
Managing habitat for the eastern tiger salamander and other Ambystomid salamander species

Ecological role of the salamander Ensatina eschscholtzii: direct impacts
on the arthropod assemblage and indirect influence on the carbon cycle
in mixed hardwood/conifer forest in Northwestern California
By
Michael Best
A Thesis
Presented to
The faculty of Humboldt State University
In Partial Fulfillment
Of the Requirements for the Degree
Masters of Science
In Natural Resources: Wildlife
August 10, 2012

iii
ABSTRACT
Ecological role of the salamander Ensatina eschscholtzii: direct impacts on the arthropod
assemblage and indirect influence on the carbon cycle in mixed hardwood/conifer forest
in Northwestern California
Michael Best
Terrestrial salamanders are the most abundant vertebrate predators in
northwestern California forests, fulfilling a vital role converting invertebrate to vertebrate
biomass. The most common of these salamanders in northwestern California is the
salamander Ensatina (Ensatina eschsccholtzii). I examined the top-down effects of
Ensatina on leaf litter invertebrates, and how these effects influence the relative amount
of leaf litter retained for decomposition, thereby fostering the input of carbon and
nutrients to the forest soil. The experiment ran during the wet season (November - May)
of two years (2007-2009) in the Mattole watershed of northwest California. In Year One
results revealed a top-down effect on multiple invertebrate taxa, resulting in a 13%
difference in litter weight. The retention of more leaf litter on salamander plots was
attributed to Ensatina’s selective removal of large invertebrate shedders (beetle and fly
larva) and grazers (beetles, springtails, and earwigs), which also enabled small grazers
(mites; barklice in year two) to become more numerous. Ensatina’s predation modified
the composition of the invertebrate assemblage by shifting the densities of members of a
key functional group (shredders) resulting in an overall increase in leaf litter retention.
Results from year two indicated that these effects were affected by moisture availability,
and that direct salamander impacts on invertebrates, and the related indirect effects on the
capacity for forest floor leaf litter retention were diminished in the second, wetter year.

iv
ACKNOWLEDGMENTS
First and foremost I must thank my parents for constantly nurturing the young
scientist within me; allowing their 6 year old son to tromp around the neighborhood with
a heavy 35 mm Nikon, enabling me to capture photos of insects and habitat. I am
inspired by their endless support and now look to my two children, ripe with a sea of
discoveries, directing my constant observation of the World from a new perspective. I
am so grateful for the humbling and educational guidance only parenthood could provide.
Next I have to thank Dr. H. Welsh Jr. for bringing me through this process of
development into fruition as I (and my work) transitioned from biologist to scientist. I
now feel prepared and motivated to tackle any scientific inquiry rigorously and
effectively. His expertise and graceful nature offered a singular gracious experience.
I am forever grateful to J. Baldwin for guiding my statistical analysis and writing
the many lines of code, enabling me to capture all the results at once rather than clumsily
stumbling through it on my own. The completion of this thesis also may not have been
possible without a writing grant from the Amphibian and Reptile Conservancy. I must
also acknowledge J. Gibbs for initially turning me on to reptile and amphibian
conservation and N. Karraker for sending me out to Northern coastal California in 2005,
wetting my appetite for the study of California amphibians.
Finally, digging trenches and collecting data in the rain, by my side on cold winter
days was my loving life partner Jada. I would not be who I am today without her
support. Her unconditional bond and universal wisdom are unprecedented and have
forever opened my eyes to the true power of love and commitment. The family she has
given me will continue to enrich our lives and inspires me to be the best I can be.

v
TABLE OF CONTENTS
Page
ABSTRACT........................................................................................................................iii
ACKNOWLEDGMENTS..................................................................................................iv
LIST OF TABLES..............................................................................................................vi
LIST OF FIGURES...........................................................................................................vii
INTRODUCTION..............................................................................................................1
STUDY SITE......................................................................................................................4
MATERIALS AND METHODS........................................................................................5
Experimental design...............................................................................................5
Timing……………….............................................................................................8
Invertebrate samples…...........................................................................................8
Leaf litter bags.......................................................................................................11
Statistical analysis.................................................................................................12
RESULTS..........................................................................................................................15
Ensatina effects on invertebrate taxa.....................................................................21
Leaf-litter...............................................................................................................34
DISCUSSION....................................................................................................................36
The influence of moisture and prey density……………………………...............40
Ensatina and optimal foraging theory....................................................................41
CONCLUSIONS AND RECOMMENDATIONS.............................................................44
LITERATURE CITED.......................................................................................................45
APPENDIX A....................................................................................................................50

vi
LIST OF TABLES
Table Page
1 General linear model equation terms and their explanations; used in the
Analysis comparing invertebrate samples across plots in each year.………… 14
2 Analysis of the effects of salamander predation, moisture, month, and the
interaction of month*moisture, on invertebrate functional groups by size
class in two years using a general linear model. Data were analyzed
separately by group, size, and year. Results indicated with – were not
statistically significant at α = 0.1……………………………………………… 20
3 Analysis of the effects of salamander presence (Control_Treatment), and
their interactions with moisture and time interval on invertebrate taxa in two
years using a general linear model. Data were analyzed separately by taxon,
size class, and year. Results indicated with - were not statistically significant
at α = 0.1.……………………………………………………………………… 22

vii
LIST OF FIGURES
Figure Page
1 Experimental enclosures to assess the impacts of Ensatina on invertebrate and
litter turnover in situ at the field location near Ettersburg, California. Enclosure
dimensions were 3 m x 3 m x 23 cm, plot dimensions = 1.5 m x 1.5 m x 23 cm. 6
2 Density of invertebrates by functional group, with the two most abundant taxa
within the decomposers, mites and springtails represented separately.
Invertebrates were extracted from litter samples collected within experimental
plots near Ettersburg, California in 2007-2008. Values above bars are relative
composition out of all invertebrates found represented as percentage of 100…. 16
3 Density of invertebrates by functional group, with the two most abundant taxa
within the decomposers, mites and springtails represented separately.
Invertebrates were extracted from litter samples collected within experimental
plots near Ettersburg, California in 2008-2009. Values above bars are relative
composition out of all invertebrates found represented as percentage of 100…. 17
4 Mean density of: invertebrates and invertebrate decomposers <1mm on control
and treatment plots sampled at 5 monthly intervals in 2007-2008 from Mattole
field sites near Ettersburg, California. The blue line represents the percent litter
moisture in 2007-2008. Error bars are ± one standard error…………………... 18
5 Mean density of: invertebrates and invertebrate decomposers <1mm on control
and treatment plots sampled at 5 monthly intervals in 2008-2009 from Mattole
field sites near Ettersburg, California. The blue line represents the percent litter
moisture in 2008-2009. Error bars are ± one standard error….......................... 19
6 Mean density of: Entomobryidae springtails <2mm, beetles <2mm, larvae
>2mm, and larvae <2mm on control and treatment plots sampled at 5 monthly
intervals in 2007-2008 from Mattole field sites near Ettersburg, California.
Error bars are ± one standard error………..……………………………………. 23
7 Mean density of earwigs on control and treatment plots sampled at 5 monthly
intervals in 2007-2008 from Mattole field sites near Ettersburg, California.
Error bars are ± one standard error……………………………………………... 24
8 Mean density of Orabatidae mites <1mm on control and treatment plots
sampled at 5 monthly intervals in 2007-2008 from Mattole field sites near
Ettersburg, California. Error bars are ± one standard error…………………… 25

viii
LIST OF FIGURES (CONTINUED)
Figure Page
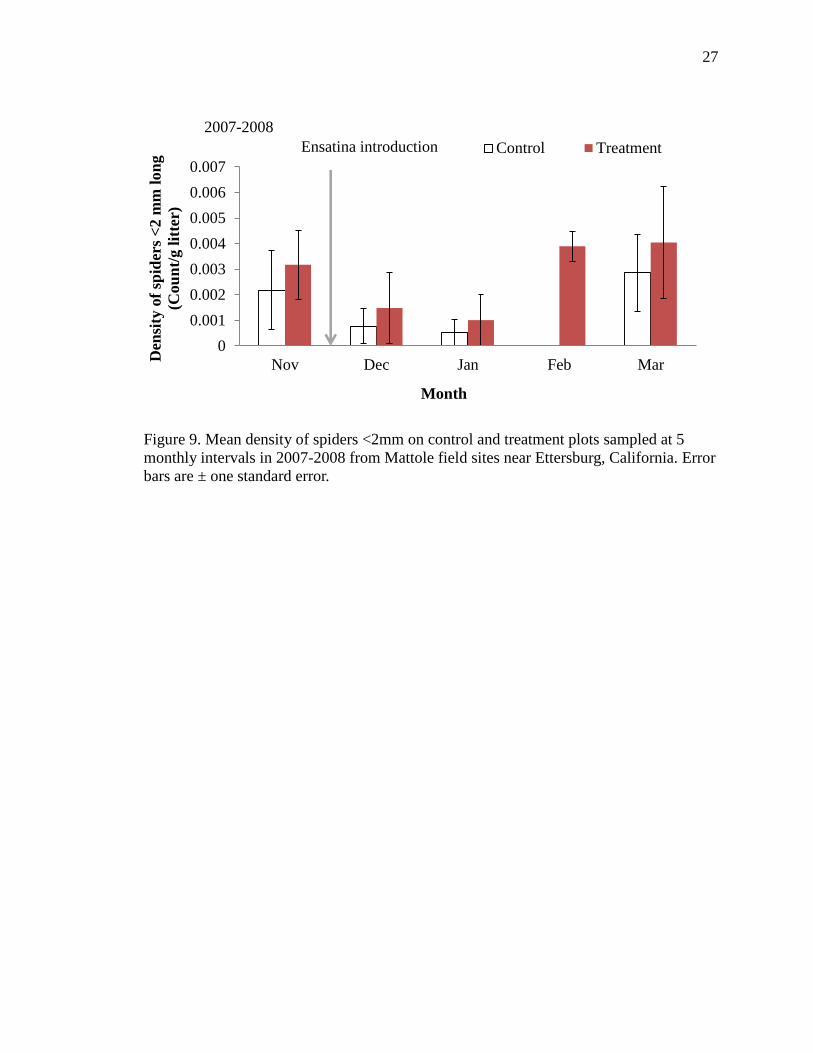
9 Mean density of spiders <2mm on control and treatment plots sampled at 5
monthly intervals in 2007-2008 from Mattole field sites near Ettersburg,
California. Error bars are ± one standard error………………………………… 27
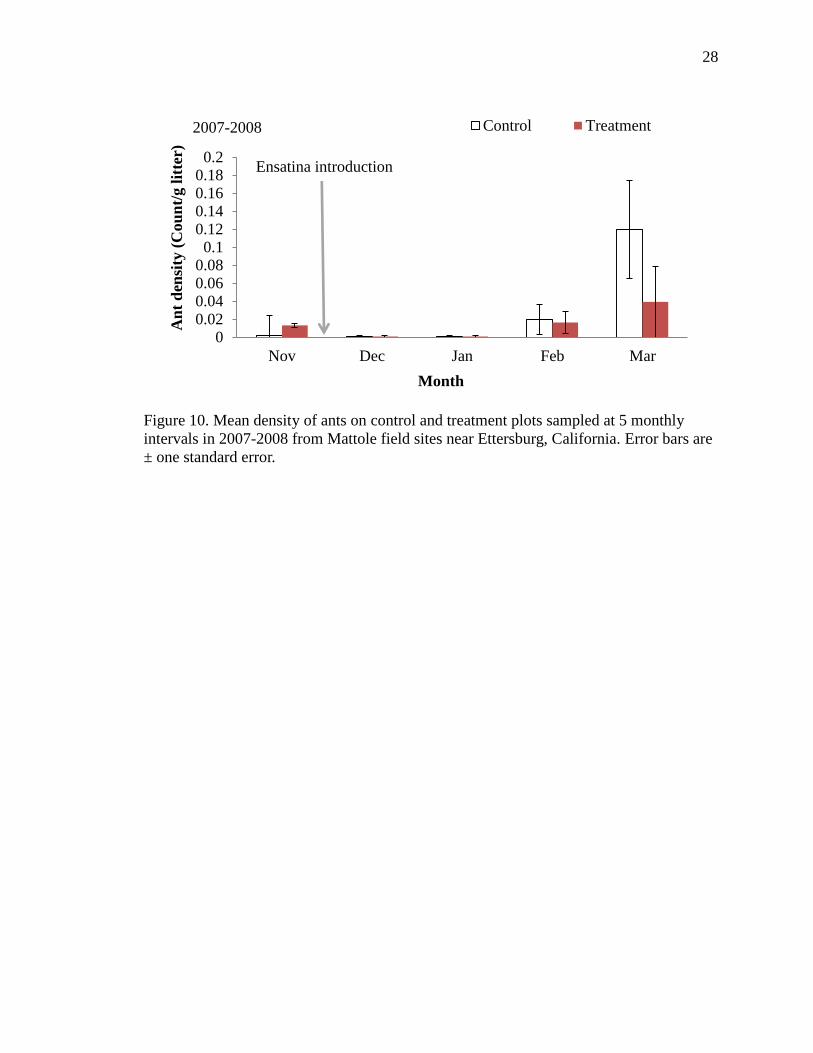
10 Mean density of ants on control and treatment plots sampled at 5 monthly
intervals in 2007-2008 from Mattole field sites near Ettersburg, California.
Error bars are ± one standard error…………………………………………….. 28
11 Mean density of millipedes on control and treatment plots sampled at 5
monthly intervals in 2007-2008 from Mattole field sites near Ettersburg,
California. Error bars are ± one standard error……………………………….. 29
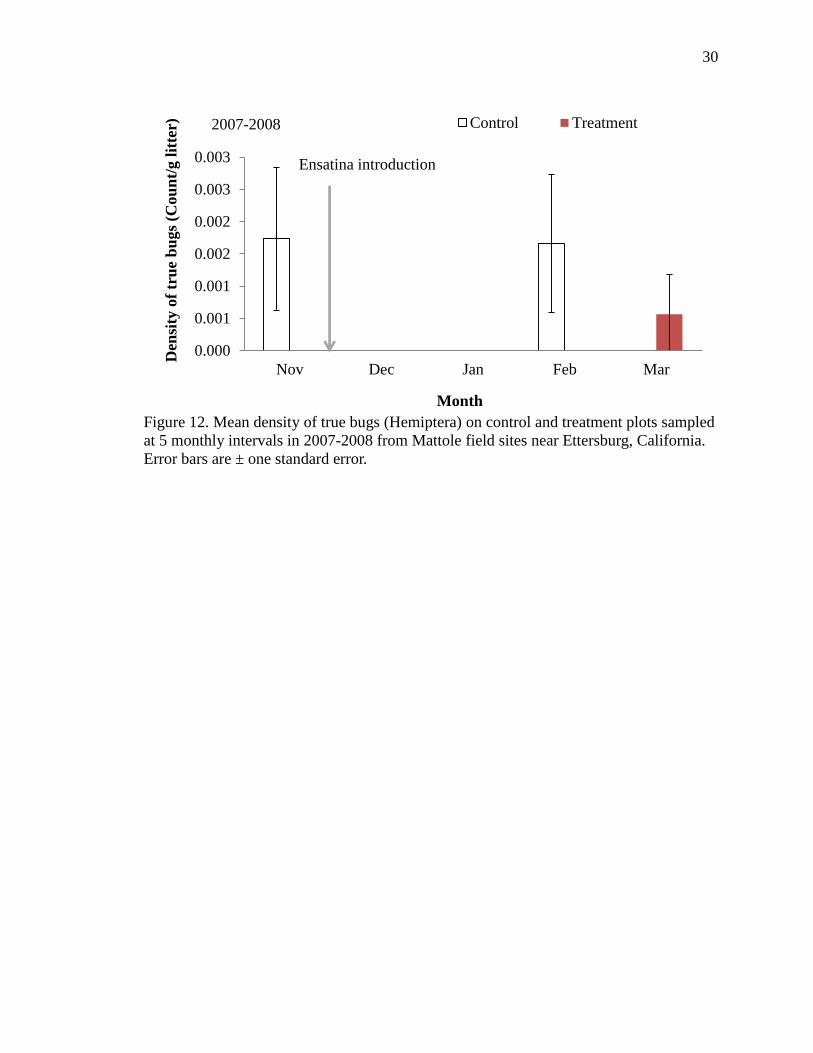
12 Mean density of true bugs (Hemiptera) on control and treatment plots sampled
at 5 monthly intervals in 2007-2008 from Mattole field sites near Ettersburg,
California. Error bars are ± one standard error……………………………….. 30
13 Mean density of: barklice (Psocoptera) and worms (Annelida) on control and
treatment plots sampled at 5 monthly intervals in 2008-2009 from Mattole
field sites near Ettersburg, California. Error bars are ± one standard error…... 31
14 Mean density of Pseudoscorpions on control and treatment plots sampled at 5
monthly intervals in 2008-2009 from Mattole field sites near Ettersburg,
California. Error bars are ± one standard error.…………………….…………. 32
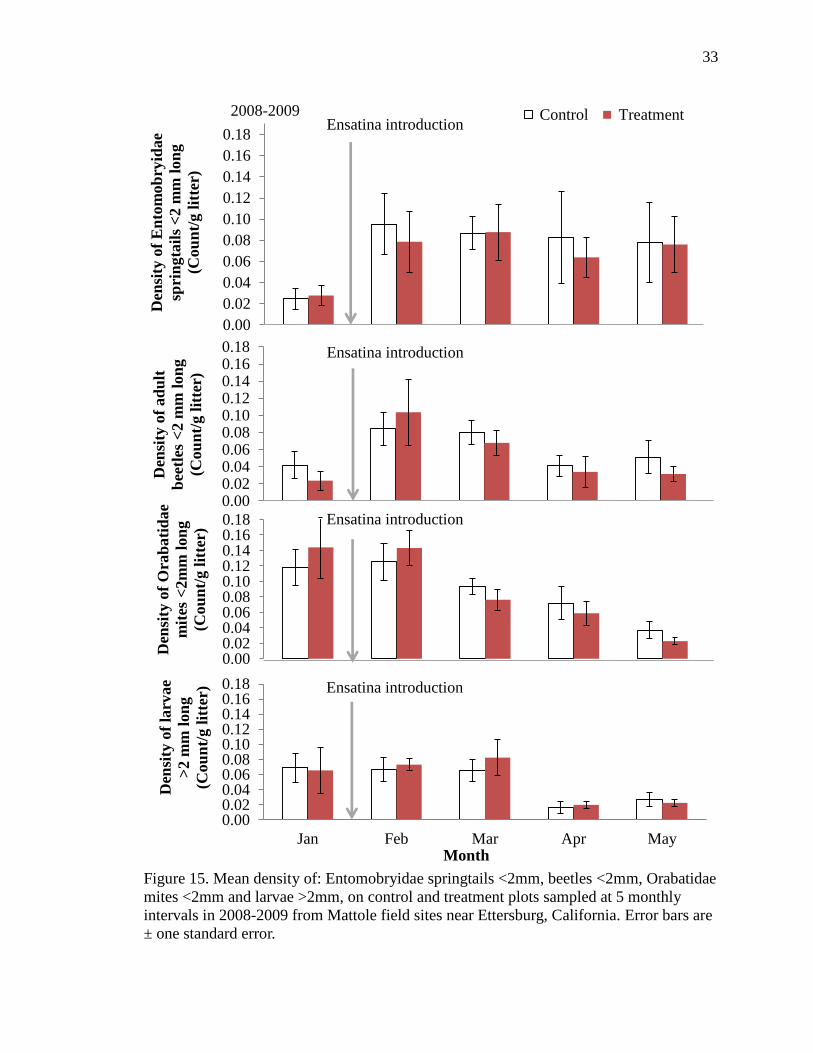
15 Mean density of: Entomobryidae springtails <2mm, beetles <2mm,
Orabatidae mites <2mm and larvae >2mm on control and treatment plots
sampled at 5 monthly intervals in 2008-2009 from Mattole field sites near
Ettersburg, California. Error bars are ± one standard error................................ 33
16 Mean leaf litter weight (g) on treatment and control plots in: Year One (2007-
2008); and Year Two (2008-2009), at the Mattole field sites near Ettersburg,
California. Error bars are ± one standard deviation.…………………………. 35

1
INTRODUCTION
Woodland (plethodontid) salamanders are the most abundant vertebrate predators
in northwestern California forests (Welsh and Lind 1991), where based on their numbers
and biomass they are ecologically dominant, filling a key nutrient cycling role:
converting invertebrate to vertebrate biomass (Burton and Likens 1975a). Burton and
Likens (1975b) found one species of plethodontid salamander (Plethodon cinereus) in an
eastern U.S. hardwood forest comprised greater biomass than all the songbirds and equal
to that of all the small mammals combined. At a North Carolina forest site Petranka and
Murray (2001) reported the entire plethodontid assemblage comprised six times the
number of individuals and 14 times the biomass reported by Burton and Likens (1975b).
Welsh and Lind (1992) reported an extremely high density for a single species of
plethodontid salamander at a mixed conifer/hardwood forest site in northern California.
In a New York forest, Wyman (1998) found that the consumption of invertebrates by the
most abundant plethodontid salamander Plethodon cinereus increased forest floor carbon
retention by reducing leaf-litter breakdown 11-17%. Walton (2005) found that this
process was not straight-forward, and might be affected by variability in both leaf litter
mass and moisture content. Results from other studies have provided evidence that
invertebrate densities might have increased in the presence of terrestrial salamanders
(Rooney 2000, Walton et al. 2006) or have had negligible effects on invertebrates
(Homyack et al. 2010), however, having no significant effect on leaf litter breakdown.
Complex food webs can be regulated both from below, where abiotic factors (e.g.,
nutrients, moisture, etc.) control the potential for productivity (bottom-up effects), and

2
simultaneously from above by predation, disease, parasitism, etc. (top-down effects) (e.g.,
Benrong and Wise 1999, Gruner 2004, Bridgeland et al. 2010). The bottom-up effects
can both promote or restrict the higher trophic levels from below, while predation or its
absence can similarly influence the system from above (McCay and Storm 1997,
Benrong and Wise 1999, Kagata and Ohgushi 2006).
Wyman (1998) found all invertebrate groups he sampled were reduced by
salamanders in field enclosures, while other studies using both lab and field enclosures
found Podomorphic springtails (Collembola) increased while other invertebrate taxa
decreased, in the presence of salamanders (Rooney 2000, Walton and Steckler 2005,
Walton et al. 2006). Walton and Steckler (2005) attributed the increase in Podomorphic
springtails to selection by salamanders for larger prey, releasing smaller arthropods from
competition and depredation by invertebrate predators, along with the effect of enhancing
their microfloral food base via salamander feces deposition. In contrast, avoiding the use
of enclosures, Walton (2005) found no significant top-down effects from salamanders
over the first year and instead found moisture and litter mass influenced invertebrate
densities. However, in the following year, salamander presence was the single significant
factor that influenced invertebrate densities in the spring; the combination of salamanders
and litter mass significantly influenced invertebrate densities in the fall (Walton 2005).
The Ensatina salamander (Ensatina eschscholtzii; hereafter Ensatina), a member
of the family Plethodontidae, is the most abundant terrestrial salamander in the mixed
hardwood/ Douglas-fir forests of Northwestern California (Welsh and Lind 1991).
Terrestrial salamanders feed on most arthropods including springtails, mites, and beetles
(Bury and Martin 1973, Lynch 1985, Rooney 2000). Many small arthropods are

3
important decomposers of forest leaf litter, an assemblage dominated by mites and
springtails (Gist and Crossley 1975, Singh 1977). An analysis of stomach content of
Ensatina and another common plethodontid species, the California slender salamander
(Batrachoseps attenuatus), in redwood forest (Sequoia sempervirens) found springtails
were the most common prey consumed, followed by mites, which were equal in number
to springtails in the slender salamander (Bury and Martin 1973). The importance of
small invertebrate decomposers in the diets of these abundant salamanders indicates the
ecologically dominant influence they can have on the capacity of the forest litter layer to
sequester or release carbon and cycle important nutrients at the litter-soil interface (Gist
and Crossley 1975, Singh 1977).
Studies of the roles of terrestrial salamanders in forest detrital food web dynamics
have not been conducted in the Western United States. The objective of my study was to
determine how Ensatina predation impacted the densities of members of the forest
invertebrate assemblage, and how this may have affected the breakdown of leaf-litter;
which indirectly influenced the relative amount of carbon and nutrients retained at the
litter/soil interface in mixed hardwood-conifer forests in Northwestern California.
I evaluated the following hypotheses in a mixed hardwood/conifer forest of
northern California: (1) Ensatina had a top down influence via predation on the
composition and densities of invertebrates dwelling in the forest floor litter; (2) leaf litter
breakdown in this forest floor food web was reduced via this predation on plots with
Ensatina compared to controls; and (3) available moisture affected these dynamics by
influencing the relative abundances of the invertebrates and the rate of litter breakdown
in this leaf litter food web.

4
STUDY SITE
I conducted a field experiment in the Mattole river watershed of northwestern
California near the village of Ettersberg (40° 6'3.21"N, 123°58'42.31"W). The study site
was 400 meters above sea level on a forested ridge that divides two Mattole River
tributaries. This forest is dominated by tanoak (Lithocarpus densiflorus) and Douglas-fir
(Pseudotsuga menziesii), and also contains madrone (Arbutus menziesii), black oak
(Quercus velutina), canyon live-oak (Quercus chrysolepis), and Bay Laurel (Laurus
nobilis). The understory consists mostly of huckleberry (Vaccinium ovatum), but this was
uncommon in the direct vicinity of the field site. This mixed forest type results in leaf
deposition throughout the year; with litter at the study site dominated by madrone leaves
in the summer and tanoak leaves in the fall. The area receives little precipitation during
spring and summer, with rainfall during the fall and winter months high, averaging over
250 cm (100 inches) and exceeding 500 cm (200 inches) during wet years (Welsh et al.
2005). Summer temperatures often exceed 32OC (90
OF); winters are cool with occasional
freezing nights and snowfall along ridges (Welsh 2007, personal communication).

5
MATERIALS AND METHODS
To test the effects of Ensatina predation on invertebrates I conducted a controlled
salamander housing experiment over four winter months during 2007-2008 and 2008-
2009. In order to incorporate the bottom up effects of the living forest floor, salamander
barriers were installed directly into the ground (15 cm deep) where they remained,
uncovered and exposed. These barriers created 12 similar plots utilized as either controls
or treatments (six of each). I collected leaf-litter cores to quantify the invertebrates in
each plot before conducting the experiment and then at the end of each 30 day period
following the introduction of salamander treatment individuals. Leaf litter bags of a
similar composition and pre-determined dry weight were placed within each plot at the
initiation of salamander introductions and were removed 120 days later to assess the
breakdown of leaf litter in both control and treatment plots.
Experimental design
I used three experimental enclosures, each divided into four plots (1.5 m2);
creating a total of six treatment and six control plots (Figure 1). The enclosures were
constructed in situ on the forest floor. The walls were buried 15 cm deep into the litter
layer down to the mineral soil to ensure that salamanders could not escape beneath the
walls. The three enclosures were within a 12 m2 area and all three were enclosed by a
chicken wire fence 2.5 meters in height to prevent destruction or predation of Ensatina by
wildlife. Enclosures were constructed using 30 cm high metal sheets folded and attached

6
Figure 1. Experimental enclosures to assess the impacts of Ensatina on invertebrate and
litter turnover in situ at the field location near Ettersburg, California. Enclosure
dimensions were 3 m x 3 m x 23 cm, plot dimensions = 1.5 m x 1.5 m x 23 cm.

7
at the corners with steel bolts to form the perimeter and transected with a pair of metal
sheets through the center as a perpendicular bisector of each exterior wall to create four
equal-sized plots in each enclosure (Figure 1). The use of experimental field enclosures
to study food web dynamics has been criticized because it may confound predator-prey
interactions, including predator avoidance, and influx of prey from neighboring sites
(Walton 2005, Walton et al. 2006). To address these issues, the four interior walls were
equipped with two hardware cloth windows (1.5 cm mesh) measuring 10 cm in height
and 35 cm in length fastened to the sheet metal with screws to allow for arthropod
migration between the plots while preventing salamanders from moving between them.
Enclosures were equipped with a 10 cm aluminum lip around all edges fastened with
binder clips silicone glued to the underside, using duct-tape to seal the adjoining surfaces;
preventing salamanders from climbing out. The enclosures remained open at the top to
allow rainfall and leaf-litter deposition to occur naturally.
Each four-unit enclosure was systematically assigned two treatment and two
control plots with a random start, but so that no two treatment plots were adjacent. One
male Ensatina was placed in each treatment plot. Ensatina weighed at least 3.5 grams but
not more than 4.5 grams. Each plot was provided with three rough-cut Douglas-fir
boards measuring: 45 cm by 15 cm and between five and eight cm high for surface cover.
Salamanders found to already exist within enclosures were removed at the beginning of
the experiment. One treatment plot was omitted from the analysis on December 7, 2007
due to escape of the Ensatina salamander. Methodologies were approved by the

8
International Animal Care and Use Committee at Humboldt State University; protocol
number was 06/07.W.150.A.
Timing
The salamander predation experiment was initiated on November 1, 2007 with the
onset of seasonal rainfall and when Ensatina salamanders were present in the surface
litter of the adjacent forest. The salamander treatment plots were populated over the
following 3 weeks and were removed on March 22, 2008, 120 days after the last plot was
populated. In the winter of 2008 rainfall was delayed and salamander introductions
occurred from December 30, 2008 through January 11, 2009. Salamanders were
removed from the enclosures on May 15, 2009, 120 days after salamander introductions.
This timing was also consistent with the appearance of local salamanders on the surface
of the forest floor, which in both years allowed me to find individuals from the
surrounding forest to populate the experimental plots (Welsh unpublished data). HOBO
Pendant data loggers enclosed in water-tight PVC canisters were randomly assigned to
one plot within each enclosure to measure air temperature during the experiment.
Invertebrate samples
Five leaf litter cores were extracted from each plot in each time interval (month)
to provide invertebrate samples. The litter cores (sub-samples) were collected using a
soup can (486 cm3) with both ends removed, pushed firmly down through the leaf-litter
until contact with the mineral soil beneath. Sub-sample locations were determined using
a random number generator and a 100-point grid, but stratified to encompass three sub-
samples along the plot edge and two sub-samples within the interior. It was presumed

9
that the plot edge might accumulate invertebrates by promoting travel along this barrier
and as such it would be important to sample this area. Sub-sample locations were not
reused. Holes generated from removing litter cores were first measured to determine
litter depth then gently collapsed to prevent soil drying and to minimize disturbance to
soil strata. The five sub-samples from each plot were combined to create the invertebrate
sample for each plot, each month. The litter removal associated with this sampling
method totaled approximately 5% of the surface area of each plot in each sampling event;
resulting in approximately 25% of the plot surface area sampled over the experiment.
The material removed from a sub-sample core was immediately placed into a
labeled quart-sized Ziploc© bag and then into a cooler to prevent the samples from
drying out or mobile invertebrates from escaping. The sub-samples were chilled but
above freezing (1-5 O
C) until they could be processed in Berlese-funnel extractions to
collect the arthropods they contained, within 48 hours. The Berlese funnels were setup
within a wooden frame approximately one square meter with 30 seven centimeter
diameter holes bored into wooden slats fitted atop the frame; each hole with a plastic
funnel attached below. The structure was designed to fit a sub-sample can within each
position such that the can would rest securely within the funnel. A hardware cloth barrier
(1.5 cm mesh) was placed at the bottom of each can to prevent soil from clogging the
funnel. A string of seven watt lights hung above the frame with one light placed within
the top of each sampling can. Each can was wrapped shut with aluminum foil to promote
the drying of samples and the downward migration of invertebrates. One dram vials
filled with 95% ethanol were placed underneath each funnel to collect invertebrates as
they fell from the litter sample. Berlese-funnel extractions received continuous heat and

10
light from the seven watt bulbs for four days (96 hours) after which the samples were
completely dry (adapted for small, wet samples; see Wyman 1998). The leaf litter sub-
samples were weighed before and after the drying extraction process to determine percent
moisture; the percent moisture calculations from the sub-samples were averaged for each
plot and time. For the statistical analysis invertebrate abundances in each sub-sample
were divided by the dry weight of the litter in their respective sub-sample to correct for
variable litter depths and to generate invertebrate densities (count/gram of dry leaf litter).
Ensatina has the widest gape of the plethondontid salamanders in the western
U.S., allowing it to consume a variety of prey, with moderate sized adult salamanders
(35-49 mm, snout-vent-length) consuming mostly small (<0.3 mm3) and medium (<19
mm3) sized invertebrate prey; 45% of their diet by volume was small and 55% medium,
with less than 0.2% of prey greater than 19 mm3 (Lynch 1985). In order to evaluate
differences between control and treatment plots based on both prey type and prey size, I
examined invertebrates under a dissecting microscope, identifying them to family, and
assigning them to one of three size classes: less than one millimeter, between one and two
millimeters, greater than two millimeters (small, medium, and large, respectively). These
three length categories captured the variability of prey size consumed by Ensatina defined
in Lynch (1985). Invertebrates were organized into: decomposers (shredders/grazers),
predators, herbivores, omnivores (ants) (see Peterson 1982). Fly and beetle larvae
identified as shredders were combined into a single group (larvae), due to low sample
sizes. Invertebrate larvae identified as predators based on morphology were included in:
predators; these were comprised of immature stages from the orders Coleoptera, and
Neuroptera. The identification of mites was simplified into two groups: Orabatidae (fully

11
sclerotized) and non-Orabatidae (not or partially sclerotized). Appendix A contains a
complete list of invertebrates detected.
Leaf litter bags
Each experimental plot was provided with three mesh bags made of metal
window screen (3.0 mm mesh) open at one end and containing 3.0 g of leaf litter (equal
amounts of Madrone and Tanoak) which had not begun decomposition and was fully
intact. Douglas fir needles were excluded from these procedures as their small size
enabled them to slip through the window screen easily thus generating a large source of
error. The leaves were collected from an area approximately 50 m2 centered on the
location of the experiment. Once a sufficient quantity of leaves was collected they were
dried in an oven at 93OC (200
OF) for two hours (until completely dry and brittle) and
weighed out to 3.0 g increments containing leaves from both dominant species. The 36
litter bags were filled and deployed at one time so that drying and weighing conditions
would be consistent for all bags. Leaf-litter bags were collected after the experiment and
re-dried at 93OC for two hours and carefully re-weighed immediately upon drying to
ensure the accuracy of dry-weight measurements. The change in weight from the initial
3.0 g to final dry weight was averaged across the three litter bags in each plot and used to
compare the change in mean leaf weight between control and treatment plots over the
course of the experiment. Hardwood leaves are composed of approximately 50% carbon
by weight (carbon mass=0.475 * mass of oven dried leaf; Schlesinger 1991) so we
assumed that half the change in leaf weight quantified here would serve as a surrogate for
the amount of carbon either retained or lost over the four months of the experiment.

12
Statistical analysis
The samples of invertebrates yielded counts of their densities from each of the 12
plots: initially (prior to salamander introductions), and after each 30 day period following
their introductions (4 months), replicated in both years. Because invertebrate sampling
occurred within the same plots over time the samples were considered repeated measures
within each year. Walton (2005) demonstrated the significant influence of moisture on
invertebrate densities and differences in the effects of top-down regulation by a
salamander predator, therefore the distinct differences in rainfall regime between the two
years at the site warranted the analysis of each year separately. The depth of each litter
core and the amount of moisture each contained were highly correlated, requiring the
choice of only one (i.e. percent moisture) in the analysis. Air temperatures were not
included in the model because invertebrate samples were collected only once a month
which truncated temperature measurements to a scale too coarse to be meaningful.
However, the variable “month” included all effects other than moisture and
control/treatment, including temperature extremes (freezing nights, very warm days) and
trophic interactions (birth/recruitment, predation, parasitism/disease, movement).
I used a general linear model (GLM) analysis of variance that accommodated
repeated measures to test for significant effects of the three independent variables
(treatment, moisture, month), and their interactions, on each invertebrate group. The
analysis was conducted using SAS 9.2 (2008). A generalized linear model was not used
because the residual error was too large to be considered a good fit; residuals increased
consistently with increasing counts. Invertebrate counts were log transformed
[log(Count+1)] to meet the assumptions of normality. Residuals were examined to assess

13
the adequacy of these transformations. The residuals were approximately normal and
relatively constant across the predicted values following the transformation.
The response variables used in the general linear model analysis were the log
transformed counts of the density of each invertebrate family. Invertebrate families
commonly consumed by Ensatina (Bury and Martin 1973) were each broken into three
response variables (each size class) to increase the resolution of impacts to these groups.
Invertebrate families not commonly consumed by Ensatina (Bury and Martin 1973) were
each analyzed as a single response variable which included all sizes. Invertebrate
families identified as Ensatina prey, but which contained insufficient data to be analyzed
separately by size class were each treated as a single response variable that included all
sizes. Invertebrate families were also combined into functional groups to assess Ensatina
effects at a coarse level of resolution; the invertebrate decomposer group contained an
adequate sampling size which enabled the analysis of this functional group by size class.
The GLM terms for each response variable were as follows (see Table 1 for definitions):
The plot was the sampling level of the analysis, with 12 plots in total divided
equally between control and treatment, N=6. In Year One N=5 due to removal of one
treatment plot from analysis due to salamander escape. The GLM was applied once to
each invertebrate group in each year in an exploratory and not confirmatory analysis,
therefore, Bonferroni adjustments for multiple comparisons were not deemed appropriate.
The change in dry weight of the leaf litter bags from start to end were compared (control
vs. treatment) for each year using ANOVA in NCSS (Hintze 2001).

14
Table 1. General linear model equation terms and their explanations; used in the analysis
comparing invertebrate densities between control and treatment plots in each year.
Equation term (Effect) Data type Explanation
Month categorical Month of experiment (0-4)
Control_Treatment categorical Treatment of plot/ or Control
Percent Moisture continuous Percent moisture by weight
Month*Control_Treatment categorical Interaction: treatment and month
Month*Moisture continuous Interaction: moisture and month
Moisture*Control_Treatment continuous Interaction: moisture and treatment
Moisture * Month * Treatment continuous 3- interaction: moisture, month, treatment
ra
continuous Repeated measures error
e continuous Residual error, random error aSAS 9.2 2008: PROC MIXED, autoregressive covariance structure of order 1 was used

15
RESULTS
In Year One (2007-2008), leaf litter cores from the 10 experimental plots yielded
14,401 individual invertebrates (57.4 individuals/gram of leaf litter) from 38 families. In
Year Two (2008-2009), litter cores from the 12 plots yielded 32,721 invertebrates (253.1
individuals/gram of litter) from 48 families (Appendix A). Invertebrates were about half
as dense in leaf litter cores during the 2007-2008 sampling year (Figure 2) as they were in
the 2008-2009 samples (Figure 3); however, the relative composition of the functional
groups was nearly identical in the two years. The majority of invertebrates found in the
leaf litter were decomposers (95%); herbivores, predators, and omnivores (ants) together
constituted a minority, comprising about 5% of all invertebrates found in litter samples in
each year. The decomposers were comprised overwhelmingly of mites (~65%), which
were almost 3 times as dense as springtails (~25%); all others constituted about 10%.
Invertebrate density was similar between control and treatment plots prior to
salamander introductions in 2007 (t=0.12, df=190, p=0.90, Figure 4) and 2008 (t=-0.05,
df=230, p=0.96, Figure 5). Invertebrate density was significantly influenced by percent
moisture of litter samples in 2007-2008 (f=17.48, df=190, p<0.0001) and 2008-2009
(f=18.52, df=230, p<0.0001, Table 2). The density of invertebrates appeared to fluctuate
with available moisture over time, in each year, with the majority of these fluctuations
attributed to small invertebrate decomposers (Table 2, Figures 4 and 5). The presence of
Ensatina did not significantly affect overall invertebrate density in 2007-2008 (f=0.03,
df=190, p=0.87) or 2008-2009 (f=0.73, df=230, p=0.39; Table 2), however, Ensatina did
influence the densities of individual invertebrate taxa in each year (see below).
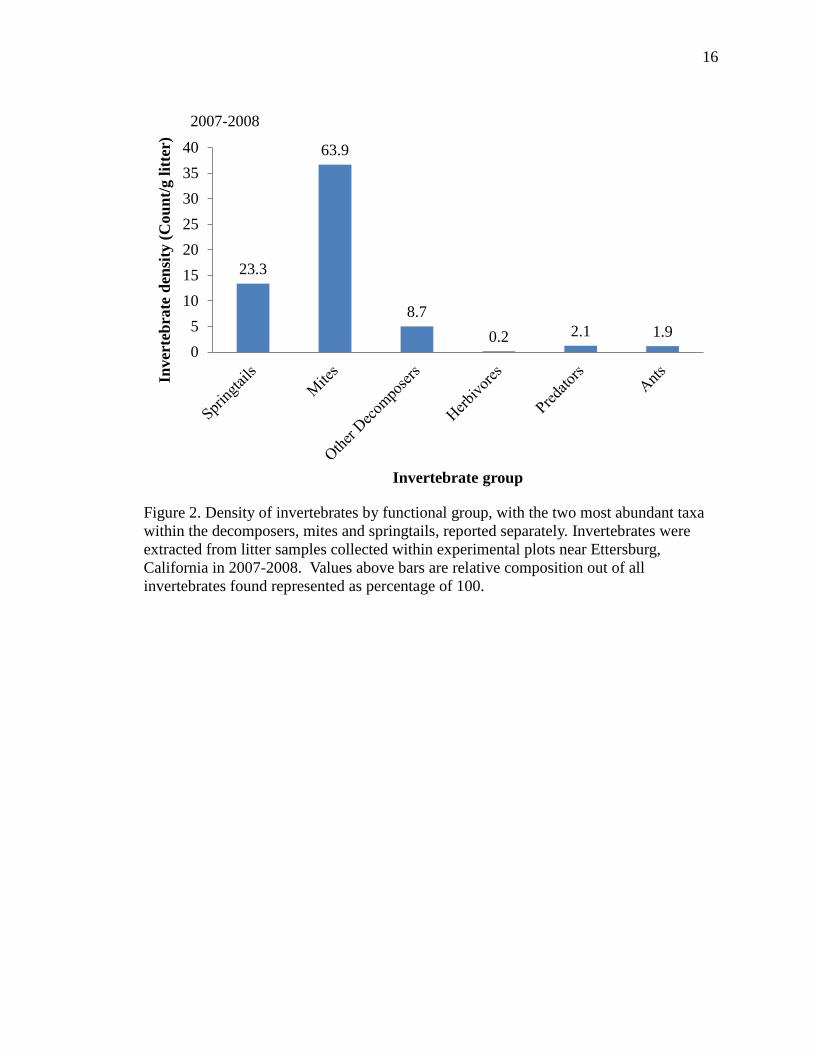
16
Figure 2. Density of invertebrates by functional group, with the two most abundant taxa
within the decomposers, mites and springtails, reported separately. Invertebrates were
extracted from litter samples collected within experimental plots near Ettersburg,
California in 2007-2008. Values above bars are relative composition out of all
invertebrates found represented as percentage of 100.
23.3
63.9
8.7
0.2 2.1 1.9
0
5
10
15
20
25
30
35
40In
ver
teb
rate
den
sity
(C
ou
nt/
g l
itte
r)
Invertebrate group
2007-2008

17
Figure 3. Density of invertebrates by functional group, with the two most abundant taxa
within the decomposers, mites and springtails, reported separately. Invertebrates were
extracted from litter samples collected within experimental plots near Ettersburg,
California in 2008-2009. Values above bars are relative composition out of all
invertebrates found represented as percentage of 100.
21.7
68.8
6.1 1.1 1.8 1.4
0
25
50
75
100
125
150
175
200
Inver
teb
rate
den
sity
(C
ou
nt/
g l
itte
r)
Invertebrate group
2008-2009

18
0
0.5
1
1.5
2
2.5
3
Nov Dec Jan Feb Mar
Den
sity
of
dec
om
pose
rs
<1m
m l
on
g (
Cou
nt/
g l
itte
r)
Month
Per
cen
t li
tter
mois
ture
Ensatina introduction
0
0.5
1
1.5
2
2.5
3In
ver
teb
rate
den
sity
(C
ou
nt/
g l
itte
r)
Control Treatment
Figure 4. Mean density of all invertebrates and invertebrate decomposers <1mm on
control and treatment plots sampled at 5 monthly intervals in 2007-2008 from Mattole
field sites near Ettersburg, California. The blue line represents the percent litter moisture
in 2007-2008. Error bars are ± one standard error.
Per
cen
t li
tter
mois
ture
Moisture
Ensatina introduction
0
0.05
0.1
0.15
0.2
0.25
2007-2008
0
0.05
0.1
0.15
0.2
0.25

19
0
1
2
3
4
5
6
7
8In
ver
teb
rate
den
sity
(Cou
nt/
gli
tter
)
Control Treatment
0
1
2
3
4
5
6
7
8
Jan Feb Mar Apr May
Den
sity
of
dec
om
pose
rs
<1m
m l
on
g (
Cou
nt/
g l
itte
r)
Month
Per
cen
t li
tter
mois
ture
Ensatina introduction
Per
cen
t li
tter
mois
ture
Moisture
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
Figure 5. Mean density of all invertebrates and invertebrate decomposers <1mm on
control and treatment plots sampled at 5 monthly intervals in 2008-2009 from Mattole
field sites near Ettersburg, California. The blue line represents the percent litter moisture
in 2008-2009. Error bars are ± one standard error.
2008-2009
Ensatina introduction

20
Table 2. Analysis of the effects of salamander predation (Control_Treatment), moisture, month, and the interaction of
month*moisture, on invertebrate functional groups by size class in two years using a general linear model. Data were analyzed
separately by group, size, and year. Results indicated with – were not statistically significant at α = 0.1.
Moisture Month Month*Moisture Control_Treatment
Group Size Year F-value P-value F-value P-value F-value P-value F-value P-value
Decomposers <1 mm 1 10.31 0.002 - - 2.09 0.083 - -
<2 mm 1 9.91 0.002 - - - - - -
>2 mm 1 - - - - - - - -
<1 mm 2 20.94 0.000007 5.01 0.001 5.56 0.0003 - -
<2 mm 2 3.92 0.049 3.18 0.014 4.41 0.002 - -
>2 mm 2 22.66 0.000003 2.56 0.039 2.37 0.053 - -
Omnivores (Ants) All sizes 1 14.24 0.0002 - - - - - -
All sizes 2 - - - - - - - -
Predators All sizes 1 3.64 0.058 - - - - - -
All sizes 2 - - - - 2.02 0.092 - -
Herbivores All sizes 1 - - - - - - - -
All sizes 2 - - 9.81 0.000 - - - -
All invertebrates All sizes 1 17.48 0.00004 2.09 0.08 2.23 0.07 - -
All sizes 2 18.52 0.00002 4.84 0.0009 6.06 0.0001 - -

21
Ensatina effects on invertebrate taxa
In 2007-2008 the densities of 10 invertebrate taxa were significantly affected by
Ensatina presence (6 decreased, 4 increased; Table 3); in contrast, only three taxa were
affected by Ensatina in 2008-2009, all of which increased on treatment compared to
control plots. Two of the taxa which declined on treatment plots in Year One, rebounded
to densities higher than controls following the initial declines (Figure 6). There were no
significant declines on treatment plots in Year Two, although two taxa that had declined
significantly in Year One showed declining trends in the Year Two but did not achieve
statistical significance: medium Entomobryidae springtails and beetles (see Figure 15).
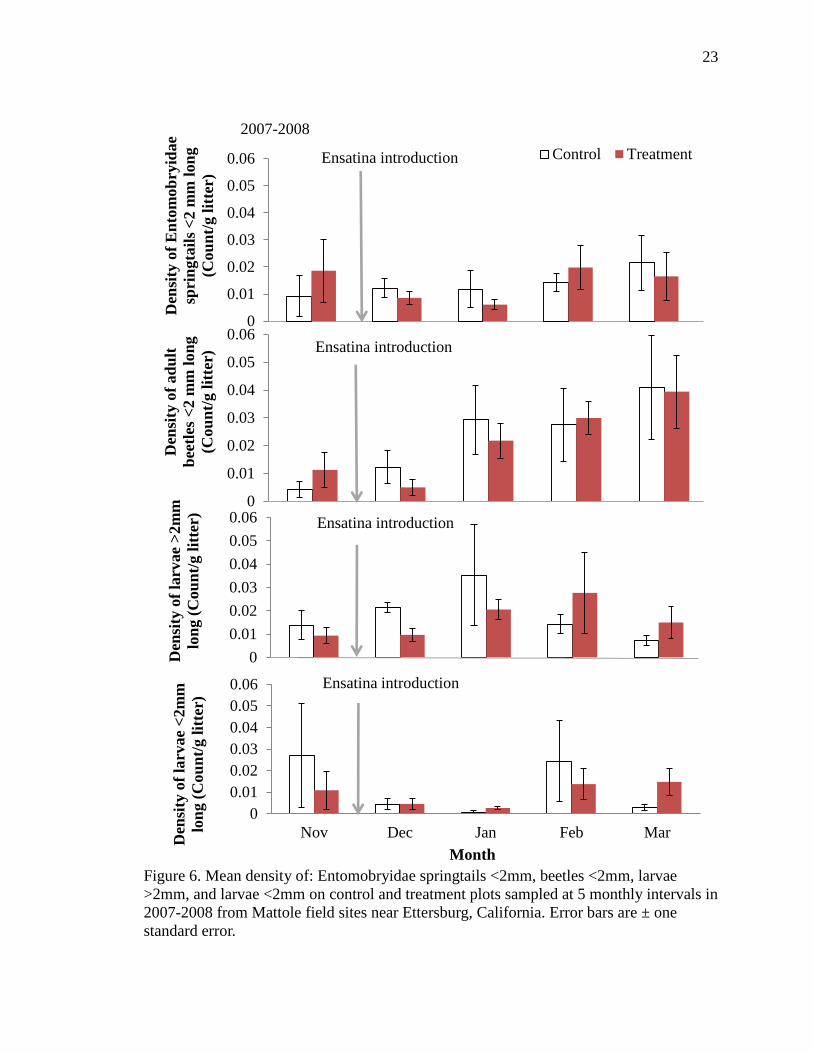
In Year One (2007-2008) the density of medium Entomobryidae springtails
(f=3.32, df=190, p=0.07), medium beetles (f=6.39, df=190, p=0.01), and large larval
shredders (f=2.79, df=190, p=0.09) each declined significantly on plots with Ensatina in
month one and two (Table 3, Figure 6). The density of medium Entomobryidae
springtails and large larval shredders each increased in month three following their
declines, to a level greater than adjacent control sites (Figure 6). Large larval shredders
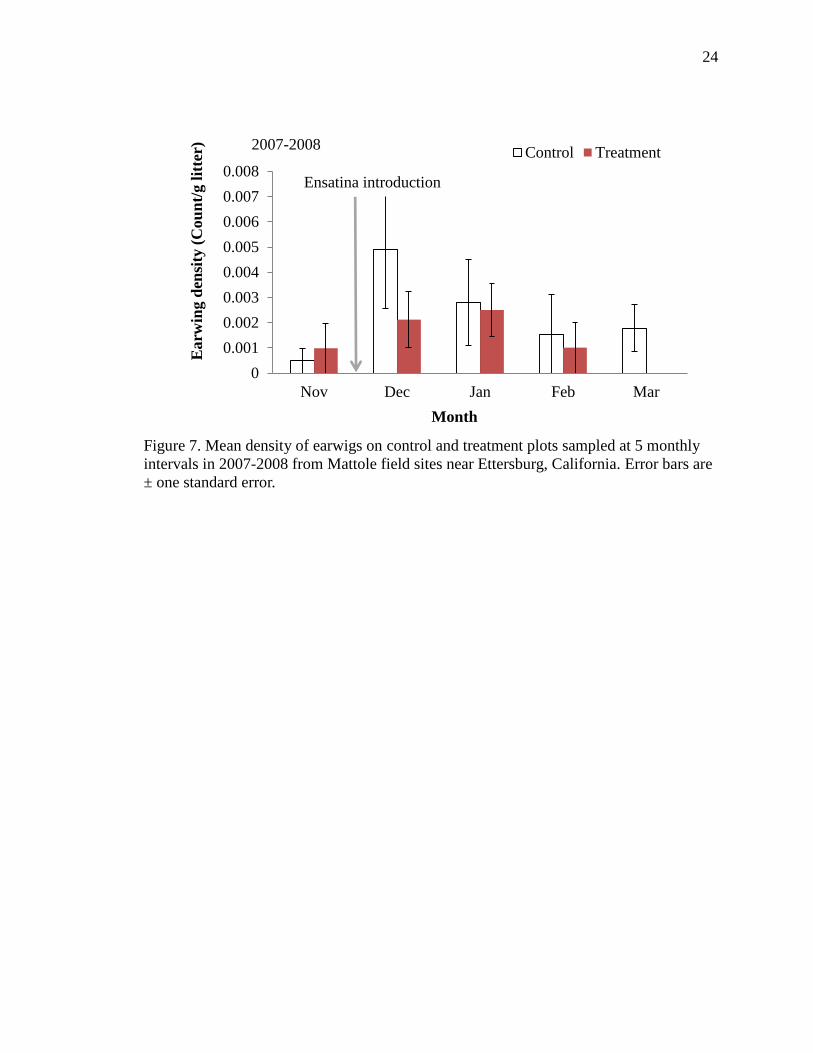
remained greater than controls into month four (Figure 6). The density of earwigs
declined significantly on treatments in months one and four (f=3.57, df=190, p=0.06,
Figure 7). The density of medium larval shredders remained similar between control and
treatment plots in month one, but were found in significantly higher densities on
treatments in months two and four (f=3.85, df=190, p=0.05, Table 3, Figure 6).
The density of small Orabatidae mites increased significantly on plots with
Ensatina within the first month following Ensatina introduction and again in month four
(f=2.69, df=190, p=0.32, Table 3, Figure 8); interaction between treatment and month.
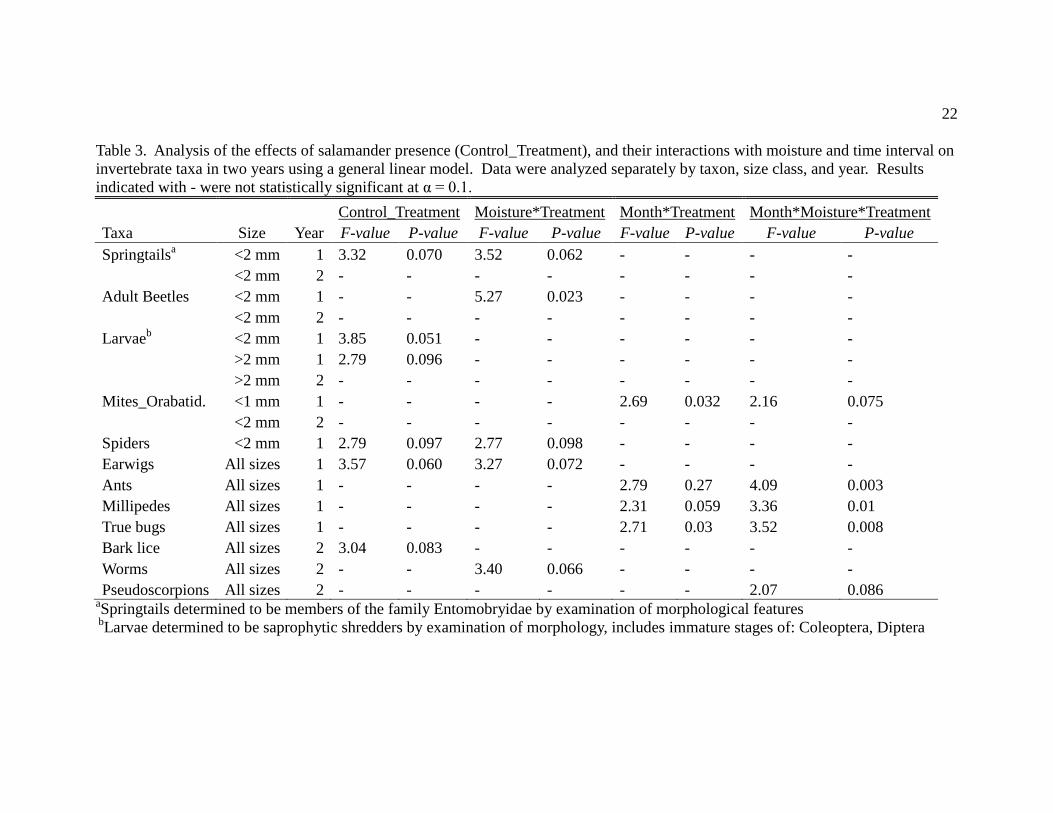
22
Table 3. Analysis of the effects of salamander presence (Control_Treatment), and their interactions with moisture and time interval on
invertebrate taxa in two years using a general linear model. Data were analyzed separately by taxon, size class, and year. Results
indicated with - were not statistically significant at α = 0.1.
Control_Treatment Moisture*Treatment Month*Treatment Month*Moisture*Treatment
Taxa Size Year F-value P-value F-value P-value F-value P-value F-value P-value
Springtailsa <2 mm 1 3.32 0.070 3.52 0.062 - - - -
<2 mm 2 - - - - - - - -
Adult Beetles <2 mm 1 - - 5.27 0.023 - - - -
<2 mm 2 - - - - - - - -
Larvaeb
<2 mm 1 3.85 0.051 - - - - - -
>2 mm 1 2.79 0.096 - - - - - -
>2 mm 2 - - - - - - - -
Mites_Orabatid. <1 mm 1 - - - - 2.69 0.032 2.16 0.075
<2 mm 2 - - - - - - - -
Spiders <2 mm 1 2.79 0.097 2.77 0.098 - - - -
Earwigs All sizes 1 3.57 0.060 3.27 0.072 - - - -
Ants All sizes 1 - - - - 2.79 0.27 4.09 0.003
Millipedes All sizes 1 - - - - 2.31 0.059 3.36 0.01
True bugs All sizes 1 - - - - 2.71 0.03 3.52 0.008
Bark lice All sizes 2 3.04 0.083 - - - - - -
Worms All sizes 2 - - 3.40 0.066 - - - -
Pseudoscorpions All sizes 2 - - - - - - 2.07 0.086 aSpringtails determined to be members of the family Entomobryidae by examination of morphological features
bLarvae determined to be saprophytic shredders by examination of morphology, includes immature stages of: Coleoptera, Diptera
23
0
0.01
0.02
0.03
0.04
0.05
0.06
Den
sity
of
ad
ult
bee
tles
<2 m
m l
on
g
(Cou
nt/
g l
itte
r) Ensatina introduction
0
0.01
0.02
0.03
0.04
0.05
0.06
Den
sity
of
larv
ae
>2m
m
lon
g (
Cou
nt/
g l
itte
r)
0
0.01
0.02
0.03
0.04
0.05
0.06
Nov Dec Jan Feb Mar
Den
sity
of
larv
ae
<2m
m
lon
g (
Cou
nt/
g l
itte
r)
Month
Ensatina introduction
0
0.01
0.02
0.03
0.04
0.05
0.06D
ensi
ty o
f E
nto
mob
ryid
ae
spri
ngta
ils
<2 m
m l
on
g
(Cou
nt/
g l
itte
r)
Control TreatmentEnsatina introduction
Figure 6. Mean density of: Entomobryidae springtails <2mm, beetles <2mm, larvae
>2mm, and larvae <2mm on control and treatment plots sampled at 5 monthly intervals in
2007-2008 from Mattole field sites near Ettersburg, California. Error bars are ± one
standard error.
Ensatina introduction
2007-2008
24
Figure 7. Mean density of earwigs on control and treatment plots sampled at 5 monthly
intervals in 2007-2008 from Mattole field sites near Ettersburg, California. Error bars are
± one standard error.
0
0.001
0.002
0.003
0.004
0.005
0.006
0.007
0.008
Nov Dec Jan Feb Mar
Earw
ing d
ensi
ty (
Cou
nt/
g l
itte
r)
Month
Control Treatment
Ensatina introduction
2007-2008

25
Figure 8. Mean density of Orabatidae mites <1mm on control and treatment plots
sampled at 5 monthly intervals in 2007-2008 from Mattole field sites near Ettersburg,
California. Error bars are ± one standard error.
0
0.2
0.4
0.6
0.8
1
1.2
Nov Dec Jan Feb Mar
Den
sity
of
Ora
ba
tid
ae
mit
es
<1 m
m l
on
g (
Cou
nt/
g l
itte
r)
Month
Control Treatment
Ensatina introduction
2007-2008

26
In Year One medium spiders were found at greater density on plots with Ensatina
compared to controls throughout the four months of the experiment (f=2.79, df=190,
p=0.097, Table 3, Figure 9). The density of ants was low on all plots through the first
three months of Year One, however decreased significantly on plots with Ensatina in the
final month of the experiment, indicating the significant interaction between month,
moisture, and treatment (f=4.09, df=190, p=0.003, Figure 10). The density of millipedes
was greater on treatment plots in months one, two, and four (f=3.36, df=190, p=0.01,
Figure 11). The density of true bugs (Hemiptera) was significantly lower on treatments in
Year One (f=3.52, df=190, p=0.008; Table 3, Figure 12), however, also rare (0.00005%
of all invertebrates), likely influencing comparisons and significance.
In Year Two the density of barklice (Psocoptera) increased significantly on plots
with Ensatina compared to controls in months two and four (f=3.04, df=230, p=0.08;
Table 3, Figure 13). The density of worms (Annelida) increased significantly on plots
with Ensatina over controls during months two and four, indicating an interaction
between treatment and moisture (f=3.4, df=230, p=0.07; Table 3, Figure 13). The density
of Pseudoscorpions increased significantly on plots with Ensatina in month one and again
in months three and four of Year Two (f=2.07, df=230, p=0.09; Table 3, Figure 14);
indicating the interaction between month, moisture, and treatment.
During the latter three months of Year Two the densities of four taxa which
differed significantly between control and treatment in Year One (Fig. 6, Table 3) showed
a similar trend in the second year: medium Entomobryidae springtails, medium beetles,
medium Orabatidae mites, and large larval shredders. The differences in densities of
these taxa did not achieve statistical significance in the second year (Figure 15, Table 3).
27
Figure 9. Mean density of spiders <2mm on control and treatment plots sampled at 5
monthly intervals in 2007-2008 from Mattole field sites near Ettersburg, California. Error
bars are ± one standard error.
0
0.001
0.002
0.003
0.004
0.005
0.006
0.007
Nov Dec Jan Feb Mar
Den
sity
of
spid
ers
<2 m
m l
on
g
(Co
un
t/g
lit
ter)
Month
Control Treatment
2007-2008
Ensatina introduction
28
Figure 10. Mean density of ants on control and treatment plots sampled at 5 monthly
intervals in 2007-2008 from Mattole field sites near Ettersburg, California. Error bars are
± one standard error.
00.020.040.060.08
0.10.120.140.160.18
0.2
Nov Dec Jan Feb Mar
An
t d
ensi
ty (
Cou
nt/
g l
itte
r)
Month
Control Treatment
Ensatina introduction
2007-2008

29
Figure 11. Mean density of millipedes on control and treatment plots sampled at 5
monthly intervals in 2007-2008 from Mattole field sites near Ettersburg, California. Error
bars are ± one standard error.
0
0.005
0.01
0.015
0.02
0.025
0.03
0.035
0.04
Nov Dec Jan Feb Mar
Mil
lip
ede
den
sity
(C
ou
nt/
g l
itte
r)
Month
Control Treatment
Ensatina introduction
2007-2008
30
Figure 12. Mean density of true bugs (Hemiptera) on control and treatment plots sampled
at 5 monthly intervals in 2007-2008 from Mattole field sites near Ettersburg, California.
Error bars are ± one standard error.
0.000
0.001
0.001
0.002
0.002
0.003
0.003
Nov Dec Jan Feb Mar
Den
sity
of
tru
e b
ugs
(Cou
nt/
g l
itte
r)
Month
Control Treatment
Ensatina introduction
2007-2008

31
0
0.01
0.02
0.03
0.04
0.05
Jan Feb Mar Apr May
Worm
den
sity
(C
ou
nt/
g l
itte
r)
Month
0
0.01
0.02
0.03
0.04
0.05B
ark
lice
den
sity
(C
ou
nt/
g l
itte
r) Control Treatment
Ensatina introduction
Figure 13. Mean density of: barklice (Psocoptera) and worms (Annelida) on control and
treatment plots sampled at 5 monthly intervals in 2008-2009 from Mattole field sites near
Ettersburg, California. Error bars are ± one standard error.
2008-2009
Ensatina introduction

32
Figure 14. Mean density of Pseudoscorpions on control and treatment plots sampled at 5
monthly intervals in 2008-2009 from Mattole field sites near Ettersburg, California. Error
bars are ± one standard error.
0.000
0.002
0.004
0.006
0.008
0.010
0.012
0.014
0.016
Jan Feb Mar Apr May
Pse
ud
osc
orp
ion
den
sity
(Co
un
t/g
lit
ter)
Month
Control TreatmentEnsatina introduction
2008-2009
33
0.000.020.040.060.080.100.120.140.160.18
Den
sity
of
ad
ult
bee
tles
<2 m
m l
on
g
(Cou
nt/
g l
itte
r)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
Den
sity
of
En
tom
ob
ryid
ae
spri
ngta
ils
<2 m
m l
on
g
(Co
un
t/g
lit
ter)
Control Treatment
0.000.020.040.060.080.100.120.140.160.18
Den
sity
of
Ora
bati
dae
mit
es <
2m
m l
on
g
(Cou
nt/
g l
itte
r)
0.000.020.040.060.080.100.120.140.160.18
Jan Feb Mar Apr May
Den
sity
of
larv
ae
>2 m
m l
on
g
(Cou
nt/
g l
itte
r)
Month
Figure 15. Mean density of: Entomobryidae springtails <2mm, beetles <2mm, Orabatidae
mites <2mm and larvae >2mm, on control and treatment plots sampled at 5 monthly
intervals in 2008-2009 from Mattole field sites near Ettersburg, California. Error bars are
± one standard error.
Ensatina introduction
Ensatina introduction
Ensatina introduction
Ensatina introduction 2008-2009

34
Leaf-litter
In Year One (2007-2008) mean leaf litter mass (Χ± SE) was significantly greater
on treatments (2.68 g ± 0.06 g) compared with controls (2.28 g ± 0.06 g) after the four
months of the experiment (t=-5.32, df=28, p<0.0001; Figure 16). The retention of leaf-
litter was significantly greater (leaf-litter breakdown reduced) by 13.3% (± 0.2%) on
treatments than on controls in 2007-2008.
In Year Two (2008-2009) the mean litter mass was not significantly different
between controls (2.06 g ± 0.07 g) and treatments (2.23 g ± 0.09 g) over the four months
of the experiment (t=-1.43, df=34, p=0.16; Figure 16). The average retention of litter
mass across all plots (control and treatment) in 2008-2009 (2.04 g ± 0.05 g) was
significantly less (t=4.80, df=64, p=0.00001) than it was in 2007-2008 (2.51 g ± 0.05 g
(Figure 16); there was significantly more leaf litter breakdown across all plots in the
second year.
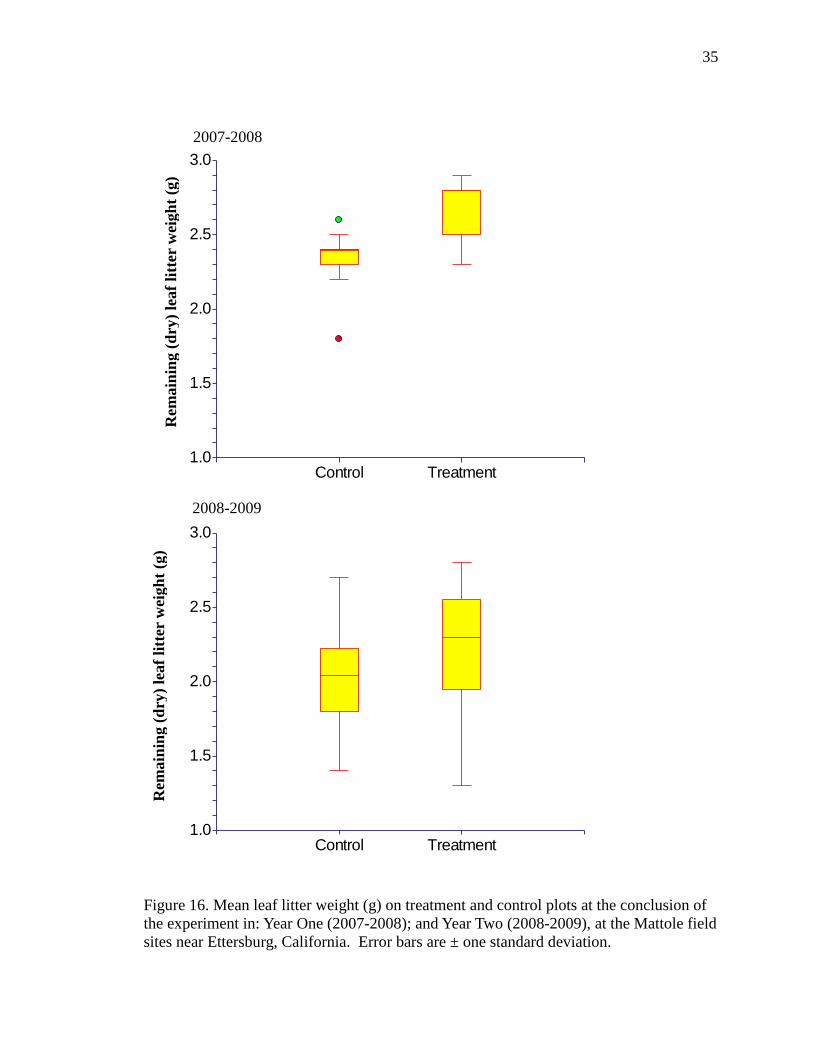
35
Figure 16. Mean leaf litter weight (g) on treatment and control plots at the conclusion of
the experiment in: Year One (2007-2008); and Year Two (2008-2009), at the Mattole field
sites near Ettersburg, California. Error bars are ± one standard deviation.
1.0
1.5
2.0
2.5
3.0
Control Treatment
1.0
1.5
2.0
2.5
3.0
Control Treatment
Rem
ain
ing (
dry
) le
af
litt
er w
eigh
t (g
) R
em
ain
ing (
dry
) le
af
litt
er w
eigh
t (g
)
2007-2008
2008-2009

36
DISCUSSION
The great abundances of terrestrial salamanders in North American forests (e.g.,
Burton and Likens 1978a, Welsh and Lind 1992, Petranka and Murray 1999) implies an
important role in forest floor food web dynamics through predation on invertebrate
assemblages including larval shredders and detrital grazers (beetles and springtails).
Members of the invertebrate shredder guild (larvae and worms) physically tear organic
material such as leaf litter, conifer needles, and wood on the forest floor into smaller
pieces, which are then processed through their gut, inoculated with microflora, and
utilized by microfauna (Gist and Crossley 1975). The small decomposers (mites,
springtails, and nematodes [together microfauna]) directly mediate the primary
productivity of this microfloral resource by fulfilling three ecological functions: grazing,
spreading propogules, and preying on one another (McBrayer and Reichle 1971, Singh
1977). Ultimately, this often dense microscopic layer of primary and secondary
consumers converts primary microfloral productivity and waste into invertebrate
biomass, effectively transferring energy from forest materials up the food web and
simultaneously recharging soil nutrients, including carbon and nitrogen, for plant growth
(McBrayer and Reichle 1971, Gist and Crossley 1975, Singh 1977). Ensatina predation
on these two critical components of the decomposition food web (grazers, shredders)
indicates the potential for terrestrial salamanders to have a top down influence on the
processes of nutrient cycling and carbon storage in forests.
While Ensatina preys on many of these invertebrates, exerting a top-down
influence on the rate of litter breakdown, this relationship can be affected by moisture.

37
Detrital food web processes respond directly to moisture by increasing the densities of
invertebrate shredders, grazers, and microfloral growth (Wardle 2002), which increases
the rate of leaf litter breakdown (Gist and Crossley 1975, Singh 1977). In Year Two
(2008-2009) there was an initial high pulse of moisture within the first two months of the
experiment, probably resulting in the detection of twice as many invertebrates in litter
samples than in Year One. Consistent with this pulse of enhanced activity, leaf litter
breakdown was greater in Year Two than it was in Year One. Furthermore, significant
salamander effects on invertebrate densities were fewer in Year Two: three taxa increased
in Ensatina presence (barklice, worms, pseudoscorpions; indirect effects); compared to
six taxa which decreased (medium Entomobryidae springtails, medium beetles, earwigs,
large larval shredders, ants, true bugs; direct effects) and four taxa which increased (small
Orabatidae mites, medium spiders, medium larva, millipedes; indirect effects), Year One.
In Year One, the direct and indirect effects of Ensatina presence on the
invertebrate assemblage was apparent within the first month of the experiment. The
densities of three microfloral grazers: medium Entomobryidae springtails, medium
beetles and earwigs each decreased immediately following Ensatina introduction. While
simultaneously the density of a small microfloral grazer increased: Orabatidae mites less
than one millimeter long. The removal of many of these larger grazers by Ensatina
would have likely opened up primary resources for the smaller and more numerous mites,
allowing them to capitalize on these resources and increase in density (competitive
release). A similar relationship was found with Plethodon cinereus where decreases in
the abundance of Entomobryidae springtails on salamander plots resulted in an increase
in mites and Podomorphic springtails (Walton and Steckler 2005). Rooney (2000) and

38
Walton et al. (2006) also described a similar release of Podomorphic springtails in the
presence of this salamander. In my study, the larger and highly mobile millipedes (a
microfloral grazer) also apparently capitalized on available microfloral resources,
increasing significantly in density on Ensatina plots. Large larval shredders decreased on
Ensatina plots in the first two months, allowing medium larval shredders to increase.
Consistent with the reduced densities of large larval shredders, and microfloral
grazers (springtails, beetles, earwigs) on treatments in Year One was an increase to mean
leaf litter retention by 13.3%, compared to control plots. Consistent with the pulse of
moisture early in the experiment in Year Two, and the high invertebrate densities in that
year, was an increase in the breakdown of litter across all plots, compared to Year One.
Consequently, the salamander treatment plots showed a lack of statistically significant
declines to invertebrate densities and an insignificant retention of litter on treatment plots
in Year Two. While mean dry weight of litter retained on treatments was 5% greater than
controls in Year Two, which may indicate the consumption of invertebrate decomposers
by Ensatina, there was an insufficient effect to achieve statistical significance.
With a single Ensatina salamander per 1.5 m2 treatment plot, the increased
invertebrate densities in Year Two probably decreased the likelihood of a single
salamander being able to consume enough invertebrates to achieve a statistically
significant decline. However, it appears that these Ensatina did consume high numbers of
medium Entomobryidae springtails, beetles, and mites in Year Two despite the lack of
statistically significant differences. Evidence of this is apparent in the indirect effect on a
small microfloral grazer: barklice (Psocoptera) which increased in density on plots with
Ensatina in month two; precisely when the densities of these three grazers (springtails,

39
beetles, mites) began to fluctuate on treatments. By month four barklice density was
three times greater on plots with Ensatina than on controls, indicating the compounding
indirect effects of Ensatina predation on larger microfloral grazers (medium-sized
Entomobryidae springtails, beetles, and mites) in the last three months of the experiment
in Year Two (2008-2009). Walton (2005) also observed increases in barklice on treatment
plots, apparently due to this competitive release phenomenon.
Gnaedinger and Reed (1948) reported the stomachs of 21 Ensatina contained in
decreasing frequency: springtails, spiders, millipedes, centipedes, beetles, larvae, and
mites, followed by pill bugs, thrips, and wasps. Bury and Martin (1973) found the
stomachs of 37 Ensatina contained in decreasing frequency: springtails, spiders, beetles,
millipedes, pill bugs, larvae, ants, and mites; with several other taxa including centipedes
and pseudoscorpions found in fewer than 5% of stomachs. In both studies the most
frequently consumed invertebrates included springtails, spiders, beetles, and larvae; the
densities of each differed in the presence of Ensatina in this study.
While I did not detect significant declines to invertebrate predators (possibly due
to small sample sizes), there was evidence of an indirect increase to an intermediate
invertebrate predator in each year (meso-predator release [Richie and Johnson 2009]). In
Year One medium spiders occurred in higher densities on plots with Ensatina compared
to controls. In Year Two Pseudoscorpions occurred in higher densities on plots with
Ensatina compared to controls. Gnaedinger and Reed (1948) and Bury and Martin (1973)
indicated spiders as an important food source for Ensatina; Ensatina predation on large
spiders may explain the increase in density of medium spiders on treatment plots in Year
One (2007-2008) and Pseudoscorpions in Year Two (2008-2009).

40
The influence of moisture and prey density
In Year One the density of invertebrates was high prior to introduction of
Ensatina, and was quite low on all plots in the first month following Ensatina population.
The density of invertebrates gradually increased from a low in month one through month
four of 2007-2008, apparently, as the percent moisture of litter samples increased. In the
first two months of the first year Ensatina appeared to capitalize on select invertebrates:
large larval shredders, medium Entomobryidae springtails, medium beetles, and earwigs.
By month three the significant differences (control vs. treatment) to these four taxa were
no longer apparent and at least two taxa: medium Entomobryidae springtails and large
larval shredders began to increase to densities higher than controls. In months three and
four invertebrate densities continued to increase and significant differences to these select
taxa disappeared: only earwigs and ants were less dense on treatments in month four.
In Year Two (2008-2009) invertebrate density was much higher than in Year One
(2007-2008) and I failed to detect any statistically significant declines to invertebrate taxa
on the salamander plots in Year Two (Tables 2 and 3). However, a statistically significant
indirect effect of Ensatina presence (e. g. increased density of barklice) suggests Ensatina
did consume invertebrate microfloral grazers (e. g. medium Entomobryidae springtails,
beetles, and Orabatidae mites) during months two-four of Year Two. Furthermore, these
latter three months of Year Two coincide with the decline of moisture in litter samples
and declining invertebrate density. This may have forced Ensatina to be more selective of
prey species as availability of prey decreased (Jaeger and Barnard 1981); similar to
Stamps et al. (1981) and Diaz and Carrascal (1993) whom found prey selection decrease
and eventually disappear as prey availability increased for insectivorous lizards.

41
Ensatina and optimal foraging theory
Optimal foraging theory states that predators will maximize gains and minimize
efforts by first selecting prey which provide the most energy gained per energy invested
(most profitable) and then broadening that selection to include less profitable prey as the
more preferred prey decline in density (Emlen 1966). This is based on an energy limited
model developed primarily for endothermic predators with high caloric requirements.
Salamanders are poikilotherms with low energetic requirements (Pough 1980); Ensatina
is more efficient than an endothermic insectivore (e. g. birds utilize 90% of calories for
respiration) at converting ingested calories into biomass (Burton and Likens 1975, Pough
1983). The low energetic requirements of Ensatina allow it to include abundant prey
even when energetically less profitable (Jaeger and Barnard 1981, Stamps et al. 1981,
Diaz and carascal 1993), and consider the relative nutritional qualities (complimentary
amino acids, etc.) of different taxa (Pulliam 1975, Stamps et al. 1981, Mayntz and Toft
2001). These circumstances enabled Ensatina to modulate foraging behaviors (prey
selection) in response to environmental conditions (moisture, prey density).
Ensatina is a sit-and-wait predator that invests little energy in foraging behavior so
a majority of foraging cost is inherent in the relative percentage of exoskeleton chitin in
each invertebrate taxa consumed (Jaeger 1990, Diaz and Carrascal 1993). A regression
analysis comparing handling time to prey size (mean dry mass) found relatively shallow
slopes (slow increase in handling time with size) for particularly round and/or soft bodied
taxa: true bugs, larvae, flies, and spiders but steep slopes (handling times increased
rapidly with size) for highly chitinized and elongated taxa: crickets, beetles, ants, etc.
(Jaeger 1990, Diaz and Carrascal 1993). Entomobryidae springtails are armored with

42
hairs and an enlarged dorsal segment which may place them in the high chitin group,
along with beetles and ants. Medium Entomobryidae springtails and beetles seem to be
important prey types for Ensatina in forests of Northern coastal California as they were
consumed in each year. Large beetles and springtails although common in samples
(appendix A) were not significantly reduced by Ensatina, compared to the medium size
class of each which were significantly reduced in the first two months of Year One.
Large larval shredders were important prey for Ensatina during the first two months of
Year One in particular, when prey density was the lowest recorded during this study. This
is in contrast to medium larval shredders which increased on treatments at the same time.
Temperate invertebrate communities are highly skewed towards smaller species
which can provide an ample prey base for small insectivores (Whitaker 1952, Stamps et
al. 1981). Jaeger (1980) confirms that prey are only very rarely limiting for terrestrial
salamanders, but rather may become temporarily unavailable due to low moisture or high
temperatures which can threaten salamanders with desiccation, suggesting moisture
availability may influence Ensatina foraging behaviors. During the first two months of
Year One, invertebrate density was slowly increasing from a low point and Ensatina
consumed prey within energetically favorable taxa (large larval shredders, medium
Entomobryidae springtails and beetles). In month three of Year One Ensatina did not
significantly impact these groups, and as moisture and invertebrate density approached a
peak for the experiment in month four, Ensatina consumed ants and earwigs (high chitin).
It may be that relatively limited prey availability in Year One influenced Ensatina
to consume more energetically advantageous prey (calories/cost) to maintain a positive
energy budget (e.g., Jaeger 1990, Diaz and Carrascal 1993). This may have become less

43
important in months three and four as prey density increased. Alternatively, but not
exclusively, it may be that the consumption of prey under cover objects during periods of
relatively low moisture (Jaeger 1980) influenced Ensatina to capitalize on particular taxa
commonly encountered under the cover provided (beetles, Entomobryidae springtails,
larvae). The increased moisture of months three and four of Year One may have enabled
Ensatina to explore more ground during foraging and consume a wider variety of prey.
These two phenomena were likely included in the dynamics of Year Two as well,
where top-down salamander effects did not significantly decrease the densities of any
invertebrate taxa: with moisture and invertebrate densities high there was nothing to limit
the foraging behaviors of Ensatina. Furthermore, the significant indirect effect of
Ensatina presence, which increased the density of barklice, first occurred in month two
and again in month four when moisture and invertebrate density were each most limited
in Year Two. The early pulse of moisture in Year Two may have limited the ability of
Ensatina to regulate the invertebrate community (bottom-up versus top-down forces) and
indirectly influence a greater retention of leaf litter. The differences I found between
years in the regulation of invertebrate densities by salamanders due to variation in
moisture is consistent with Walton (2005), who also found the downward effects of a
woodland salamander on invertebrates to be ameliorated by moisture. The ability of
moisture to influence the effects of top-down regulation (top-down cascades) has been
well documented for several other insectivorous predators (e.g., Anolis lizards [Spiller
and Schoner 1995]; arboreal birds [Bridgeland et al. 2010]), including terrestrial
salamanders in the Midwest (Walton 2005).

44
CONCLUSIONS AND RECOMMENDATIONS
Soils are the third largest active carbon pool globally (2,400 Pg of carbon in the
top 2 m) after the lithosphere and hydrosphere (Eshel et al. 2007); representing the largest
terrestrial reservoir of carbon (Zhou 2006, Hungate et al. 2009), especially in temperate
forests of the northern hemisphere (Beedlow 2004), where woodland salamanders are so
abundant and diverse. My results and others (Wyman 1998) indicate that terrestrial
salamanders play an ecologically dominant role at the soil-leaf litter interface of forested
ecosystems of temperate North America by promoting nutrient cycling and increasing the
retention of litter which stores energy, nutrients (nitrogen), and minerals including carbon
(Davic and Welsh 2004). The critical role of predators in maintaining ecosystem
functionality is now recognized (Richie and Johnson 2009, Estes et al. 2011). Terrestrial
salamanders clearly serve as predators in forested ecosystems, with the ability to generate
top-down effects on the invertebrate assemblage and increase the retention of leaf litter,
fostering storage of material for decomposition and increasing carbon buildup in the soil.
Further research is needed on the relative influences of sympatric terrestrial
salamanders on the detrital food web and on each other, as they relate to environmental
factors and leaf litter retention/turnover. As climatic variables continue to respond to the
effects of global climate change we need to continue to address the role of ecologically
dominant species like Ensatina in response to the impacts of climate extremes on forested
ecosystems. Increasing our understanding of the ecological linkages within these forests
can enhance our ability to better manage this resource while mitigating the effects of
anthropogenic climate change, in order to maintain these life-sustaining environments.

45
LITERATURE CITED
Beard, K. H., A. K. Eschtruth, K. A. Vogt, D. J. Vogt, and N. Scatena. 2003. The effects
of the frog Eleutherodactylus coqui on invertebrates and ecosystem processes at
two scales in the Luquillo experimental forest, Puerto Rico. Journal of Tropical
Ecology 19:607-617.
Beedlow, P. A., D. T. Tingey, D. L. Phillips, W. E. Hogsett, and D. M Olszyk. 2004.
Rising atmospheric CO2 and carbon sequestration in forests. Frontiers in Ecology
and Environment 2:315-322.
Benrong, C. and D. H. Wise. 1999. Bottom up limitation of predaceous arthropods in a
detritus based terrestrial food web. Ecological Society of America 80:761-772.
Bridgeland, W. T., P. Beier, T. Kolb, and T. G. Whitham. 2010. A conditional trophic
cascade: birds benefit faster growing trees with strong links between predators
and plants. Ecological Society of America 91(1):73-84.
Bury, R. B. and M. Martin. 1973. Comparative studies on the distribution and foods of
Plethodontid salamanders in the redwood region on Northern California. Journal
of Herpetology 7:331-335
Burton, T. M. and G. E. Likens. 1975a. Energy flow and nutrient cycling in salamander
populations in the Hubbard Brook experimental forest, New Hampshire. Ecology
56:1068-1080.
Burton, T. M. and G. E. Likens. 1975b. Salamander Populations and Biomass in the
Hubbard Brook Experimental Forest, New Hampshire. Copeia 3:541-546.
Chernova, N. M., A. I. Bokova, E. V. Varshav, N. P. Goloshcapova, and Y. Y. Savenkova.
2007. Zoophagy in Collembola. Entomological Review 87:799-811.
Davic, R. D. and H. Welsh, Jr. 2004. On the Ecological Roles of Salamanders. Annual
Review of Ecology and Evolutionary Systems 35:405-435.
Diaz, J. A. and L. M. Carrascal. 1993. Variation in the effect of profitability on prey size
selection by the Lacertid lizard Psammodromus algirus. Oecologia 94(1): 23-29.
Drift, J. v. d. and E. Jansen. 1977. Grazing of springtails on hyphal mats and its
influence on fungal growth and respiration. Ecological Bulletins 25:203-209.
Emlen, J. M. 1966. The role of time and energy in food preference. American Naturalist
100:611-617.

46
Eshel, G., P. Fine, and M. J. Singer. 2007. Total soil carbon and water quality: an
implication for carbon sequestration. Soil and Water Management and
Conservation 71: 397-405.
Gist C. S. and D. A. Crossley Jr. 1975. The litter arthropod community in a Southern
Appalachian hardwood forest: numbers, biomass, and mineral element content.
American Midland Naturalist, 93(1):107-122.
Gnaedinger, L. M. and C. A. Reed. 1948. Contribution to the natural history of the
Plethodont salamander Ensatina eschscholtzii. Copeia 3:187-196.
Gruner, D. S. 2004. Attenuation of top down and bottom up forces in a complex
terrestrial community. Ecology 85:3010-3022.
Hairston, N. G. sr., F. E. Smith, and L. B. Slobodkin. 1960. Community structure,
population control, and competition. American Naturalist 94:421-425.
Hairston N. G. Jr. and N. G. Hairston Sr. 1997. Does food web complexity eliminate
trophic level dynamics? The American Naturalist 149:1001-1007.
Halaj, J. and D. H. Wise. 2002. Impact of a detrital subsidy on trophic cascades in a
terrestrial grazing food web. Ecology 83:3141-3151.
Hickerson, C.-A. M. C. D. Anthony, and G. A, Wicknick. 2004. Behavioral interactions
between salamanders and centipedes: competition in divergent taxa. Behavioral
Ecology 15:679-686
Hintze, J. 2001. NCSS and PASS. Number Cruncher Statistical Systems. Kaysville,
Utah.
Homyak, J. A., E. B. Sucre, C. A. Haas, and T. R. Fox. 2010. Does Plethodon cinereus
affect leaf litter decomposition and invertebrate abundances in mixed oak forest?
Journal of Herpetology 44:447-456.
Hungate B. A., K.-J. v. Groenigen, J. Six, J. D. Jastrows, Yiqu Luo, M. A. de Graaf, C.
van Kessel and C. W. Osenberg. 2009. Assessing the effect of elevated carbon
dioxide on soil carbon: a comparison of four meta-analyses. Global Change
Biology 15:2020-2034.
Jaeger, R. G. 1980. Fluctuations in prey availability and food limitation in a terrestrial
salamander. Oecologia 44:335-341.
Jaeger, R. G. and D. E. Barnard. 1981. Foraging tactics of a terrestrial salamander:
choice of diet in structurally simple environments. The American Naturalist
117:639-664.

47
Jaeger, R. G. 1990. Terrestrial salamanders evaluate size and chitinous content of
arthropod prey. Pages 111-126, in R. N. Hughes, Editor. Behavioral mechanisms
of food selection. NATO ASI Series, Subseries G: Ecological Sciences. Springer,
Heidelberg, Germany.
Karraker, N. E. and H. H. Welsh Jr. 2006. Long-term impacts of even-aged timber
management on abundance and body condition of terrestrial amphibians in
Northwestern California. Biological Conservation 131:132-140.
Koagata, H. and T. Ogushi. 2006. Bottom-up trophic cascades and material transfer in
terrestrial food webs. Ecological Research 21:26-34.
Lynch, J. F. 1985. The feeding ecology of Aneides flavipunctatus and sympatric
Plethodontid salamanders in Northwester California. Journal of Herpetology
19:328-352.
Mayntz, D. and S. Toft. 2001. Nutrient composition of a prey’s diet affects the growth
and survivorship of a generalist predator. Oecologia 127:207-213.
McBrayer, J. F. and D. E, Reichle. 1971. Trophic structure and feeding rates of forest
soil invertebrate populations. Oikos 22(3):381-388.
McCay, T. S. and G. S. Storm. 1997. Masked shrew (Sorex cinereus) abundance, diet
and prey selection in an irrigated forest. American Midland Naturalist,
138(2):268-275.
Paine, R. T. 1980. Food webs: Linkage, interaction strength and community
infrastructure. Journal of Animal Ecology 94:666-685.
Petranka, J. W. 1998. Salamanders of the United States and Canada. Smithsonian Press,
Washington D. C.
Petranka, J. W., and S. S. Murray. 2001. Effectiveness of removal sampling for
determining salamander density and biomass: a case study in an Appalachian
streamside community. Journal of Herpetology 35:36-44.
Pough F. H. 1980. The advantages of ectothermy for tetrapods. American Naturalist
115:92-112.
Pough, F. H. 1983. Amphibians and reptiles as low energy systems. Pages 141-188 in
W. P. Aspey and S. I. Lustick, editors. Behavioral energetics; the cost of survival
in vertebrates. Ohio State University Press, Columbus, Ohio.
Pulliam, H. R. 1975. Diet optimization with Nutrient constraints. The American
Naturalist 109(970):65-768.

48
Ritchie, E. G. and C. N. Johnson. 2009. Predator interactions, mesopredator release and
biodiversity conservation. Ecological Letters 12:982-998.
Rooney, T. P., C. Antolik, and M. D. Morgan. 2000. The impact of salamander predation
on Collembola abundance. Proceedings of the Entomological Society in
Washington 102:308-312.
SAS version 9.2. 2008. SAS Institute Inc. Cary, North Carolina, USA.
Schlesinger, W. H. 1991. Biogeochemistry, an Analysis of Global Change. New York,
USA, Academic Press.
Singh, U. R. 1977. Relationship between the population density of soil microarthropods
and mycoflaura associated with litter and the total litter respiration on the floor of
a sal forest in Varanasi, India. Ecological Bulletins 25: 463-470.
Spiller, D. A. and T. W. Schoener. 1995. Long-term variation in the effect of lizards on
spider density is linked to rainfall. Oecologia 103:133-139.
Stamps, J., S. Tanaka, and V. V. Krishnan. 1981. The relationship between selectivity
and food abundance in a juvenile lizard. Ecology 62:1079-1092.
Walton, M. B. and S. Steckler. 2005. Contrasting effects of salamanders on forest floor
macro- and mesofauna in laboratory microcosms. Pedobiologia 49:51-60
Walton, M. B. 2005. Salamanders in forest-floor food webs: Environmental
heterogeneity affects the strength of top-down effects. Pedobiologia 49:381-393
Walton, M. B., D. Tsatiris, and M. Rivera-Sostre. 2006. Salamanders in forest-floor food
webs: Invertebrate species composition influences top-down effects.
Pedobiologia 50:313-321
Wardle, D. A. 2002. Communities and ecosystems: linking the above ground and the
below ground components. Princton University Press, Princton, NJ.
Welsh, H. H. and A. J. Lind. 1991. The structure of the herpetofaunal Assemblage in the
Douglas-fir/hardwood forests of the Northwestern California and Southwestern
Oregon. US Department of Agriculture, Forest Service Pacific Northwest
Research Station, Portland, Oregon, general technical report PNW-GTR-285:394-
413.
Welsh, H. H. and A. J. Lind. 1992. Population ecology of two relictual salamanders
from the Klamath Mountains of northwestern California. Pp 419-437, in:
McCulloch, D. R., and R. H. Barrett (Eds.), Wildlife 2001: populations. Elsevier
Science Publishers, London, England.

49
Welsh, H. H. Jr., Hodgson G. R., and Lind A. J. 2005. Ecogeography of the
herpetofauna of a northern California watershed: linking species patterns to
landscape processes. Ecogeography 28:521-536.
Welsh H. H. Jr. 2007. Personal communication. USDA Forest Service, Redwood
Sciences Laboratory, 1700 Bayview Drive, Arcata, CA 95521.
Whittaker, R. H. 1952. A study of summer foliage insect communities in the Great
Smokey Mountains. Ecological Monographs 22:1-44.
Wyman, R. L. 1998. Experimental Assessment of Salamanders as Predators of detrital
food webs: effects on invertebrates, decomposition and the carbon cycle.
Biodiversity and Conservation 7:641-650.
Zhou, G., S. Liu, Z. Li, D. Zhang, X. Tang, C. Zhou, J. Yan, J. Mo. 2006. Old-growth
forests can accumulate carbon in soils. Science 314:1417.
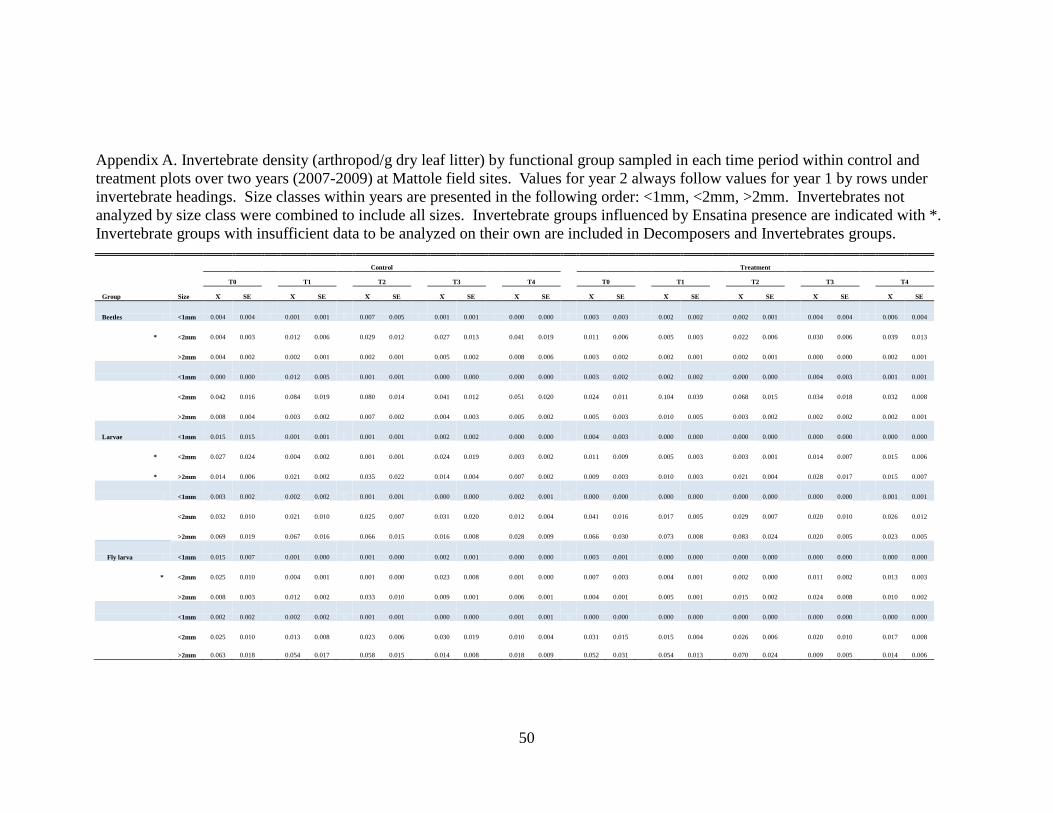
50
Appendix A. Invertebrate density (arthropod/g dry leaf litter) by functional group sampled in each time period within control and
treatment plots over two years (2007-2009) at Mattole field sites. Values for year 2 always follow values for year 1 by rows under
invertebrate headings. Size classes within years are presented in the following order: <1mm, <2mm, >2mm. Invertebrates not
analyzed by size class were combined to include all sizes. Invertebrate groups influenced by Ensatina presence are indicated with *.
Invertebrate groups with insufficient data to be analyzed on their own are included in Decomposers and Invertebrates groups.
Control Treatment
T0
T1
T2
T3
T4
T0
T1
T2
T3
T4
Group Size Χ SE
Χ SE
Χ SE
Χ SE
Χ SE
Χ SE
Χ SE
Χ SE
Χ SE
Χ SE
Beetles <1mm 0.004 0.004 0.001 0.001 0.007 0.005 0.001 0.001 0.000 0.000 0.003 0.003 0.002 0.002 0.002 0.001 0.004 0.004 0.006 0.004
* <2mm 0.004 0.003
0.012 0.006
0.029 0.012
0.027 0.013
0.041 0.019
0.011 0.006
0.005 0.003
0.022 0.006
0.030 0.006
0.039 0.013
>2mm 0.004 0.002
0.002 0.001
0.002 0.001
0.005 0.002
0.008 0.006
0.003 0.002
0.002 0.001
0.002 0.001
0.000 0.000
0.002 0.001
<1mm 0.000 0.000 0.012 0.005 0.001 0.001 0.000 0.000 0.000 0.000 0.003 0.002 0.002 0.002 0.000 0.000 0.004 0.003 0.001 0.001
<2mm 0.042 0.016
0.084 0.019
0.080 0.014
0.041 0.012
0.051 0.020
0.024 0.011
0.104 0.039
0.068 0.015
0.034 0.018
0.032 0.008
>2mm 0.008 0.004
0.003 0.002
0.007 0.002
0.004 0.003
0.005 0.002
0.005 0.003
0.010 0.005
0.003 0.002
0.002 0.002
0.002 0.001
Larvae <1mm 0.015 0.015 0.001 0.001 0.001 0.001 0.002 0.002 0.000 0.000 0.004 0.003 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
* <2mm 0.027 0.024
0.004 0.002
0.001 0.001
0.024 0.019
0.003 0.002
0.011 0.009
0.005 0.003
0.003 0.001
0.014 0.007
0.015 0.006
* >2mm 0.014 0.006
0.021 0.002
0.035 0.022
0.014 0.004
0.007 0.002
0.009 0.003
0.010 0.003
0.021 0.004
0.028 0.017
0.015 0.007
<1mm 0.003 0.002 0.002 0.002 0.001 0.001 0.000 0.000 0.002 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.001 0.001
<2mm 0.032 0.010
0.021 0.010
0.025 0.007
0.031 0.020
0.012 0.004
0.041 0.016
0.017 0.005
0.029 0.007
0.020 0.010
0.026 0.012
>2mm 0.069 0.019
0.067 0.016
0.066 0.015
0.016 0.008
0.028 0.009
0.066 0.030
0.073 0.008
0.083 0.024
0.020 0.005
0.023 0.005
Fly larva <1mm 0.015 0.007 0.001 0.000 0.001 0.000 0.002 0.001 0.000 0.000 0.003 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
* <2mm 0.025 0.010
0.004 0.001
0.001 0.000
0.023 0.008
0.001 0.000
0.007 0.003
0.004 0.001
0.002 0.000
0.011 0.002
0.013 0.003
>2mm 0.008 0.003
0.012 0.002
0.033 0.010
0.009 0.001
0.006 0.001
0.004 0.001
0.005 0.001
0.015 0.002
0.024 0.008
0.010 0.002
<1mm 0.002 0.002 0.002 0.002 0.001 0.001 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
<2mm 0.025 0.010
0.013 0.008
0.023 0.006
0.030 0.019
0.010 0.004
0.031 0.015
0.015 0.004
0.026 0.006
0.020 0.010
0.017 0.008
>2mm 0.063 0.018
0.054 0.017
0.058 0.015
0.014 0.008
0.018 0.009
0.052 0.031
0.054 0.013
0.070 0.024
0.009 0.005
0.014 0.006

51
Control Treatment
T0
T1
T2
T3
T4
T0
T1
T2
T3
T4
Group Size Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE
Beetle Larva <1mm 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
* <2mm 0.002 0.002
0.001 0.001
0.000 0.000
0.001 0.001
0.002 0.001
0.004 0.003
0.001 0.001
0.000 0.000
0.003 0.002
0.002 0.002
* >2mm 0.005 0.001
0.009 0.002
0.002 0.001
0.004 0.002
0.001 0.001
0.006 0.002
0.005 0.002
0.003 0.002
0.004 0.002
0.004 0.003
<1mm 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
* <2mm 0.004 0.002
0.002 0.002
0.001 0.001
0.000 0.000
0.000 0.000
0.011 0.007
0.002 0.002
0.002 0.002
0.000 0.000
0.000 0.000
>2mm 0.005 0.002
0.003 0.002
0.004 0.002
0.000 0.000
0.001 0.001
0.011 0.005
0.012 0.009
0.006 0.004
0.004 0.003
0.003 0.002
Springtails <1mm 0.431 0.296 0.073 0.035 0.073 0.023 0.076 0.045 0.208 0.026 0.473 0.400 0.076 0.028 0.077 0.029 0.162 0.081 0.300 0.082
<2mm 0.099 0.056
0.033 0.006
0.044 0.013
0.057 0.023
0.057 0.010
0.098 0.063
0.034 0.009
0.038 0.011
0.055 0.020
0.103 0.031
>2mm 0.013 0.007
0.005 0.002
0.005 0.003
0.004 0.002
0.002 0.001
0.020 0.013
0.011 0.004
0.008 0.003
0.006 0.004
0.007 0.004
<1mm 0.527 0.121 1.020 0.112 0.578 0.159 0.737 0.369 0.382 0.134 0.466 0.102 0.857 0.301 0.721 0.149 0.509 0.212 0.475 0.066
<2mm 0.252 0.055
0.511 0.071
0.301 0.055
0.188 0.077
0.136 0.054
0.303 0.060
0.437 0.092
0.287 0.068
0.134 0.030
0.165 0.040
>2mm 0.027 0.009
0.038 0.006
0.007 0.002
0.002 0.001
0.000 0.000
0.025 0.008
0.025 0.009
0.013 0.003
0.005 0.003
0.004 0.002
Entomobryidae <1mm 0.011 0.009 0.010 0.007 0.015 0.007 0.012 0.006 0.038 0.015 0.037 0.025 0.019 0.008 0.009 0.003 0.007 0.003 0.030 0.025
* <2mm 0.009 0.008
0.012 0.003
0.012 0.007
0.014 0.003
0.022 0.010
0.019 0.012
0.009 0.002
0.006 0.002
0.020 0.008
0.017 0.009
>2mm 0.006 0.003
0.004 0.002
0.002 0.001
0.002 0.001
0.001 0.001
0.010 0.007
0.006 0.002
0.006 0.003
0.003 0.002
0.003 0.002
<1mm 0.052 0.028 0.106 0.031 0.070 0.015 0.138 0.056 0.122 0.074 0.029 0.011 0.118 0.056 0.081 0.018 0.051 0.016 0.114 0.044
<2mm 0.025 0.010
0.095 0.029
0.087 0.016
0.082 0.044
0.078 0.038
0.028 0.009
0.079 0.029
0.088 0.027
0.064 0.019
0.076 0.026
>2mm 0.013 0.009
0.013 0.005
0.003 0.002
0.002 0.001
0.000 0.000
0.006 0.004
0.014 0.004
0.009 0.002
0.004 0.003
0.004 0.002
Isotomidae <1mm 0.329 0.234 0.058 0.027 0.040 0.015 0.020 0.017 0.057 0.021 0.282 0.244 0.053 0.023 0.059 0.027 0.067 0.044 0.124 0.030
<2mm 0.062 0.033
0.012 0.003
0.016 0.006
0.011 0.010
0.006 0.003
0.025 0.010
0.015 0.006
0.016 0.006
0.012 0.005
0.033 0.009
>2mm 0.004 0.003
0.000 0.000
0.002 0.001
0.001 0.001
0.001 0.001
0.010 0.006
0.003 0.002
0.002 0.001
0.002 0.002
0.004 0.002
<1mm 0.162 0.031 0.661 0.103 0.384 0.128 0.471 0.289 0.120 0.030 0.200 0.065 0.569 0.288 0.465 0.100 0.322 0.151 0.203 0.035
<2mm 0.055 0.007
0.164 0.034
0.108 0.026
0.043 0.023
0.013 0.006
0.065 0.011
0.157 0.043
0.128 0.051
0.024 0.009
0.028 0.009
>2mm 0.008 0.002
0.024 0.006
0.003 0.001
0.000 0.000
0.000 0.000
0.017 0.009
0.007 0.004
0.005 0.003
0.000 0.000
0.000 0.000
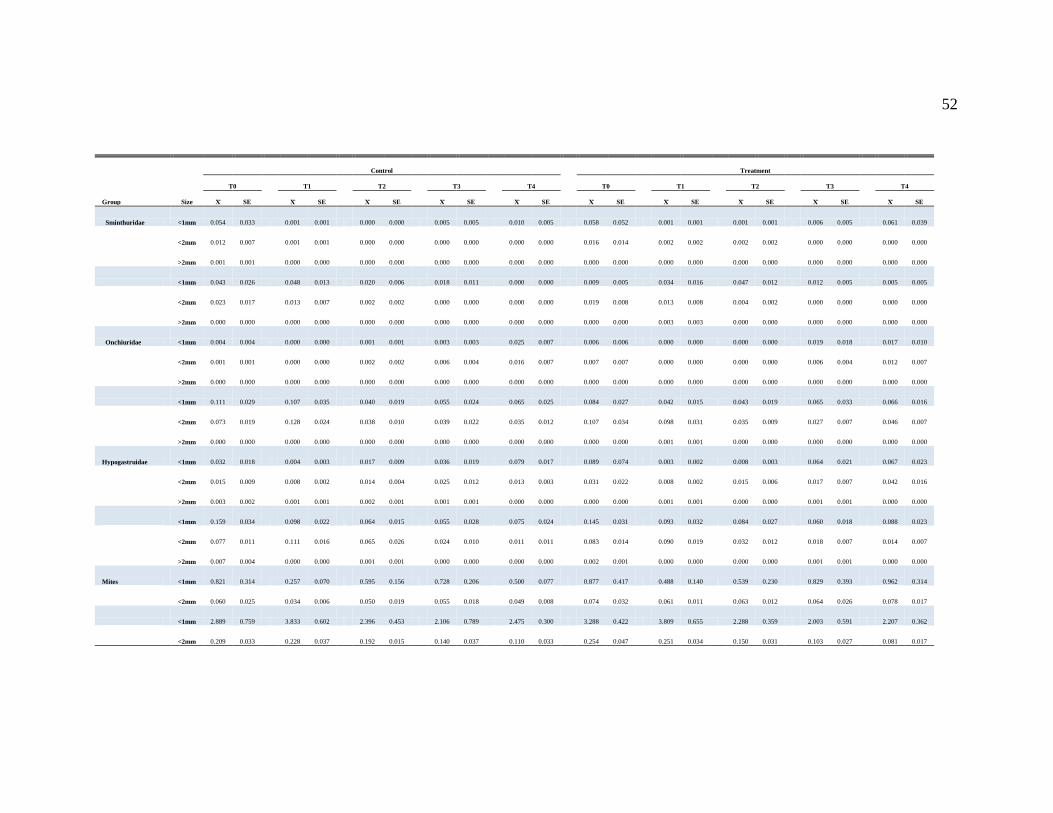
52
Control Treatment
T0
T1
T2
T3
T4
T0
T1
T2
T3
T4
Group Size Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE
Sminthuridae <1mm 0.054 0.033 0.001 0.001 0.000 0.000 0.005 0.005 0.010 0.005 0.058 0.052 0.001 0.001 0.001 0.001 0.006 0.005 0.061 0.039
<2mm 0.012 0.007
0.001 0.001
0.000 0.000
0.000 0.000
0.000 0.000
0.016 0.014
0.002 0.002
0.002 0.002
0.000 0.000
0.000 0.000
>2mm 0.001 0.001
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
<1mm 0.043 0.026 0.048 0.013 0.020 0.006 0.018 0.011 0.000 0.000 0.009 0.005 0.034 0.016 0.047 0.012 0.012 0.005 0.005 0.005
<2mm 0.023 0.017
0.013 0.007
0.002 0.002
0.000 0.000
0.000 0.000
0.019 0.008
0.013 0.008
0.004 0.002
0.000 0.000
0.000 0.000
>2mm 0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.003 0.003
0.000 0.000
0.000 0.000
0.000 0.000
Onchiuridae <1mm 0.004 0.004 0.000 0.000 0.001 0.001 0.003 0.003 0.025 0.007 0.006 0.006 0.000 0.000 0.000 0.000 0.019 0.018 0.017 0.010
<2mm 0.001 0.001
0.000 0.000
0.002 0.002
0.006 0.004
0.016 0.007
0.007 0.007
0.000 0.000
0.000 0.000
0.006 0.004
0.012 0.007
>2mm 0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
<1mm 0.111 0.029 0.107 0.035 0.040 0.019 0.055 0.024 0.065 0.025 0.084 0.027 0.042 0.015 0.043 0.019 0.065 0.033 0.066 0.016
<2mm 0.073 0.019
0.128 0.024
0.038 0.010
0.039 0.022
0.035 0.012
0.107 0.034
0.098 0.031
0.035 0.009
0.027 0.007
0.046 0.007
>2mm 0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.001 0.001
0.000 0.000
0.000 0.000
0.000 0.000
Hypogastruidae <1mm 0.032 0.018 0.004 0.003 0.017 0.009 0.036 0.019 0.079 0.017 0.089 0.074 0.003 0.002 0.008 0.003 0.064 0.021 0.067 0.023
<2mm 0.015 0.009
0.008 0.002
0.014 0.004
0.025 0.012
0.013 0.003
0.031 0.022
0.008 0.002
0.015 0.006
0.017 0.007
0.042 0.016
>2mm 0.003 0.002
0.001 0.001
0.002 0.001
0.001 0.001
0.000 0.000
0.000 0.000
0.001 0.001
0.000 0.000
0.001 0.001
0.000 0.000
<1mm 0.159 0.034 0.098 0.022 0.064 0.015 0.055 0.028 0.075 0.024 0.145 0.031 0.093 0.032 0.084 0.027 0.060 0.018 0.088 0.023
<2mm 0.077 0.011
0.111 0.016
0.065 0.026
0.024 0.010
0.011 0.011
0.083 0.014
0.090 0.019
0.032 0.012
0.018 0.007
0.014 0.007
>2mm 0.007 0.004
0.000 0.000
0.001 0.001
0.000 0.000
0.000 0.000
0.002 0.001
0.000 0.000
0.000 0.000
0.001 0.001
0.000 0.000
Mites <1mm 0.821 0.314 0.257 0.070 0.595 0.156 0.728 0.206 0.500 0.077 0.877 0.417 0.488 0.140 0.539 0.230 0.829 0.393 0.962 0.314
<2mm 0.060 0.025
0.034 0.006
0.050 0.019
0.055 0.018
0.049 0.008
0.074 0.032
0.061 0.011
0.063 0.012
0.064 0.026
0.078 0.017
<1mm 2.889 0.759 3.833 0.602 2.396 0.453 2.106 0.789 2.475 0.300 3.288 0.422 3.809 0.655 2.288 0.359 2.003 0.591 2.207 0.362
<2mm 0.209 0.033
0.228 0.037
0.192 0.015
0.140 0.037
0.110 0.033
0.254 0.047
0.251 0.034
0.150 0.031
0.103 0.027
0.081 0.017

53
Control Treatment
T0
T1
T2
T3
T4
T0
T1
T2
T3
T4
Group Size Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE
Orabatidae * <1mm 0.647 0.256 0.178 0.047 0.363 0.107 0.470 0.133 0.339 0.062 0.693 0.331 0.372 0.115 0.314 0.129 0.442 0.224 0.575 0.166
<2mm 0.046 0.022 0.019 0.002 0.027 0.010 0.036 0.012 0.026 0.002 0.048 0.023 0.043 0.012 0.044 0.009 0.046 0.020 0.050 0.014
<1mm 1.870 0.565 2.277 0.307 1.488 0.273 1.303 0.451 1.518 0.202 2.104 0.304 2.226 0.310 1.246 0.203 1.203 0.328 1.237 0.159
<2mm 0.118 0.024 0.125 0.024 0.093 0.010 0.072 0.021 0.037 0.011 0.144 0.041 0.143 0.023 0.076 0.014 0.059 0.015 0.023 0.005
Non-Orabatidae <1mm 0.174 0.059 0.078 0.024 0.232 0.065 0.258 0.076 0.161 0.017 0.184 0.096 0.116 0.031 0.224 0.105 0.387 0.171 0.387 0.159
<2mm 0.015 0.004
0.015 0.006
0.023 0.010
0.019 0.006
0.023 0.007
0.026 0.009
0.018 0.004
0.018 0.004
0.018 0.006
0.028 0.005
<1mm 1.019 0.206 1.557 0.330 0.908 0.206 0.803 0.347 0.958 0.147 1.184 0.139 1.583 0.372 1.042 0.178 0.799 0.277 0.970 0.206
<2mm 0.091 0.013
0.103 0.025
0.098 0.020
0.068 0.019
0.073 0.023
0.110 0.015
0.108 0.018
0.073 0.018
0.044 0.014
0.058 0.015
Milipedes * All 0.006 0.003 0.001 0.001 0.000 0.000 0.005 0.004 0.000 0.000 0.000 0.000 0.003 0.002 0.002 0.002 0.002 0.001 0.019 0.019
All 0.002 0.002
0.004 0.002
0.000 0.000
0.003 0.002
0.004 0.003
0.002 0.002
0.000 0.000
0.003 0.002
0.003 0.001
0.000 0.000
Worms All 0.002 0.002 0.000 0.000 0.001 0.001 0.001 0.001 0.003 0.002 0.007 0.005 0.002 0.002 0.007 0.004 0.002 0.001 0.003 0.002
* All 0.003 0.002
0.015 0.006
0.010 0.004
0.021 0.009
0.006 0.003
0.013 0.008
0.015 0.005
0.021 0.008
0.025 0.013
0.013 0.002
Diplura All 0.020 0.011 0.001 0.001 0.006 0.003 0.004 0.002 0.005 0.004 0.015 0.009 0.002 0.002 0.005 0.001 0.001 0.001 0.012 0.008
All 0.006 0.002
0.009 0.005
0.011 0.004
0.008 0.006
0.016 0.003
0.015 0.005
0.005 0.003
0.020 0.009
0.014 0.002
0.012 0.005
Protura All 0.066 0.037 0.005 0.003 0.024 0.018 0.012 0.005 0.004 0.002 0.047 0.020 0.016 0.007 0.021 0.010 0.014 0.010 0.004 0.002
All 0.039 0.020
0.013 0.006
0.025 0.014
0.036 0.019
0.019 0.012
0.050 0.019
0.017 0.006
0.015 0.006
0.047 0.038
0.026 0.009
Barklice All 0.001 0.001 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.002 0.001 0.000 0.000 0.000 0.000 0.001 0.001 0.001 0.001
* All 0.003 0.002
0.008 0.003
0.004 0.002
0.021 0.005
0.012 0.007
0.003 0.002
0.008 0.002
0.007 0.004
0.020 0.008
0.037 0.011
Termites All 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
* All 0.000 0.000
0.000 0.000
0.001 0.001
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
Symphyla All 0.005 0.002 0.000 0.000 0.005 0.003 0.001 0.001 0.003 0.001 0.007 0.002 0.004 0.001 0.006 0.004 0.007 0.004 0.003 0.001
All 0.022 0.005
0.002 0.002
0.005 0.002
0.009 0.005
0.007 0.005
0.013 0.006
0.004 0.003
0.019 0.008
0.011 0.006
0.021 0.010
Earwigs * All 0.000 0.000 0.005 0.002 0.003 0.002 0.002 0.002 0.002 0.001 0.001 0.001 0.002 0.001 0.003 0.001 0.001 0.001 0.000 0.000
All 0.003 0.001
0.000 0.000
0.000 0.000
0.002 0.002
0.006 0.003
0.004 0.004
0.002 0.002
0.001 0.001
0.000 0.000
0.003 0.002

54
Control Treatment
T0
T1
T2
T3
T4
T0
T1
T2
T3
T4
Group Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE
Decomposers <1mm 1.324 0.627 0.334 0.102 0.686 0.179 0.787 0.251 0.710 0.062 1.410 0.822 0.574 0.157 0.632 0.259 0.997 0.420 1.270 0.341
<2mm 0.222 0.120 0.087 0.013 0.142 0.048 0.174 0.074 0.155 0.026 0.208 0.095 0.116 0.017 0.135 0.028 0.182 0.064 0.246 0.052
>2mm 0.047 0.018 0.033 0.002 0.051 0.024 0.028 0.007 0.019 0.006 0.043 0.020 0.031 0.004 0.048 0.011 0.039 0.018 0.037 0.013
<1mm 3.435 0.876 4.873 0.675 2.977 0.600 2.880 1.162 2.876 0.407 3.780 0.444 4.672 0.924 3.022 0.500 2.556 0.820 2.709 0.400
<2mm 0.576 0.091 0.870 0.105 0.646 0.064 0.432 0.151 0.343 0.065 0.662 0.084 0.837 0.145 0.575 0.113 0.352 0.085 0.363 0.065
>2mm 0.122 0.027 0.130 0.016 0.090 0.015 0.046 0.019 0.056 0.017 0.130 0.040 0.123 0.002 0.130 0.032 0.050 0.015 0.058 0.014
Predators All 0.040 0.017 0.018 0.002 0.022 0.004 0.030 0.010 0.026 0.010 0.025 0.005 0.025 0.006 0.009 0.003 0.023 0.010 0.024 0.006
All 0.076 0.017
0.092 0.009
0.076 0.009
0.060 0.022
0.082 0.029
0.089 0.019
0.075 0.015
0.055 0.009
0.088 0.029
0.071 0.013
Centipedes All 0.011 0.003 0.007 0.003 0.013 0.002 0.011 0.005 0.009 0.002 0.009 0.004 0.008 0.001 0.004 0.002 0.009 0.004 0.009 0.002
All 0.016 0.003
0.035 0.010
0.033 0.008
0.018 0.006
0.031 0.004
0.032 0.008
0.037 0.011
0.019 0.006
0.031 0.010
0.026 0.007
Pseudoscrpions All 0.007 0.003 0.002 0.002 0.002 0.002 0.006 0.004 0.001 0.001 0.004 0.002 0.006 0.005 0.002 0.001 0.002 0.002 0.004 0.002
* All 0.009 0.005
0.004 0.002
0.008 0.003
0.004 0.002
0.006 0.004
0.006 0.002
0.009 0.002
0.007 0.003
0.005 0.003
0.010 0.003
Predatory larva <1mm 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
<2mm 0.002 0.002
0.000 0.000
0.000 0.000
0.001 0.001
0.000 0.000
0.001 0.001
0.000 0.000
0.000 0.000
0.003 0.002
0.000 0.000
>2mm 0.003 0.002
0.004 0.001
0.003 0.002
0.005 0.002
0.001 0.001
0.003 0.001
0.004 0.002
0.001 0.001
0.002 0.001
0.003 0.002
<1mm 0.000 0.000 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
<2mm 0.007 0.003
0.012 0.004
0.003 0.002
0.005 0.003
0.006 0.004
0.011 0.003
0.002 0.002
0.003 0.002
0.005 0.002
0.010 0.006
>2mm 0.012 0.006
0.012 0.003
0.009 0.005
0.013 0.006
0.006 0.002
0.013 0.006
0.007 0.003
0.005 0.002
0.014 0.009
0.011 0.005
Spiders <1mm 0.008 0.008 0.000 0.000 0.001 0.001 0.002 0.001 0.000 0.000 0.001 0.001 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000
* <2mm 0.002 0.002
0.001 0.001
0.001 0.001
0.000 0.000
0.003 0.001
0.003 0.001
0.001 0.002
0.001 0.001
0.004 0.001
0.004 0.002
>2mm 0.003 0.003
0.002 0.001
0.001 0.001
0.001 0.001
0.004 0.003
0.003 0.002
0.001 0.001
0.000 0.000
0.001 0.001
0.004 0.002
<1mm 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.002 0.002 0.001 0.001 0.000 0.000 0.002 0.002
<2mm 0.019 0.005
0.015 0.004
0.012 0.003
0.012 0.007
0.023 0.014
0.012 0.005
0.008 0.003
0.012 0.001
0.019 0.007
0.005 0.003
>2mm 0.001 0.001
0.013 0.006
0.003 0.001
0.001 0.001
0.004 0.002
0.009 0.005
0.001 0.001
0.002 0.001
0.007 0.002
0.004 0.002
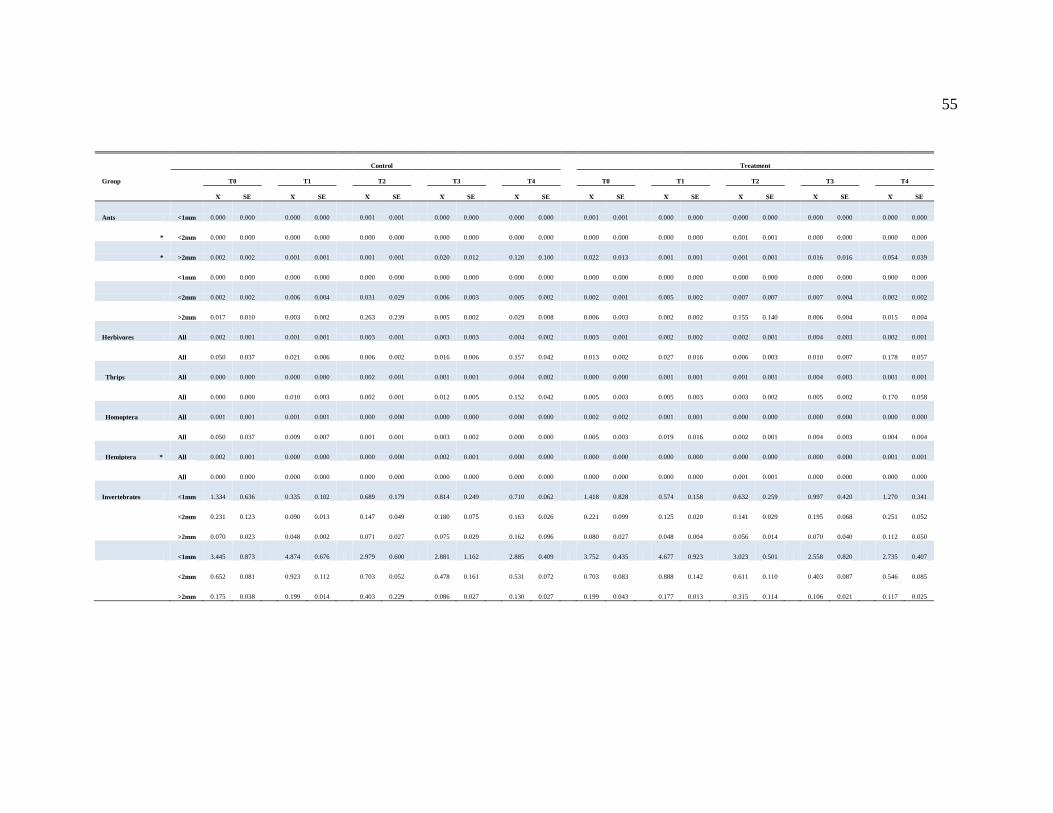
55
Control Treatment
Group
T0
T1
T2
T3
T4
T0
T1
T2
T3
T4
Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE Χ SE
Ants <1mm 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
* <2mm 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000
* >2mm 0.002 0.002 0.001 0.001 0.001 0.001 0.020 0.012 0.120 0.100 0.022 0.013 0.001 0.001 0.001 0.001 0.016 0.016 0.054 0.039
<1mm 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
<2mm 0.002 0.002 0.006 0.004 0.031 0.029 0.006 0.003 0.005 0.002 0.002 0.001 0.005 0.002 0.007 0.007 0.007 0.004 0.002 0.002
>2mm 0.017 0.010 0.003 0.002 0.263 0.239 0.005 0.002 0.029 0.008 0.006 0.003 0.002 0.002 0.155 0.140 0.006 0.004 0.015 0.004
Herbivores All 0.002 0.001 0.001 0.001 0.003 0.001 0.003 0.003 0.004 0.002 0.003 0.001 0.002 0.002 0.002 0.001 0.004 0.003 0.002 0.001
All 0.050 0.037 0.021 0.006 0.006 0.002 0.016 0.006 0.157 0.042 0.013 0.002 0.027 0.016 0.006 0.003 0.010 0.007 0.178 0.057
Thrips All 0.000 0.000 0.000 0.000 0.002 0.001 0.001 0.001 0.004 0.002 0.000 0.000 0.001 0.001 0.001 0.001 0.004 0.003 0.001 0.001
All 0.000 0.000
0.010 0.003
0.002 0.001
0.012 0.005
0.152 0.042
0.005 0.003
0.005 0.003
0.003 0.002
0.005 0.002
0.170 0.058
Homoptera All 0.001 0.001 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.002 0.002 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000
All 0.050 0.037
0.009 0.007
0.001 0.001
0.003 0.002
0.000 0.000
0.005 0.003
0.019 0.016
0.002 0.001
0.004 0.003
0.004 0.004
Hemiptera * All 0.002 0.001 0.000 0.000 0.000 0.000 0.002 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.001 0.001
All 0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.000 0.000
0.001 0.001
0.000 0.000
0.000 0.000
Invertebrates <1mm 1.334 0.636 0.335 0.102 0.689 0.179 0.814 0.249 0.710 0.062 1.418 0.828 0.574 0.158 0.632 0.259 0.997 0.420 1.270 0.341
<2mm 0.231 0.123
0.090 0.013
0.147 0.049
0.180 0.075
0.163 0.026
0.221 0.099
0.125 0.020
0.141 0.029
0.195 0.068
0.251 0.052
>2mm 0.070 0.023
0.048 0.002
0.071 0.027
0.075 0.029
0.162 0.096
0.080 0.027
0.048 0.004
0.056 0.014
0.070 0.040
0.112 0.050
<1mm 3.445 0.873 4.874 0.676 2.979 0.600 2.881 1.162 2.885 0.409 3.752 0.435 4.677 0.923 3.023 0.501 2.558 0.820 2.735 0.407
<2mm 0.652 0.081
0.923 0.112
0.703 0.052
0.478 0.161
0.531 0.072
0.703 0.083
0.888 0.142
0.611 0.110
0.403 0.087
0.546 0.085
>2mm 0.175 0.038 0.199 0.014 0.403 0.229 0.086 0.027 0.130 0.027 0.199 0.043 0.177 0.013 0.315 0.114 0.106 0.021 0.117 0.025


















