
Early Stages in Bacillus subtilis Transformation: Association
10
JOURNAL OF BACTERIOLOGY, Sept. 1978, p. 731-740 0021-9193/78/0135-0731$02.00/0 Copyright © 1978 American Society for Microbiology Vol. 135, No. 3 Printed in U.S.A. Early Stages in Bacillus subtilis Transformation: Association Between Homologous DNA and Surface Structures E. GARCIA, PALOMA LOPEZ, M. TERESA PEREZ URENA, AND M. ESPINOSA* Instiuto de Inmunologia y Biologia Microbiana, Madrid 6, Spain Received for publication 6 June 1978 The addition of ethylenediaminetetraacetate to competent cultures of Bacillus subtilis irreversibly inhibited the transformability as well as the cellular binding of DNA. Our results show that the inhibition of DNA binding by ethylenedia- minetetraacetate in whole cells, protoplasts, and membrane vesicles is mainly due to a permanent alteration of the DNA receptors. Transformation absolutely requires free magnesium ions, whereas DNA binding is a magnesium-independent step. In contrast to ethylenediaminetetraacetate, the absence of Mg2e does not irreversibly affect the capacity of the competent cells to be transformed. DNA- binding receptors located at the cell surface remain associated with the plasma membrane after protoplasting and after isolation of membrane vesicles. A Mg2+- dependent endonucleolytic activity associated with the membrane appears to be responsible for the lower levels of binding by protoplasts in the presence of this ion. In the transformation system of Bacillus sub- tilis, several succesive steps have been demon- strated after the addition of donor DNA to com- petent cultures. The attachment of donor DNA to the outer surface of the recipient cell, where it is still sensitive to added deoxyribonuclease (DNase), is defined as binding (8). Entry occurs when donor DNA penetrates into the recipient cell so that it is no longer susceptible to pan- creatic DNase. It has been suggested that the binding of DNA precedes its exonucleolytic breakdown by competent bacteria (7). Lacks et al. (9) have pointed out that- a nuclease is in- volved in the entry of donor DNA in Diplococ- cus pneumoniae. A similar conclusion has been claimed by Joenje and Venema (7) to occur in B. subtilis. There is evidence indicating that ethylenedi- aminetetraacetate (EDTA) is an inhibitor of genetic transformation both in B. subtilis (12) and in D. pneumoniae (14). In both transfor- mation systems, the DNA attached to EDTA- treated cells is in a DNase-sensitive form (2, 12, 14). Furthermore, in B. subtilis it has also been demonstrated that the activity of the Mg2e-de- pendent competence-specific exonuclease is completely inhibited by the presence of EDTA, but not by ethyleneglycol-bis(aminoethyl ether)-N,N-tetraacetic acid, which also suggests that this nuclease may be involved in the trans- formation process (2, 7). In the pneumococcal system, EDTA considerably increases the bind- ing of exogenous DNA (14). In B. subtilis, Joenje and Venema (7) have shown that EDTA inhibits the binding of donor DNA to an extent of about 40%. However, in a more recent report (2) it has been concluded that because of the absence of breakdown, DNA binding is approximately five times higher in the presence of EDTA than in its absence. The apparent conflicts observed about the role of EDTA in genetic transforma- tion led us to study, in a more detailed way, the effect of this agent on DNA binding and trans- formation. In the present paper, we show that EDTA inhibits the binding of homologous native DNA to competent cells, protoplasts, and membrane vesicles of B. subtilis. We further show that this inhibition is probably due to an alteration of the DNA receptors on the cell surface rather than to the complexing of Mg2e by EDTA. Our results suggest that, in B. subtilis, EDTA is not an appropriate agent to use for the study of the early stages in DNA-mediated transformation because of its inhibitory effect upon DNA bind- ing. In addition, we conclude that DNA binding is a separate step in the transformation system of B. subtilis and one that does not require divalent cations. Magnesium, but not calcium, is required for the processing of reversibly bound DNA. MATERIALS AND METHODS Strains. B. subtilis RUB 300 (lys-3 metBlO hisB2) was the recipient strain and was used as a source of unlabeled competing DNA, kindly provided by F. E. Young. B. subtilis OG1, a prototrophic strain, was used as a source of unlabeled transforming DNA; B. subtilis 1G22 (thy) was employed for the isolation of 731
Transcript of Early Stages in Bacillus subtilis Transformation: Association

JOURNAL OF BACTERIOLOGY, Sept. 1978, p. 731-7400021-9193/78/0135-0731$02.00/0Copyright © 1978 American Society for Microbiology
Vol. 135, No. 3
Printed in U.S.A.
Early Stages in Bacillus subtilis Transformation: AssociationBetween Homologous DNA and Surface StructuresE. GARCIA, PALOMA LOPEZ, M. TERESA PEREZ URENA, AND M. ESPINOSA*
Instiuto de Inmunologia y Biologia Microbiana, Madrid 6, Spain
Received for publication 6 June 1978
The addition of ethylenediaminetetraacetate to competent cultures of Bacillussubtilis irreversibly inhibited the transformability as well as the cellular bindingof DNA. Our results show that the inhibition of DNA binding by ethylenedia-minetetraacetate in whole cells, protoplasts, and membrane vesicles is mainly dueto a permanent alteration of the DNA receptors. Transformation absolutelyrequires free magnesium ions, whereas DNA binding is a magnesium-independentstep. In contrast to ethylenediaminetetraacetate, the absence of Mg2e does notirreversibly affect the capacity of the competent cells to be transformed. DNA-binding receptors located at the cell surface remain associated with the plasmamembrane after protoplasting and after isolation of membrane vesicles. A Mg2+-dependent endonucleolytic activity associated with the membrane appears to beresponsible for the lower levels of binding by protoplasts in the presence of thision.
In the transformation system of Bacillus sub-tilis, several succesive steps have been demon-strated after the addition of donor DNA to com-petent cultures. The attachment of donor DNAto the outer surface of the recipient cell, whereit is still sensitive to added deoxyribonuclease(DNase), is defined as binding (8). Entry occurswhen donor DNA penetrates into the recipientcell so that it is no longer susceptible to pan-creatic DNase. It has been suggested that thebinding of DNA precedes its exonucleolyticbreakdown by competent bacteria (7). Lacks etal. (9) have pointed out that- a nuclease is in-volved in the entry of donor DNA in Diplococ-cus pneumoniae. A similar conclusion has beenclaimed by Joenje and Venema (7) to occur inB. subtilis.There is evidence indicating that ethylenedi-
aminetetraacetate (EDTA) is an inhibitor ofgenetic transformation both in B. subtilis (12)and in D. pneumoniae (14). In both transfor-mation systems, the DNA attached to EDTA-treated cells is in a DNase-sensitive form (2, 12,14). Furthermore, in B. subtilis it has also beendemonstrated that the activity of the Mg2e-de-pendent competence-specific exonuclease iscompletely inhibited by the presence of EDTA,but not by ethyleneglycol-bis(aminoethylether)-N,N-tetraacetic acid, which also suggeststhat this nuclease may be involved in the trans-formation process (2, 7). In the pneumococcalsystem, EDTA considerably increases the bind-ing ofexogenous DNA (14). In B. subtilis, Joenjeand Venema (7) have shown that EDTA inhibits
the binding of donor DNA to an extent of about40%. However, in a more recent report (2) it hasbeen concluded that because of the absence ofbreakdown, DNA binding is approximately fivetimes higher in the presence of EDTA than inits absence. The apparent conflicts observedabout the role of EDTA in genetic transforma-tion led us to study, in a more detailed way, theeffect of this agent on DNA binding and trans-formation.
In the present paper, we show that EDTAinhibits the binding of homologous native DNAto competent cells, protoplasts, and membranevesicles of B. subtilis. We further show that thisinhibition is probably due to an alteration of theDNA receptors on the cell surface rather thanto the complexing of Mg2e by EDTA. Our resultssuggest that, in B. subtilis, EDTA is not anappropriate agent to use for the study of theearly stages in DNA-mediated transformationbecause of its inhibitory effect upon DNA bind-ing. In addition, we conclude that DNA bindingis a separate step in the transformation systemof B. subtilis and one that does not requiredivalent cations. Magnesium, but not calcium, isrequired for the processing of reversibly boundDNA.
MATERIALS AND METHODSStrains. B. subtilis RUB 300 (lys-3 metBlO hisB2)
was the recipient strain and was used as a source ofunlabeled competing DNA, kindly provided by F. E.Young. B. subtilis OG1, a prototrophic strain, wasused as a source of unlabeled transforming DNA; B.subtilis 1G22 (thy) was employed for the isolation of
731

732 GARCIA ET AL.
[3H]DNA. Both strains were kindly supplied by G.Venema.Media and competence development The recip-
ient strain was grown to competence in GM1 medium(17). At 90 min after the culture left the exponentialphase of growth, 5% dimethyl sulfoxide was added,and cells were quickly frozen in alcohol-dry ice andstored at -70°C. Samples were rapidly thawed anddiluted 1:10 either in GM2 medium (17) or in Bott andWilson medium (1) supplemented with the compe-
tence-stimulating amino acids (BWC) according toWilson and Bott (16). In experiments in which Mg2"was omitted or its concentration was changed, freshlythawed cells were centrifuged, washed twice with sa-
line, and suspended either in GM1 or in BWC mediumcontaining the appropriate amount of Mg2e. After-wards, the cultures were incubated at 37°C with vig-orous shaking (300 rpm) for 60 min unless otherwisestated.
Plating was done in minimal agar: 3% agar (Difco)in Bott and Wilson minimal salts (1) plus 1% glucose,the growth requirements (20 yg/ml), and competence-enhancing amino acids (50 ,ig of each per ml). Allmedia were prepared with glass-distilled and demin-eralized water.DNA and transformation procedure. Unlabeled
and 3H-labeled transforming DNA were isolated fromstrains OGI and 1G22, respectively, and competinghomologous DNA was isolated from strain RUB300,following the procedure described by Joenje et al. (6).To minimize radiochemical damage, labeled DNA wasused for no longer than 3 weeks after preparation.Incubation of competent cells with transforming DNA(1 ug/ml) was performed for 45 min unless otherwisestated. Uptake ofDNA was terminated by the additionof DNase I (specific activity, 90 U/gLg) (Worthington)in 0.5 M MgCl2 and 5 min of additional incubation.Transformants were scored for met+.Transformation in the presence of EDTA. Dif-
ferent concentrations of EDTA were added to maxi-mally competent cells. After 5 min of incubation at370C, DNA (1 gig/ml) was added and incubation pro-
ceeded for 20 min more. Mg2e was then added atdifferent concentrations and, at the times indicated inthe results, the reaction was stopped by the additionof DNase I. Controls were done using distilled anddemineralized water instead of EDTA. In some exper-iments, EDTA was removed by centrifugation andresuspension of bacterial cells in fresh medium beforethe addition of Mg2e.DNA binding to competent cells. Competent
cultures were centrifuged and concentrated to giveabout 108 viable cells per ml. [3H]DNA (1 gig/ml) wasadded, and samples were incubated at 370C for 10 minunless otherwise stated. After exposure to DNA,chilled calf thymus DNA (100,ug/ml) was added (1:1,vol/vol), and samples were immediately cooled. Thesuspensions were centrifuged in the cold (12,000 x g,
5 min) and washed twice with cold Mg2e-free medium.Pellets were lysed in the same medium containingDNase I (100 U/ml) plus lysozyme (500 ,ig/ml). Frac-tions of 0.5 ml were counted by adding suitableamounts of scintillation fluid (Instagel, Packard In-struments Co.). Samples were counted in an Intertech-nique scintillation counter. Background counts (be-
tween 20 and 50 cpm) were subtracted from the countsof each assay. Total DNA binding refers to the totalamount of donor DNA associated with the cell (i.e.,binding plus entry).DNA binding to protoplasts. Cells were grown to
competence, centrifuged, and concentrated 10 times(to give 2 x 108 to 3 x 108 viable cells per ml) in thesame medium plus 20% sucrose. Lysozyme (500 ug/ml)was added, and the culture was incubated at 37°C withoccasional shaking.
Protoplasting was monitored by phase-contrast mi-croscopy. When more than 95% of the cells wereconverted to protoplasts (about 20 min), the culturewas centrifuged at 12,000 x g for 10 min, and the pelletwas suspended in prewarmed 20% sucrose-containingGM1 medium (with or without Mg2+). [3H]DNA (1pg/ml) was added, and the suspension was incubatedwith vigorous shaking for 10 min (unless stated oth-erwise). At the end of this time, the same volume ofcold Mg2e-free medium (plus sucrose) was added, theculture was centrifuged immediately (12,000 x g, 10min, 20C) and washed twice, and the pellet was lysedwith water. Samples of 0.5 ml of the lysate werecounted as above.
Sucrose gradients. [3H]DNA (1 yg/ml) was addedto protoplast suspensions and incubated for the timesindicated. At the end of these times, EDTA (pH 7.6;final concentration, 40 mM) was added, samples werecentrifuged immediately at 12,000 x g for 10 min, andthe supernatants were kept on ice.
Neutral sucrose gradients (5 to 20%) were preparedas described (14). A 0.15-ml volume of the superna-tants was layered on the top of the gradient, andcentrifugation was carried out at 40,000 rpm for 105min in an SW50.1 rotor. Fractions (15 drops each)were collected by puncturing the bottom of the tubesand counted as above.
Preparation of the samples and alkaline sucrosegradients (5 to 20%) was performed as described (3). A0.15-ml volume of each sample was layered on the topof the gradient, and centrifugation was carried out at40,000 rpm for 45 min in an SW50.1 rotor. Fractions(15 drops each) were collected and counted as above.Membrane vesicles: isolation and DNA bind-
ing. Membrane vesicles were isolated from maximallycompetent cultures following the procedure describedby Joenje et al. (6), except that EDTA was omitted.Membrane vesicles were washed with 0.1 M potassiumphosphate buffer (pH 7.0) and frozen at -70°C. Pro-tein concentrations were measured as described byLowry et al. (10).[3H]DNA binding to membrane vesicles was per-
formed as previously described (6).
RESULTSEffect of EDTA on transformation and
binding. To determine the pattern of inhibitionof the transformability by EDTA, we testeddifferent amounts of this chemical. Concentra-tions of EDTA as low as 0.1 mM reduced theappearance of transformants about 50% (Fig. 1).Concentrations of EDTA higher than 1 mMdrastically inhibited the transfonnability of thecultures.
J. BACTERIOL.

DNA BINDING IN B. SUBTILIS 733
lo,E
id" I l0 102
Concentration of EDTA(mM)FIG. 1. Effect of different concentrations ofEDTA
on transformability. Frozen ceUs in GMI mediumwere quickly thawed and suspended in GM2 medium.After 60min ofincubation at 37°C, 1-ml samples werewithdrawn, and EDTA was added to the samples.After 5 min of additional incubation, 1 pg of coldDNA per ml was added, and incubation proceeded20 min more. At the end of this time, Mg2+ was addedto a final concentration of 5 mM together with 1,OOOU ofDNase Iper ml, and 5 min later transfornantswere scored in aU samples. No changes in the nunberof viable cells were observed.
Table 1 depicts the effect of different experi-mental conditions on the transformability ofstrain RUB300. As can be observed in experi-ments 1 and 2,EDTA drastically inhibited trans-formation frequency (about 95%) when 100 U ofDNase I per ml were used. The inhibitory effectofEDTA was even more pronounced (more than99%) when the amount ofDNase I was increasedup to 1,000 U/ml (experiment 4). When EDTA-treated cultures loaded with DNA were diluted100-fold into miniinal medium containing DNaseI (1,000 U/mi) plus Mge+ (10 mM), no furtherdecrease in transformation was observed (notshown).Only a partial recovery of transformation (be-
tween 40 and 50% of the control values) was
achieved when Mg2+ was added to culturestreated with 10 mM EDTA, and the incubationproceeded for 10 min more before the additionofDNase I (Table 1, experiments 3 and 4). Whenthe concentration ofEDTA was increased up to15 mM, the inhibitory effect ofEDTA was moremarked, and the recovery of transformants wasonly 2.7% even though the incubation with Mg2+proceeded for 46 mMi before DNase addition(experiment 5). The slight decrease in the recov-
ery of transformation observed when 100 mM
Mg2e was used could be due to the frequentappearance of precipitate in these samples.We investigated the effect of 10 mM EDTA
on the total DNA binding in competent andnoncompetent cultures. In competent cultures,the amount of DNA associated with the cells inthe presence of EDTA varied between 20 and45% of the control values (Table 2), whereasEDTA had no effect on the DNA bound bynoncompetent cultures.Effect of divalent cations. EDTA causes
the release of several membrane-associated en-zymes (e.g. NADH2 dehydrogenase, malic aciddehydrogenase, and adenosinetriphosphatase)and a partial destruction of membranes of sev-eral microorganisms (4). Consequently, the in-hibition ofDNA binding by EDTA could be dueeither to a chelation of divalent cations and/orto an alteration of the DNA receptors located inthe membrane. To distinguish between thesetwo possibilities, we studied the effect ofremovalof Ca2' and Mg2e on the development of trans-formability. A 40-fold reduction in the compe-tence level as compared to the control was ob-served after 60 min of incubation when Mg2ewas omitted from GM1 medium (not shown).Nevertheless, a relatively high level of transfor-mation was still observed, which suggests thatsome traces of Mg2e could be present in somecomponents of the medium (e.g., yeast extract).When we used a more defined medium, such asBWC medium, supplemented with knownamounts of Ca2' and Mg2+, we found that Mg2ewas an absolute requirement for the develop-ment of transformability, whereas Ca2+ showedsome inhibitory effect when added simultane-ously with Mg2e (not shown).To test whether the lack of Mg2e in the me-
dium irreversibly affects the ability of recipientcells to be transformed, competent cells wereincubated in Mg2-free medium with transform-ing DNA (Fig. 2). At different times, sampleswere withdrawn and either Mg2e (curve A) orMg2e plus competing homologous DNA (curveB) was added; the incubation proceeded to reacha total incubation time of 45 min, when all thesamples received DNase. The addition of Mg2ealone to the culture increased the transforma-bility up to control levels when the ion waspresent for only 10 min (curve A). We found asimilar response when the samples receivedMg2e and competing DNA simultaneously ifcells had been incubated with transformingDNA for 25 min before the additions. Transfor-mation was inhibited when Mg2e and competingDNA were added at shorter times of exposure ofcells to transforming DNA (curve B). No signif-icant bansformability was detected when trans-
VOL. 135, 1978

TABLE 1. Effect of different experimental conditions on transformation
xPt EDTA concn OuM) me+ concn Time of DNase Transformation fre- Residual per-(mM) addition (mm)a quencyb centage
1 Control: 0 10 0 0.86 1005 50 0 0.034 3.910 50 0 0.022 2.6
2 Control: 0 10 0 0.83 10010 100 0 0.04 4.810 100 10 0.33 39.7
3 Control: 0 10 0 0.84 10010 50 10 0.46 54.710 75 10 0.46 54.710 100 10 0.35 41.6
4 Control: 0 50 0 0.11 1005 50 0 0.0008 0.7310 50c 0 0.00030c 0.2710 50 0 0.00023 0.20
Control: 0 50 10 0.13 10010 50 0 0.00030 0.2310 50 10 0.063 48.5
5d Control: 0 10 0 0.6 10015 30 0 0.00020 0.03315 30 45 0.010 1.715 50 0 0.00025 0.0415 50 45 0.016 2.7
a Time after addition of Mg2e. Transformation was performed as described in the text; 100 U of DNase perml was used in experiments 1 to 3, and 1,000 U/ml was used in experiments 4 and 5.
b Transformation frequency - (met' transformants per nl/viable cells per ml) x 100.c EDTA was removed by centrifugation before the addition of Mg2e.d Experimental conditions as described by Buitenwerf and Venema (2).
TABLE 2. Effect of 10mMEDTA on total CHIDNAbinding to competent and noncompetent culturesa
Transform- [3H]DNA PercentageExpt tion frequency bound u of DNA
M per 10" bound()cells)
Expt lb-EDTA 0.5 0.010 100
0.010+EDTA 0.005 0.004 45
0.005
Expt 2b-EDTA 1.5 0.023 100
0.020+EDTA 0.03 0.0040 20.2
0.0047
Expt 3C-EDTA 8.2 x 10- 0.0007 <10+EDTA 2 x 10- 0.0010 <10a Specific activity of [3H]DNA, 71,200 cpm/,sg.
Binding was performed as indicated in the text, incu-bating the cultures with or without EDTA (10 mM) inthe presence of [3H]DNA for 10 min.
b Competent cultures, grown in GM2 medium.c Noncompetent cultures, grown in Penassay broth
(antibiotic medium no. 3, Difco).
forming and competing DNAs were added si-multaneously (curve C).To investigate the influence of the absence of
Mg2e on DNA binding, we tested the ability ofrecipient cells to be transformed and to bind[3H]DNA in relation to the amount of Mg2epresent in the medium. A linear response toMg2e concentration was observed in the trans-formation levels reached by the cultures (Fig. 3).However, the amount of[3H]DNA bound by thecells is independent of the concentration of Mg2epresent in the medium.We performed a multiple time point study of
the total binding and of the entry of [3H]DNAunder different conditions. EDTA always in-hibited the total binding as compared with thatobtained in GM1 Mg2e-free medium (Table 3),which demonstrates that EDTA alters the ca-pacity of competent cells to bind donor DNA.The kinetics of total DNA binding in the pres-ence or in the absence of Mg2e are very similar.However, in the absence of Mg2e, about 95% ofthe DNA associated with the cells is removedafter treatment with pancreatic DNase I.Interaction between DNA and mem-
branes. To determine howDNA associates with
734 GARCIA ET AL. J. BACTERIOL.

DNA BINDING IN B. SUBTILIS 735
0~~~~~~~~~~~
@ 1040 10 20 |I~~~~~~~~~~~~
Tim of contact with MOtI+nin)FIG. 2. Transformability of a Mg'+-fi-ee culture
after the addition of compewting homologous DNAand/or Mg2+ (10 mMJ. Cells incubated (as describedin the text) in GM1 without Mg2+ for 60 min wereloaded with transformning DNA (I pg/ml) alone(curves A and B) or with transforming DNA (1 pglml) pluw competing DNA (10 jig/ml) (curve C). At 0, 5,10, 15, 20, 30, 35, 40, 42.5, and 45 min of incubation,0.5-ml samples of each culture uwere removed, andMg2+ (curves A and C) or Mg2+ plus compweting DNA(10 pg/ml) (curve B) was added. The total time ofincubation with transforming DNA was 45 min foraU samples. Consequently, the samples that receivedMg2+ (curvesA and C) or Mg2+ plu8 competing DNA(curve B) at 10 min were in contact with Mg2+ for 35min, etc. At the end of the incubation time (45 min),DNase I (1,000 U/ml) was added, and transformantswere scored in plates without methionine.
the cell membrane in the absence ofme2, weperformned the set of experiments described inTable 4. As can be observed, the addition ofEDTA to competent cells or protoplats resultedin an inhibition of 75% in the amount of DNAbound as compared to the control values,whereas EDTA had no effect on samples fromnoncompetent cultures. When the DNA wasbound to whole competent cells and then theywere converted to protoplasts, there werelosses in the amount of DNA bound of 65 and50% in the presence or in the absence ofme2,respectively. In the presence of Mg2', the nu-
cleases released during protoplasting (7) mayaccount for that difference (50 to 35%) in theamount ofDNA bound. This reduction could beexpected to be greater if lysozyme were notpresent, since this enzyme is known to protectDNA against nucleolytic degradation (7).
In Mg2e-free medium, there was an increasein the amount of DNA bound by protoplasts ascompared to whole cells, which suggests that
4-0
0
EU)
I
or
cwr:0
Concentration of Mg 6(M)FIG. 3. Effect of different concentrations of Mg2+
on DNA binding and transformation. Specific activ-ity of[3HJDNA, 95,000 cpm/pg.
TABLE 3. Kinetics of total binding and entry of[HIDNA in cells growing in GMI medium
Length of 3 TotalA Entry ofMedia et(o DN)Ae bipding (3HIDNA' Percent-Media
o DNA nd'ngb(mig per 10' agec(mn" (Pg per 108 02()0 cells, x102) x1)
GM1 + 5 mM 0 0.08Mg2e 10 1.0 0.10 10.0
15 1.2 0.22 14.730 2.0 0.82 37.345 2.7 1.8 45.0
GM1-Mg2+ 0 0.08310 1.0 0.04 4.015 1.5 0.05 4.230 2.2 0.045 2.245 4.0 0.05 1.8
GM1 + 5 mM 0 0.075Mg2+ + 10 10 0.6 0.04 6.6mM EDTA 15 0.7 0.05 7.1
30 1.3 0.045 3.545 2.2 0.04 1.9
a Binding to competent cells was performed as described inthe text. Entry was determined by the addition of 1,000 U ofDNase I (in 50 mM Mg2+) per ml at the times indicated, andthe incubation proceeded for 15 min. At the end of this time,radioactivities were determined as described in the text.
b Each figure is the average of five experiments.'Percentage - (Entry/total binding) x 100.
VOL. 135, 1978
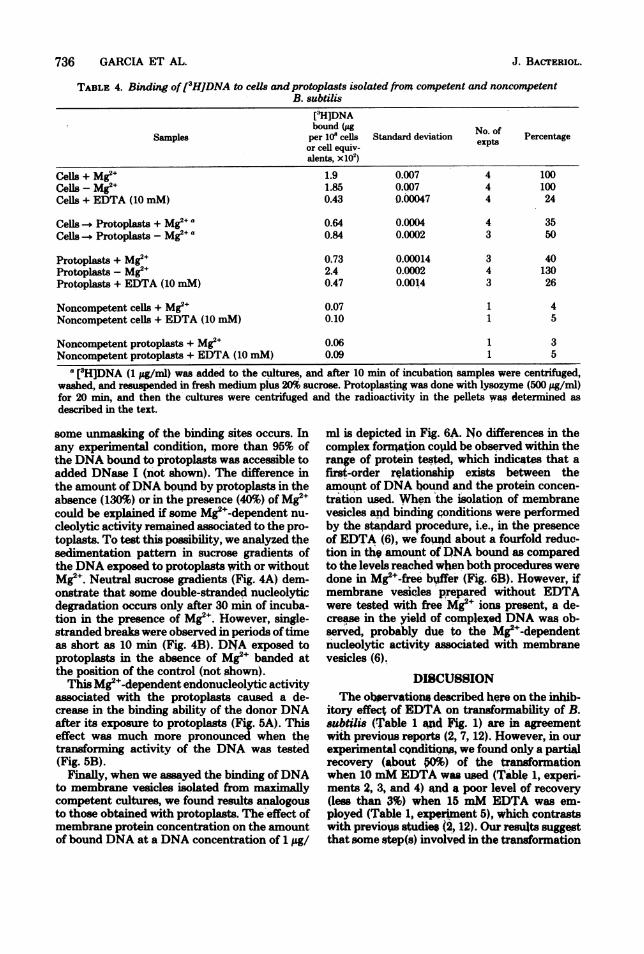
TABLE 4. Binding of[3HJDNA to cells and protoplasts isolated from competent and noncompetentB. subtilis
[3H]DNAbound (ugper 10' cells Standard deviation Neop.f Percentageor cell equiv-alents, xl02)
Cells + Mg2+ 1.9 0.007 4 100Cells - Mg2+ 1.85 0.007 4 100CeUs + EDTA (10 mM) 0.43 0.00047 4 24
Cells Protoplasts + Mg2+a 0.64 0.0004 4 35Cells Protoplats - Mg2+ a 0.84 0.0002 3 50
Protoplasts + Mg2+ 0.73 0.00014 3 40Protoplasts -Mg2+ 2.4 0.0002 4 130Protoplasts + EDTA (10 mM) 0.47 0.0014 3 26
Noncompetent cells + Mg2+ 0.07 1 4Noncompetent cells + EDTA (10 mM) 0.10 1 5
Noncompetent protoplasts + Mg2+ 0.06 1 3Noncompetent protoplasts + EDTA (10 mM) 0.09 1 5
a (3H]DNA (1 jLg/ml) was added to the cultures, and after 10 min of incubation' samples were centrifuged,washed, and resuspended in fresh medium plus 20% sucrose. Protoplasting was done with lysozyme (500 yg/ml)for 20 min, and then the cultures were centrifuged and the radioactivity in the pellets was determined asdescribed in the text.
some unmasking of the binding sites occurs. Inany experimental condition, more than 95% ofthe DNA bound to protoplasts was accessible toadded DNase I (not shown). The difference inthe amount ofDNA bound by protoplasts in theabsence (130%) or in the presence (40%) of Mg2ecould be explained if some Mg2e-dependent nu-
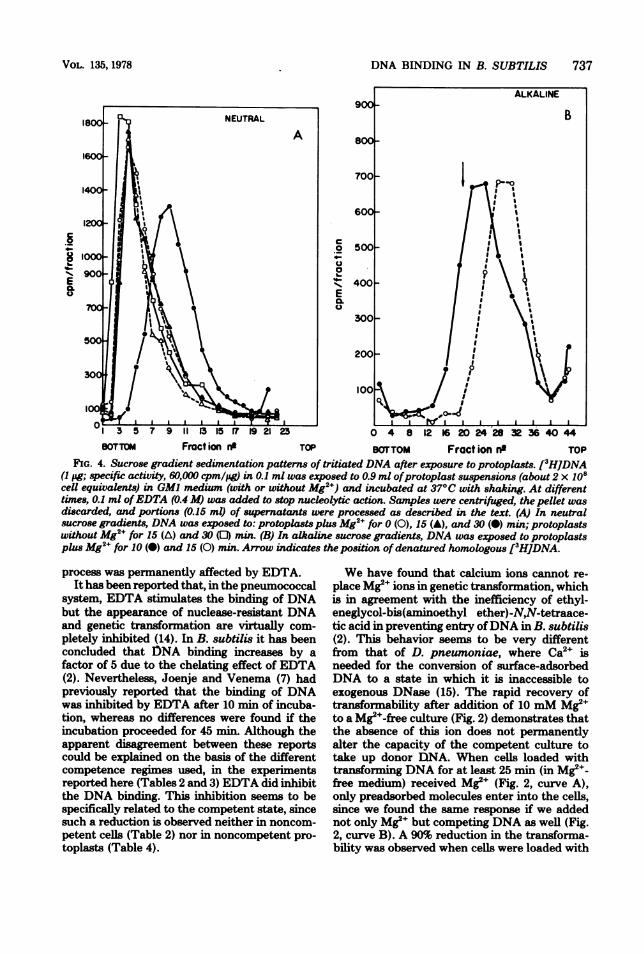
cleolytic activity remained associated to the pro-toplasts. To test this possibility, we analyzed thesedimentation pattern in sucrose gradients ofthe DNA exposed to protoplasts with or withoutMg2e. Neutral sucrose gradients (Fig. 4A) dem-onstrate that some double-stranded nucleolyticdegradation occurs only after 30 min of incuba-tion in the presence of Mg2e. Hiowever, single-
stranded breaks were observed in periods oftimeas short as 10 min (Fig. 4B). DNA exposed toprotoplasts in the absence of Mg2e banded atthe position of the control (not shown).This Mg2+-dependent endonucleolytic activity
associated with the protoplasts caused a de-crease in the binding ability of the donor DNAafter its exposure to protoplasts (Fig. 5A). Thiseffect was much more pronounced when thetransforming activity of the DNA was tested(Fig. 5B).
Finally, when we assayed the binding ofDNAto membrane vesicles isolated from maximally
competent cultures, we found results analogousto those obtained with protoplasts. The effect ofmembrane protein concentration on the amountof bound DNA at a DNA concentration of 1 itg/
ml is depicted in Fig. 6A. No differences in thecomplex forat,ion could be observed within therange of protein tested, which indicates that afirst-order relationship exists between theamount of DNA bound and the protein concen-tration used. ,Wh.en the isolation of membranevesicles and binding ponditions were performedby the standard procedure, i.e., in the presenceof EDTA (6), we found about a fourfold reduc-tion in thp amount of DNA bound as comparedto the levels reached when both procedures weredone in Mg2+-free buffer (Fig. 6B). However, ifmembrane vesiwles prepared without EDTAwere tested with free Mg2+ ions present, a de-cre#ose in the yield of complexed DNA was ob-served, probably due to the Mg2'-dependentnucleolytic activity associated with membranevesicles (6).
DISCUSSIONThe observations described here on the inhib-
itory effect of EDTA on transformability of B.subtilis (Table 1 sond Fig. 1) are in agreementwith previous reports (2, 7, 12). However, in ourexperimental condition, we found only a partialrecovery (about *0%) of the transformationwhen 10 mM EDTA was used (Table 1, experi,ments 2, 3, and 4) and a poor level of recovery(less than 3%) when 15 mM EDTA was em-ployed (Table 1, experiment 5), which contrastswith previous studies (2, 12). Our results suggestthat some step(s) involved in the transformation
736 GARCIA ET AL. J. BACTERIOL.
DNA BINDING IN B. SUBTILIS 737
8
E06
900
800
700
600
C
0
E
500
400
300
200
100
-I 3 5 7 9 11 3 IS 7 9 21 23 0 4 8 12 16 20 24 28 32 36 40 44
BOTTOM Fraction nt TOP BOTTOM Fraction ni TOP
FIG. 4. Sucrose gradient sedimentation patterns of tritiated DNA after exposure to protoplasts. [3HJDNA(1 pg; specific activity, 60,000 cpm/pg) in 0.1 ml was exposed to 0.9 ml ofprotoplast suspensions (about 2 x 108cell equivalents) in GM) medium (with or without Mg2") and incubated at 37°C with shaking. At differenttimes, 0.1 ml ofEDTA (0.4 M) was added to stop nucleolytic action. Samples were centrifuged, the pelkt wasdiscarded, and portions (0.15 ml) of supernatants were processed as described in the text. (A) In neutralsucrose gradients, DNA was exposed to: protoplasts plus Mg2" for 0 (0), 15 (A), and 30 (0) min; protoplastswithout Mg2+ for 15 (A) and 30 (0) min. (B) In alkaline sucrose gradients, DNA was exposed to protoplastsplus Mg2+ for 10 (0) and 15 (0) min. Arrow indicates the position ofdenatured homologous [3HJDNA.
process was permanently affected by EDTA.It has been reported that, in the pneumococcal
system, EDTA stimulates the binding of DNAbut the appearance of nuclease-reistant DNAand genetic transformation are virtually com-pletely inhibited (14). In B. subtilis it has beenconcluded that DNA binding increases by afactor of 5 due to the chelating effect of EDTA(2). Nevertheless, Joenje and Venema (7) hadpreviously reported that the binding of DNAwas inhibited by EDTA after 10 min of incuba-tion, whereas no differences were found if theincubation proceeded for 45 min. Although theapparent disagreement between these reportscould be explained on the basis of the differentcompetence regimes used, in the experimentsreported here (Tables 2 and 3) EDTA did inhibitthe DNA binding. This inhibition seems to bespecifically related to the competent state, sincesuch a reduction is observed neither in noncom-petent cells (Table 2) nor in noncompetent pro-toplasts (Table 4).
We have found that calcium ions cannot re-place Mg2e ions in genetic transformation, whichis in agreement with the inefficiency of ethyl-eneglycol-bis(aminoethyl ether)-N,N-tetraace-tic acid in preventing entry ofDNA in B. subtilis(2). This behavior seems to be very differentfrom that of D. pneumoniae, where Ca2" isneeded for the conversion of surface-adsorbedDNA to a state in which it is inaccessible toexogenous DNase (15). The rapid recovery oftransformability after addition of 10 mM Mg2eto a Mg2e-free culture (Fig. 2) demonstrates thatthe absence of this ion does not permanentlyalter the capacity of the competent culture totake up donor DNA. When cells loaded withtransforming DNA for at least 25 min (in Mg2+-free medium) received Mg2e (Fig. 2, curve A),only preadsorbed molecules enter into the cells,since we found the same response if we addednot only Mg2e but competing DNA as well (Fig.2, curve B). A 90% reduction in the transforma-bility was observed when cells were loaded with
ALKALINE
B
II
a II* I* I
VIIa III II I
I 1~ ~~~~~~
VOL. 135, 1978

738 GARCIA ET AL.
Aoo~~~ ~>D* 2+I00&% 0
%%080 a%%
%%+g0
60-
40
20-
a * v0 5 10 15
TIME (min)
4-._
IEP
0
60[
401
20o
0 5 1 15
TIME (min)FIG. 5. Residual biological activity of [3HJDNA exposed to protoplasts. Protoplasts prepared from a
competent culture of strain RUB300 received 1 pg of[3H]DNA (specific activity, 60,000 cpm/pg) per ml in thepresence or in the absence ofMg2+, and incubation (370C, with shaking) proceeded for the times indicated. Atthese times, sampes (1 ml) were withdrawn and centrifuged in the cold, and the supernatants were collected.The amounts ofDNA present in the supernatants were calculated from their radioactivities. All the samplesreceived the same amount of DNA, and they were used to assay binding (A) and transformation (B) in asecond competent culture of the same stratn, as indicated in the text.
transforming DNA for 5 min and then Mg2" pluscompeting DNA was added (Fig. 2, curve B), ascompared with the values obtained when onlyMg2e was added (curve A). This reduction slowsdown as time goes on. This finding indicates thata reinitiation of the entire process takes place,which seems to start about 20 min after theaddition of Mg2+. This further supports the con-
cept that there is no damage to the DNA recep-tors by the absence of Mg+.
It has been pointed out (7) that the conversionof bound DNA into a DNase I-rosistapt state isa magnesium-dependent step in which some cel-lular exonucleolytic activity is involved. On thebasis of the results presented here (Table 3; Fig.3), we can conclude that Mg2e is not required forDNA binding to competent cells. Furthermore,higher levels of binding were reached in Mg2+-free medium after 45 min of incubation, as com-pared to the results obtained in the presence ofmagnesium or EDTA (Table 3). In the last twoconditions the final amount ofDNA bound wasapproximately the same, which agrees with pre-vious results (7). These findings indicate thatthe DNA receptors are different from the exo-nuclease specifically associated with competentcelLs (7) that would be involved in the processingof transforniing DNA during the entry (i.e., con-version of the cell-bound DNA to a DNase I-
resistant form). Consequently, DNA binding canbe separated as an early step in the transfor-mation process in which DNA molecules attachto the binding sites on the cell surface withoutconsecutive processing. It is interesting that ourexperimental system allows a clear separation ofthis stage without any damage to the competentcells.
In the pneumococcal system, it has been con-cluded that protoplasts do not contain the DNAbinding sites involved in genetic transformationin vivo (13). In B. subtilis, it has been suggestedthat, after protoplasting of competent cultures,no DNA binding sites remained associated withthe membrane (11). However, in this latter re-port the authors did not distinguish betweenbinding and entry of donor DNA. Our resultsshow that in the absence of Mg2e 50% of theDNA bound to the celLs is lost during protoplast-ing (Table 4). This is not due to a release of thesites, since naked protoplasts are able to bindmore DNA than intact competent cells (Table4). The loss of the previously bound DNA islikely due to mechanical forces generated duringprotoplasting. These results demonstrate thatDNA receptors are not lost during protoplastformation and indicate that unmasking of preex-isting DNA binding sites may occur.The low level ofDNA bound to protoplasts in
CX
._
4za
if?:
It4-
ca._
B- , - Mgil\ el
Q
'4~~~~~~~~~I
'4 ~ % WI 1
J. BACTERIOL.
0-%
0- lt%e#"

VOL. 135, 1978
4
en
9
to
DNA BINDING IN B. SUBTILIS 739
10
A5-
5 10 50100 a00,membre protecinc. r (Ig/fmi)
0.01 o0 l 10DNA concentrotion (gg/mi)
FIG. 6. Dose-response relationships of membrane vesicles-[3HJDNA association. (A) Effect of differentamounts ofmembrane vesicles on the complex formation. The reaction mixture (0.5 ml) contained increasingamounts ofproteins in 0.1 M potassium phosphate (pH 7.0) and 0.5 pg of [3HJDNA. (B) Effect of differentamounts ofDNA on the binding assay. The reaction mixture (0.5 ml) contained 25 pg of membrane proteinsin 0.1 Mpotassium phosphate (pH 7.0) and increasing amounts of[3H]DNA. (0, 0) Membrane vesicles wereprepared in the absence of EDTA, and DNA binding was performed in the absence (0) or in the presence(0) of Mg2 . (A) Membrane vesicles were isolated in buffer containing EDTA (15 mM), and binding wasassayed in the presence ofEDTA (final concentration 15 mM). Straight line represents a first-order line witha slope of 450.
the presence of EDTA again supports the hy-pothesis that this chemical alters the DNA re-ceptors.A Mg2e-dependent endonucleolytic activity
remains associated to the protoplast mem-branes. The DNA suffers endonucleolytic cleav-age (Fig. 4) and loses binding capacity and trans-forming activity (Fig. 5). However, the reductionin molecular weight alone does not seem to besufficient to account for the loss of biologicalactivity. The inactivated DNA had completely
lost its capacity to transform competent cultures(Fig. 5B). This phenomenon resembles theDNA-inactivating activity which is associatedwith the noncompetent fraction (B cells) (5).The binding assays to membrane vesicles (Fig.
6), in addition to the experiments with cells andprotoplasts, also demonstrate that the inhibitionofDNA binding by EDTA is due to an alterationof the DNA receptors and not to chelation ofdivalent cations. In our conditions of binding(i.e., in Mg2e-free buffer), we found that mem-

740 GARCIA ET AL.
brane vesicles prepared from competent-cultureshave not lost the DNA receptors. It has beencalculated (13) that, in B. 8ubt1i18, 1010 cel equiv-alents (yielding 0.1 mg of membrane protein)would bind about 1 itg of DNA, at a DNAconcentration of 3 jig/ml. In our experiments,we have found that 0.1 mg ofmembrane proteinscorresponds to 4 x 109 cells, approximately, andthis amount of membrane vesicles binds 0.5 jgof DNA at the above DNA concentration (Fig.6). Thus, 1010 cell equivalents bind 1.25 jig ofDNA, which is in agreement with the calcula-tions of Seto et al. (13). Consequently, we canconclude that DNA receptors are not lost duringpreparation ofmembrane vesicles. The low levelof binding obtained by Joenje and Venema (7)can be reconciled with the calculations of Setoet al. (13) if it is considered that EDTA waspresent during the isolation and DNA bindingof the membrane vesicles in the experimentswith B. subtilis (7). It seems that our experi-mental approach would help to examine thenature of coupling between the donor DNA andthe cell surface in B. subtilis.
ACKNOW]LEDGMENTSWe wish to acknowledge the technical assistance of M. T.
Alda and M. L. del Pozo. We are grateful to R. Lpez for hisinvaluable suggestions and to A. Portoles for his cntical read-ing of the manusript.
LITERATURE CITED1. Bott, K. F., and G. A. Wilson. 1968. Metabolic and
nutritional factors influencing the development of com-petence for trbnfection of Bacilus subtilis. Bacteriol.Rev. 82:370-378.
2. Buitenwerf, J., and G. Venema. 1977. Transformationin BaciUus subtilis. Biological and physical evidencefor a novel DNA-intermediate in synchronously transforming cells. Mol. Gen. Genet. 156:145-155.
3. Dubnau, D., and R. Davidoff-Abelson. 1971. Fate oftraorming DNA following uptake by competent Ba-cilus subtilis. I. Formation and propertie ofthe donor-recipient complex. J. Mol. Biol. 6:209-221.
4. Gel'man, N. S., AL A. Lukoyanova, and D. N. Ostrov-IL. 1975. Bacterial membranes and the respiratory
chain, p. 55-117. In N. S. Gel'man, M. A. Lukoyanova,and D. N. Ostrovskii (ed.), Biomembranes, vol. 6.Plenum Press, New York.
5. Hasltne, F. P., and ML S. Fox 1971. Bacterial inacti-vation of transforming deoxyribonucleate. J. Bacteriol.107:89-89.
6. Joenje, H., W. N. Konsng, and G. Venema. 1974.Interactions between exogenous deoxyribonucleic acidand membrane vesicles isolated from Bacilus subtilu168. J. Bacteriol. 119: 784-794.
7. Joenje, H., and G. Venema. 1975. Different nucleaseactivities in competent and noncompetent Bacillus sub-tilis. J. Bacteriol. 122:2-33.
& Lcks, S. 1977. Binding and entry ofDNA in pneumococ-cal transormation, p. 36-44. In A. Portoles, R L6pez,and M. Espinosa (ed.), Modern trends in bacterial transformation and transfection. North-Holland PublishingCo., Amsterdam.
9. Lacks, S., B. Greenberg, and N. Neuberger. 1974. Roleof a deoxyribonuclease in the genetic transformation ofDiplococcuspneumontae. Proc. Natl. Acad. Sci. U.S.A.71:2305-2309.
10. Lowry, 0. IL, N. J. Rosebrough, A. L Farr, and R. J.Randall. 1961. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:266-275.
11. Mller, L L., C. D. Palmer, and 0. E. Landman 1972.Comparison of deoxyribonucleic acid uptake andmarker integration in bacilli and protoplasts ofBaciUussubtilis. J. Bacteriol. 110:661-666.
12. Morrison, D. A. 1971. Early intermediate state of trans-forming deoxyribonucleic acid during uptake by Bacil-lus subtilis. J. Bacteriol. 108:38-44.
13. Seto, IL, R. L6pez, and A. Toma. 1975. Cell surface-located deoxyribonucleic acid receptors in transform-able pneumococci. J. Bacteriol. 122:1339-1360.
14. Seto, H., and A. Tomas 1974. Early stages in DNAbinding and uptake during genetic transformation ofpneumococci. Proc. Natl. Acad. Sci. U.S.A. 71:1493-1498.
15. Seto, H., and A. Tomsz. 1976. Calcium-requiring stepin the uptake of deoxyribonucleic acid moleculesthrough the surface of competent pneumococci. J. Bac-teriol. 126:1113-1118.
16. Wison, G. A., and K. F. BotL 1968. Nutritional factorsinfluencing the development of competence in the Ba-ciUus subtilis transformation system. J. Bacteriol.96:1439-1449.
17. Young, F. E., and G. A. Wilson. 1974. Bacilus subtilis,p. 69-114. In C. King (ed.), Handbook of genetics, vol.1. Plenum Publishing Corp., New York.
J. BACTERIOL.


















