
Electromagnetic exposure effects the hippocampal dentate...
7
Indian Journal of Experimental Biology Vol. 39, December 2001, pp.I220-1226 Electromagnetic exposure effects the hippocampal dentate cell proliferation in gerbils (Meriones unguiculatus) K Hoffmann, F Bagorda, A F G Stevenson* & G Teuchert-Noodtt Department of Neuroanatomy, Faculty of Biology, University of Bielefeld, 33615 Bielefeld, Germany *Radiobiology Unit, Institute for Toxicology, University of Kiel, Brunswiker Str., D-24105 Kiel, Germany Received II September, 2000; revised 13 August, 2001 The chronic effect on hippocampal neurogenesis after exposure (30 min/day for 14 days) to a high frequency (35,53 kHz) electromagnetic field, double modulated at extremely low frequencies (ELF; I, 8, I2, 29 and 50 Hz), was studied in young adult gerbils. Immediately after the last exposure proliferation of dentate granule cells was identified by in vivo labeling with 5-bromo-2-desoxyuridine (BrdU). Exposure to I, 29 and 50 Hz resulted in a statistically signifucant reduction of cell proliferation rates, but only the 50Hz-group manifested the effect highly significantly (-29,3 %). On rhe other hand, gerbils exposed to 8 and I 2 Hz showed no significant change of postmitotic cell proliferation as compared with the sham treated controls. The results suggest that the effects of ELF on the granule cell proliferation are mediated by neurotmnsmitters and hormones which regulate hippocampal neurogenesis. Biological effects of exposure to magnetic, electric and electromagnetic fields are yet to be adequately investigated. In extremely low frequency (ELF) range, the magnetic and electric components of the field separate and may be expected to act independently on biological systems. As far as biological effects are concerned it is believed that the magnetic effects predominate overwhelmingly. Epidemiological studies from chronic exposure to magnetic fields showing increased risks for childhood brain tumors 1 and mental disorders 2 , indicate that the brain may be highly sensitive to ELF. Brain sensitivity to ELF exposure by an alteration of EEG (electro-encephalograph) 3 , which is a reflection of the electrical activity of the brain cortex has been reported. The implications are, however, not yet known. Most importantly, evidence on the perturbation of the neuroendocrine system by an exposure to ELF impairs neurotransmitters 5 also. Central to this issue is a suppression 6 or alternatively, an increase 7 of epiphyseal melatonin secretion. The function of the pineal organ-in particular the circadian rhythmic release of melatonin - is associated with other central neurotransmitter functions. In this context the influences of ELF on the homeostasis of tcorrespondent author Phone: +49-521106-5700. Fax: +49-521106-5844 e-mail: G.Teuchert @ Biologie.Uni- Bielefeld.DE the cholinergic 8 • 9 , serotonergic 10 and doparninergic 11 central systems are of vital importance. Experimental studies on rodents indicated adaptive changes of short-term memory 12 and open field behavior 13 to ELF exposure-the two patterns in which the limbic system is essentially involved. Behavioural studies on rodents under the influence of magnetic fields revealed that the spatial learning ability, which is a specific function of the limbic hippocampus, was affected 14 • 15 • Recently, the hifpocampal dentate gyrus (gyrus dentatus) of rodents 1 and humans 17 has been shown to be strongly implicated in neuroplasticity and learning on the basis of neurogenesis persisting for life. The above mentioned central neurotransmitter systems have been shown to take an essential part in the control of renewal of the granular cell Iayer 18 · 19 • Moreover, the glutamatergic NMDA (N-methyl-0- aspartate) receptor system, which i .s the main relay station of input to the hippocampal dentate gyrus, plays a primary role in the regulation of this cell proliferation 20 • Thus, it is important to know if post- partal neurogenesis may be affected by ELF exposure. Influence of ELF on the mitotic activity of cells is known 21 .2 2 • While one study has primarily been confined to the influence of mitogens on isolated Iymphocytes 23 , another study involved the response of T-cells in rats 24 • With the exception of these, there is
Transcript of Electromagnetic exposure effects the hippocampal dentate...

Indian Journal of Experimental Biology Vol. 39, December 2001, pp.I220-1226
Electromagnetic exposure effects the hippocampal dentate cell proliferation in gerbils (Meriones unguiculatus)
K Hoffmann, F Bagorda, A F G Stevenson* & G Teuchert-Noodtt Department of Neuroanatomy, Faculty of Biology, University of Bielefeld,
33615 Bielefeld, Germany
*Radiobiology Unit, Institute for Toxicology, University of Kiel, Brunswiker Str., D-24105 Kiel, Germany
Received II September, 2000; revised 13 August, 2001
The chronic effect on hippocampal neurogenesis after exposure (30 min/day for 14 days) to a high frequency (35,53 kHz) electromagnetic field, double modulated at extremely low frequencies (ELF; I, 8, I 2, 29 and 50 Hz), was studied in young adult gerbils. Immediately after the last exposure proliferation of dentate granule cells was identified by in vivo labeling with 5-bromo-2-desoxyuridine (BrdU). Exposure to I, 29 and 50 Hz resulted in a statistically signifucant reduction of cell proliferation rates, but only the 50Hz-group manifested the effect highly significantly (-29,3 %). On rhe other hand, gerbils exposed to 8 and I 2 Hz showed no significant change of postmitotic cell proliferation as compared with the sham treated controls. The results suggest that the effects of ELF on the granule cell proliferation are mediated by neurotmnsmitters and hormones which regulate hippocampal neurogenesis.
Biological effects of exposure to magnetic, electric and electromagnetic fields are yet to be adequately investigated. In extremely low frequency (ELF) range, the magnetic and electric components of the field separate and may be expected to act independently on biological systems. As far as biological effects are concerned it is believed that the magnetic effects predominate overwhelmingly.
Epidemiological studies from chronic exposure to magnetic fields showing increased risks for childhood brain tumors 1 and mental disorders2
, indicate that the brain may be highly sensitive to ELF. Brain sensitivity to ELF exposure by an alteration of EEG (electro-encephalograph)3
, which is a reflection of the electrical activity of the brain cortex has been reported. The implications are, however, not yet known. Most importantly, evidence on the perturbation of the neuroendocrine system by an exposure to ELF impairs neurotransmitters5 also. Central to this issue is a suppression6 or alternatively, an increase7 of epiphyseal melatonin secretion. The function of the pineal organ-in particular the circadian rhythmic release of melatonin - is associated with other central neurotransmitter functions. In this context the influences of ELF on the homeostasis of
tcorrespondent author Phone: +49-521106-5700. Fax: +49-521106-5844 e-mail: G.Teuchert @ Biologie.Uni-Bielefeld.DE
the cholinergic8•9
, serotonergic10 and doparninergic 11
central systems are of vital importance. Experimental studies on rodents indicated adaptive
changes of short-term memory 12 and open field behavior13 to ELF exposure-the two patterns in which the limbic system is essentially involved. Behavioural studies on rodents under the influence of magnetic fields revealed that the spatial learning ability, which is a specific function of the limbic hippocampus, was affected14
•15
• Recently, the hifpocampal dentate gyrus (gyrus dentatus) of rodents 1 and humans 17 has been shown to be strongly implicated in neuroplasticity and learning on the basis of neurogenesis persisting for life. The above mentioned central neurotransmitter systems have been shown to take an essential part in the control of renewal of the granular cell Iayer18
·19
•
Moreover, the glutamatergic NMDA (N-methyl-0-aspartate) receptor system, which i.s the main relay station of input to the hippocampal dentate gyrus, plays a primary role in the regulation of this cell proliferation20
• Thus, it is important to know if postpartal neurogenesis may be affected by ELF exposure.
Influence of ELF on the mitotic activity of cells is known21
.22
• While one study has primarily been confined to the influence of mitogens on isolated Iymphocytes23
, another study involved the response of T-cells in rats24
• With the exception of these, there is

HOFFMANN et al.: ELECTROMAGNETIC EFFECfS ON HIPPOCAMPAL DENTATE CELL PROLIFERATION 1221
no report on a cell renewal system in vivo. The present study has been undertaken to quantify effects of ELF on the granule cell proliferation against the background of those neurotransmitters and hormones, which are known to influence hippocampal neurogenesis. With regard to hippocampal functions ELF results may have implications for neuroplasticity.
Materials and Methods Male gerbils (Meriones unguiculatus) were bred
and maintained under standard conventional animal housing conditions. The animals were kept in macrolon cages under natural day-night conditions with water and food available ad libitum. Sexual maturity is acquired at about 60 days of age.
Exposure to magnetic fields--From day 75 postpartum, the animals received daily 30 min exposures to a high frequency (35.53 kHz) electromagnetic field modulated at very low frequencies. The total exposure period was 15 days. The high frequency was pulsemodulated and settings of the coil for magnetic fields at 1 Hz (n=IO), 8 Hz (n=8), 12 Hz (n=l2), 29 Hz (n=IO) and 50 Hz (n=IO) produced in each case a rlouble modulated signal, as indicated by the frequency pulses obtained from a spectrum analyser (Advantest R3131, Rohde & Schwanz, Munich, Germany). The time difference between successive pulses was not identical but constant. During exposures the animals remained in their cages which were placed in the exposure-device made of a drum (Helrnholz double-coil) open at both ends [Biomedical Systems (Model MOMS 2000) Wiesbaden, Germany].
BrdU-labelin~The method of quantitative evaluation of cell proliferative activity in the dentate gyrus by immuno-cytochemical means has been described previously 19
"25
• In short, after 2 hr of the last exposure, the animals received 30 mglkg BrdU (Cell Proliferation Kit RPN, Amersham, Braunschweig, Germany) by intraperitoneal injection. The animals were sacrificed 7 days later, brains removed and freeze-sectioned at 12 ~m horizontal sections from the septal to temporal pole of the hippocampus (Fig. I). Every third section (in sum 38) per brain was collected on slides and processed for immunocytochemical stammg of nuclei which had incorporated the BrdU during scheduled DNA synthesis. The total number of labeled cell profiles in the granular and subgranular areas of the dentate
gyrus were counted under a Zeiss microscope at 250x.
Statistical evaluation--Mean values were computed as arithmetic means ± SD and compared by a two-tailed t-test after checking the dispersion of variances using Fisher's test. The distribution along the septa-temporal axis was subjected to a computeraided regression analysis. Additionally, the results were compared between groups with a two-way analysis of variance with repeated measurements (ANOV A). Level of significance was set at P<0.05.
Results A distinct proportion of cell nuclei in the
germinative zone of the subgranular layer in all groups showed BrdU-labeling. There were, however, differences in the pattern of distribution of the labeling. While in the controls and in the groups exposed to 8 and 12 Hz, the labelings were denser and appeared in clusters, in the groups exposed to 1 Hz, 29 Hz and 50 Hz, the labelings were less dense and not clustered (Fig. 2).
Fig. 1-Horizontal planes from the septal to temporal pole in the brain from which the sections were taken (A). B, C Gross structural illustrations of typical sections from a septal and a temporal plane, respectively. The double-lined rectangle indicates the area (infrapyramidal blade) at which the BrdU-nuclei were photomicrographed (see Fig. 2). The dotted lines represent the germinal layer in the subgranular zone where counting was done.

1222
m ~
.. ' \ '. .
:g •.r:.
.; . .... . ~~ i . ~ ..
h
m
'· ..
:.. .t ... . . . ,. ..
h
m
.. ~
r ,. '· " ' ~ ... It
h -
INDIAN J EXP BIOL, DECEMBER 2001
1 Hz
..
29Hz
50 Hz
8Hz m
P,< . . -· g __ . f• ~ :0
~ • • ... ........
"' . ... • y ••
12.Hz • m
• ; , -,. -~· .
• .., • h • ..
J
m. control •
' • :<. • • • ~# . 1 ' f~ ' ..,.,. .... ' \ ""' ..
7 ~ ~ ... - . 6' .. ( ' ~.. • , . •, J • , I a~.,. . •c. •,. ). . ·• . ~ . -- ~ .. " ~ . . ... .
• ~ • & .,-; . • .. ' • .. \ . « • ' 1- ) .... . • ·'• •,.• ,· _.. f• • . ,A .~ ;_# 6 "'' ., . . ,. . ._.,.,
• "f' • , ''• ·• -1 f II"\. •' ! • •• • • . ... .... a: , .. '. • ' . 4 c.• .... .. .. ... ·** . . '
....
•• ....
• ' .
' !
• ,
Fig. 2- Photomicrographs of representative BrdU-Iabeled nuclei in the germinative zone at the base of the granular layer in the dentate gyrus from exposed and control groups. (m = molecular layer, g =granular layer, h = hilar layer, horizontal bar= 80 11m)
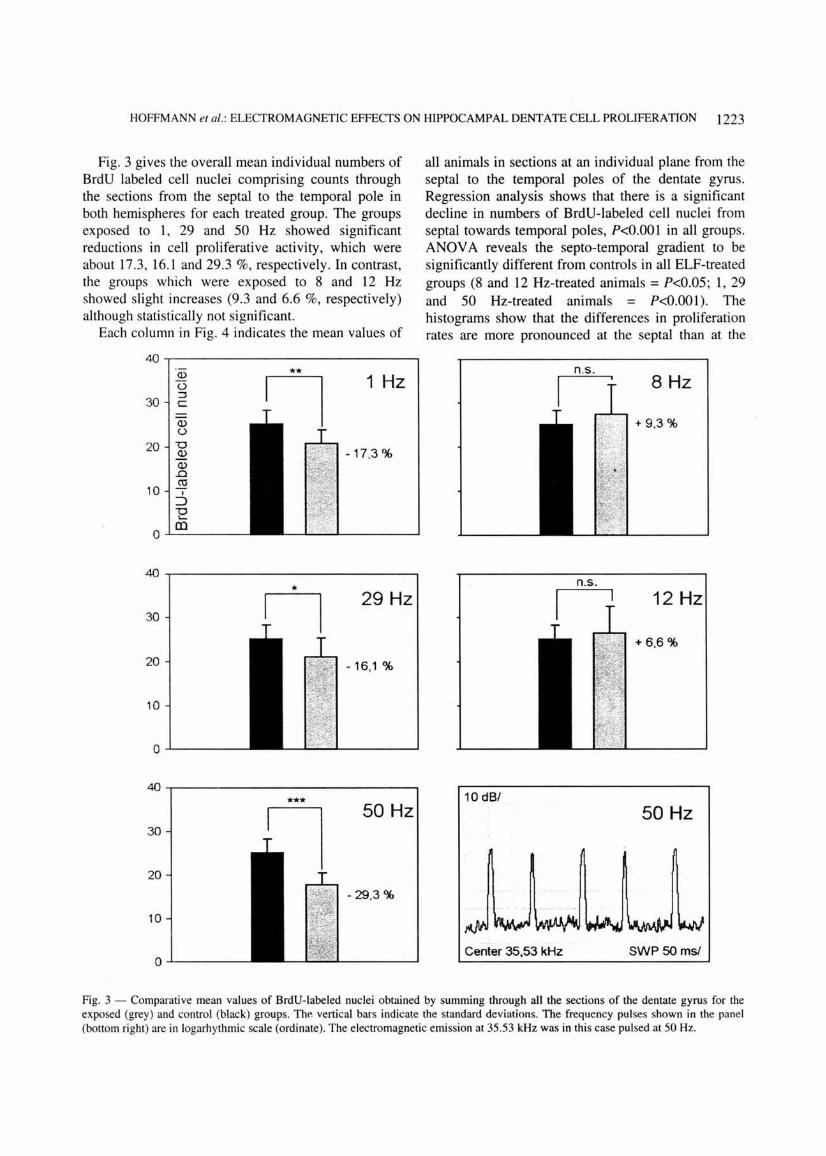
HOFFMANN et al. : ELECTROMAGNETIC EFFECTS ON HIPPOCAMPAL DENTATE CELL PROLIFERATION 1223
Fig. 3 gives the overall mean individual numbers of BrdU labeled cell nuclei comprising counts through the sections from the septal to the temporal pole in both hemispheres for each treated group. The groups exposed to 1, 29 and 50 Hz showed significant reductions in cell proliferative activity, which were about 17.3, 16.1 and 29.3 %, respectively. In contrast, the groups which were exposed to 8 and 12 Hz showed slight increases (9.3 and 6.6 %, respectively) although statistically not significant.
Each column in Fig. 4 indicates the mean values of
40 ~----------------------------~ Q)
0 1 Hz 30
::J c
Q) 0
20 u Q) - 17,3% Q) .0
10 co
I
::J "b <....
0 (])
40
29Hz 30
20 - 16,1 %
10
0
40
*** 50 Hz 30
20
-29,3%
10
0
all animals in sections at an individual plane from the septal to the temporal poles of the dentate gyrus. Regression analysis shows that there is a significant decline in numbers of BrdU-labeled cell nuclei from septal towards temporal poles, P<0.001 in all groups . ANOV A reveals the septa-temporal gradient to be significantly different from controls in all ELF-treated groups (8 and 12 Hz-treated animals = P<0.05; 1, 29 and 50 Hz-treated animals = P<0.001). The histograms show that the differences in proliferation rates are more pronounced at the septal than at the
8Hz
+9,3%
12Hz
+ 6,6%
10 dB/ 50 Hz
Center 35,53 kHz SWP 50 msl
Fig. 3 - Comparative mean values of BrdU-labeled nuclei obtained by summing through all the sections of the dentate gyrus for the exposed (grey) and control (black) groups. The vertical bars indicate the standard deviations. The frequency pulses shown in the panel (bottom right) are in logarhythmic scale (ordinate). The electromagnetic emission at 35.53 kHz was in this case pulsed at 50 Hz.

1224 INDIAN J EXP BIOL, DECEMBER 2001
temporal pole in 1 Hz (P<O.OOI vs. <0.05), 12 Hz (P<O.OS vs. n.s.) and 50 Hz-treated animals (P<O.OO 1 vs. <0.01). Discussion
The seemingly contradictory effects--on the one hand a decrease and on the other a tendential increase in the granule cell proliferative activity--draw attention and is a feature which is often pointed at to argue against the occurrence of biological effects owing to exposures to electric, magnetic and electromagnetic fields (which shall collectively be
referred to here as EM fields). The basis of argumentation is the obvious lack of a clearcut doseeffect relationship, as it is known in radiobiology and general toxicology. However, this general principle cannot be applied indiscriminately to EM fields as these operate differently. The principle which has been recognised is the Window Effect. Studies26 have shown that windows exist for frequencies as well as for amplitudes i.e. the studied effects alternately appear and disappear in dependence on either frequencies and/or amplitudes. The seeming
80 .-----------------------------~
60 1Hz 8 Hz
40
20
0
"(ii 80
() ::::s c 60
29 Hz 12 Hz (l) ()
"0 40 .E;? (l) .c ro 20
I
:J "E (]) 0
80
60 50 Hz control
40
20
0 4 7 1 0 13 16 19 22 25 28 31 34 37 4 7 10 13 16 19 22 25 28 31 34 37
septal+- slice number - • temporal
Fig. 4 --- Mean values of the BrdU-Iabeled nuclei of each section (1-38) from septal to temporal poles for exposed and control groups. Each column represents the mean of all animals obtained from all sections at an individual plane from the septal to the temporal pole. Regression analysis shows significant gradients (P<O.OO I) in all groups. In all ELF groups the septo-temporal gradient is significantly different from controls (ANOYA). Statistical signi ficance was calculated by two-way analysis of variance.

HOFFMANN eta/.: ELECTROMAGNETIC EFFECTS ON HIPPOCAMPAL DENTATE CELL PROLIFERATION 1225
discrepancy in the results presented here may in fact be just an expression of the Window Effect.
The neurogenic activity in the hippocampal dentate gyrus determined in terms of cell proliferation (scheduled DNA synthesis marked by BrdU-label) in the germinative layer, may be influenced by neuroregulatory substances via the glutamatergic NMDA-receptor system. Some of these may be neurotransmitters 18
. We have recently shown that acute doses of the dopaminergic agonist methamphetamine decrease granule cell proliferation in the dentate gyrus 19 and acute doses of the antagonist haloperidol increase ie5
. Hence, we have argued that the dopaminergic system might be involved in the in vivo regulation of hippocampal plasticity. Since different investigations indicate that .the activity of dopaminergic neurons in the brainstem is sensitive to magnetic stimulation 11
, we have reason to assume a direct influence of EM fields via the meso-limbic dopamine system on hippocampal neurogenesis, mediated by the glutamatergic NMDAreceptor system. The direct influence of EM fields on melatonin secretion and the interaction of melatonin with dopamine have already been mentioned5
-7
•
Moreover, it should be noted that melatonin generally inhibits the secretion of neuronal releasing factors (RFs) which affect the hypophyseal secretion of specific endocrine gland stimulatory hormones27
. To what extent the proliferative activities of the dentate germinative layer, as indicated by these experiments, have been affected either directly through the glutamatergic and dopaminergic systems, respectively, or alternatively over the endocrine system, cannot be assessed here.
The implications of these measurements remain a matter of speculation. The fact that there was topological preference in the inhibitory but not in the stimulatory effect is interesting but cannot at the moment be interpreted. Such an effect could bear profound implications, depending as to whether specific hippocampal functions are lodged in discrete topological locations. The behavior of the exposed animals (observations made during the course of experiments) was altered to an extent, which may be shortly characterised as follows . Aberrant behavior was observed only during the period of exposure. Exposure at 50 Hz evoked aggression denoted by scratching and escape attempts. The groups exposed to 1, 8 and 12 Hz showed essentially the same but Jess intense behavioral change. On the other hand, the animals exposed to 29 Hz showed calming and lapses into stupor lasting several minutes. Since the learning
ability is thought to be affected by altered neurogenesis in the dentate gyrus28
·29
, it would be of interest to appraise such eventual effects in exposed animals particularly in view of pu~orted changes in hippocampal associated behaviors 2
-15 subjected to
ELF exposures. The continual increase in the commercial use of ELF-pulsed signals in the field of electronic communications warrants such studies .
Acknowledgement The dosimetric estimations and advice of Dr. L von
Klitzing and R Anderson, Department of Experimental Medicine, University of Luebeck, are greatly appreciated.
References I Savitz D A, John F M & Kleckner R C , Magnetic fie ld
exposure appliances and -childhood cancer, Am J Epidemio/, 131 (1990) 763.
2 Wilson B W, Chronic exposure to ELF field s may induce depression , Bioelectromagnetics. 9 (1988) 195.
3 Bell G B, Marino A A & Chesson A L, Alterations in brain electrical activity caused by magnetic fields: detecting the detection process, Electroencephalography Clinic Neuropysiol, 83 ( 1992) 389.
4 Wilson B W, Matt K S, Morri s J E, Sasser L B, Miller D L & Anderson L E, Effects of 60 Hz magnetic field exposure on the pineal and hypothalamic-pituitary-gonadal axis in the siberian hamster (Phodopus sungorus), BioelectromagneTics, 20 (1999) 224.
5 Kun-Ruey Shieh, Yey-Shiu Chu & Jenn-Tser Pan , Circadian change of dopaminergic neuron activity: effects of co nstant light and melatonin, Neuro Report, 8 (1997) 2283.
6 Selmaoui B & Touitou Y, Sinusoidal 50-Hz magnetic fie lds depress rat pineal NAT activity and serum melatonin. Role of duration and intensity of exposure, Life Sci, 57/14 ( 1995) 1351 .
7 Niehaus M, Bruggemeyer H, Behre H M & Lerch! A, Growth retardation, testicular Djungarian stimulation and increased melatonin synthesis by weak magnetic field s (50 Hz) in hamsters , Phodopus sungorus, Biochem Biophys Res Commun, 234 (1997) 171.
8 Lai H, Carino M, Horita A & Guy A W, Effects of 60 Hz magnetic field on central cholinergic systems of the rat, Bioelectromagnetics, 14 (1993) 5.
9 Lai H & Carino M, 60 Hz magnetic fields and central
cholinergic activity : effects of exposure, intensity and
duration, Bioelectromagnetics, 20 ( 1999) 284. 10 Lerch! A, Nonaka K 0, Stokkan K A & Reiter R J, Marked
rapid alterations in nocturnal pineal serotonin metabolism in mice and rats exposed to weak intermittent magnetic fie lds, Biochem Biophys Res Commun , 169 (1990) 102.
II Orlova T V, Sidyakin V G, Kulichenko A M & Pavlenko V B, Activity of the neurons of the parietal associative cortex and the region of the substantia nigra of the cat on exposure to magnetic fields of the frequency of 8 Hz, Biophysics, 4015 (1995) 977.
12 Lyskov Y B, Chernysev M V, Mikhailov V 0 , Kozlov A P, Makarova T M, Vasii'Yeva Y V, Druzin MY, Sokolov G V

1226 INDIAN J EXP BIOL, DECEMBER 200 I
& Vishnevskii A M, The effect of a magnetic field with the frequency of 50 Hz on behavior in rats depends on the value of the constant magnetic field, Biophysics, 4114 ( 1996) 881 .
13 Deryugina 0 N, Pisachenko T M & Zhadin M N, Combined action of alternating and constant magnetic fields on the behavior in rats in the open " field", Biophysics, 4113 (1996) 769.
14 Lai H, Spatial learning deficit in the rat after exposure to a 60Hz magnetic field, Bioelectromagnetics, 17/6 (1996) 494.
15 Sienkiewics Z J, Haylock R G, Bartrum R & Saunders R D, 50 Hz magnetic field effects on the performance of a spatial learning task by mice, Bioelectromagnetics, 19/8 (1998) 486.
16 BayerS A, Changes in total number of dentate granule cells in juvenile and adult rats: a correlated volumetric and 3Hthymidine autoradiographic study, Exp Brain Res, 46 (1982) 315.
17 Eriksson P S, Perfilieva E, Bjork-Eriksson T, Alborn A-M, Nordberg C, Peterson D A & Gage F H, Neurogenesis in ihe adult human brain, Nature Medicine, 4/JJ (1998) 1313.
18 Cameron H A, Hazel T G & Me Kay R D G, Regulation of neurogenesis by growth factors and neurotransmitters, J Neurobiol, 36 ( 1998) 287.
19 Teuchert-Noodt G, Dawirs R R & Hildebrandt K, Adult treatment with methamphetamine transiently decreases dentate granule cell proliferation in the gerbil, J Neural Transm, 107 (2000) 133.
20 Cameron H A, Gould E & McEwen B S, Regulation of adult neurogenesis by excitatory input and NMDA-receptor activation in the dentate gyrus, J Neurosci, 15 (1995) 4687.
2 1 Kwee S & Raskmark P, Changes in cell proliferation due to environmental non-ionizing radiation. I. ELF electromagnetic fields, Bioelectrochem Bioenerg, 36 (1995) 109.
22 Goodman E M, Greenebaum B & Marron M T, Effects of electromagnetic fields on molecules and cells, lnt Rev Cytol, !58 (1995) 279.
23 Cadossi R, Bersani F, Cossarizza A, Zucchini P, Emilia G, Tortelli G & Franceschi C, Lymphocytes and low-frequency electromagnetic fields, Faseb J, 6 (1992) 2667.
24 Mevissen M, Haussler M, Szamel M, Emmendorffer A, Thun-Buttersby S & Loscher W, Compl<:x effects of longterm 50 Hz magnetic field exposer in vivo on immune functions in female sprague-dawley rats depend on duration of exposer, Bioelectromagnetics, 19 ( 1998) 259.
25 Dawirs R R, Hildebrandt K & Teuchert-Noodt G, Adult treatment with haloperidol increases dentate granule cell proli feration in the gerbil hippocampus, J Neural Transm, 105 (1998) 317.
26 Liboff A R, McLeod B R, Smith S D, Ion cyclotron
resonance effects of ELF fields in biological systems, in
Extremely low frequency electromagnetic f ields: The question
of cancers, edited by B W Wilson, R G Stevens & L E
Anderson (Battelle Press, Columbus, Ohio ) 1990, 383. 27 Vazquez D M, Stress and the developing limbic
hypothalamic-pituitary-adrenal axis, Psychoneuroendocrino/ogy, 23/7 (1998) 663.
28 Gould E, Beylin A, Tanapat P, Reeves A & Shors T J,
Learning enhances adult neurogenesis in the hippocampal
formation, Nature Neurosci, 213 ( 1999) 260.
29 Teuchert-Noodt G, Neuronal degeneration and reorgani
zation: a mutual prinicple in pathological and healthy
interactions of limbic and prefrontal circuits, J Neural
Transm , (Suppl) 60 (2000) 315.


















