
Effect of Lactobacillus brevis-based bioingredient and bran on microbiological, physico-chemical and...
30
Effect of Lactobacillus brevis - based bioingredient and bran on microbio- logical, physico-chemical and textural quality of yeast leavened bread during storage Valerio Francesca, Di Biase Mariaelena, Caputo Leonardo, Creanza Teresa M., Ancona Nicola, Visconti Angelo, Lavermicocca Paola PII: S1466-8564(13)00141-0 DOI: doi: 10.1016/j.ifset.2013.09.003 Reference: INNFOO 1059 To appear in: Innovative Food Science and Emerging Technologies Received date: 30 April 2013 Accepted date: 11 September 2013 Please cite this article as: Francesca, V., Mariaelena, D.B., Leonardo, C., M., C.T., Nicola, A., Angelo, V. & Paola, L., Effect of Lactobacillus brevis - based bioingredi- ent and bran on microbiological, physico-chemical and textural quality of yeast leav- ened bread during storage, Innovative Food Science and Emerging Technologies (2013), doi: 10.1016/j.ifset.2013.09.003 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Transcript of Effect of Lactobacillus brevis-based bioingredient and bran on microbiological, physico-chemical and...

�������� ����� ��
Effect of Lactobacillus brevis - based bioingredient and bran on microbio-logical, physico-chemical and textural quality of yeast leavened bread duringstorage
Valerio Francesca, Di Biase Mariaelena, Caputo Leonardo, Creanza TeresaM., Ancona Nicola, Visconti Angelo, Lavermicocca Paola
PII: S1466-8564(13)00141-0DOI: doi: 10.1016/j.ifset.2013.09.003Reference: INNFOO 1059
To appear in: Innovative Food Science and Emerging Technologies
Received date: 30 April 2013Accepted date: 11 September 2013
Please cite this article as: Francesca, V., Mariaelena, D.B., Leonardo, C., M., C.T.,Nicola, A., Angelo, V. & Paola, L., Effect of Lactobacillus brevis - based bioingredi-ent and bran on microbiological, physico-chemical and textural quality of yeast leav-ened bread during storage, Innovative Food Science and Emerging Technologies (2013), doi:10.1016/j.ifset.2013.09.003
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
1
Effect of Lactobacillus brevis - based bioingredient and bran on microbiological, physico-
chemical and textural quality of yeast leavened bread during storage
Valerio Francescaa, Di Biase Mariaelena
a, Caputo Leonardo
a, Creanza Teresa M.
b, Ancona Nicola
b,
Visconti Angeloa and Lavermicocca Paola
a*,
a Institute of Sciences of Food Production, National Research Council, Via Amendola 122/O, 70126
– Bari
b Institute of Intelligent Systems for Automation, National Research Council, Via Amendola 122/D-
I - 70126 – Bari
*Corresponding author:
E-mail: [email protected]
Phone: +39 080 5929356; Fax : +39 080 5929374
Mailing address: Istituto di Scienze delle Produzioni Alimentari – CNR, Via Amendola
122/O, 70126 – Bari, Italy

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
2
Abstract
The effects of wheat bran and of a Lactobacillus brevis based-bioingredient (LbBio), obtained after
growth in flour-based medium, on quality of yeast leavened wheat bread (WWB) were investigated.
Bran was used in bread formulation by substituting a part (20 g/100 g) of white wheat flour (WBB),
while LbBio was used instead of the water content (WWB+LbBio and WBB+LbBio). The use of
LbBio in WWB resulted in the biological acidification of the dough due to lactic, phenyllactic and
OH-phenyllactic acid contents determining a high fermentation quotient value and an improved
bread texture and microbiological quality. Conversely, wheat bran reduced the specific volume and
crumb hardness during storage at 25 °C, and affected the antibacterial ability of LbBio during 30°C
storage. Our findings demonstrated that LbBio counteracted the negative effects of bran and
allowed to obtain an enriched fibre bread with specific volume and soft crumb comparable to bread
without bran.
Industrial relevance
Bread is a perishable food with a short microbiological and physico-chemical shelf-life. The main
microbiological alteration occurring into few days after baking is the “rope spoilage” caused by
spore-forming bacteria originating from raw materials. This phenomenon, often misinterpreted as a
sign of unsuccessful dough leavening and not visible from outside, is more common under
industrial production conditions during the hot season causing large economic losses in the warm
climates of Mediterranean countries, Africa and Australia. The use of sourdough often controls this
alteration even if the industrial application of this traditional process is limited by the long
leavening times. In this study an innovative procedure for the preparation of yeast-leavened bread
comprising the addition of a fermentation product from Lactobacillus brevis grown in a flour-
based medium, has been applied. The resulting fementation product (LbBio bioingredient) acts as a

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
3
sourdough acidifying the dough and improving the textural, physico-chemical and microbiological
properties of the resulting bread. The application of bioingredient LbBio could represent an
innovative strategy in industrial bread production to obtain acidified yeast leavened products
thus preventing the ropy spoilage and reducing the negative effects of bran addition.
Keywords: ropy spoilage, firmness, spore-forming bacteria, organic acids, bioingredient; two-ways
ANOVA.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
4
1. Introduction
Bread is one of the principal components of the human diet, but it generally undergoes staling
process and microbial contamination within few days from its production. In particular, after baking
bread can be spoiled by moulds (mainly Penicillium and Aspergillus species) and heat resistant
spore-forming bacteria, naturally occurring in raw materials and foods of vegetable origin, and
surviving to the cooking process (Rosenkvist & Hansen, 1995). A recurrent microbiological issue
for bakery industries is represented by the ropy spoilage mainly associated to the presence of spores
of Bacillus species in raw materials (Pepe, Blaiotta, Moschetti, Greco & Villani, 2003; Valerio, De
Bellis, Di Biase, Lonigro, Giussani, Visconti, Lavermicocca, & Sisto, 2012). The spores of these
microorganisms survive in the central part of baked bread, where the temperature values reach up to
97-101°C for some minutes. Even though the spore-forming loads are very low in flour (about 2.0
log spores/g), the baking process and the subsequent storage conditions of bread (temperature ≥ 25
°C, water activity≥0.95, pH>5) favour the germination of heat-resistant spores and their increase in
total viable counts up to about 7.0 log CFU/g in bread crumb within two days, causing the loss of
freshness and worsening of the bread quality (Rosenkvist & Hansen, 1995; Viedma, Abriouel,
Omar, López, Gálvez, 2011). As recently demonstrated (Valerio et al., 2012), flour and other raw
materials (brewer yeast, improvers, etc) used to make bread are contaminated by a great variety of
spore-forming bacteria mainly belonging to the genus Bacillus and which include also potential
toxigenic species (Bacillus cereus group). Recently, some authors (De Jonghe, Coorevits, De Block,
Van Coillie, Grijspeerdt, Herman, De Vos & Heyndrickx, 2010) demonstrated the ability of
Bacillus amyloliquefaciens species, a common bread contaminant, to produce heat-labile cytotoxic
substances and a heat-stable cytotoxic component. Generally, microbial contamination levels of
bread higher than 5 log CFU/g are associated to the onset of spoilage process and the elevated risk
of foodborne illness when the causative agent is a toxigenic species (Kramer & Gilbert, 1989).
Spore-forming bacteria are naturally occurring in soil and contaminate wheat and other cereal grain
flours. In particular, bran, arising from the outer part of the grain kernels, contains a higher content

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
5
of different microorganisms than those found in endosperm flours; among these microbes the spore-
forming bacteria could be related to the bread spoilage over its storage (Rosenkvist & Hansen,
1995).
In certain cases, bran has been used to replace part of the formulated flours in order to increase the
dietary fibres (DF) content in the bakery products, since wheat bran typically contains
approximately 45% of dietary fibre, of which about 95% is non-soluble fibre (Pomeranz, 1988;
Cornell & Hoveling, 1998). Nevertheless, health claims related to the ability of wheat bran fibres to
increase faecal bulk and to reduce intestinal transit time were accepted (EFSA, 2010) for labelling
food high in fibres in agreement to the European Regulation (EC) No 1924/2006. Besides DF, other
compounds concentrated in the outer layers of the grains, such as oligosaccharides and
phytochemicals, are gaining more and more importance in improving the nutritional and functional
quality of bread (Chavan & Chavan, 2011).
Furthermore, wheat bran-based flour blend can affect the rheological properties of bread dough
(Katina, Salmenkallio-Marttila, Partanen, Forssell & Autio, 2006a) and final bread quality attributes
that are consistent with reduction in volume, increasing in crumb firmness, and changing in flavour
(Laurikainen, Härkönen, Autio & Poutanen, 1998; Chavan & Chavan , 2011). In bread-making
these drawbacks are usually overcome by adding commercial enzyme mixtures or fermented wheat
bran to the dough during bread-making (Laurikainen et al., 1998; Katina, Laitila, Juvonen,
Liukkonen, Kariluoto, Piironen, Landberg, Åman, & Poutanen, 2007; Damen, Pollet, Dornez,
Broekaert, Van Haesendonck, Trogh, Arnaut, Delcour, Courtin, 2012). Well studied is also the
application of sourdough that, owing to fermentation by Lactobacillus sanfranciscensis,
Lactobacillus brevis and Lactobacillus plantarum, efficiently affects bread crumb properties and
controls moulds and bacterial spoilers (Niku-Paavola, Laitila, Mattila-Sandholm & Hikara, 1999;
Lavermicocca, Valerio, Evidente, Lazzaroni, Corsetti & Gobbetti, 2000; Katina, Sauri, Alakomi &
Mattila-Sandholm, 2002; Ström, Sjögren, Broberg & Schnürer, 2002; Pepe et al., 2003; Sjögren,
Magnusson, Broberg & Schnürer, 2003; Katina, Heiniö, Autio & Poutanen, 2006b; Corsetti &

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
6
Settanni, 2007; Valerio, De Bellis, Lonigro, Visconti & Lavermicocca, 2008; Gerez, Torino,
Obregozo & Font de Valdez, 2010; Coda, Cassone, Rizzello, Nionelli, Cardinali & Gobbetti, 2011;
Wang, Yan, Wang, Zhang & Qi, 2012). Recently, some authors (Komlenić, Ugarčić-Hardi, Jukić,
Planinić, Bucić-Kojić & Strelec, 2010) observed a modification of the rheological properties of
wheat flour dough, and in particular a reduction of bread hardness and an increase in specific
volume after the addition of biological (L. brevis preferment and sourdough) acidifiers. In fact, the
fermentation of dough with LAB enhances the level of organic acids that was reported to be
involved in the reduction of dough mixing time and in a significant desirable weakening of dough
(Delcour & Hoseney, 2010). The resulted bread has greater volume, lower density, softer crumb and
higher slice height (Arendt, Ryan & Dal Bello, 2007). On the other hand, a moderate dough
acidification also enhanced wheat flour proteinase activities that, at the optimal pH values (3.8 -
4.1), influence the extensibility of gluten and the final quality of bread (Thiele, Gänzle & Vogel,
2002; Schober, Dockery & Arendt, 2003). The beneficial effect of LAB observed in sourdough can
be obtained by the addition of LAB-derived acidifiers, even if their effects on physico-chemical,
textural and microbiological quality of yeast-leavened bread formulated with wheat bran, needs to
be further investigated. Thus, the aim of the current work was to apply a L. brevis-based
bioingredient in yeast-leavened wheat bread containing bran to improve the final quality of enriched
fibre bread.
2 Materials and Methods
2.1 Bacterial cultures
Lactobacillus brevis LMG P-25726 was isolated from sourdough and deposited in the Belgian
Coordinated Collections of Microorganisms (BCCM/LGM, Gent, Belgium). For long-term storage,
stock cultures were prepared by mixing 8 mL of a culture with 2 mL of Bacto glycerol (Difco,
Becton Dickinson Co., Sparks, MD, USA) and freezing 1 mL portions of this mixture at -80 °C.
Culture was stored frozen (-80°C) in MRS broth (Oxoid LTD, Basinstoke Hampshire, England)
supplemented with 20% Bacto glycerol (Difco) and subcultured twice before use.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
7
2.2 LbBio bioingredient preparation
The bioingredient (LbBio) was prepared inoculating 5 mL of an overnight (37°C, 150 rpm) L.
brevis LMG P-25726 culture in a flour-based medium obtained by a mixture of white wheat flour
(100 g), water (500 mL), demineralised whey powder W714 (5 g) (ProfileTM
90, Kerry Ingredients,
Listowel, Ireland) and fructose (5 g) and incubated at 37 °C, 150 rpm for 18 hours. The final
product was combined, instead of water amount, with ingredients of wheat bread according the
formulations reported in Table 1. As a control the flour-based medium (FBM) incubated in the same
conditions (37°C, 150 rpm, 18 hours) but not inoculated with the L. brevis strain LMG P-25726,
was used.
2.3 Bread production
The bread formulation was optimized and standardized within the European Seventh Framework
Programme project DREAM (Design and development of realistic food models with well
characterized micro- and macro-structure and composition,
http://dream.aaeuropae.org/AboutDREAM/tabid/56/Default.aspx). All ingredients were mixed and
cooked in a kneading machine (Princess
Home Breadmaker, type 1936; Princess Household
Appliance BV, Breda, Netherlands). Bread types were prepared according to a standard recipe as
reported in Table 1. Breads containing the FBM instead of the LbBio or water were also made as
controls (WWB+FBM and WBB+FBM). Finally, dough pH and total titratable acidity (TTA) were
determined after the leavening step. TTA was measured according to AOAC Method No.: 981.12
(AOAC, 1990). Loaves were stored for 3 days at 30°C, for microbiological analysis, and at 25°C
for the textural analysis.
2.4 Organic acid quantification in the bioingredient and in the dough
The bioingredient was centrifuged (8422 × g, 10 min) and the supernatant was freeze-dried,
resuspended in HPLC mobile phase (0.005 mol/L H2SO4, Fluka, Deisenhofen, Germany) and
passed through a micro-concentrator (Ultracel-3k, Amicon, Danvers, MA, USA) with a molecular-
mass cut-off of 3000 Da, by centrifugation (7000 × g, 2°C, 1 h). Solutions were loaded onto the

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
8
column. In the case of the dough, 10-gram portions were homogenized with 90 mL of sterile tap
water in a blender for 2 min. Ten millilitre-aliquots were centrifuged (8422 × g, 10 min, 10 °C) and
the supernatant was freeze-dried, resuspended in mobile phase (0.005 mol/L H2SO4) and then
passed through a 3000 Da cut-off micro-concentrator by centrifugation (7000 × g, 2°C, 1 h). The
resulting solutions were loaded onto the column. The analysis of organic acids (lactic, acetic,
propionic, phenyllactic, hydroxyl-phenyllactic, valeric, isovaleric, butyric, isobutyric acids), in the
bioingredient and in the dough, was performed by HPLC (AKTABasic10, P-900 series pump,
Amersham Biosciences AB, Uppsala, Sweden), using a Rezex ROAorganic acid H+ (8%) column
(7.80×300 mm, Phenomenex, Torrance, CA, USA) and a 3-channel UV detector (Amersham
Biosciences 900) set at 210 and 220 nm. The mobile phase was pumped at a flow rate of 0.6
mL/min through the column heated to 70°C. Quantification of the organic acids were performed by
integrating calibration curves obtained from the relevant standards.
2.5 Microbiological quality of raw materials and bread
Wheat flour and bran batches used in bread-making were checked for the presence of natural spore
–forming contaminants as reported in Valerio et al. (2012) with slight modifications. Briefly, 20 g
of each sample was diluted with 180 g of a sterile Maximum Recovery Diluent (MRD, Oxoid) and
homogenized in a stomacher (Seward, London, United Kingdom) for 2 min. The suspension was
filtered through sterile Whatman paper No 4 (Whatman, Maidstone, United Kingdom) to remove
the coarse material and heat treated for 10 min at 80 °C to select spores. Therefore, the suspension
was pour plated (1 mL) and decimally diluted and spread plated (100 μl) on Plate Count Agar
(PCA, Oxoid). In the case of raw materials expected to be contaminated at low levels (<100
spores/g), the suspension was centrifuged (8422 × g, 10 min) after heat treatment and the pellet
resuspended in one-tenth of the original volume.
To enumerate bacterial contaminants of bread samples produced in laboratory and stored at 30°C,
loaves were aseptically sliced after baking and cooling and slices were individually sealed in sterile
polyethylene bags. At each sampling time (T0, T1day and T3days), 20 g of each sample was diluted

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
9
with 180 g of sterile MRD and homogenized in a stomacher for 2 min. The food homogenate was
decimally diluted and suspensions plated on PCA agar.
Plates were incubated for 24 h at 30 °C and the number colony forming units (CFU) was counted.
Detection limit of bacterial count was 1 CFU/g.
Bread slices were also monitored during storage for rope appearance, by evaluating sweet rope
odor, discoloration of the crumb and sticky threads.
2.6 Physicochemical and firmness properties of bread
After cooking the bread loaves not subjected to microbiological analysis were cooled for 2 h and
evaluated for their weight and for volume and specific volume using the rapeseed displacement
approved method 10-05.01 (AACC International 2010). Furthermore, the loaves were sliced to
obtain six transversal slices (25 mm-thickness) that were analyzed for crumb firmness (defined as
the force required to compress the bread slice) by a Zwicki-line uniaxial testing machine (Zwick,
Ulm, Germany) with a 500 N load cell and TestXpert version 6.01 software was used. Briefly, all
slices from each loaf were compressed by 6.25 and 10 mm (25 and 40% deformation, respectively)
with a crosshead speed of 1.7 mm/s to measure the crumb hardness according to the approved
method 74-09, 2000 (AACC International 2000). These strains correspond to the common practice
to squeeze bread between the forefinger and opposed thumb to evaluate the crumb freshness or
between incisor teeth at the first bite (Bourne, 2002). Water activity values were measured on the
crumb of cooked loaves after cooling using a Decagon AquaLab Serie 3 (Steroglass, Perugia, Italy)
and the pH was recorded with a portable 110 pH meter (Oakton Instruments, Vernon Hills, IL,
USA) supplied with Double Pore D electrode (Hamilton, Bonaduz, Switzerland).
2.7 Statistical analyses
Data are presented as mean values ± standard deviation. Comparisons during time were done by
using the one-way Analysis of Variance (ANOVA), while bread types were compared by using the
two-ways ANOVA (Hogg & Ledolter, 1987). The function “anovan” of the software Matlab
R2012a (The MathWorks, Inc.) was used for testing the effects of the two factors (presence of bran

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
10
and/or LbBio) and their interactions on the mean values of the measured variables. To determine
which pairs of levels of the factors were significantly different, the Tukey’s test was adopted
(Hochberg & Tamhane, 1987). Statistical significance was assessed at a level of 5%.
3 Results
3.1 Microbiological quality of bread types during storage and organic acid quantification
Microbiological analysis of flour and bran batches used in this work indicated a low level of
bacterial spores (<103 spores/g of flour or bran) even though, the total bacterial count in baked
control bread types with and without bran (WBB and WWB) reached a concentration level close to
5 log CFU/g after only one day storage at 30°C (Table 2). After baking (T0) some differences in
bacterial counts among bread types were observed and could be related to the different batch used
for each experiment. When bread formulation was modified replacing water with a relevant amount
of the liquid bioingredient LbBio, the total bacterial count as well as the ropy spoilage appearance
were reduced after 3 days only in bread prepared with white wheat flour (WWB+LbBio), while the
efficacy of LbBio was loweredd when bread was prepared with bran (WBB+LbBio) (Table 2).
When a flour-based medium (FBM) was used as a control instead of LbBio, no inhibition of
bacterial count was observed.
The production of lactic, acetic, phenyllactic (PLA) and OH-phenyllactic (OH-PLA) acids was
evaluated in LbBio, after growing L. brevis LMG P-25726 for 18 h, in FBM and in the resulting
dough samples (Table 3). No trace of propionic, valeric, isovaleric, butyric, isobutyric acids was
found both in the LbBio, FBM and in dough samples. The concentration of lactic acid, acetic acid,
PLA and OH-PLA produced by L. brevis LMG P-25726 in the bioingredient was found to be
significantly higher (p>0.05) than in the flour based medium. Dough samples with bioingredient
contained all the acids and showed the higher molar lactic/acetic acid ratio (fermentation quotient,
FQ). Conversely, dough samples not prepared with LbBio contained only acetic acid (except for
WWB) and showed FQ values lower than 1 (Table 3).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
11
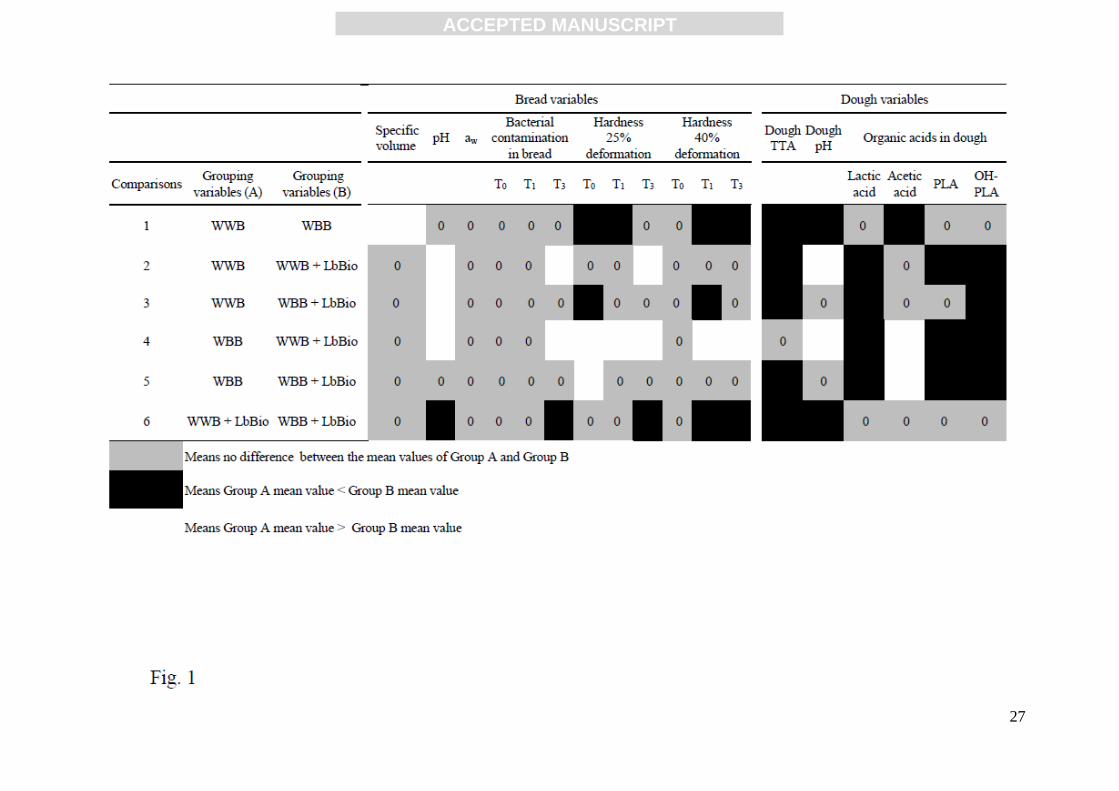
In order to assess the influence of each factor (bran and/or LbBio) and their interaction on bread
quality, a two ways ANOVA test was performed (Table 4). The analysis demonstrated the ability of
LbBio to significantly affect the dough content of lactic acid, PLA and OH-PLA regardless of the
presence of bran and this effect determined the reduction of bacterial count after three days storage.
Whereas, significant effect of bran and interaction between the two factors (LbBio and bran) were
observed for the acetic acid in dough and for bread contamination (three day-storage), although in
these bread formulations (WBB and WBB+LbBio) bacterial counts were not modified (Table 2 and
Table 4). These observations were explained by the multiple comparison Tukey’s test performed to
establish which pairs of levels of the factors were significantly different for each variable (Fig. 1).
The analysis confirmed that a significant inhibitory effect on bread contamination was exerted by
the LbBio (Table 2; Fig. 1, comparison 2), but this ability was influenced by the presence of bran.
In fact, the addition of LbBio to dough together with bran did not produce any significant changes
on bread contamination (Table 2; Fig. 1, comparison 5). The addition of bran to the dough without
LbBio did not modify its contamination level (Table 2; Fig. 1, comparison 1), whereas bran added
to dough with LbBio (WBB+LbBio) led to an increase in the bacterial count of baked bread with
respect to the formulation lacking bran (WWB+LbBio) (Table 2; Fig. 1, comparison 6). At the same
time LbBio significantly reduced the effect of bran on the acetic acid content in dough (Table 3 and
Fig. 1, comparisons 1, 6).
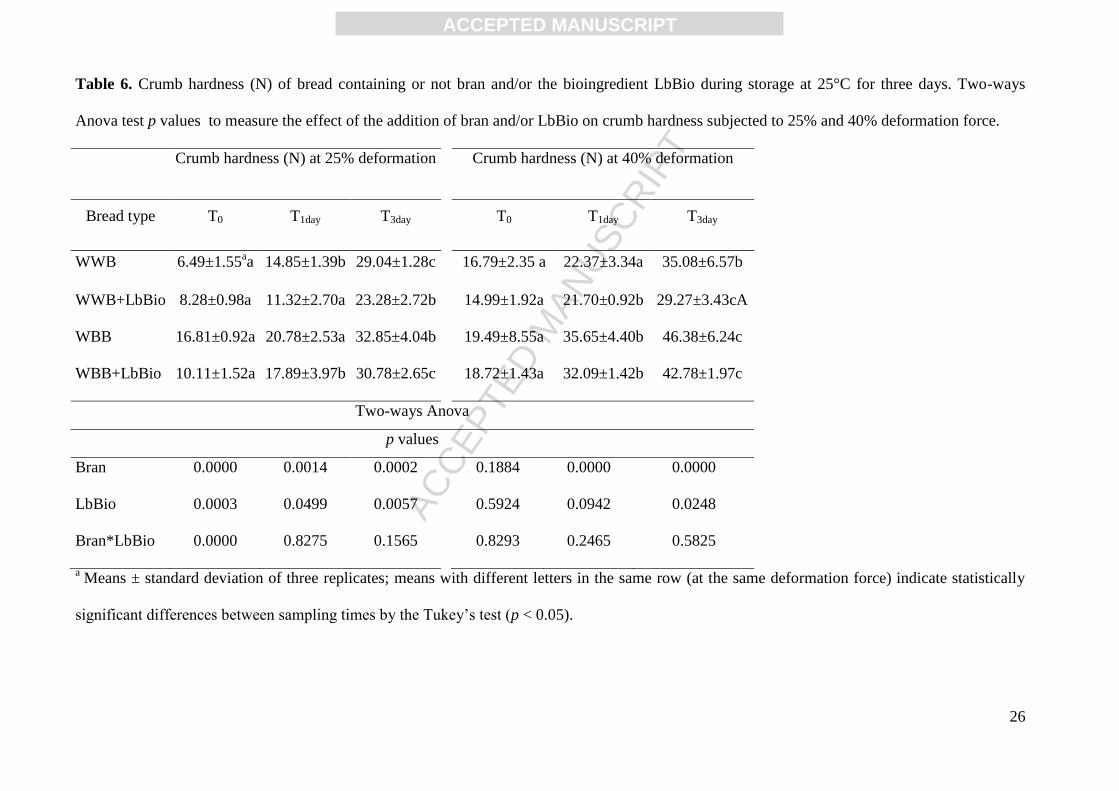
3.2 Physico-chemical and textural properties of bread
The effect of bran and/or LbBio on some physico-chemical and textural properties of bread is
reported in Tables 5 and 6, Figures 1 and 2. Results indicated that white wheat bread (WWB) was
significantly different from bread containing wheat bran (WBB) in all properties except for water
activity, bread pH and hardness at 25% deformation (Tables 5 and 6, Fig. 1, comparison 1). The use
of LbBio significantly affected dough and bread pH values in WWB which resulted most acidified
while this effect was limited by the presence of bran (Table 5, Fig. 1, comparisons 2 and 5). At the
same time, a significant increase in the TTA values was observed in both dough types after LbBio

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
12
application (WWB+LbBio and WBB+LbBio). The two ways ANOVA confirmed that the addition
of LbBio significantly affected dough and bread pH and dough TTA values (Table 5) and that this
effect was significantly influenced by bran addition (see the interaction between factors
Bran*LbBio in Table 5).
Discussion
Results of this work indicated an effective role of Lactobacillus brevis LMG P-25726 in the LbBio
technological and antimicrobial efficacy and confirmed the applicability of LAB-based
fermentation products as bioingredients in yeast-leavened bread.
The microbiological quality of breads during storage resulted improved and the rope spoilage was
delayed or hampered, in the presence of LbBio, confirming the synergistic role of organic acids as
previously observed (Valerio et al., 2008; Mantzourani, Plessas, Saxami, Alexopoulos, Galanis &
Bezirtzoglou, 2014). Although a high acetic acid content and FQ down to 1 can be associated to
enhanced inhibitory efficacy against rope producing bacteria (Röcken, 1996), the presence of only
acetic acid in breads not prepared with LbBio did not warranty bacterial inhibition. Additionally,
other authors demonstrated that acetic acid can determine a shorter and harder gluten and that the
production of lactic acid is advisable to increase elasticity of dough (Gobbetti, Corsetti & Rossi,
1995). In fact, the high molar lactic/acetic acid ratio (FQ>1) generally obtained in sourdough is
associated to pleasant odour, sensory properties and improved shelf-life. In our yeast-leavened
bread types, the presence of the LbBio allowed to obtain optimal FQ, TTA and pH values
comparable to those previously reported for sourdough bread (FQ>1, TTA: 4.8-6.2; pH: 4.8-5.1)
(Katina et al., 2002; Mantzourani et al., 2014).
Additionally to the influence on microbiological quality, the application of LAB bioingredients in
bread-making could have positive effects on some physico-chemical and textural properties of
bread as widely reported (Komlenić et al., 2010; Pepe, Ventorino, Cavella, Fagnano & Brugno,
2013). Acids produced during fermentation can influence the mixing behaviour of dough, since the
gluten protein solubility was proved to cause an increase in softness and elasticity of gluten

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
13
(Wehrle, Grau& Arendt, 1997; Takeda, Matsumura & Shimizu, 2001). In addition, the dough
acidification could enhance enzymatic activity of wheat proteases, causing an increase in free amino
acid content and improved bread flavour (Thiele et al., 2002; Clarke, Schober, Dockery, O’Sullivan
& Arendt, 2004; Rizzello, Coda, Mazzacane, Minervini & Gobbetti, 2012). In our study, a
significant increase in TTA values was observed in both dough types containing LbBio and values
were comparable to those obtained in L. brevis sourdough (Komlenić et al., 2010). Furthermore, the
addition of bran significantly influenced the effect of LbBio on pH and TTA values (see the
interaction between factors Bran*LbBio in Table 6). This result is consistent with the acidity
increase of dough when sourdough was prepared with bran fractions, than confirming an interaction
occurring between LAB and dietary fibres (Rizzello et al., 2012). Recently, Pepe et al. (2013)
demonstrated the contribute of LAB producing exopolysaccharides (EPS) in combination with
immature wheat grain flour containing fructo-olgosaccharides (FOS), in improving the nutritional
and technological characteristics of breads. The ability of LAB to produce EPS is frequently
exploited in bread technology to reduce the amount of additives since these metabolites influence
the viscoelastic characteristics and stabilize the rheological properties of dough (Palomba, Cavella,
Torrieri, Piccolo, Mazzei, Blaiotta, Ventorino & Pepe, 2012). Moreover, the use of rich-FOS
prebiotics can stimulate the microbial metabolism leading to an increase in TTA (Pepe et al., 2013).
4 Conclusions
The bioingredient Lactobacillus brevis–based LbBio, acting as a sourdough, counteract the negative
effects of bran allowing to obtain an enriched-fibre bread with overall quality comparable to that of
the reference white wheat bread. The study also indicated that bread formulation can modulate
bacterial behaviour during storage and consequently the bread spoilage. Further investigations are
needed to better evaluate the positive and negative interactions on bread quality occurring when
different levels of dietary fibers are used in the presence of the bioingredient LbBio.
Acknowledgments

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
14
This research has received funding from the European Community's Seventh Framework
Programme (FP7/ 2007-2013) under the grant agreement no FP7-222 654-DREAM.
N.A. and T.M.C were supported by Project FIRB CAROMICS RBAP11B2SX.
References
AACC International (2000). Approved Methods of Analysis, 10th Ed. Method 74–09. AACC
International: St. Paul, MN. AACC method 74-09, 2000.
AACC International (2010). Approved Methods of Analysis, 11th Ed. Methods 10-05.01. Available
online only. AACC International: St. Paul, MN. AACC method 10-05.01, 2010.
AOAC (1990). Official methods of analysis. Washington, DC: Association of Official Analytical
Chemists. TTA, AOAC method No.: 981.12, 1990.
Arendt, E.K., Ryan, L.A.M. & Dal Bello, F. (2007). Impact of sourdough on the texture of bread.
Food Microbiology, 24, 165-174.
Bourne, M.C., 2002. Sensory methods of texture and viscosity measurement. In: Bourne M.C. (Ed)
Food texture and viscosity: concept and measurement (chapter 7, pp. 257-291), 2nd
edition.
Academic Press, London UK.
Chavan, R.S. & Chavan, S.R. (2011). Sourdough technology - a traditional way for wholesome
foods: a review. Comprehensive Reviewers in Food Science and Food Safety, 10, 169-182.
Clarke, C.I., Schober, T.J., Dockery, P., O'Sullivan, K. & Arendt, E.K., (2004). Wheat sourdough
fermentation: effect of time and acidification on fundamental rheological properties. Cereal
Chemistry Journal, 81, 409-417.
Coda, R., Cassone, A., Rizzello, C.G., Nionelli, L., Cardinali, G. & Gobbetti, M. (2011). Antifungal
activity of Wickerhamomyces anomalus and Lactobacillus plantarum during sourdough
fermentation: identification of novel compounds and long-term effect during storage of
wheat bread. Applied and Environmental Microbiology, 77, 3484-3492.
Cornell, H.J. & Hoveling, A.W. (1998). Wheat: chemistry and utilization. Technomic Publishing
Company, CRC Pressl Llc.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
15
Corsetti, A. & Settanni, L. (2007). Lactobacilli in sourdough fermentation. Food Reseach
International, 40, 539–558.
Damen B., Pollet A., Dornez E., Broekaert W.F., Van Haesendonck I., Trogh I., Arnaut F., Delcour
J.A., Courtin C.M., (2012). Xylanase-mediated in situ production of arabinoxylan
oligosaccharides with prebiotic potential in whole meal breads and breads enriched with
arabinoxylan rich materials. Food Chemistry, 131, 111-118.
Delcour J. A. & Hoseney R.C., (2010). Rheology of Doughs and Batters. In: Delcour J. A. and
Hoseney R.C. (Eds). Principles of Cereal Science and Technology, (pp. 87-96). Third
edition, American Association of Cereal Chemists, Inc. St Paul, Minnesota.
De Jonghe, V., Coorevits, A., De Block, J., Van Coillie, E., Grijspeerdt, K., Herman, L., De Vos, P.
& Heyndrickx, M. (2010). Toxinogenic and spoilage potential of aerobic spore-formers
isolated from raw milk. International Journal of Food Microbiology, 136, 318–325.
EFSA 2010. Scientific Opinion on the substantiation of health claims related to dietary fibre (ID
744, 745, 746, 748, 749, 753, 803, 810, 855, 1415, 1416, 4308, 4330) pursuant to Article
13(1) of Regulation (EC) No 1924/2006. EFSA Journal, 8(10): 1817-1835.
Gerez, C.L., Torino, M.I., Obregozo, M.D. & Font de Valdez, G. (2010). A ready-to-use antifungal
starter culture improves the shelf life of packaged bread. Journal of Food Protection, 73,
758-762.
Gobbetti, M., Corsetti, A. & Rossi, J. (1995). Interaction between lactic acid bacteria and yeasts in
sourdough using a rheofermentometer. World Journal Microbiology and Biotechnology, 11,
625-630.
Hochberg, Y. & Tamhane, A. C. (1987). Multiple Comparison Procedures. New York: John Wiley
& Sons.
Hogg, R. V. & Ledolter, J. (1987). Engineering Statistics. New York: MacMillan.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
16
Katina, K., Sauri, M., Alakomi, H.-L. & Mattila-Sandholm, T. (2002). Potential of lactic acid
bacteria to inhibit rope spoilage in wheat sourdough bread. LWT – Food Science and
Technology, 35, 38-45.
Katina, K., Salmenkallio-Marttila, M., Partanen, R., Forssell, P. & Autio, K. (2006a). Effects of
sourdough and enzymes on staling of high-fibre wheat bread. LWT – Food Science and
Technology, 39, 479-491.
Katina, K., Heiniö, R.-L., Autio, K. & Poutanen, K. (2006b). Optimization of sourdough process for
improved sensory profile and texture of wheat bread. LWT – Food Science and Technology,
39, 1189-1202.
Katina, K., Laitila, A., Juvonen, R., Liukkonen, K.-H., Kariluoto, S., Piironen, V., Landberg, R.,
Åman, P. & Poutanen, K. (2007). Bran fermentation as a means to enhance technological
properties and bioactivity of rye. Food Microbiology, 24, 175-186.
Komlenić., D. K., Ugarčić-Hardi, Ž., Jukić, M., Planinić, M., Bucić-Kojić, A. & Strelec, I. (2010).
Wheat dough rheology and bread quality effected by Lactobacillus brevis preferment, dry
sourdough and lactic acid addition. International Journal of Food Science & Technology,
45, 1417-1425.
Kramer, J.M. & Gilbert, R.J. (1989). Bacillus cereus and other Bacillus species. In: Doyle, M.P.
(Ed.). Foodborne bacterial pathogens (pp. 21–70) Marcel Dekker, New York.
Laurikainen, T., Härkönen, H., Autio, K. & Poutanen, K. (1998). Effects of enzymes in fibre-
enriched baking. Journal of the Science of Food and Agriculture, 76, 239-249.
Lavermicocca, P., Valerio, F., Evidente, A., Lazzaroni, S., Corsetti, A. & Gobbetti, M. (2000).
Purification and characterization of novel antifungal compounds from the sourdough
Lactobacillus plantarum strain 21B. Applied and Environmental Microbiology, 66, 4084-
4090.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
17
Mantzourani, I., Plessas, S., Saxami, G., Alexopoulos, A., Galanis, A. & Bezirtzoglou, E. (2014).
Study of kefir grains application in sourdough bread regarding rope spoilage caused by
Bacillus spp. Food Chemistry, 143, 17-21.
Niku-Paavola, M., Laitila L.A., Mattila-Sandholm, T. & Hikara, A. (1999). New types of
antimicrobial compounds produced by Lactobacillus plantarum. Journal of Applied
Microbiology, 86, 29-35.
Palomba, S., Cavella, S., Torrieri, E., Piccolo, A., Mazzei, P., Blaiotta, G., Ventorino,V., & Pepe, O
(2012). Polyphasic screening, homopolysaccharide composition, and viscoelastic behavior
of wheat sourdough from sourdough from a Leuconostoc lactis and Lactobacillus curvatus
exopolysaccharide-producing starter culture. Applied and Environmental Microbiology, 78,
2737–2747.
Pepe, O., Blaiotta, G., Moschetti, G., Greco, T. & Villani, F. (2003). Rope-producing strains of
Bacillus spp. from wheat bread and strategy for their control by lactic acid bacteria. Applied
and Environmental Microbiology, 69, 2321–2329.
Pepe, O., Ventorino, V., Cavella, S., Fagnano M., & Brugno, R. (2013). Prebiotic contento f bread
prepared with flour from immature wheat grain and selected dextran-producing lactic acid
bacteria. Applied and Environmental Microbiology, 79, 3779–3785.
Pomeranz, Y. (1988). Wheat: Chemistry and Technology. American Association of Cereal
Chemists, Inc. St Paul, Minnesota.
Rizzello, C.G., Coda, R., Mazzacane, F., Minervini, D. & Gobbetti, M. (2012). Micronized by-
products from debranned durum wheat and sourdough fermentation enhanced the
nutritional, textural and sensory features of bread. Food Research International, 46, 304-
313.
Röcken, W. (1996). Applied aspects of sourdough fermentation. Advances in Food Science, 18,
212-216.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
18
Rosenkvist, H. & Hansen, Å. (1995). Contamination profiles and characterization of Bacillus
species in wheat bread and raw materials for bread production. International Journal of
Food Microbiology, 26, 353–363.
Schober, T.J., Dockery, P. & Arendt, E.K. (2003). Model studies for wheat sourdough systems
using gluten, lactate buffer and sodium chloride. European Food Research & Technology,
217, 235-243.
Sjögren, J., Magnusson, J., Broberg, A. & Schnürer, J. (2003). Antifungal 3-hydroxy fatty acids
from Lactobacillus plantarum MiLAB 14. Applied and Environmental Microbiology, 69,
7554-7557.
Ström, K., Sjögren, J., Broberg, A. & Schnürer, J. (2002). Lactobacillus plantarum MiLAB 393
produces the antifungal cyclic dipeptides cyclo(L-Phe-L-Pro) and cyclo(L-Phe-trans-4-OH-
L-Pro) and 3-phenyllactic acid. Applied and Environmental Microbiology, 68, 4322-4327.
Takeda, K., Matsumura, Y., Shimizu, M. (2001). Emulsifying and surface properties of wheat
gluten under acidic conditions. Journal of Food Science, 66, 393–399.
Thiele, C., Gänzle, M.G. & Vogel, R.F. (2002). Contribution of sourdough lactobacilli, yeast and
cereal enzymes to the generation of amino acids in dough relevant for bread flavour. Cereal
Chemistry, 79, 45–51.
Valerio, F., De Bellis, P., Lonigro, S.L., Visconti, A. & Lavermicocca, P. (2008). Use of
Lactobacillus plantarum fermentation products in bread-making to prevent Bacillus subtilis
ropy spoilage. International Journal of Food Microbiology, 122, 328-332.
Valerio, F., De Bellis, P., Di Biase, M., Lonigro, S.L., Giussani, B., Visconti, A., Lavermicocca, P.
& Sisto, A. (2012). Diversity of spore-forming bacteria and identification of Bacillus
amyloliquefaciens as a species frequently associated with the ropy spoilage of bread.
International Journal of Food Microbiology, 156, 278-285.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
19
Viedma, P.M., Abriouel, H., Omar, N.B., López, R.L. & Gálvez, A. (2011). Inhibition of spoilage
and toxigenic Bacillus species in dough from wheat flour by the cyclic peptide enterocin
AS-48. Food Control, 22, 756-761.
Wang, H.K., Yan, Y.H, Wang, J.M., Zhang, H.P & Qi, W. (2012). Production and characterization
of antifungal compounds produced by Lactobacillus plantarum IMAU10014. PLOS ONE,
7(1), e29452.
Wehrle, K., Grau, H. & Arendt, E.K. (1997). Effects of lactic acid, acetic acid and table salt on
fundamental rheological properties of wheat dough. Cereal Chemistry, 74, 739–744.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
20
Figure 1. Multiple comparison test on each pair of factor levels.
Figure 2. Slices of bread loaves produced with A) white wheat flour (WWB), B) white wheat flour
and wheat bran (20% w/w of wheat flour) (WBB), C) white wheat flour and LbBio (WWB+LbBio),
and D) white wheat flour, wheat bran (20% w/w of wheat flour) and LbBio (WBB+LbBio).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
21
Table 1. Bread formulations containing or not wheat bran and/or Lactobacillus brevis-based
bioingredient (LbBio) and/or the flour-based medium (FBM) as control.
Bread typea
WWB
WBB WWB+LbBio WBB+LbBio WWB+ FBM WBB+ FBM
Ingredient Mass (g)
White wheat flour 350 280 350 280 350 280
Wheat bran - 70 - 70 - 70
Salt 6.3 6.3 6.3 6.3 6.3 6.3
Margarine 10.5 10.5 10.5 10.5 10.5 10.5
Dry yeast 5.25 5.25 5.25 5.25 5.25 5.25
Tap water 210 245 - 35 - 35
LbBio - - 210 210 - -
FBM - - - - 210 210
a White wheat bread (WWB), White bran bread (WBB), White wheat bread+LbBio (WWB+LbBio),
Wheat bran bread+LbBio (WBB+LbBio), White wheat bread+ FBM (WWB+ FBM), Wheat bran
bread+ FBM (WBB+ FBM).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
22
Table 2. Rope spoilage and total viable count (log CFU/g) of bacterial contaminants naturally
occurring in white wheat bread (WWB) and wheat bran bread (WBB) containing or not the
bioingredient (WWB+LbBio or WBB+LbBio) or the flour based medium (WWB+FBM or
WBB+FBM) and stored at 30°C for different times.
Bread type T0 T1day T3day Rope (T3day)
WWB 2.96±1.10aa
4.61±1.10a 6.93±1.03b ++b
WWB+LbBio 2.67±1.32a 2.74±0.90ab 4.52±1.01b -
WWB+FBM 2.58±0.26a 2.61±0.09a 6.10±0.43b +
WBB 1.15±0.21a 4.80±1.17b 7.34±0.89c ++
WBB+LbBio 2.62±2.13a 3.08±1.53ab 6.63±0.98b +
WBB+FBM 1.38±0.12a 3.52±0.26a 7.17±0.54b ++
a Means ± standard deviation of three replicates; means with different letters in the same row
indicate statistically significant differences between samples by the Tukey test (p < 0.05).
b Rope occurrence in bread crumb: -: no ropey alteration; +: typical ropey smell; ++: ropey slime.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
23
Table 3. Organic acid content in the bioingredient LbBio, in the flour-based medium (FBM) and in
the dough samples prepared with white wheat flour (WWB) or a mixture of white wheat flour and
wheat bran (WBB) and containing or not the LbBio or the FBM.
Lactic acid Acetic acid PLA OH-PLA FQa
Ingredient mmol/L
LbBio 30.43 ± 5.23 ba 4.37 ± 1.01a 0.05 ± 0.02a 0.02 ± 0.01a
FBMc
3.90± 0.84b 9.11± 0.06b -d b - b
Dough sample mmol/Kg
WWB <DLe a <DLa <DL a <DLa -
WWB +LbBio 9.31±2.71b 3.43±1.47a 0.02 ± 0.01b 0.01± 0.00b 2.71
WWB+FBM <DLa 2.58±2.40a <DLa <DLa 0.18
WBB <DLa 11.13±1.7b <DLa <DLa -
WBB+LbBio 9.36±5.39b 4.60±2.74ac 0.02±0.01b 0.01±0.01b 2.03
WBB+FBM <DLa 8.32±2.66bc <DLa <DLa 0.07
a Fermentation Quotient: molar ratio between lactic and acetic acids
b Means ± standard deviation of three replicates; means with different letters in the same column
indicate statistically significant differences between samples by the Tukey test (p < 0.05).
c Flour-based medium not inoculated with Lactobacillus brevis LMG P-25726 and used as control.
d not found.
e DL: detection limit. In LbBio and FBM DL were: 3.60 mM (lactic acid), 1.62 mM (acetic acid),
0.038 mM (PLA), 0.015 mM (OH-PLA). In dough DL were: 2.26 mmol/Kg (lactic acid), 1.37
mmol/Kg (acetic acid), 0.0004 mmol/Kg (PLA), 0.0008 mmol/Kg (OH-PLA).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
24
Table 4. Two-ways Anova test p values to measure the effect of the addition of bran and/or LbBio and their interaction on bacterial contamination
of bread during storage at 30°C for different times and on the organic acids content of dough.
Bacterial
contamination of
bread
Organic acids content in dough
factors T0 T1day T3day Lactic acid Acetic acid PLA OH-PLA
Bran 0.1616 0.7735 0.0008 0.9875 0.0001 0.8048 0.2396
LbBio 0.3673 0.1081 0.0000 0.0009 0.1507 0.0012 0.0003
Bran*LbBio 0.1842 0.9319 0.0195 0.9875 0.0005 0.8048 0.2396

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
25
Table 5. Physico-chemical properties of bread types and two-ways Anova test p values to measure the effect of the addition of bran and/or LbBio
on physicochemical properties of bread and dough.
Bread dough Bread
Bread type Dough pH TTAa Specific volume sample
(cm3/g)
Water activity
(aw)
Bread pH
WWB 5.24± 0.37ba 1.96 ± 0.51 3.20 ± 0.51 0.94 ± 0.03 5.56 ± 0.50b
WWB+LbBio 4.57± 0.14a 4.45 ± 0.07 3.10± 0.00 0.94 ± 0.01 4.82 ± 0.18b
WBB 5.43± 0.15a 3.96 ± 0.06 2.65 ± 0.23 0.96 ± 0.01 5.49 ± 0.14a
WBB+LbBio 5.17± 0.11a 5.70 ± 0.14 2.65 ± 0.35 0.95 ± 0.01 5.20 ± 0.16a
Two-ways Anova
p values
factors
Bran 0.0000 0.0000 0.0341 0.0678 0.0705
LbBio 0.0000 0.0000 0.8036 0.7384 0.000
Bran*LbBio 0.0490 0.0452 0.8229 0.3396 0.0041
avalues represent volume (mL) of NaOH 0.1 N. TTA was determined using the AOAC method N° 981.12
b Means ± standard deviation of three replicates; means with different letters in the same row indicate statistically significant differences between
samples by the Tukey’s test (p < 0.05).
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
26
Table 6. Crumb hardness (N) of bread containing or not bran and/or the bioingredient LbBio during storage at 25°C for three days. Two-ways
Anova test p values to measure the effect of the addition of bran and/or LbBio on crumb hardness subjected to 25% and 40% deformation force.
Crumb hardness (N) at 25% deformation Crumb hardness (N) at 40% deformation
Bread type T0 T1day T3day T0 T1day T3day
WWB 6.49±1.55aa 14.85±1.39b 29.04±1.28c 16.79±2.35 a 22.37±3.34a 35.08±6.57b
WWB+LbBio 8.28±0.98a 11.32±2.70a 23.28±2.72b 14.99±1.92a 21.70±0.92b 29.27±3.43cA
WBB 16.81±0.92a 20.78±2.53a 32.85±4.04b 19.49±8.55a 35.65±4.40b 46.38±6.24c
WBB+LbBio 10.11±1.52a 17.89±3.97b 30.78±2.65c 18.72±1.43a 32.09±1.42b 42.78±1.97c
Two-ways Anova
p values
Bran 0.0000 0.0014 0.0002 0.1884 0.0000 0.0000
LbBio 0.0003 0.0499 0.0057 0.5924 0.0942 0.0248
Bran*LbBio 0.0000 0.8275 0.1565 0.8293 0.2465 0.5825
a Means ± standard deviation of three replicates; means with different letters in the same row (at the same deformation force) indicate statistically
significant differences between sampling times by the Tukey’s test (p < 0.05).
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
27

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
28

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
29
Highlights
Lactobacillus brevis-based bioingredient (LbBio) improves bread texture and quality
Bread formulation influences bacterial behavior in bread and the consequent spoilage
LbBio counteracts the negative effects of bran


















