
DualPathwaysforCopperUptakebyMethanotrophic … ... port process. Cu-Mb from M ... Cu-Mb labeled...
8
Dual Pathways for Copper Uptake by Methanotrophic Bacteria * □ S Received for publication, July 21, 2011, and in revised form, August 31, 2011 Published, JBC Papers in Press, September 7, 2011, DOI 10.1074/jbc.M111.284984 Ramakrishnan Balasubramanian, Grace E. Kenney 1 , and Amy C. Rosenzweig 2 From the Departments of Molecular Biosciences and of Chemistry, Northwestern University, Evanston, Illinois 60208 Background: Methanobactin is a copper-binding molecule produced by methanotrophic bacteria. Results: The copper-loaded and apo (copper-free) forms of methanobactin are taken up by methanotroph cells, and uptake is inhibited by uncoupling agents. Conclusion: Methanobactin plays a key role in copper uptake by methanotrophs. Significance: Copper acquisition via methanobactin has important implications for methanotrophs in the environment. Methanobactin (Mb), a 1217-Da copper chelator produced by the methanotroph Methylosinus trichosporium OB3b, is hypothesized to mediate copper acquisition from the environ- ment, particularly from insoluble copper mineral sources. Although indirect evidence suggests that Mb provides copper for the regulation and activity of methane monooxygenase enzymes, experimental data for direct uptake of copper loaded Mb (Cu-Mb) are lacking. Uptake of intact Cu-Mb by M. tricho- sporium OB3b was demonstrated by isotopic and fluorescent labeling experiments. Confocal microscopy data indicate that Cu-Mb is localized in the cytoplasm. Both Cu-Mb and unche- lated Cu are taken up by M. trichosporium OB3b, but by differ- ent mechanisms. Uptake of unchelated Cu is inhibited by spermine, suggesting a porin-dependent passive transport proc- ess. By contrast, uptake of Cu-Mb is inhibited by the uncoupling agents carbonyl cyanide m-chlorophenylhydrazone and meth- ylamine, but not by spermine, consistent with an active trans- port process. Cu-Mb from M. trichosporium OB3b can also be internalized by other strains of methanotroph, but not by Esch- erichia coli, suggesting that Cu-Mb uptake is specific to metha- notrophic bacteria. These findings are consistent with a key role for Cu-Mb in copper acquisition by methanotrophs and have important implications for further investigation of the copper uptake machinery. Copper is central to the physiology and metabolism of methanotrophic bacteria, methane-oxidizing organisms with potential applications in bioremediation and greenhouse gas removal (1). Methanotrophs have a particularly high require- ment for copper in part because a primary metabolic enzyme, particulate methane monooxygenase (pMMO), 3 is copper-de- pendent (2). pMMO comprises 20% of the total cellular pro- tein (3) and catalyzes the first step in methanotroph metabo- lism, the oxidation of methane to methanol. Under conditions of copper starvation, some methanotrophs produce an alterna- tive, soluble methane monooxygenase (sMMO) that utilizes a diiron active site (4). In these methanotroph strains, copper mediates a switch between MMOs, repressing transcription of the sMMO genes and enhancing pMMO expression and the formation of intracytoplasmic membranes (5, 6). Copper also regulates the expression of a number of other proteins (1). The molecular mechanisms of this copper switch have not been elucidated. Methanotrophs are believed to meet their high requirement for copper by secretion of methanobactin (Mb), a 1217-Da mol- ecule that chelates Cu(I) with high affinity (7–10) and has been investigated recently as a possible treatment for Wilson disease (11). The structure of Mb from Methylosinus trichosporium OB3b includes seven amino acid residues, two oxazolone rings, two neighboring thioamide groups, a 3-methylbutanoyl group, and a pyrollidinyl group (Fig. 1) (10, 12, 13). The peptidic nature of Mb is analogous to the structures of many iron siderophores (14 –16), suggesting that it is a chalkophore (“sidero” is Greek for iron; “chalko” is Greek for copper) (13). Other metha- notrophs, including Methylococcus capsulatus (Bath), Methylo- microbium album BG8, Methylocystis species strain M, and Methylocystis strain SB2, have been reported to produce Mb (17–20), but it remains unclear whether there are different structural classes of Mb. It is not known how Mb is biosynthe- sized, and both nonribosomal and ribosomal peptide synthesis pathways have been suggested (1, 12, 18, 21). It has been proposed that Mb is secreted in its apo form (apo-Mb) to retrieve copper from the environment and then recognized and reinternalized in its copper-loaded form (Cu- Mb) (1, 8). In support of this functional model, addition of Cu-Mb can initiate the switch between sMMO and pMMO expression (10, 22). Notably, although both CuCl 2 and Cu-Mb * This work was supported by National Science Foundation Grant MCB0842366. Imaging work was performed at the Northwestern Univer- sity Cell Imaging Facility supported by National Institutes of Health Grant CCSG P30 CA060553 through the NCI, awarded to the Robert H. Lurie Com- prehensive Cancer Center. □ S The on-line version of this article (available at http://www.jbc.org) contains supplemental Tables S1–S5 and Figs. S1–S6. 1 Supported in part by National Institutes of Health Training Grant GM08061. 2 To whom correspondence should be addressed. Tel.: 847-467-5301; Fax: 847-467-6489; E-mail: [email protected]. 3 The abbreviations used are: pMMO, particulate methane monooxygenase; sMMO, soluble methane monooxygenase; Mb, methanobactin; apo-Mb, metal-free form of Mb; Cu-Mb, copper-bound form of Mb; mBBr, monobro- mobimane; mBBr-Cu-Mb, Cu-Mb labeled with mBBr; mBBr-Mb, apo-Mb labeled with mBBr; CCCP, carbonyl cyanide m-chlorophenylhydrazone; TCEP, tris(2-carboxyethyl)phosphine; TBDT, TonB-dependent transporter; FCCP, carbonyl cyanide p-trifluoromethoxyphenylhydrazone. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 43, pp. 37313–37319, October 28, 2011 © 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. OCTOBER 28, 2011 • VOLUME 286 • NUMBER 43 JOURNAL OF BIOLOGICAL CHEMISTRY 37313 by guest on June 3, 2018 http://www.jbc.org/ Downloaded from
-
Upload
truonghanh -
Category
Documents
-
view
218 -
download
3
Transcript of DualPathwaysforCopperUptakebyMethanotrophic … ... port process. Cu-Mb from M ... Cu-Mb labeled...

Dual Pathways for Copper Uptake by MethanotrophicBacteria*□S
Received for publication, July 21, 2011, and in revised form, August 31, 2011 Published, JBC Papers in Press, September 7, 2011, DOI 10.1074/jbc.M111.284984
Ramakrishnan Balasubramanian, Grace E. Kenney1, and Amy C. Rosenzweig2
From the Departments of Molecular Biosciences and of Chemistry, Northwestern University, Evanston, Illinois 60208
Background:Methanobactin is a copper-binding molecule produced by methanotrophic bacteria.Results: The copper-loaded and apo (copper-free) forms of methanobactin are taken up by methanotroph cells, and uptake isinhibited by uncoupling agents.Conclusion:Methanobactin plays a key role in copper uptake by methanotrophs.Significance: Copper acquisition via methanobactin has important implications for methanotrophs in the environment.
Methanobactin (Mb), a 1217-Da copper chelator produced bythe methanotroph Methylosinus trichosporium OB3b, ishypothesized to mediate copper acquisition from the environ-ment, particularly from insoluble copper mineral sources.Although indirect evidence suggests that Mb provides copperfor the regulation and activity of methane monooxygenaseenzymes, experimental data for direct uptake of copper loadedMb (Cu-Mb) are lacking. Uptake of intact Cu-Mb byM. tricho-sporium OB3b was demonstrated by isotopic and fluorescentlabeling experiments. Confocal microscopy data indicate thatCu-Mb is localized in the cytoplasm. Both Cu-Mb and unche-lated Cu are taken up byM. trichosporiumOB3b, but by differ-ent mechanisms. Uptake of unchelated Cu is inhibited byspermine, suggesting a porin-dependent passive transport proc-ess. By contrast, uptake of Cu-Mb is inhibited by the uncouplingagents carbonyl cyanide m-chlorophenylhydrazone and meth-ylamine, but not by spermine, consistent with an active trans-port process. Cu-Mb from M. trichosporium OB3b can also beinternalized by other strains of methanotroph, but not by Esch-erichia coli, suggesting that Cu-Mb uptake is specific to metha-notrophic bacteria. These findings are consistent with a key rolefor Cu-Mb in copper acquisition by methanotrophs and haveimportant implications for further investigation of the copperuptake machinery.
Copper is central to the physiology and metabolism ofmethanotrophic bacteria, methane-oxidizing organisms withpotential applications in bioremediation and greenhouse gasremoval (1). Methanotrophs have a particularly high require-ment for copper in part because a primary metabolic enzyme,particulate methane monooxygenase (pMMO),3 is copper-de-
pendent (2). pMMO comprises �20% of the total cellular pro-tein (3) and catalyzes the first step in methanotroph metabo-lism, the oxidation of methane to methanol. Under conditionsof copper starvation, somemethanotrophs produce an alterna-tive, soluble methane monooxygenase (sMMO) that utilizes adiiron active site (4). In these methanotroph strains, coppermediates a switch between MMOs, repressing transcription ofthe sMMO genes and enhancing pMMO expression and theformation of intracytoplasmic membranes (5, 6). Copper alsoregulates the expression of a number of other proteins (1). Themolecular mechanisms of this copper switch have not beenelucidated.Methanotrophs are believed to meet their high requirement
for copper by secretion ofmethanobactin (Mb), a 1217-Damol-ecule that chelates Cu(I) with high affinity (7–10) and has beeninvestigated recently as a possible treatment forWilson disease(11). The structure of Mb from Methylosinus trichosporiumOB3b includes seven amino acid residues, two oxazolone rings,two neighboring thioamide groups, a 3-methylbutanoyl group,and a pyrollidinyl group (Fig. 1) (10, 12, 13). The peptidic natureof Mb is analogous to the structures of many iron siderophores(14–16), suggesting that it is a chalkophore (“sidero” is Greekfor iron; “chalko” is Greek for copper) (13). Other metha-notrophs, includingMethylococcus capsulatus (Bath),Methylo-microbium album BG8, Methylocystis species strain M, andMethylocystis strain SB2, have been reported to produce Mb(17–20), but it remains unclear whether there are differentstructural classes of Mb. It is not known howMb is biosynthe-sized, and both nonribosomal and ribosomal peptide synthesispathways have been suggested (1, 12, 18, 21).It has been proposed that Mb is secreted in its apo form
(apo-Mb) to retrieve copper from the environment and thenrecognized and reinternalized in its copper-loaded form (Cu-Mb) (1, 8). In support of this functional model, addition ofCu-Mb can initiate the switch between sMMO and pMMOexpression (10, 22). Notably, although both CuCl2 and Cu-Mb
* This work was supported by National Science Foundation GrantMCB0842366. Imaging work was performed at the Northwestern Univer-sity Cell Imaging Facility supported by National Institutes of Health GrantCCSG P30 CA060553 through the NCI, awarded to the Robert H. Lurie Com-prehensive Cancer Center.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Tables S1–S5 and Figs. S1–S6.
1 Supported in part by National Institutes of Health Training Grant GM08061.2 To whom correspondence should be addressed. Tel.: 847-467-5301; Fax:
847-467-6489; E-mail: [email protected] The abbreviations used are: pMMO, particulate methane monooxygenase;
sMMO, soluble methane monooxygenase; Mb, methanobactin; apo-Mb,metal-free form of Mb; Cu-Mb, copper-bound form of Mb; mBBr, monobro-mobimane; mBBr-Cu-Mb, Cu-Mb labeled with mBBr; mBBr-Mb, apo-Mblabeled with mBBr; CCCP, carbonyl cyanide m-chlorophenylhydrazone;TCEP, tris(2-carboxyethyl)phosphine; TBDT, TonB-dependent transporter;FCCP, carbonyl cyanide p-trifluoromethoxyphenylhydrazone.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 43, pp. 37313–37319, October 28, 2011© 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
OCTOBER 28, 2011 • VOLUME 286 • NUMBER 43 JOURNAL OF BIOLOGICAL CHEMISTRY 37313
by guest on June 3, 2018http://w
ww
.jbc.org/D
ownloaded from

have this effect, addition of Cu-Mb can also promote the switchto pMMOwhen cells only have access to insoluble coppermin-eral sources (22).Mb-mediated release of copper from glass hasbeen correlatedwithmethane oxidation activity aswell (23, 24).However, experimental evidence for the underlying tenet ofthis model, the uptake of Mb by methanotrophs, is lacking. Toaddress this fundamental question, we have tracked copper andMb uptake by isotopic and fluorescent labeling studies. Thesedata provide direct evidence for Mb uptake by methanotrophs.Using inhibitors, we have also demonstrated thatMb transportis not mediated by porins, but by an active transport process.These findings provide a solid framework for further investiga-tion of the Mb handling machinery.
EXPERIMENTAL PROCEDURES
Growth ofM. trichosporiumOB3b—Cultures ofM. trichospo-rium OB3b were grown either in 250-ml Erlenmeyer flasks fit-ted with gas-tight rubber septa or in a 1.25-liter BioFlo 3000bioreactor. Cells were grown at 30 °C with a constant agitationrate of 200–300 rpm. For the flasks, 50ml of air was withdrawnand replaced with 50ml of methane each day. For cells growingin the bioreactor, a 3:1 methane:air mixture was supplied con-stantly at a flow rate of �2 liters/min. Cells were harvested inmid-exponential phase at an absorbance at 600 nm (A600) of0.5–0.9 for flask growths and 2.0–4.0 for bioreactor growths.The components of the growth media for normal and copper-starved conditions were the same as described previously (25,26).Isolation of Mb—M. trichosporium OB3b was grown under
limiting copper concentrations as described previously (26).The spent mediumwas harvested by centrifugation at 9000� gfor 30 min to remove the cells. Purification of apo-Mb wasperformed as described previously (26). Cu-Mb was preparedby an overnight incubation with Cu(II), provided as 3 g ofCuSO4 � 5H2O/liter of spent medium. Purification was per-formed using a DSC-18 HPLC columnwithmethanol or aceto-nitrile as the elution solvent. Fractions containing Cu-Mb werelyophilized and stored at�80 °C until further use. Further puri-fication of Cu-Mb was performed using a Bio-Sil SEC 125 size
exclusion HPLC column. For isotopic labeling, 24mg of 65CuO(Cambridge Isotopes) was dissolved in 1.2 ml of 0.5 N H2SO4and heated at 80 °C for 1 h to yield 65CuSO4 at a final concen-tration of 0.25 M. The resulting 65CuSO4 solution was added toapo-Mb, incubated for 1 h at room temperature in the dark, andthe 65Cu-Mb was then purified using a Sep-Pak Plus tC18 col-umn (Waters).Mass Spectrometry—MALDI-TOF MS was performed on
Cu-Mb and 65Cu-Mb using a Bruker Autoflex III smart beamMALDI-TOF mass spectrometer in the negative ion mode(supplemental Fig. S1). One �l of 20 mM p-nitroaniline wasadded to 1 �l of Mb (�1 �M, concentration measured by theA280 using an extinction coefficient of 1.65 � 104 M�1 cm�1)and spotted on aMALDI target plate. A laser power of 30–50%was used for excitation of the sample with detection in thereflector mode.Fluorescent Labeling of Mb—Cu-Mb was incubated with a
10-fold molar excess of TCEP at room temperature for 1 h.Upon TCEP addition, the Cu-Mb color changed from brown toyellow. Reduced Cu-Mb was purified on a Sep-Pak Plus tC18column using 60% methanol as the elution solvent. The elutedsamples were incubated with a 5-fold molar excess of eithermonobromobimane (mBBr) or bodipy FL L-cysteine (Invitro-gen) in 50 mM Tris, pH 8.0, overnight at room temperature.Excess label was removed by purification using a Sep-Pak PlustC18 column, and thiol-specific labeling was confirmed byMALDI-TOF MS.Mb Uptake Experiments—Uptake experiments were per-
formed on cells grown to mid-exponential growth. In a typicalexperiment, 1 ml of cells was harvested at an A600 of 1.0 bycentrifugation at 18,000 � g for 5 min, washed twice in growthmedium containing only salts and phosphate buffer (salt solu-tion contains 0.85 g/liter NaNO3, 0.17 g/liter K2SO4, 0.037 g/li-ter MgSO4 � 7H2O, and 0.01 g/liter CaCl2 � 2H2O in 1 liter ofwater, and phosphate buffer contains 48.06 g/liter Na2HPO4 �7H2O and 23.4 g/liter KH2PO4 in 1 liter of water). No tracemetals or additional copper and iron were added to this washbuffer. The twice-washed, pelleted cells were resuspended in 1ml of the wash buffer. Cu-Mbwas then added to a final concen-tration of 5 �M, and the mixture was incubated in the darkeither in a culture tube with a rubber stopper or in a 1.5-mlEppendorf tube. Incubation times depended on the experimentand ranged from 40 min to 16 h.Inhibition of Cu and Cu-Mb uptake was performed by the
addition of 3% methylamine (Sigma-Aldrich) or 0.2–0.5 mM
carbonyl cyanidem-chlorophenylhydrazone (CCCP; Enzo LifeSciences) or 0.25–1 mM spermine (Alfa Aesar) to the cells. Fol-lowing a 10–15 min incubation, cells were either analyzeddirectly or washed at least three times in wash buffer followingthe addition of 5�MCuSO4 or 5�MCu-Mb after incubation for45 min prior to analysis. The effective concentrations of meth-ylamine and CCCP were determined using an ethidium bro-mide accumulation assay. Cells were harvested during mid-ex-ponential growth, washed twice in 20 mM phosphate, pH 7.2,and resuspended in the same buffer to an A600 of �5.0.Ethidium bromide was added to a final concentration of 100�g/ml, and the cells were incubated for 60 min at room tem-perature. Published protocols (27–29) were adapted for use in a
FIGURE 1. Schematic diagram of the M. trichosporium OB3b Cu-Mbstructure.
Uptake of Methanobactin
37314 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 43 • OCTOBER 28, 2011
by guest on June 3, 2018http://w
ww
.jbc.org/D
ownloaded from

96-well plate reader. To each well, 150 �l of 100 mM phosphatebuffer, pH 7.2, and 500–1 mM inhibitors were added. Theethidium bromide-treated cells (20 ml) were then added. Fluo-rescence of this mixture was either monitored continuously(emission at 600 nm after excitation at 388 nm) for 8 h, or thestarting and final spectra in the 420–700 nm range (excitationat 388 nm) were collected for 10-min incubations with inhibi-tors. Normalization was performed against fluorescence ofethidium bromide-treated cells with no added inhibitors.Estimation of Metal Content in Whole Cells—After incuba-
tion with Cu-Mb or CuSO4,M. trichosporiumOB3b cells werewashed three times with wash buffer, and 100 �l of 60% tracemetal grade nitric acid was added. The cells were then incu-bated in a heating water bath for 1 h at 80 °C. After 1 h, theEppendorf (microcentrifuge) tubes were centrifuged, and met-al-free water was used to dilute the sample such that the finalconcentration of the nitric acid was 5%. Metal content wasmeasured using a Varian Vista-MPX CCD Simultaneous ICP-OES instrument and copper and iron atomic absorption stand-ards (Sigma-Aldrich) prepared in 5% nitric acid. To determinethe isotopic ratios of 63Cu to 65Cu, ICP-MS was performed onacid hydrolyzed samples using an X Series 2 ICP-MS (ThermoFisher Scientific). Indium was added to a final concentration of20 ppb as an internal mass standard. Because of the variabilityassociatedwith the estimation of themetal content and cell lossduring the wash procedures, the determined values for Cu con-tent were normalized against intracellular Fe content.Competition for the Uptake of Apo-Mb and Cu-Mb—Com-
petition experiments inM. trichosporiumOB3b cells were per-formed by incubating copper-starved cells at anA600 of 1.0 with5�MCu-Mb and varying concentrations (0–10�M) of apo-Mb.The mixtures were incubated for 45 min at room temperature,washed three times in wash buffer, and the copper and ironcontents were measured as described above.Confocal and Fluorescence Microscopy—Copper-starved
cells were washed three times in wash buffer and resuspendedto a final A600 of 1.0. After incubation with 28.1–140.6 �M
(final) mBBr-Mb ormBBr-Cu-Mb for 1 h at room temperature,cells were mounted on a poly-L-lysine-coated glass-bottomPetri dish with FM 4-64 at 5–15 �g/ml (Invitrogen) and 10%DABCO anti-fading solution (Sigma-Aldrich). Live cell imag-ing was performed using a Zeiss LSM510 META systemequipped with a 100�/1.46 Plan-Apochromat oil-immersionTIRF objective. A 25-mW laser diode was used to excitemBBr-Mb at 405 nm, and a 1-mW HeNe laser was used toexcite FM 4-64 at 543 nm. For experiments involving Mblabeled with bodipy FL L-cysteine, a 488-nm argon laser wasused. Images were collected using Zeiss LSM 5 software, andfurther analysis was performed using Volocity (PerkinElmerLife Sciences). Images produced using the 405-nm laser wereslightly offset along the x, y, and z axes compared with imagesproduced using the 543-nm laser. For image merging and fur-ther data analysis, registration correction along the x and y axeswas performed using Volocity. Conventional fluorescencemicroscopy was performed using a Zeiss fluorescence micro-scope equipped with a halogen lamp and band pass filters. Asample preparation procedure similar to that described above,with the exception of poly-L-lysine glass-bottom Petri dishes,
was used for all experiments. A 100� oil immersion objectivewas used for visualization. Imageswere recorded and processedusing the AxioVision software.
RESULTS AND DISCUSSION
WhenM. trichosporiumOB3b cells are presented with 5 �M
copper as unchelated copper (Cu, supplied as CuSO4) orCu-Mb and incubated for 45 min at room temperature, theintracellular copper content, compared with the normalizedcopper content of cells with no added Cu or Cu-Mb, increasesby �2-fold (Fig. 2A and supplemental Table S1). These datashow that both unchelated Cu and Cu associated with Mb canbe taken up by the cells. To track the added Cu, we performeduptake studies using 65Cu and 65Cu-Mb. Natural copper con-tains 69% 63Cu and 31% 65Cu (63Cu/65Cu ratio of 2.2), althoughfractionation of copper isotopes in biological systems is notwellunderstood (30). The observed 63Cu/65Cu ratio of cells with noadded Cu or Cu-Mb (control in Fig. 2B) is �2.0. This deviationis likely due to biological and instrumental fractionation as well
FIGURE 2. Uptake of Cu and Cu-Mb by M. trichosporium OB3b cells. A,increase in intracellular copper content upon addition of Cu and Cu-Mb. B,effect of 65Cu and 65Cu-Mb addition on the intracellular 63Cu/65Cu copperisotope ratio. The control corresponds to the normalized copper content orintracellular 63Cu/65Cu copper isotope ratio of cells with no added Cu orCu-Mb. The error bars represent averages � S.D. of three independent exper-iments. Each single experiment is an average of triplicates.
Uptake of Methanobactin
OCTOBER 28, 2011 • VOLUME 286 • NUMBER 43 JOURNAL OF BIOLOGICAL CHEMISTRY 37315
by guest on June 3, 2018http://w
ww
.jbc.org/D
ownloaded from

as the relatively poor analytical precision of ICP-MS (31).Replacing the natural isotopic copper mixture with 65Cu doesnot affect the ability of cells to take up Cu or Cu-Mb (Fig. 2Aand supplemental Table S1), but the intracellular 63Cu/65Curatio decreases significantly to �0.5 and �0.7 for 65Cu and65Cu-Mb (Fig. 2B). Given that the affinity of Mb for Cu(I) is(6–7)� 1020 M�1 (10), it is likely that when 65Cu is presented as65Cu-Mb, the intact 65Cu-Mb molecule is internalized ratherthan 65Cu-dissociating and then entering the cell. Addition ofeither 65Cu and Cu-Mb or Cu and 65Cu-Mb in combinationleads to an increase in intracellular copper content (Fig. 2A andsupplemental Table S1) and a reduction in the intracellular63Cu/65Cu ratio to�1.0 (Fig. 2B). This value is in between thoseobtained for natural Cu and fully labeled 65Cu, indicating thatnatural Cu and 65Cu are taken up simultaneously, consistentwith uptake of both Cu and Cu-Mb.To determine whether Cu-Mb enters the cell as an intact
complex or whether Cu dissociates from Cu-Mb before enter-ing the cell, Cu-Mb was fluorescently labeled with the thiol-specific probe mBBr. This probe is only fluorescent when con-jugated with thiols. Reduction of the Mb disulfide bondfollowed by overnight incubationwithmBBr yielded a predom-inantly doubly labeled species (mBBr-Cu-Mb)with amolecularmass of 1599.23Da (predictedmass, 1599.27Da) (supplementalFig. S2). Incubation of cells with mBBr-Cu-Mb leads to anincrease in intracellular copper content comparable with whatis observedwithCu-Mb, indicating that labeling does not inter-ferewithCu-Mbuptake (supplemental Fig. S3). Cells incubatedwith mBBr-Cu-Mb were examined using both confocal andconventional fluorescencemicroscopy. Confocal images clearlyshow thatmBBr-Cu-Mb is inside the cell (Fig. 3A). To probe thelocalization of mBBr-Cu-Mb, the membrane-specific fluores-cent dye FM 4-64 (32–34) was used. The merged images sug-gest that mBBr-Cu-Mb is localized primarily in the cytoplasm.The intrinsic fluorescence of Cu-Mb (35, 36) was also detecta-ble by confocal microscopy (supplemental Fig. S4), confirming
that copper uptake mediated by Mb occurs via import of theintactCu-Mbcomplex.Notably, Cu-Mb labeledwith bodipy FLL-cysteine appears to localize at the membrane (supplementalFig. S4), suggesting that the larger size or chemical properties ofthis label may preclude uptake. Uptake of apo-Mb labeled withmBBr was also investigated, and fluorescence is detected in thecytoplasm (Fig. 3B). Although the copper-starved cells used forthese experiments produce their own apo-Mb, the increase insignal intensity for cells incubated with apomBBr-Mb suggeststhat the added Mb is taken up into the cell.To assess the specificity of Mb uptake, uptake experiments
were also performed using Escherichia coli and several otherstrains of methanotroph, including Methylocystis sp. strain M,M. capsulatus (Bath), andMethylomicrobiumalcaliphilum 20Z(supplemental Fig. S5). For all three methanotroph strains,uptake was detected by an increase in intracellular copper con-tent (normalized to the copper content of cells with no addi-tions). It would not be surprising if Methylocystis sp. strain Mcould internalize Cu-Mb purified fromM. trichosporiumOB3bbecause it appears to produce a Mb of identical mass (20). Thechemical composition ofM. capsulatus (Bath)Mb is not known(17), and it is not clear whetherM. alcaliphilum 20Z producesMb. No uptake of Mb-associated copper was observed forE. coli, suggesting that uptake of Mb may be specific to metha-notrophic bacteria.We next investigated possible mechanisms of unchelated Cu
and Cu-Mb uptake. In Gram-negative bacteria, transport ofnutrients across the outer membrane can occur via passive dif-fusion or by active transport (37). Porins typically allow diffu-sion of smaller solutes, including cations and anions, and are aprobable route for unchelated Cu uptake intoM. trichosporiumOB3b cells.However, the 1217-DaCu-Mbmolecule is likely toolarge for classical porins. By analogy to iron siderophores,cobalamin, and putative nickel complexes, we suggested previ-ously (21) that Cu-Mb uptake might be mediated by outermembrane TonB-dependent transporters (TBDTs) (38–41).
FIGURE 3. Uptake of mBBr-Mb and mBBr-Cu-Mb by M. trichosporium OB3b cells observed via confocal microscopy. Fluorescence images were taken atwavelengths of 405 nm (cyan, mBBr) and 543 nm (red, the lipophilic membrane dye FM 4-64). The corresponding phase-contrast image is shown in gray. Themerged images illustrate the distribution of both signals, and the intensity profile (y axis, in arbitrary units) of both dyes across the center of a representativecell is also shown. The scale bar corresponds to 4 mm. Cells incubated with either (A) mBBr-Cu-Mb or (B) mBBr-Mb exhibit cytoplasmic localization.
Uptake of Methanobactin
37316 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 43 • OCTOBER 28, 2011
by guest on June 3, 2018http://w
ww
.jbc.org/D
ownloaded from

In TonB-dependent uptake systems, energy from the innermembrane proton motive force is transduced by interaction ofthe TBDTs with the inner membrane TonB-ExbB-ExbD pro-tein complex.To test the hypothesis that uptake of Mb is via specific outer
membrane receptors, the possibility of apo-Mb competingwithCu-Mb uptake was investigated (Fig. 4 and supplemental TableS2). A 3-fold increase in intracellular copper upon addition of 5�M Cu-Mb to copper-starved cells decreases in the presence ofapo-Mb and is reduced to �0.6 upon addition of 10 �M apo-Mb. Thus, apo-Mb and Cu-Mbmay compete for uptake, whichlikely involves the same surface receptors. Similarly, competi-tion for binding to the FpvA TBDT is observed for apo- andholo-pyoverdin (42–44).To probe further the involvement of porins or TBDTs in
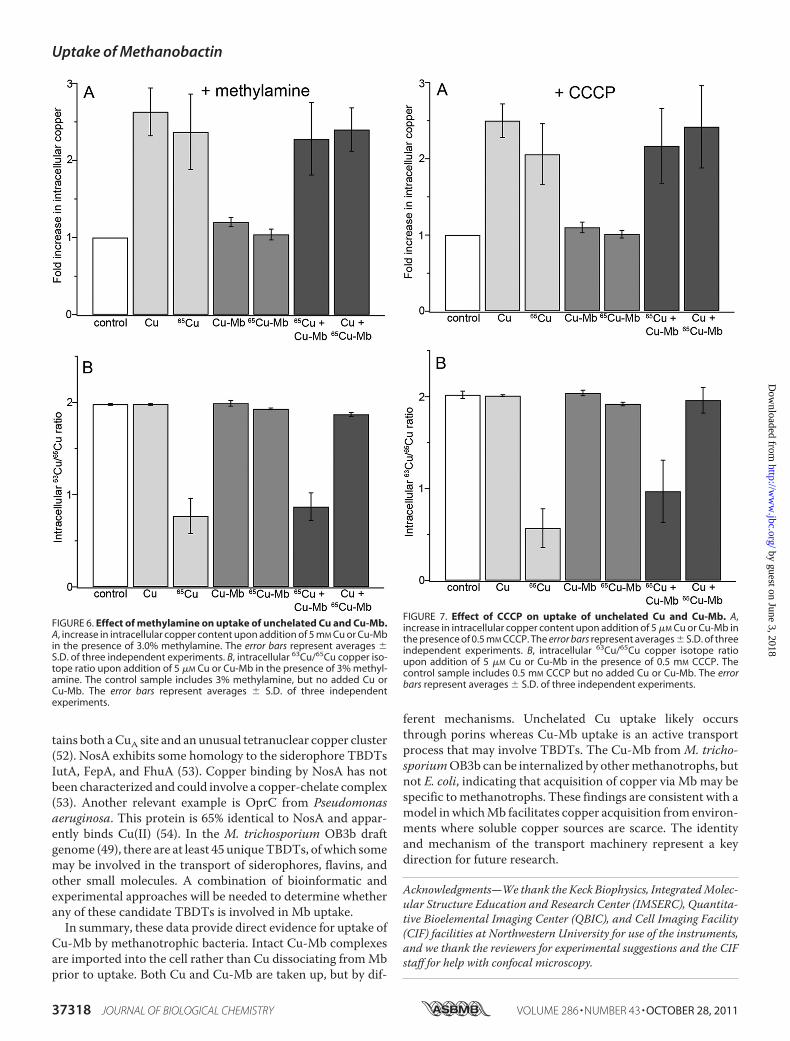
copper uptake byM. trichosporiumOB3b, we employed trans-port inhibitors. Polyamines such as spermine and putrescineare known to inhibit passive transport by porins (45, 46). Treat-ment of cells with 0.5 mM spermine inhibits Cu uptake but hasno effect on Cu-Mb uptake (Fig. 5A and supplemental TableS3). Therefore, unchelated Cu likely diffuses into the cell viaporins whereas Cu-Mb is transported by a different mecha-nism, consistent with uptake of the intact Cu-Mb molecule.Uncouplers such as CCCP, FCCP, dinitrophenol, and methyla-mine dissipate the inner membrane protonmotive force, inter-fering with active transport. In particular, CCCP and FCCPhave been used to block TBDT-mediated uptake of iron sidero-phores (42, 47, 48). Of seven uncoupling agents tested byethidium bromide accumulation assays (27–29), CCCP andmethylamine were found to inhibit active transport in M. tri-chosporium OB3b (supplemental Fig. S6). Treatment of cellswith 3% methylamine (Fig. 6A and supplemental Table S4) or0.5 mM CCCP (Fig. 7A and supplemental Table S5) inhibitsCu-Mb uptake, but not Cu uptake, indicating that Cu-Mbtransport is an energy-dependent process.Isotopic labeling experiments are also consistent with inde-
pendent pathways for Cu and Cu-Mb uptake. The intracellular63Cu/65Cu ratio is �0.5 (Fig. 2B) upon incubation of cells with65Cu and is�2.0 in the presence of 65Cu and spermine, indicat-
ing that very little 65Cu uptake is occurring (Fig. 5B). By con-trast, spermine does not affect the uptake of 65Cu-Mb (Fig. 5B).The converse is observed with methylamine (Fig. 6B) andCCCP (Fig. 7B). Addition of methylamine or CCCP prevents adecrease in the intracellular 63Cu/65Cu ratio upon addition of65Cu-Mb but has no effect on 65Cu uptake. Isotopic ratios ofless than �1.0 are observed whenmixtures of 65Cu and Cu-Mbor Cu and 65Cu-Mb are provided in the presence of each inhib-itor (Figs. 5B, 6B, and 7B), consistent with selective targeting oftwo different pathways.The observations that Cu-Mb uptake is energy-dependent
and that apo-Mb competes for uptake with Cu-Mb suggest thatTBDTs could facilitate transport of Cu-Mb across the outermembrane, similar to uptake of iron siderophores (21). There issome precedent in the literature for TBDTs participating incopper uptake (41). The NosA protein from Pseudomonasstutzeri is an outer membrane protein necessary for formationof copper-loaded nitrous-oxide reductase (50, 51) which con-
FIGURE 4. Effect of apo-Mb on uptake of Cu-Mb by M. trichosporium OB3bcells. Cells were incubated with varying concentrations of apo-Mb in thepresence of 5 �M Cu-Mb. The error bars represent averages � S.D. of threeindependent experiments.
FIGURE 5. Effect of spermine on uptake of unchelated Cu and Cu-Mb. A,increase in intracellular copper content upon addition of 5 �M Cu or Cu-Mb inthe presence of 0.5 mM spermine. B, intracellular 63Cu/65Cu copper isotoperatio upon addition of 5 �M Cu or Cu-Mb in the presence of 0.5 mM spermine.The control sample includes 0.5 mM spermine, but no added Cu or Cu-Mb. Theerror bars represent averages � S.D. of three independent experiments.
Uptake of Methanobactin
OCTOBER 28, 2011 • VOLUME 286 • NUMBER 43 JOURNAL OF BIOLOGICAL CHEMISTRY 37317
by guest on June 3, 2018http://w
ww
.jbc.org/D
ownloaded from
tains both aCuA site and an unusual tetranuclear copper cluster(52). NosA exhibits some homology to the siderophore TBDTsIutA, FepA, and FhuA (53). Copper binding by NosA has notbeen characterized and could involve a copper-chelate complex(53). Another relevant example is OprC from Pseudomonasaeruginosa. This protein is 65% identical to NosA and appar-ently binds Cu(II) (54). In the M. trichosporium OB3b draftgenome (49), there are at least 45 uniqueTBDTs, ofwhich somemay be involved in the transport of siderophores, flavins, andother small molecules. A combination of bioinformatic andexperimental approaches will be needed to determine whetherany of these candidate TBDTs is involved in Mb uptake.In summary, these data provide direct evidence for uptake of
Cu-Mb by methanotrophic bacteria. Intact Cu-Mb complexesare imported into the cell rather than Cu dissociating fromMbprior to uptake. Both Cu and Cu-Mb are taken up, but by dif-
ferent mechanisms. Unchelated Cu uptake likely occursthrough porins whereas Cu-Mb uptake is an active transportprocess that may involve TBDTs. The Cu-Mb from M. tricho-sporiumOB3b can be internalized by othermethanotrophs, butnot E. coli, indicating that acquisition of copper via Mb may bespecific tomethanotrophs. These findings are consistent with amodel inwhichMb facilitates copper acquisition from environ-ments where soluble copper sources are scarce. The identityand mechanism of the transport machinery represent a keydirection for future research.
Acknowledgments—We thank the Keck Biophysics, IntegratedMolec-ular Structure Education and Research Center (IMSERC), Quantita-tive Bioelemental Imaging Center (QBIC), and Cell Imaging Facility(CIF) facilities at Northwestern University for use of the instruments,and we thank the reviewers for experimental suggestions and the CIFstaff for help with confocal microscopy.
FIGURE 6. Effect of methylamine on uptake of unchelated Cu and Cu-Mb.A, increase in intracellular copper content upon addition of 5 mM Cu or Cu-Mbin the presence of 3.0% methylamine. The error bars represent averages �S.D. of three independent experiments. B, intracellular 63Cu/65Cu copper iso-tope ratio upon addition of 5 �M Cu or Cu-Mb in the presence of 3% methyl-amine. The control sample includes 3% methylamine, but no added Cu orCu-Mb. The error bars represent averages � S.D. of three independentexperiments.
FIGURE 7. Effect of CCCP on uptake of unchelated Cu and Cu-Mb. A,increase in intracellular copper content upon addition of 5 �M Cu or Cu-Mb inthe presence of 0.5 mM CCCP. The error bars represent averages � S.D. of threeindependent experiments. B, intracellular 63Cu/65Cu copper isotope ratioupon addition of 5 �M Cu or Cu-Mb in the presence of 0.5 mM CCCP. Thecontrol sample includes 0.5 mM CCCP but no added Cu or Cu-Mb. The errorbars represent averages � S.D. of three independent experiments.
Uptake of Methanobactin
37318 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 43 • OCTOBER 28, 2011
by guest on June 3, 2018http://w
ww
.jbc.org/D
ownloaded from

REFERENCES1. Semrau, J. D., Dispirito, A. A., and Yoon, S. (2010) FEMS Microbiol. Lett.
34, 496–5312. Balasubramanian, R., Smith, S. M., Rawat, S., Yatsunyk, L. A., Stemmler,
T. L., and Rosenzweig, A. C. (2010) Nature 465, 115–1193. Martinho, M., Choi, D. W., DiSpirito, A. A., Antholine, W. E., Semrau,
J. D., and Münck, E. (2007) J. Am. Chem. Soc. 129, 15783–157854. Rosenzweig, A. C., Frederick, C. A., Lippard, S. J., and Nordlund, P. (1993)
Nature 366, 537–5435. Murrell, J. C., McDonald, I. R., and Gilbert, B. (2000) Trends Microbiol. 8,
221–2256. Hakemian, A. S., and Rosenzweig, A. C. (2007) Annu. Rev. Biochem. 76,
223–2417. DiSpirito, A. A., Zahn, J. A., Graham, D. W., Kim, H. J., Larive, C. K.,
Derrick, T. S., Cox, C. D., and Taylor, A. B. (1998) J. Bacteriol. 180,3606–3613
8. Téllez, C.M.,Gaus, K. P., Graham,D.W., Arnold, R.G., andGuzman, R. Z.(1998) Appl. Environ. Microbiol. 64, 1115–1122
9. Choi, D.W., Zea, C. J., Do, Y. S., Semrau, J. D., Antholine,W. E., Hargrove,M. S., Pohl, N. L., Boyd, E. S., Geesey, G. G., Hartsel, S. C., Shafe, P. H.,McEllistrem,M. T., Kisting, C. J., Campbell, D., Rao, V., de laMora, A.M.,and DiSpirito, A. A. (2006) Biochemistry 45, 1442–1453
10. El Ghazouani, A., Baslé, A., Firbank, S. J., Knapp, C. W., Gray, J., Graham,D. W., and Dennison, C. (2011) Inorg. Chem. 50, 1378–1391
11. Summer, K. H., Lichtmannegger, J., Bandow, N., Choi, D. W., DiSpirito,A. A., and Michalke, B. (2011) J. Trace Elem. Med. Biol. 25, 36–41
12. Behling, L. A., Hartsel, S. C., Lewis, D. E., DiSpirito, A. A., Choi, D. W.,Masterson, L. R., Veglia, G., andGallagher,W. H. (2008) J. Am. Chem. Soc.130, 12604–12605
13. Kim, H. J., Graham, D. W., DiSpirito, A. A., Alterman, M. A., Galeva, N.,Larive, C. K., Asunskis, D., and Sherwood, P. M. A. (2004) Science 305,1612–1615
14. Crosa, J. H., andWalsh, C. T. (2002)Microbiol.Mol. Biol. Rev. 66, 223–24915. Demange, P., Bateman, A., Mertz, C., Dell, A., Piémont, Y., and Abdallah,
M. A. (1990) Biochemistry 29, 11041–1105116. Drechsel, H., and Jung, G. (1998) J. Pept. Sci. 4, 147–18117. Choi, D. W., Bandow, N. L., McEllistrem, M. T., Semrau, J. D., Antholine,
W. E., Hartsel, S. C., Gallagher, W., Zea, C. J., Pohl, N. L., Zahn, J. A., andDiSpirito, A. A. (2010) J. Inorg. Biochem. 104, 1240–1247
18. Krentz, B. D., Mulheron, H. J., Semrau, J. D., DiSpirito, A. A., Bandow,N. L., Haft, D. H., Vuilleumier, S., Murrell, J. C., McEllistrem, M. T., Hart-sel, S. C., and Gallagher, W. H. (2010) Biochemistry 49, 10117–10130
19. Yoon, S., Kraemer, S.M., Dispirito, A. A., and Semrau, J. D. (2010)Environ.Microbiol. Rep. 2, 295–303
20. Balasubramanian, R., Levinson, B. T., and Rosenzweig, A. C. (2010) Appl.Environ. Microbiol. 76, 7356–7358
21. Balasubramanian, R., and Rosenzweig, A. C. (2008) Curr. Opin. Chem.Biol. 12, 245–249
22. Knapp, C. W., Fowle, D. A., Kulczycki, E., Roberts, J. A., and Graham,D. W. (2007) Proc. Natl. Acad. Sci. U.S.A. 104, 12040–12045
23. Kulczycki, E., Fowle, D. A., Kenward, P. A., Leslie, K., Graham, D.W., andRoberts, J. A. (2011) Geomicrobiol. J. 28, 1–10
24. Kulczycki, E., Fowle, DA., Knapp, C., Graham, DW., Roberts JA. (2007)Geobiology 5, 251–263
25. Hakemian, A. S., Kondapalli, K. C., Telser, J., Hoffman, B. M., Stemmler,T. L., and Rosenzweig, A. C. (2008) Biochemistry 47, 6793–6801
26. Hakemian, A. S., Tinberg, C. E., Kondapalli, K. C., Telser, J., Hoffman,B. M., Stemmler, T. L., and Rosenzweig, A. C. (2005) J. Am. Chem. Soc.127, 17142–17143
27. Minato, Y., Shahcheraghi, F., Ogawa, W., Kuroda, T., and Tsuchiya, T.(2008) Biol. Pharm. Bull. 31, 516–519
28. Paixão, L., Rodrigues, L., Couto, I., Martins, M., Fernandes, P., de Car-valho, C. C., Monteiro, G. A., Sansonetty, F., Amaral, L., and Viveiros, M.(2009) J. Biol. Eng. 3, 18
29. Turovskiy, Y., Ludescher, R. D., Aroutcheva, A. A., Faro, S., andChikindas,M. L. (2009) Probiotics Antimicrob. Proteins 1, 67–74
30. Navarrete, J. U., Viveros, M., Elizey, J. T., and Borrok, D. M. (2011) Appl.Geochem. 26, S319–321
31. Maréchal, C. N., Télouk, P., and Albarède, F. (1999) Chem. Geol. 156,251–273
32. Jelínková, A., Malínská, K., Simon, S., Kleine-Vehn, J., Parezová, M.,Pejchar, P., Kubes, M., Martinec, J., Friml, J., Zazímalová, E., and Petrásek,J. (2010) Plant J. 61, 883–892
33. Journo, D., Winter, G., and Abeliovich, H. (2008)Methods Enzymol. 451,79–88
34. Muchová, K., Jamroskovic, J., and Barák, I. (2010) Res. Microbiol. 161,783–790
35. Kim, H. J., Galeva, N., Larive, C. K., Alterman, M., and Graham, D. W.(2005) Biochemistry 44, 5140–5148
36. Choi, D. W., Do, Y. S., Zea, C. J., McEllistrem, M. T., Lee, S. W., Semrau,J. D., Pohl, N. L., Kisting, C. J., Scardino, L. L., Hartsel, S. C., Boyd, E. S.,Geesey, G. G., Riedel, T. P., Shafe, P. H., Kranski, K. A., Tritsch, J. R.,Antholine, W. E., and DiSpirito, A. A. (2006) J. Inorg. Biochem. 100,2150–2161
37. Nikaido, H. (2003)Microbiol. Mol. Biol. Rev. 67, 593–65638. Postle, K., and Kadner, R. J. (2003)Mol. Microbiol. 49, 869–88239. Krewulak, K. D., and Vogel, H. J. (2011) Biochem. Cell Biol. 89, 87–9740. Noinaj, N., Guillier, M., Barnard, T. J., and Buchanan, S. K. (2010) Annu.
Rev. Microbiol. 64, 43–6041. Schauer, K., Rodionov, D. A., and de Reuse, H. (2008)Trends Biochem. Sci.
33, 330–33842. Clément, E., Mesini, P. J., Pattus, F., and Schalk, I. J. (2004) Biochemistry
43, 7954–796543. Schalk, I. J., Hennard, C., Dugave, C., Poole, K., Abdallah, M. A., and
Pattus, F. (2001)Mol. Microbiol. 39, 351–36044. Schalk, I. J., Yue, W. W., and Buchanan, S. K. (2004) Mol. Microbiol. 54,
14–2245. Iyer, R., and Delcour, A. H. (1997) J. Biol. Chem. 272, 18595–1860146. Pagès, J. M., James, C. E., andWinterhalter, M. (2008)Nat. Rev. Microbiol.
6, 893–90347. Larsen, R. A., Thomas, M. G., and Postle, K. (1999) Mol. Microbiol. 31,
1809–182448. Braun, M., Killmann, H., and Braun, V. (1999) Mol. Microbiol. 33,
1037–104949. Stein, L. Y., Yoon, S., Semrau, J. D., DiSpirito, A. A., Crombie, A., Murrell,
J. C., Vuilleumier, S., Kalyuzhnaya, M. G., Op den Camp, H. J., Bringel, F.,Bruce, D., Cheng, J. F., Copeland, A., Goodwin, L., Han, S., Hauser, L.,Jetten, M. S., Lajus, A., Land, M. L., Lapidus, A., Lucas, S., Médigue, C.,Pitluck, S.,Woyke, T., Zeytun, A., and Klotz,M.G. (2010) J. Bacteriol. 192,6497–6498
50. Lee, H. S., Hancock, R. E., and Ingraham, J. L. (1989) J. Bacteriol. 171,2096–2100
51. Mokhele, H., Tang, Y. J., Clark, M. A., and Ingraham, J. L. (1987) J. Bacte-riol. 169, 5721–5726
52. Zumft, W. G. (2005) J. Mol. Microbiol. Biotechnol. 10, 154–16653. Lee, H. S., Abdelal, A. H., Clark, M. A., and Ingraham, J. L. (1991) J. Bac-
teriol. 173, 5406–541354. Yoneyama, H., and Nakae, T. (1996)Microbiol. 142, 2137–2144 .
Uptake of Methanobactin
OCTOBER 28, 2011 • VOLUME 286 • NUMBER 43 JOURNAL OF BIOLOGICAL CHEMISTRY 37319
by guest on June 3, 2018http://w
ww
.jbc.org/D
ownloaded from

Ramakrishnan Balasubramanian, Grace E. Kenney and Amy C. RosenzweigDual Pathways for Copper Uptake by Methanotrophic Bacteria
doi: 10.1074/jbc.M111.284984 originally published online September 7, 20112011, 286:37313-37319.J. Biol. Chem.
10.1074/jbc.M111.284984Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2011/09/07/M111.284984.DC1
http://www.jbc.org/content/286/43/37313.full.html#ref-list-1
This article cites 54 references, 12 of which can be accessed free at
by guest on June 3, 2018http://w
ww
.jbc.org/D
ownloaded from


















