
Ecology is the study of interactions among organisms (biotic factors) and their physical environment...
43
Ecology is the study of interactions among organisms (biotic factors) and their physical environment (abiotic factors). Jaguar in Brazilian rain forest Solomon et. al. 1999 Population Ecology Humans on a crowded urban street Freeman 2002
-
Upload
isabel-norris -
Category
Documents
-
view
213 -
download
0
Transcript of Ecology is the study of interactions among organisms (biotic factors) and their physical environment...

Ecology is the study of interactions among organisms (biotic factors) and their physical environment (abiotic factors).
Jaguar in Brazilian rain forest
Solomon et. al. 1999
Population Ecology
Humans on a crowded urban street
Freeman 2002

Organism
Community
Ecosystem
Population
Raven & Johnson 1999
Population refers to individuals within a species living in the same place at the same time

Populations as Units of Structure and Function
•The population has unique, discernable properties and is an important unit of biological investigation.
•Interests of population ecologists include
•to understand how and why population sizes change.
•to understand populations as functional subunits of communities.
•The population is an important level of organization for the study of evolution
•a population has common gene pool
•evolutionary change happens to populations
Population ecology has important applications:
•Conservation biology - predicting extinction risk, managing populations
•Understanding, predicting human population growth
Population of Mexican poppies (Eschscholzia mexiana) blooming in the desert after the winter rains
Solomon et al 2002)

Density and Dispersion are important attributes of
populations
Species Range Geographic extent of the occurrence of individuals in a species
Population Individuals within a species living in the same place at the same time;
Species consist of populations of varying size and number, with varying degrees of proximity and connectedness (think gene flow)
Population Density Number of conspecific individuals per unit area or volume at a given time
Dispersion Distribution, or spacing of individuals within a population, relative to each other
Aerial census for African Buffalo (Syncerus caffer) in the Serengeti of East Africa. (Campbell 2002)

Clumped
Uniform dispersion Not common in general, but is not uncommon among birds nesting on small islands. This is a breeding colony of King Penguins on South Georgia Island in the South Atlantic Ocean.
Clumped dispersion Many fishes, including these butterfly fish, often clump in schools. Adaptive function may relate to hydrodynamic efficiency of swimming, decreased predation risk, increased feeding efficiency. Within a school, individuals are fairly evenly spaced
Random dispersion Populations of trees in tropical rain forests are often randomly dispersed, but in general, this dispersion pattern is rather rare in nature
Patterns of dispersion of individuals in a population

Uniform dispersion: Individuals are dispersed more evenly than expected from random occurrence in a habitat. Explanations include:
•uniform territory sizes in relatively homogenous environments (e.g. penguins)
•allelopathy -- production of toxins that inhibit growth of nearby plants (e.g. desert creosote bush and saltbush)
Campbell 1993
(Solomon et. al. 1999)
Campbell 1993

Clumped dispersion (aggregation) by far most common in nature.
•Environmental conditions seldom uniform throughout even relatively small area. Clumping of individuals often a response to clumping of resources
Clumped distribution of “Microhabitats” often explains clumping; local soil moisture, rotting log, limestone outrcrop…
•Reproductive behavior patterns including sexual attraction, often favor clumping
•Nonreproductive behavior patterns often lead to active congregation in loose groups or in more organized colonies, schools, flocks, or herds

Population demographics affect increase (growth) or decrease (decline) in population size
Demography; study of changes in size and structure of populations
Important demographic variables include:
•Sex Ratio
•Age structure (cohort=individuals of same age, age class)
•Age specific fecundity (birth) rate and mortality (death) rate
Ecologists assemble such demographic data in life tables, then use the life tables as the basis for further analysis of population dynamics

Population Dynamics; Changes in population size and demographics over time
Populations size changes in response to births and deaths, and to the movement of individuals into (immigration) our out of (emigration) the population
N1=N0 + B - D + I - E

•Mortality high during first year of life; mortality then dropped for several years, followed by a general increase
•Some of the fluctuation between years was probably related to annual rainfall; survival is related to seed production, which is closely correlated with rainfall
Life Table of the 1978 Cohort of Ground Finches on Isla Daphne
In drought years on the archipelago, seed production is low, nesting is low (most adults don’t breed), and adult survival is low
In years of heavy rainfall, seed production is high, most birds breed several times, and adult survival is high

Survivorship curves are graphical representations of age-specific survivorship
Survivorship = 1-mortality, = percent of cohort surviving to a given age
Top left figure shows generalized, hypothetical survivorship curves across a range of possibilities, and that are recurring in nature
Curve I characterizes humans, other large mammals, that have high parental care but produce relatively few young
Curve III characterizes some species with low parental care, large number of young (eg many fishes, marine invertebrates
Curve II Individuals roughly equally likely to die at any age. Characterizes some annual plants, some invertebrates (eg Hydra), some lizards, rodents, birds…

•Deer live up to 16, and females can breed at 4
•Type I survival curve indicates a relatively consistent increase in the risk of mortality with age
•The growth rate for most populations is strongly dependent on age structure
Life table for Red Deer on island of Rhum, Scotland.
http://cervid.forsci.ualberta.ca/library/taxonomy/cervus_elaphus.htm
Raven and Johnson 1999
Raven and Johnson 1999

Mathematical modeling is an important and widely-used approach to studying population dynamics
• Using equations to simulate population dynamics over time
•Illuminate complex processes and guide further research
•Vary in predictive ability; rarely are perfect approximations

Biotic Potential and the Exponential Growth Model
•All species have potential for explosive, exponential growth; absent resource limitations, growth would be exponential
•Biotic potential (rmax) is the maximum rate at which a population could increase under ideal conditions
•Exponential growth has been demonstrated experimentally in bacterial and protist cultures and in some insects.
•At some point, environmental resisitance will curtail exponential growth; environmental limitations cause decreases in birth rates and increases in death rates Time
Pop
ulat
ion
Siz
eExample of rapidly increasing population. European loosestrife is now naturalized over thousands of square miles of North American wetlands. Introduced in 1860, it has had a negative effect on may native plant and animal species
Exponential growth in bacteria. Bacteria, dividing every 20 minutes, experience exponential population growth
Raven & Johnson 1999
Solomon et al 1999

Biotic potential and the Exponential Growth Model
Time
Pop
ulat
ion
Siz
e
•Rate of increase (slope of the curve) steadily accelerates, population increases exponentially.
b=per capita birth rated=per capita death rate
Think about (bN-dN) ...
•difference between births and deaths in absolute numbers
•determines if population will grow, be stable, or decline
Keeton & Gould 1993
dN = (bN-dN) dt
dN/dt = rate of change in size of population
dt
dN

Time
Pop
ulat
ion
Siz
e
(Keeton & Gould 1993)
(b-d) is the per capita growth rate. It is the net rate of population change per individual
factor N out on right side of equation dN = (b-d)N dt
divide both sides by N to “see” the per capita growth rate (b-d)
dN dt = (b-d)----- N
back to...
dN = (b-d)N dt

Time
Pop
ulat
ion
Siz
edN = (b-d)N dt
substitute r for b-d
dN = (r)N dt
r = b-d = net rate of population change per individual at a given moment
Under these conditions of maximum birth rate and minimum death rate r is designated rmax
dN = rmaxN dt
• rmax represents the intrinsic rate of increase, or biotic potential of the population
• rmax varies widely among species
(Keeton & Gould 1993)

Time
Pop
ulat
ion
Siz
e
dN = rmaxN dt
Rate of population growth is a function of r and N; N is related to the number of breeders
N increases with each generation; therefore so does the rate of increase -- dN/ d t
Its due to this accelerating rate of increase that the slope of the curve becomes steeper and steeper
(Keeton & Gould 1993)

Population growth predicted by the exponential model. The exponential growth model predicts unlimited populations increase under conditions of unlimited resources. This graph compares growth in populations with two different values of rmax:1.0 and 0.5
•All other things being equal, a population with a higher intrinsic rate of increase will grow faster than one with a lower rate of increase
•The value of rmax for a population is influenced by life history features, such as
•age at onset of reproductive capability
•number of young produced
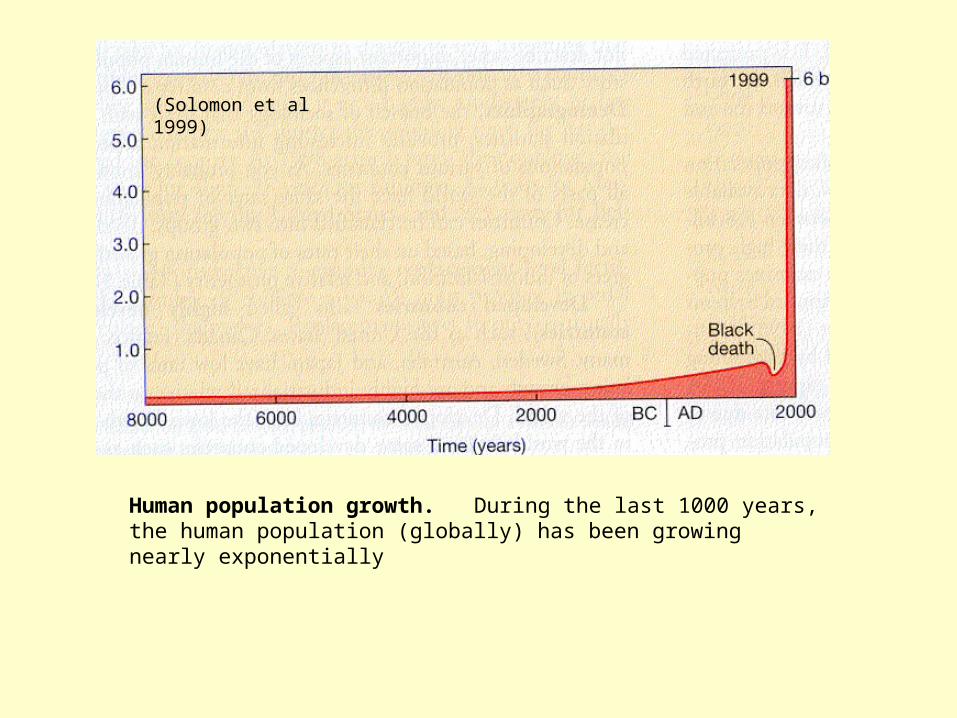
Human population growth. During the last 1000 years, the human population (globally) has been growing nearly exponentially
(Solomon et al 1999)

No population can continue to increase exponentially indefinitely
Environmental resistance:
•Environment imposes limits on population growth
•Food; water; disease; shelter from elements, predators…...
Carrying capacity (K):
•Theoretical maximum population size that can be maintained indefinitely (assumes unchanging environment)
•In reality, K changes with changes in environmental conditions
Logistic population growth:
•Populations can be modeled taking carrying capacity of environment into account using the “logistic growth equation”
•dN/dt = rN [(K-N)/K]

Num
ber
of In
divi
dual
s (N
)
The term (K-N)/K causes growth in the simulated population to respond to environmental resistance
•When N is small compared to K, [(K-N)/K] is close to 1 and growth is nearly exponential
•When N is large compared to K, [(K-N)/K] approaches 0, as does
population growth
Time

Some assumptions and simplifications of the logistic model are either not true for most populations or do not apply equally to all populations
•Each individual added to a population at a low level (N) has the same negative effect on population growth rate at low population at a high level (N)
•Each individual exerts its negative effects immediately at birth
•All individuals have equal effect on the population
•Populations approach carrying capacity smoothly – don’t overshoot it
•Carrying capacity is constant

How well does the logistic growth model fit the growth of real populations?
Experimental populations (bacteria, yeast, Paramecia…)
•Some show sigmoidal growth fairly well, but conditions do not approximate nature (predators, competitors lacking).
•Some, not all, experimental populations stabilize at some carrying capacity, and most experimental populations deviate unpredictably from a smooth sigmoidal curve
Natural populations
•Introduced populations and decimated, recovering populations show growth patterns that generally support the concept of carrying capacity that underlies logistic population growth

Logistic Population Growth
http://www.pinnipeds.fsnet.co.uk/species/species.htm
A fur seal population on St. Paul Island, Alaska The numbers of male fur seals with harems were reduced to very low numbers due to hunting until 1911. After hunting was banned, the population increased dramatically and now oscillates around an equilibrium number, presumably the islands carrying capacity for this species (Campbell 2000)
Raven & Johnson 1999
Raven & Johnson 1999

Logistic Population Growth – Overshooting K
•Lag time in many populations before negative effects of increasing population are realized
•Hypothetical example: food becomes limiting, but birthrate not immediately affected because females use energy reserves to continue producing eggs for a period; population may then overshoot carrying capacity
•Real life: In many of the populations that show sigmoidal-type growth, they oscillate around K, or at least overshoot it the first time
Num
ber
of s
heep
(in
thou
sand
s)Growth curve of the sheep population of Australia. Smooth curve is the hypothetical curve about which real curve fluctuates
(Keeton & Gould 1993)

Population Growth & Life Histories*
•Conditions of high population density may favor life history traits different from those favored at low population density
•High population size and life history
•High population size; limited resources, slow or zero population growth
•Traits favored may be those that enable organisms to survive and reproduce with few resources
•Competitive ability and high efficiency at resource use may be favored in populations that tend to remain at or near their carrying capacity
•Low population size and life history
•Low population size; abundant resources, rapid population growth
•Traits favored may be those that promote rapid reproduction; ie high fecundity, early maturity; efficiency of resource use not as important
* Life history
•Life history of an organism includes birth, growth to reproductive maturity, reproduction, and death
•“Life history traits” are characteristics that affect an organisms schedule of reproduction and death.

Life History
•Life history of an organism includes birth, growth to reproductive maturity, reproduction, and death
•“Life history traits” are characteristics that affect an organisms schedule of reproduction and death.
•Life history of any individual will include these traits;
-size & energy supply at birth-rate and pattern of growth and development-number and timing of dispersal events-number and timing of reproductive events-number, size and sex ratio of offspring-age at death
•Life History varies among organisms, lineages, based on variation in the allocation of time, effort, energy, etc, to activities and stages from birth, growth to maturity, reproduction, death
•Consider the importance of life history traits in explaining demographic populations statistics…age-specific fecundity, mortality…
Salmon spawning. Chugack National Forest, AK. Copyright J. Robert Stottlemyer/BPS.
Flowering stalks of century plants (Agave shawii). Copyright G. J. James/BPS.

•“r-Selected populations” likely to be found in variable environments in which population densities fluctuate, or in “open” habitats where individuals likely to face little competition
•“K-selected populations” likely to be living at a density near the limit imposed by their resources
•Life history traits do often vary in ways shown in table
•No demonstration of direct relationship between population growth rate and specific life history traits; concepts of r and K selection are mainly useful as hypothetical models
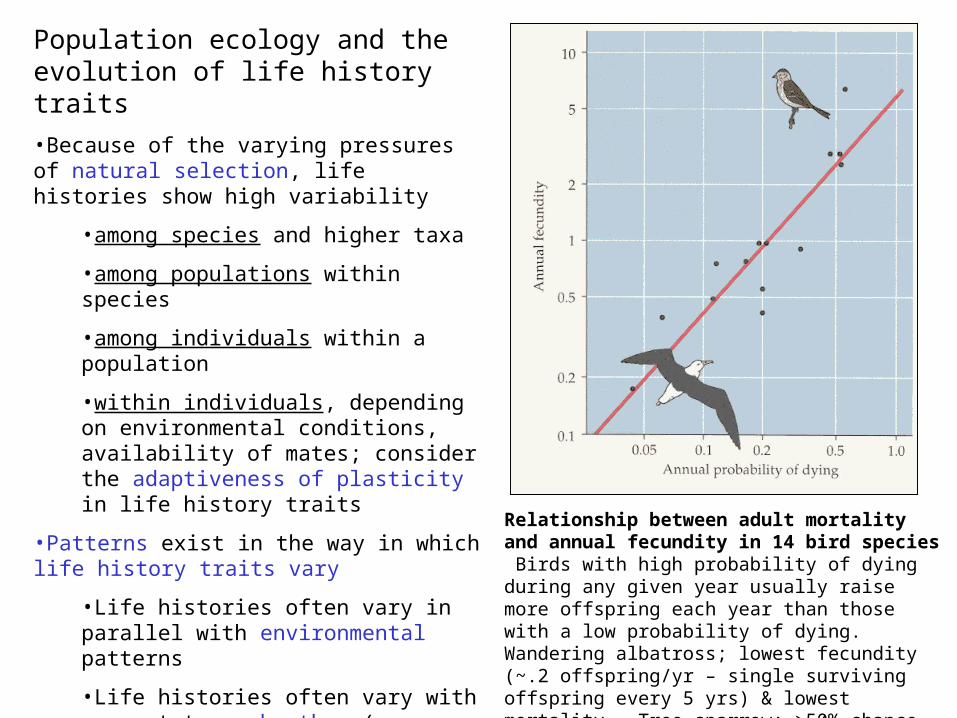
Population ecology and the evolution of life history traits
•Because of the varying pressures of natural selection, life histories show high variability
•among species and higher taxa
•among populations within species
•among individuals within a population
•within individuals, depending on environmental conditions, availability of mates; consider the adaptiveness of plasticity in life history traits
•Patterns exist in the way in which life history traits vary
•Life histories often vary in parallel with environmental patterns
•Life histories often vary with respect to each other (eg, delayed maturity & high parental investment tend to correlate with low fecundity and low mortality); such relationships between life history traits often reflect “trade-offs”…
Relationship between adult mortality and annual fecundity in 14 bird species Birds with high probability of dying during any given year usually raise more offspring each year than those with a low probability of dying. Wandering albatross; lowest fecundity (~.2 offspring/yr – single surviving offspring every 5 yrs) & lowest mortality. Tree sparrow; >50% chance of dying from one breeding season to another, produces average of 6 offspring per year

Organisms have finite resources to invest in components of their life history; Trade-offs between investments in reproduction and survival are a consequence
•Selection favors (heritable) life history traits that allow individuals to maximize lifetime reproductive success; these traits will become more common in a population
•Natural selection can not simultaneously “maximize” all the life history traits that can potentially contribute to the greatest lifetime reproductive output, because organisms have finite resources to invest; this mandates trade-offs.
•Trade-offs occur between:-number & size of young; -number of young & parental care per young; -reproduction and growth; -reproduction and survival; -current reproduction and future reproduction--between investing in current reproduction and future reproduction
Experimental manipulation demonstrates trade-off between investing in current reproduction and survival Manipulating fecundity of female seed beetles by denying access to males or egg-laying sites causes a trade off in adult longevity and fecundity

Experimental manipulation demonstrates trade-off between investing in current reproduction and survival Manipulating clutch size in collared flycatchers (add or remove eggs) results in direct trade off between number of chicks raised that year and the next year’s fecundity (no effect of current fecundity on adult survival in this study)

Populations dynamics may be influenced by factors operating independent of population density, or in a manner that is dependent on population density.
Density-independent factors Factors that affect per capita birth rate (b) or per capita death rate (d), with the degree of the effect not influenced by (dependent on), population density
•Typically abiotic, often weather-related; e.g. in insects, winters kill off all individuals except eggs and dormant larvae
•Often random (unpredictable) e.g., blizzard, flood, fire
•Effects may be indirectly related to density; e.g., social animals often able to endure weather by collective behavior -- sheep huddling in snow storm
Density-dependent factors Factors that affect the per capita birth rate (b) or per capita death rate (d), with the degree of the effect influenced by (dependent on), population density
•Important density dependent factors; Competition, predation, disease
-increasing density may attract predators (more successful, efficient, hunting prey at high density)-increasing density may increase foster spread of contagious disease-increasing density may lead to depleted food supplies
Effects are proportional to population density; Density-dependent factors exert stronger effect as population increases
Fire may constitute a density-independent factor for some populations
(Solomon et. al. 1999)

Solomon et. al. 1999
Effect of lizard presence on spider density. Tropical islands with lizards typically have few spiders. Spiller and Schoener (UC-Davis) tested the effect of presence of lizards on spider population density on Bahamian Islands
•12 Bahamian islands; all islands have native spider populations, 4 have lizards, 8 do not have lizards
•Lizards introduced in enclosures on 4 islands without native lizard populations.
•After 7 years, spider densities were higher in lizard-free islands (enclosures).
•Species diversity of spiders was also greater on lizard free islands.
•Due to predation (lizards eat spiders), or competition (lizards and spiders compete for insect food)?

Hypothetical population dynamics in regulated population. In these models, if birth or death rates or both are density dependent, population responds to increases or decreases in density by returning toward equilbrium density (zero positive or negative growth

Population “Regulation” and Reality
A regulated population is one whose dynamics are influenced primarily by density-dependent factors
Regulated populations experience interactions between density and carrying capacity
In reality, many populations are probably affected by both density-dependent and density-independent factors

Number of song sparrows on Mandarte Island (B.C.) is a consequence of density-independent (winter weather) and density dependent factors

Age interval (in months)
Number alive at beginning of age interval
Proportion alive at beginning of age interval
Number dying during age interval
Death rate for age interval
No. seeds produced per ind. during age interval
•Age-specific demographics for 9-12 month cohort•death rate = 172/316=.544•per capita birth rate = 430

Age interval (in months)
Number alive at beginning of age interval
Proportion alive at beginning of age interval
Number dying during age interval
Death rate for age interval
No. seeds produced per ind. during age interval
•Cohort members began reproducing after 3 months of age; •Then reproduced throughout life; all dead by 2 years of age•Probability of age-specific survival decreases with age•Can calculate age-specific birth rates and death rates

Time
Pop
ulat
ion
Siz
e
factor N out on right side of equation
N = (b-d) t N
(Keeton & Gould 1993)
…and dividing both sides by N gives the per capita growth rate (b-d)...

Time
Pop
ulat
ion
Siz
e
N = rN t
Under these conditions of maximum birth rate and minimum death rate:
r is designated rmax
rmax represents the intrinsic rate of increase, or biotic potential of the population
rmax varies widely among species
(Keeton & Gould 1993)
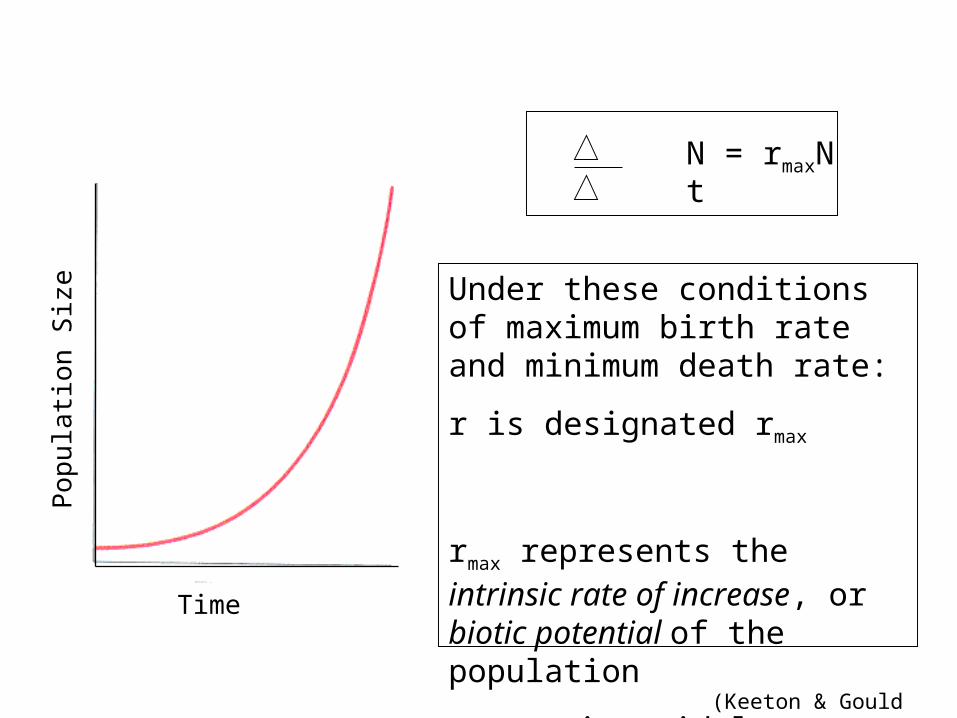
Time
Pop
ulat
ion
Siz
e
N = rmaxN t
Under these conditions of maximum birth rate and minimum death rate:
r is designated rmax
rmax represents the intrinsic rate of increase, or biotic potential of the population
rmax varies widely among species
(Keeton & Gould 1993)



















