of firing is high.In many cases in the present study the rate of firing is low compared with that reportedfor other insects and may well be too low for the coincidence effect to be apparent. Itis therefore impossible to ensure that the traces shown in Fig. 4, and others like them,in fact represent the activity of the number of neurones that they seem to indicate.
Further, the terminal sensilla each contain at least six neurones (Blaney et al. 1971);the total range of amplitude from the largest to the smallest impulses recorded com-monly did not exceed 20 mm, and the range of variation in impulses of one amplitudecategory was seldom less than 6 mm, hence the chance of 6 neurones firing withoutoverlap is remote.
Firm conclusions regarding the number of cells responding at any time are there-fore unwarranted; and it appears that, while it is fairly certain that in some cases(during stimulation with o-1 M sodium chloride) at least three neurones are firing, innone of the records is there unequivocal evidence for less than three neurones firing.
It is arguable that some of these neurones were firing spontaneously at a tonic levelunrelated to the application of the stimulus under investigation, but Figs. 6 and 7show that all the neurones, identified on the basis of impulse-amplitude categories,initially show rapid rates of adaptation consistent with a response to the stimulus(Rees, 1968).
The same holds true for other stimulating substances; in every case the responseappears to involve a number of neurones.
The variation of impulse-amplitude categories associated with the application ofdifferent concentrations of sodium chloride solution to a given sensillum (Fig. 9 a)might indicate the activity of different neurones at different concentrations. However
19-2
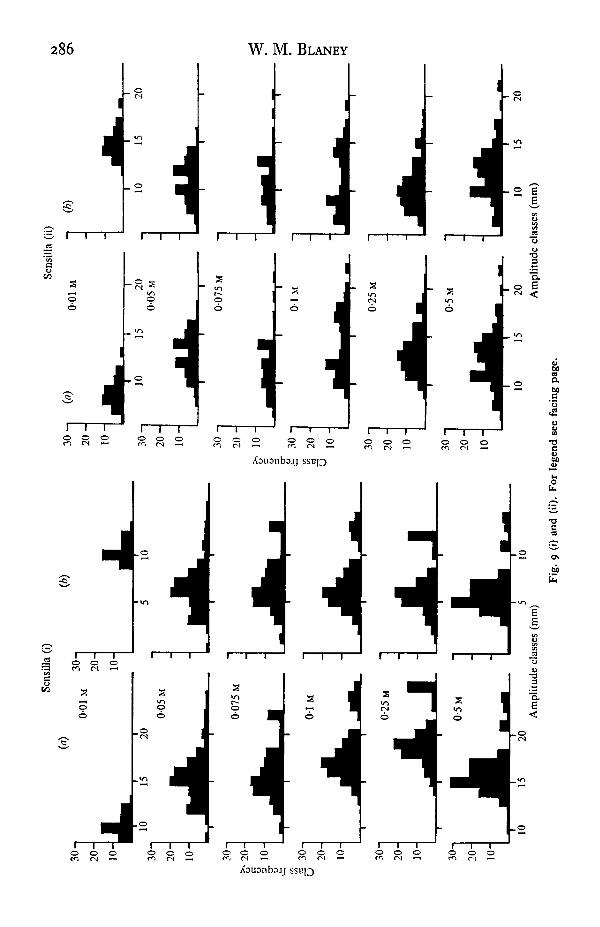
Sens
illa
(i)
Sens
illa
(ii)
h)
00
30L:, 0.
01;
I>-,
20
10
10
15
20
10
30
i &
0.05 M &
:- iL
0.05
; L':
2r 20
10
30 7
0.07
5 M
1
- -
Am
plit
ude
clas
ses
(mm
) A
mpl
itud
e cl
asse
s (m
m)
Fig
. 9
(i) a
nd (
ii).
For
leg
end
see
faci
ng p
age.

Am
plitu
de c
lass
es (mm)
Am
plitu
de c
lass
es (
mm
)
Fig
. g.
Fre
quen
cy h
isto
gram
s of
impu
lse
ampl
itud
es d
urin
g th
e fir
st s
econ
d of
sti
mul
atio
n of
fou
r se
nsill
a, (i
)-(i
v), w
ith
sodi
um c
hlor
ide
solu
tion
at t
he c
once
ntra
tion
s in
dica
ted.
(a) I
mpu
lse
ampl
itud
es a
s re
cord
ed; (
b) a
mpl
itud
e co
rrec
ted
by t
he c
alib
rati
on fa
ctor
. For
fur
ther
det
ails
see
tex
t.

288 W. M. BLANEY
120 r
i no
105
aJ
5g
001 002 005M-NaCl
0075i ?
0-1
Fig. io. Comparison of equivalent conductance (•—9) of various concentrations of sodiumchloride (Weast, 1964) with the calibration-pulse amplitude (y—v) recorded by placing theindifferent and recording electrodes to a fixed depth, and at a fixed distance apart, into eachsolution.
when the calibration pulse was passed through the preparation during recording andits height used as a correction factor to standardize the impulse amplitudes, the varia-tion was largely eliminated (Fig. gb), and must have arisen through some character-istic of the preparation/stimulus system which varied with concentration of electrolyte.There is, however, still considerable misalignment when the recorded calibrationpulse was of low amplitude, i.e. especially at o-oi M concentration.
That this was not an effect of low electrolyte concentration on the recording of thecalibration pulse was shown by recording the amplitude of the calibration pulsewithout the preparation in circuit (Fig. 10). The amplitude increases at low con-centrations in parallel with the equivalent conductance curve (Weast, 1964). Thus themisalignment is associated with the preparation.
The correlation between impulse frequency and calibration-pulse amplitude ishigh (Z = 0-89, Fig. 11). It may be that the recorded changes in amplitude ofcalibration pulse and of nerve impulses are related to changes in the dendritic mem-brane resistance which is in series with the recording electrode (Bernard & Guillet,1972; Rees, 1967). Thus a high rate of firing would reflect a high degree of depolariza-tion of the receptor membrane and increased electrical conductivity of the dendriticmembrane. That would reduce the series resistance in the recording pathway andincrease the amplitude of the recorded events.
Alternatively Morita & Takeda (1957) working on tarsal receptors of Vanessa andStiirckow (1971) on chemoreceptors of Calliphora suggest that changes in resistancemay be related to different amounts of viscous material present at the tip of thesensillum. This material is also present in Locusta but the apparent changes in resist-ance observed here occur too rapidly to be accounted for by changes in the amountof viscous material. The electrical properties of this material may vary when subjectedto different osmotic potentials but no evidence of this is presently available.
Varying osmotic potential might also have an effect on resistance pathways by

Electrophysiological responses of terminal sensilla of maxillary palps 289
30 r
25
20
?• 15
10
0-25 M
0-5 M
001 M,
10 15 20 25
Impulses/sec
30 35
Fig. 11. The average number of impulses occurring in the first second of stimulation withsodium chloride at six different concentrations plotted against the corresponding averagecalibration-pulse amplitude obtained by recording through the sensilla. (Data from seveninsects.) The calculated regression for impulse frequency on calibration pulse amplitude isdrawn (Z = O'8o.).
opening or closing of the distal pore of the sensilla. Scanning electron-microscopestudies (Blaney & Chapman (1969), on Schistocerca gregaria and (unpublished) onLocusta) have shown that the distal pores on these sensilla are capable of being openedand closed, and Stiirckow, Holbert & Adams (1967) describe a similar phenomenonin chemoreceptor hairs of Phormta regina. However, no evidence was found here ofany change in degree of conductivity during stimulation when solutions of differentosmotic potential were applied successively (see also Bernays et al. 1972). Thus itseems highly unlikely that the degree of opening could be affected by the osmoticpotential of the stimulus solution since this would take some measurable time to haveits effect.
It is apparent that certain difficulties of interpretation remain unresolved. It wouldbe possible, by rejection of anomalies and conservative selection of reproducible data(Gillary, 1969), to present a picture of chemoreception by the terminal sensilla alongconventional lines (see, for example, Haskell & Schoonhoven, 1969); but in so doingmuch usable and valuable information would be lost, not least the magnitude andextent of occurrence of variability.
It seems best to use analysis at the neuronal level where it is warranted, but other-wise analysis at the level of total output of individual sensilla is probably valid (seealso Dethier & Kuch, 1971), particularly where the equivalent of 'across-fibre'analysis is being considered.
DISCUSSION
This study confirms the original findings of Haskell & Schoonhoven (1969) that theterminal sensilla on the maxillary palps of Locusta migratoria act as contact chemo-receptors, responding to a wide range of chemicals of different chemical classes. It isalso clear that, in some cases at least, individual neurones are capable of responding tomore than one type of compound. This characteristic, together with the variability

290 W. M. BLANEY
of results following stimulation with a constant stimulus, suggests that these sensillado not respond in the manner commonly accepted for chemoreceptors.
It is generally considered (Rees, 1968; Haskell & Schoonhoven, 1969) that theresponse of sensilla to a salt solution involves different neurones for each of themodalities (the 'salt' and 'water' neurons of most authors and the 'Type 1' and'Type 3' neurones of Rees). In the present study it is clear that salt and water invokeactivity from more than two neurones in many cases, and there is no evidence tosuggest that this is not so in all cases.
Similarly, den Otter & van der Starre (1967) and van der Starre (1972), working onCalliphora, and van der Starre (1970), on Phormia, have shown that water alone canelicit activity in more than one neurone in sensilla where it was previously thoughtthat only one water-sensitive neurone existed. Further, Dethier (1973), studying theresponses of the maxillary taste receptors of seven species of caterpillars, commonlyfound that in addition to a 'major salt receptor' neurone, up to three other neuronesalso responded to salt and that some ' salt receptors' responded to other chemicals aswell, e.g. amino acids. Other reports of compounds stimulating more than one cellhave been given by Schoonhoven (1969) and by Dethier & Kuch (1971).
These data do not suggest general applicability of the concept of specializedneurones for each stimulus modality. It seems more likely that many neurones respondto a range of stimuli, though perhaps with differential sensitivity. Instances of neuronesresponding to only a single modality would then be regarded as extremes of thisdifferentiation.
Such a situation would yield less precise information than that commonly describedfor the blowflies and other insects but this limitation may be overcome in the case ofthe terminal sensilla on the maxillary palps of Locusta by virtue of the large numberof sensillaavailable to test the environment at any one time. This, and the effect ofpalpation (see Blaney, in preparation), may allow the acquisition of adequatelydetailed information, particularly if some form of ' across-fibre' analysis is used bythe insect. Further, the lack of differentiation may have a positive advantage in that,by having broad band sensitivity, the palpal chemoreceptors are able to perceive awide range of potential food materials.
Such specificity as exists in this system appears to be more at the level of sensillathan at the level of neurones; some sensilla have a notably vigorous over-all responseto salt, some to water, some to sugar (see also Blaney, in preparation, and Stiirckow,1970). This would appear to reflect the presence in these sensilla of one or moreneurones having their differential sensitivity particularly biased towards a specificmodality. The significance of this in terms of neural integration is not yet clear.
The mechanism by which the insect determines different modalities and differentdegrees of the same modality and how it distinguishes between these two conditionsis not clear from this study, but the indications are that the code for taste qualityconsists of the relative amounts of activity produced in many neurones according tothe model proposed by Pfaffmann (1941, 1955) and developed by Erickson (1963,1967) and Doetsch et al. (1969) in respect of mammalian gustatory systems. Aninvestigation of these problems is the subject of further studies (in preparation).
It is surprising that there was not a more distinct response to neem which is knownto be a powerful feeding deterrent for locusts (Sinha & Gulati, 1965). Haskell &

Electrophysiological responses of terminal sensilla of maxillary palps 291
Schoonhoven (1969) have reported that the terminal sensilla contain a neurone specifi-cally sensitive to neem. However, the response to neem is less marked when it isapplied to the palps than to the sensilla of the cibarial cavity (Blaney, unpublished).If, as seems likely, sensitivity to a particular compound is not universally dis-tributed among sensilla, the small number of sensilla tested with neem in the presentstudy may account for the failure to obtain an unequivocal response.
Gilbert, Baker & Norris (1967, 1968) and Norris (1969) have established the role ofcertain quinones as inhibitors of gustation in a number of insects. In the present studythe effect of anthroquinone was to reduce the activity of neurones previously responsiveto sodium chloride and water. This cannot, however, be regarded as the typicalresponse elicited by a deterrent substance as other such substances had a markedlydifferent effect. Similarly Ishikawa (1966) working on Bombyx mori, and Dethier (1973)with seven species of lepidopterous larvae, have shown that inhibition is not a unitarymodality.
Haskell & Schoonhoven (1969) have reported that all of the terminal sensilla theytested could act as mechanoreceptors; in the present study this property has only beenfound in 47 % of the sensilla tested. Blaney et al. (1971) and Blaney & Chapman (1969)studying the fine structure of these sensilla and similar sensilla on Schistocercagregaria found a differentiated region of dendrite near the base of some sensilla and,following other authors (Thurm, 1965; Nichlaus, Lunquist & Wersall, 1967) associatethis with a mechanoreceptor neurone. This region was not found in all sensillaexamined by these authors, but failure to find it cannot be regarded as significant.Haskell & Schoonhoven (1969) do not record how many sensilla they tested for themechanoreceptor response. It may be that the present findings reflect the absence ofa mechanoreceptor neurone in many sensilla, or alternatively it may result from afailure of response in many sensilla at the time of recording. This would be in generalagreement with the variability of response shown by these sensilla. In either event, thedensity of sensilla on the palp tip is such that mechanoreceptor sensitivity, say as ameans of monitoring hardness of grass (Haskell & Schoonhoven, 1969), is unlikely tobe impaired significantly by the non-functioning of 50% of sensilla.
SUMMARY
1. Three morphologically and functionally different types of sensilla on the maxillarypalp tips of Locusta migratoria are distinguished and an electrophysiological study ismade of the most commonly occurring type.
2. Responses to stimulation with o-i M sodium chloride show impulses of one, twoor three amplitude categories in different sensilla.
3. Variability of response to a constant stimulus is appreciable, both between andwithin sensilla.
4. Tests with a range of compounds indicate that individual neurones are capableof responses to compounds of different chemical classes.
5. Specificity of sensilla is more marked than specificity of neurones.6. Methods of interpretation of results are evaluated and the results are discussed
in the context of discrimination of food plant materials.

292 W. M. BLANEY
I wish to thank Mrs V. E. Bone for technical assistance, Dr R. C. Lethbridge forhelp with photography and Dr R. F. Chapman for comments on the manuscript andfor many useful discussions. I am also grateful to the Royal Society and the CentralResearch Fund for grants in support of this work and to the Centre for Overseas PestResearch for an unfailing supply of locusts.
REFERENCES
BERNARD, J. & GUILLET, J. C. (197a). Changes in the receptor potential under polarizing currents intwo insect receptors. J. Insect Physiol. 18, 2173-87.
BERNAYS, E. A., BLANEY, W. M. & CHAPMAN, R. F. (1972). Changes in the chemoreceptor sensilla onthe maxillary palps of Locusta migratoria in relation to feeding. X exp. Biol. 57, 745-53.
BLANEY, W. M. & CHAPMAN, R. F. (1969). The fine structure of the terminal sensilla on the maxillary-palps of Schistocerca gregaria (Forskol) (Orthoptera, Acrididae). Z. Zellforsch. mikrosk. Anat. 99,74-97-
BLANEY, W. M. & CHAPMAN, R. F. (1970). The functions of the maxillary palps of Acrididae Ortho-ptera). Entomologia exp. appl. 13, 363-76.
BLANEY, W. M., CHAPMAN, R. F. & COOK, A. G. (1971). The structure of the terminal sensilla on themaxillary palps of Locusta migratoria (L.) and changes associated with moulting. Z. Zellforsch.mikrosk. Anat. 121, 48-68.
BLANEY, W. M., CHAPMAN, R. F. & WILSON, A. (1973). The pattern of feeding of Locusta migratoria(Orthoptera, Acrididae). Acrida (in the Press).
DETHIER, V. G. (1973). Electrophysiological studies of gustation in lepidopterous larvae. II. Tastespectra in relation to food-plant discrimination. .7. comp. Physiol. 82, 103-34.
DETHIER, V. G. & HANSON, F. E. (1965). Taste papillae of the blowfly. J. cell. comp. Physiol. 65, 93-100.DETHIER, V. G. & HANSON, F. E. (1968). Electrophysiological responses of the chemoreceptors of the
blowfly to sodium salts of fatty acids. Proc. natn. Acad. Science U.S.A. 60, 1296-303.DETHIER, V. G. & KUCH, J. H. (1971). Electrophysiological studies of gustation in lepidopterous larvae.
I. Comparative sensitivity to sugars, amino acids and glycosides. Z. vergl. Physiol, 72, 343-63.DOETSCH, G. S., GANCHROW, J. J., NELSON, L. M. & ERICKSON, R. P. (1969). Information processing
in the taste system of the rat. In Olfaction and Taste (ed. C. Pfaffmann), pp. 492-511. New York:Rockefeller University Press.
ERICKSON, R. P. (1963). Sensory neural patterns and gustation. In Olfaction and Taste (ed. Y. Zotter-man), pp. 205—13. London, New York: Pergamon Press.
ERICKSON, R. P. (1967). Neural coding of taste quality. In: The Chemical Senses and Nutrition (ed.M. R. Kare and O. Mailer), pp. 313-27. Baltimore, Md.: The John Hopkins Press.
GILBERT, B. L., BAKER, J. E. & NORRIS, D. M. (1967). Juglone (s-hydroxy-i,4-naphthoquinone) fromCarya ovata, a deterrent to feeding by Scolytus multistriatus. J. Insect Physiol. 13, 1453—9.
GILBERT, B. L. & NORRIS, D. M. (1968). A chemical basis for bark beetle (Scolytus) distinction betweenhost and non-host trees. J. Insect Physiol. 14, 1063-8.
GILLARY, H. L. (1969). Stimulation of an insect taste receptor. In Experiments in Physiology and Bio-chemistry (ed. G. A. Kerkut), pp. 41-57. London, New York: Academic Press.
HASKELL, P. T. & ScHOONHOVEN, L. M. (1969). The function of certain mouthpart receptors in relationto feeding in Schistocerca gregaria and Locusta migratoria migratorioides. Entomologia exp. appl. 12,423-40.
HODGSON, E. S., LETTVIN, J. Y. & ROEDER, K. D. (1955). Physiology of a primary chemoreceptor unit.Science, N.Y. 122, 417-18.
HODGSON, E. S. (1968). Taste receptors of arthropods. Invertebrate receptors. Symp. Zool. Soc. Lond.no. 23 (ed. J. D. Carthy and G. G. Newell), pp. 269-77.
HUNTER-JONES, P. (1961). Rearing and Breeding Locusts in the Laboratory. London.ISHIKAWA, S. (1966). Electrical response and function of a bitter substance receptor associated with the
maxillary sensilla of the silkworm, Bombyx mori L. J. cell Physiol. 67, 1—11.L E BERRE, J. R., SINOIR, Y. & BOULAY, C. (1967). Etude de l'equipment sensoriel de 1'article distal des
palps chez la larve de Locusta migratoria migratorioides (R. et F.). C. r. hebd. Se"anc. Acad. Sci.,Paris D 265, 1717-20.
LEWIS, C. T. (1970). Structure and function in some external receptors. Symp. R. ent. Soc. Lond. 5,59-76.
MCCUTCHAN, M. C. (1969). Responses of tarsal chemoreceptive hairs of the blowfly, Phormia regina.jf. Insect Physiol. 15, 2059-68.
MORITA, H. (1959). Initiation of spike potentials in contact chemosensory hairs of insects. III. D.C.stimulation and generator potential of labellar chemoreceptors of Calliphora. J. cell. comp. Physiol.54, 189-204.

Journal of Experimental Biology, Vol. 60, No. 2 Plate 1
W. M. BLANEY (Facing p. 292)

Journal of Experimental Biology, Vol. 60, No. 2 Plate 2
W. M. BLANEY

Electrophysiological responses of terminal sensilla of maxillary palps 293MORITA, H. & TAKEDA, K. (1957). The electrical resistance of the tarsal chemosensory hair of the
butterfly, Vanessa indica. J. fac. Set. Hokhaido Univ. (series 6) 13, 465-69.NICKLAUS, R., LUNQUIST, P.-G. & WERSXLL, J. (1967). Elektronmikroskopie am sensorischen Apparat
der Fadenhaare auf den Cerci der Schabe Periplaneta americana. Z. vergl. Physiol. 56, 412-15.NORRIS, D. M. (1969). Transduction mechanism in olfaction and gustation. Nature, Lond. 222, 1263-4.DEN OTTER, C. J. & VAN DER STARRE, H. (1967). Responses of tarsal hairs of the bluebottle, Calliphora
erythrocephala Meig., to sugar and water. J. Insect Physiol. 13, 1177-88.PFAFFMANN, C. (1941). Gustatory afferent impulses..?, cell. comp. Physiol. 17, 243-58.PFAFFMANN, C. (1955). Gustatory nerve impulses in rat, cat and rabbit. J. Neurophysiol. 18, 429-40.REES, C. J. C. (1967). Transmission of receptor potential in dipteran chemoreceptors. Nature, Lond.
215, 301-2.REES, C. J. C. (1968). The effect of aqueous solutions of some 1:1 electrolytes on the electrical response
of the type 1 ('salt') chemoreceptor cell in the labella of Phormia. J. Insect Physiol. 14, 1331-64.REES, C. J. C. (1969). Chemoreceptor specificity associated with choice of feeding site by the beetle,
Chrysolina brunsvicensis on its foodplant, Hypericum hirsutum. Entomologia exp. appl. 12, 565-83.SCHOONHOVEN, L. M. (1969). Gustation and foodplant selection in some lepidopterous larvae. Ento-
mologia exp. appl. 12, 555-64.SINHA, W. P. & GULATI, K. C. (1965). Neem seed cake as a source of pest control chemical. III.
Repellency against locusts. Proc. natn. Acad. Sci. India B 35, 338-43.SLIFER, E. H. (1970). The structure of arthropod chemoreceptors. A. Rev. Ent. 15, 121-42.VAN DER STARRE, H. (1970). Tarsal water receptor cell responses in the blowfly Phormia regina. Neth.
J. Zool. 20, 289-90.VAN DER STARRE, H. (1972). Tarsal taste discrimination in the blowfly Calliphora vicina Robineau-
Desvoidy. Neth. J. Zool. 22 (3), 227-82.STORCKOW, B. (1970). Responses of olfactory and gustatory receptor cells in insects. In Advances in
chentoreception, vol. 1 (ed. J. W. Johnston Jr., D. G. Moulton and A. Turk), pp. 107-59. New York:Appleton-Century-Crofts.
STORCKOW, B. (1971). Electrical impedance of the labellar taste hair of the blowfly Calliphora erythro-cephala Mg. Z. vergl. Physiol. 72, 131-43.
STORCKOW, B., HOLBERT, P. E. & ADAMS, J. R. (1967). Fine structure of the tip of chemosensitive hairsin two blowflies and the stable fly. Experientia 23, 780-2.
THURM, U. (1965). An insect mechanoreceptor. I. Fine structure and adequate stimulus. Cold Spr. Harb.Syrnp. quant. Biol. 30, 75-82.
WEAST, R. C. (ed.) (1964). In Handbook of Chemistry and Physics. Ohio: The Chemical Rubber Co.
EXPLANATION OF PLATES
PLATE I
(a) Microscope stage to show cradle (C), locust (L), indifferent electrode (I.E.) and recording electrode(R.E.). (b) Locust in position for recording, showing method of restraint, (c) Palp tip exposed forrecording.
PLATE 2
Scanning electron-micrograph of part of tip of maxillary palp showing several normal sensilla (N.and one small sensillum (S.S.). so = socket of sensillum, c = crest.



















