
Effect of the molar ratio of an energy source to the substrate on yeast-mediated production of...
5
JOURNAL OF BIOSCIENCE AND BIOENGINEERING Vol. 89, No. 4, 329-333. 2000 Effect of the Molar Ratio of an Energy Source to the Substrate on Yeast-Mediated Production of 2-Chloro-a-Methylbenzyl Alcohol YURI HASEGAWA, SHUJI ADACHI,* AND RYUICHI MATSUNO Division of Applied Life Sciences, Graduate School of Agriculture, Kyoto University, Sakyo-ku, Kyoto 6068502, Japan Received 21 October 1999/Accepted 27 December 1999 Pachysolen tannophilus cells immobilized in Ca-alginate gels were shown to catalyze the asymmetric reduc- tion of acetophenone (AP) and chloroacetophenones (CLAPS) to their corresponding alcohols. The position of the Cl-group on the aromatic ring of AP greatly affected the reaction rate, and o-CLAP was the most readily reduced. For the reduction of o-CLAP to 2-chloro-a-methylbenzyl alcohol, the effect of the molar ratio of the energy source, glucose, to the substrate was examined in both batch and continuous operations, and a molar ratio much lower than that conventionally used was found to be sufficient for the reduction. [Key words: energy source, asymmetric reduction, chloroacetophenone, 2-chloro-n-methylbenzyl alcohol, Pachysolen tannophilus] In recent decades, viable cells have been recognized as catalysts for chemical synthesis. Cells intrinsically have a wide variety of enzymes which have the potential to cata- lyze many kinds of reactions. Microorganisms are consi- dered suitable for this purpose in view of their rapid growth rate and the convenience of their treatment, com- pared to animal or plant cells. Among the many known species, organic chemists have traditionally used bakers’ yeast, which can be cultivated without special equipment for strict sterilization or knowledge of advanced tech- niques in microbiology, and is regarded as being nontox- ic to the environment (1, 2). Yeast cells have a good abil- ity to asymmetrically reduce a ketone group in a single- step reaction to form a homochiral alcohol, while a series of complex reactions is usually involved in organ- ic-metal-catalyzed chemical reactions (3, 4). Hence, yeast cells have been harnessed to introduce a chiral center into achiral compounds (5-S). In the yeast-mediated asymmetric reduction of ketones, the cells require some energy source to regenerate the cofactor NAD(P)H, and, in general, sugars or ethanol are added to the reaction mixture as the energy source. Little attention has, however, been paid to the quantum of the energy source input; quite often, a large excess of energy source has been added to the reaction mixture with the result that the product is contaminated with a large amount of un- reacted energy source. Microorganisms would form a cer- tain amount of product per unit consumed of a given energy source. A large excess of the residual energy source in the reaction mixture would make product recovery from the solution and further purification proc- esses difficult and expensive. A proper, quantitatively- controlled investment of the energy source could be a key to the cost-effective operation of microbial processes for the production of useful materials and the industrializa- tion of such processes. We investigated the relationship between the amount of product formed and the quantum of energy source input in regard to microbial asymmetric reduction of o- chloroacetophenone (o-Cl-AP) to produce 2-chloro-cw- methylbenzyl alcohol (o-chloro-1-phenylethanol, o-Cl-l- PA) as a model reaction (Fig. 1). We previously reported * Corresponding author. that the yeast Pachysolen tannophilus IF0 1007 could catalyze the enantioselective oxidoreduction of l- phenylethanol/acetophenone (9); in this study, the reac- tion system was applied to the chloro-derivatives using the same yeast. The yeast species P. tannophilus is reported to utilize xylose as a carbon source (10, ll), but our preliminary study revealed that glucose could be a better potential energy source than xylose or ethanol for this reaction system. Hence, we adopted glucose as the energy source in this study. We examined the amount of glucose required to efficiently convert a given amount of o-Cl-AP in batch and continuous operation modes. MATERIALS AND METHODS Microorganism and materials The yeast P. tannophi- lus IF0 1007 was supplied by the Institute for Fermenta- tion, Osaka. The cells were stored at 4°C on a 1.5X- agarose slant of YM medium (0.3% (w/v) yeast extract, 0.3% malt extract, 0.5% peptone and 1% D-glucose; pH6.0). o-, m-, p-Chloroacetophenones (o-, m-, p-Cl- APs) and their corresponding alcohols, 2-, 3-, 4-chloro- a-methylbenzyl alcohols (o-, m-, p-Cl-~-PAS) were ob- tained from Tokyo Kasei Kogyo Co. Ltd., Tokyo. Aceto- phenone (AP) and 1-phenylethanol (a-phenethyl alcohol, l-PA) were purchased from Nacalai Tesque, Kyoto. m- Cl-l-PA was of analytical grade, and o-, p-Cl-~-PAS and Cl-APs were of reagent grade. All other chemicals used were of analytical grade and were obtained from either Wako Pure Chemical Industries (Osaka) or Nacalai Tes- que. Preparation of biocatalyst A seed culture grown on an agarose slant of YM medium at 24°C for 48 h was in- oculated into two test tubes each containing 3.5 ml of the YPD medium (1% yeast extract, 2% peptone and Asymmetric P o-Chloroacetophenone 2-Chloro-a-methylbenzyl alcohol (o-CLAP) (cdhloro-1-phenylethanol, NZI-l-PA) FIG. 1. Reaction scheme. 329
-
Upload
yuri-hasegawa -
Category
Documents
-
view
212 -
download
0
Transcript of Effect of the molar ratio of an energy source to the substrate on yeast-mediated production of...

JOURNAL OF BIOSCIENCE AND BIOENGINEERING Vol. 89, No. 4, 329-333. 2000
Effect of the Molar Ratio of an Energy Source to the Substrate on Yeast-Mediated Production of 2-Chloro-a-Methylbenzyl Alcohol
YURI HASEGAWA, SHUJI ADACHI,* AND RYUICHI MATSUNO Division of Applied Life Sciences, Graduate School of Agriculture, Kyoto University, Sakyo-ku, Kyoto 6068502, Japan
Received 21 October 1999/Accepted 27 December 1999
Pachysolen tannophilus cells immobilized in Ca-alginate gels were shown to catalyze the asymmetric reduc- tion of acetophenone (AP) and chloroacetophenones (CLAPS) to their corresponding alcohols. The position of the Cl-group on the aromatic ring of AP greatly affected the reaction rate, and o-CLAP was the most readily reduced. For the reduction of o-CLAP to 2-chloro-a-methylbenzyl alcohol, the effect of the molar ratio of the energy source, glucose, to the substrate was examined in both batch and continuous operations, and a molar ratio much lower than that conventionally used was found to be sufficient for the reduction.
[Key words: energy source, asymmetric reduction, chloroacetophenone, 2-chloro-n-methylbenzyl alcohol, Pachysolen tannophilus]
In recent decades, viable cells have been recognized as catalysts for chemical synthesis. Cells intrinsically have a wide variety of enzymes which have the potential to cata- lyze many kinds of reactions. Microorganisms are consi- dered suitable for this purpose in view of their rapid growth rate and the convenience of their treatment, com- pared to animal or plant cells. Among the many known species, organic chemists have traditionally used bakers’ yeast, which can be cultivated without special equipment for strict sterilization or knowledge of advanced tech- niques in microbiology, and is regarded as being nontox- ic to the environment (1, 2). Yeast cells have a good abil- ity to asymmetrically reduce a ketone group in a single- step reaction to form a homochiral alcohol, while a series of complex reactions is usually involved in organ- ic-metal-catalyzed chemical reactions (3, 4). Hence, yeast cells have been harnessed to introduce a chiral center into achiral compounds (5-S). In the yeast-mediated asymmetric reduction of ketones, the cells require some energy source to regenerate the cofactor NAD(P)H, and, in general, sugars or ethanol are added to the reaction mixture as the energy source. Little attention has, however, been paid to the quantum of the energy source input; quite often, a large excess of energy source has been added to the reaction mixture with the result that the product is contaminated with a large amount of un- reacted energy source. Microorganisms would form a cer- tain amount of product per unit consumed of a given energy source. A large excess of the residual energy source in the reaction mixture would make product recovery from the solution and further purification proc- esses difficult and expensive. A proper, quantitatively- controlled investment of the energy source could be a key to the cost-effective operation of microbial processes for the production of useful materials and the industrializa- tion of such processes.
We investigated the relationship between the amount of product formed and the quantum of energy source input in regard to microbial asymmetric reduction of o- chloroacetophenone (o-Cl-AP) to produce 2-chloro-cw- methylbenzyl alcohol (o-chloro-1-phenylethanol, o-Cl-l- PA) as a model reaction (Fig. 1). We previously reported
* Corresponding author.
that the yeast Pachysolen tannophilus IF0 1007 could catalyze the enantioselective oxidoreduction of l- phenylethanol/acetophenone (9); in this study, the reac- tion system was applied to the chloro-derivatives using the same yeast. The yeast species P. tannophilus is reported to utilize xylose as a carbon source (10, ll), but our preliminary study revealed that glucose could be a better potential energy source than xylose or ethanol for this reaction system. Hence, we adopted glucose as the energy source in this study. We examined the amount of glucose required to efficiently convert a given amount of o-Cl-AP in batch and continuous operation modes.
MATERIALS AND METHODS
Microorganism and materials The yeast P. tannophi- lus IF0 1007 was supplied by the Institute for Fermenta- tion, Osaka. The cells were stored at 4°C on a 1.5X- agarose slant of YM medium (0.3% (w/v) yeast extract, 0.3% malt extract, 0.5% peptone and 1% D-glucose; pH6.0). o-, m-, p-Chloroacetophenones (o-, m-, p-Cl- APs) and their corresponding alcohols, 2-, 3-, 4-chloro- a-methylbenzyl alcohols (o-, m-, p-Cl-~-PAS) were ob- tained from Tokyo Kasei Kogyo Co. Ltd., Tokyo. Aceto- phenone (AP) and 1-phenylethanol (a-phenethyl alcohol, l-PA) were purchased from Nacalai Tesque, Kyoto. m- Cl-l-PA was of analytical grade, and o-, p-Cl-~-PAS and Cl-APs were of reagent grade. All other chemicals used were of analytical grade and were obtained from either Wako Pure Chemical Industries (Osaka) or Nacalai Tes- que.
Preparation of biocatalyst A seed culture grown on an agarose slant of YM medium at 24°C for 48 h was in- oculated into two test tubes each containing 3.5 ml of the YPD medium (1% yeast extract, 2% peptone and
Asymmetr ic P
o-Chloroacetophenone 2-Chloro-a-methylbenzyl alcohol (o-CLAP) (cdhloro-1-phenylethanol, NZI-l-PA)
FIG. 1. Reaction scheme.
329
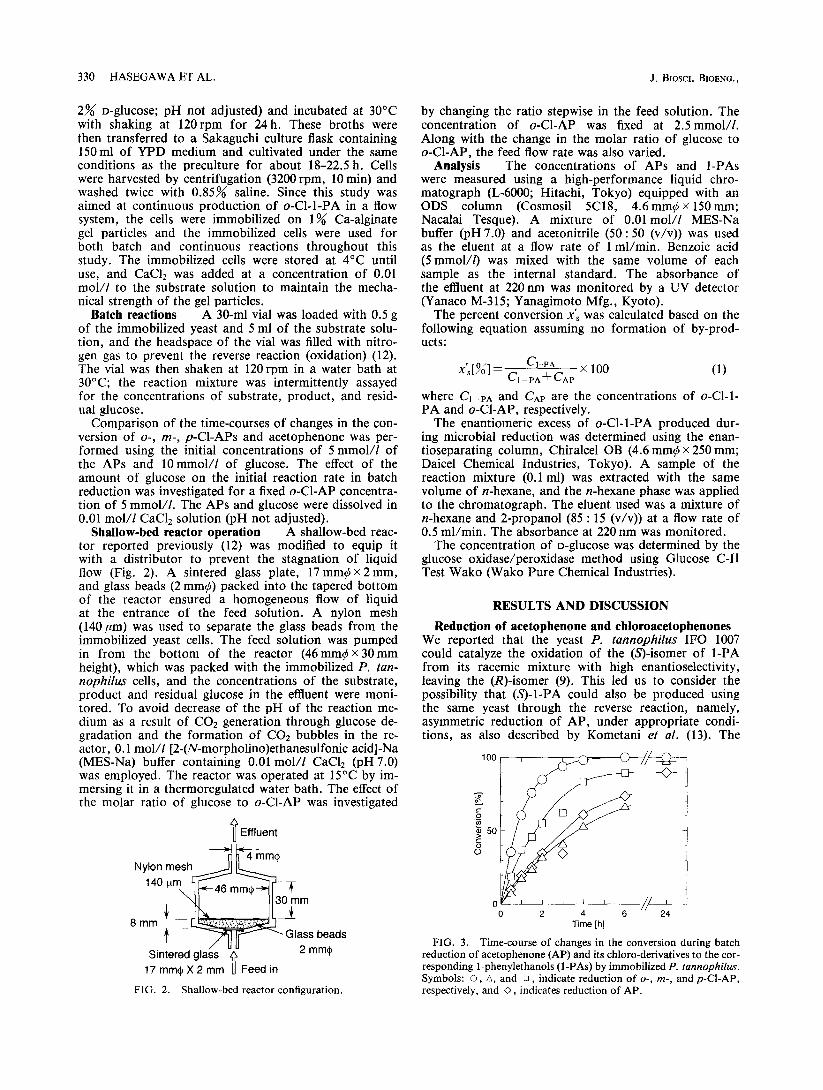
330 HASEGAWA ET AL. J. BIOSCI. BIOENG.,
2% n-glucose; pH not adjusted) and incubated at 30°C with shaking at 120rpm for 24 h. These broths were then transferred to a Sakaguchi culture flask containing 150 ml of YPD medium and cultivated under the same conditions as the preculture for about 18-22.5 h. Cells were harvested by centrifugation (3200 rpm, 10 min) and washed twice with 0.85% saline. Since this study was aimed at continuous production of o-Cl-l-PA in a flow system, the cells were immobilized on 1% Ca-alginate gel particles and the immobilized cells were used for both batch and continuous reactions throughout this study. The immobilized cells were stored at 4°C until use, and CaC& was added at a concentration of 0.01 mol/l to the substrate solution to maintain the mecha- nical strength of the gel particles.
Batch reactions A 30-ml vial was loaded with 0.5 g of the immobilized yeast and 5 ml of the substrate solu- tion, and the headspace of the vial was filled with nitro- gen gas to prevent the reverse reaction (oxidation) (12). The vial was then shaken at 120rpm in a water bath at 30°C; the reaction mixture was intermittently assayed for the concentrations of substrate, product, and resid- ual glucose.
Comparison of the time-courses of changes in the con- version of o-, m-, p-Cl-APs and acetophenone was per- formed using the initial concentrations of 5 mmol/l of the APs and lOmmol/l of glucose. The effect of the amount of glucose on the initial reaction rate in batch reduction was investigated for a fixed o-Cl-AP concentra- tion of 5 mmol/l. The APs and glucose were dissolved in 0.01 mol/l CaCl* solution (pH not adjusted).
Shallow-bed reactor operation A shallow-bed reac- tor reported previously (12) was modified to equip it with a distributor to prevent the stagnation of liquid flow (Fig. 2). A sintered glass plate, 17 mm4 X 2mm, and glass beads (2mm$) packed into the tapered bottom of the reactor ensured a homogeneous flow of liquid at the entrance of the feed solution. A nylon mesh (140/zm) was used to separate the glass beads from the immobilized yeast cells. The feed solution was pumped in from the bottom of the reactor (46mm$x 30mm height), which was packed with the immobilized P. tan- nophilus cells, and the concentrations of the substrate, product and residual glucose in the effluent were moni- tored. To avoid decrease of the pH of the reaction me- dium as a result of CO* generation through glucose de- gradation and the formation of CO2 bubbles in the re- actor, 0.1 mol/f [2-(N-morpholino)ethanesulfonic acid]-Na (MES-Na) buffer containing 0.01 mol/f CaClz (pH 7.0) was employed. The reactor was operated at 15°C by im- mersing it in a thermoregulated water bath. The effect of the molar ratio of glucose to o-Cl-AP was investigated
v Effluent
/ uu Sintered glass V 2 mm+
17mm+X2mm Feed in
FIG. 2. Shallow-bed reactor configuration.
by changing the ratio stepwise in the feed solution. The concentration of a-Cl-AP was fixed at 2.5 mmol/l. Along with the change in the molar ratio of glucose to o-Cl-AP, the feed flow rate was also varied.
Analysis The concentrations of APs and I-PAS were measured using a high-performance liquid chro- matograph (L-6000; Hitachi, Tokyo) equipped with an ODS column (Cosmosil 5C18, 4.6 mm+ x 150 mm; Nacalai Tesque). A mixture of 0.01 mol/f MES-Na buffer (pH 7.0) and acetonitrile (50 : 50 (v/v)) was used as the eluent at a flow rate of 1 ml/min. Benzoic acid (5 mmol/r) was mixed with the same volume of each sample as the internal standard. The absorbance of the effluent at 220 nm was monitored by a UV detector (Yanaco M-315; Yanagimoto Mfg., Kyoto).
The percent conversion x’~ was calculated based on the following equation assuming no formation of by-prod- ucts:
Cl--PA xLI%l= ClpPA+CAP x 100 (1)
where Cl-PA and CAP are the concentrations of o-cl-l- PA and a-Cl-AP, respectively.
The enantiomeric excess of o-Cl-l-PA produced dur- ing microbial reduction was determined using the enan- tioseparating column, Chiralcel OB (4.6 mm$ x 250 mm; Daicel Chemical Industries, Tokyo). A sample of the reaction mixture (0.1 ml) was extracted with the same volume of n-hexane, and the n-hexane phase was applied to the chromatograph. The eluent used was a mixture of n-hexane and 2-propanol (85 : 15 (v/v)) at a flow rate of 0.5 ml/min. The absorbance at 220 nm was monitored.
The concentration of n-glucose was determined by the glucose oxidase/peroxidase method using Glucose C-II Test Wako (Wako Pure Chemical Industries).
RESULTS AND DISCUSSION Reduction of acetophenone and chloroacetophenones
We reported that the yeast P. tannophilus IF0 1007 could catalyze the oxidation of the @)-isomer of I-PA from its racemic mixture with high enantioselectivity, leaving the @)-isomer (9). This led us to consider the possibility that @)-l-PA could also be produced using the same yeast through the reverse reaction, namely, asymmetric reduction of AP, under appropriate condi- tions, as also described by Kometani et al. (13). The
0 2 ‘de [h]
6
FIG. 3. Time-course of changes in the conversion during batch reduction of acetophenone (AP) and its chloro-derivatives to the cor- responding I-phenylethanols (I-PAS) by immobilized P. tunnophilus. Symbols: 0, A, and 0, indicate reduction of o-, m-, and P-CLAP, respectively, and 0, indicates reduction of AP.
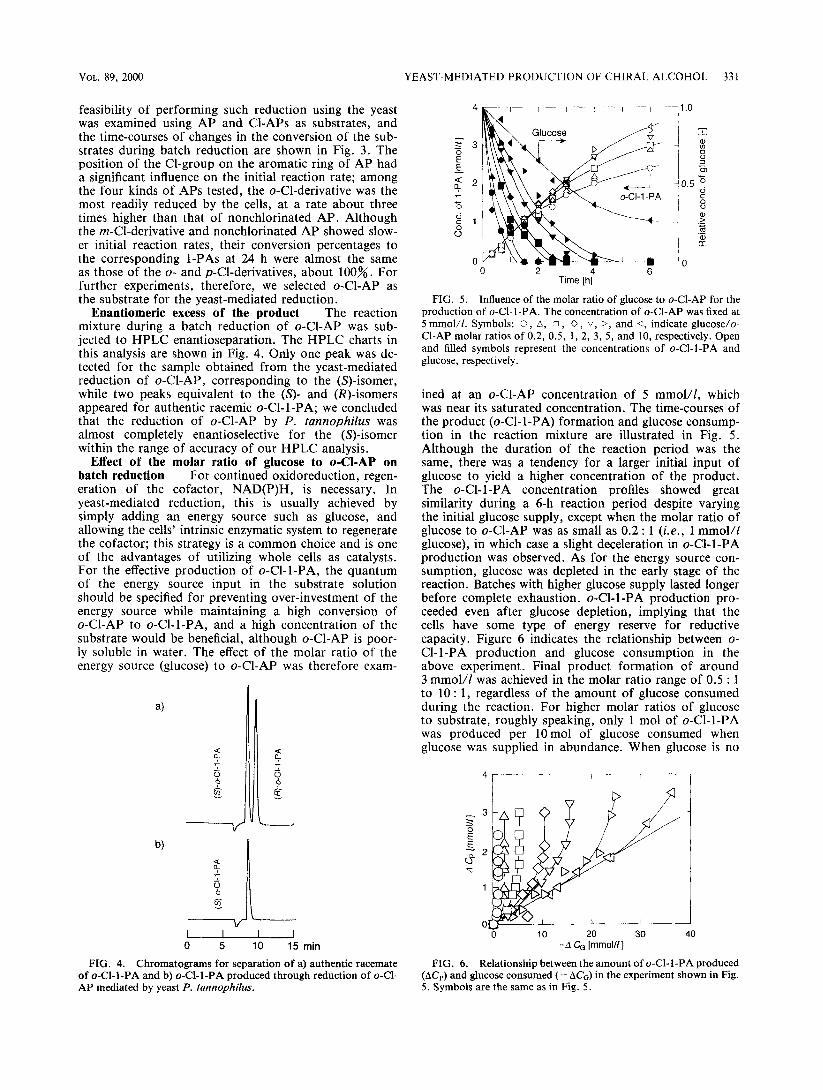
VOL. 89, 2000 YEAST-MEDIATED PRODUCTION OF CHIRAL ALCOHOL 331
feasibility of performing such reduction using the yeast was examined using AP and Cl-APs as substrates, and the time-courses of changes in the conversion of the sub- strates during batch reduction are shown in Fig. 3. The position of the Cl-group on the aromatic ring of AP had a significant influence on the initial reaction rate; among the four kinds of APs tested, the o-Cl-derivative was the most readily reduced by the cells, at a rate about three times higher than that of nonchlorinated AP. Although the m-Cl-derivative and nonchlorinated AP showed slow- er initial reaction rates, their conversion percentages to the corresponding ~-PAS at 24 h were almost the same as those of the o- and p-Cl-derivatives, about 100%. For further experiments, therefore, we selected o-Cl-AP as the substrate for the yeast-mediated reduction.
Enantiomeric excess of the product The reaction mixture during a batch reduction of o-Cl-AP was sub- jected to HPLC enantioseparation. The HPLC charts in this analysis are shown in Fig. 4. Only one peak was de- tected for the sample obtained from the yeast-mediated reduction of o-Cl-AP, corresponding to the @ ‘)-isomer, while two peaks equivalent to the (s)- and (@-isomers appeared for authentic racemic o-Cl-l-PA; we concluded that the reduction of o-Cl-AP by P. tannophilus was almost completely enantioselective for the @)-isomer within the range of accuracy of our HPLC analysis.
Effect of the molar ratio of glucose to o-GAP on batch reduction For continued oxidoreduction, regen- eration of the cofactor, NAD(P)H, is necessary. In yeast-mediated reduction, this is usually achieved by simply adding an energy source such as glucose, and allowing the cells’ intrinsic enzymatic system to regenerate the cofactor; this strategy is a common choice and is one of the advantages of utilizing whole cells as catalysts. For the effective production of o-Cl-l-PA, the quantum of the energy source input in the substrate solution should be specified for preventing over-investment of the energy source while maintaining a high conversion of o-Cl-AP to o-Cl-l-PA, and a high concentration of the substrate would be beneficial, although o-Cl-AP is poor- ly soluble in water. The effect of the molar ratio of the energy source (glucose) to o-Cl-AP was therefore exam-
b) 1
------I/- -
0 5 10 15 min
FIG. 4. Chromatograms for separat ion of a) authentic racemate of o-Cl-l-PA and b) o-Cl-l-PA produced through reduct ion of o-Cl- AP mediated by yeast P. tannophilus.
4
Time [h]
FIG. 5. Inf luence of the molar ratio of g lucose to o-Cl-AP for the product ion of o-Cl-l-PA. The concentrat ion of o-Cl-AP was fixed at 5 mmol/l. Symbols: 0, A, 0, 0, 0, b, and 4, indicate glucose/o- Cl-AP molar ratios of 0.2, 0.5, 1, 2, 3, 5, and 10, respectively. Open and filled symbols represent the concentrat ions of o-Cl-l-PA and glucose, respectively.
ined at an o-Cl-AP concentration of 5 mmol/l, which was near its saturated concentration. The time-courses of the product (o-Cl-l-PA) formation and glucose consump- tion in the reaction mixture are illustrated in Fig. 5. Although the duration of the reaction period was the same, there was a tendency for a larger initial input of glucose to yield a higher concentration of the product. The o-Cl-l-PA concentration profiles showed great similarity during a 6-h reaction period despite varying the initial glucose supply, except when the molar ratio of glucose to o-Cl-AP was as small as 0.2 : 1 (i.e., 1 mmol// glucose), in which case a slight deceleration in o-Cl-l-PA production was observed. As for the energy source con- sumption, glucose was depleted in the early stage of the reaction. Batches with higher glucose supply lasted longer before complete exhaustion. o-Cl-l-PA production pro- ceeded even after glucose depletion, implying that the cells have some type of energy reserve for reductive capacity. Figure 6 indicates the relationship between o- Cl-l-PA production and glucose consumption in the above experiment. Final product formation of around 3 mmol/l was achieved in the molar ratio range of 0.5 : 1 to 10 : 1, regardless of the amount of glucose consumed during the reaction. For higher molar ratios of glucose to substrate, roughly speaking, only 1 mol of o-Cl-l-PA was produced per 10mol of glucose consumed when glucose was supplied in abundance. When glucose is no
-TO
g 3
E” E 4 2
D
1
0 10 20 30 40
-A C, [mmol/l]
FIG. 6. Relationship between the amount of o-Cl-l-PA produced (AC,) and glucose consumed (-AC,) in the experiment shown in Fig. 5. Symbols are the same as in Fig. 5.
332 HASEGAWA ET AL. J. BIOSCI. BIOENG..
longer available, glucose uptake must inevitably stop, but the product formation in the reaction still continues, which is the reason for the vertical alignment of the sym- bols in the figure after glucose depletion. This result strongly indicates that the cells have the potential to con- vert o-Cl-AP to o-Cl-l-PA to some extent even in the absence of energy supply, once a stockpile of energy is formed within the cells.
We then examined whether NADH or NADPH was involved in the reaction by batch reduction of AP. The crude enzyme obtained from the disrupted P. tannophi- fus cells was added to a substrate solution containing NADH or NADPH as a reducing agent. After a l-h reac- tion period, although more product seemed to be form- ed in the presence of NADH as a cofactor than that form- ed in the presence of NADPH, l-PA was produced in the presence of either of the cofactors (data not shown). Thus, unfortunately, neither NADH nor NADPH could be specified as the more useful cofactor.
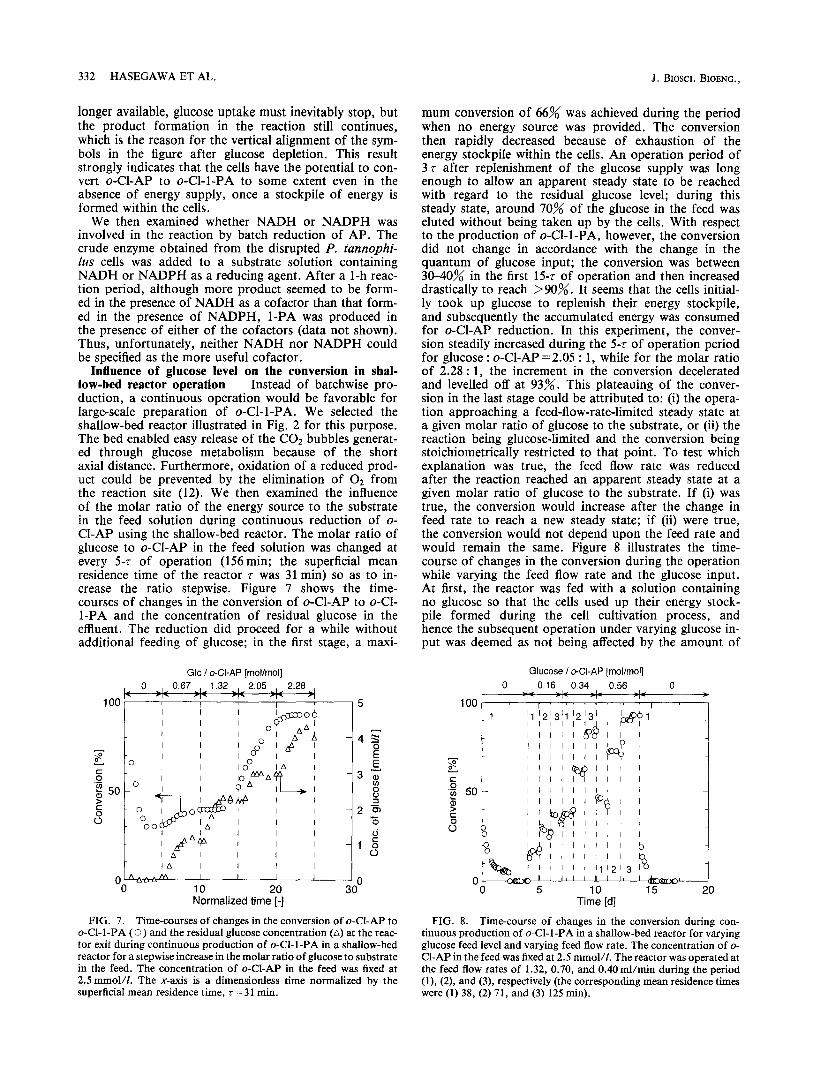
Influence of glucose level on the conversion in shal- low-bed reactor operation Instead of batchwise pro- duction, a continuous operation would be favorable for large-scale preparation of o-Cl-l-PA. We selected the shallow-bed reactor illustrated in Fig. 2 for this purpose. The bed enabled easy release of the COz bubbles generat- ed through glucose metabolism because of the short axial distance. Furthermore, oxidation of a reduced prod- uct could be prevented by the elimination of O2 from the reaction site (12). We then examined the influence of the molar ratio of the energy source to the substrate in the feed solution during continuous reduction of o- CLAP using the shallow-bed reactor. The molar ratio of glucose to o-CLAP in the feed solution was changed at every 5-r of operation (156 m in; the superficial mean residence time of the reactor r was 31 m in) so as to in- crease the ratio stepwise. Figure 7 shows the time- courses of changes in the conversion of o-Cl-AP to o-Cl- l-PA and the concentration of residual glucose in the effluent. The reduction did proceed for a while without additional feeding of glucose; in the first stage, a maxi-
Glc / o-CLAP [mol/mol] 0 , 0.67 1.32 , 2.05 , 2.28
I- I- I- I- I I 15
IA- I I I I
Normalized time [-]
FIG. 7. Time-courses of changes in the conversion of o-CLAP to o-Cl-I-PA (0) and the residual glucose concentration (a) at the reac- tor exit during continuous production of o-Cl-l-PA in a shallow-bed reactor for a stepwise increase in the molar ratio of glucose to substrate in the feed. The concentration of o-Cl-AP in the feed was fixed at 2.5mmoVl. The x-axis is a dimensionless time normalized by the superficial mean residence time, r = 3 1 min.
mum conversion of 66% was achieved during the period when no energy source was provided. The conversion then rapidly decreased because of exhaustion of the energy stockpile within the cells. An operation period of 3 r after replenishment of the glucose supply was long enough to allow an apparent steady state to be reached with regard to the residual glucose level; during this steady state, around 70% of the glucose in the feed was eluted without being taken up by the cells. With respect to the production of o-Cl-l-PA, however, the conversion did not change in accordance with the change in the quantum of glucose input; the conversion was between 30-40% in the first 15-r of operation and then increased drastically to reach >90%. It seems that the cells initial- ly took up glucose to replenish their energy stockpile, and subsequently the accumulated energy was consumed for o-CLAP reduction. In this experiment, the conver- sion steadily increased during the 5-r of operation period for glucose : o-Cl-AP=2.05 : 1, while for the molar ratio of 2.28 : 1, the increment in the conversion decelerated and levelled off at 93%. This plateauing of the conver- sion in the last stage could be attributed to: (i) the opera- tion approaching a feed-flow-rate-limited steady state at a given molar ratio of glucose to the substrate, or (ii) the reaction being glucose-limited and the conversion being stoichiometrically restricted to that point. To test which explanation was true, the feed flow rate was reduced after the reaction reached an apparent steady state at a given molar ratio of glucose to the substrate. If (i) was true, the conversion would increase after the change in feed rate to reach a new steady state; if (ii) were true, the conversion would not depend upon the feed rate and would remain the same. Figure 8 illustrates the time- course of changes in the conversion during the operation while varying the feed flow rate and the glucose input. At first, the reactor was fed with a solution containing no glucose so that the cells used up their energy stock- pile formed during the cell cultivation process, and hence the subsequent operation under varying glucose in- put was deemed as not being affected by the amount of
Glucose / CHZI-AP [mol/mol] 0 0.16 034 0.56 0
1OOr I- -I- -l- I- *
r “’ I ‘I I I
7 I I I I I I /
0
.s 50 I//IIIIl I I
ti z
s
I I I / I I I I I I I I I
0 0 5 10 15 20
Time [d]
FIG. 8. Time-course of changes in the conversion during con- tinuous production of o-Cl-l-PA in a shallow-bed reactor for varying glucose feed level and varying feed flow rate. The concentration of o- CLAP in the feed was fixed at 2.5 mmol/l. The reactor was operated at the feed flow rates of 1.32, 0.70, and 0.40 ml/min during the period (l), (2), and (3), respectively (the corresponding mean residence times were (1) 38, (2) 71, and (3) 125 min).

VOL. 89, 2000 YEAST-MEDIATED PRODUCTION OF CHIRAL ALCOHOL 333
initially-accumulated energy within the cells. For each glucose input, the conversion changed drastically in response to a decrease in the feed rate to a higher value, implying that the reactor operation was feed-flow-rate- limited rather than stoichiometrically-restricted. Glucose was not eluted from the reactor throughout the opera- tion and was considered to be completely consumed by the cells because of the low glucose input. At this time also, the glucose uptake by the cells showed a behavior out of harmony with the changes in the conversion. It is likely that the cells established an energy stockpile dur- ing the operation independent of the conversion of o-Cl- AP. After the manipulation of the glucose molar ratio and feed flow rate, a feed solution with no glucose was supplied on day 14. The conversion dropped to zero very rapidly within a day compared to the operation under the same conditions as at the beginning of the ex- periment, where zero-conversion was reached after 3-d operation. It could be deduced that the accumulation of energy during the reactor operation was not very significant because of complete glucose deprivation in the first stage and the subsequent energy-exhaustive o-Cl- AP reduction process, although the energy accumulated during cultivation for cell preparation was considerable.
The amount of cofactor NAD(P)H regenerated for a unit amount of glucose consumption was then examined in this experiment. The following were assumed: (i) the reaction rate is proportional to the concentrations of the substrate and glucose (second-order reaction) because of their relatively low concentrations, (ii) the effectiveness factor of the immobilized yeast cells is unity, (iii) the liquid flow in the reactor is the piston-flow type, (iv) glucose taken up by the cells is exclusively used as an energy source for the regeneration of the cofactor NAD(P)H which is involved in the bioconversion of o- Cl-AP to o-Cl-l-PA, and (v) 1 mol of the cofactor yields 1 mol of o-Cl-l-PA. Under these assumptions, the mass balance equations of the substrate and glucose in the steady state are given by Eqs. 2 and 3, respectively.
s= p(l ~&&zkCsCo u” dz (2)
a= -(l -&lJkCsCo ” dz (3)
where Cs is the substrate concentration, Co is the glu- cose concentration, ug is the superficial velocity of the liquid in the reactor, z is the axial distance, k is the rate
T (nRGs - I) X 1 W4 [s]
FIG. 9. Estimation of the amount of the cofactor (n mol) regenerated per mol of glucose consumed according to Eq. 4. The case for n=8. The symbols 0, A and 0 represent the molar ratios of glucose to o-Cl-AP, Rojs, of 0.16, 0.34 and 0.56, respectively.
constant, &b is the bed voidage, and n is the moles of the reduced cofactor regenerated through consumption of 1 mol glucose. Using the boundary conditions that Cs = Cso and Co= Co, at z=O, Eqs. 2 and 3 can be easily solved to give
In 1 -xs 1 --wwbs)
= - (1 - &,,)kd&,,(n&j,$ ~- 1) (4)
where xs is the conversion, Cs, is the concentration of o- Cl-AP in the feed solution, and RGIS is the molar ratio of glucose to o-Cl-AP in the feed solution. The left term of Eq. 4 is plotted against r(nRo,s-1) for various values of IZ, and the case for n=8 is illustrated in Fig. 9. Similar linearity was observed in the plots for values of II from 7 to 10, indicating that l/7 to l/10 of glu- cose against o-Cl-AP would be sufficient for cofactor NAD(P)H regeneration in this reaction system.
As shown here, a low molar ratio of glucose to sub- strate was sufficient for continuous production of o-cl-l- PA through asymmetric reduction of o-Cl-AP mediated by immobilized P. tannophilus cells in a shallow-bed reactor, which is a type of plug-flow reactor.
REFERENCES
1. Faber, K.: Biotransformations in organic chemistry, 2nd ed. Springer-Verlag, Berlin (1992).
2. Nakamura, K., Ushio, K., Oka, S., and Ohoo, A.: Stereochem- ical control in yeast reduction. Tetrahedron Lett.. 25. 3979- 3982 (1984). -
3. Carry, S. W. and Neilson, D. G.: The preparation and absolute configuration of the optically active forms of the diastereoiso- mers of 2-(1-phenylethyl)amino-1-phenylethanol and their use as chiral auxiliaries in the asymmetric reduction of acetophe- none with modified lithium aluminium hydrides. J. Chem. Sot. Perkin Trans. I, 1987, 601-605 (1987).
4. Mtiler, D., Umbricht, G., Web&, B:, and Pfaltz, A.: C2-Sym- metric 4,4’,5,5’-tetrahydrobi(oxazoles) and 4,4’,5,5’-tetrahydro- 2,2’-methylenebis[oxazoles] as chiral ligands for enantioselective catalysis. Helv. Chim. Acta, 74, 232-240 (1991).
5. MacLeod, R., Prosser, H., Fikentscher, L., Lanyi, J., and Mosher, H. S.: Asymmetric reductions. XII. Stereoselective ke- tone reductions by fermenting yeast. Biochemistry, 3, 838-846 (1964).
6. Sih, C. J. and Chen, C.-S.: Microbial asymmetric catalysis: enantioselective reduction of ketones. Angew. Chem. Int. Ed. Engl., 23, 570-$78 (1984).
7. Gillois, J., Buisson, D., Azerad, R., and Jaouen, G.: Enan- tioselective microbial reduction of transition-metal-complexed aromatic ketones. J. Chem. Sot., Chem. Commun., 1988, 1224-1225 (1988).
8. Young, C. S. and Ward, 0. P.: Studies of the reductive bio- transformation of selected carbonyl comuounds by whole cells and extracts of baker’s yeast, SQ&hQrOkIyCeS ceievisiae. Bio- technol. Bioeng., 38, 1280-1284 (1991).
9. Haseeawa, Y., Uosaki, H., Adachi, S., and Matsuno, R.: Prod<ction of (R)-1-phenylethanol through enantioselective oxi- dation of (S’)-isomer from their racemic mixture by pachysolen tannophil&IFO 1007. Biotechnol. Lett., 18, 367-372 (1996).
10. Roebuch. K.. Brundin. A.. and Johns. M.: Resnonse surface optimization ‘of temperature and pH for’the growth of Pachyso- len tQnno,ohihm. Enzyme Microb. Technol., 17, 75-78 (1995).
II. Slininger; P. J., Bole& P. L., and Kurtzman, C. P.: Pachysolen tannophi/us: properties and process considerations for ethanol production from n-xylose. Enzyme Microb. Technol., 9, 5-15 (1987).
12. Hasegawa, Y., Adachi, S., and Matsuno, R.: Asymmetric reduction of acetophenone by immobilized Hansenuia capsu- lata cells. J. Ferment. Bioeng., 85, 322-327 (1998).
13. Kometani, T., Yoshii, H., and Matsuno, R.: Large-scale production of chiral alcohols with bakers’ yeast. J. Mol. Catal. B: Enzymatic, 1, 45-52 (1996).


















