
Effect of changes in the fractal structure of a littoral ... fileseral stage, the most important...
26
Submitted 24 May 2018 Accepted 29 August 2018 Published 26 September 2018 Corresponding author Joanna Pakulnicka, [email protected] Academic editor María Ángeles Esteban Additional Information and Declarations can be found on page 20 DOI 10.7717/peerj.5662 Copyright 2018 Pakulnicka and Zawal Distributed under Creative Commons CC-BY 4.0 OPEN ACCESS Effect of changes in the fractal structure of a littoral zone in the course of lake succession on the abundance, body size sequence and biomass of beetles Joanna Pakulnicka 1 and Andrzej Zawal 2 1 Department of Ecology and Environmental Protection, Faculty of Biology and Biotechnology, University of Warmia and Mazury Olsztyn, Olsztyn, Poland 2 Department of Invertebrate Zoology and Limnology, Institute for Research for Biodiversity, Centre of Molecular Biology and Biotechnology, Faculty of Biology, University of Szczecin, Szczecin, Poland ABSTRACT Dystrophic lakes undergo natural disharmonic succession, in the course of which an increasingly complex and diverse, mosaic-like pattern of habitats evolves. In the final seral stage, the most important role is played by a spreading Sphagnum mat, which gradually reduces the lake’s open water surface area. Long-term transformations in the primary structure of lakes cause changes in the structure of lake-dwelling fauna assemblages. Knowledge of the succession mechanisms in lake fauna is essential for proper lake management. The use of fractal concepts helps to explain the character of fauna in relation to other aspects of the changing complexity of habitats. Our 12-year-long study into the succession of water beetles has covered habitats of 40 selected lakes which are diverse in terms of the fractal dimension. The taxonomic diversity and density of lake beetles increase parallel to an increase in the fractal dimension. An in-depth analysis of the fractal structure proved to be helpful in explaining the directional changes in fauna induced by the natural succession of lakes. Negative correlations appear between the body size and abundance. An increase in the density of beetles within the higher dimension fractals is counterbalanced by a change in the size of individual organisms. As a result, the biomass is constant, regardless of the fractal dimension. Subjects Biodiversity, Conservation Biology, Ecology, Entomology, Freshwater Biology Keywords Water beetles, Lakes, Disharmonic succession, Fractal dimension, Body size INTRODUCTION Ecological succession is a natural course of events that occurs in lakes (Kajak, 1998). Lake succession manifests itself in the growth of macrophytes along lake shores, which initially create increasingly diverse and compact communities (McFarland, Carse & Sandin, 2009; Drinan et al., 2013; Beadle, Brown & Holden, 2015; Šiling & Urbanič, 2016; Stryjecki et al., 2017), only to have the littoral zone ultimately dominated by a single species. If the succession is disharmonic, the dominant species is Sphagnum sp. (Kajak, 1998), whereas in lakes undergoing harmonic succession the prevalent species are most often Phragmites australis (Ciecierska, 2008; Pakulnicka, Górski & Bielecki, 2015b), and less frequently Carex How to cite this article Pakulnicka and Zawal (2018), Effect of changes in the fractal structure of a littoral zone in the course of lake suc- cession on the abundance, body size sequence and biomass of beetles. PeerJ 6:e5662; DOI 10.7717/peerj.5662
Transcript of Effect of changes in the fractal structure of a littoral ... fileseral stage, the most important...

Submitted 24 May 2018Accepted 29 August 2018Published 26 September 2018
Corresponding authorJoanna Pakulnickajoannapakulnickauwmedupl
Academic editorMariacutea Aacutengeles Esteban
Additional Information andDeclarations can be found onpage 20
DOI 107717peerj5662
Copyright2018 Pakulnicka and Zawal
Distributed underCreative Commons CC-BY 40
OPEN ACCESS
Effect of changes in the fractal structureof a littoral zone in the course of lakesuccession on the abundance body sizesequence and biomass of beetlesJoanna Pakulnicka1 and Andrzej Zawal2
1Department of Ecology and Environmental Protection Faculty of Biology and Biotechnology University ofWarmia and Mazury Olsztyn Olsztyn Poland
2Department of Invertebrate Zoology and Limnology Institute for Research for Biodiversity Centre ofMolecular Biology and Biotechnology Faculty of Biology University of Szczecin Szczecin Poland
ABSTRACTDystrophic lakes undergo natural disharmonic succession in the course of which anincreasingly complex and diverse mosaic-like pattern of habitats evolves In the finalseral stage the most important role is played by a spreading Sphagnum mat whichgradually reduces the lakersquos open water surface area Long-term transformations inthe primary structure of lakes cause changes in the structure of lake-dwelling faunaassemblages Knowledge of the succession mechanisms in lake fauna is essential forproper lake management The use of fractal concepts helps to explain the characterof fauna in relation to other aspects of the changing complexity of habitats Our12-year-long study into the succession of water beetles has covered habitats of 40selected lakes which are diverse in terms of the fractal dimension The taxonomicdiversity and density of lake beetles increase parallel to an increase in the fractaldimension An in-depth analysis of the fractal structure proved to be helpful inexplaining the directional changes in fauna induced by the natural succession of lakesNegative correlations appear between the body size and abundance An increase in thedensity of beetles within the higher dimension fractals is counterbalanced by a changein the size of individual organisms As a result the biomass is constant regardless ofthe fractal dimension
Subjects Biodiversity Conservation Biology Ecology Entomology Freshwater BiologyKeywords Water beetles Lakes Disharmonic succession Fractal dimension Body size
INTRODUCTIONEcological succession is a natural course of events that occurs in lakes (Kajak 1998)Lake succession manifests itself in the growth of macrophytes along lake shores whichinitially create increasingly diverse and compact communities (McFarland Carse amp Sandin2009 Drinan et al 2013 Beadle Brown amp Holden 2015 Šiling amp Urbanič 2016 Stryjeckiet al 2017) only to have the littoral zone ultimately dominated by a single species If thesuccession is disharmonic the dominant species is Sphagnum sp (Kajak 1998) whereasin lakes undergoing harmonic succession the prevalent species are most often Phragmitesaustralis (Ciecierska 2008 Pakulnicka Goacuterski amp Bielecki 2015b) and less frequently Carex
How to cite this article Pakulnicka and Zawal (2018) Effect of changes in the fractal structure of a littoral zone in the course of lake suc-cession on the abundance body size sequence and biomass of beetles PeerJ 6e5662 DOI 107717peerj5662
sp Sparganium sp or Acorus sp Many hydrobiologists emphasize the important role ofthe littoral zone in the secondary production of lakes and as a zone having the highestspecies richness and density of macroinvertebrates (Cremona Planas amp Lucotte 2008Timm ampMoumlls 2012Mieczan et al 2014 Płaska amp Mieczan 2018) The littoral zone is alsoconsidered to be the most sensitive part of a lake and its character provides evidence onthe ecological condition of the whole lake (Czachorowski 1998 Šiling amp Urbanič 2016)
The succession of lakes entails changes in the lake-dwelling fauna (Kowalik 1968Pakulnicka amp Bartnik 1999 Ranta 1985 Kordylas 1990 Lundkvist Landin amp Milberg2001 Pakulnicka amp Zawal 2007) This problem is rarely raised in research and when itis it most often consists of brief studies into zooplankton (eg Demetraki-Paleolog 2012Kuczyńska-Kippen 2008 Jasser 1997 Lepistouml amp Saura 1998 Odland amp Del Moral 2002Fiłoc amp Kupryjanowicz 2015) More detailed investigations pertaining to changes in lakefauna during consecutive seral stages are conducted on anthropogenic rather than naturalecosystems (Pakulnicka 2008 Bloechl et al 2010 Buczyński 2015 Pakulnicka Goacuterski ampBielecki 2015b) Hence despite the wealth of references our knowledge of the successionmechanisms in lake fauna ismodest and fragmentary even though it is extremely importantfor developing proper lake management practice (Šiling amp Urbanič 2016 Shadrin et al2016)
Another challenge for researchers is to find an adequate measure for the determinationof changes in fauna induced by the succession of water bodies Hydrobiologists mostoften draw attention to changes in the abundance and species richness while comparingsmall groups of lakes with different trophic states (Kowalik 1968 Ranta 1985 Kordylas1990 Lundkvist Landin amp Milberg 2001 Pakulnicka amp Zawal 2007 Soldaacuten et al 2012Barndt 2012 Drinan et al 2013 Pakulnicka et al 2013 Baars et al 2014 Beadle Brownamp Holden 2015) A measure that has been gaining popularity over the last twenty yearsconsists of an analysis of quantitative relations between components (generalists anddiversified specialists) distinguished on the basis of their affinity towards specific habitatconditions (eg Kowalik 1968 Kordylas 1990 Czachorowski 1998 Pakulnicka amp Zawal2007 Płaska amp Tarkowska-Kukuryk 2014 Pakulnicka et al 2016a Pakulnicka et al 2016bPłaska et al 2016 Stryjecki et al 2017) However the results obtained from this approachare discrepant and therefore not highly reliable often because of a small number of samplesor analysed objects
An important addition to research consists of analyses based onbiometricmeasurementsfound also in studies on small water organisms including beetles (Jeffries 1993McAbendroth et al 2005 Ulrich 2007 Vamosi Naydani amp Vamosi 2007 Cremona Planasamp Lucotte 2008 Scheffer et al 2015 Scheffer amp Van Nes 2006 Tokeshi amp Arakaki 2012Deacutesamoreacute et al 2018) In recent years biometric measures have become a tool applied inevolutionary ecology Some hydrobiologists eg Scheffer amp Van Nes (2006) and Schefferet al (2015) analyse the structure of body sizes of organisms in specific communities fromthe viewpoint of coevolution of concurrent species Others eg Deacutesamoreacute et al (2018)search for relationships between the evolution of a body size and the environment as wellas the diversification of species in various water habitats
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 226
Data regarding the body mass and body size of macroinvertebrates often appear in thecontext of studies on a fractal structure a notion which still awaits a better understanding(Jeffries 1993 Tokeshi amp Arakaki 2012 Barnes Vaughan amp Ormerod 2013) and a fractaldimension a borrowing from mathematical sciences (Mandelbrot 1983) According toAndrejczuk (2014) all landscapes with inner diversity demonstrate fractality ie they arecomposed of smaller fragments self-similar fractals which are self-reproducing duplicatorsof parameters on a different scale Thus a fragment of any system should contain all thesystemrsquos characteristics in a nutshell (on a smaller scale) The littoral zone of ecologicallyyoung lakes can serve as an example of a linear structure of spatial organisation whichdemonstrates properties of fractality However as the succession progresses the shorelinebecomes more diverse and turns into a multifractal system ie heterogeneous composedof fragments (separate subsets) with different local characteristics each presenting self-similar properties (Miałdun 2010 Miałdun amp Ostrowski 2010) Thus similar fragments(fractals) of the littoral zone of different objects should be characterised by similar habitattraits offering the same niches to the organisms which populate them A convenient andvaluable object in studies on the fractal structure are dystrophic lakes in which varioushabitats (fragments of the littoral zone) appear during the course of succession from aplant-free zone to zones overgrown with macrophyte communities of various compactnessto a compact Sphagnum mat with small pools
In our studywe looked at water beetles because they are particularly numerous organismsin the littoral zone as well as being very sensitive and responsive to any unfavourable changesin the environment (Eyre Foster amp Foster 1992 Foster amp Eyre 1992 Corbet et al 1980Winfield Fairchild Faulds amp Matta 2000 Bosi 2001 Menetrey et al 2005 McFarlandCarse amp Sandin 2009 Gioria Bacaro amp Feehan 2010a Gioria et al 2010b Yee 2014)Furthermore many species are predators which defines their important role as organismsregulating the abundance and species richness of concurrent taxa (Yee 2014 Frelik ampPakulnicka 2015 Frelik amp Pakulnicka 2015 Perissinotto Bird amp Bilton 2016 Miller ampBergsten 2016 Płaska amp Mieczan 2018) Water beetles penetrate both the ground andthe water column so according to McAbendroth et al (2005) their ecological niche isthree-dimensional Consequently the fractal dimension of the habitats they occupy iswithin the range of 2ltDlt 3 (increasing from the least to the most compact habitats)(Cremona Planas amp Lucotte 2008 Tokeshi amp Arakaki 2012)
In the light of the above considerations our aim has been to investigate whether (1)the distinguished habitats are populated by similar assemblages of organisms with respectto body size and trophic preferences (2) if there is a relationship between the body massand abundance of water beetles (3) if the fractal dimension of a habitat has an effect onthe total biomass of beetles and whether the total biomass of beetles in particular habitatsis the same and finally (4) if there are changes in the sequence of body size of organismsoccurring in the course of disharmonic succession
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 326
Figure 1 Study area Location of lakes (1ndash40) 1 Żabie 2 Szare 3 Wygoda 4 Krypko 5 Pałsznik6 Małe Gacno 7 Wielkie Gacno 8 Długie 9 Moczadło 10 Sosnoacutewek 11 Żabionek 12 Klimontek13 Nierybno 14 Małe Łowne 15 Czarne 16 Piecki 17 Babionek Duży 18 Babionek Mały 19 Białe20 Kociołek 21 Żabie 22 Motylek 23 Purdka 24 Jonkowo 25 Borkowskie 26 Bobroacutewko 27Gryżewskie 28 Skarp 29 Zakręt 30 Kruczy Stawek 31 Kruczek Duży 32 Kruczek Mały 33 KruczeOko 34 Kruczy Staw 35 Suchar Wielki 36 Suchar 1 Lake 37 Suchar 2 38 Suchar 3 39 Suchar 4 40Suchar 5
Full-size DOI 107717peerj5662fig-1
MATERIAL amp METHODSStudy area and field studiesThe study covered 40 dystrophic lakes located in northern Poland the South BalticCoastland the West Pomeranian Olsztyn Mrągowo Lakelands and the Suwałki Lakeland(Fig 1 Appendix S1) The lakes chosen for the study were of various surface areas andhaving a floating peat mat growth belonging to Sphagnum sp that varied in size The lakesrepresented various succession stages in a disharmonic seriesmdashfrom oligo- to polyhumiclakes The oligohumic lakes were inhabited by Juncus bulbosus Eleocharis palustrisPhragmites australis Typha angustifolia Typha latiffolia Sphagnum sp Lobelia dortmannaIsoeumltes lacustris Drosera rotundifolia D anglica D intermedia Sparganium angustifoliumLycopodiella inundata In polyhumic lakes the dominant species included Sphagnum spOxycoccus quadripelatus Andromeda polifonia Eriophorum vaginatum Calluna vulgarisErica tetralix Empetrum nigrum The lakes were arranged a priori according to the seralstages of succession (Bloechl et al 2010) and the point of reference was assumed to be the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 426
coverage of a lakersquos surface by a Sphagnum mat (0ndash75) (Appendix S1) The percentagecover by a Sphagnum mat in each lake was calculated in the GIS system supported byArcMap 931 software For charting the research sites data made available throughGeoportal 2 in the WMS format were employed
The study was carried out from 2002 to 2014 in spring summer and autumn The faunasamples were collected with a dip net from a surface area of about 1 m2 In the compactedSphagnum mat environment a sample comprised 10 subsamples (aggregated afterwards)which were collected using a 01 m2 strainer Sampling sites were chosen so as to representthe biggest array of the littoral habitat diversity and areas of individual lakes Thus fivedifferent littoral components (habitats) were identified (1) the Sphagnum mat (2) sparsemacrophyte zone (3) dense macrophyte zone (4) sandy bottom zone and (5) pockets andponds within a Sphagnum mat The vegetation cover was assessed using Braun-Blanquet(1964) phytosociological records All lakes were identified with respect to the surface areaextent of the lakersquos surface covered with a Sphagnummat which corresponds to successionstages (1ndash3) percent share of individual habitats in the littoral zone (1ndash4) and a seralstage (1ndash40) (Appendix S1) It was assumed that these habitats due to different degreesof complexity represent different fractal dimensions all within an interval of 20 gtD gt30(Tokeshi amp Arakaki 2012) A schematic representation of fractals different in size can beseen in Fig 2 The smallest size (1) corresponds to the sandy bottom zone while the largestone (5) is assigned to the Sphagnummat The intermediate sizes belong to (2) pockets andponds within the Sphagnum mat (3) sparse macrophyte zone and (4) dense macrophytezone Field studies were conducted in accordance with field-study-approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP Certificate (PNE 08-070 18) and ZPKCertificate (06012018)
In total 499 samples were collected Subsequently the collected samples were describedaccording to the chosen environmental parameters habitat (1mdashSphagnummat 2mdashsparsemacrophytes 3mdashdense macrophytes 4mdashsandy bottom 5mdashpockets and ponds withinSphagnummat) place (location) (1ndashecotone 2mdashpockets and ponds within the Sphagnummat 3mdashcompacted Sphagnummat) bottom (soil substrate) (1mdashsand 2mdashsand and mud3mdashSphagnum) depth (1mdash0ndash10 cm 2mdash0ndash20 cm 3mdash20ndash40 cm 4mdash40ndash60 cm) nympheids(1mdashnone 2mdashpresent) elodeids (1mdashnone 2mdashpresent) (detritus (1mdashnone 2mdashscarce andfine 3mdashabundant and fine 4mdashabundant and coarse) debris (1 ndashnone 2mdashpresent) fractaldimension (1ndash5) stagemdashseral stage of succession (1ndash40)
Ecological and statistical analysesSpecies diversity was calculated using Smdashnumber of species Nmdashnumber of individualsand Dmdashpercent share All collected beetles were divided into five classes of different bodysize (where a body size meant the total length of an organisms) ie 1mdashvery small beetles(lt30 mm) 2mdashsmall beetles (30ndash50 mm) 3mdashmedium beetles (51ndash100 mm) 4mdashlargebeetles (101ndash200 mm) and 5mdashvery large beetles (gt200 mm) In addition the bodyweight was measured within each of the five classes The ranges of body weight values ofbeetles in the distinguished body size classes are shown in Table 1 Three functional groupswere distinguished in the trophic structure of beetles predators (families Gyrinidae
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 526
Figure 2 Distinguished habitats in the littoral zone of lakes (A) and hypothetical fractal dimensions(B) (after Tokeshi amp Arakaki (2012) modified)
Full-size DOI 107717peerj5662fig-2
Table 1 General description of the lake beetles
Body size class Abundance Number of species Body size (mm) Body weight (mg)
(meanplusmn SD) minndashmax meanplusmn (SD) minndashmax
Very small 2953 32 225plusmn 038 130ndash295 589plusmn 041 49ndash66Small 5640 37 347plusmn 056 300ndash490 790plusmn 078 710ndash889Medium 945 26 582plusmn 112 500ndash980 3828plusmn 376 2580ndash4216Large 565 24 1306plusmn 237 105ndash186 23426plusmn 8316 19028ndash31526Very large 36 5 3179plusmn 209 275ndash351 222802plusmn 1804 21302ndash29501
NotesMin minimum value max maximum value mean average value SD standard deviation
Dytiscidae and Noteridae) polyphages (Haliplidae) and saprophages (HelophoridaeHydrophilidae and Hydraenidae) (Appendix S2) Collected adult water beetles wereidentified using standard identification keys (Galewski 1990 Galewski amp Tranda 1978Friday 1988 Hansen 1987 Holmen 1987)
Because the samples of water beetle fauna were collected several times (taking intoaccount the phenological aspect) from the same lakes and from the habitats distinguishedwithin these lakes we used a GLM (Generalized Linear Model) for repeated measuresANOVA (Hocking 1996) to determine the significance of differences in the number
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 626
of beetles within each functional group in the distinguished habitats and to identifydependencies between the type of habitats body size abundance and biodiversity ofColeoptera First we checked the assumptions of normality (the ShapirondashWilk test) andhomogeneity of variances (the Levenersquos test) respectively The GLM repeated measuremodels were calculated on the basis of Type III sums of squares so as to take the unbalanceddesign into account Significant results were tested for pair-wise comparisons by theTukeyrsquos HSD post-hoc tests Dependent variables (abundance and number of species) weretransformed where necessary to fulfil the requirements of parametric tests (Saint-Germainet al 2007 Cremona Planas amp Lucotte 2008)
In four separate analyses we calculated the Pearsonrsquos correlation coefficients (rp) in orderto determine correlations between (1) body size and counts of beetles in the distinguishedhabitats (2) cover of the Sphagnum mat and counts of beetles (3) cover of the Sphagnummat and functional groups and (4) species and environmental variables
Linear regression analysis was performed to determine the influence of (1) the body sizeon the abundance of beetles (2) fractal dimension on the mean body size on (3) fractaldimension on the mean weight and (4) the seral stage of succession on the mean body sizeMultidimensional correspondence analysis (MCA) (Clausen 1998) served to determinedependencies between the abundance of water beetles within the identified body sizeclasses the share of the distinguished habitats in the littoral zones and in the analysed lakesMCA has been used in similar studies eg Usseglio-Polatera Bournaud amp Tachet 2000Obolewski Glińska-Lewczuk amp Kobus 2009 The average density of body size classes in asample was adjusted by the value corresponding to the total contribution of a given littoralzone type in individual lakes thus giving the weighted average The analysis included twodimensions of which the first explained the biggest part of the general chi-squared statistics( of inertia) whereas the inclusion of the other dimension increased the percentage ofexplained inertia Similarities in the fauna between particular lakes were also analysed bythe single linkage method for the hierarchical clustering of objects The distance measureis an Euclidean distance The results are presented in the form of a dendrogram drawn for40 lakes
Relationships between the presence of beetles and the analysed environmentalparameters in individual lakes were determined with the help of a principal componentanalysis PCA All calculations were performed in Statistica ver 130 (StatSoft Tulsa USA)
RESULTSGeneral description of the collected materialThe collected material comprised 10139 specimens representing 124 species classified toseven families (Gyrinidae Haliplidae Noteridae Dytiscidae Hydraenidae Hydrochidaeand Hydrophilidae) (Appendix S1) Most species (556) belonged to the two smallestbody size classes In total they composed 8475 of all collected specimens The remainingbeetles represented the consecutively higher body size classes of which the largest onehad the fewest representatives (Table 1) Most beetles (288 of the entire material) werecollected in pockets and ponds of the Sphagnum mat It was also there that the highest
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 726
Figure 3 The structure of body size of Coleoptera in distinguished habitatsN abundance S numberof species
Full-size DOI 107717peerj5662fig-3
species richness was determined (91 spp) The species diversity was lower in the densemacrophytes zone (266 74 spp) sparsemacrophytes zone (1964 66 spp) compactedSphagnum mat (189 64 spp) and finally in the sandy bottom (59 39 spp) Verysmall beetles which represented the first body size class demonstrated the highest speciesrichness in the compacted Sphagnummat (415 of the whole collected material) (Fig 3)while being the most numerous in the dense macrophyte zone Small beetles (class 2 of thebody size) were most numerously caught in the pockets and ponds within the Sphagnummat (734) The presence of very large beetles was notable in the sparse macrophyteszone pockets and ponds in the Sphagnum mat and in the sandy bottom zone althoughthey made up a small share of the collected material (around 1)
In the trophic structure the highest percentage (719) was achieved by predatorybeetles represented mainly by Dytiscidae (Fig 4) Although their species diversity wassimilar in all the habitats they were most numerous in the dense macropytes zone (913)and in the sandy bottom zone (845) Most saprophages (472) were noted in thecompacted Sphagnum mat while the number of species representing polyphages waslow in all of the habitats with the highest score (six spp) determined in the compactedSphagnum mat The GLM Repeated Measure Anova results (F(2499)= 674 p= 00089)indicate significant differences in counts of the distinguished trophic groups in theparticular habitats The counts of predatory beetles in the habitats in question did notdiffer significantly (the Tukeyrsquos HSD test p= 018) Conversely differences in the countsof predators and polyphages were significant (p= 00003)
Dependencies between types of habitats body size and abundanceas well as biodiversity of ColeopteraThe results of the GLM repeated measure Anova analysis point to a significant influence ofthe synergistic effect between the type of habitat and body size class on the abundance ofbeetles (F(161996)= 725 p= 00000002) Significant differences (the Tukeyrsquos HSD post -hoc
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 826
Figure 4 The trophic structure of Coleoptera in distinguished habitatsN abundance S number ofspecies
Full-size DOI 107717peerj5662fig-4
test) between the analysed subclasses are illustrated in Fig 5A (cf Appendix S3) Moreovera significant influence of the synergistic effect was determined between the indicated factors(habitat and body size class) and the number of species (F(161996)= 1622 p= 000002)(Appendix S3) Results of the GLM repeated measure also point to a significant influenceof the type of habitat on the number of beetles F(4499) = 675 p= 000003 (Fig 5B)Significant differences (the Tukeyrsquos HSD post -hoc test) were determined between thecompacted Sphagnum mat and sparse macrophytes zone (p= 0031) dense macrophyteszone and sparse macrophytes zone (p= 000014) dense macrophytes zone and pockets(p= 00479) Moreover significant differences were demonstrated in the numbers ofbeetles representing the distinguished body size classes (F(41996)= 7761 p= 000002) TheTukeyrsquos post-hoc test reveals significant differences in the counts of beetles representing allthe classes of body size (p= 000002) except 3 and 4 class (p= 077) Both factors the typeof habitat (F(4499)= 1104 p= 0000003) and body size (F(41996)= 37561 p= 0000001)also had a significant impact on the number of species determined With respect to thespecies diversity significant differences were found between the Sphagnummat and sparsemacrophytes (p= 00048) and between the pockets and ponds in the Sphagnum mat andsandy bottom zone (p= 00046) and between the Sphagnummat and sparse macrophytes(p= 000018) and dense macrophytes (p= 00014) Significant differences in the numberof species were shown between all body size classes (p= 000002) except 1 and 3 (p= 024)
Negative correlations were observed in all the habitats between the body size and thenumber of beetles It was only in the sandy bottom zone that these correlations were notsignificant (p= 047) (Table 2) Similarly negative correlations appeared between the bodysize and species richness in the compacted Sphagnum mat (p= 001) and in the sparsemacrophytes zone (p= 00008) (Table 2)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 926
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

sp Sparganium sp or Acorus sp Many hydrobiologists emphasize the important role ofthe littoral zone in the secondary production of lakes and as a zone having the highestspecies richness and density of macroinvertebrates (Cremona Planas amp Lucotte 2008Timm ampMoumlls 2012Mieczan et al 2014 Płaska amp Mieczan 2018) The littoral zone is alsoconsidered to be the most sensitive part of a lake and its character provides evidence onthe ecological condition of the whole lake (Czachorowski 1998 Šiling amp Urbanič 2016)
The succession of lakes entails changes in the lake-dwelling fauna (Kowalik 1968Pakulnicka amp Bartnik 1999 Ranta 1985 Kordylas 1990 Lundkvist Landin amp Milberg2001 Pakulnicka amp Zawal 2007) This problem is rarely raised in research and when itis it most often consists of brief studies into zooplankton (eg Demetraki-Paleolog 2012Kuczyńska-Kippen 2008 Jasser 1997 Lepistouml amp Saura 1998 Odland amp Del Moral 2002Fiłoc amp Kupryjanowicz 2015) More detailed investigations pertaining to changes in lakefauna during consecutive seral stages are conducted on anthropogenic rather than naturalecosystems (Pakulnicka 2008 Bloechl et al 2010 Buczyński 2015 Pakulnicka Goacuterski ampBielecki 2015b) Hence despite the wealth of references our knowledge of the successionmechanisms in lake fauna ismodest and fragmentary even though it is extremely importantfor developing proper lake management practice (Šiling amp Urbanič 2016 Shadrin et al2016)
Another challenge for researchers is to find an adequate measure for the determinationof changes in fauna induced by the succession of water bodies Hydrobiologists mostoften draw attention to changes in the abundance and species richness while comparingsmall groups of lakes with different trophic states (Kowalik 1968 Ranta 1985 Kordylas1990 Lundkvist Landin amp Milberg 2001 Pakulnicka amp Zawal 2007 Soldaacuten et al 2012Barndt 2012 Drinan et al 2013 Pakulnicka et al 2013 Baars et al 2014 Beadle Brownamp Holden 2015) A measure that has been gaining popularity over the last twenty yearsconsists of an analysis of quantitative relations between components (generalists anddiversified specialists) distinguished on the basis of their affinity towards specific habitatconditions (eg Kowalik 1968 Kordylas 1990 Czachorowski 1998 Pakulnicka amp Zawal2007 Płaska amp Tarkowska-Kukuryk 2014 Pakulnicka et al 2016a Pakulnicka et al 2016bPłaska et al 2016 Stryjecki et al 2017) However the results obtained from this approachare discrepant and therefore not highly reliable often because of a small number of samplesor analysed objects
An important addition to research consists of analyses based onbiometricmeasurementsfound also in studies on small water organisms including beetles (Jeffries 1993McAbendroth et al 2005 Ulrich 2007 Vamosi Naydani amp Vamosi 2007 Cremona Planasamp Lucotte 2008 Scheffer et al 2015 Scheffer amp Van Nes 2006 Tokeshi amp Arakaki 2012Deacutesamoreacute et al 2018) In recent years biometric measures have become a tool applied inevolutionary ecology Some hydrobiologists eg Scheffer amp Van Nes (2006) and Schefferet al (2015) analyse the structure of body sizes of organisms in specific communities fromthe viewpoint of coevolution of concurrent species Others eg Deacutesamoreacute et al (2018)search for relationships between the evolution of a body size and the environment as wellas the diversification of species in various water habitats
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 226
Data regarding the body mass and body size of macroinvertebrates often appear in thecontext of studies on a fractal structure a notion which still awaits a better understanding(Jeffries 1993 Tokeshi amp Arakaki 2012 Barnes Vaughan amp Ormerod 2013) and a fractaldimension a borrowing from mathematical sciences (Mandelbrot 1983) According toAndrejczuk (2014) all landscapes with inner diversity demonstrate fractality ie they arecomposed of smaller fragments self-similar fractals which are self-reproducing duplicatorsof parameters on a different scale Thus a fragment of any system should contain all thesystemrsquos characteristics in a nutshell (on a smaller scale) The littoral zone of ecologicallyyoung lakes can serve as an example of a linear structure of spatial organisation whichdemonstrates properties of fractality However as the succession progresses the shorelinebecomes more diverse and turns into a multifractal system ie heterogeneous composedof fragments (separate subsets) with different local characteristics each presenting self-similar properties (Miałdun 2010 Miałdun amp Ostrowski 2010) Thus similar fragments(fractals) of the littoral zone of different objects should be characterised by similar habitattraits offering the same niches to the organisms which populate them A convenient andvaluable object in studies on the fractal structure are dystrophic lakes in which varioushabitats (fragments of the littoral zone) appear during the course of succession from aplant-free zone to zones overgrown with macrophyte communities of various compactnessto a compact Sphagnum mat with small pools
In our studywe looked at water beetles because they are particularly numerous organismsin the littoral zone as well as being very sensitive and responsive to any unfavourable changesin the environment (Eyre Foster amp Foster 1992 Foster amp Eyre 1992 Corbet et al 1980Winfield Fairchild Faulds amp Matta 2000 Bosi 2001 Menetrey et al 2005 McFarlandCarse amp Sandin 2009 Gioria Bacaro amp Feehan 2010a Gioria et al 2010b Yee 2014)Furthermore many species are predators which defines their important role as organismsregulating the abundance and species richness of concurrent taxa (Yee 2014 Frelik ampPakulnicka 2015 Frelik amp Pakulnicka 2015 Perissinotto Bird amp Bilton 2016 Miller ampBergsten 2016 Płaska amp Mieczan 2018) Water beetles penetrate both the ground andthe water column so according to McAbendroth et al (2005) their ecological niche isthree-dimensional Consequently the fractal dimension of the habitats they occupy iswithin the range of 2ltDlt 3 (increasing from the least to the most compact habitats)(Cremona Planas amp Lucotte 2008 Tokeshi amp Arakaki 2012)
In the light of the above considerations our aim has been to investigate whether (1)the distinguished habitats are populated by similar assemblages of organisms with respectto body size and trophic preferences (2) if there is a relationship between the body massand abundance of water beetles (3) if the fractal dimension of a habitat has an effect onthe total biomass of beetles and whether the total biomass of beetles in particular habitatsis the same and finally (4) if there are changes in the sequence of body size of organismsoccurring in the course of disharmonic succession
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 326
Figure 1 Study area Location of lakes (1ndash40) 1 Żabie 2 Szare 3 Wygoda 4 Krypko 5 Pałsznik6 Małe Gacno 7 Wielkie Gacno 8 Długie 9 Moczadło 10 Sosnoacutewek 11 Żabionek 12 Klimontek13 Nierybno 14 Małe Łowne 15 Czarne 16 Piecki 17 Babionek Duży 18 Babionek Mały 19 Białe20 Kociołek 21 Żabie 22 Motylek 23 Purdka 24 Jonkowo 25 Borkowskie 26 Bobroacutewko 27Gryżewskie 28 Skarp 29 Zakręt 30 Kruczy Stawek 31 Kruczek Duży 32 Kruczek Mały 33 KruczeOko 34 Kruczy Staw 35 Suchar Wielki 36 Suchar 1 Lake 37 Suchar 2 38 Suchar 3 39 Suchar 4 40Suchar 5
Full-size DOI 107717peerj5662fig-1
MATERIAL amp METHODSStudy area and field studiesThe study covered 40 dystrophic lakes located in northern Poland the South BalticCoastland the West Pomeranian Olsztyn Mrągowo Lakelands and the Suwałki Lakeland(Fig 1 Appendix S1) The lakes chosen for the study were of various surface areas andhaving a floating peat mat growth belonging to Sphagnum sp that varied in size The lakesrepresented various succession stages in a disharmonic seriesmdashfrom oligo- to polyhumiclakes The oligohumic lakes were inhabited by Juncus bulbosus Eleocharis palustrisPhragmites australis Typha angustifolia Typha latiffolia Sphagnum sp Lobelia dortmannaIsoeumltes lacustris Drosera rotundifolia D anglica D intermedia Sparganium angustifoliumLycopodiella inundata In polyhumic lakes the dominant species included Sphagnum spOxycoccus quadripelatus Andromeda polifonia Eriophorum vaginatum Calluna vulgarisErica tetralix Empetrum nigrum The lakes were arranged a priori according to the seralstages of succession (Bloechl et al 2010) and the point of reference was assumed to be the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 426
coverage of a lakersquos surface by a Sphagnum mat (0ndash75) (Appendix S1) The percentagecover by a Sphagnum mat in each lake was calculated in the GIS system supported byArcMap 931 software For charting the research sites data made available throughGeoportal 2 in the WMS format were employed
The study was carried out from 2002 to 2014 in spring summer and autumn The faunasamples were collected with a dip net from a surface area of about 1 m2 In the compactedSphagnum mat environment a sample comprised 10 subsamples (aggregated afterwards)which were collected using a 01 m2 strainer Sampling sites were chosen so as to representthe biggest array of the littoral habitat diversity and areas of individual lakes Thus fivedifferent littoral components (habitats) were identified (1) the Sphagnum mat (2) sparsemacrophyte zone (3) dense macrophyte zone (4) sandy bottom zone and (5) pockets andponds within a Sphagnum mat The vegetation cover was assessed using Braun-Blanquet(1964) phytosociological records All lakes were identified with respect to the surface areaextent of the lakersquos surface covered with a Sphagnummat which corresponds to successionstages (1ndash3) percent share of individual habitats in the littoral zone (1ndash4) and a seralstage (1ndash40) (Appendix S1) It was assumed that these habitats due to different degreesof complexity represent different fractal dimensions all within an interval of 20 gtD gt30(Tokeshi amp Arakaki 2012) A schematic representation of fractals different in size can beseen in Fig 2 The smallest size (1) corresponds to the sandy bottom zone while the largestone (5) is assigned to the Sphagnummat The intermediate sizes belong to (2) pockets andponds within the Sphagnum mat (3) sparse macrophyte zone and (4) dense macrophytezone Field studies were conducted in accordance with field-study-approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP Certificate (PNE 08-070 18) and ZPKCertificate (06012018)
In total 499 samples were collected Subsequently the collected samples were describedaccording to the chosen environmental parameters habitat (1mdashSphagnummat 2mdashsparsemacrophytes 3mdashdense macrophytes 4mdashsandy bottom 5mdashpockets and ponds withinSphagnummat) place (location) (1ndashecotone 2mdashpockets and ponds within the Sphagnummat 3mdashcompacted Sphagnummat) bottom (soil substrate) (1mdashsand 2mdashsand and mud3mdashSphagnum) depth (1mdash0ndash10 cm 2mdash0ndash20 cm 3mdash20ndash40 cm 4mdash40ndash60 cm) nympheids(1mdashnone 2mdashpresent) elodeids (1mdashnone 2mdashpresent) (detritus (1mdashnone 2mdashscarce andfine 3mdashabundant and fine 4mdashabundant and coarse) debris (1 ndashnone 2mdashpresent) fractaldimension (1ndash5) stagemdashseral stage of succession (1ndash40)
Ecological and statistical analysesSpecies diversity was calculated using Smdashnumber of species Nmdashnumber of individualsand Dmdashpercent share All collected beetles were divided into five classes of different bodysize (where a body size meant the total length of an organisms) ie 1mdashvery small beetles(lt30 mm) 2mdashsmall beetles (30ndash50 mm) 3mdashmedium beetles (51ndash100 mm) 4mdashlargebeetles (101ndash200 mm) and 5mdashvery large beetles (gt200 mm) In addition the bodyweight was measured within each of the five classes The ranges of body weight values ofbeetles in the distinguished body size classes are shown in Table 1 Three functional groupswere distinguished in the trophic structure of beetles predators (families Gyrinidae
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 526
Figure 2 Distinguished habitats in the littoral zone of lakes (A) and hypothetical fractal dimensions(B) (after Tokeshi amp Arakaki (2012) modified)
Full-size DOI 107717peerj5662fig-2
Table 1 General description of the lake beetles
Body size class Abundance Number of species Body size (mm) Body weight (mg)
(meanplusmn SD) minndashmax meanplusmn (SD) minndashmax
Very small 2953 32 225plusmn 038 130ndash295 589plusmn 041 49ndash66Small 5640 37 347plusmn 056 300ndash490 790plusmn 078 710ndash889Medium 945 26 582plusmn 112 500ndash980 3828plusmn 376 2580ndash4216Large 565 24 1306plusmn 237 105ndash186 23426plusmn 8316 19028ndash31526Very large 36 5 3179plusmn 209 275ndash351 222802plusmn 1804 21302ndash29501
NotesMin minimum value max maximum value mean average value SD standard deviation
Dytiscidae and Noteridae) polyphages (Haliplidae) and saprophages (HelophoridaeHydrophilidae and Hydraenidae) (Appendix S2) Collected adult water beetles wereidentified using standard identification keys (Galewski 1990 Galewski amp Tranda 1978Friday 1988 Hansen 1987 Holmen 1987)
Because the samples of water beetle fauna were collected several times (taking intoaccount the phenological aspect) from the same lakes and from the habitats distinguishedwithin these lakes we used a GLM (Generalized Linear Model) for repeated measuresANOVA (Hocking 1996) to determine the significance of differences in the number
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 626
of beetles within each functional group in the distinguished habitats and to identifydependencies between the type of habitats body size abundance and biodiversity ofColeoptera First we checked the assumptions of normality (the ShapirondashWilk test) andhomogeneity of variances (the Levenersquos test) respectively The GLM repeated measuremodels were calculated on the basis of Type III sums of squares so as to take the unbalanceddesign into account Significant results were tested for pair-wise comparisons by theTukeyrsquos HSD post-hoc tests Dependent variables (abundance and number of species) weretransformed where necessary to fulfil the requirements of parametric tests (Saint-Germainet al 2007 Cremona Planas amp Lucotte 2008)
In four separate analyses we calculated the Pearsonrsquos correlation coefficients (rp) in orderto determine correlations between (1) body size and counts of beetles in the distinguishedhabitats (2) cover of the Sphagnum mat and counts of beetles (3) cover of the Sphagnummat and functional groups and (4) species and environmental variables
Linear regression analysis was performed to determine the influence of (1) the body sizeon the abundance of beetles (2) fractal dimension on the mean body size on (3) fractaldimension on the mean weight and (4) the seral stage of succession on the mean body sizeMultidimensional correspondence analysis (MCA) (Clausen 1998) served to determinedependencies between the abundance of water beetles within the identified body sizeclasses the share of the distinguished habitats in the littoral zones and in the analysed lakesMCA has been used in similar studies eg Usseglio-Polatera Bournaud amp Tachet 2000Obolewski Glińska-Lewczuk amp Kobus 2009 The average density of body size classes in asample was adjusted by the value corresponding to the total contribution of a given littoralzone type in individual lakes thus giving the weighted average The analysis included twodimensions of which the first explained the biggest part of the general chi-squared statistics( of inertia) whereas the inclusion of the other dimension increased the percentage ofexplained inertia Similarities in the fauna between particular lakes were also analysed bythe single linkage method for the hierarchical clustering of objects The distance measureis an Euclidean distance The results are presented in the form of a dendrogram drawn for40 lakes
Relationships between the presence of beetles and the analysed environmentalparameters in individual lakes were determined with the help of a principal componentanalysis PCA All calculations were performed in Statistica ver 130 (StatSoft Tulsa USA)
RESULTSGeneral description of the collected materialThe collected material comprised 10139 specimens representing 124 species classified toseven families (Gyrinidae Haliplidae Noteridae Dytiscidae Hydraenidae Hydrochidaeand Hydrophilidae) (Appendix S1) Most species (556) belonged to the two smallestbody size classes In total they composed 8475 of all collected specimens The remainingbeetles represented the consecutively higher body size classes of which the largest onehad the fewest representatives (Table 1) Most beetles (288 of the entire material) werecollected in pockets and ponds of the Sphagnum mat It was also there that the highest
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 726
Figure 3 The structure of body size of Coleoptera in distinguished habitatsN abundance S numberof species
Full-size DOI 107717peerj5662fig-3
species richness was determined (91 spp) The species diversity was lower in the densemacrophytes zone (266 74 spp) sparsemacrophytes zone (1964 66 spp) compactedSphagnum mat (189 64 spp) and finally in the sandy bottom (59 39 spp) Verysmall beetles which represented the first body size class demonstrated the highest speciesrichness in the compacted Sphagnummat (415 of the whole collected material) (Fig 3)while being the most numerous in the dense macrophyte zone Small beetles (class 2 of thebody size) were most numerously caught in the pockets and ponds within the Sphagnummat (734) The presence of very large beetles was notable in the sparse macrophyteszone pockets and ponds in the Sphagnum mat and in the sandy bottom zone althoughthey made up a small share of the collected material (around 1)
In the trophic structure the highest percentage (719) was achieved by predatorybeetles represented mainly by Dytiscidae (Fig 4) Although their species diversity wassimilar in all the habitats they were most numerous in the dense macropytes zone (913)and in the sandy bottom zone (845) Most saprophages (472) were noted in thecompacted Sphagnum mat while the number of species representing polyphages waslow in all of the habitats with the highest score (six spp) determined in the compactedSphagnum mat The GLM Repeated Measure Anova results (F(2499)= 674 p= 00089)indicate significant differences in counts of the distinguished trophic groups in theparticular habitats The counts of predatory beetles in the habitats in question did notdiffer significantly (the Tukeyrsquos HSD test p= 018) Conversely differences in the countsof predators and polyphages were significant (p= 00003)
Dependencies between types of habitats body size and abundanceas well as biodiversity of ColeopteraThe results of the GLM repeated measure Anova analysis point to a significant influence ofthe synergistic effect between the type of habitat and body size class on the abundance ofbeetles (F(161996)= 725 p= 00000002) Significant differences (the Tukeyrsquos HSD post -hoc
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 826
Figure 4 The trophic structure of Coleoptera in distinguished habitatsN abundance S number ofspecies
Full-size DOI 107717peerj5662fig-4
test) between the analysed subclasses are illustrated in Fig 5A (cf Appendix S3) Moreovera significant influence of the synergistic effect was determined between the indicated factors(habitat and body size class) and the number of species (F(161996)= 1622 p= 000002)(Appendix S3) Results of the GLM repeated measure also point to a significant influenceof the type of habitat on the number of beetles F(4499) = 675 p= 000003 (Fig 5B)Significant differences (the Tukeyrsquos HSD post -hoc test) were determined between thecompacted Sphagnum mat and sparse macrophytes zone (p= 0031) dense macrophyteszone and sparse macrophytes zone (p= 000014) dense macrophytes zone and pockets(p= 00479) Moreover significant differences were demonstrated in the numbers ofbeetles representing the distinguished body size classes (F(41996)= 7761 p= 000002) TheTukeyrsquos post-hoc test reveals significant differences in the counts of beetles representing allthe classes of body size (p= 000002) except 3 and 4 class (p= 077) Both factors the typeof habitat (F(4499)= 1104 p= 0000003) and body size (F(41996)= 37561 p= 0000001)also had a significant impact on the number of species determined With respect to thespecies diversity significant differences were found between the Sphagnummat and sparsemacrophytes (p= 00048) and between the pockets and ponds in the Sphagnum mat andsandy bottom zone (p= 00046) and between the Sphagnummat and sparse macrophytes(p= 000018) and dense macrophytes (p= 00014) Significant differences in the numberof species were shown between all body size classes (p= 000002) except 1 and 3 (p= 024)
Negative correlations were observed in all the habitats between the body size and thenumber of beetles It was only in the sandy bottom zone that these correlations were notsignificant (p= 047) (Table 2) Similarly negative correlations appeared between the bodysize and species richness in the compacted Sphagnum mat (p= 001) and in the sparsemacrophytes zone (p= 00008) (Table 2)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 926
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

Data regarding the body mass and body size of macroinvertebrates often appear in thecontext of studies on a fractal structure a notion which still awaits a better understanding(Jeffries 1993 Tokeshi amp Arakaki 2012 Barnes Vaughan amp Ormerod 2013) and a fractaldimension a borrowing from mathematical sciences (Mandelbrot 1983) According toAndrejczuk (2014) all landscapes with inner diversity demonstrate fractality ie they arecomposed of smaller fragments self-similar fractals which are self-reproducing duplicatorsof parameters on a different scale Thus a fragment of any system should contain all thesystemrsquos characteristics in a nutshell (on a smaller scale) The littoral zone of ecologicallyyoung lakes can serve as an example of a linear structure of spatial organisation whichdemonstrates properties of fractality However as the succession progresses the shorelinebecomes more diverse and turns into a multifractal system ie heterogeneous composedof fragments (separate subsets) with different local characteristics each presenting self-similar properties (Miałdun 2010 Miałdun amp Ostrowski 2010) Thus similar fragments(fractals) of the littoral zone of different objects should be characterised by similar habitattraits offering the same niches to the organisms which populate them A convenient andvaluable object in studies on the fractal structure are dystrophic lakes in which varioushabitats (fragments of the littoral zone) appear during the course of succession from aplant-free zone to zones overgrown with macrophyte communities of various compactnessto a compact Sphagnum mat with small pools
In our studywe looked at water beetles because they are particularly numerous organismsin the littoral zone as well as being very sensitive and responsive to any unfavourable changesin the environment (Eyre Foster amp Foster 1992 Foster amp Eyre 1992 Corbet et al 1980Winfield Fairchild Faulds amp Matta 2000 Bosi 2001 Menetrey et al 2005 McFarlandCarse amp Sandin 2009 Gioria Bacaro amp Feehan 2010a Gioria et al 2010b Yee 2014)Furthermore many species are predators which defines their important role as organismsregulating the abundance and species richness of concurrent taxa (Yee 2014 Frelik ampPakulnicka 2015 Frelik amp Pakulnicka 2015 Perissinotto Bird amp Bilton 2016 Miller ampBergsten 2016 Płaska amp Mieczan 2018) Water beetles penetrate both the ground andthe water column so according to McAbendroth et al (2005) their ecological niche isthree-dimensional Consequently the fractal dimension of the habitats they occupy iswithin the range of 2ltDlt 3 (increasing from the least to the most compact habitats)(Cremona Planas amp Lucotte 2008 Tokeshi amp Arakaki 2012)
In the light of the above considerations our aim has been to investigate whether (1)the distinguished habitats are populated by similar assemblages of organisms with respectto body size and trophic preferences (2) if there is a relationship between the body massand abundance of water beetles (3) if the fractal dimension of a habitat has an effect onthe total biomass of beetles and whether the total biomass of beetles in particular habitatsis the same and finally (4) if there are changes in the sequence of body size of organismsoccurring in the course of disharmonic succession
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 326
Figure 1 Study area Location of lakes (1ndash40) 1 Żabie 2 Szare 3 Wygoda 4 Krypko 5 Pałsznik6 Małe Gacno 7 Wielkie Gacno 8 Długie 9 Moczadło 10 Sosnoacutewek 11 Żabionek 12 Klimontek13 Nierybno 14 Małe Łowne 15 Czarne 16 Piecki 17 Babionek Duży 18 Babionek Mały 19 Białe20 Kociołek 21 Żabie 22 Motylek 23 Purdka 24 Jonkowo 25 Borkowskie 26 Bobroacutewko 27Gryżewskie 28 Skarp 29 Zakręt 30 Kruczy Stawek 31 Kruczek Duży 32 Kruczek Mały 33 KruczeOko 34 Kruczy Staw 35 Suchar Wielki 36 Suchar 1 Lake 37 Suchar 2 38 Suchar 3 39 Suchar 4 40Suchar 5
Full-size DOI 107717peerj5662fig-1
MATERIAL amp METHODSStudy area and field studiesThe study covered 40 dystrophic lakes located in northern Poland the South BalticCoastland the West Pomeranian Olsztyn Mrągowo Lakelands and the Suwałki Lakeland(Fig 1 Appendix S1) The lakes chosen for the study were of various surface areas andhaving a floating peat mat growth belonging to Sphagnum sp that varied in size The lakesrepresented various succession stages in a disharmonic seriesmdashfrom oligo- to polyhumiclakes The oligohumic lakes were inhabited by Juncus bulbosus Eleocharis palustrisPhragmites australis Typha angustifolia Typha latiffolia Sphagnum sp Lobelia dortmannaIsoeumltes lacustris Drosera rotundifolia D anglica D intermedia Sparganium angustifoliumLycopodiella inundata In polyhumic lakes the dominant species included Sphagnum spOxycoccus quadripelatus Andromeda polifonia Eriophorum vaginatum Calluna vulgarisErica tetralix Empetrum nigrum The lakes were arranged a priori according to the seralstages of succession (Bloechl et al 2010) and the point of reference was assumed to be the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 426
coverage of a lakersquos surface by a Sphagnum mat (0ndash75) (Appendix S1) The percentagecover by a Sphagnum mat in each lake was calculated in the GIS system supported byArcMap 931 software For charting the research sites data made available throughGeoportal 2 in the WMS format were employed
The study was carried out from 2002 to 2014 in spring summer and autumn The faunasamples were collected with a dip net from a surface area of about 1 m2 In the compactedSphagnum mat environment a sample comprised 10 subsamples (aggregated afterwards)which were collected using a 01 m2 strainer Sampling sites were chosen so as to representthe biggest array of the littoral habitat diversity and areas of individual lakes Thus fivedifferent littoral components (habitats) were identified (1) the Sphagnum mat (2) sparsemacrophyte zone (3) dense macrophyte zone (4) sandy bottom zone and (5) pockets andponds within a Sphagnum mat The vegetation cover was assessed using Braun-Blanquet(1964) phytosociological records All lakes were identified with respect to the surface areaextent of the lakersquos surface covered with a Sphagnummat which corresponds to successionstages (1ndash3) percent share of individual habitats in the littoral zone (1ndash4) and a seralstage (1ndash40) (Appendix S1) It was assumed that these habitats due to different degreesof complexity represent different fractal dimensions all within an interval of 20 gtD gt30(Tokeshi amp Arakaki 2012) A schematic representation of fractals different in size can beseen in Fig 2 The smallest size (1) corresponds to the sandy bottom zone while the largestone (5) is assigned to the Sphagnummat The intermediate sizes belong to (2) pockets andponds within the Sphagnum mat (3) sparse macrophyte zone and (4) dense macrophytezone Field studies were conducted in accordance with field-study-approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP Certificate (PNE 08-070 18) and ZPKCertificate (06012018)
In total 499 samples were collected Subsequently the collected samples were describedaccording to the chosen environmental parameters habitat (1mdashSphagnummat 2mdashsparsemacrophytes 3mdashdense macrophytes 4mdashsandy bottom 5mdashpockets and ponds withinSphagnummat) place (location) (1ndashecotone 2mdashpockets and ponds within the Sphagnummat 3mdashcompacted Sphagnummat) bottom (soil substrate) (1mdashsand 2mdashsand and mud3mdashSphagnum) depth (1mdash0ndash10 cm 2mdash0ndash20 cm 3mdash20ndash40 cm 4mdash40ndash60 cm) nympheids(1mdashnone 2mdashpresent) elodeids (1mdashnone 2mdashpresent) (detritus (1mdashnone 2mdashscarce andfine 3mdashabundant and fine 4mdashabundant and coarse) debris (1 ndashnone 2mdashpresent) fractaldimension (1ndash5) stagemdashseral stage of succession (1ndash40)
Ecological and statistical analysesSpecies diversity was calculated using Smdashnumber of species Nmdashnumber of individualsand Dmdashpercent share All collected beetles were divided into five classes of different bodysize (where a body size meant the total length of an organisms) ie 1mdashvery small beetles(lt30 mm) 2mdashsmall beetles (30ndash50 mm) 3mdashmedium beetles (51ndash100 mm) 4mdashlargebeetles (101ndash200 mm) and 5mdashvery large beetles (gt200 mm) In addition the bodyweight was measured within each of the five classes The ranges of body weight values ofbeetles in the distinguished body size classes are shown in Table 1 Three functional groupswere distinguished in the trophic structure of beetles predators (families Gyrinidae
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 526
Figure 2 Distinguished habitats in the littoral zone of lakes (A) and hypothetical fractal dimensions(B) (after Tokeshi amp Arakaki (2012) modified)
Full-size DOI 107717peerj5662fig-2
Table 1 General description of the lake beetles
Body size class Abundance Number of species Body size (mm) Body weight (mg)
(meanplusmn SD) minndashmax meanplusmn (SD) minndashmax
Very small 2953 32 225plusmn 038 130ndash295 589plusmn 041 49ndash66Small 5640 37 347plusmn 056 300ndash490 790plusmn 078 710ndash889Medium 945 26 582plusmn 112 500ndash980 3828plusmn 376 2580ndash4216Large 565 24 1306plusmn 237 105ndash186 23426plusmn 8316 19028ndash31526Very large 36 5 3179plusmn 209 275ndash351 222802plusmn 1804 21302ndash29501
NotesMin minimum value max maximum value mean average value SD standard deviation
Dytiscidae and Noteridae) polyphages (Haliplidae) and saprophages (HelophoridaeHydrophilidae and Hydraenidae) (Appendix S2) Collected adult water beetles wereidentified using standard identification keys (Galewski 1990 Galewski amp Tranda 1978Friday 1988 Hansen 1987 Holmen 1987)
Because the samples of water beetle fauna were collected several times (taking intoaccount the phenological aspect) from the same lakes and from the habitats distinguishedwithin these lakes we used a GLM (Generalized Linear Model) for repeated measuresANOVA (Hocking 1996) to determine the significance of differences in the number
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 626
of beetles within each functional group in the distinguished habitats and to identifydependencies between the type of habitats body size abundance and biodiversity ofColeoptera First we checked the assumptions of normality (the ShapirondashWilk test) andhomogeneity of variances (the Levenersquos test) respectively The GLM repeated measuremodels were calculated on the basis of Type III sums of squares so as to take the unbalanceddesign into account Significant results were tested for pair-wise comparisons by theTukeyrsquos HSD post-hoc tests Dependent variables (abundance and number of species) weretransformed where necessary to fulfil the requirements of parametric tests (Saint-Germainet al 2007 Cremona Planas amp Lucotte 2008)
In four separate analyses we calculated the Pearsonrsquos correlation coefficients (rp) in orderto determine correlations between (1) body size and counts of beetles in the distinguishedhabitats (2) cover of the Sphagnum mat and counts of beetles (3) cover of the Sphagnummat and functional groups and (4) species and environmental variables
Linear regression analysis was performed to determine the influence of (1) the body sizeon the abundance of beetles (2) fractal dimension on the mean body size on (3) fractaldimension on the mean weight and (4) the seral stage of succession on the mean body sizeMultidimensional correspondence analysis (MCA) (Clausen 1998) served to determinedependencies between the abundance of water beetles within the identified body sizeclasses the share of the distinguished habitats in the littoral zones and in the analysed lakesMCA has been used in similar studies eg Usseglio-Polatera Bournaud amp Tachet 2000Obolewski Glińska-Lewczuk amp Kobus 2009 The average density of body size classes in asample was adjusted by the value corresponding to the total contribution of a given littoralzone type in individual lakes thus giving the weighted average The analysis included twodimensions of which the first explained the biggest part of the general chi-squared statistics( of inertia) whereas the inclusion of the other dimension increased the percentage ofexplained inertia Similarities in the fauna between particular lakes were also analysed bythe single linkage method for the hierarchical clustering of objects The distance measureis an Euclidean distance The results are presented in the form of a dendrogram drawn for40 lakes
Relationships between the presence of beetles and the analysed environmentalparameters in individual lakes were determined with the help of a principal componentanalysis PCA All calculations were performed in Statistica ver 130 (StatSoft Tulsa USA)
RESULTSGeneral description of the collected materialThe collected material comprised 10139 specimens representing 124 species classified toseven families (Gyrinidae Haliplidae Noteridae Dytiscidae Hydraenidae Hydrochidaeand Hydrophilidae) (Appendix S1) Most species (556) belonged to the two smallestbody size classes In total they composed 8475 of all collected specimens The remainingbeetles represented the consecutively higher body size classes of which the largest onehad the fewest representatives (Table 1) Most beetles (288 of the entire material) werecollected in pockets and ponds of the Sphagnum mat It was also there that the highest
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 726
Figure 3 The structure of body size of Coleoptera in distinguished habitatsN abundance S numberof species
Full-size DOI 107717peerj5662fig-3
species richness was determined (91 spp) The species diversity was lower in the densemacrophytes zone (266 74 spp) sparsemacrophytes zone (1964 66 spp) compactedSphagnum mat (189 64 spp) and finally in the sandy bottom (59 39 spp) Verysmall beetles which represented the first body size class demonstrated the highest speciesrichness in the compacted Sphagnummat (415 of the whole collected material) (Fig 3)while being the most numerous in the dense macrophyte zone Small beetles (class 2 of thebody size) were most numerously caught in the pockets and ponds within the Sphagnummat (734) The presence of very large beetles was notable in the sparse macrophyteszone pockets and ponds in the Sphagnum mat and in the sandy bottom zone althoughthey made up a small share of the collected material (around 1)
In the trophic structure the highest percentage (719) was achieved by predatorybeetles represented mainly by Dytiscidae (Fig 4) Although their species diversity wassimilar in all the habitats they were most numerous in the dense macropytes zone (913)and in the sandy bottom zone (845) Most saprophages (472) were noted in thecompacted Sphagnum mat while the number of species representing polyphages waslow in all of the habitats with the highest score (six spp) determined in the compactedSphagnum mat The GLM Repeated Measure Anova results (F(2499)= 674 p= 00089)indicate significant differences in counts of the distinguished trophic groups in theparticular habitats The counts of predatory beetles in the habitats in question did notdiffer significantly (the Tukeyrsquos HSD test p= 018) Conversely differences in the countsof predators and polyphages were significant (p= 00003)
Dependencies between types of habitats body size and abundanceas well as biodiversity of ColeopteraThe results of the GLM repeated measure Anova analysis point to a significant influence ofthe synergistic effect between the type of habitat and body size class on the abundance ofbeetles (F(161996)= 725 p= 00000002) Significant differences (the Tukeyrsquos HSD post -hoc
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 826
Figure 4 The trophic structure of Coleoptera in distinguished habitatsN abundance S number ofspecies
Full-size DOI 107717peerj5662fig-4
test) between the analysed subclasses are illustrated in Fig 5A (cf Appendix S3) Moreovera significant influence of the synergistic effect was determined between the indicated factors(habitat and body size class) and the number of species (F(161996)= 1622 p= 000002)(Appendix S3) Results of the GLM repeated measure also point to a significant influenceof the type of habitat on the number of beetles F(4499) = 675 p= 000003 (Fig 5B)Significant differences (the Tukeyrsquos HSD post -hoc test) were determined between thecompacted Sphagnum mat and sparse macrophytes zone (p= 0031) dense macrophyteszone and sparse macrophytes zone (p= 000014) dense macrophytes zone and pockets(p= 00479) Moreover significant differences were demonstrated in the numbers ofbeetles representing the distinguished body size classes (F(41996)= 7761 p= 000002) TheTukeyrsquos post-hoc test reveals significant differences in the counts of beetles representing allthe classes of body size (p= 000002) except 3 and 4 class (p= 077) Both factors the typeof habitat (F(4499)= 1104 p= 0000003) and body size (F(41996)= 37561 p= 0000001)also had a significant impact on the number of species determined With respect to thespecies diversity significant differences were found between the Sphagnummat and sparsemacrophytes (p= 00048) and between the pockets and ponds in the Sphagnum mat andsandy bottom zone (p= 00046) and between the Sphagnummat and sparse macrophytes(p= 000018) and dense macrophytes (p= 00014) Significant differences in the numberof species were shown between all body size classes (p= 000002) except 1 and 3 (p= 024)
Negative correlations were observed in all the habitats between the body size and thenumber of beetles It was only in the sandy bottom zone that these correlations were notsignificant (p= 047) (Table 2) Similarly negative correlations appeared between the bodysize and species richness in the compacted Sphagnum mat (p= 001) and in the sparsemacrophytes zone (p= 00008) (Table 2)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 926
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

Figure 1 Study area Location of lakes (1ndash40) 1 Żabie 2 Szare 3 Wygoda 4 Krypko 5 Pałsznik6 Małe Gacno 7 Wielkie Gacno 8 Długie 9 Moczadło 10 Sosnoacutewek 11 Żabionek 12 Klimontek13 Nierybno 14 Małe Łowne 15 Czarne 16 Piecki 17 Babionek Duży 18 Babionek Mały 19 Białe20 Kociołek 21 Żabie 22 Motylek 23 Purdka 24 Jonkowo 25 Borkowskie 26 Bobroacutewko 27Gryżewskie 28 Skarp 29 Zakręt 30 Kruczy Stawek 31 Kruczek Duży 32 Kruczek Mały 33 KruczeOko 34 Kruczy Staw 35 Suchar Wielki 36 Suchar 1 Lake 37 Suchar 2 38 Suchar 3 39 Suchar 4 40Suchar 5
Full-size DOI 107717peerj5662fig-1
MATERIAL amp METHODSStudy area and field studiesThe study covered 40 dystrophic lakes located in northern Poland the South BalticCoastland the West Pomeranian Olsztyn Mrągowo Lakelands and the Suwałki Lakeland(Fig 1 Appendix S1) The lakes chosen for the study were of various surface areas andhaving a floating peat mat growth belonging to Sphagnum sp that varied in size The lakesrepresented various succession stages in a disharmonic seriesmdashfrom oligo- to polyhumiclakes The oligohumic lakes were inhabited by Juncus bulbosus Eleocharis palustrisPhragmites australis Typha angustifolia Typha latiffolia Sphagnum sp Lobelia dortmannaIsoeumltes lacustris Drosera rotundifolia D anglica D intermedia Sparganium angustifoliumLycopodiella inundata In polyhumic lakes the dominant species included Sphagnum spOxycoccus quadripelatus Andromeda polifonia Eriophorum vaginatum Calluna vulgarisErica tetralix Empetrum nigrum The lakes were arranged a priori according to the seralstages of succession (Bloechl et al 2010) and the point of reference was assumed to be the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 426
coverage of a lakersquos surface by a Sphagnum mat (0ndash75) (Appendix S1) The percentagecover by a Sphagnum mat in each lake was calculated in the GIS system supported byArcMap 931 software For charting the research sites data made available throughGeoportal 2 in the WMS format were employed
The study was carried out from 2002 to 2014 in spring summer and autumn The faunasamples were collected with a dip net from a surface area of about 1 m2 In the compactedSphagnum mat environment a sample comprised 10 subsamples (aggregated afterwards)which were collected using a 01 m2 strainer Sampling sites were chosen so as to representthe biggest array of the littoral habitat diversity and areas of individual lakes Thus fivedifferent littoral components (habitats) were identified (1) the Sphagnum mat (2) sparsemacrophyte zone (3) dense macrophyte zone (4) sandy bottom zone and (5) pockets andponds within a Sphagnum mat The vegetation cover was assessed using Braun-Blanquet(1964) phytosociological records All lakes were identified with respect to the surface areaextent of the lakersquos surface covered with a Sphagnummat which corresponds to successionstages (1ndash3) percent share of individual habitats in the littoral zone (1ndash4) and a seralstage (1ndash40) (Appendix S1) It was assumed that these habitats due to different degreesof complexity represent different fractal dimensions all within an interval of 20 gtD gt30(Tokeshi amp Arakaki 2012) A schematic representation of fractals different in size can beseen in Fig 2 The smallest size (1) corresponds to the sandy bottom zone while the largestone (5) is assigned to the Sphagnummat The intermediate sizes belong to (2) pockets andponds within the Sphagnum mat (3) sparse macrophyte zone and (4) dense macrophytezone Field studies were conducted in accordance with field-study-approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP Certificate (PNE 08-070 18) and ZPKCertificate (06012018)
In total 499 samples were collected Subsequently the collected samples were describedaccording to the chosen environmental parameters habitat (1mdashSphagnummat 2mdashsparsemacrophytes 3mdashdense macrophytes 4mdashsandy bottom 5mdashpockets and ponds withinSphagnummat) place (location) (1ndashecotone 2mdashpockets and ponds within the Sphagnummat 3mdashcompacted Sphagnummat) bottom (soil substrate) (1mdashsand 2mdashsand and mud3mdashSphagnum) depth (1mdash0ndash10 cm 2mdash0ndash20 cm 3mdash20ndash40 cm 4mdash40ndash60 cm) nympheids(1mdashnone 2mdashpresent) elodeids (1mdashnone 2mdashpresent) (detritus (1mdashnone 2mdashscarce andfine 3mdashabundant and fine 4mdashabundant and coarse) debris (1 ndashnone 2mdashpresent) fractaldimension (1ndash5) stagemdashseral stage of succession (1ndash40)
Ecological and statistical analysesSpecies diversity was calculated using Smdashnumber of species Nmdashnumber of individualsand Dmdashpercent share All collected beetles were divided into five classes of different bodysize (where a body size meant the total length of an organisms) ie 1mdashvery small beetles(lt30 mm) 2mdashsmall beetles (30ndash50 mm) 3mdashmedium beetles (51ndash100 mm) 4mdashlargebeetles (101ndash200 mm) and 5mdashvery large beetles (gt200 mm) In addition the bodyweight was measured within each of the five classes The ranges of body weight values ofbeetles in the distinguished body size classes are shown in Table 1 Three functional groupswere distinguished in the trophic structure of beetles predators (families Gyrinidae
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 526
Figure 2 Distinguished habitats in the littoral zone of lakes (A) and hypothetical fractal dimensions(B) (after Tokeshi amp Arakaki (2012) modified)
Full-size DOI 107717peerj5662fig-2
Table 1 General description of the lake beetles
Body size class Abundance Number of species Body size (mm) Body weight (mg)
(meanplusmn SD) minndashmax meanplusmn (SD) minndashmax
Very small 2953 32 225plusmn 038 130ndash295 589plusmn 041 49ndash66Small 5640 37 347plusmn 056 300ndash490 790plusmn 078 710ndash889Medium 945 26 582plusmn 112 500ndash980 3828plusmn 376 2580ndash4216Large 565 24 1306plusmn 237 105ndash186 23426plusmn 8316 19028ndash31526Very large 36 5 3179plusmn 209 275ndash351 222802plusmn 1804 21302ndash29501
NotesMin minimum value max maximum value mean average value SD standard deviation
Dytiscidae and Noteridae) polyphages (Haliplidae) and saprophages (HelophoridaeHydrophilidae and Hydraenidae) (Appendix S2) Collected adult water beetles wereidentified using standard identification keys (Galewski 1990 Galewski amp Tranda 1978Friday 1988 Hansen 1987 Holmen 1987)
Because the samples of water beetle fauna were collected several times (taking intoaccount the phenological aspect) from the same lakes and from the habitats distinguishedwithin these lakes we used a GLM (Generalized Linear Model) for repeated measuresANOVA (Hocking 1996) to determine the significance of differences in the number
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 626
of beetles within each functional group in the distinguished habitats and to identifydependencies between the type of habitats body size abundance and biodiversity ofColeoptera First we checked the assumptions of normality (the ShapirondashWilk test) andhomogeneity of variances (the Levenersquos test) respectively The GLM repeated measuremodels were calculated on the basis of Type III sums of squares so as to take the unbalanceddesign into account Significant results were tested for pair-wise comparisons by theTukeyrsquos HSD post-hoc tests Dependent variables (abundance and number of species) weretransformed where necessary to fulfil the requirements of parametric tests (Saint-Germainet al 2007 Cremona Planas amp Lucotte 2008)
In four separate analyses we calculated the Pearsonrsquos correlation coefficients (rp) in orderto determine correlations between (1) body size and counts of beetles in the distinguishedhabitats (2) cover of the Sphagnum mat and counts of beetles (3) cover of the Sphagnummat and functional groups and (4) species and environmental variables
Linear regression analysis was performed to determine the influence of (1) the body sizeon the abundance of beetles (2) fractal dimension on the mean body size on (3) fractaldimension on the mean weight and (4) the seral stage of succession on the mean body sizeMultidimensional correspondence analysis (MCA) (Clausen 1998) served to determinedependencies between the abundance of water beetles within the identified body sizeclasses the share of the distinguished habitats in the littoral zones and in the analysed lakesMCA has been used in similar studies eg Usseglio-Polatera Bournaud amp Tachet 2000Obolewski Glińska-Lewczuk amp Kobus 2009 The average density of body size classes in asample was adjusted by the value corresponding to the total contribution of a given littoralzone type in individual lakes thus giving the weighted average The analysis included twodimensions of which the first explained the biggest part of the general chi-squared statistics( of inertia) whereas the inclusion of the other dimension increased the percentage ofexplained inertia Similarities in the fauna between particular lakes were also analysed bythe single linkage method for the hierarchical clustering of objects The distance measureis an Euclidean distance The results are presented in the form of a dendrogram drawn for40 lakes
Relationships between the presence of beetles and the analysed environmentalparameters in individual lakes were determined with the help of a principal componentanalysis PCA All calculations were performed in Statistica ver 130 (StatSoft Tulsa USA)
RESULTSGeneral description of the collected materialThe collected material comprised 10139 specimens representing 124 species classified toseven families (Gyrinidae Haliplidae Noteridae Dytiscidae Hydraenidae Hydrochidaeand Hydrophilidae) (Appendix S1) Most species (556) belonged to the two smallestbody size classes In total they composed 8475 of all collected specimens The remainingbeetles represented the consecutively higher body size classes of which the largest onehad the fewest representatives (Table 1) Most beetles (288 of the entire material) werecollected in pockets and ponds of the Sphagnum mat It was also there that the highest
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 726
Figure 3 The structure of body size of Coleoptera in distinguished habitatsN abundance S numberof species
Full-size DOI 107717peerj5662fig-3
species richness was determined (91 spp) The species diversity was lower in the densemacrophytes zone (266 74 spp) sparsemacrophytes zone (1964 66 spp) compactedSphagnum mat (189 64 spp) and finally in the sandy bottom (59 39 spp) Verysmall beetles which represented the first body size class demonstrated the highest speciesrichness in the compacted Sphagnummat (415 of the whole collected material) (Fig 3)while being the most numerous in the dense macrophyte zone Small beetles (class 2 of thebody size) were most numerously caught in the pockets and ponds within the Sphagnummat (734) The presence of very large beetles was notable in the sparse macrophyteszone pockets and ponds in the Sphagnum mat and in the sandy bottom zone althoughthey made up a small share of the collected material (around 1)
In the trophic structure the highest percentage (719) was achieved by predatorybeetles represented mainly by Dytiscidae (Fig 4) Although their species diversity wassimilar in all the habitats they were most numerous in the dense macropytes zone (913)and in the sandy bottom zone (845) Most saprophages (472) were noted in thecompacted Sphagnum mat while the number of species representing polyphages waslow in all of the habitats with the highest score (six spp) determined in the compactedSphagnum mat The GLM Repeated Measure Anova results (F(2499)= 674 p= 00089)indicate significant differences in counts of the distinguished trophic groups in theparticular habitats The counts of predatory beetles in the habitats in question did notdiffer significantly (the Tukeyrsquos HSD test p= 018) Conversely differences in the countsof predators and polyphages were significant (p= 00003)
Dependencies between types of habitats body size and abundanceas well as biodiversity of ColeopteraThe results of the GLM repeated measure Anova analysis point to a significant influence ofthe synergistic effect between the type of habitat and body size class on the abundance ofbeetles (F(161996)= 725 p= 00000002) Significant differences (the Tukeyrsquos HSD post -hoc
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 826
Figure 4 The trophic structure of Coleoptera in distinguished habitatsN abundance S number ofspecies
Full-size DOI 107717peerj5662fig-4
test) between the analysed subclasses are illustrated in Fig 5A (cf Appendix S3) Moreovera significant influence of the synergistic effect was determined between the indicated factors(habitat and body size class) and the number of species (F(161996)= 1622 p= 000002)(Appendix S3) Results of the GLM repeated measure also point to a significant influenceof the type of habitat on the number of beetles F(4499) = 675 p= 000003 (Fig 5B)Significant differences (the Tukeyrsquos HSD post -hoc test) were determined between thecompacted Sphagnum mat and sparse macrophytes zone (p= 0031) dense macrophyteszone and sparse macrophytes zone (p= 000014) dense macrophytes zone and pockets(p= 00479) Moreover significant differences were demonstrated in the numbers ofbeetles representing the distinguished body size classes (F(41996)= 7761 p= 000002) TheTukeyrsquos post-hoc test reveals significant differences in the counts of beetles representing allthe classes of body size (p= 000002) except 3 and 4 class (p= 077) Both factors the typeof habitat (F(4499)= 1104 p= 0000003) and body size (F(41996)= 37561 p= 0000001)also had a significant impact on the number of species determined With respect to thespecies diversity significant differences were found between the Sphagnummat and sparsemacrophytes (p= 00048) and between the pockets and ponds in the Sphagnum mat andsandy bottom zone (p= 00046) and between the Sphagnummat and sparse macrophytes(p= 000018) and dense macrophytes (p= 00014) Significant differences in the numberof species were shown between all body size classes (p= 000002) except 1 and 3 (p= 024)
Negative correlations were observed in all the habitats between the body size and thenumber of beetles It was only in the sandy bottom zone that these correlations were notsignificant (p= 047) (Table 2) Similarly negative correlations appeared between the bodysize and species richness in the compacted Sphagnum mat (p= 001) and in the sparsemacrophytes zone (p= 00008) (Table 2)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 926
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

coverage of a lakersquos surface by a Sphagnum mat (0ndash75) (Appendix S1) The percentagecover by a Sphagnum mat in each lake was calculated in the GIS system supported byArcMap 931 software For charting the research sites data made available throughGeoportal 2 in the WMS format were employed
The study was carried out from 2002 to 2014 in spring summer and autumn The faunasamples were collected with a dip net from a surface area of about 1 m2 In the compactedSphagnum mat environment a sample comprised 10 subsamples (aggregated afterwards)which were collected using a 01 m2 strainer Sampling sites were chosen so as to representthe biggest array of the littoral habitat diversity and areas of individual lakes Thus fivedifferent littoral components (habitats) were identified (1) the Sphagnum mat (2) sparsemacrophyte zone (3) dense macrophyte zone (4) sandy bottom zone and (5) pockets andponds within a Sphagnum mat The vegetation cover was assessed using Braun-Blanquet(1964) phytosociological records All lakes were identified with respect to the surface areaextent of the lakersquos surface covered with a Sphagnummat which corresponds to successionstages (1ndash3) percent share of individual habitats in the littoral zone (1ndash4) and a seralstage (1ndash40) (Appendix S1) It was assumed that these habitats due to different degreesof complexity represent different fractal dimensions all within an interval of 20 gtD gt30(Tokeshi amp Arakaki 2012) A schematic representation of fractals different in size can beseen in Fig 2 The smallest size (1) corresponds to the sandy bottom zone while the largestone (5) is assigned to the Sphagnummat The intermediate sizes belong to (2) pockets andponds within the Sphagnum mat (3) sparse macrophyte zone and (4) dense macrophytezone Field studies were conducted in accordance with field-study-approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP Certificate (PNE 08-070 18) and ZPKCertificate (06012018)
In total 499 samples were collected Subsequently the collected samples were describedaccording to the chosen environmental parameters habitat (1mdashSphagnummat 2mdashsparsemacrophytes 3mdashdense macrophytes 4mdashsandy bottom 5mdashpockets and ponds withinSphagnummat) place (location) (1ndashecotone 2mdashpockets and ponds within the Sphagnummat 3mdashcompacted Sphagnummat) bottom (soil substrate) (1mdashsand 2mdashsand and mud3mdashSphagnum) depth (1mdash0ndash10 cm 2mdash0ndash20 cm 3mdash20ndash40 cm 4mdash40ndash60 cm) nympheids(1mdashnone 2mdashpresent) elodeids (1mdashnone 2mdashpresent) (detritus (1mdashnone 2mdashscarce andfine 3mdashabundant and fine 4mdashabundant and coarse) debris (1 ndashnone 2mdashpresent) fractaldimension (1ndash5) stagemdashseral stage of succession (1ndash40)
Ecological and statistical analysesSpecies diversity was calculated using Smdashnumber of species Nmdashnumber of individualsand Dmdashpercent share All collected beetles were divided into five classes of different bodysize (where a body size meant the total length of an organisms) ie 1mdashvery small beetles(lt30 mm) 2mdashsmall beetles (30ndash50 mm) 3mdashmedium beetles (51ndash100 mm) 4mdashlargebeetles (101ndash200 mm) and 5mdashvery large beetles (gt200 mm) In addition the bodyweight was measured within each of the five classes The ranges of body weight values ofbeetles in the distinguished body size classes are shown in Table 1 Three functional groupswere distinguished in the trophic structure of beetles predators (families Gyrinidae
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 526
Figure 2 Distinguished habitats in the littoral zone of lakes (A) and hypothetical fractal dimensions(B) (after Tokeshi amp Arakaki (2012) modified)
Full-size DOI 107717peerj5662fig-2
Table 1 General description of the lake beetles
Body size class Abundance Number of species Body size (mm) Body weight (mg)
(meanplusmn SD) minndashmax meanplusmn (SD) minndashmax
Very small 2953 32 225plusmn 038 130ndash295 589plusmn 041 49ndash66Small 5640 37 347plusmn 056 300ndash490 790plusmn 078 710ndash889Medium 945 26 582plusmn 112 500ndash980 3828plusmn 376 2580ndash4216Large 565 24 1306plusmn 237 105ndash186 23426plusmn 8316 19028ndash31526Very large 36 5 3179plusmn 209 275ndash351 222802plusmn 1804 21302ndash29501
NotesMin minimum value max maximum value mean average value SD standard deviation
Dytiscidae and Noteridae) polyphages (Haliplidae) and saprophages (HelophoridaeHydrophilidae and Hydraenidae) (Appendix S2) Collected adult water beetles wereidentified using standard identification keys (Galewski 1990 Galewski amp Tranda 1978Friday 1988 Hansen 1987 Holmen 1987)
Because the samples of water beetle fauna were collected several times (taking intoaccount the phenological aspect) from the same lakes and from the habitats distinguishedwithin these lakes we used a GLM (Generalized Linear Model) for repeated measuresANOVA (Hocking 1996) to determine the significance of differences in the number
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 626
of beetles within each functional group in the distinguished habitats and to identifydependencies between the type of habitats body size abundance and biodiversity ofColeoptera First we checked the assumptions of normality (the ShapirondashWilk test) andhomogeneity of variances (the Levenersquos test) respectively The GLM repeated measuremodels were calculated on the basis of Type III sums of squares so as to take the unbalanceddesign into account Significant results were tested for pair-wise comparisons by theTukeyrsquos HSD post-hoc tests Dependent variables (abundance and number of species) weretransformed where necessary to fulfil the requirements of parametric tests (Saint-Germainet al 2007 Cremona Planas amp Lucotte 2008)
In four separate analyses we calculated the Pearsonrsquos correlation coefficients (rp) in orderto determine correlations between (1) body size and counts of beetles in the distinguishedhabitats (2) cover of the Sphagnum mat and counts of beetles (3) cover of the Sphagnummat and functional groups and (4) species and environmental variables
Linear regression analysis was performed to determine the influence of (1) the body sizeon the abundance of beetles (2) fractal dimension on the mean body size on (3) fractaldimension on the mean weight and (4) the seral stage of succession on the mean body sizeMultidimensional correspondence analysis (MCA) (Clausen 1998) served to determinedependencies between the abundance of water beetles within the identified body sizeclasses the share of the distinguished habitats in the littoral zones and in the analysed lakesMCA has been used in similar studies eg Usseglio-Polatera Bournaud amp Tachet 2000Obolewski Glińska-Lewczuk amp Kobus 2009 The average density of body size classes in asample was adjusted by the value corresponding to the total contribution of a given littoralzone type in individual lakes thus giving the weighted average The analysis included twodimensions of which the first explained the biggest part of the general chi-squared statistics( of inertia) whereas the inclusion of the other dimension increased the percentage ofexplained inertia Similarities in the fauna between particular lakes were also analysed bythe single linkage method for the hierarchical clustering of objects The distance measureis an Euclidean distance The results are presented in the form of a dendrogram drawn for40 lakes
Relationships between the presence of beetles and the analysed environmentalparameters in individual lakes were determined with the help of a principal componentanalysis PCA All calculations were performed in Statistica ver 130 (StatSoft Tulsa USA)
RESULTSGeneral description of the collected materialThe collected material comprised 10139 specimens representing 124 species classified toseven families (Gyrinidae Haliplidae Noteridae Dytiscidae Hydraenidae Hydrochidaeand Hydrophilidae) (Appendix S1) Most species (556) belonged to the two smallestbody size classes In total they composed 8475 of all collected specimens The remainingbeetles represented the consecutively higher body size classes of which the largest onehad the fewest representatives (Table 1) Most beetles (288 of the entire material) werecollected in pockets and ponds of the Sphagnum mat It was also there that the highest
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 726
Figure 3 The structure of body size of Coleoptera in distinguished habitatsN abundance S numberof species
Full-size DOI 107717peerj5662fig-3
species richness was determined (91 spp) The species diversity was lower in the densemacrophytes zone (266 74 spp) sparsemacrophytes zone (1964 66 spp) compactedSphagnum mat (189 64 spp) and finally in the sandy bottom (59 39 spp) Verysmall beetles which represented the first body size class demonstrated the highest speciesrichness in the compacted Sphagnummat (415 of the whole collected material) (Fig 3)while being the most numerous in the dense macrophyte zone Small beetles (class 2 of thebody size) were most numerously caught in the pockets and ponds within the Sphagnummat (734) The presence of very large beetles was notable in the sparse macrophyteszone pockets and ponds in the Sphagnum mat and in the sandy bottom zone althoughthey made up a small share of the collected material (around 1)
In the trophic structure the highest percentage (719) was achieved by predatorybeetles represented mainly by Dytiscidae (Fig 4) Although their species diversity wassimilar in all the habitats they were most numerous in the dense macropytes zone (913)and in the sandy bottom zone (845) Most saprophages (472) were noted in thecompacted Sphagnum mat while the number of species representing polyphages waslow in all of the habitats with the highest score (six spp) determined in the compactedSphagnum mat The GLM Repeated Measure Anova results (F(2499)= 674 p= 00089)indicate significant differences in counts of the distinguished trophic groups in theparticular habitats The counts of predatory beetles in the habitats in question did notdiffer significantly (the Tukeyrsquos HSD test p= 018) Conversely differences in the countsof predators and polyphages were significant (p= 00003)
Dependencies between types of habitats body size and abundanceas well as biodiversity of ColeopteraThe results of the GLM repeated measure Anova analysis point to a significant influence ofthe synergistic effect between the type of habitat and body size class on the abundance ofbeetles (F(161996)= 725 p= 00000002) Significant differences (the Tukeyrsquos HSD post -hoc
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 826
Figure 4 The trophic structure of Coleoptera in distinguished habitatsN abundance S number ofspecies
Full-size DOI 107717peerj5662fig-4
test) between the analysed subclasses are illustrated in Fig 5A (cf Appendix S3) Moreovera significant influence of the synergistic effect was determined between the indicated factors(habitat and body size class) and the number of species (F(161996)= 1622 p= 000002)(Appendix S3) Results of the GLM repeated measure also point to a significant influenceof the type of habitat on the number of beetles F(4499) = 675 p= 000003 (Fig 5B)Significant differences (the Tukeyrsquos HSD post -hoc test) were determined between thecompacted Sphagnum mat and sparse macrophytes zone (p= 0031) dense macrophyteszone and sparse macrophytes zone (p= 000014) dense macrophytes zone and pockets(p= 00479) Moreover significant differences were demonstrated in the numbers ofbeetles representing the distinguished body size classes (F(41996)= 7761 p= 000002) TheTukeyrsquos post-hoc test reveals significant differences in the counts of beetles representing allthe classes of body size (p= 000002) except 3 and 4 class (p= 077) Both factors the typeof habitat (F(4499)= 1104 p= 0000003) and body size (F(41996)= 37561 p= 0000001)also had a significant impact on the number of species determined With respect to thespecies diversity significant differences were found between the Sphagnummat and sparsemacrophytes (p= 00048) and between the pockets and ponds in the Sphagnum mat andsandy bottom zone (p= 00046) and between the Sphagnummat and sparse macrophytes(p= 000018) and dense macrophytes (p= 00014) Significant differences in the numberof species were shown between all body size classes (p= 000002) except 1 and 3 (p= 024)
Negative correlations were observed in all the habitats between the body size and thenumber of beetles It was only in the sandy bottom zone that these correlations were notsignificant (p= 047) (Table 2) Similarly negative correlations appeared between the bodysize and species richness in the compacted Sphagnum mat (p= 001) and in the sparsemacrophytes zone (p= 00008) (Table 2)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 926
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
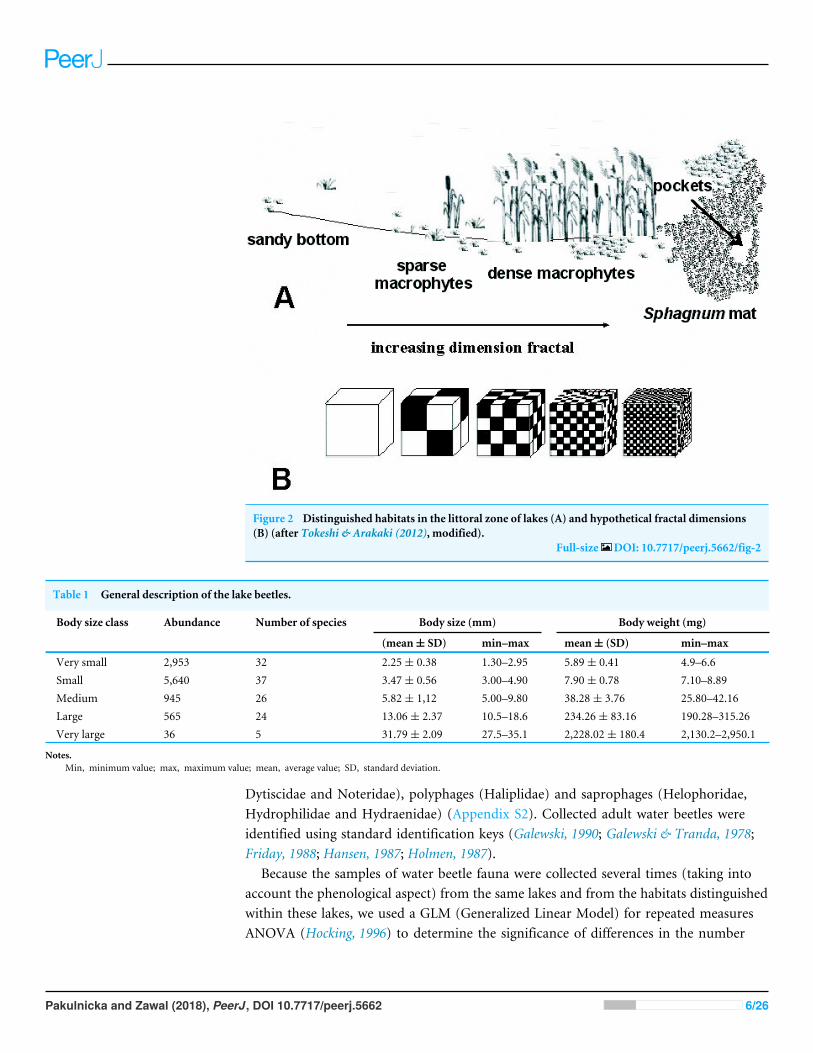
Figure 2 Distinguished habitats in the littoral zone of lakes (A) and hypothetical fractal dimensions(B) (after Tokeshi amp Arakaki (2012) modified)
Full-size DOI 107717peerj5662fig-2
Table 1 General description of the lake beetles
Body size class Abundance Number of species Body size (mm) Body weight (mg)
(meanplusmn SD) minndashmax meanplusmn (SD) minndashmax
Very small 2953 32 225plusmn 038 130ndash295 589plusmn 041 49ndash66Small 5640 37 347plusmn 056 300ndash490 790plusmn 078 710ndash889Medium 945 26 582plusmn 112 500ndash980 3828plusmn 376 2580ndash4216Large 565 24 1306plusmn 237 105ndash186 23426plusmn 8316 19028ndash31526Very large 36 5 3179plusmn 209 275ndash351 222802plusmn 1804 21302ndash29501
NotesMin minimum value max maximum value mean average value SD standard deviation
Dytiscidae and Noteridae) polyphages (Haliplidae) and saprophages (HelophoridaeHydrophilidae and Hydraenidae) (Appendix S2) Collected adult water beetles wereidentified using standard identification keys (Galewski 1990 Galewski amp Tranda 1978Friday 1988 Hansen 1987 Holmen 1987)
Because the samples of water beetle fauna were collected several times (taking intoaccount the phenological aspect) from the same lakes and from the habitats distinguishedwithin these lakes we used a GLM (Generalized Linear Model) for repeated measuresANOVA (Hocking 1996) to determine the significance of differences in the number
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 626
of beetles within each functional group in the distinguished habitats and to identifydependencies between the type of habitats body size abundance and biodiversity ofColeoptera First we checked the assumptions of normality (the ShapirondashWilk test) andhomogeneity of variances (the Levenersquos test) respectively The GLM repeated measuremodels were calculated on the basis of Type III sums of squares so as to take the unbalanceddesign into account Significant results were tested for pair-wise comparisons by theTukeyrsquos HSD post-hoc tests Dependent variables (abundance and number of species) weretransformed where necessary to fulfil the requirements of parametric tests (Saint-Germainet al 2007 Cremona Planas amp Lucotte 2008)
In four separate analyses we calculated the Pearsonrsquos correlation coefficients (rp) in orderto determine correlations between (1) body size and counts of beetles in the distinguishedhabitats (2) cover of the Sphagnum mat and counts of beetles (3) cover of the Sphagnummat and functional groups and (4) species and environmental variables
Linear regression analysis was performed to determine the influence of (1) the body sizeon the abundance of beetles (2) fractal dimension on the mean body size on (3) fractaldimension on the mean weight and (4) the seral stage of succession on the mean body sizeMultidimensional correspondence analysis (MCA) (Clausen 1998) served to determinedependencies between the abundance of water beetles within the identified body sizeclasses the share of the distinguished habitats in the littoral zones and in the analysed lakesMCA has been used in similar studies eg Usseglio-Polatera Bournaud amp Tachet 2000Obolewski Glińska-Lewczuk amp Kobus 2009 The average density of body size classes in asample was adjusted by the value corresponding to the total contribution of a given littoralzone type in individual lakes thus giving the weighted average The analysis included twodimensions of which the first explained the biggest part of the general chi-squared statistics( of inertia) whereas the inclusion of the other dimension increased the percentage ofexplained inertia Similarities in the fauna between particular lakes were also analysed bythe single linkage method for the hierarchical clustering of objects The distance measureis an Euclidean distance The results are presented in the form of a dendrogram drawn for40 lakes
Relationships between the presence of beetles and the analysed environmentalparameters in individual lakes were determined with the help of a principal componentanalysis PCA All calculations were performed in Statistica ver 130 (StatSoft Tulsa USA)
RESULTSGeneral description of the collected materialThe collected material comprised 10139 specimens representing 124 species classified toseven families (Gyrinidae Haliplidae Noteridae Dytiscidae Hydraenidae Hydrochidaeand Hydrophilidae) (Appendix S1) Most species (556) belonged to the two smallestbody size classes In total they composed 8475 of all collected specimens The remainingbeetles represented the consecutively higher body size classes of which the largest onehad the fewest representatives (Table 1) Most beetles (288 of the entire material) werecollected in pockets and ponds of the Sphagnum mat It was also there that the highest
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 726
Figure 3 The structure of body size of Coleoptera in distinguished habitatsN abundance S numberof species
Full-size DOI 107717peerj5662fig-3
species richness was determined (91 spp) The species diversity was lower in the densemacrophytes zone (266 74 spp) sparsemacrophytes zone (1964 66 spp) compactedSphagnum mat (189 64 spp) and finally in the sandy bottom (59 39 spp) Verysmall beetles which represented the first body size class demonstrated the highest speciesrichness in the compacted Sphagnummat (415 of the whole collected material) (Fig 3)while being the most numerous in the dense macrophyte zone Small beetles (class 2 of thebody size) were most numerously caught in the pockets and ponds within the Sphagnummat (734) The presence of very large beetles was notable in the sparse macrophyteszone pockets and ponds in the Sphagnum mat and in the sandy bottom zone althoughthey made up a small share of the collected material (around 1)
In the trophic structure the highest percentage (719) was achieved by predatorybeetles represented mainly by Dytiscidae (Fig 4) Although their species diversity wassimilar in all the habitats they were most numerous in the dense macropytes zone (913)and in the sandy bottom zone (845) Most saprophages (472) were noted in thecompacted Sphagnum mat while the number of species representing polyphages waslow in all of the habitats with the highest score (six spp) determined in the compactedSphagnum mat The GLM Repeated Measure Anova results (F(2499)= 674 p= 00089)indicate significant differences in counts of the distinguished trophic groups in theparticular habitats The counts of predatory beetles in the habitats in question did notdiffer significantly (the Tukeyrsquos HSD test p= 018) Conversely differences in the countsof predators and polyphages were significant (p= 00003)
Dependencies between types of habitats body size and abundanceas well as biodiversity of ColeopteraThe results of the GLM repeated measure Anova analysis point to a significant influence ofthe synergistic effect between the type of habitat and body size class on the abundance ofbeetles (F(161996)= 725 p= 00000002) Significant differences (the Tukeyrsquos HSD post -hoc
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 826
Figure 4 The trophic structure of Coleoptera in distinguished habitatsN abundance S number ofspecies
Full-size DOI 107717peerj5662fig-4
test) between the analysed subclasses are illustrated in Fig 5A (cf Appendix S3) Moreovera significant influence of the synergistic effect was determined between the indicated factors(habitat and body size class) and the number of species (F(161996)= 1622 p= 000002)(Appendix S3) Results of the GLM repeated measure also point to a significant influenceof the type of habitat on the number of beetles F(4499) = 675 p= 000003 (Fig 5B)Significant differences (the Tukeyrsquos HSD post -hoc test) were determined between thecompacted Sphagnum mat and sparse macrophytes zone (p= 0031) dense macrophyteszone and sparse macrophytes zone (p= 000014) dense macrophytes zone and pockets(p= 00479) Moreover significant differences were demonstrated in the numbers ofbeetles representing the distinguished body size classes (F(41996)= 7761 p= 000002) TheTukeyrsquos post-hoc test reveals significant differences in the counts of beetles representing allthe classes of body size (p= 000002) except 3 and 4 class (p= 077) Both factors the typeof habitat (F(4499)= 1104 p= 0000003) and body size (F(41996)= 37561 p= 0000001)also had a significant impact on the number of species determined With respect to thespecies diversity significant differences were found between the Sphagnummat and sparsemacrophytes (p= 00048) and between the pockets and ponds in the Sphagnum mat andsandy bottom zone (p= 00046) and between the Sphagnummat and sparse macrophytes(p= 000018) and dense macrophytes (p= 00014) Significant differences in the numberof species were shown between all body size classes (p= 000002) except 1 and 3 (p= 024)
Negative correlations were observed in all the habitats between the body size and thenumber of beetles It was only in the sandy bottom zone that these correlations were notsignificant (p= 047) (Table 2) Similarly negative correlations appeared between the bodysize and species richness in the compacted Sphagnum mat (p= 001) and in the sparsemacrophytes zone (p= 00008) (Table 2)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 926
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

of beetles within each functional group in the distinguished habitats and to identifydependencies between the type of habitats body size abundance and biodiversity ofColeoptera First we checked the assumptions of normality (the ShapirondashWilk test) andhomogeneity of variances (the Levenersquos test) respectively The GLM repeated measuremodels were calculated on the basis of Type III sums of squares so as to take the unbalanceddesign into account Significant results were tested for pair-wise comparisons by theTukeyrsquos HSD post-hoc tests Dependent variables (abundance and number of species) weretransformed where necessary to fulfil the requirements of parametric tests (Saint-Germainet al 2007 Cremona Planas amp Lucotte 2008)
In four separate analyses we calculated the Pearsonrsquos correlation coefficients (rp) in orderto determine correlations between (1) body size and counts of beetles in the distinguishedhabitats (2) cover of the Sphagnum mat and counts of beetles (3) cover of the Sphagnummat and functional groups and (4) species and environmental variables
Linear regression analysis was performed to determine the influence of (1) the body sizeon the abundance of beetles (2) fractal dimension on the mean body size on (3) fractaldimension on the mean weight and (4) the seral stage of succession on the mean body sizeMultidimensional correspondence analysis (MCA) (Clausen 1998) served to determinedependencies between the abundance of water beetles within the identified body sizeclasses the share of the distinguished habitats in the littoral zones and in the analysed lakesMCA has been used in similar studies eg Usseglio-Polatera Bournaud amp Tachet 2000Obolewski Glińska-Lewczuk amp Kobus 2009 The average density of body size classes in asample was adjusted by the value corresponding to the total contribution of a given littoralzone type in individual lakes thus giving the weighted average The analysis included twodimensions of which the first explained the biggest part of the general chi-squared statistics( of inertia) whereas the inclusion of the other dimension increased the percentage ofexplained inertia Similarities in the fauna between particular lakes were also analysed bythe single linkage method for the hierarchical clustering of objects The distance measureis an Euclidean distance The results are presented in the form of a dendrogram drawn for40 lakes
Relationships between the presence of beetles and the analysed environmentalparameters in individual lakes were determined with the help of a principal componentanalysis PCA All calculations were performed in Statistica ver 130 (StatSoft Tulsa USA)
RESULTSGeneral description of the collected materialThe collected material comprised 10139 specimens representing 124 species classified toseven families (Gyrinidae Haliplidae Noteridae Dytiscidae Hydraenidae Hydrochidaeand Hydrophilidae) (Appendix S1) Most species (556) belonged to the two smallestbody size classes In total they composed 8475 of all collected specimens The remainingbeetles represented the consecutively higher body size classes of which the largest onehad the fewest representatives (Table 1) Most beetles (288 of the entire material) werecollected in pockets and ponds of the Sphagnum mat It was also there that the highest
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 726
Figure 3 The structure of body size of Coleoptera in distinguished habitatsN abundance S numberof species
Full-size DOI 107717peerj5662fig-3
species richness was determined (91 spp) The species diversity was lower in the densemacrophytes zone (266 74 spp) sparsemacrophytes zone (1964 66 spp) compactedSphagnum mat (189 64 spp) and finally in the sandy bottom (59 39 spp) Verysmall beetles which represented the first body size class demonstrated the highest speciesrichness in the compacted Sphagnummat (415 of the whole collected material) (Fig 3)while being the most numerous in the dense macrophyte zone Small beetles (class 2 of thebody size) were most numerously caught in the pockets and ponds within the Sphagnummat (734) The presence of very large beetles was notable in the sparse macrophyteszone pockets and ponds in the Sphagnum mat and in the sandy bottom zone althoughthey made up a small share of the collected material (around 1)
In the trophic structure the highest percentage (719) was achieved by predatorybeetles represented mainly by Dytiscidae (Fig 4) Although their species diversity wassimilar in all the habitats they were most numerous in the dense macropytes zone (913)and in the sandy bottom zone (845) Most saprophages (472) were noted in thecompacted Sphagnum mat while the number of species representing polyphages waslow in all of the habitats with the highest score (six spp) determined in the compactedSphagnum mat The GLM Repeated Measure Anova results (F(2499)= 674 p= 00089)indicate significant differences in counts of the distinguished trophic groups in theparticular habitats The counts of predatory beetles in the habitats in question did notdiffer significantly (the Tukeyrsquos HSD test p= 018) Conversely differences in the countsof predators and polyphages were significant (p= 00003)
Dependencies between types of habitats body size and abundanceas well as biodiversity of ColeopteraThe results of the GLM repeated measure Anova analysis point to a significant influence ofthe synergistic effect between the type of habitat and body size class on the abundance ofbeetles (F(161996)= 725 p= 00000002) Significant differences (the Tukeyrsquos HSD post -hoc
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 826
Figure 4 The trophic structure of Coleoptera in distinguished habitatsN abundance S number ofspecies
Full-size DOI 107717peerj5662fig-4
test) between the analysed subclasses are illustrated in Fig 5A (cf Appendix S3) Moreovera significant influence of the synergistic effect was determined between the indicated factors(habitat and body size class) and the number of species (F(161996)= 1622 p= 000002)(Appendix S3) Results of the GLM repeated measure also point to a significant influenceof the type of habitat on the number of beetles F(4499) = 675 p= 000003 (Fig 5B)Significant differences (the Tukeyrsquos HSD post -hoc test) were determined between thecompacted Sphagnum mat and sparse macrophytes zone (p= 0031) dense macrophyteszone and sparse macrophytes zone (p= 000014) dense macrophytes zone and pockets(p= 00479) Moreover significant differences were demonstrated in the numbers ofbeetles representing the distinguished body size classes (F(41996)= 7761 p= 000002) TheTukeyrsquos post-hoc test reveals significant differences in the counts of beetles representing allthe classes of body size (p= 000002) except 3 and 4 class (p= 077) Both factors the typeof habitat (F(4499)= 1104 p= 0000003) and body size (F(41996)= 37561 p= 0000001)also had a significant impact on the number of species determined With respect to thespecies diversity significant differences were found between the Sphagnummat and sparsemacrophytes (p= 00048) and between the pockets and ponds in the Sphagnum mat andsandy bottom zone (p= 00046) and between the Sphagnummat and sparse macrophytes(p= 000018) and dense macrophytes (p= 00014) Significant differences in the numberof species were shown between all body size classes (p= 000002) except 1 and 3 (p= 024)
Negative correlations were observed in all the habitats between the body size and thenumber of beetles It was only in the sandy bottom zone that these correlations were notsignificant (p= 047) (Table 2) Similarly negative correlations appeared between the bodysize and species richness in the compacted Sphagnum mat (p= 001) and in the sparsemacrophytes zone (p= 00008) (Table 2)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 926
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
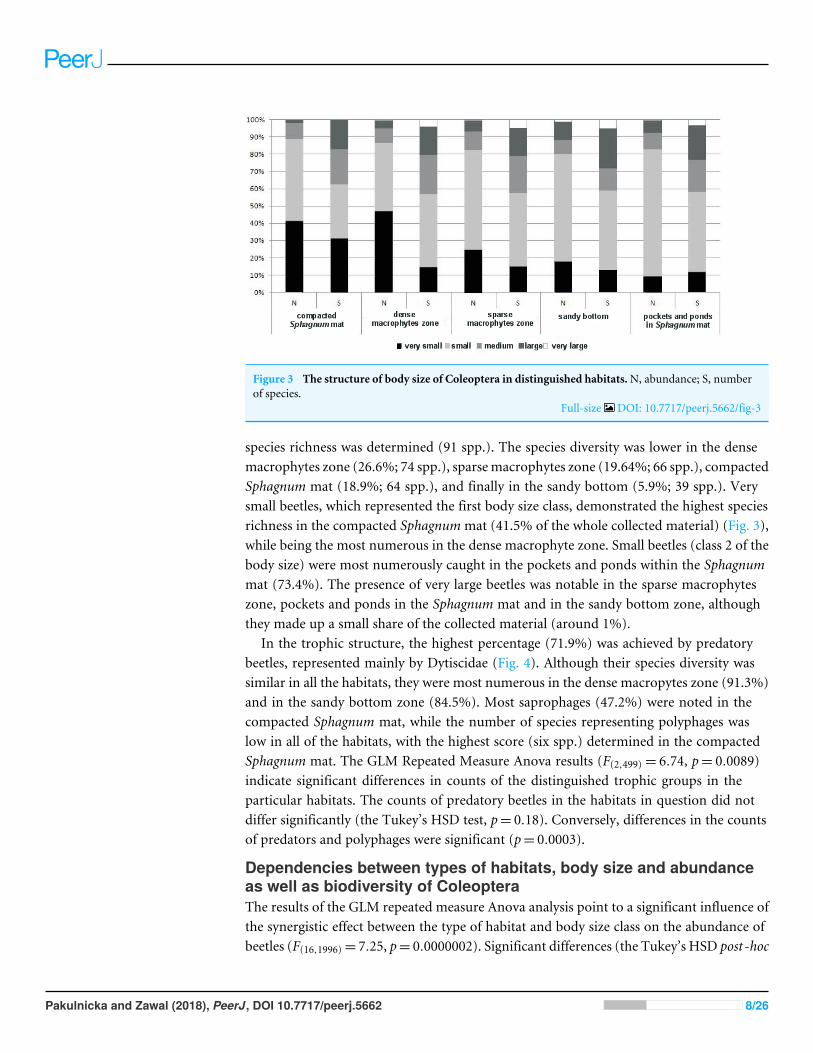
Figure 3 The structure of body size of Coleoptera in distinguished habitatsN abundance S numberof species
Full-size DOI 107717peerj5662fig-3
species richness was determined (91 spp) The species diversity was lower in the densemacrophytes zone (266 74 spp) sparsemacrophytes zone (1964 66 spp) compactedSphagnum mat (189 64 spp) and finally in the sandy bottom (59 39 spp) Verysmall beetles which represented the first body size class demonstrated the highest speciesrichness in the compacted Sphagnummat (415 of the whole collected material) (Fig 3)while being the most numerous in the dense macrophyte zone Small beetles (class 2 of thebody size) were most numerously caught in the pockets and ponds within the Sphagnummat (734) The presence of very large beetles was notable in the sparse macrophyteszone pockets and ponds in the Sphagnum mat and in the sandy bottom zone althoughthey made up a small share of the collected material (around 1)
In the trophic structure the highest percentage (719) was achieved by predatorybeetles represented mainly by Dytiscidae (Fig 4) Although their species diversity wassimilar in all the habitats they were most numerous in the dense macropytes zone (913)and in the sandy bottom zone (845) Most saprophages (472) were noted in thecompacted Sphagnum mat while the number of species representing polyphages waslow in all of the habitats with the highest score (six spp) determined in the compactedSphagnum mat The GLM Repeated Measure Anova results (F(2499)= 674 p= 00089)indicate significant differences in counts of the distinguished trophic groups in theparticular habitats The counts of predatory beetles in the habitats in question did notdiffer significantly (the Tukeyrsquos HSD test p= 018) Conversely differences in the countsof predators and polyphages were significant (p= 00003)
Dependencies between types of habitats body size and abundanceas well as biodiversity of ColeopteraThe results of the GLM repeated measure Anova analysis point to a significant influence ofthe synergistic effect between the type of habitat and body size class on the abundance ofbeetles (F(161996)= 725 p= 00000002) Significant differences (the Tukeyrsquos HSD post -hoc
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 826
Figure 4 The trophic structure of Coleoptera in distinguished habitatsN abundance S number ofspecies
Full-size DOI 107717peerj5662fig-4
test) between the analysed subclasses are illustrated in Fig 5A (cf Appendix S3) Moreovera significant influence of the synergistic effect was determined between the indicated factors(habitat and body size class) and the number of species (F(161996)= 1622 p= 000002)(Appendix S3) Results of the GLM repeated measure also point to a significant influenceof the type of habitat on the number of beetles F(4499) = 675 p= 000003 (Fig 5B)Significant differences (the Tukeyrsquos HSD post -hoc test) were determined between thecompacted Sphagnum mat and sparse macrophytes zone (p= 0031) dense macrophyteszone and sparse macrophytes zone (p= 000014) dense macrophytes zone and pockets(p= 00479) Moreover significant differences were demonstrated in the numbers ofbeetles representing the distinguished body size classes (F(41996)= 7761 p= 000002) TheTukeyrsquos post-hoc test reveals significant differences in the counts of beetles representing allthe classes of body size (p= 000002) except 3 and 4 class (p= 077) Both factors the typeof habitat (F(4499)= 1104 p= 0000003) and body size (F(41996)= 37561 p= 0000001)also had a significant impact on the number of species determined With respect to thespecies diversity significant differences were found between the Sphagnummat and sparsemacrophytes (p= 00048) and between the pockets and ponds in the Sphagnum mat andsandy bottom zone (p= 00046) and between the Sphagnummat and sparse macrophytes(p= 000018) and dense macrophytes (p= 00014) Significant differences in the numberof species were shown between all body size classes (p= 000002) except 1 and 3 (p= 024)
Negative correlations were observed in all the habitats between the body size and thenumber of beetles It was only in the sandy bottom zone that these correlations were notsignificant (p= 047) (Table 2) Similarly negative correlations appeared between the bodysize and species richness in the compacted Sphagnum mat (p= 001) and in the sparsemacrophytes zone (p= 00008) (Table 2)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 926
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
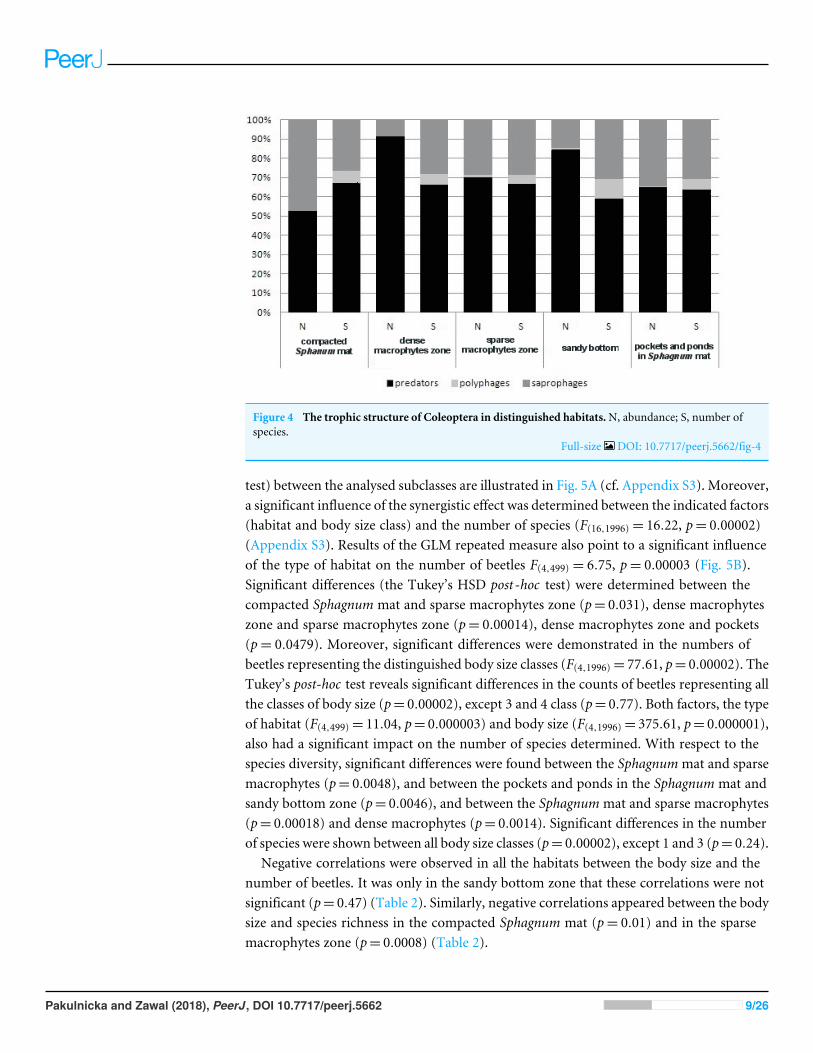
Figure 4 The trophic structure of Coleoptera in distinguished habitatsN abundance S number ofspecies
Full-size DOI 107717peerj5662fig-4
test) between the analysed subclasses are illustrated in Fig 5A (cf Appendix S3) Moreovera significant influence of the synergistic effect was determined between the indicated factors(habitat and body size class) and the number of species (F(161996)= 1622 p= 000002)(Appendix S3) Results of the GLM repeated measure also point to a significant influenceof the type of habitat on the number of beetles F(4499) = 675 p= 000003 (Fig 5B)Significant differences (the Tukeyrsquos HSD post -hoc test) were determined between thecompacted Sphagnum mat and sparse macrophytes zone (p= 0031) dense macrophyteszone and sparse macrophytes zone (p= 000014) dense macrophytes zone and pockets(p= 00479) Moreover significant differences were demonstrated in the numbers ofbeetles representing the distinguished body size classes (F(41996)= 7761 p= 000002) TheTukeyrsquos post-hoc test reveals significant differences in the counts of beetles representing allthe classes of body size (p= 000002) except 3 and 4 class (p= 077) Both factors the typeof habitat (F(4499)= 1104 p= 0000003) and body size (F(41996)= 37561 p= 0000001)also had a significant impact on the number of species determined With respect to thespecies diversity significant differences were found between the Sphagnummat and sparsemacrophytes (p= 00048) and between the pockets and ponds in the Sphagnum mat andsandy bottom zone (p= 00046) and between the Sphagnummat and sparse macrophytes(p= 000018) and dense macrophytes (p= 00014) Significant differences in the numberof species were shown between all body size classes (p= 000002) except 1 and 3 (p= 024)
Negative correlations were observed in all the habitats between the body size and thenumber of beetles It was only in the sandy bottom zone that these correlations were notsignificant (p= 047) (Table 2) Similarly negative correlations appeared between the bodysize and species richness in the compacted Sphagnum mat (p= 001) and in the sparsemacrophytes zone (p= 00008) (Table 2)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 926
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
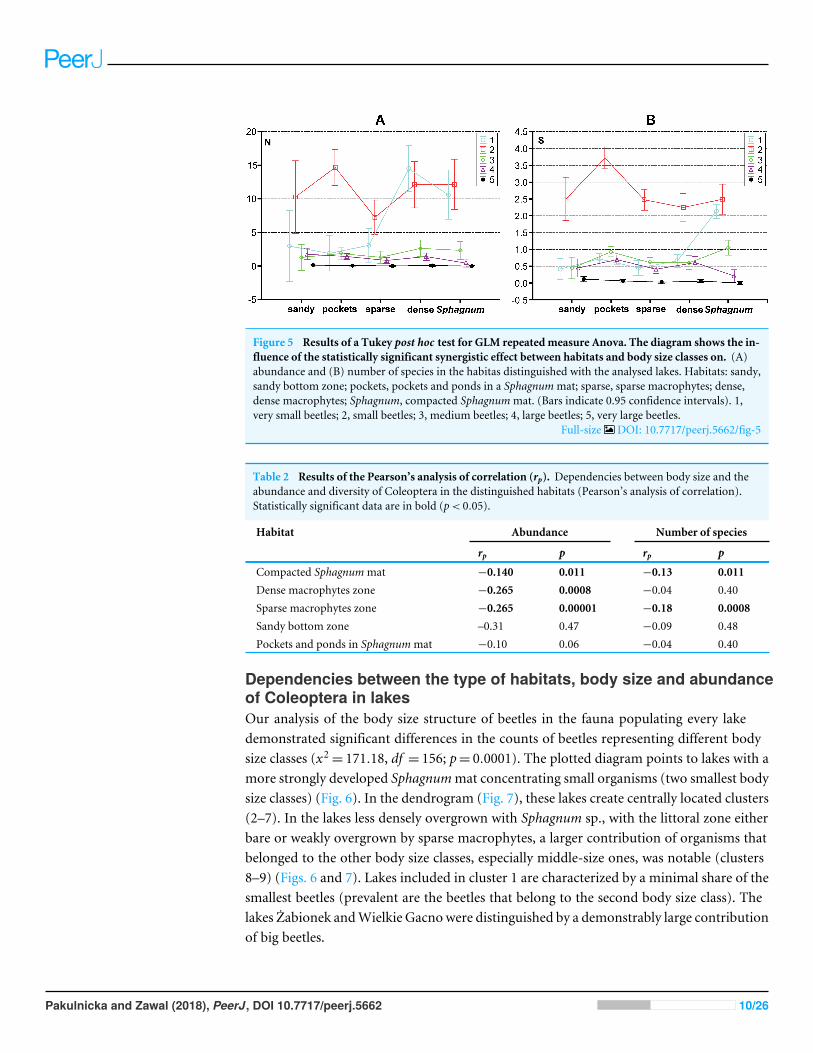
Figure 5 Results of a Tukey post hoc test for GLM repeated measure Anova The diagram shows the in-fluence of the statistically significant synergistic effect between habitats and body size classes on (A)abundance and (B) number of species in the habitas distinguished with the analysed lakes Habitats sandysandy bottom zone pockets pockets and ponds in a Sphagnummat sparse sparse macrophytes densedense macrophytes Sphagnum compacted Sphagnummat (Bars indicate 095 confidence intervals) 1very small beetles 2 small beetles 3 medium beetles 4 large beetles 5 very large beetles
Full-size DOI 107717peerj5662fig-5
Table 2 Results of the Pearsonrsquos analysis of correlation (rp) Dependencies between body size and theabundance and diversity of Coleoptera in the distinguished habitats (Pearsonrsquos analysis of correlation)Statistically significant data are in bold (plt 005)
Habitat Abundance Number of species
rp p rp p
Compacted Sphagnummat minus0140 0011 minus013 0011Dense macrophytes zone minus0265 00008 minus004 040Sparse macrophytes zone minus0265 000001 minus018 00008Sandy bottom zone ndash031 047 minus009 048Pockets and ponds in Sphagnummat minus010 006 minus004 040
Dependencies between the type of habitats body size and abundanceof Coleoptera in lakesOur analysis of the body size structure of beetles in the fauna populating every lakedemonstrated significant differences in the counts of beetles representing different bodysize classes (x2= 17118 df = 156 p= 00001) The plotted diagram points to lakes with amore strongly developed Sphagnummat concentrating small organisms (two smallest bodysize classes) (Fig 6) In the dendrogram (Fig 7) these lakes create centrally located clusters(2ndash7) In the lakes less densely overgrown with Sphagnum sp with the littoral zone eitherbare or weakly overgrown by sparse macrophytes a larger contribution of organisms thatbelonged to the other body size classes especially middle-size ones was notable (clusters8ndash9) (Figs 6 and 7) Lakes included in cluster 1 are characterized by a minimal share of thesmallest beetles (prevalent are the beetles that belong to the second body size class) Thelakes Żabionek andWielkie Gacnowere distinguished by a demonstrably large contributionof big beetles
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1026
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
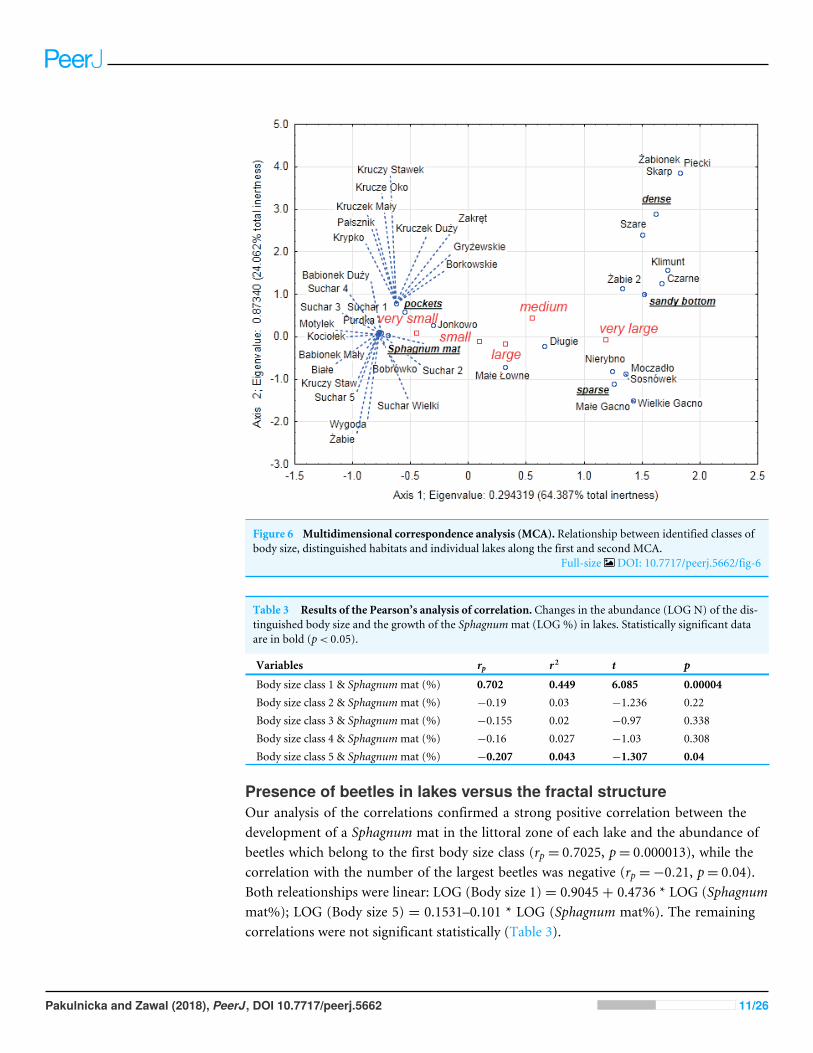
Figure 6 Multidimensional correspondence analysis (MCA) Relationship between identified classes ofbody size distinguished habitats and individual lakes along the first and second MCA
Full-size DOI 107717peerj5662fig-6
Table 3 Results of the Pearsonrsquos analysis of correlation Changes in the abundance (LOG N) of the dis-tinguished body size and the growth of the Sphagnummat (LOG ) in lakes Statistically significant dataare in bold (plt 005)
Variables rp r2 t p
Body size class 1 amp Sphagnummat () 0702 0449 6085 000004Body size class 2 amp Sphagnummat () minus019 003 minus1236 022Body size class 3 amp Sphagnummat () minus0155 002 minus097 0338Body size class 4 amp Sphagnummat () minus016 0027 minus103 0308Body size class 5 amp Sphagnummat () minus0207 0043 minus1307 004
Presence of beetles in lakes versus the fractal structureOur analysis of the correlations confirmed a strong positive correlation between thedevelopment of a Sphagnum mat in the littoral zone of each lake and the abundance ofbeetles which belong to the first body size class (rp = 07025 p= 0000013) while thecorrelation with the number of the largest beetles was negative (rp =minus021 p= 004)Both releationships were linear LOG (Body size 1) = 09045 + 04736 LOG (Sphagnummat) LOG (Body size 5) = 01531ndash0101 LOG (Sphagnum mat) The remainingcorrelations were not significant statistically (Table 3)
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1126
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
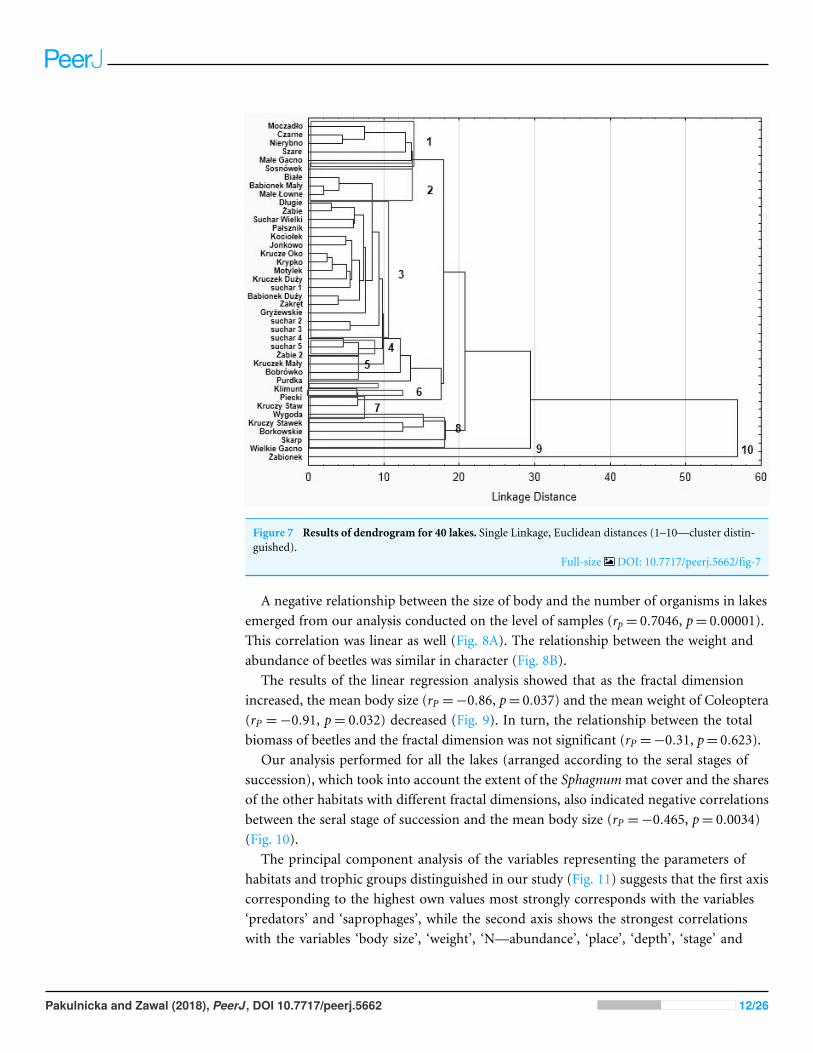
Figure 7 Results of dendrogram for 40 lakes Single Linkage Euclidean distances (1ndash10mdashcluster distin-guished)
Full-size DOI 107717peerj5662fig-7
A negative relationship between the size of body and the number of organisms in lakesemerged from our analysis conducted on the level of samples (rp= 07046 p= 000001)This correlation was linear as well (Fig 8A) The relationship between the weight andabundance of beetles was similar in character (Fig 8B)
The results of the linear regression analysis showed that as the fractal dimensionincreased the mean body size (rP =minus086 p= 0037) and the mean weight of Coleoptera(rP =minus091 p= 0032) decreased (Fig 9) In turn the relationship between the totalbiomass of beetles and the fractal dimension was not significant (rP =minus031 p= 0623)
Our analysis performed for all the lakes (arranged according to the seral stages ofsuccession) which took into account the extent of the Sphagnummat cover and the sharesof the other habitats with different fractal dimensions also indicated negative correlationsbetween the seral stage of succession and the mean body size (rP =minus0465 p= 00034)(Fig 10)
The principal component analysis of the variables representing the parameters ofhabitats and trophic groups distinguished in our study (Fig 11) suggests that the first axiscorresponding to the highest own values most strongly corresponds with the variableslsquopredatorsrsquo and lsquosaprophagesrsquo while the second axis shows the strongest correlationswith the variables lsquobody sizersquo lsquoweightrsquo lsquoNmdashabundancersquo lsquoplacersquo lsquodepthrsquo lsquostagersquo and
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1226
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
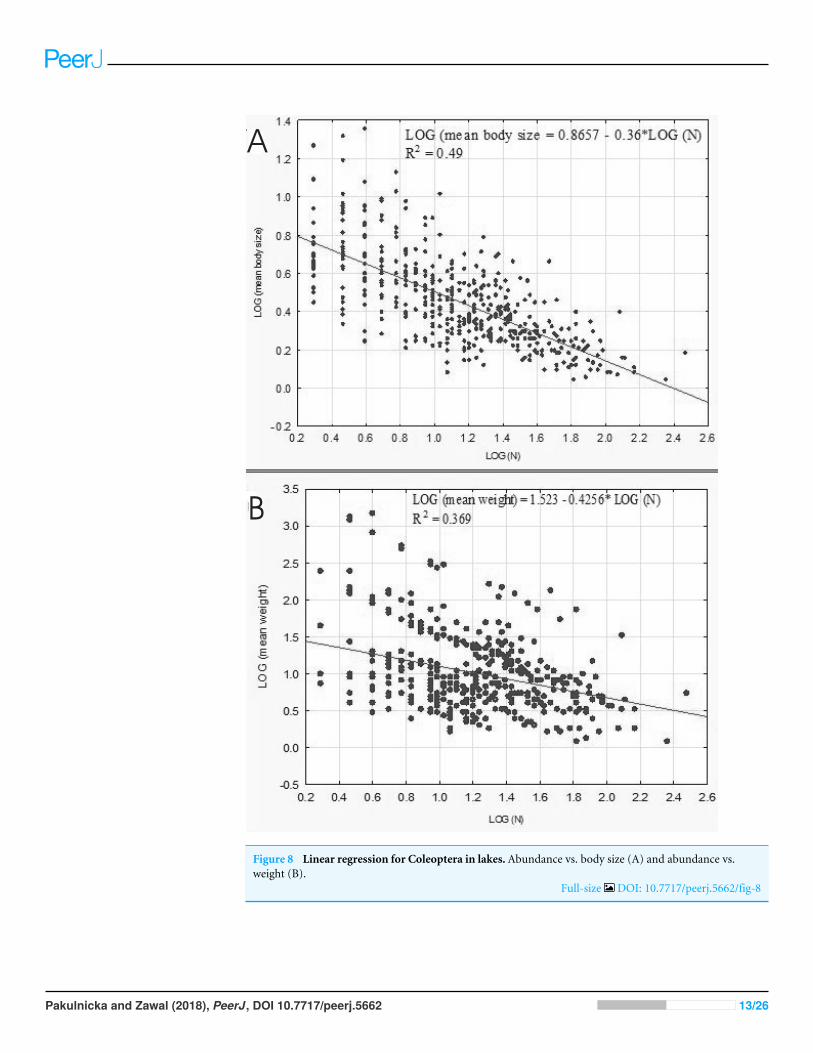
A
B
Figure 8 Linear regression for Coleoptera in lakes Abundance vs body size (A) and abundance vsweight (B)
Full-size DOI 107717peerj5662fig-8
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1326
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
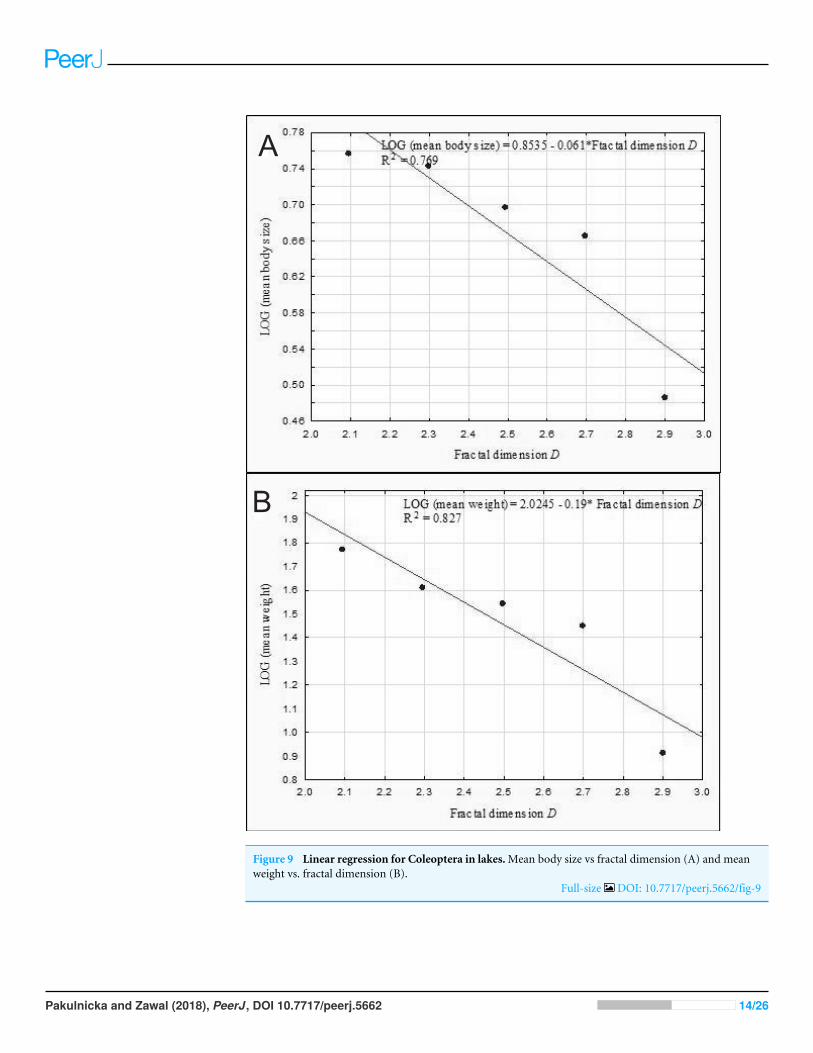
A
B
Figure 9 Linear regression for Coleoptera in lakesMean body size vs fractal dimension (A) and meanweight vs fractal dimension (B)
Full-size DOI 107717peerj5662fig-9
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1426
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
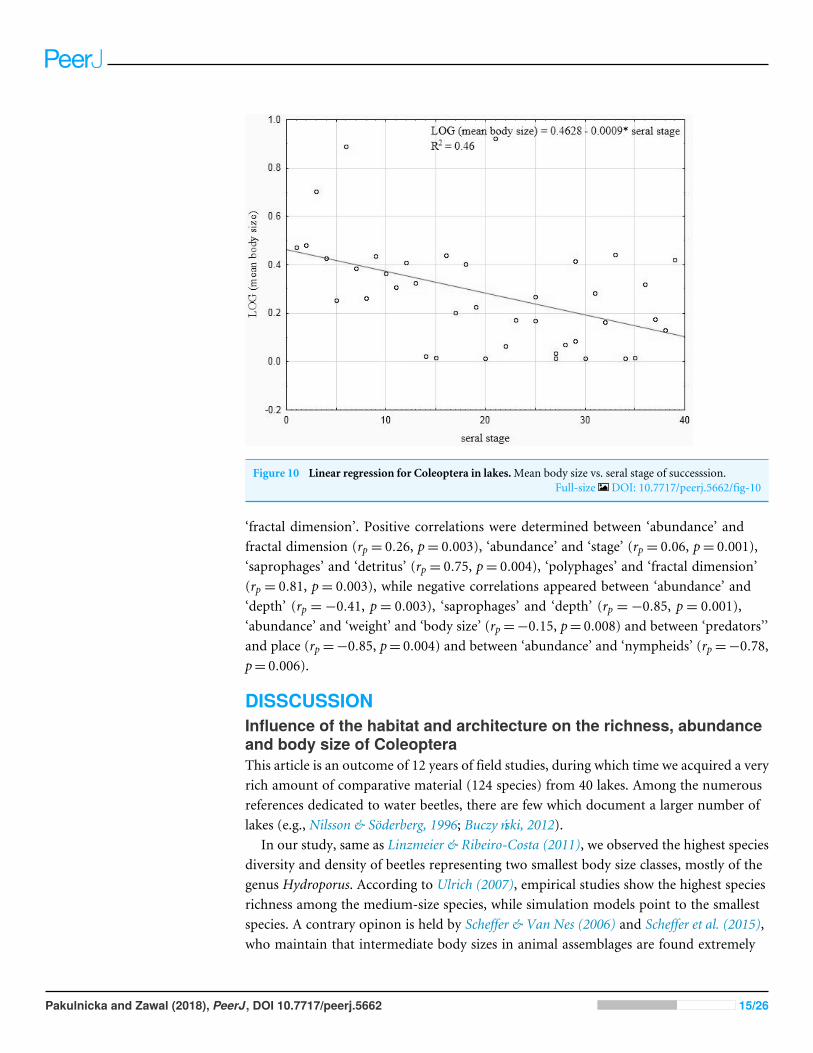
Figure 10 Linear regression for Coleoptera in lakesMean body size vs seral stage of successsionFull-size DOI 107717peerj5662fig-10
lsquofractal dimensionrsquo Positive correlations were determined between lsquoabundancersquo andfractal dimension (rp = 026 p= 0003) lsquoabundancersquo and lsquostagersquo (rp = 006 p= 0001)lsquosaprophagesrsquo and lsquodetritusrsquo (rp = 075 p= 0004) lsquopolyphagesrsquo and lsquofractal dimensionrsquo(rp = 081 p= 0003) while negative correlations appeared between lsquoabundancersquo andlsquodepthrsquo (rp =minus041 p= 0003) lsquosaprophagesrsquo and lsquodepthrsquo (rp =minus085 p= 0001)lsquoabundancersquo and lsquoweightrsquo and lsquobody sizersquo (rp=minus015 p= 0008) and between lsquopredatorsrsquorsquoand place (rp=minus085 p= 0004) and between lsquoabundancersquo and lsquonympheidsrsquo (rp=minus078p= 0006)
DISSCUSSIONInfluence of the habitat and architecture on the richness abundanceand body size of ColeopteraThis article is an outcome of 12 years of field studies during which time we acquired a veryrich amount of comparative material (124 species) from 40 lakes Among the numerousreferences dedicated to water beetles there are few which document a larger number oflakes (eg Nilsson amp Soumlderberg 1996 Buczyński 2012)
In our study same as Linzmeier amp Ribeiro-Costa (2011) we observed the highest speciesdiversity and density of beetles representing two smallest body size classes mostly of thegenus Hydroporus According to Ulrich (2007) empirical studies show the highest speciesrichness among the medium-size species while simulation models point to the smallestspecies A contrary opinon is held by Scheffer amp Van Nes (2006) and Scheffer et al (2015)who maintain that intermediate body sizes in animal assemblages are found extremely
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1526
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626
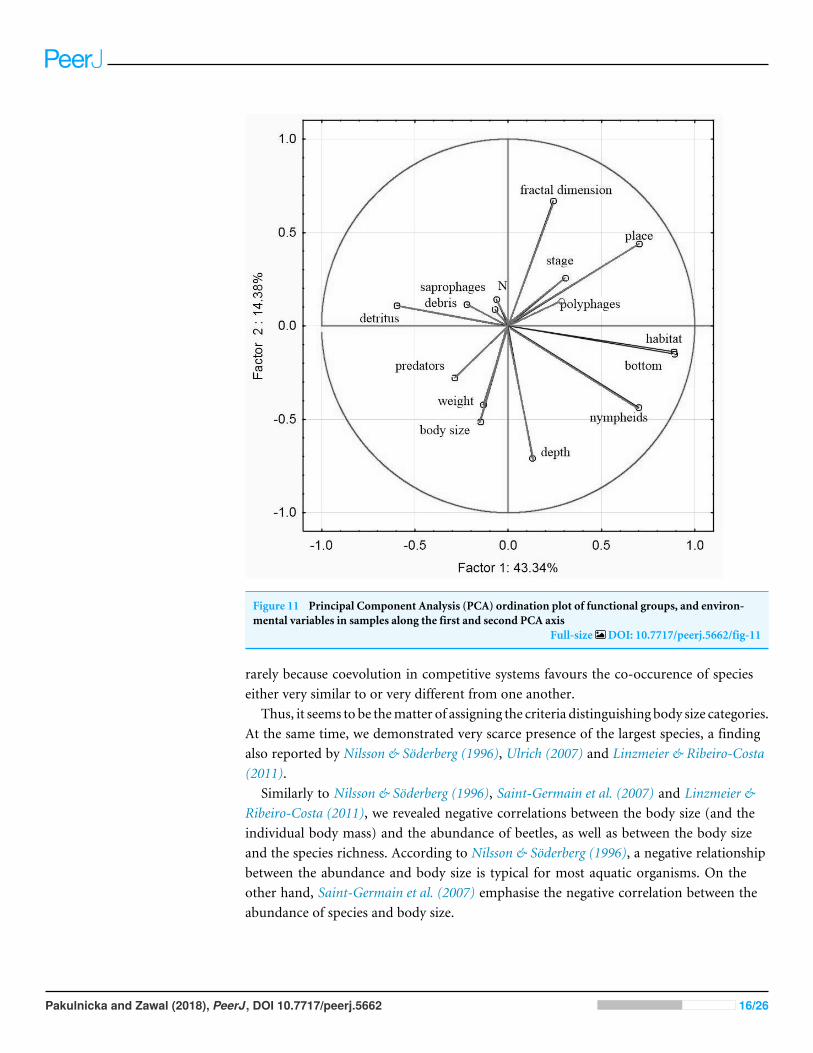
Figure 11 Principal Component Analysis (PCA) ordination plot of functional groups and environ-mental variables in samples along the first and second PCA axis
Full-size DOI 107717peerj5662fig-11
rarely because coevolution in competitive systems favours the co-occurence of specieseither very similar to or very different from one another
Thus it seems to be thematter of assigning the criteria distinguishing body size categoriesAt the same time we demonstrated very scarce presence of the largest species a findingalso reported by Nilsson amp Soumlderberg (1996) Ulrich (2007) and Linzmeier amp Ribeiro-Costa(2011)
Similarly to Nilsson amp Soumlderberg (1996) Saint-Germain et al (2007) and Linzmeier ampRibeiro-Costa (2011) we revealed negative correlations between the body size (and theindividual body mass) and the abundance of beetles as well as between the body sizeand the species richness According to Nilsson amp Soumlderberg (1996) a negative relationshipbetween the abundance and body size is typical for most aquatic organisms On theother hand Saint-Germain et al (2007) emphasise the negative correlation between theabundance of species and body size
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1626
For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

For clarification we made an in-depth analysis of the dependencies between the bodysize and the number of beetles relative to the aspects defining the complex nature of eachhabitat Such an approach enjoys a long tradition as suggested by Ulrich (2007) althoughit is rarely implemented in hydrobiological research (Cremona Planas amp Lucotte 2008Tokeshi amp Arakaki 2012 Deacutesamoreacute et al 2018) We demonstrated significant differencesbetween the abundance and species richness of beetles representing different body sizeclasses in the particular habitats Figure 5 shows that the highest abundance and speciesrichness occurred in the Sphagnummat andmostly with respect to the smallest beetles (bodysize classes 1 and 2) mainly of the genera Anacaena and Hydroporus In the same habitatthe least numerous were large beetles and the largest ones such asDytiscus or Cybister werecompletely absent A reverse situation was observed in the sandy bottom zone and in thepockets and ponds within the Sphagnum mat where the smallest beetles (class 1) were theleast numerous while the largest ones were more abundant than elsewhere Medium-sizebeetles (classes 2 and 3) eg Noterus Agabus and Ilybius were numerous in all habitatsThese relationships are explained perfectly well by the fractal structure Should we take intoconsideration an increase in the fractal dimension from the smallest one (the less complexform of a habitat) in the sandy bottom zone to the highest (the more complex form ofa habitat with increasingly small structural elements) in the Sphagnum mat then ourobservations are in accord with the ones reported by other hydrobiologists who concludethat the species diversity and density increase as the fractal dimensions increases (egVerberk et al 2001 Tokeshi amp Arakaki 2012) Meanwhile small spaces between leaves ina Sphagnum mat inhibit the presence of very large beetles a conclusion supported by theresults provided by Tokeshi amp Arakaki (2012) Similar conclusions were drawn by Schefferet al (2015) who suggest that the maximum body size is limited by the available space inwhich beetles could move
In our study negative correlations between the body size weight or abundance of waterbeetles were determined in particular habitats Considering the total biomass of beetlesin individual habitats we were unable to identify any significant differences which againagrees with the conclusions drawn by Tokeshi amp Arakaki (2012) namely that biomass doesnot change in the fractal dimensions An increase in the density of beetles is offset by adecrease in the individual size of the body which is concordant with the results of Tokeshiamp Arakakirsquos research (2012)
A very compact structure of vegetation creates niches for small beetles offering themegg-laying safety food for larvae (much humus) and a shelter frompredators (Verberk et al2001) How is the co-occurrence of these smallest beetles possible if according to Schefferet al (2015) specimens of the same size compete with one another most strongly Somehydrobiologists eg Scheffer amp Van Nes (2006) and Scheffer et al (2015) draw attentionto the fact that these organisms create self-organising assemblages the presence of whichis the result of co-evolution between potential competitors
In the trophic structure we analysed we discovered that 70 of beetles were predatorsbeing the most numerous groups in all habitats According to Bloechl et al (2010) thenumber of predatory beetles depends on the amount of their prey In our study thesmallest quantitative impact of predatory beetles (same as the number of saprophages) was
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1726
identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

identified in the Sphagnum mat There they represented classes of organisms with smallbody size egHydroporus which do not limit the presence of otherwater beetles as they useother food resources eg zooplankton smaller insects like mayflies or insect eggs (Frelikamp Pakulnicka 2015 Frelik Koszałka amp Pakulnicka 2016 Perissinotto Bird amp Bilton 2016)According to Scheffer amp Van Nes (2006) Pakulnicka et al (2013) and Scheffer et al (2015)what happens here is the evident division of functions which relies on the principle ofminimising similarity (being similar albeit different) as this minimizes competitiveness AsScheffer et al (2015) maintain these mechanisms ensure a certain measure of redundancyof similar species in the environment which is essential for the functioning of ecosystemsduring unfavourable changes
Implications of the disharmonic succession of lakes on coleopteranfaunaThe species diversity and density of organisms are the properties most often appliedto measuring both biodiversity and quality of the environment also in the context ofsuccession changes (Kowalik 1968 Ranta 1985Kordylas 1990 Linzmeier amp Ribeiro-Costa2011) According to Vamosi Naydani amp Vamosi (2007) species richness and body sizechange predictably along the spatial gradient whereas Linzmeier amp Ribeiro-Costa (2011)suggest that changes in the body size can be an important indicator while Saint-Germainet al (2007) claim that body mass is a better measure than abundance
Lake succession is a phenomenon that occurs in a given place and over a certainperiod of time and with time (in subsequent stages) habitat conditions mostly shaped bymacrophytes change as well For particular organisms optimal living conditions appearhere and now and then they disappear Thus changes in the primary structure of lakesresult in changes in the secondary structure formed by assemblages of various organismsMany ecologists including Siemann Haarstad amp Tilman (1999) Brown (2003) White etal (2007) and Linzmeier amp Ribeiro-Costa (2011) underline that body size is correlatedwith numerous morphological physiological and ecological features such as an abilityto disperse metabolic capacity digestion capacity reproduction rate and duration of ageneration as well as biodiversity Scheffer amp Van Nes (2006) claim that these characteristicsact as specific mechanisms which prevent competition between species especially oneswith similar body size
We have demonstrated that body size and body mass are also useful measurementsin investigations into the mechanisms of succession of beetles in dystrophic lakesCommunities of lake-dwelling beetles representing different seral stages are characterisedby diverse shares of species representing different body size classes This is a consequenceof changes in the fractal structure that is the representativeness of habitats (fractals) withdifferent fractal dimensions Hence in young lakes (see the Dendrogram) where the littoralzone is very modestly overgrown with plants we noted very few smallest beetles associatedwith peats (tyrphophiles) eg Anacaena Helophorus or Hydroporus which agrees withthe results reported by Bloechl et al (2010) However these beetles were very numerous inmature lakes where a Sphagnummat dominates Thus we determined negative correlationsbetween the body size and the percentage of an area overgrown with Sphagnummoss in the
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1826
littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

littoral zone of lakes as well as between the body size (andmean weight) and the abundanceof beetles in lakes Also we observed that as the fractal dimension of lakes increased themean body size (and mean weight) decreased As spaces between the habitatrsquos componentsie plants decrease in size and become more and more complex smaller organisms areevidently preferred even though larger ones may occur in such habitats as well becausethey can push away or move plant stems small shoots or leaves an observation also madebyMcAbendroth et al (2005) This could be the reason why large beetles were found in thepockets and ponds within the Sphagnum mat and in the ecotone between the Sphagnummat and the open water surface
We support the view of other researchers eg McAbendroth et al (2005) Bloechl etal (2010) Linzmeier amp Ribeiro-Costa (2011) and believe that large species gain benefitsduring the early stages of succession while smaller and specialised ones are at an advantagein the final stages We have shown (see Fig 11) that the abundance of beetles is correlatedwith the age of lakes (the seral stage) (contrary to Bloechl et al (2010) and with thefractal dimension (McAbendroth et al 2005) These relationships are connected with thedevelopment of plants which offer a shelter from predators places for laying eggs and foodsupply Like Bloechl et al (2010) and Cremona Planas amp Lucotte (2008) we demonstrateda relationship between polyphages (Haliplidae) and submerged more compact plantswhile a negative correlation was proven with nympheids which restrain the growth offood (algae) eaten by polyphages In turn the presence of saprophages is correlated withdetritus and drifters (which additionally contribute to a growth in the fractal dimension bycreating microhabitats that can be used by smaller organisms) which has been confirmedby Verberk et al (2001) and Cremona Planas amp Lucotte (2008) Moreover we found outthat the abundant presence of predatory beetles in all lakes and especially among loosevegetation is favoured by the availability of potential prey a conclusion also supported bythe findings made by Bloechl et al (2010) whereas a more compact structure is clearly abarrier to the occurrence of the largest organisms Another factor that limits the numberof beetles is the depth This suggestion is confirmed by the research completed by Brittain(1978) Pakulnicka Goacuterski amp Bielecki (2015b) Pakulnicka et al (2013) and Pakulnicka et al(2015a) Among the lakes which we analysed the ones that contained dark brown waterwith acidic reaction (Pakulnicka amp Zawal 2018) there were no fish or amphibians whichare usually considered as one of the major factors limiting the presence of water beetles(Eriksson et al 1980 Soleacute amp Roumldder 2010)
CONCLUSIONSOur research has revealed that an application of fractal concepts enables a morecomprehensive explanation regarding the character of fauna relative to the complexnatural aspects of habitats This approach is also helpful in explaining directional changesin fauna induced by the natural succession of lakes The taxonomic diversity and variousdensities of beetles in lakes increase as the fractal dimension increases There are negativecorrelations between the body size and abundance An increase in the density of beetlesin fractals with a higher dimension is offset by a lower individual body size of specimensConsequently the biomass is constant regardless of the fractal dimension
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 1926
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

ADDITIONAL INFORMATION AND DECLARATIONS
FundingThe authors received no funding for this work
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Joanna Pakulnicka conceived and designed the experiments performed the experimentsanalyzed the data contributed reagentsmaterialsanalysis tools prepared figures andortables authored or reviewed drafts of the paper approved the final draftbull Andrzej Zawal performed the experiments analyzed the data approved the final draft
Field Study PermissionsThe following information was supplied relating to field study approvals (ie approvingbody and any reference numbers)
Field studies were conducted in accordance with field study approvals Certificate ofRDOS in Olsztyn (WOPN070212018AKI) WNP certificate (PNE 08-070 18) and ZPKCertificate (06012018)
Data AvailabilityThe following information was supplied regarding data availability
The raw data are provided in Appendix S4
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5662supplemental-information
REFERENCESAndrejczukW 2014 About the landscape space Dissertations of Cultural Landscape
Commission 2445ndash64Baars JR Murray DA Hannigan E Kelly-QuinnM 2014Macroinvertebrate assem-
blages of small upland peatland lakes in Ireland Biology amp Environment Proceedingsof the Royal Irish Academy 114B233ndash248 DOI 103318BIOE201431
Barndt D 2012 Beitrag zur Kenntnis der Arthropodenfauna der ZwischenmooreButzener Bagen Trockenes Luch und Moumlllnsee bei Lieberose (Land Brandenburg)(Coleoptera Heteroptera Hymenoptera part Auchenorrhyncha Saltatoria Dipterapart Diplopoda Chilopoda Araneae Opiliones ua)Maumlrkischen EntomologischenNachrichten 14147ndash200
Barnes JB Vaughan IP Ormerod SJ 2013 Reappraising the effects of habitatstructure on river macroinvertebrates Freshwater Biology 582154ndash2167DOI 101111fwb12198
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2026
Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

Beadle JM Brown LE Holden J 2015 Biodiversity and ecosystem functioning innatural bog pools and those created by rewetting schemesWIREs Water 265ndash84DOI 101002wat21063
Bloechl A Koenemann S Philippi B Melber A 2010 Abundance diversity and suc-cession of aquatic Coleoptera and Heteroptera in a cluster of artificial ponds in theNorth German Lowlands Limnologica 4015ndash225 DOI 101016jlimno200908001
Bosi G 2001 Abundance diversity and seasonal succession of dytiscid and noteridbeetles (Coleoptera Adephaga) in two marshes of the Eastern Po Plain (Italy)Hydrobiologia 4591ndash7 DOI 101023A1012594615880
Braun-Blanquet J 1964 Pflanzensociologie Grundzuumlge der Vegetationskunde ThirdEdition Wien New York Springer-Verlag 1ndash607
Brittain JE 1978 The aquatic Coleoptera of Oslashvre Heimdalsvatn Holarctic Ecology1266ndash270
Brown JH 2003Macroecologiacutea Mexico Fondo de Cultura Econoacutemica 1ndash397Buczyński P 2012 VI 2 Aquatic beatles (Coleoptera Adephaga Hydrophiloidea
Staphylinoidea Byrrhoidea) In Kornijoacutew R Buczyński P eds Lake Skomielno(ŁęcznamdashWłodawa Lakeland Eastern Poland) Environment monograph OlsztynWydawnictwo Mantis 257ndash272
Buczyński P 2015Dragonflies (Odonata) of anthropogenic waters in middle-easternPoland Olsztyn Wydawnictwo Mantis 1ndash272
Ciecierska H 2008 Macrophytes as indicators of the ecological state of lakes InDissertations and monographs 139 Olsztyn Wydawnictwo Mantis 1ndash202
Clausen SE 1998 Applied correspondence analysis Thousand Oaks Sage Publications1ndash215
Corbet S Perrin R Hartley D Lancashire P Mace H Mc Clay A Morton J ParfittR Tomiak HWheatley KWillmer RWillows R 1980 Diel changes inplankton and water chemistry in Wicken Brickpit Hydrobiologia 74249ndash271DOI 101007BF00008758
Cremona F Planas D Lucotte M 2008 Biomass and composition of macroinvertebratecommunities associated with different types of macrophyte architectures andhabitats in a large fluvial lake Fundamental and Applied Limnology 1712119ndash213DOI 1011271863-913520080171-0119
Czachorowski S 1998 Caddisflies (Trichoptera) of Polish lakesmdashdescription of thedistribution of larvae Olsztyn Wydawnictwo WSP 1ndash155
Demetraki-Paleolog A 2012 Long term changes of planktonic rotifers of ten lakesof different trophic status Teka Komisji Ochrony i Kształtowania ŚrodowiskaPrzyrodniczego OL PAN 944ndash57
Deacutesamoreacute A Laenen B Miller KB Bergsten J 2018 Early burst in body size evolutionis uncoupled from species diversification in diving beetles (Dytiscidae)MolecularEcology 27(4)979ndash993 DOI 101111mec14492
Drinan TJ Foster GN Nelson BH OrsquoHalloran J Harrison SSC 2013Macroin-vertebrate assemblages of peatland lakes assessment of conservation value with
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2126
respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

respect to anthropogenic land-cover change Biological Conservation 158175ndash187DOI 101016jbiocon201210001
ErikssonMOG Henrikson L Nilsson B-I Nyman G Oscarson HG Stenson AELarsson K 1980 Predator-prey relations important for the biotic changes inacidified lakes Ambio 9248ndash249
Eyre MD Foster GN Foster AP 1992 Factors affecting the distribution of water beetlespecies assemblages in drains of eastern England Journal of Applied Entomology109217ndash225 DOI 101111j1439-04181990tb00043x
Fiłoc M Kupryjanowicz M 2015 Non-pollen palynomorphs characteristic or thedystrophic stage of humic lakes in the Wigry National Park NE Poland StudiaQuaternaria 3231ndash41 DOI 101515squa-2015-0003
Foster GN Eyre MD 1992 Classification ranking of water beetle communities UK natureconservation 1 Peterborough Joint Nature Conservation Committee 1ndash110
Frelik A Koszałka J Pakulnicka J 2016 Trophic relations between adult water beetlesfrom the Dytiscidae family and fly larvae from the Chironomidae family Biologia71931ndash940 DOI 101515biolog-2016-0115
Frelik A Pakulnicka J 2015 Relations between the structure of benthic macro-invertebrates and the composition of adult water beetle diets from the Dytiscidaefamily Environmental Entomology 441348ndash1357 DOI 101093eenvv113
Friday LE 1988 A key to the adults of British water beetles Field Studies 71ndash151Galewski K 1990 Chrząszcze (Coleoptera) Rodzina Kałużnicowate (Hydrophilidae)
Fauna Słodkowodna Polski Warsaw PWN 1ndash261Galewski K Tranda E 1978 Fauna słodkowodna Polski Chrząszcze (Coleoptera)
Rodziny Pływakowate (Dytiscidae) Flisakowate (Haliplidae) Mokrzelicowate (Hy-grobiidae) Krętakowate (Gyrinidae) Fauna Słodkowodna Polski Warsaw PWNmdashPoznań 1ndash396
Gioria M Bacaro G Feehan J 2010a Identifying the drivers of pond biodiversity theagony of model selection Community Ecology 11179ndash186DOI 101556ComEc11201026
Gioria M Schaffers A Bacaro G Feehan J 2010b The conservation value of farmlandponds predicting water beetle assemblages using vascular plants as a surrogategroup Biological Conservation 1431125ndash1133 DOI 101016jbiocon201002007
HansenM 1987 The Hydrophiloidea (Coleoptera) of Fennoskandia and DenmarkFauna Entomologica Scandinavica 181ndash254 DOI 101163187631287X00016
Hocking RR 1996Methods and applications of linear models Regression and theanalysis of variance Journal of the American Statistical Association 70707ndash712
HolmenM 1987 The aquatic Adephaga (Coleoptera) of Fennoskandia and Denmark IGyrinidae Haliplidae Hygrobiidae and Noteridae Fauna Entomologica Scandinavica201ndash168
Jasser I 1997 The dynamics and importance of picoplankton in shallow dystrophic lakein comparison with surface waters of two deep lakes with contrasting trophic statusHydrobiologia 34234387ndash93 DOI 101007978-94-011-5648-6_10
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2226
Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

Jeffries M 1993 Invertebrate colonization of artificial pondweeds of differing fractaldimension Oikos 67142ndash148 DOI 1023073545104
Kajak Z 1998 Hydrobiologymdashlimnology In Ecosystems of inland water bodiesWarsaw Wydawnictwo Naukowe PWN 1ndash356
Kordylas A 1990Water beetles (Coleoptera) of the lobelian Krzemno Lake FragmentaFaunistica 3371ndash81 DOI 10316100159301FF1990335071
KowalikW 1968Water beetles (Coleoptera aquatica) in the Sosnowickie Lakes in theŁęczna-Włodawa Lake District Annales Universitatis Mariae Curie-Skłodowska18283ndash300
Kuczyńska-Kippen N 2008 Spatial distribution of zooplankton communities betweenthe sphagnum mat and open water in a dystrophic lake Polish Journal of Ecology5657ndash64
Lepistouml L SauraM 1998 Effects of forest fertilization on phytoplankton in a borealbrown-water lake Boreal Environment Research 333ndash43
Linzmeier AM Ribeiro-Costa CS 2011 Body size of Chrysomelidae (ColeopteraInsecta) in areas with different levels of conservation in South Brazil In Jolivet PSantiago-Blay J Schmitt M eds Research on Chrysomelidae 3 ZooKeys 1571ndash14DOI 103897zookeys1571083
Lundkvist E Landin J Milberg P 2001 Diving beetle (Dytiscidae) assemblages alongenvironmental gradients in an agricultural landscape in southeastern SwedenWET-LANDS 2148ndash58DOI 1016720277-5212(2001)021[0048DBDAAE]20CO2
Mandelbrot B 1983 The fractal geometry of nature New York WH Freeman andCompany 1ndash468
McAbendroth L Ramsay PM Foggo A Rundle SD Bilton DT 2005 Does macrophytefractal complexity drive invertebrate diversity biomass and body size distributionsOikos 111279ndash290 DOI 101111j0030-1299200513804x
McFarland B Carse F Sandin L 2009 Littoral macroinvertebrates as indicators of lakeacidification within the UK Aquatic Conservation Marine and Freshwater Ecosystems20105ndash116 DOI 101002aqc1064
Menetrey N Sager L Oertli B Lachavanne JB 2005 Looking for metrics to assess thetrophic state of ponds Macroinvertebrates and amphibians Aquatic ConservationMarine and Freshwater Ecosystems 15653ndash664 DOI 101002aqc746
Miałdun J 2010 Effect of ground sample distance and impulse noise on fractal dimen-sion of littoral vegetation of Lake Łuknajno Archiwum Fotogrametrii Kartografii iTeledetekcji 21257ndash266
Miałdun J Ostrowski M 2010 Fractal dimension of fragments of aerial images ofshoreline zones of the Lakes Mikołajskie Śniardwy and Łuknajno ArchiwumFotogrametrii Kartografii i Teledetekcji 21267ndash279
Mieczan T Tarkowska-KukurykM PłaskaW Rechulicz J 2014 Abiotic predictors offaunal communities in an ombrotrophic peatland lagg and an open peat bog IsraelJournal of Ecology amp Evolution 6062ndash74 DOI 101080156598012014978149
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2326
Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

Miller KB Bergsten J 2016 Diving beetles of the world In Systematics and biology of theDytiscidae Baltimore Johns Hopkins University Press
Nilsson AN Soumlderberg H 1996 Abundance and species richness patterns of divingbeetles (Coleoptera Dytiscidae) from exposed and protected sites in 98 northernSwedish lakes Hydrobiologia 32183ndash88 DOI 101007BF00018680
Obolewski K Glińska-Lewczuk K Kobus S 2009 Effect of hydrological connectivity onthe molluscan community structure in oxbow lakes of the Łyna River Oceanologicaland Hydrobiological Studies 3875ndash88 DOI 102478v10009-009-0045-1
Odland A Del Moral R 2002 Thirteen years of wetland vegetation succession followinga permanent drawdown Myrkdalen Lake Norway Plant Ecology 162185ndash198DOI 101023A1020388910724
Pakulnicka J 2008 The formation of water beetle fauna in anthropogenic water bodiesOceanological and Hydrobiological Studies 3731ndash42 DOI 102478v10009-007-0037-y
Pakulnicka J BartnikW 1999 Changes in the fauna of aquatic beetles (Coleopteraaquatica) in Lake Luterskie (Olsztyn Lake District) in 1981ndash1993 FragmentaFaunistica 4271ndash93 DOI 10316100159301FF1999428071
Pakulnicka J Buczyńska E Buczyński P Czachorowski S Kurzątkowska ALewandowski K Stryjecki R Frelik A 2015a Are beetles good indicators of insectdiversity in freshwater lakes Oceanological and Hydrobiological Studies 44487ndash499DOI 101515ohs-2015-0046
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki R Szlauer-Łukaszewska A Zawal A 2016a Aquatic beetles (Coleoptera) in springs situated inthe valley of a small lowland river habitat factors vs landscape factors Knowledge andManagement of Aquatic Ecosystems 417(29)29 DOI 101051kmae2016016
Pakulnicka J Buczyński P Dąbkowski P Buczyńska E Stępień E Stryjecki RSzlauer-Łukaszewska A Zawal A 2016b Development of fauna of water beetles(Coleoptera) in waters bodies of a river valley habitat factors landscape andgeomorphology Knowledge and Management of Aquatic Ecosystems 417(40)1ndash23DOI 101051kmae2016027
Pakulnicka J Goacuterski A Bielecki A 2015b Environmental factors associated withbiodiversity and the occurrence of rare threatened thermophilous species of aquaticbeetles in the anthropogenic ponds of the Masurian Lake District Biodiversity andConservation 24429ndash445 DOI 101007s10531-014-0774-7
Pakulnicka J Goacuterski A Bielecki A Buczyński P Tończyk G Cichocka JM 2013Relationships within aquatic beetle (Coleoptera) communities in the lightof ecological theories Fundamental and Applied Limnology 183249ndash258DOI 1011271863-913520130413
Pakulnicka J Zawal A 2007Water beetles (Coleoptera) of lsquoSzare Lakersquo Reserve andwater bodies situated in its vicinity Parki Narodowe i Rezerwaty Przyrody 25121ndash133[in Polish]
Pakulnicka J Zawal A 2018Model of disharmonic succession of dystrophic lakes basedon aquatic beetle fauna (Coleoptera)Marine and Freshwater Research Epub ahead ofprint Sep 11 2018 DOI 101071MF17050
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2426
Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

Perissinotto R BirdMS Bilton D 2016 Predaceous water beetles (ColeopteraHydradephaga) of the Lake St Lucia system South Africa biodiversity communityecology and conservation implications ZooKeys 59585ndash135DOI 103897zookeys5958614
PłaskaW Kurzątkowska A Stępień A Buczyńska E Pakulnicka J Szlauer-Lukaszewska A Zawal A 2016 The effect of dredging of a small lowaland river onaquatic Heteroptera Annales Zoologici Fennici 53139ndash153 DOI 1057350860530403
PłaskaWMieczan T 2018 Effects of water bugs on crustacean zooplankton in ashallow littoral zone Knowledge and Management of Aquatic Ecosystems 419(16)1ndash7DOI 101051kmae2018012
PłaskaW Tarkowska-KukurykM 2014 Influence of abiotic factors on species spec-trum of zoopleuston in different types of peatlands Polish Journal of EnvironmentalStudies 23(2)441ndash447
Ranta E 1985 Communities of water-beetles in different kinds of waters in FinlandProceedings of the Academy of Natural Sciences of Philadelphia 13733ndash45
Saint-GermainM Buddle CM Larriveacutee M Mercado A Motchula T Reichert E SackettTE Sylvain ZWebb A 2007 Should biomass be considered more frequently as acurrency in terrestrial arthropod community analyses Journal of Applied Ecology44330ndash339 DOI 101111j1365-2664200601269x
Scheffer M Van Nes EH 2006 Self-organized similarity the evolutionary emergenceof groups of similar species Proceedings of the National Academy of Sciences of theUnited States of America 103(16)6230ndash6235 DOI 101073pnas0508024103
Scheffer M Vergnon R Van Nes EH Cuppen JGM Peeters ETHM Leijs R Nilsson AN2015 The evolution of functionally redundant species evidence from beetles PLOSONE 10(10)e0137974 DOI 101371journalpone0137974
Shadrin NV EL-Shabrawy GM Anufriieva EV Goher ME Ragab E 2016 Long-termchanges of physicochemical parameters and benthos in Lake Qarun (Egypt) can wemake a correct forecast of ecosystem future Knowledge and Management of AquaticEcosystems 417(18)1ndash11 DOI 101051kmae2016005
Siemann E Haarstad J Tilman D 1999 Dynamics of plant and arthropod diversity dur-ing old field succession Ecography 22406ndash414DOI 101111j1600-05871999tb00577x
Soldaacuten T Bojkovaacute J Vrba J Bitušiacutek P Chvojka P PapaacutečekM Peltanovaacute J Sychra JTaacutetosovaacute J 2012 Aquatic insects of the Bohemian Forest glacial lakes diversitylong-term changes and influence of acidification Silva Gabreta 18123ndash283
Soleacute M Roumldder D 2010 Dietary assessments of adult amphibians In Dodd Jr CK edAmphibian ecology and conservation a handbook of techniques Oxford UniversityPress 1ndash180
Stryjecki R Zawal A Szlauer-Łukaszewska A Michoński G Buczyńska E BuczyńskiP Pakulnicka J Śmietana P 2017Water mite (Acari Hydrachnidia) as-semblages in relation to differentiation of mesohabitats in a shallow lake andits margins Turkish Journal of Fisheries and Aquatic Sciences 17323ndash332DOI 1041941303-2712-v17_2_11
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2526
TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626

TimmHMoumlls T 2012 Littoral macroinvertebrates in Estonian lowland lakes the effectsof habitat season eutrophication and land use on some metrics of biological qualityFundamental Applied Limnology 180145ndash156 DOI 1011271863-913520120203
Tokeshi M Arakaki S 2012Habitat complexity in aquatic systems fractals and beyondHydrobiologia 68527ndash47 DOI 101007s10750-011-0832-z
UlrichW 2007 Body weight distributions of central European Coleoptera EuropeanJournal of Entomology 104769ndash776 DOI 1014411eje2007098
Usseglio-Polatera P BournaudM Tachet H 2000 Biological and ecological traits ofbenthic freshwater macroinvertebrates relationships and definition of groups withsimilar traits Freshwater Biology 43175ndash205 DOI 101046j1365-2427200000535x
Vamosi SM Naydani CJ Vamosi JC 2007 Body size and species richness along geo-graphical gradients in Albertan diving beetle (Coleoptera Dytiscidae) communitiesCanadian Journal of Zoology 85443ndash449 DOI 101139Z07-021
VerberkWCEP Van Duinen GJA Peeters TMJ Esselink H 2001 Importance ofvariation in water-types for water beetle fauna (Coleoptera) in Korenburgerveen abog remnant in the Netherlands Proceedings of the Section Experimental and AppliedEntomology of the Netherlands Entomological Society 12121ndash128
Šiling R Urbanič G 2016 Do lake littoral benthic invertebrates respond differ-ently to eutrophication hydromorphological alteration land use and fishstocking Knowledge and Management of Aquatic Ecosystems 417(35)1ndash31DOI 101051kmae2016022
White EP Ernest SKM Kerkhoff AJ Enquist BJ 2007 Relationships between bodysize and abundance in ecology Trends in Ecology and Evolution 22323ndash330DOI 101016jtree200703007
Winfield Fairchild G Faulds A Matta JF 2000 Beetle assemblages in ponds effects ofhabitat and site age Freshwater Biology 44523ndash534DOI 101046j1365-2427200000601x
Yee DA (ed) 2014 Ecology systematics and the natural history of predaceous diving beetles(Coleoptera Dytiscidae) Dordrecht Springer 1ndash468
Pakulnicka and Zawal (2018) PeerJ DOI 107717peerj5662 2626


















