
Effect of saprophytic phylloplane fungi on germination and development of Melampsora larici-populina
7
Trans. Br. mycol, Soc. 72 (2) 225-231 (1979) [ 225 ] Printed in Great Britain EFFECT OF SAPROPHYTIC PHYLLOPLANE FUNGI ON GERMINATION AND DEVELOPMENT OF MELAMPSORA LARICI-POPULINA By MOHAMMED OMAR* AND W. A. HEATHER Department of Forestry, Australian National University, Canberra, A.C.T. 2600, Australia The common phylloplane fungi of poplar leaves, Cladosporium sp., Alternaria sp. and Penicillium sp. were shown to reduce germination and limit development of Melampsora larici-populina Kleb. to varying degrees. Alternaria and Cladosporium species were hyper- parasitic, penetrating and causing lysis of the urediniospores. The significance of this study in biological control is discussed. The leaf surface is a good site for both sapro- phytic and parasitic fungi. Among the phylloplane fungi, a few have been reported as hyperparasites (McKenzie & Hudson, 1976). Of these, Clado- sporium sp. have been reported parasitizing various Uredinales (Smith, 1905; Keener, 1954; Bolland, 1973; McKenzie & Hudson, 1976; Heather & Sharma, 1977; Sharma & Heather, 1978). In Australia, in late summer, uredinial pustules of both leaf rusts of poplar, Melampsora larici- populina Kleb. and M. medusae Thurn. are ex- tensively colonized by hyperparasitic fungi, particularly species of Darluca, Cladosporium, Penicillium, Alternaria and Fusarium (Bolland, 1973; Sharma & Heather, 1978; Omar, 1978). This study reports the effects of three commonly occurring poplar phylloplane fungi, namely Alter- naria, Penicillium and Cladosporium species, on the germination of urediniospores of M. larici-populina Kleb. and on the development of rust on leaf disks of poplar clone Populus x euramericana cv. '1-48 8'. MA TERIALS AND METHODS Urediniospores were collected from field samples of infected poplar leaves and plated on potato dextrose agar (PDA). Colonies of contaminating phylloplane fungi developed and these were sub- cultured on PDA to obtain pure cultures of Alternaria sp., Cladosporium sp. and Penicillium sp. Conidia were brushed from these pure cultures and dried at 25°C for use in the following experiments. Treatment A - saprophyte then parasite Five replicates each of 13 mm diam coverslips and surface sterilized, 13 mm diam leaf disks of the poplar clone '1-488', were separately inoculated * Present address: Microbiology Unit, Science Faculty, National University of Malaysia, Bangi, Selangor, Malaysia. with conidia of each saprophyte in a spore settling tower (Omar, 1978). Leaf disks were inoculated on their abaxial surface. The coverslips and leaf disks were then placed in separate humidity chambers (Fig. 1) at 100 % r.h, and sealed with greased 20 x 20 mm cover- slips. Water was used to provide the required humidity. The chambers were incubated in the dark at 16 ± 1°. After 60 min coverslips and leaf disks were reinoculated in the settling tower with 20 mg of urediniospores of M. larici-populina. The chambers were resealed and incubated for a further 5 h. Treatment B - parasite then saprophyte The procedure was similar except that coverslips and leaf disks were first inoculated with uredinio- spores and 60 min later with conidia of each saprophyte. Total incubation period of the urediniospores was 5 h. Treatment C- parasite and saprophyte simultaneously Procedure was as described in Treatment A except that coverslips and leaf disks were inoculated with separate mixtures of urediniospores and conidia of each saprophyte; these were incubated without interruption for 5 h. Coverslips and leaf disks, inoculated with urediniospores only, and incubated under the same conditions for 5 h, were included as controls. At the end of the incubation period, coverslips were inverted over a drop of polyvinyl alcohol cotton blue stain (PVACB) (Omar, 1978) and the percentage germination assessed using a Zeiss W.L. Research Microscope. The criterion of germination was as described previously (Manners, 1966; Omar & Heather, 1975). Since the PVACB sets in a few hours to give a permanent mount, specimens need not be assessed immediately. The 0007-1536179/2828-4790 $01.00 © 1979 The British Mycological Society
-
Upload
mohammed-omar -
Category
Documents
-
view
214 -
download
0
Transcript of Effect of saprophytic phylloplane fungi on germination and development of Melampsora larici-populina

Trans. Br. mycol, Soc. 72 (2) 225-231 (1979)
[ 225 ]
Printed in Great Britain
EFFECT OF SAPROPHYTIC PHYLLOPLANE FUNGI ONGERMINATION AND DEVELOPMENT OF MELAMPSORA
LARICI-POPULINA
By MOHAMMED OMAR* AND W. A. HEATHER
Department of Forestry, Australian National University, Canberra, A.C.T. 2600, Australia
The common phylloplane fungi of poplar leaves, Cladosporium sp., Alternaria sp. andPenicillium sp. were shown to reduce germination and limit development of Melampsoralarici-populina Kleb. to varying degrees. Alternaria and Cladosporium species were hyper-parasitic, penetrating and causing lysis of the urediniospores. The significance of thisstudy in biological control is discussed.
The leaf surface is a good site for both sapro-phytic and parasitic fungi. Among the phylloplanefungi, a few have been reported as hyperparasites(McKenzie & Hudson, 1976). Of these, Clado-sporium sp. have been reported parasitizing variousUredinales (Smith, 1905; Keener, 1954; Bolland,1973; McKenzie & Hudson, 1976; Heather &Sharma, 1977; Sharma & Heather, 1978). InAustralia, in late summer, uredinial pustules ofboth leaf rusts of poplar, Melampsora larici-populina Kleb. and M. medusae Thurn. are ex-tensively colonized by hyperparasitic fungi,particularly species of Darluca, Cladosporium,Penicillium, Alternaria and Fusarium (Bolland,1973; Sharma & Heather, 1978; Omar, 1978).
This study reports the effects of three commonlyoccurring poplar phylloplane fungi, namely Alter-naria, Penicillium and Cladosporium species, on thegermination of urediniospores of M. larici-populinaKleb. and on the development of rust on leafdisks of poplar clone Populus x euramericana cv.'1-488'.
MA TERIALS AND METHODS
Urediniospores were collected from field samplesof infected poplar leaves and plated on potatodextrose agar (PDA). Colonies of contaminatingphylloplane fungi developed and these were sub-cultured on PDA to obtain pure cultures ofAlternaria sp., Cladosporium sp. and Penicillium sp.Conidia were brushed from these pure cultures anddried at 25°C for use in the following experiments.
Treatment A - saprophyte then parasite
Five replicates each of 13 mm diam coverslips andsurface sterilized, 13 mm diam leaf disks of thepoplar clone '1-488', were separately inoculated
* Present address: Microbiology Unit, ScienceFaculty, National University of Malaysia, Bangi,Selangor, Malaysia.
with conidia of each saprophyte in a spore settlingtower (Omar, 1978). Leaf disks were inoculated ontheir abaxial surface.
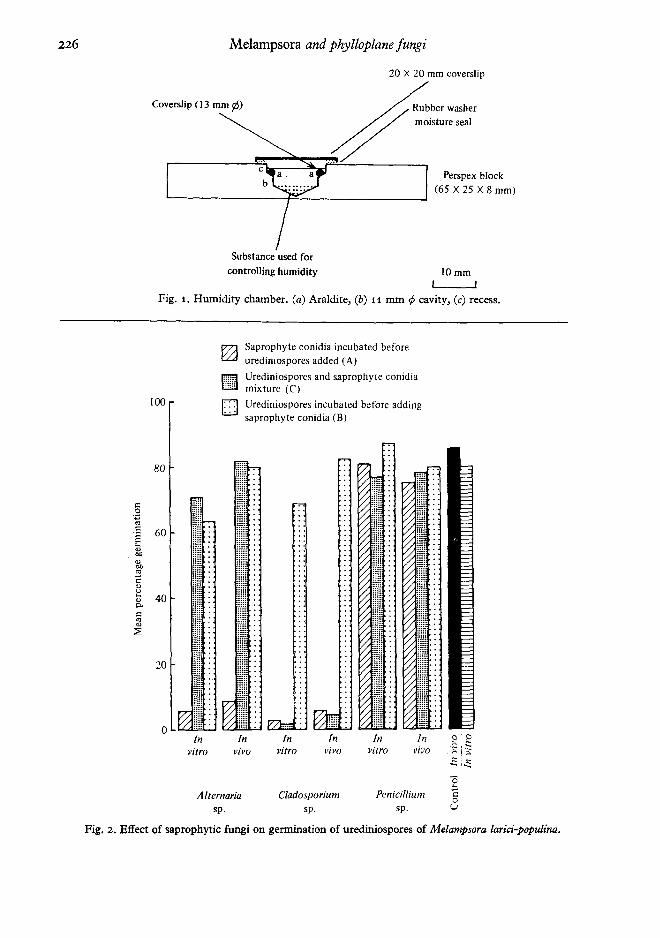
The coverslips and leaf disks were then placedin separate humidity chambers (Fig. 1) at 100 %r.h, and sealed with greased 20 x 20 mm cover-slips. Water was used to provide the requiredhumidity. The chambers were incubated in thedark at 16 ± 1°. After 60 min coverslips and leafdisks were reinoculated in the settling tower with20 mg of urediniospores of M. larici-populina. Thechambers were resealed and incubated for a further5 h.
Treatment B - parasite then saprophyte
The procedure was similar except that coverslipsand leaf disks were first inoculated with uredinio-spores and 60 min later with conidia of eachsaprophyte. Total incubation period of theurediniospores was 5 h.
Treatment C - parasite and saprophytesimultaneously
Procedure was as described in Treatment A exceptthat coverslips and leaf disks were inoculated withseparate mixtures of urediniospores and conidia ofeach saprophyte; these were incubated withoutinterruption for 5 h. Coverslips and leaf disks,inoculated with urediniospores only, and incubatedunder the same conditions for 5 h, were includedas controls.
At the end of the incubation period, coverslipswere inverted over a drop of polyvinyl alcoholcotton blue stain (PVACB) (Omar, 1978) and thepercentage germination assessed using a ZeissW.L. Research Microscope. The criterion ofgermination was as described previously (Manners,1966; Omar & Heather, 1975). Since the PVACBsets in a few hours to give a permanent mount,specimens need not be assessed immediately. The
0007-1536179/2828-4790 $01.00 © 1979The British Mycological Society
226 Melampsora and phylloplanefungi
20 X 20 mm co verslip
Coverslip (\3 mm ¢)
c
Rubber washer
...~ moisture ~I
. Perspe x block
(65 X 25 X 8 mrn )
Substance used for
controlling humidity 10 mmI I
Fig. 1. Humidity chamber. (a) Araldite, (b) 11 mm </J cavity, (c) recess.
100
~
I]jillj]no
Saprophyt e conidia incub at ed beforeured iniospores added ( A )
Urediniospores and sapro phy te conidiamixture (e)
Uredin iospores incubated before add ingsaprophy te conidia (B )
80
60
40
20
Invitro
Invivo
Invitro
Invivo
Invitro
Invivo
Alternariasp .
Cladosporiumsp.
Penicilliumsp .
'2co
l.J
Fig. 2. Effect of saprophytic fungi on germination of urediniospores of Melampsora larici-populina .

M. Omar and W. A. Heather 227
germination on leaf disks was assessed immediatelyusing a Leitz Incident Light Microscope.
A few coverslips and leaf disks were quenched inliquid nitrogen and placed in a Dynavac TissueFreeze Drier. Specimens were then coated with a20-30 nm layer of gold and examined using aCambridge Stereoscan 180 scanning electronmicroscope at 30 kV accelerating voltage.
Following assessment of germination, five leafdisks of each treatment were floated on 5 mg/]gibberellic acid solution. They were incubated in acontrolled environment chamber (16 ± 1°, 16 hphoto-period at 5000 lux). The numbers ofuredinialpustules which developed on the disks wereassessed after 15 days incubation. A few leaf diskswere also treated as described previously forexamination under the scanning electron micro-scope.
RESUL TS AND DISCUSSION
The results of the germination experiments areillustrated in Fig. 2. The germination of uredinio-spores was significantly reduced (P = 0'001) whenthese were deposited on coverslips or leaf disks onwhich conidia of the saprophytes had previouslybeen deposited and incubated for 1 h. The pre-sence of conidia of Alternaria sp. and Cladosporiumsp. caused large reductions in urediniosporegermination while Penicillium sp. had only mar-ginal effects (P = 0'05) under these conditions.The saprophyte conidia had not germinated duringthe initial 60 min incubation. In contrast theurediniospores in Treatment B had germinatedduring this time and therefore conidia of Clado-sporium sp. and Alternaria sp. depressed uredinio-spore germination only marginally (P = 0'05) inthis treatment. However, conidia of Penicilliumdidnot affect urediniospore germination in TreatmentB. Cladosporium sp, also reduced urediniosporegermination (P = 0'001) when its conidia weredeposited and incubated together (Treatment C),while Alternaria sp. and Penicillium sp. had nosignificant effect. Overall, presence of conidia ofPenicillium sp. had minimal effects on the germina-tion of urediniospores. Significant reduction ingermination of urediniospores of Melampsoraoccidentalis Jacks. in contact with suspensions ofleaf saprophytes has also been reported (Bier,1964).
Alternaria and Cladosporium species developedappressoria which parasitised urediniospores (Fig.4,6-8). Elsewhere Locci et al. (1971) have demon-strated close adherence of hyphae of Verticilliumhemileiae Bour. to;.hrediniospores of Hemileiauastatrix Berk. & Br.
Lysis of urediniospores in contact with Clado-sporium mycelium was commonly noticed (Fig. 3,
4, 7, 8) which agrees with the earlier observationsof Heather & Sharma (1977) and Sharma &Heather (1978). Lysis was not detected in thePenicillium-urediniospore association.
Parasitisms of fungi have been classified intotwo broad divisions (Hashioka, 1973). These arebiotrophic which includes haustorial, non hausto-rial and indirect reactions, and necrotrophic. Innecrotrophic or destructive mycoparasitism, theparasite destroys the host after or even beforeinvasion and absorbs nutrients from the disor-ganized or dead host cells. It appears from thisstudy that Alternaria sp, is abiotrophic haustorialparasite and that Cladosporium sp. is both haus-torial and necrotrophic causing lysis of host cellsafter contact. Possibly, exudates from the conidiaof Cladosporium sp. caused lysis of urediniospores.
Conidia of the three saprophytes, irrespective ofmethod of inoculation (Treatment A, B or C),caused significant reduction (P = 0'001) in num-bers of uredinia per em" developed on leaf disks(Fig. 11). The reduction was particularly pro-nounced with Alternaria and Cladosporium species.Scanning electron microscope studies showed thecolonization of uredinia by the mycelium of thelatter (Fig. 9) and also that the fungus penetratesthe stomata (Fig. 10). While species of Clado-sporium (Agnihothrudu, 1974; Sutton, 1970) andAlternaria (Chaturvedi, 1972) have been reportedto cause diseases of poplars, the relationship of thespecies used in these experiments to those patho-genic on poplar leaves is unknown.
The results confirm earlier reports indicatinginhibition of germination of spores of pathogenicfungi and consequent reduction in the infectionlevel by saprophytic leaf colonizers. Cladosporiumherbarum (Pers.) Lk ex S. F. Gray and Alternariaalternata (Fr.) Keissl, have been shown to inhibitgermination of Botrytis cinerea Pers. ex Fr.conidia and once established, prevented thecolonization of the leaves by B. cinerea (Newhook,1957). Aureobasidium pullulans (de Bary) Arnaud,commonly isolated from bean leaves, reduced theinfection of beans by Alternaria zinniae Pape (vanden Heuvel, 1970). Fokkema & Lorbeer (1974)found that the growth of germ-tubes of Alternariaporri (E 11) Neergaard on onion leaves was re-duced by the presence of Aureobasidium pullulansand Sporobolomyces roseus Kluyver &van Niel. Theantagonists reduced the number of lesions by c.50 %.
While reduction of germination of uredinio-spores induced by saprophytes is a possible explana-tion for the decreased rust incidence on inoculatedleaf disks, in these experiments other mechanismsare also possible. Removal of nutrients from thephylloplane by the saprophytes could constitute an

22'3
m
3
30"m 8.!~1II30I'ID~l
Melampsora and phylloplane fungi
IOO"m •L-..-JJ,VV,....J.H

M. Omar and W. A. Heather 229
antagonism similar to the mechanism of soilfungistasis formulated by Ko & Lockwood (1968,1970). However, this explanation is unlikely withM. larici-populina which enters the leaf eitherdirectly through the epidermis or the stomata andusually does not require exogenous substrates forgermination (Omar, 1978).
Inhibition of germination of urediniospores
could possibly be caused by phytoalexins orinduced inhibitors formed by the leaves understimulation of the saprophytes. This theory hasbeen cited to explain reduced incidence of Alter-naria zinniae on bean leaves in the presence of asaprophyte, Aureobasidium pullulans (van denHeuvel, 1970).
Cladosporium spp. have been reported to grow
sp.PenicilliumCladosporium
sp.sp.
Saprophyte conidia incubated beforeurediniospores added (A)
Urediniospores and saprophyte conidiamixture (e)
Urediniospores incubated before addingsaprophyte conidia (8)
Alternaria
~
~
D··...
o
10
70 r
6J~
"2cov
Fig. 11. Effect of saprophytic fungi on development of uredinial pustules of Melampsora larici-populina.
Fig. 3. Light micrograph of lysed urediniospores (ud) and associated hypha of Cladosporium after 5 hincubation.
Fig. 4. Scanning electron micrograph showing the relationship between Cladosporium mycelium andurediniospores. Urediniospores in contact with the Cladosporium mycelium were lysed (ud).
Fig. 5. SEM of a conidium of Cladosporium sp. Note the warty appearance of the wall.
Fig. 6. SEM showing the effect of Alternaria sp. Note the presence of lysed urediniospores (ud) andappressoria (h) in contact with saprophyte. Many urediniospores fail to germinate.
Fig. 7. SEM showing the close association of Cladosporium mycelium with urediniospore. The penetra-tion of the urediniospore probably occurred at the appressorium (h).
Fig. 8. SEM showing production of conidia (c) by the hyperparasite Cladosporium sp.
Fig. 9. SEM of uredinium (ur) on the surface of a leaf disk showing the presence of the fungus.
Fig. 10. SEM of stomata (s):showing apparent penetration by mycelium of Cladosporium sp. c, Conidium;gt, germ-tube; h, appressorium of saprophytic fungus; m, mycelium of saprophytic fungus ; s, stromata;11, urediniospore; ud, lysed urediniospore; ui, intact urediniospore; ur, uredinium.

Melampsora and phylloplane fungi
faster on leaves in the presence of urediniosporesof M. larici-populina (Sharma & Heather, 1978).This observation agrees with that for Ramulariauredinis (Voss.) Sacco and Melampsora allii-populinaKleb, (Magnani, 1970). More propagules ofsaprophytes were shown to occur on rust-infectedthan on rust-free leaves of poplar (McKenzie &Hudson, 1976). They suggested that the rust mayhave provided nutrients directly for the sapro-phytes through decay and liberation of nutrientsfrom the rust hyphae. The total carbon content ofrust-infected material is often higher than non-infected material as biotrophic parasites frequentlycaused decreased export from, and increasedimport to, infected parts (L ewis, 1974). Relativesuccess or failure of some saprophytes on rust-infected material may be related to their ability toutilize fungal carbohydrates.
It has been suggested that hyperparasites playalimited role in reducing epidemics of rust in plants(Darpaux, 1960; Bier, 1964; Bean, 1968; Sharma &Heather, 1978). This study confirrns previousobservations that simultaneous inoculation ofleaves with conidia of Cladosporium and uredinio-spores of M. larici-populina reduced the infectionlevel of the rust and the number of urediniosporesper uredinia (Sharma & Heather, 1978).
The significance of hyperparasites of poplar rustfungi in biological control of rust has been can-vassed (Magnani, 1967; Morelet & Pinon, 1973;Heather & Sharma, 1977). In an envirorunent-con-scious society control of pathogens by the use ofhyperparasites, rather than with chemical sprays,is an attractive proposal. A strong case for biolo-gical control has been advanced by Baker & Cook(1974). With a forest crop, characterized by anextensive form of culture, a long rotation and loweconomic return, vis-a-vis certain food crops,biological control is an even more attractive pro-position. However, the field implementation ofsuch a proposal would involve long term experi-mental investigations of pathogen/hyperparasiteinteractions (possibly including race specificity), ofmethods of hyperparasite introduction and condi-tions favouring its multiplication and of the econo-mics of the whole procedure. It is unlikely to be aviable proposition in poplar rust for a considerableperiod.
REFEREN CES
AGNIHOTHRUDU, V. (1974). A leaf disease of poplarfrom Kashmir. Current Science 45, 389-390 •
BAKER, K . F . & COOK, R. J. (1974). Biological Controlof Plant Pathogens. San Francisco : W. H. Freeman.
BEAN, G. A. (1968). Growth of the hyperparasiteDarluca filum on chemically defined medium. Phyto-pathology 58, 252-253.
BIER, J. E. (1964). The possibility of microbiologicaltypes with different degrees of disease resistance with
a tree species on clone. In Breeding Pest ResistantTrees (ed , H. D. Gerhold, E. J. Schreiner, R. E.McDermott and J. A. Winieski), pp. 257-270.Toronto: Pergamon Press .
BOLLAND, L. (1973). Poplar rust in Queensland .A ustralian Plant Pathology Society N ewsletter 2, 28.
CHATURVEDI, C. (1972). A new leaf spot of poplarincited by Alternaria alternata (Fr. ) Keissler. IndianPhytopathology 25, 316-318.
DARPAUX, H. (1960). Biological interferences in theepidemics. In Plant Pathology : An Advan ced Treat ise,vol. III (ed . ]. G. Horsfall and A. E. Dimond), pp ,521-566. New York : Academic Press.
FOKKEMA, N. J. & LORBEER, J. W. (1974) . Interactionsbetween Alternaria porri and the saprophytic micro-flora of onion leaves. Phytopathology 64, 1128-1133.
HASHIOKA, Y. (1973). Mycoparasiti sm in relation tophytopathogens, Shokubutsu Byogai Kenkyu 8, 179-190.
HEATHER, W. A. & SHARMA, J. K. (1977). Some aspectsof poplar rust research in Australia. AustralianForestry 40, 28-43 .
KEENER, P. D. (1954). Cladosporium aecidiicola. Thuem.and Tuberculina persicina (D itm .) Sacco associatedwith Puccinia conspicua (Arth .) Mains on Heliniumhoopesii A. Gray in Arizona . Plant Disease Reporter38, 690-694.
Ko, W. H. & LOCKWOOD, J. L. (1968). Soil fungistasis :relation to fungal spore nutrition. Phytopathology 57,894-901.
Ko, W. H. & LOCKWOOD, J. L. (1970) . Mechanism oflysis of fungal mycelia in soil. Phy topa thology 60,148-154.
LEWIS, O. H. (1974). Microorganisms and plants. Theevolution of parasitism and mutualism. Symposiumof the Society of General Microbiology 24, 367-392.
LOCCI, R., FERRANTE, G. M. & RODRIGUES, C. J. (1971).Studies by transmission and scanning electronmicroscopy on the Hem ileia oastatrix- V erticilliumhemileiae association. R itnsta di Pato logia Vegetale 7,127-140.
MAGNANI, G. (1967). A hyperparasite of poplar rust.Cellulosa e Carta 18, 37-39.
MAGNANI, G. (1970). A poplar rust hyperparasite.Publicasioni del Centro di Sperimenta zioni agricola efor estale 11, 27-35.
MANNERS, J. G. (1966). Assessment of germination.In The Fungus Spore (ed. M. F. Madelin), pp. 165-174. London: Butterworth.
McKENZIE, E. H. C. & HUDSON, H. J. (1976). Myco-flora of rust-infected and non-infected plant materialduring decay. Tran sactions of the Brit ish MycologicalSociety 66, 223-238.
MORELET, M. & PINON, J. (1973). Darlucafilum hyper-paras ite of the genus Melampsora on poplar andwillow. Revue [orestiere francoise 25, 378-379.
NEWHOOK, F. J. (1957). The relationship of saprophyticantagonism to control of Botrytis cinerea Pers. ontomatoes. New Zealand fournal of Science and Tech-nology A 38, 473-481.
OMAR, M. B. (1978). Pre- and post-penetration phe-nomena in Melampsora larici-populina Kleb. Ph.D.Thesis, Australian National University.

M. Omar and W. A. Heather
OMAR, M. B. & HEATHER, W. A. (1975). Effects oftemperature and humidity on the in vitro germina-tion of uredospores of Melampsora larici-populina.Proceedings of the XVII Session, F.A.O.jI .P.C.Working Group on Diseases, Novisad . 28 Septemberto 4 October 1975·
SHARMA,]. K. & HEATHER, W. A. (1978). Paras itism ofuredospores of Melampsora larici-populina Kleb. byCladosporium sp, European'[ournal of Forest Pathology8,48-54.
SMITH, R. E. (1905). Asparagus and asparagus rust in
California. California Agricultural Experiment Sta-tion Bulletin 165, 1-99.
SUTTON, B. C. (1970). Forest Microfungi. IV. A leafspot of Populus caused by Cladosporium subsessile.Canadian Journal of Botany 48.471-477.
VAN DEN HEUVEL, J. (1970). Antagonistic effects ofepiphytic micro-organisms on infection of dwarfbean leaves by Alternaria zinniae, Phytopatholog ischLaboratorium ' Willie Commelin S cholten', Baarn,Mededeling 84, 1-84.
(Accepted for publication 14 July 1978)














