
Ecotoxicologie et écologie de la réussite d’incubation...
45
1 Institut National Agronomique Paris-Grignon Université Paris-Sud 11 Université Pierre et Marie Curie Paris 6 Ecole Normale Supérieure de Paris MASTER Sciences et technologie du vivant, Mention Biologie Spécialité Ecologie, Biodiversité, Evolution (EBE) Elodie GUIRLET Stage réalisé sous la direction de Marc Girondot et Hélène Roche Juin 2005 Laboratoire Ecologie, Systématique, Evolution Université Paris Sud UMR 8079, Bât.362, 91405 Orsay Cedex Ecotoxicologie et écologie de la réussite d’incubation chez la tortue luth, Dermochelys coriacea, en Guyane française
Transcript of Ecotoxicologie et écologie de la réussite d’incubation...

- 1 - 1
Institut National Agronomique Paris-Grignon Université Paris-Sud 11 Université Pierre et Marie Curie Paris 6 Ecole Normale Supérieure de Paris
MASTER Sciences et technologie du vivant, Mention Biologie Spécialité Ecologie, Biodiversité, Evolution (EBE)
Elodie GUIRLET Stage réalisé sous la direction de Marc Girondot et Hélène Roche Juin 2005 Laboratoire Ecologie, Systématique, Evolution Université Paris Sud UMR 8079, Bât.362, 91405 Orsay Cedex
Ecotoxicologie et écologie de la réussite d’incubation chez la tortue luth, Dermochelys coriacea,
en Guyane française

- 2 - 2
Remerciements Je tiens à remercier Monsieur Pierre-Henri Gouyon, directeur du laboratoire d'Ecologie,
Systématique, Evolution et Monsieur Paul Leadley, directeur du département d'Ecologie des
populations et des communautés, de m'avoir accueillie au sein de leur équipe.
J'adresse tout particulièrement mes remerciements à Monsieur Marc Girondot, mon maître de stage,
pour m'avoir accueillie dans l'équipe de Conservation des populations et des communautés, de
m'avoir encadrée tout au long de ce stage de MASTER et de m'avoir donné l'opportunité de
découvrir la Guyane et les tortues luths.
Je remercie également Madame Hélène Roche pour tout le temps qu'elle a consacré à m'initier au
monde de l'écotoxicologie.
J'adresse également mes remerciements à Johan Chevalier qui m'a très efficacement aidé lors de ma
campagne de prélèvement et qui m'a fait découvrir la Guyane avec beaucoup de gentillesse.
Mes remerciements vont aussi à la communauté d’Awala-Yalimapo pour m’avoir accueillie lors de
mon séjour en Guyane.
Je remercie également Benjamin, pour sa patience et son écoute au téléphone.
Enfin, je remercie Audrey pour son soutien et ses conseils, tout simplement.

- 3 - 3
Table des matières
TABLE DES ILLUSTRATIONS................................................................................................. 5
LISTE DES ABREVIATIONS .................................................................................................... 6
INTRODUCTION ........................................................................................................................ 7
ETUDE BIBLIOGRAPHIQUE ................................................................................................... 9
1 POLLUANTS ORGANIQUES PERSISTANTS ET PESTICIDES ORGANOCHLORES ................................... 9
2 EFFETS DES PESTICIDES CHEZ LES REPTILES ............................................................................... 9
2.1- Les substances perturbatrices du système endocrinien (SPSE) ........................................10
2.1.1- Effets sur la croissance et la masse corporelle des jeunes à l'éclosion .......................10
2.1.2- Effets sur la reproduction .........................................................................................10
2.1.3- Altération de la détermination du sexe......................................................................11
2.2- Effet sur la réussite d'incubation .....................................................................................12
3. PROPRIETES PHYSICO-CHIMIQUES DES ORGANOCHLORES..........................................................12
3.1- Bioaccumulation.............................................................................................................13
3.2- Mobilité ..........................................................................................................................13
3.3- Persistance .....................................................................................................................13
4. DEVENIR DANS L’ENVIRONNEMENT .........................................................................................14
4.1- Dégradation....................................................................................................................14
4.2- Formation des métabolites : exemple du DDT.................................................................14
4.3- Evolution des concentrations à long terme......................................................................15
MATERIELS ET METHODES..................................................................................................16
1. LA TORTUE LUTH, DERMOCHELYS CORIACEA ............................................................................16
2. SITE D'ETUDE..........................................................................................................................16
3. CARTOGRAPHIE DE LA PLAGE ..................................................................................................17
3.1- Géoréférencement...........................................................................................................17
3.1.1- Carte IGN de Ya:lima:po .........................................................................................17
3.1.2- Photo aérienne de la plage : calage affiné .................................................................18
3.2- Calcul des coordonnées des nids étudiés et des points de prélèvements de sable .............18
4. ANALYSE DES PESTICIDES DANS LE SABLE ...............................................................................19

- 4 - 4
4.1- Prélèvements ..................................................................................................................19
4.2- Matériel chimique : les pesticides analysés .....................................................................19
4.3- Extraction des lipides et des pesticides organochlorés lipophiles ....................................20
4.4- Purification (Clean-up)...................................................................................................20
4.5- Détermination et quantification des micropolluants par chromatographie ......................21
4.6- Distribution spatiale des pesticides : test de Mantel ........................................................22
5. DONNEES RELATIVES AU SUIVI DES NIDS..................................................................................22
6. ANALYSE STATISTIQUE ...........................................................................................................23
RESULTATS ...............................................................................................................................24
1. ANALYSE SPATIALE : POSITION DES PRELEVEMENTS ET DES NIDS SUR LA PLAGE .......................24
2. ANALYSE DES PESTICIDES DU SABLE DE LA PLAGE D’AWA:LA-YA:LIMA:PO ..............................25
2.1- Pesticides identifiés.........................................................................................................25
2.2- Test de Mantel ................................................................................................................27
3. DONNEES RELATIVES AU SUIVI DES NIDS..................................................................................28
3.1- Composition de la ponte et réussite de l'incubation des œufs à Ya:lima:po......................28
3-2 Position du nid sur la plage .............................................................................................28
3.3- Fréquentation de la plage par les femelles ......................................................................29
3.4- Taille des femelles...........................................................................................................30
3.5- Analyse des facteurs ayant un effet sur la réussite d'incubation.......................................30
3.5.1- Modèle binomial ......................................................................................................30
3.5.2- Modèle multinomial.................................................................................................31
3.6- Distribution de la réussite d’incubation ..........................................................................32
DISCUSSION ..............................................................................................................................33
CONCLUSION............................................................................................................................37
REFERENCES BIBLIOGRAPHIQUES....................................................................................38
ANNEXES....................................................................................................................................41

- 5 - 5
Table des illustrations Liste des figures
Figure 1 : Voies de dégradation du DDT dans l'eau et les sédiments. .............................................15
Figure 2 : Tortue luth, Dermochelys coriacea ................................................................................16
Figure 3 : Réserve naturelle de l'Amana en Guyane française.........................................................17
Figure 4 : Carte de la portion de plage étudiée avec les positions des prélèvements de sable et des
nids........................................................................................................................................24
Figure 5 : Proportion des différentes catégories d'œufs dans le nid à la fin de la période d'incubation
..............................................................................................................................................28
Figure 6 a et b : Fréquence de distribution des nids par rapport à la ligne de marée haute (a) et à la
ligne de végétation du bord de plage (b).................................................................................29
Figure 7 : Fréquence de distribution du nombre de pontes pour la saison 2002...............................29
Liste des tableaux
Tableau 1 : Niveaux de contamination du sable par les pesticides identifiés dans les échantillons ..26
Tableau 2 : Niveaux d'imprégnation du sable pour les groupes de pesticides (molécule mère +
métabolites) dont les concentrations sont supérieures à 500ng/kg de sable sec .......................27
Tableau 3 : Résultats du test de Mantel pour les six groupes de pesticides retenus .........................27
Tableau 4 : GLM (Generalised Linear Model) pour le modèle œufs éclos (A), œufs avortés (B),
œufs prédatés par les courtilières (C), œufs prédatés par les crabes (D) et œufs pourris (E). ...31
Tableau 5 : résultat du modèle multinomial : vraisemblance, AIC, c et QAIC pour le modèle saturé
le modèle complet et les 21 modèles contraints. .....................................................................32

- 6 - 6
Liste des abréviations AIC : Akaike Information Criterion
ASE : Accelerated Solvant Extraction
CPG : Chromatographie en Phase Gazeuse
DDASS : Direction Départementale de l'Action Sanitaire et Sociale DDD : dichloro-diphényl-dichloéthane DDE : dicholoro-diphényl-dichloroéthylène DDT : dichloro-diphényl-trichloroéthane
DIREN : Direction Régionale de l'Environnement
DistMH : distance à la ligne de marée haute
DistVeg : distance à la ligne de végétation
ECD : détecteur à capture d’électron
EE : oestrogènes Environnementaux
ET : Ecart-Type
GLM : Modèle Linéaire Général
GPS : Global Positionning System
HCH : hexachlorocyclohexane
IGN : Institut Géographique National
LMH : ligne de marée haute
LV : ligne végétation
MB : milieu de plage bas
MH : milieu de plage haut
MO : Matière Organique
Ninf : Nombre d'œufs sans jaune
Ocs : OrganoChlorés
PCB :Polychloro-biphényles
PIT : Passive Integrated Transponder
POPs : Polluants Organiques Persistants
QAIC : Quasi Akaike Information Criterion
RGFG : Réseau Géodésique Français de Guyane
SPSE : Substances Perturbatrices du Système Endocrinien
TSD : Temperature-dependent Sex Determination

- 7 - 7
Introduction
Les pesticides organochlorés synthétiques ont été largement utilisés dès les années 1940. Leur
utilisation est répandue en raison de leurs avantages dans le domaine agricole (accroissement des
rendements des cultures) ainsi que pour la protection de la santé publique (lutte contre les insectes
vecteurs de maladies). Toutefois, une vingtaine d’années après le début de leur utilisation intensive
à grande échelle, des problèmes environnementaux ont commencé à être observés. L’utilisation
massive des pesticides et l’évolution permanente de leurs caractéristiques chimiques constituent en
effet une menace pour la qualité de l’environnement et pour les écosystèmes aquatiques en
particulier (Voldner et Li 1995).
La présence de DDT et de ses métabolites a été détectée par hasard récemment sur les plages de
Guyane française. Ces produits sont surtout utilisés pour leur efficacité dans la lutte contre les
insectes ravageurs des cultures ou vecteurs de maladies (paludisme). Toxiques pour les organismes,
ces produits présentent en plus la particularité d’être féminisant chez les reptiles (Crain et al. 1997 ,
Willingham et Crews 1999).
Or le littoral de la Guyane est le plus gros site de ponte au monde de la tortue luth, Dermochelys
coriacea. Près de la moitié des femelles de l'espèce vient pondre, de mars à août, sur les plages de
Guyane et du Surinam (Spotila et al. 1996). La plage de Awa:la-Ya:lima:po, située à l’embouchure
des fleuves Mana et Maroni dans l’Ouest guyanais recevait à elle seule, à la fin des années 80, 90 à
95% des femelles nidifiant en Guyane française (Girondot et Fretey 1996).
Cependant, des estimations des effectifs de femelles de la population du Pacifique ces vingt
dernières années, ont montré un fort déclin dans plusieurs lieux de nidification. La population
Atlantique semblerait plus stable, bien que le nombre de femelles nidifiant soit variable d'une année
à l'autre, rendant les tendances difficiles à dégager. Etant donné l'importance de cette population, sa
viabilité est d'intérêt majeur pour la persistance de l’espèce au niveau mondial. L'étude et la
compréhension des menaces qui pèsent sur cette population, pour l'instant stable, sont donc
primordiales (Spotila et al. 2000).
Les pontes de tortues luths ont un succès d’incubation moins élevé que les autres tortues marines,
sans que les causes de cette forte mortalité embryonnaire ne soient identifiées (Wallace et al. 2004).
A Awa:la-Ya:lima:po, la productivité est très faible avec un taux de réussite d’incubation
(pourcentage des œufs fertiles donnant un nouveau-né à l’émergence) inférieur à 40 % (Girondot et
al., sous presse). Or cette proportion peut atteindre sur d'autres plages de ponte 40 % à 75 % (Leslie
et al. 1996) et jusqu'à 90 % en conditions expérimentales. Le recrutement au niveau de cette plage
est donc très faible. Dans la mesure où la tortue luth pourrait être l’espèce de tortue marine la plus
menacée (Spotila et al. 1996) et que la plage de Yalimapo est la plus fréquentée pour cette espèce, il

- 8 - 8
est important de s’interroger sur l’origine de cette faible réussite d’incubation, dans une optique de
conservation de l’espèce.
Plusieurs facteurs ayant un effet sur le succès de la ponte ont déjà été identifiés ; ainsi, la position
du nid par rapport à la ligne de végétation et la ligne de marée haute ou le nombre d’œufs sans
jaune (longtemps appelés œufs infertiles) influencent le taux de prédation des œufs (Caut et
Girondot, non publié). D’autres facteurs physico-chimiques pourraient affecter l’environnement du
nid et influencer l’incubation des œufs comme l’humidité ou la taille des grains de sable (Wallace et
al. 2004). Il est donc intéressant de continuer à s’interroger sur l’impact de certains facteurs
notamment les pesticides, sur la réussite de la ponte.
La connaissance du taux de réussite des pontes et la compréhension de son faible niveau à
Ya:lima:po pourrait également aider à envisager et gérer la revendication des Amérindiens au sujet
d'un éventuel prélèvement d'œufs à des fins de consommation. En effet si le taux de succès de
l'incubation est aussi faible, un prélèvement d'œufs choisis parmi ceux qui n'ont que peu de chances
d'arriver à terme ne peut mettre en péril la population de tortues luths.
L’équipe de Conservation des Populations et des Communautés de l’Université Paris Sud, CNRS et
ENGREF travaille sur la biologie de la conservation de la tortue luth en tentant de cerner la
démographie de l’espèce. C’est dans ce contexte que nous nous sommes intéressés au
fonctionnement de cette plage de pontes afin d’évaluer la réussite d’incubation et les facteurs qui
l’influencent.
Cette étude a pour objectif :
- d'évaluer les concentrations de pesticides sur la plage d'Awa:la-Ya:lima:po par analyse
chromatographique,
- de construire une carte de la distribution spatiale des concentrations en pesticides sur la plage,
en utilisant des cartes géoréférencées à partir d’un logiciel de Système d’Informatique
Géographique,
- de déterminer quels sont les facteurs pouvant expliquer une partie de la variabilité du succès des
pontes.

- 9 - 9
Etude bibliographique
1 Polluants organiques persistants et pesticides organochlorés
Les polluants organiques persistants (POPs) forment un groupe de produits chimiques qui se
dégradent lentement dans l’environnement (faible biodégradabilité), s’accumulent dans les
organismes (bioaccumulation) et le long des chaînes alimentaires (bioamplification), possèdent des
propriétés toxiques à faible dose et sont omniprésents dans l’environnement. Parmi les POPs
existants, cette étude s’intéresse aux pesticides organochlorés en particulier. Un pesticide est une
substance ou une préparation utilisée pour lutter contre des êtres vivants considérés comme
nuisibles. Les pesticides organochlorés représentent une catégorie extrêmement vaste puisqu’elle
prend en compte tout dérivé organique dans lequel un ou plusieurs atomes d’hydrogène ont été
remplacés par des atomes de chlore.
Synthétisés à partir des années 40, les pesticides organochlorés ont été massivement utilisés dans
l’agriculture pour le traitement des cultures, la prévention de la détérioration des récoltes après
moisson et lors du stockage et pour la lutte contre certains vecteurs de maladies. Les membres de ce
groupe présentent généralement une très grande stabilité chimique et leur accumulation progressive
dans l'environnement a entraîné leur interdiction dans la plupart des pays. Ils restent cependant
largement utilisés dans certaines régions tropicales pour le contrôle des insectes (Bidlan 2002). Des
doses considérables ont été et sont encore largement déversées dans l’environnement. Fortement
persistants, ils sont aujourd’hui de redoutables polluants des sols et des milieux aquatiques. Ces
produits chimiques ont été associés à toutes sortes d’effets néfastes sur la santé d’organismes
non-cibles, notamment des déficiences reproductives chez les animaux et le cancer chez les
humains. L’effet toxique des composés organochlorés semble être lié à leur déchlorination
graduelle dans le corps (réaction en chaîne) et à la formation de radicaux libres interférant avec la
structure cellulaire (Veningerova et al. 1997).
2 Effets des pesticides chez les reptiles
Il y a eu récemment un grand intérêt porté aux nombreux xénobiotiques largement répandus dans
l'environnement. Il a été mis en évidence que des agents environnementaux (synthétiques ou
naturels) avaient le potentiel de perturber la fonction endocrine d'un large éventail d'espèces en
milieu naturel, entraînant tout une gamme d'effets néfastes. Les reptiles pourraient être plus exposés
à ces dangers car étant au sommet de la chaîne-alimentaire, ils accumuleraient les polluants.

- 10 - 10
2.1- Les substances perturbatrices du système endocrinien (SPSE) Les substances perturbatrices du système endocrinien sont des substances qui peuvent causer des
effets néfastes en interférant d'une manière ou d'une autre avec les hormones du corps ou les
messagers chimiques. Comme les hormones jouent un rôle crucial dans la différenciation des
cellules à des stades précoces, une exposition à ces substances dans l'œuf peut altérer le processus
normal de développement (Lyons 1999).
Ces substances comprennent une grande variété de contaminants environnementaux qui peuvent
avoir divers effets sur la croissance, le développement et la reproduction. Ces effets peuvent se
manifester à des concentrations extrêmement faibles et s’exprimer dans la génération suivante, bien
après l’exposition initiale. Les pesticides organochlorés peuvent affecter la fonction endocrine en se
comportant comme des antagonistes d'hormones sur les récepteurs ou en inhibant les enzymes
responsables de la synthèse des hormones ou de leur dégradation (Vonier et al. 1996). Un
dérèglement de la fonction endocrine n'est cependant pas une surprise étant donné qu'un certain
nombre de pesticides ont été synthétisés pour fonctionner comme des régulateurs d'hormones et de
croissance pour contrôler les populations d'espèces nuisibles (Crisp et al. 1998).
2.1.1- Effets sur la croissance et la masse corporelle des jeunes à l'éclosion
Les pesticides peuvent affecter la masse du jeune à l'éclosion, la manière d'utiliser les réserves de
l'œuf et le taux de croissance des jeunes. Chez Trachemys scripta elegans, des expositions à des
doses moyennes de pp'DDE et de tansnonachlor ont provoqué une diminution de la masse
corporelle des jeunes pendant le premier mois après l'éclosion (Willingham 2001).
2.1.2- Effets sur la reproduction
Si l'organisme est exposé à des pesticides mimant l'action d'hormones sexuelles, les effets peuvent
toucher le comportement sexuel ou causer des anormalités de l’appareil reproducteur, incluant des
individus inter sexués ainsi que des effets sur le ratio des sexes à la naissance par avortement
sélectif ou perturbation de la détermination du sexe.
L'altération du développement des organes reproducteurs (taille réduite) ainsi que des caractères
sexuels secondaires peut être causée par le DDE, le DDT, le transnonachlor, le dicofol et par
l'atrazine chez l'alligator commun, Alligator mississipiensis (Crain et al. 1997 , Portelli et al. 1999).
Le DDE a aussi un effet chez Chelydra serpentina serpentina sur la longueur précloacale (longueur
entre le cloaque et le plastron, qui correspond à l'emplacement du penis). Les adultes mâles de lacs
contaminés près des grands lacs aux Etats-Unis ont des longueurs précloacales significativement
plus petites que les individus de sites non contaminés. Dans les régions contaminées, les
observateurs ne peuvent plus identifier correctement le sexe en se basant seulement sur ce caractère
sexuel secondaire (Solla et al. 1998). Une réduction de la fertilité et une altération des

- 11 - 11
comportements sexuels des adultes ont également été reportées chez Alligator mississipiensis
(Guillette et al. 1994).
2.1.3- Altération de la détermination du sexe
2.1.3.1- Détermination du sexe chez les reptiles
De nombreux reptiles présentent une sensibilité de la détermination du sexe à la température
d'incubation des oeufs (TSD pour temperature-dependent sex determination). La température joue
un rôle dans la production d’oestradiol et d’oestrone, hormones féminisantes (oestrogènes). Par
exemple, chez les tortues, des mâles sont produits à basses températures et des femelles à hautes
températures. En effet, à des niveaux faibles d'oestradiol, les testicules se différencient et le cortex
ovarien ne se développe pas. A des niveaux plus élevés, le développement des testicules est inhibé
et le cortex ovarien est stimulé. La quantité d'oestradiol et d’oestrone endogènes dépend de l'activité
de l'aromatase. Cette enzyme convertit la testostérone en oestradiol et l’androstenedione en
oestrone. Or la régulation de l'expression de l'aromatase est sensible à la température ; en effet, il est
suggéré qu'un facteur thermosensible intervient, directement ou indirectement, dans la régulation de
la transcription du gène de l'aromatase chez les reptiles à TSD. A faible température, le niveau
d'aromatase reste faible, donc le niveau d'oestrogènes aussi et il y a production de mâles.
Inversement, à haute température, il y a production de femelles. On parlera alors de températures
masculinisante ou féminisante (Pieau et al.).
Les reptiles pourraient donc être sensibles aux composés exogènes qui miment ou altèrent le milieu
œstrogénique de l'embryon pendant son développement (Guillette et Iguchi 2003). Il en résulte
parfois des féminisations d'individus ayant été incubés à des températures masculinisantes.
2.1.3.2- Mécanismes d'action
Ces pesticides peuvent agir de différentes manières. Ils peuvent se lier aux récepteurs hormonaux et
mimer l'hormone (action agoniste), ou bloquer l'action de cette hormone (action antagoniste). Ils
peuvent aussi stimuler ou inhiber les enzymes responsables de la synthèse ou de la dégradation de
ces hormones et donc entraîner une augmentation ou une diminution de l'action de l'hormone.
Les composés qui imitent le phénomène dépendant des oestrogènes sont appelés oestrogènes
environnementaux (EE). L'exposition des embryons à des composés chimiques exogènes peut donc
mimer les effets de la température sur la détermination du sexe. Le risque qu'induirait alors la
présence des pesticides est donc un sexe ratio biaisé vers les femelles.
Chez Alligator mississipiensis, le DDD, le DDT, le transnonachlor, le dicofol et l'atrazine ont été
identifiés comme étant capables de ce mimétisme (Crain et al. 1997). Chez Trachemys scripta
elegans, le transnonachlor et le chlordane sont également capables de mimer les oestrogènes
causant de la même manière des féminisations (Willingham et Crews 1999). Cependant, chez

- 12 - 12
Chelydra serpentina serpentina, le pp'-DDE, à des taux qui existent dans la nature, ne cause pas de
féminisation pendant le développement embryonnaire lors d’incubation à des températures très
masculinisantes.
Il serait utile de connaître les concentrations pour lesquelles des féminisations commencent à être
observées, pour évaluer la menace potentielle de ces contaminants plus précisément.
2.2- Effet sur la réussite d'incubation Les embryons exposés dans les œufs via une contamination maternelle ou exposés constamment à
un environnement contaminé sont plus à risque.
Les œufs incubant dans un nid dont l'environnement est pollué par des pesticides tels que le DDE,
le chlordane, la dieldrine ou encore le toxaphène et ses métabolites sembleraient montrer une
mortalité des embryons plus forte et donc un succès de ponte plus faible chez Alligator
mississipiensis (Rauschenberger et al. 2004) et chez Chelydra serpentina serpentina (Bishop et al.
1998). Chez Alligator mississipiensis, le fait que les contaminants contribuent largement à la faible
viabilité des œufs a été mis en évidence suite à un déversement accidentel de produits chimiques
dans le lac Apopka en 1980, qui a entraîné un rapide déclin dans la population de juvénile et dans la
viabilité des œufs (Guillette et al. 1994).
En plus des pesticides qui se trouvent dans l'environnement du nid, il peut également y avoir un
transfert de contaminants de la mère vers l'œuf en développement. L'exposition maternelle peut
donc aussi être associée à une réduction du succès de la ponte et une augmentation de la mortalité
embryonnaire (Rauschenberger et al. 2004).
3. Propriétés physico-chimiques des organochlorés
La plupart des polluants arrivent au sol où leur comportement chimique va définir leur dispersion
vers d'autres compartiments de l'environnement. Le sol occupe une position centrale dans la
régulation des polluants avec un double rôle d'épuration et de stockage des polluants. Il va pouvoir
participer à leur élimination, leur transformation vers d’autres espèces chimiques plus ou moins
toxiques, diminuer ou retarder leurs impacts sur les différentes cibles.
Les propriétés physico-chimiques des pesticides vont déterminer ce comportement dans
l’environnement. Différents processus vont entraîner leur transport (par les végétaux, la faune, par
lixiviation, lessivage, ruissellement), leur rétention (absorption par les végétaux ou par la microflore
du sol, adsorption-désorption) et leur dégradation (métabolisme, photolyse, catalyse) (Barriuso et
al. 1996).

- 13 - 13
3.1- Bioaccumulation Le statut de substances lipophiles des composés organochlorés leur confère la capacité de se
bioaccumuler dans les organismes vivants, où leur concentration dépasse souvent celle du milieu
extérieur. Ils peuvent ensuite être transférés par voie alimentaire dans les chaînes trophiques. Il en
résulte que la concentration en organochlorés augmente au fur et à mesure qu’ils circulent vers les
maillons supérieurs d’un réseau trophique ; il s’agit de biomagnification. Au niveau des prédateurs
supérieurs, les concentrations peuvent alors atteindre des niveaux considérables, voire toxiques.
3.2- Mobilité La diminution de la charge des pesticides des sols peut se faire par voie atmosphérique via la
volatilisation, qui est largement influencée par la température, les propriétés physico-chimiques du
composé et celles du sol, et par les conditions météorologiques (vent, pression). L’exposition des
sols aux rayons lumineux du soleil et au vent augmente les pertes volatiles de DDT (Samuel et
Pillai 1989). Une fois dans l’atmosphère, les POPs peuvent alors être transportés sur de longues
distance avant de se déposer ; il en résulte une distribution globale de ces composés (Ockenden et
al. 2003).
Les pesticides peuvent également être transportés par les cours d’eau ; les précipitations lessivent
les sols et entraînent les composés vers le réseau hydrique qui les acheminent jusqu’aux océans,
sorte de réservoir, où ils s’accumulent (Racke et al. 1997).
L’utilisation des pesticides dans les cultures ouvertes facilite leur transport par l’air, l’eau et les
particules du sol vers les eaux de surface ou souterraines (Shimizu et al. 2005). Ils sont alors
devenus des contaminants ubiquistes, particulièrement dans l’environnement marin, qui est l’étape
finale de leur cheminement (Aguilar et Borrell 2005). Le DDT en particulier, peut être détecté,
avec ses produits de dégradation, dans l'eau, l'air et le sol (Bidlan, 2002). En Antarctique, zone
pourtant très éloignés des zones de contamination, de nombreux OCs ont été détectés chez plusieurs
espèces d’oiseaux marins (fulmar, pétrel, manchot) révélant l’étendue de la distribution des
composés (VanderBrink et DeRuiter-Dijkman 1997).
3.3- Persistance Outre la toxicité propre du polluant, sa rétention par le sol et sa persistance sont les deux facteurs
fondamentaux conditionnant le caractère polluant et/ou sa manifestation. La rétention par le sol est
le résultat d'un ensemble de phénomènes impliquant des interactions avec les constituants
organiques et minéraux des sols. La persistance, elle, est la résultante d'un ensemble de processus
de dissipation, physico-chimiques et biologiques, qui font diminuer la concentration du polluant.

- 14 - 14
La rétention conditionne la disponibilité des polluants et peut jouer un rôle de protection à l'égard
de la dégradation biologique. Parmi les caractéristiques du sol, la teneur en matière organique est le
paramètre le mieux corrélé avec les coefficients d'adsorption. L'augmentation de la rétention des
polluants sur la phase solide du sol diminue les risques de dispersion du polluant mais peut rendre
difficile sa complète élimination (Barriuso et al. 1996).
La persistance est exprimée par la demi-vie du polluant (temps nécessaire à la dégradation de la
moitié de la quantité du composé). Le DDT est considéré comme l’un des contaminants
organochlorés les plus dangereux à cause de sa longue persistance ; en effet, après 15 ans, plus de la
moitié des quantités de DDT demeure encore dans les sols traités par cet insecticide (Serrano et al.
2003).
4. Devenir dans l’environnement
4.1- Dégradation Dès leur arrivée dans le sol, les pesticides se distribuent dans trois phases : solide, liquide, vapeur.
La concentration du polluant dans chacune des phases n'est pas figée dans le temps, elle évolue en
fonction des conditions physico-chimiques (température, pH, humidité). La part de polluant la plus
mobile est celle localisée dans les phases gazeuse et liquide, et constitue la part de polluant
disponible. Ces produits disponibles pourront être transformés, ou entraînés vers les profondeurs.
Leur transformation peut être due à des phénomènes abiotiques ou biologiques. Normalement, la
dégradation biologique est quantitativement plus importante que les transformations abiotiques. La
dégradation des polluants s'accompagne de l'apparition de métabolites, avec un changement de la
structure chimique, ce qui provoque des modifications de leur toxicité et de leur comportement dans
les sols par rapport à celui de la molécule mère. D'un point de vue chimique, la plupart des
transformations abiotiques et biologiques obéissent aux mêmes types de réaction: oxydation,
réduction, hydrolyse, déchlorination. Ces réactions seront initiées ou catalysées par les constituants
organiques ou minéraux du sol ou par les systèmes enzymatiques cellulaires (Barriuso et al. 1996).
4.2- Formation des métabolites : exemple du DDT
Le DDD est le métabolite majeur dans les sédiments ; il se forme par déchlorination dans les
sédiments en condition anaérobie (Kale et al. 1999). Le DDE se forme plutôt dans l’eau et en
condition aérobie. D’autres transformations existent à partir de ces deux premiers métabolites. Le
DDMU dérive du DDE par déchlorination réductive dans les sédiments marins et le DDA est formé
par biodégradation à partir du DDD. Le DDA peut à son tour se dégrader en DDM et le DDMU en
DBP. La formation de DDA et DDM intervient quand le potentiel redox est similaire à celui du

- 15 - 15
DDE et celle de DDMU et DBP quand le potentiel redox est similaire à celui du DDD (Wan et al.
2004). La synthèse de ces réactions est montrée sur la Figure 1.
Figure 1 : Voies de dégradation du DDT dans l'eau et les sédiments ; dégradation directe ,- - > dégradation en passant par d'autres métabolites.
4.3- Evolution des concentrations à long terme En région tropicale, les conditions de température et de radiations lumineuses sont favorables à la
dégradation rapide du DDT et de ses métabolites (Samuel et Pillai 1989) mais les apports continuels
de ce polluant peuvent freiner la diminution de la charge dans l'environnement. Il est donc
important de suivre ce polluant dans les différents compartiments de l'environnement, ainsi que ses
produits de dégradation. Le ratio DDT/DDE peut être utilisé pour une estimation grossière de la
période d’utilisation du DDT (Kale et al. 1999). La dégradation progressive du DDT et l’absence
de nouvel apport dans l’environnement étudié entraîneront une augmentation relative de DDE et le
ratio DDT/DDE diminuera. Ce rapport est utilisé comme indicateur de la dégradation du DDT et
permet de savoir si l’apport est récent ou non (Aguilar et Borrell 2005).

- 16 - 16
Matériels et méthodes 1. La tortue luth, Dermochelys coriacea
La tortue luth est la plus grande des sept espèces de tortues marines (figure 2). Sa masse corporelle
est habituellement inférieure à 500 kg pour une longueur de carapace d'environ 160 cm. Elle est la
seule représentante de la famille des Dermochelidae, avec sa pseudo-carapace recouverte d’une
peau lisse et brillante. Les femelles effectuent de grandes migrations avant de venir pondre tous les
deux ou trois ans sur les plages tropicales.
En Guyane, la saison de ponte s’étale de mars à août ; les femelles pondent en moyenne 115 œufs
par nid dont 80 environ sont fertiles Le nombre de ponte par femelle et par saison peut être
supérieur à 10, avec une dizaine de jours d’intervalle entre chaque ponte (Bell et al. 2003). Le nid
est creusé dans le sable à 80 cm de profondeur. L’incubation dure entre 50 et 70 jours selon la
température du nid. Durant ces deux mois, les œufs sont exposés à de nombreux risques liés à la
présence de prédateurs, notamment les courtilières, orthoptères de la famille des Gryllotalpidae
(Maros et al. 2003), et les crabes, ou à des conditions physico-chimiques sub-optimales (humidité,
granulométrie) qui influencent la réussite d'incubation.
Figure 2 : Tortue luth, Dermochelys coriacea
2. Site d'étude
La Guyane se situe entre le 2e et le 6e degré de latitude nord, au Nord Est de l’Amérique du Sud. Le
fleuve Maroni définit la frontière avec le Surinam au Nord Ouest, et l’Oyapock, celle avec le Brésil
au Sud Est. Le climat est de type tropical humide. La température moyenne annuelle dans la journée
est de 27°C. Les pluies (de 2500 à 4000 mm/an) diminuent d'intensité vers l'Ouest. Le trait essentiel
est évidemment l'humidité relative dont le taux descend rarement en dessous de 80 %.

- 17 - 17
Située dans l'Ouest guyanais entre l’embouchure du fleuve Maroni et celle de la rivière Mana (05°
44' N, 53° 56' W), la plage de Ya:lima:po est actuellement le site de ponte le plus important au
monde pour les tortues luths (Girondot et Fretey 1996). La zone littorale est riche en sédiments
provenant de l’Amazone. Ils sont charriés par le courant des Guyanes le long de la côte, dont le
profil se modifie assez rapidement. Cependant, la plage de Ya:lima:po est relativement stable,
comparée aux autres de la région (Lefebvre et al. 2004) et actuellement, les tortues marines
l’utilisent activement pour y creuser leurs nids.
La plage s’étend sur une longueur de 4 km environ, sa largeur varie selon les années. Elle fait partie
de la réserve naturelle de l’Amana, créée en 1998, qui couvre une superficie de 14 800 hectares et
qui s’étend de l’embouchure du Maroni jusqu’à celle de l’Organabo. Ce milieu naturel protégé est
bordé de rizières et de cultures maraîchères (Figure 3).
Figure 3 : Réserve naturelle de l'Amana en Guyane française.
3. Cartographie de la plage
3.1- Géoréférencement 3.1.1- Carte IGN de Ya:lima:po
La carte IGN n°4701, utilisée comme référence pour la zone étudiée, est géoréférencée dans le
système CSG 1967 (Centre Spatial Guyanais 1967) qui est une réalisation bidimensionnelle
effectuée par mesure angulaire en 1967. Pour faire face à l’émergence du positionnement satellitaire
par GPS (Global Positionning System), l’IGN (Institut Géographique National) a mis en place un
nouveau canevas local, le RGFG 95 (Réseau Géodésique Français de Guyane). Ce système
correspond à la détermination d’un canevas GPS de 26 points, homogène et précis réalisé en 1995
(exactitude de l’ordre du centimètre).

- 18 - 18
Le logiciel CIRCE 2000 Antilles-Guyane a été utilisé pour transformer les coordonnées longitude-
latitude de la carte IGN (CSG 67) en RGFG 95. La carte IGN servant de référence a alors pu être
géoréférencée avec ces nouvelles coordonnées à l’aide du logiciel ArcMap (ArcGis 8.3)
3.1.2- Photo aérienne de la plage : calage affiné
Nous disposons d’une photo aérienne prise en octobre 2001 sur laquelle figurent la plage de
Awa:la-Ya:lima:po, la ligne de végétation le long de cette plage, les villages de Ya:lima:po et
d’Awa:la et la zone de prélèvement des échantillons de sable. Pour la géoréférencer, la photo est
calée sur la carte IGN de référence, en utilisant des éléments caractéristiques du paysage (carrefour,
bâtiments, végétation). Ce calage est également réalisé à l’aide d’ArcMap et permet d’obtenir une
photo aérienne géoréférencée dans le système RGFG 95.
3.2- Calcul des coordonnées des nids étudiés et des points de prélèvements de sable La position des prélèvements de sable et des nids sur la portion de plage de 300 mètres étudiée a été
définie par triangulation lors de l’étude en 2002. Une carte où figurent ces emplacements ainsi que
la ligne de végétation a été réalisée à la main sur le terrain. Pour positionner les nids et les
prélèvements sur la photo, il a fallu transformer les coordonnées de la carte en coordonnées RGFG
95. Pour cela, la ligne de végétation, figurant sur le relevé de triangulation et sur la photo, a servi de
point de repère. Des points caractéristiques visibles à la fois sur le relevé et la photo ont été utilisés
pour construire un modèle qui permet de passer, pour un point donné, des coordonnées de la carte
de terrain (XRELEVE , YRELEVE) à de nouvelles coordonnées dans le système RGFG 95 (XCALCULE ,
YCALCULE ).
XCALCULE = a YRELEVE + b XRELEVE + c
YCALCULE = a’ XRELEVE + b’ YRELEVE + c’
Pour chaque point de la végétation utilisé, les valeurs suivantes sont calculées :
!Ln L = Ln " + 0, 5Ln 2#( ) +XPHOTO
! XRELEVE( )
2
2"2
!Ln L = Ln " + 0, 5Ln 2#( ) +YPHOTO
!YRELEVE( )
2
2"2
Les paramètres a, b, c, a’, b’, c’ du modèle sont estimés en minimisant la somme des Ln L! de
chaque point, c'est à dire en maximisant l'ajustement des coordonnées du relevé à celles de la photo.
Le modèle le mieux ajusté est celui qui a l'AIC le plus faible, avec AIC = -2 Ln L + 2K, où K est le
nombre de paramètres.

- 19 - 19
4. Analyse des pesticides dans le sable
4.1- Prélèvements Les prélèvements de sable ont été effectués sur une portion de 300 m de plage, suffisamment
fréquentée par les tortues. Tous les 20 m, le sable est prélevé à quatre niveaux de la plage le long
d'un transect linéaire pour échantillonner sur toute la largeur de la plage : ligne de végétation, ligne
de marée haute et deux autres points entre ces deux lignes nommés milieu haut et milieu bas. Le
sable est prélevé à 10 cm de profondeur.
4.2- Matériel chimique : les pesticides analysés Les pesticides recherchés sont ceux pour lesquels un effet chez les reptiles a été reporté dans la
littérature : DDT et métabolites (DDD et DDE), trans-Nonachlor, dicofol, chlordane, dieldrine et
atrazine, ainsi que ceux qui sont susceptibles d'être utilisés en Guyane: diuron, mirex.
Atrazine : C8H14ClN5 ; herbicide systémique du groupe des triazines, utilisé dans les cultures de
bananes, ananas, canne à sucre. Jusqu'en 2002, la famille des triazines était le produit phytosanitaire
le plus employé en France. Elle a été introduite en 1962 et est caractérisée par une excellente
efficacité et un faible coût. En raison de sa toxicité et de sa pollution rémanente dans les eaux
(molécule très peu biodégradable), l'atrazine et la famille des triazines a été bannie en France en
2001 (mise en application en juin 2003) après des années d'utilisation (1962-2003).
Chlordane : C10H6Cl8 ; insecticide organochloré du groupe des dérivés du cyclopentadiene non-
systémique avec action de contact et respiratoire, très persistant (demi-vie en années voire en
décennies) ; en France, son utilisation en agriculture a été interdite en 1972.
DDD (dichloro-diphényl-dichloéthane) : C14H10Cl4 ; métabolite du DDT
DDE (dicholoro-diphényl-dichloréthylène) : C14H8Cl4 ; métabolite du DDT
DDT (Dichloro-diphényl-trichloroéthane) : C14H9Cl5 ; insecticide organochloré peu coûteux et
ayant un large spectre d'activité mais particulièrement toxique et persistant ; son utilisation a été
interdite dans les pays industrialisés dès les années 1970 mais il est encore largement utilisé dans
les régions tropicales dans la lutte contre le paludisme.
Dicofol : C14H9Cl5O ; acaricide non systémique avec action de contact, utilisé dans les cultures de
maïs, soja, arbres fruitiers; usage autorisé.
Dieldrine : C12H8Cl6O ; insecticide du groupe des cyclodiènes chlorés très toxique et persistant
(demi vie en décennies) ; en France, son utilisation est interdite dans les traitements agricoles
depuis 1972 mais il reste utilisé dans la lutte contre les termites et les insectes xylophages.
Diuron : C9H10Cl2N2O ; herbicide de la famille des urées ; pénètre dans le végétal par les racines et
agit au niveau foliaire ; depuis 2002, son utilisation est strictement limitée en France, interdite pour

- 20 - 20
usage non agricole, mais tolérée pour le désherbage des exploitations de bananes, de canne à sucre
et d'ananas.
Mirex : C10Cl12 ; insecticide organochloré d'ingestion, vaste usage dans de nombreuses cultures
tropicales.
Trans-Nonachlor : insecticide organochloré lié au chlordane.
D'après le dictionnaire encyclopédique des pollutions (F. Ramade, 2000) et The agrochemicals
handbook third edition
4.3- Extraction des lipides et des pesticides organochlorés lipophiles L'extraction des substances liposolubles est réalisée par un extracteur ASE 200 (Dionex)
(Accelerated Solvant Extraction). Les échantillons de sable à analyser sont placés dans les cellules
d’extraction de 33 ml. Les cellules sont introduites automatiquement une par une dans le four où
elles sont remplies de dichlorométhane. La température et la pression augmentent jusqu’à 120°C et
100 bars en 6 minutes puis se stabilisent pendant 5 minutes (phase statique de l’extraction). La
température modifie les propriétés du solvant en augmentant le pouvoir de diffusion dans les
matrices solides et entraîne un accroissement de la solubilité des analytes ; cela permet d’obtenir de
meilleurs rendements d’extraction ainsi qu’une réduction considérable de temps d’extraction et de
volume de solvant. La pression permet de maintenir le solvant sous forme liquide à températures
élevées. Le rôle de la température est également d’augmenter la vitesse de l’extraction. Les
échantillons sont récupérés dans les fioles de récupération et les solvants sont évaporés. Chaque
échantillon est repris dans 2 ml d’hexane.
4.4- Purification (Clean-up) Le but de cette étape est de séparer les micropolluants OCs lipophiles de la fraction lipidique. En
effet, les lipides ont tendance à s’adsorber sur la colonne du chromatographe, entraînant une faible
performance de l’analyse chromatographique (Hong et al. 2004). Cette séparation a été effectuée
par chromatographie d’absorption, sur phase solide à l’aide d’une colonne dont la phase stationnaire
est du florisil (Bond Elut Varian). Cette méthode est très utilisée car elle ne demande pas de gros
volume de solvant. Pour les pesticides organochlorés recherchés, deux solvants différents sont
nécessaires pour l'extraction de la totalité des substances et/ou pour l'élimination des autres
molécules organochlorés non désirées (comme les PCB). La cartouche est d'abord activée en
l'imbibant avec 3mL d’hexane. L'extrait (lipides + pesticides) en solution dans l'hexane est déposé
au sommet de la colonne, avant d’être élué par deux solvants de polarité différente, qui entraînent
avec eux les deux phases d’OCs:
a) les substances les moins polaires (DDT et métabolites) : 7 ml d’hexane,
b) les polluants les plus polaires ( les autres pesticides OCs ) : 5 ml de dichlorométhane.

- 21 - 21
Les éluats sont récupérés dans deux ballons différents. Les solvants sont évaporés et les fractions
d’OCs remis en solution dans 1ml d’hexane.
4.5- Détermination et quantification des micropolluants par chromatographie Les composés organochlorés présents dans l’extrait final ont besoin d’être séparés et détectés
individuellement pour pouvoir être identifiés et quantifiés correctement. La chromatographie en
phase gazeuse (CPG) sur colonne capillaire permet la séparation des constituants organiques.
Couplée à un détecteur à capture d’électron (ECD), cette technique se révèle être la plus adaptée à
la détection de composés organochlorés (Veningerova et al. 1997).
La CPG repose sur la coïncidence des temps de rétention d’une substance détectée avec celui d’un
étalon analysé dans les mêmes conditions ; cette technique fonctionne donc par analogie.
Les échantillons purifiés et contenus dans 1ml d'hexane sont évaporés sous azote pour les
concentrer, puis repris dans 100 µl d’hexane. Chaque échantillon à analyser est injecté à l'aide d'une
micro-seringue de 5 µl dans l'injecteur, où la température qui y règne (250°C) permet la
vaporisation immédiate des substances injectées. Les OCs sont entraînés par de l’azote ultrapur (gaz
vecteur ; débit de 20 ml/min) dans une colonne capillaire de 30 mètres de long et de 0,25mm de
diamètre interne ; la phase stationnaire (PE-5, 5% phenyl-methylpolysiloxane) tapisse la colonne
sur une épaisseur de 0,25µm. Pour chaque échantillon, une seule injection de 5 µl est réalisée et le
programme du four est le suivant : la température de départ est de 200°C pendant 40 minutes puis
augmente de 10°C/min jusqu'à 240°C où elle se stabilise pendant 15 minutes. La température du
détecteur est de 380°C.
L’analyse quantitative des différents pesticides OCs est réalisée à l’aide d'étalons ; ce sont des
solutions de référence, de concentration connue et dont l’aire des pics sortant à des temps de
rétention (RT) donnés sera comparée à l’aire des pics sortant au même RT pour les échantillons
analysés. Les références utilisées pour l’analyse chromatographique (Cil Cluzeau labo) sont les
suivantes :
Pesticide-Mix 1: aldrin, pp'DDD, pp'DDE, pp'DDT, Dieldrin, a-endosulfan, b-endosulfan,
Endosulfan sulfate, Endrin, Endrin aldehyde, a-HCH, b-HCH, g-HCH, d-HCH, Heptachlor,
Heptachlor-endo-epoxide.
Pesticide-Mix 15 : atrazine, atrazine-desethyl, atrazine desisopropyl, propazine, simazine et
terbuthylazine. (10ng/µl)
Pesticide-Mix 25 : alachlor, aldrin, atrazine, cis-chlordane (alpha), trans-chlordane (gamma),
endrin, gamma-HCH, heptachlor, heptachlor-exo-epoxide (cis-, isomer B), methoxychlor, trans-
nonachlor et simazine. ( 10 ng/µl)
Pesticide-Mix 164 : 2,4' DDD, 4,4' DDD, 2,4' DDE, 4,4' DDE, 2,4' DDT, 4,4' DDT. (10 ng/µl)

- 22 - 22
Atrazine (30 ng/µl) ; Dicofol (30 ng/µl) ; Mirex : (10ng/µl) ; MPCA (10ng/µl)
4.6- Distribution spatiale des pesticides : test de Mantel Pour construire la carte de la plage pour les concentrations en pesticides, il est nécessaire de savoir
comment les concentrations sont distribuées spatialement. Le test de Mantel est une méthode
permettant de comparer des matrices d'association (ici une matrice pour les concentrations et une
pour les distances entre échantillon). Ce type d'approche permet de répondre à la question suivante :
des localités proches dans l'espace sont-elles également proches du point de vue des concentrations
en pesticide ?
Les tests sont réalisés pour chaque pesticide et par niveau de plage (ligne végétation LV, milieu de
plage haut MH, milieu de plage bas MB, ligne de marée haute LMH).
Le logiciel Permute version 3.4 alpha 9 est utilisé pour réaliser ce test. Ce programme évalue la
probabilité des coefficients de régression et le coefficient associé en utilisant une méthode de
permutation de matrice, où la variable dépendante est la concentration et la variable indépendante
est la distance.
5. Données relatives au suivi des nids
Soixante-seize tortues et nids associés ont été étudiés lors d’une campagne datant de 2002 en même
temps que le prélèvement des échantillons de sable utilisé pour l’extraction a été réalisé. Pour
chaque femelle, la taille est mesurée et le numéro de PIT (Passive Integrated Transponder) est
relevé, ce qui permet d’identifier les individus. Il existe une base de données qui répertorie toutes
les femelles (nidifiant en Guyane) ayant un PIT et qui permet d’avoir accès à des informations sur
la fréquence de leurs visites en Guyane. Par exemple, il est possible de savoir, pour chaque femelle
de l’étude :
- si elle a déjà pondu sur Ya:lima:po avant 2002. Les femelles sont séparées en deux catégories :
celles qui ont déjà été vues sur cette plage et les nouvelles (déjà vue/première observation).
- combien de fois chaque femelle a été vue pondant sur cette plage pendant la saison de ponte de
2002.
Pour chaque nid, les distances nid / ligne de végétation et nid / ligne de haute mer sont relevées.
Enfin, la réussite d’incubation de chaque nid est quantifiée. Pour cela, chaque nid est repéré au
moment de la ponte. La position est relevée à 1 m près par triangulation par rapport à des poteaux
placés le long de la ligne de végétation tous les 10 m. Après 50 jours d'incubation, les nids sont
contrôlés tous les jours jusqu'à un signe d'émergence. Les nids sont déterrés après 48h du premier
signe d'émergence et le nombre d'œufs éclos, avortés, pourris, prédatés par des courtilières, par des

- 23 - 23
crabes, œufs sans jaune (plus petits que les autres) est compté. La réussite d’incubation est calculée
en divisant le nombre d'œufs éclos par le nombre d’œufs fertiles total. L’objectif est de relier cette
réussite d’incubation aux différents paramètres disponibles : taille de la femelle, distances à la
végétation et la ligne de haute mer, fréquence des pontes à Ya:lima:po et concentrations en
pesticides.
6. Analyse statistique La régression logistique peut être utilisée pour prédire une variable dépendante (réussite
d’incubation) sur la base de variables indépendantes (taille de la femelle, distance du nid à
différents repères,…) et de déterminer le pourcentage de variance de cette variable dépendante
expliquée par chacune des variables indépendantes. La réussite d'incubation peut être analysée en
modèle binomial ou multinomial.
Dans la régression logistique binomiale, cinq modèles (éclos, avorté, pourri, prédaté crabe, prédaté
courtilière) ont été testés. Cette analyse est réalisée sous GLMstat 5.7.4. L’analyse débute avec les
six facteurs à tester : nombre de pontes en 2002 (nbponte), distance à la ligne de végétation
(DistVeg), distance à la ligne de marée haute (DistMH), nombre d'œufs sans jaune (Ninf), taille de
la femelle (taille), première observation à Ya:lima:po (1ere obs), ainsi que les interactions de
premier ordre. Puis, on procède à une élimination des facteurs non significatifs, c’est-à-dire que le
facteur qui a la probabilité non significative la plus grande est éliminé du modèle. Le modèle final
est atteint lorsque toutes les variables sont statistiquement significatives (P<0,05). L'erreur est
binomiale, la fonction lien est la fonction logistique. La surdispersion des données est corrigée par
la déviance.
Dans la régression logistique multinomiale, la réponse du modèle est subdivisée en cinq catégories
d'œufs : éclos, avorté, pourri, prédaté par les courtilières, prédaté par les crabes. Cette analyse est
réalisée sous Jump 5.1. Dans Jump, la surdispersion des données ne peut pas être corrigée. Pour
comparer les modèles entre eux, l'AIC est alors utilisée. La surdispersion des données est corrigée
sous Excel en utilisant c qui permet de calculer le QAIC pour chaque modèle. C'est cette valeur qui
est utilisée pour déterminer quels sont les facteurs qui ont un effet sur la réussite d'incubation en
comparant les QAIC des différents modèles contraints au QAIC du modèle saturé. Le modèle qui a
le QAIC le plus faible est retenu.
AIC = -2Ln L + 2K avec K = nombre de paramètres du modèle
QAIC = AIC / c + 2K
c = -2Ln L MODELE SATURE / ddl avec ddl = ddl modèle saturé - ddl modèle complet

- 24 - 24
Résultats
1. Analyse spatiale : Position des prélèvements et des nids sur la plage Le modèle retenu pour calculer les coordonnées RGFG 95 des nids et des points de prélèvement de
sable est le suivant :
XCALCULE = a YRELEVE + b XRELEVE + c ; a = 0,10394221 ; b = 0,95845509 ; c = 174581,727
YCALCULE = a’ XRELEVE + b’ YRELEVE + c’ ; a' = 0,02975576 ; b' = 0,88005842 ; c' = 636048,59
Ces 6 paramètres ont été retenus car ils ont été obtenus avec le modèle qui présentait le plus faible
AIC (AIC= 58.54).
Les points de prélèvements de sable, répartis en seize transects et sur quatre niveaux de plage, ainsi
que la position des nids sont représentés sur la Figure 4 .
Figure 4 : Carte de la portion de plage étudiée avec les positions des prélèvements de sable et des nids

- 25 - 25
2. Analyse des pesticides du sable de la plage d’Awa:la-Ya:lima:po
2.1- Pesticides identifiés Tous les pesticides recherchés pour leurs effets chez les reptiles ainsi que ceux suspectés d'être
utilisés en Guyane ont été détectés dans le sable de la plage de Ya:lima:po. Ce sont par ordre
d'émergence sur les chromatogrammes : l'atrazine, le dicofol; le g-chlordane (molécule active), le
trans-nonaclhlor, la dieldrine et le DDT (sous sa forme op' et pp'). Certains métabolites de ces
molécules mères ont également été détectés : l’atrazine-deisopropyl, l’atrazine-desethyl, l’aldrine,
l’a-chlordane (isomère), l’op'DDE, le pp'DDE, le pp'DDD. Pour certains pesticides, les
concentrations moyennes pour la plage sont très faibles (inférieure à 15 ng/kg de sable sec) : a-
HCH, heptachlor, aldrine, op'DDE, a-endosulfan, trans-nonachlor, dieldrine, endrine, endosulfan
sulfate et mirex. La plus forte concentration moyenne est observée pour l'atrazine-deisopropopyl
avec 1715 ng/kg (2012) de sable sec.
Les étalons utilisés pour l’analyse chromatographique ont permis de détecter d’autres substances
présentes sur la plage. Ainsi ce sont 37 molécules (molécules mères et métabolites confondus) au
total qui sont présentes dans le sable. Le nombre de pic par chromatogramme était donc élevé, avec
des temps de rétention (RT) souvent proches ou confondus, comme pour le pp'DDE et la dieldrine
dont le RT est compris entre 21,578 et 21,94 minutes, rendant l’identification parfois difficile. Trois
molécules n’ont pas pu être identifiées avec les étalons utilisés. Ces molécules sont nommées X, Y,
Z pour la suite de l’analyse. Les pesticides ne sont pas détectés systématiquement dans tous les
échantillons analysés. Seul, le MCPA est présent dans les 64 prélèvements. Le g-HCH et la
molécule Z sont également fréquents (présents dans 93,8 et 90,6 % des échantillons analysés,
respectivement). Au contraire, l'heptachlor et son métabolite l'heptachlor exo-epoxide sont présents
dans moins de 10 % des échantillons. Le Tableau 1 récapitule les pesticides rencontrés et leur
valeur moyenne sur la plage, les valeurs minimum et maximum rencontrées, la fréquence de leur
présence dans les échantillons analysés et les RT utilisés pour l' identification des pics. Ils sont
classés dans l'ordre de leur apparition sur le chromatogramme. En raison de la grande variabilité des
données et du nombre réduit d'injection, les moyennes ne donnent qu'un niveau d'imprégnation et
les valeurs extrêmes un aperçu de l'étalement des données.
Parmi les pesticides identifiés dans le sable, de nombreux pesticides sont normalement interdits
d'utilisation depuis plus ou moins longtemps (aldrine, chlordane, dieldrine, endrine, heptachlor,
mirex, atrazine).

- 26 - 26
Tableau 1 : Niveaux de contamination du sable par les pesticides identifiés dans les échantillons (moyenne en ng/kg de sable sec, valeurs min et max, fréquence dans les prélèvements, ET= écart-type; RT= temps de rétention ; nd = non détectable)
pesticide moy ng/kg
ET min max fréquence (%)
RTmin RTmax
diuron 201,35 825,44 nd 4349,85 32,8 3,151 3,203
atrazine-deisopropyl 1715,13 2012,16 nd 10614,95 82,8 5,256 5,382 atrazine-desethyl 619,52 1190,61 nd 4910,56 60,9 5,383 5,519
alpha HCH <15 25,03 nd 147,85 32,8 6,145 6,287 simazine 1705,42 2346,40 nd 12451,63 67,2 6,342 6,506 atrazine 1067,52 1164,13 nd 5184,45 57,8 6,441 6,522
propazine 961,13 840,97 nd 5281,58 87,5 6,612 6,692 terbuthylazine 1687,75 1406,13 nd 6319,05 84,4 6,902 7,069
beta HCH 38,19 97,56 nd 713,26 40,6 7,111 7,145 gamma HCH 207,06 164,69 nd 780,85 93,8 7,146 7,26
delta HCH 20,20 24,66 nd 89,15 75,0 8,071 8,22 chlorothalonil 56,68 110,78 nd 417,77 45,3 8,27 8,42
alachlor 162,73 295,70 nd 1129,06 26,6 9,765 9,908 X 245,83 241,87 nd 823,75 73,4 9,97 10,07
heptachlor <15 14,23 nd 75,13 7,8 10,08 10,224 aldrine <15 19,68 nd 95,89 54,7 12,098 12,328 dicofol 299,77 1441,05 nd 10364,22 17,2 12,618 12,868
Y 140,67 282,16 nd 1577,60 78,1 14,35 14,63 Z 2228,28 2343,54 nd 7493,64 90,6 14,88 15,07
heptachlor exo epoxide 51,90 214,74 nd 1119,39 9,4 15,083 15,308 heptachlor endo epoxide 30,76 84,59 nd 475,23 34,4 15,445 15,735
g-chlordane 37,96 91,67 nd 463,42 42,2 17,299 17,561 op'DDE <15 74,40 nd 590,60 28,1 17,841 18,017
alpha endosulfan <15 20,89 nd 132,71 21,9 18,511 18,854 a-chlordane 36,25 61,58 nd 302,82 78,1 18,876 19,154
trans-nonachlor <15 19,57 nd 115,37 62,5 19,465 19,755 pp'DDE 36,49 164,38 nd 1302,75 53,1 21,578 21,78 dieldrine <15 19,13 nd 151,39 53,1 21,783 21,941 endrine <15 23,50 nd 179,80 26,6 24,551 24,955
beta endosulfan 19,08 30,04 nd 147,15 51,6 26,298 26,706 pp'DDD+op'DDT 23,13 42,04 nd 199,08 57,8 28,59 28,799 endrine aldehyde 294,39 1774,23 nd 14197,94 56,3 29,605 30,092 endosulfan sulfate <15 25,55 nd 102,22 43,8 34,511 35,079
pp'DDT 34,76 51,58 nd 317,65 87,5 35,858 36,439 MCPA 748,99 1363,93 27,34 9635,86 100,0 45,4 45,6 Mirex <15 61,16 nd 450,82 25,0 45,676 45,928
Les valeurs des concentrations étant faibles, les molécules mères et les métabolites ont été
regroupés et les pesticides qui n’atteignaient pas la valeur seuil de 500 ng/kg de sable sec ont été
écartés pour la suite de l’analyse. Le Tableau 2 donne les valeurs moyennes de concentration sur la
plage des pesticides obtenus après regroupement des pesticides.
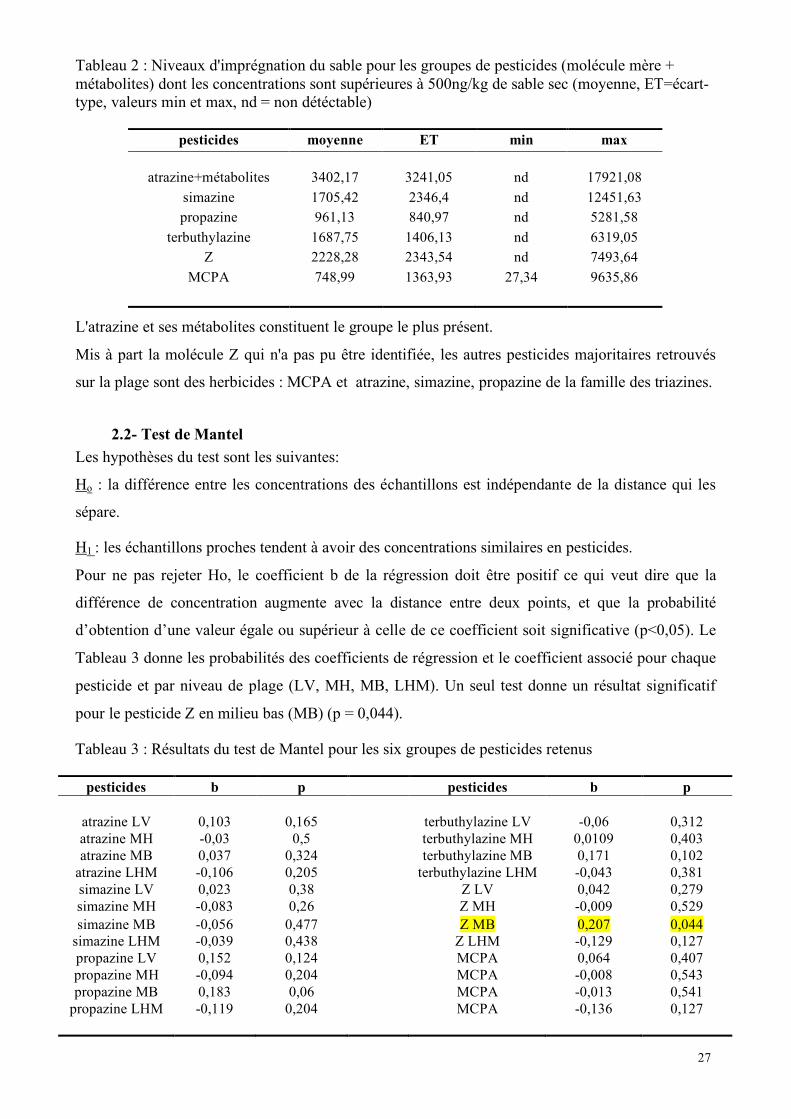
- 27 - 27
Tableau 2 : Niveaux d'imprégnation du sable pour les groupes de pesticides (molécule mère + métabolites) dont les concentrations sont supérieures à 500ng/kg de sable sec (moyenne, ET=écart-type, valeurs min et max, nd = non détéctable)
pesticides moyenne ET min max
atrazine+métabolites 3402,17 3241,05 nd 17921,08 simazine 1705,42 2346,4 nd 12451,63 propazine 961,13 840,97 nd 5281,58
terbuthylazine 1687,75 1406,13 nd 6319,05 Z 2228,28 2343,54 nd 7493,64
MCPA 748,99 1363,93 27,34 9635,86
L'atrazine et ses métabolites constituent le groupe le plus présent.
Mis à part la molécule Z qui n'a pas pu être identifiée, les autres pesticides majoritaires retrouvés
sur la plage sont des herbicides : MCPA et atrazine, simazine, propazine de la famille des triazines.
2.2- Test de Mantel Les hypothèses du test sont les suivantes:
Ho : la différence entre les concentrations des échantillons est indépendante de la distance qui les
sépare.
H1 : les échantillons proches tendent à avoir des concentrations similaires en pesticides.
Pour ne pas rejeter Ho, le coefficient b de la régression doit être positif ce qui veut dire que la
différence de concentration augmente avec la distance entre deux points, et que la probabilité
d’obtention d’une valeur égale ou supérieur à celle de ce coefficient soit significative (p<0,05). Le
Tableau 3 donne les probabilités des coefficients de régression et le coefficient associé pour chaque
pesticide et par niveau de plage (LV, MH, MB, LHM). Un seul test donne un résultat significatif
pour le pesticide Z en milieu bas (MB) (p = 0,044).
Tableau 3 : Résultats du test de Mantel pour les six groupes de pesticides retenus
pesticides b p pesticides b p
atrazine LV 0,103 0,165 terbuthylazine LV -0,06 0,312 atrazine MH -0,03 0,5 terbuthylazine MH 0,0109 0,403 atrazine MB 0,037 0,324 terbuthylazine MB 0,171 0,102
atrazine LHM -0,106 0,205 terbuthylazine LHM -0,043 0,381 simazine LV 0,023 0,38 Z LV 0,042 0,279 simazine MH -0,083 0,26 Z MH -0,009 0,529 simazine MB -0,056 0,477 Z MB 0,207 0,044
simazine LHM -0,039 0,438 Z LHM -0,129 0,127 propazine LV 0,152 0,124 MCPA 0,064 0,407 propazine MH -0,094 0,204 MCPA -0,008 0,543 propazine MB 0,183 0,06 MCPA -0,013 0,541
propazine LHM -0,119 0,204 MCPA -0,136 0,127

- 28 - 28
Cependant, le nombre important de test (4x6=24) peut être à l’origine d’un résultat significatif
(correction de Bonferonni : p = 0.002). Ce n'est donc pas suffisant pour parler de schéma de
distribution spatiale. Les concentrations sont distribuées aléatoirement sur la plage. Les
prélèvements proches ne sont pas plus semblables entre eux que les échantillons éloignés.
3. Données relatives au suivi des nids
3.1- Composition de la ponte et réussite de l'incubation des œufs à Ya:lima:po Les nids de l’étude sont composés en moyenne de 88,9 (SD 19,9) œufs fertiles et 23,9 (SD 10,4)
œufs sans jaune. Sur les 76 nids étudiés, 65 nids ont donné des nouveaux nés et 11 nids n’ont pas
abouti du tout ; le pourcentage de nids donnant au moins un nouveau né est de 85,5 %. Le taux de
réussite (correspondant à la catégorie "éclos") pour cette étude est de 37,3 % avec des valeurs
s’échelonnant de 0 à 78 %. La Figure 5 montre les moyennes des pourcentages des différentes
catégories d’œufs trouvés dans les nids.
Figure 5 : Proportion des différentes catégories d'œufs dans le nid à la fin de la période d'incubation (les barres d'erreur correspondent à l'erreur standard, le nombre d'observation n est 76) A la fin de la période d'incubation, les nids présentent en moyenne 37,3 % d'œufs éclos et 36 %
d'œufs pourris, qui représentant les catégories les plus importantes et seulement 2% d'œufs prédatés
par des crabes.
3-2 Position du nid sur la plage En moyenne, les nids se trouvent à 6,7 m (SD 4,2 ) de la ligne de marée haute. Certains nids sont
déposés en dessous de la ligne de marée haute, jusqu’à –3,8 m, c’est-à-dire qu’ils seront perdus
(œufs noyés). Les nids peuvent être pondus en revanche jusqu’à 16,4 m de la ligne de haute mer.
0
5
10
15
20
25
30
35
40
45
Œufs éclos Œuf pourri Ouef Prédaté
Courtilière
Œuf prédaté
crabe
œuf avorté
%

- 29 - 29
Par rapport à la ligne de végétation, les femelles ont creusé leur nid entre 0,1 et 15,3 m, avec une
distance moyenne de 4,8 m (SD 3,3 ). Aucune ponte observée n'a été déposé dans la végétation La Figure 6 présente les fréquences de distribution des nids par rapport à la ligne de haute mer et à
celle de la végétation. Les nids ont tendance à être déposés loin de la ligne de haute mer et près de
la ligne de végétation du haut de plage.
0,0 2,0 4,0 6,0 8,0 10,0 12,0 14,0 16,00
5
10
15
20
25
Distance à la végétation (m)
nombrede nids
-6,0 -3,0 0,0 3,0 6,0 9,0 12,0 15,0 18,00
5
10
15
20
nombrede nids
Distance à la marée haute (m)
(a) (b)
Figure 6 a et b : Fréquence de distribution des nids par rapport à la ligne de marée haute (a) et à la ligne de végétation du bord de plage (b).
3.3- Fréquentation de la plage par les femelles Durant la saison de ponte de 2002, les femelles étudiées ont pondu entre 1 et 9 fois, avec une
moyenne de 3,8 pontes (SD 2,0). Sur les 76 femelles étudiées :
- 38 ont déjà été vues sur Ya:lima:po pendant des saisons de ponte antérieures à 2002 ; certaines
sont connues depuis 1991 et ont été observées lors de six saisons de ponte différentes sur cette
plage, avec un intervalle entre deux saisons de ponte de seulement deux ans ou trois ans ;
- 38 n’existaient pas dans la base de données.
Figure 7 : Fréquence de distribution du nombre de pontes pour la saison 2002
1,00 2,00 3,00 4,00 5,00 6,00 7,00 8,00 9,000
4
8
12
16
nombre de pontes
nombrede femelles

- 30 - 30
3.4- Taille des femelles La taille des femelles étudiées est comprise entre 141 et 178 cm avec une moyenne de 161,7 cm.
3.5- Analyse des facteurs ayant un effet sur la réussite d'incubation Comme les concentrations des pesticides sont distribuées aléatoirement, il n'est pas possible d'en
déduire les concentrations au niveau des nids. La réussite d'incubation ne peut donc pas être reliée à
l'effet des pesticides. En revanche, les analyses binomiales et multinomiales peuvent quand même
être réalisée sur les autres facteurs disponibles (données relatives au suivi des nids).
3.5.1- Modèle binomial
Dans le modèle « œufs éclos », la distance du nid à la mer et la distance du nid à la végétation ne
sont pas des effets significatifs seuls mais leur interaction l'est (p=0,003) (Tableau 4). Le nombre
d'œufs éclos, donc la réussite d'incubation augmente avec l’interaction DistMH.DistVeg. Cette
interaction explique 43,1 % de la déviance du modèle.
Dans le modèle « œufs avortés », l'interaction DistVeg.DistMH est également significative
(p=0,004), et explique 31,8 % de la déviance de ce modèle.
Le modèle « œufs prédatés par les courtilières » a trois effets significatifs : le nombre de pontes de
la femelle en 2002 (p=0,0027), la distance du nid à la végétation (p=0,0242) et le statut de la
femelle (1ere observation /déjà vue) (p=0,0215). Ces facteurs expliquent respectivement 26,3, 14,4
et 15 % de la déviance du modèle. Le nombre d’œufs prédatés par les courtilières diminue avec le
nombre de pontes et la distance à la végétation et augmente quand les femelles ont déjà été
observée.
Dans le modèle « œufs prédatés par les crabes », les facteurs Distance à la végétation et nombre de
pontes ne sont pas significatifs seuls mais leur interaction l'est beaucoup (p=0,0027) et explique
43,9 % de la déviance.
Le modèle « œufs pourris » est lui influencé par l'interaction DistVeg.DistMH (p=0,0318) qui
explique 20,7 % de la déviance.
L’interaction DistVeg.DistMH a le plus d’effet sur les différents modèles testés. Le Tableau 4
récapitule les différents facteurs significatifs pour les cinq modèles testés.
Pour ces tests, la correction de Bonferroni suggère que les facteurs significatifs sont ceux pour
lesquels p < 0,0024. Dans ce cas, seule l'interaction DistMH.DistVeg dans le modèle éclos (p =
0,0003) est significative et le nombre de pontes du modèle prédation par les courtilières (p =
0,0027) et l'interaction DistMH.DistVeg du modèle prédation par les crabes (p = 0,0027) sont à la
limite de ce seuil.

- 31 - 31
Tableau 4 : GLM (Generalised Linear Model) pour le modèle œufs éclos (A), œufs avortés (B), œufs prédatés par les courtilières (C), œufs prédatés par les crabes (D) et œufs pourris (E). Le pourcentage de déviance expliquée par chaque variable est montré (%).
Estimate Deviance d.f. F ratio Prob>F % constante
A. modèle éclos Modèle 654,8 21 1,524 0,108
constante 0,8491 DistMH.DistVeg 3,58E-02 282,5 1 14,11 0,0003 43,1
B. modèle avorté
Modèle 229,9 21 8,654 0,2804 constante -1,793
DistMH.DistVeg -2,53E-02 73,01 1 8,654 0,0044 31,8
C. modèle PeCo Modèle 193,6 21 1,876 0,0328
constante -1,219 nbponte -0,144 50,89 1 9,692 0,0027 26,3 DistVeg -6,03E-02 27,85 1 5,303 0,0242 14,4 1ereObs 0,4296 28,99 1 5,521 0,0215 15,0
D. modèle PeCa
modèle 71,66 21 0,9111 0,5788 constante -4,48
DistVeg.nbponte 0,2617 31,47 1 9,694 0,0027 43,9
E. modèle pourri modèle 534,1 21 1,08 0,3959
constante -1,302 DistMH.DistVeg -2,09E-02 110,7 1 4,794 0,0318 20,7
3.5.2- Modèle multinomial
Le modèle saturé représente le modèle pour lequel la proportion de chaque catégorie d'œufs (éclos,
avortés, prédatés et pourris) est calculé à partir des observations de terrain. C'est le meilleur modèle
puisqu'il est directement basé sur les observations. Le modèle complet est celui qui prend en compte
tous les effets des différentes variables (six facteurs et les interactions simples) pour estimer ces
différentes proportions. La vraisemblance de ce modèle est de 8381,47 (tableau 5). Le manque
d'ajustement entre ces deux modèles est la différence entre les vraisemblances des modèles
(1109,575) et la déviance est deux fois cette valeur. Chaque effet est ensuite retiré et le nouveau
modèle contraint qui en résulte est comparé au modèle complet par l'intermédiaire du QAIC,
calculé à partir de l'AIC et du c . Tous les modèles contraints testés ont un QAIC inférieur au
QAIC du modèle complet, révélant que les modèles contraints sont meilleurs lorsqu'un effet est
retiré, c'est-à-dire que l'effet testé n'explique pas la réussite d'incubation. Le Tableau 5 donne la

- 32 - 32
vraisemblance, l'AIC et le QAIC de tous les modèles testés . Le QAIC du facteur DistVeg est très
proche de celui du modèle complet rappelant le résultat du modèle obtenu par la distribution
binomiale.
Tableau 5 : résultat du modèle multinomial : vraisemblance, AIC, c et QAIC pour le modèle saturé le modèle complet et les 21 modèles contraints.
Modèle -Ln L modèle ddl D residuelle AIC c QAIC
saturé 7271,894 300 1109,575 74,835 tous les effets 8381,470 84 1109,575 16930,940 224,000 392
Nb ponte 8581,298 24 1309,403 17330,595 229,341 349,341 taille 8429,265 24 1157,371 17026,530 225,277 345,277 DMH 8536,923 24 1265,029 17241,846 228,155 388,155 DVG 8666,214 24 1394,319 17500,427 231,610 391,610 Ninf 8498,846 24 1226,951 17165,691 227,137 387,137 nouv 8530,026 24 1258,132 17228,052 227,970 387,970
nbponte*taille 8394,629 4 1122,735 16957,258 224,352 384,352 nbponte*DMH 8428,483 4 1156,589 17024,966 225,256 385,256 nbponte*DVG 8414,314 4 1142,419 16996,627 224,878 384,878 nbponte*Ninf 8404,256 4 1132,362 16976,513 224,609 384,609 nbponte*nouv 8452,451 4 1180,557 17072,902 225,897 385,897 taille*DMH 8385,640 4 1113,745 16939,279 224,111 384,111 taille*DVG 8385,736 4 1113,841 16939,471 224,114 384,114 taille*Ninf 8393,420 4 1121,526 16954,840 224,319 384,319 taille*nouv 8389,510 4 1117,616 16947,020 224,215 384,215 DMH*DVG 8522,666 4 1250,772 17213,332 227,774 387,774 DMH*Ninf 8391,913 4 1120,018 16951,825 224,279 384,279 DMH*nouv 8386,297 4 1114,403 16940,595 224,129 384,129 DVG*Ninf 8408,683 4 1136,788 16985,366 224,727 384,727 DVG*nouv 8408,983 4 1137,088 16985,966 224,735 384,735 Ninf*nouv 8404,724 4 1132,829 16977,447 224,621 384,621
3.6- Distribution de la réussite d’incubation Le test de Mantel peut aussi être utilisé pour analyser la distribution des résidus du modèle « œufs
éclos » pour voir si la réussite d’incubation est liée à l’emplacement sur la plage.
b = -0,0492 et p = 0,065
On ne détecte pas de distribution spatiale du taux de réussite.

- 33 - 33
Discussion
Bien que la plage de Ya:lima:po soit incluse dans une réserve naturelle protégée, de nombreux
pesticides sont présents dans le sable de cette plage. Cette étude ne fournit qu'une information
qualitative sur ces pesticides à cause du nombre trop faible d'injections (une seule par échantillon
par manque de temps). Pour qu'une mesure soit fiable, il est nécessaire de faire au moins trois
injections. Les valeurs moyennes de concentration obtenues ne renseignent donc pas vraiment sur
l'importance de la contamination mais plutôt sur le type et le nombre de pesticides concernés,
d'autant plus que la plupart de ces pesticides sont interdits d'utilisation en France et dans de
nombreux pays industrialisés. La Guyane, bien qu'étant un département français, est dans une
situation particulière du fait de sa position géographique. Son climat tropical humide favorisant les
insectes vecteurs de maladies, certains traitements sont indispensables pour la santé de la
population. Il n'est donc pas étonnant de retrouver des pesticides comme le DDT, utilisé pour le
traitement contre le paludisme et relativement peu coûteux (Megharaj et al. 2000). Même si certains
composés sont interdits depuis longtemps (certains depuis 1972), la stabilité et la persistance de ces
composés (demi-vie de plusieurs décennies pour certains) fait que leur présence est encore
constatée aujourd'hui ; d'autre part, de nombreux pesticides, même interdits d'utilisation sont encore
massivement utilisés pour leur grande efficacité et leur faible coût de revient (facteur important
pour les pays en voie de développement ou les régions plus pauvres). Des mesures continuent d'être
prises pour limiter l'apport de ces composés toxiques dans l'environnement (Jacolot et al. 1997). Le
protocole de la convention de Stockholm sur les POPs a pris effet en octobre 2003 et interdit toute
production et utilisation de certaines substances comme l’aldrine, le chlordane, le chlordécone, la
dieldrine, l’endrine, le mirex et le toxaphène. Le protocole restreint également considérablement
l'utilisation du DDT et de l'HCH. Il n'est donc pas étonnant que le sable de la plage de Ya:lima:po
soit contaminé par de nombreux pesticides. De plus, par sa position, la plage est soumise à des
apports continuels de pesticides. En effet, la réserve naturelle de l'Amana est bordée par des
rizières. Il existe également de nombreuses cultures maraîchères (ananas, bananes). L'utilisation de
pesticides au niveau de ces terrains pourrait avoir un impact sur le milieu environnant. De
nombreux cours d'eau longent les cultures et les rizières, collectant tous les produits des traitements,
et se déversent au niveau de l’estuaire commun du Maroni et de la Mana où se situe le site de ponte
des tortues que constitue la plage de Ya:lima:po. Les éléments d'information disponibles pour la
Guyane, recueillis grâce aux rapports de situation établis par la DDASS et les DIREN, montrent un
niveau de contamination du réseau hydrique très préoccupant (Detoc 2002). Même si les mesures
contre cette pollution étaient respectées, la contamination des eaux et des sols serait encore longue
du fait de la diminution lente et progressive de la charge dans l’environnement, expliquée par la

- 34 - 34
persistance de la plupart des pesticides. Il est donc important de se préoccuper de l’effet de ces
pesticides, car leur présence risque d’être observée encore pour longtemps.
C’est pourquoi nous nous sommes intéressés à l’impact des pesticides sur la réussite d’incubation.
De nombreuses études ont déjà démontré les effets néfastes de ces composés sur la fonction
endocrine, la reproduction, le développement embryonnaire chez de nombreux vertébrés dont les
reptiles (Crisp et al. 1998). Comme la plage de Ya:lima:po présente une faible réussite d'incubation
(Girondot et al., sous presse), nous voulions savoir si la présence de pesticides pouvait être un des
facteurs expliquant cela. Le sable analysé pour l’identification des pesticides ne correspond pas au
sable à proximité des nids dont la réussite d’incubation est connue. Pour pouvoir relier les
concentrations en pesticide à la réussite d’incubation, il fallait connaître de quelle manière ces
concentrations étaient réparties pour pouvoir en déduire des concentrations aux niveaux de points
de la page non étudiés. Or, l’analyse de la distribution spatiale des pesticides identifiés n’a pas
révélé de composante spatiale. Ce résultat peut être dû à la portion de plage étudiée trop faible ; sur
300 m, les apports en pesticides par les cours d’eau ou par l’atmosphère sont les mêmes, donc il n’y
a pas de raisons de trouver un gradient. De plus, les tortues qui pondent remuent de grandes
quantités de sable (bioturbation), homogénéisant peut être ainsi la zone d’étude. Dans ce cas, les
concentrations devraient être homogènes pour un pesticide donné, et alors la variabilité de la
réussite d’incubation ne peut être expliquée par ce facteur, qui ne varie pas. Mais ce n’est pas ce qui
est observé. Les valeurs de concentration sont très disparates même pour des points très proches, et
les résultats préliminaires sur la granulométrie de la plage montrent aussi de grandes différences de
la taille des grains selon les zones (résultats en cours). Ces résultats peuvent paraître surprenants, vu
la taille du site d’étude. L’explication peut venir de l’analyse chromatographique, limitée à une
injection. La variance (ou l’ET) des concentrations est en effet très élevée ; la grande hétérogénéité
observée n’est peut être qu’en fait l’erreur entraînée par le nombre trop faible d’injections. Les
concentrations utilisées pour le test de Mantel ont été calculées à partir de cette unique injection par
échantillon et cela pourrait expliquer les valeurs énormes rencontrées à certains points de
prélèvement (exemple du dicofol dont la moyenne est de 299,77 ng/kg de sable sec avec une valeur
maximum de 10 364,22 ng/kg). La distribution des pesticides semble donc très hétérogène même
entre deux points de prélèvement très proches. Parmi les caractéristiques du sol, la teneur en matière
organique est le paramètre le mieux corrélé avec les coefficients d'adsorption, donc avec la
rétention des polluants (Barriuso et al. 1996). Une autre hypothèse pour expliquer cette
hétérogénéité pourrait donc résider dans la répartition de la matière organique dans le sable de la
plage. Si la teneur en matière organique est très hétérogène, l'hypothèse d'une distribution
homogène des pesticides n'est plus valable. Sur les 100 000 œufs pondus par km de plage et par an
à Ya:lima:po, la majorité n'aboutit pas et contribue à un apport énorme d'azote et de carbone.

- 35 - 35
L'étude de la distribution de cette MO pourrait aider dans la compréhension de la distribution des
concentrations de pesticides.
Le fait qu’il n’y ait pas de distribution spatiale nous empêche de déduire les concentrations de
chaque pesticide au niveau des nids ; l’impact des pesticides sur la réussite d’incubation ne peut
donc pas être étudié. Seuls les autres facteurs (ceux du suivi des nids) vont pouvoir être inclus dans
l’analyse. Il aurait fallu étudier les pesticides dans le sable des nids directement ; nous possédions
en effet de ce sable mais en trop faible quantité pour pouvoir être analysé correctement. Les
analyses chromatographiques ont été réalisées avec 80 g de sable et ce qui était déjà faible pour
certains pesticides à la limite de la détection. Cette étude ne pourra donc pas apporter de réponses
pour l'effet des pesticides sur la réussite d’incubation. Pour répondre à cette question, il est
nécessaire de réaliser plus d’une injection, d'étendre la portion étudiée de prélèvement pour espérer
trouver des différences selon les zone ou de prélever directement le sable au niveau de nids.
L’analyse de la réussite d’incubation ne peut donc être réalisée qu’avec les autres facteurs (distance
à la végétation, distance à la ligne de marée haute, taille de la femelle, nombre de pontes, nombre
d'œufs sans jaune, 1ere observation de la femelle). Ce type d’analyse a déjà été réalisée (Caut et
Girondot, non publié). Pour aller plus loin, les données ont été analysées d’une part en modèle
binomial en prenant en compte chaque catégorie d’œufs trouvés dans le nid (ce qui donne lieu à
cinq modèles car cinq catégories d’œufs à la fin de la période d’incubation) et d’autre part en
modèle multinomial où toutes les catégories sont traitées en même temps. Cependant, chacune de
ces méthodes présente des inconvénients limitant la possibilité d’interpréter les résultats. En analyse
binomiale, l’effet le plus significatif est l’interaction distance marée haute distance végétation dans
le modèle «œufs éclos » ; ce modèle compare les œufs éclos aux non éclos (c’est-à-dire les avortés,
les pourris, les prédatés par les courtilières et les crabes). L’effet de l’interaction DistMH.DistVeg
est très forte avant et après correction de Bonferroni. Lorsqu’un autre modèle est testé, par exemple
«œufs avortés », les oeufs avortés sont opposés aux non avortés (c’est-à-dire les pourris, les
prédatés par les courtilières et les crabes et les éclos). Or nous savons déjà que l’interaction distance
marée haute distance végétation a un effet très fort sur les éclos et cela peut donc orienter le résultat
pour ce deuxième modèle testé. En effet dans le modèle «œufs avortés », l’interaction
DistMH.DistVeg est également très significative (p=0,0044) mais moins que pour le modèle éclos
(p=0,0003) avant la correction de bonferroni mais plus après. Finalement, comme les catégories
d’œufs ne sont pas indépendantes les unes des autres car obtenues au sein d’un même nid,
l’interprétation de l’analyse binomiale est assez limitée. D’autres facteurs se sont cependant révélés
significatifs dans le modèle "prédation par les courtilières" : le nombre de pontes de la femelle et le
fait que la femelle ait été déjà observée à Ya:lima:po semblent influencer le nombre d’œufs prédatés
par les courtilères. Aucune hypothèse ne permet d’expliquer ce résultat pour le moment. Cependant
après correction de Bonferroni, seul le nombre de pontes reste significatif. Le facteur 1ère

- 36 - 36
observation des femelles est cependant limité par le protocole de suivi des tortues sur la plage de
Ya:lima:po. Les femelles qui n’existent pas dans la base de donnée ne sont pas forcément nouvelles
sur cette plage ; il se peut qu’elles soient déjà venues pondre mais qu’elles n’aient jamais été
observées ni pitées.
Le modèle multinomial ne permet pas de dégager un facteur ayant un effet sur la réussite
d’incubation. L’avantage de cette méthode était de prendre en compte tous les facteurs et toutes les
catégories d’œufs en même temps, mais l’analyse sous le logiciel Jump ne donne pas accès aux
effets des facteurs sur chaque catégorie séparément, mais seulement sur le modèle entier
comprenant les cinq catégories à la fois. Donc si les facteurs n’ont pas d’effet sur quatre des
catégories, l’effet sur la catégorie restante risque d’être masqué. C’est ce qui se passe ici ; nous
savons que l’interaction DistVeg.DistMH a un effet fort sur les œufs éclos d'après le modèle
binomial et pourtant cette interaction n’est pas significative dans le modèle multinomial.
Finalement, le résultat important qui ressort de cette étude est que la réussite d’incubation est
largement influencée par l’interaction DistVeg.DistMH. Ceci peut s’expliquer d’une part par la
présence de courtilières près de la ligne de végétation qui se nourrissent des œufs de luths (Maros et
al. 2003). Le pourcentage moyen d’œufs prédatés par les courtilières est de 13,9 % ce qui n’est pas
négligeable. La densité des courtilières augmente près de la végétation et le nombre d’œufs prédatés
par les courtilières est également plus important comme l’a montré le modèle binomial. D’autre
part, les nids déposés trop près de la ligne de haute mer peuvent être perdus par inondation ou
érosion. La réussite d’incubation est donc largement influencée par la position du nid sur la plage.
La plupart des nids de cette étude sont en effet déposés près de la ligne de végétation et très peu ont
été creusés en dessous du niveau de la marée haute (3,9 %). La position du nid sur la plage est un
compromis entre les distances par rapport à la ligne de haute mer et celle de la végétation, de façon
à limiter le risque d'érosion par les grandes marées d'une part et le risque de prédation par les
courtilières d'autre part.
Cette étude montre également que le taux de réussite observé en 2002 est du même ordre que les
taux reportés dans la littérature. Il est de 37,3 %. Ce qui peut paraître étonnant est que la réussite
d’incubation pour les tortues vertes, autre espèce de tortues marines courantes en Guyane, est
beaucoup plus élevée (environ 70 %) pour la même plage. Une des différences entre les femelles
luths et vertes résident dans l'endroit où elles décident de creuser leur nid. Alors que les vertes
pondent quasi systématiquement dans la végétation basse en haut de plage, les luths pondent entre
la ligne de haute mer et la ligne de végétation, ce qui représente une distance plus faible à parcourir.
Une hypothèse émise à ce sujet est basée sur la différence de poids et de taille entre les deux
espèces considérées. L’effort à fournir par les luths plus grosses et plus massives pour venir pondre
en haut de plage est nettement plus important. Les femelles luths déposeraient donc leur nid moins
loin en raison de l'effort à fournir pendant la montée sur la plage.

- 37 - 37
Pour compléter cette étude, l’étude de facteurs physico-chimiques comme l’humidité, la
granulométrie, la teneur des nids en oxygène ou l'étude de l'attaque par les microorganismes et les
champignons pourraient expliquer plus en détail la faible réussite d’incubation observée à
Ya:lima:po.
Conclusion L'analyse chromatographique a permis d'identifier 37 pesticides dans les échantillons de sable de la
plage de Ya:lima:po, plus gros site de ponte des tortues luths en Guyane française. La plupart de ces
pesticides sont d'une part connus pour leur toxicité, leur rémanence et leurs effets perturbateurs de
la fonction endocrine, et d'autre part pour leur interdiction d'utilisation ou au moins leur usage
restreint fortement conseillé. La grande variabilité des données de concentrations obtenues permet
cependant d'avoir seulement une idée du niveau de contamination du sable, de l'ordre de plusieurs
centaines de ng/kg de sable sec. La distribution aléatoire de ces concentrations n'a pas permis de
déduire les concentrations pour chaque pesticide aux niveaux des nids étudiés. La construction de la
carte de la plage pour les pesticides, qui était l'un des objectifs de cette étude, n'a pu être réalisée. Il
en résulte que la réussite d'incubation ne peut pas être reliée aux teneurs en pesticides du sable.
L'analyse des autres facteurs testés (taille de la femelle, nombre d'œufs sans jaune, distance du nid à
la végétation, distance du nid à la ligne de marée haute, nombre de pontes de la femelle pendant la
saison 2002, femelle observée pour la première fois ou déjà vue sur Yalimapo) montre un effet très
significatif de l'interaction distance marée haute.distance végétation sur la réussite d'incubation. Ce
résultat peut s'expliquer par la densité accrue de courtilières près de la végétation qui augmente la
pression de prédation sur les œufs, sachant que ces insectes orthoptères se nourrissent des œufs de
luths. A l'opposé, les nids déposés trop près du niveau de la marée haute sont exposés au risque
d'érosion lors des marées à grands coefficients. Le succès de la ponte réside dans un compromis
concernant sa position sur la plage. L'effet de cette interaction ne peut cependant pas expliquer
entièrement le taux de réussite d'incubation des nids. Nous sommes amenés à envisager d'autres
facteurs, qui n'ont pas encore été étudiés sur cette zone, susceptibles d'avoir aussi un effet. Ces
facteurs pourraient concerner la granulométrie de la plage, en faisant l'hypothèse que des sédiments
trop fins obstruent les pores de la coquille de l'œuf, asphyxiant l'embryon. Le sable analysé présente
en effet une grande variabilité dans la taille des grains même sur la faible portion de sable étudié.
Les microorganismes du sable pourraient également avoir un impact sur le développement des
œufs. De nombreuses bactéries et champignons ont été retrouvés dans les nids ou sur les œufs en
décomposition (Girondot et al, 1990). S'agit-il d'une cause ou d'une conséquence de la mort de
l'œuf ? Enfin d'autres facteurs physico-chimiques comme l'humidité ou la teneur en O2 du nid
pourraient fournir des éléments de réponse.

- 38 - 38
Références bibliographiques
Aguilar, A., and A. Borrell. 2005. DDT and PCB reduction in the western Mediterranean from 1987 to 2002, as shown by levels in striped dolphins (Stenella coerualba). Marine environmental research 59:391-404.
Aislabie, J. M., N. K. Richards, and H. L. Boul. 1997. Microbial degradation of DDT and its residues: a review. New Zealand Journal of Agricultural Research 40:269-282.
Barriuso, E., R. Calvet, M. Schiavon, and G. Soulas. 1996. Les pesticides et les polluants organiques des sols. Etude et Gestion des Sols:279-295.
Bell, B. A., J. R. Spotila, F. V. Paladino, and R. D. Reina. 2003. Low reproductive success of leatherback turtles, Dermochelys coriacea, is due to high embryonic mortality. Biological Conservation 115:131-138.
Bidlan, R. 2002. Aerobic degadation od dichlorodiphenyltrichloroethane (DDT) by Serratia marcescens DT-1P. Process Biochemistry 38:49-56.
Bishop, C. A., K. E. P. P.Ng., S. W. Kennedy, J. J. Stegeman, R. J. Norstrom, and R. J. Brooks. 1998. Environmental contamination and developmental abnormalities in eggs and hatchlings of the common snapping turtle (Chelydra serpentina serpentina) from the Great Lakes-St lawrence River basin (1989-91). Environmental pollution 101:143-156.
Bumpus, J. A., and S. D. Aust. 1987. Biodegradation of DDT (1,1,1-Trichloro-2,2-Bis(4-Chlorophenyl)Ethane) by the white rot fungus Phanerochaete chrysosporium. Applied and Environmental Microbiology 53:2001-2008.
Crain, D. A., L. J. G. Jr, A. A. Rooney, and D. B. Pickford. 1997. Alterations in steroidogenesis in alligators (Alligator mississippiensis) exposed naturally and experimentally to environmental contaminants. Environ Health perspect 105:528-533.
Crisp, T. M., E. D. Clegg, R. L. Cooper, and W. P. Wood. 1998. Environmental Endocrine Disruption: An Effects Assesment and Analysis. Reviews in Environmental Health 106.
Detoc, S. 2002. Les pesticides dans les eaux. Bilan annuel 2002. Institut Français de l'Environnrmrnt.
Foght, J., T. April, K. Biggar, and J. Aislabie. 2001. Bioremediation of DDT-contaminated soils: a review. Bioremediation Journal 5:225-246.
Girondot, M., and J. Fretey. 1996. Leatherback turtles, Dermochelys coriacea, nesting in French Guyane, 1978-1995. Chelonian Conservation and Biology 2:204-208.
Girondot, M., M. Godfrey, L. Ponge, and P. Rivalan. Historical records and trends of leatherbacks in French Guiana and Suriname. Sous presse.
Girondot, M., J. Fretey, I. Prouteau, J. Lescure. 1990. Hatchling success for Dermochelys coriacea in a French Guiana hatchery. In : Richardson TH, Richardson JI, Donelly M (eds) Proc 10th Annu Workshop sea turtle Biology and Conservation, 229-232, NOAA Technical Memorandum NMFS-SEFC-278, Miami
Guillette, L. J., T. S. Gross, G. R. Masson, M. J.M., H. F. Percival, and A. R. Woodward. 1994. Developmental abnormalitiesof the gonad and abnormal sex hormone concentrations in juvenile alligators from contaminated and control lakes in Florida. Environ Health perspect 102:680-688.
Guillette, L. J., and T. Iguchi. 2003. Contaminant-induced endocrine and reproductive alterations in reptiles. Pure Appl. Chemi. 75:2275-2286.
Hong, J., H. Y. Kim, D. G. Kim, J. Seo, and K. J. Kim. 2004. Rapid determination oc chlorinated pesticides in fish by freezing-lipid filtration, solid-phase extraction and gas chromatography-mass spectometry. Journal of Chromatography A 1038:27-35.
Jacolot, P., J. Charoy, and O. Gaubert. 1997. Ravageur de riz en Guyane : pour une gestion phytosanitaire plus efficace. Phytoma La Défense des végétaux 494:36-40.
Kale, S. P., N. B. K. Murthy, K. Raghu, P. D. Sherkane, and F. P. Carvalho. 1999. Studies on degradation of 14C-DDT in the marine environment. Chemosphere 39:959-968.

- 39 - 39
Keller, J. M., J. R. Kucklick, and P. D. McClellan-Green. 2004. Organochlorine contaminants in Loggerhead sea turtle blood: extraction techniques and ditribution among plasma and red blood cells. Arch. Environ. Contam. Toxicol. 46:254-264.
Kulovaara, M., P. Backlund, and N. Corin. 1995. Light-induced degradation of DDT in humic water. The Science of teh Total Environment 170:185-191.
Lefebvre, J. P., F. Dolique, and N. Gratiot. 2004. Geomorphic evolution of a coastal mudflat under oceanic influences: an example from the dynamic shoreline of French Guiana. Marine Geology 208:191-205.
Leslie, A. J., D. N. Penick, J. R. Spotila, and F. V. Paladino. 1996. Leatherback turtle, Dermochelys coriacea, nesting and nest success at Tortuguero, Costa Rica, in 1990-1991. Chelonian Conservation and Biology 2:159-168.
Llompart, M., M. Lores, M. Lourido, L. Sanchez-Prado, and R. Cela. 2003. On-fiber photodegradation after solid-phase microextraction of p,p'-DDT and two major photopruducts, p,p'-DDE and p,p'-DDD. Journal of Chromatography 985:175-183.
Lyons, G. 1999. Endocrine disrupting pesticides. Pesticides News 46:16-19. Maros, A., A. Louveaux, M. Godfrey, and M. Girondot. 2003. Scapteriscus didactylus (Orthoptera,
Gryllotalpidae), predator of leatherback turtle eggs in French Guiana. Marine Ecology Progress Series 249:289-296.
Megharaj, M., D. kantachote, I. Singleton, and R. Naidu. 2000. Effects of long-term contamination of DDT on soil microflora with special reference to soil algae and algal transformation of DDT. Environmental pollution 109:35-42.
Ockenden, W. A., K. Breivik, S. N. Meijer, E. Steinnes, A. J. Sweetman, and C. J. Jones. 2003. The global re-cycling of persistent organic pollutants is strongly retarded by soils. Environmental pollution 121:75-80.
Pieau, C., M. Girondot, G. Desvages, M. Dorizzi, N. Richard-Mercier, and P. Zaborski. Temperature variation and sex determination in reptilia.
Portelli, M. J., S. R. d. Solla, R. J. Brooks, and C. A. Bishop. 1999. Effect of dichlorodiphenyltrichloroethane on sex determination of the common snapping turtle (Chelydra serpentina serpentina). Ecotoxicology and environment safety 43:284-291.
Racke, K. D., M. W. Skidmore, D. J. Hamilton, J. B. Unsworth, J. Miyamoto, and S. Z. Cohen. 1997. Pesticide fate in tropical soils. Pure & Appl. Chem. 69:1349-1371.
Rauschenberger, R. H., J. J. Wiebe, J. E. Buckland, J. T. Smith, M. S. Sepulveda, and T. S. Gross. 2004. Achieving environmentally relevant organochlorine pesticide concentrations in eggs through maternal exposure in Alligator mississippiensis. Marine environmental research 58:851-856.
Samuel, T., and M. K. K. Pillai. 1989. the effects of temperature and solar radiations on volatilisation, mineralisation and degradation of 14C-DDT in soil. Environmental pollution 57:63-77.
Serrano, R., A. Simal-Julian, E. Pitarch, F. Hernandez, I. Varo, and J. C. Navarro. 2003. Biomagnification study on organochlorine compounds in marine aquaculture: the sea bass (Dicentrarchus labrax) as a model. Environmental Science and Technology 37:3375-3381.
Sharma, S. K., K. V. Sadasivam, and J. M. Dave. 1987. DDT degradation by bacteria from activated sludge. Environmental International 13:183-190.
Shimizu, K. I., H. Murayama, A. Nagai, and A. Shimada. 2005. Degradation of hydrophobic organic polluants by titania pillared fluorine mica as a substrate specific photocatalyst. Applied catalysis B: Environment 55:141-148.
Solla, S. R. d., C. A. Bishop, G. V. d. Kraak, and R. J. Brooks. 1998. Impact of organochlorine contamination on levels of sex hormones and external morphology of common snapping turtles (Chelydra mydas) in Ontario, Canada. Environ Health perspect 106:253-260.
Spotila, J. R., A. E. Duham, A. J. Leslie, A. C. Steyermark, P. T. Plotkin, and F. V. Paladino. 1996. Worldwide population decline of Dermochelys coriacea: Are leatherback turtles going to extinct? Chelonian Conservation and Biology 2:173-183.
Spotila, J. R., R. D. Reina, A. C. Steyermark, P. T. Plotkin, and F. V. Paladino. 2000. Leatherbacks face extinction in the Pacific. Nature 405:529-530.

- 40 - 40
VanderBrink, N. W., and E. M. DeRuiter-Dijkman. 1997. Trans-nonachlor, octachlorostyrene, mirex and ptotomirex in Antarctic seabirds. Antarctic Science 9:414-417.
VanZwieten, L., M. R. Ayres, and S. G. Morris. 2003. Influence of arsenic co-contamination on DDT breakdown and microbial activity. Environmental pollution 124.
Veningerova, M., V. Prachar, J. Kovacicova, and J. Uhnak. 1997. Analytical methods for the determination of organochlorine compounds Application to anvironnemental samples in the Slovak Republic. Journal of Chromatography 774:333-347.
Voldner, E. C., and Y. F. Li. 1995. Global usage of selected organochlorines. The Science of the Total Environment 160/161:201-210.
Vonier, P. M., J. A. McLachlan, L. J. Guillete Jr, and S. F. Arnold. 1996. Interaction of environmental chemicals with the estrogens and progesterone receptors from the oviduct of the American alligator. Environ Health perspect 104:1318-1322.
Wallace, B. P., P. R. Sotherland, J. R. Spotila, R. D. Reina, B. F. Franks, and F. V. Paladino. 2004. Biotic and abiotic factors affect the nest environment of embryonic eatherback turtles, Dermochelys coriacea. Physiological and Biochemical Zoology 77:423-432.
Wan, Y., J. Hu, J. Liu, W. An, S. Tao, and Z. Jia. 2004. Fate of DDT-related compounds in Bohai Bay and its adjacent Haihe Basin, North China. Marine pollution bulletin.
Wedemeyer, G. 1966. Dechlorination of DDT by Aerobacter aerogenes. Applied Microbiology 15. Willingham, E. 2001. Embryonic exposure to low-dose pesticides: effcts on growth rate in the
hatchling red-eared slider turtle. Journal of toxicology and environment health 64:257-272. Willingham, E., and D. Crews. 1999. Sex reversal effects of environmentally relevant xenobiotic
concentrations on the red-eared slider turtle, a species with temperature-dependant sex determination. General and comparative endocrinology 113:429-435.
Zepp, R. G., N. L. Wolfe, L. V. Azarraga, R. H. Cox, and C. W. Pape. 1977. Photochemical transformation of the DDT and methoxychlor degradation products, DDE and DDME, by sunlight. Arch. Environ. Contam. Toxicol. 6:305-314.

- 41 - 41
Annexes
Annexe 1 : Exemple du comportement du DDT en milieu naturel 1. Dégradation par des processus abiotiques
1.1- Photolyse La lumière est impliquée dans un grand nombre de dégradation dans l’atmosphère, l’eau, le sol, et
les organismes vivants (Llompart et al. 2003). L'absorption de radiations par un polluant, suivie de
réactions chimiques, est définie comme étant une photolyse directe (Kulovaara et al. 1995). La
persistance des composés hydrophobes dans l'environnement est expliquée par leur tendance à
s'adsorber sur les sédiments là où la lumière n'est pas présente (Zepp et al. 1977). D'autres études
indiquent que la photolyse directe ne cause pas de dégradation rapide perceptible des pesticides
(Llompart et al. 2003). Les mécanismes de génération des photoproduits sont des pertes successives
d’atomes de chlore pour générer des dérivés moins chlorés, des pertes d’HCl des précurseurs,
oxydation, isomérisation, formation de lien C-C entre les cycles aromatiques.
Sous les tropiques, où l’intensité lumineuse est plus forte et constante au cours de l’année, les
réactions photolytiques sont beaucoup plus importantes.
1.2- Hydrolyse La dégradation par hydrolyse des pesticides dans le sol existe ; elle est due à des réactions qui ont
lieu dans les interstices du sol ou à la surface des argiles. Le pH et la température jouent un rôle
important dans la cinétique de ces réactions ; ainsi, le taux de ces dégradations est plus important
dans les sols tropicaux, où la température est élevée et constante toute l’année (Racke et al. 1997).
1.3- Oxydation/Réduction Certains pesticides sont sensibles aux réactions d’oxydation et de réduction qui existent dans les
sols aérobie et anaérobie respectivement. Les pesticides OCs se dégradent plus rapidement en
condition anaérobie (Racke et al. 1997).
2. Dégradation par des processus biotiques La biodégradation du DDT dans les sols est un processus important, mais le taux est faible.
L’activité des bactéries, des champignons et des algues permet d’accélérer cette dégradation, si
toutefois, ces organismes ont accès au polluant (Aislabie et al. 1997).

- 42 - 42
2.1- Activité microbienne
La flore microbiologique a le potentiel de le dégrader significativement et donc de réduire les
concentrations de DDT dans le sol. Cette biodégradation résulte de mécanismes de clivage (Foght et
al. 2001). La dégradation est influencée par la concentration en DDT, le pH (favorable pour pH
neutre 6,0-8,0), la température (favorable pour des températures mésophiles 26°-30°C), par d’autres
substrats qui peuvent affecter la croissance des microorganismes et leur capacité de dégradation
ainsi que par la composition du sol et l’activité enzymatique de la microflore du sol, la disponibilité
du DDT, la présence de matière organique dans le sol comme co-substrat métabolique et l’aération
du sol (Bidlan 2002 ; Foght et al. 2001).
Les résidus de DDT persistent sous la forme de DDE et DDD (Wedemeyer 1966). De nombreuses
bactéries ont été décrites comme ayant des capacités métaboliques à dégrader le DDT, c’est à dire
capables d’utiliser le DDT comme source de carbone (Sharma et al. 1987) . Ces bactéries sont des
genres Bacillus, Micrococcus, Pseudomonas, Flavobacterium. Le plus souvent, une espèce
microbienne ne possède pas tout le système enzymatique capable de dégrader un polluant jusqu'à sa
minéralisation. Celle-ci nécessite un consortium complexe de populations microbiennes spécifiques
chacune dans des étapes spécifiques de la voie métabolique. (Barriuso et al. 1996) Parfois il y a un
seuil de concentration de DDT pour déclencher l’induction de la production d'un système
enzymatique capable de le dégrader (Aguilar and Borrell 2005 ; Bidlan 2002). L’activité
microbienne peut cependant être inhibée dans les sols contaminés depuis longtemps (VanZwieten et
al. 2003).
La minéralisation, avec la transformation du carbone organique en CO2, est le processus ultime de
dégradation : lui seul provoque la complète élimination du polluant (Barriuso et al. 1996).
2.2- Activité fongique Beaucoup de champignons transforment aussi le DDT, formant d’autres produits (Foght et al.
2001). La voie de dégradation du DDT par Phanerochaete chrysosporium est différente de celles
des bactéries ou des dégradations environnementales ; une partie est métabolisée (DDD, DBP,
dicofol…) et une autre est minéralisée en mettant en jeu le système de dégradation de la lignine
(Bumpus and Aust 1987).
2.3- Activité des organismes photosynthétiques Les cyanobactéries (fixatrices de N2) dégradent le DDT principalement en DDD. En revanche,
Clorococcum sp (algues vertes) métabolisent majoritairement le DDT en DDE (Megharaj et al.
2000).

- 43 - 43
Annexe 2 : Pesticides organochlorés étudiés : structure chimique Alachlor : C14H20ClNO2 Aldrin : C12H8Cl6 Atrazine :C8H14ClN5
Chlordane : C10H6Cl8 DDT : C14H9Cl5 Dieldrin : C12H8Cl6O
Diuron : C9H10Cl2N2O Endosulfan : C9H6Cl6O3S Endrin :C12H8Cl6O
- 44 - 44
gamma-HCH : C6H6Cl6 Heptachlor : C10H5Cl7 Lindane : C6H6Cl6
MCPA : C9H9ClO3 Mirex : C10Cl12 Propazine : C9H16ClN5
Simazine : C7H12ClN5 Terbuthylazine : C9H16ClN5

- 45 - 45
Ecotoxicologie et écologie de la réussite d’incubation chez la tortue luth , Dermochelys coriacea, en Guyane française
Résumé L'étude de la contamination du sable de la plage de Ya:lima:po en Guyane française et de l'effet de facteurs écologiques sur la réussite d'incubation de la tortue luth a eu pour objectif d'évaluer le succès des pontes de cette plage en 2002 et d'identifier les facteurs ayant un effet sur le développement des oeufs pendant la période d'incubation. L'évaluation de l'imprégnation du sable par les pesticides organochlorés a révélé la présence de 37 pesticides différents, la plupart étant connu pour leur toxicité, leur rémanence ou leur action perturbatrice du système endocrinien. En raison de la grande variabilité des résultats, les concentrations ne donnent qu'un niveau de l'imprégnation du sable, de l'ordre de plusieurs centaines de ng/kg de sable sec au niveau des points de prélèvements. En revanche, la concentration en pesticides au niveau des nids ne peut pas être connue à cause de la distribution aléatoire de ces derniers sur la plage. L'effet des pesticides sur la réussite d'incubation n'a donc pas pu être analysé. En revanche, parmi les facteurs testés en analyse GLM binomiale et multinomiale, la réussite d'incubation s'est révélée être très influencée par la distance du nid à la ligne de haute mer et à la ligne de végétation. La position du nid influence en effet la prédation des œufs par les courtilières, insectes orthoptères de la famille des Gryllotalpidae, en plus forte densité vers la ligne de végétation, et la perte des nids près de la ligne de haute mer par inondation ou érosion lors des marées à forts coefficients.
Mots clés : Dermochelys coriacea, Guyane française, réussite d'incubation, pesticides organochlorés, position du nid.
Ecotoxicology and ecology of leatherback turtle’s hatching success, Dermochelys coriacea, in French Guiana
Abstract Our study was carried out in order to study sand contamination on Ya:lima:po beach in French Guiana in 2002, and the effects of ecological factors on leatherback nest success. The aim was to assess which of those factors had a real impact on the development of eggs during the incubation period. The assessment of contamination by organochlorine pesticides made possible the identification of 37 different pesticides. Most of them are known for their toxicity, their persistence in the environment and their endocrine disrupting effect. Because high variability of data, this study gives just an idea of the level of contamination, around several hundred ng/kg of dry sand. As a result of the randomised distribution of pesticides, the nest success couldn't have been linked with the concentration, which was unknown at the nest site. Nevertheless, the binomial and multinomial analysis of ecological factors show an important effect of the nest site relative to high tide line and vegetation line. Predation by mole crickets increases due to the larger density of those insects of the family of Gryllotalpidae observed near the vegetation line. On the opposite side of the beach, nest located near the hide tide line may be lost from tidal erosion or inundation. Key-words: Dermochelys coriacea, French Guiana, nest success, organochlorine compounds, nest location


















