
Elm City CrossFit Nutrition, Fitness & Success Presented by: Vincent Lindsley.

PSYCHOLOGICAL P H E N O M E N A AND THE ELECTROENCEPHALOGRAM
DONALD B. LINDSLEY, Ph.D. Departments of Psychology and Pediatrics, Uni~,ersity of California,
Los Angeles
It is well to remind ourselves from time to time that the problem of relatin 9 electrical activity of the brain to psychological phen- omena constituted one of the main interests of the founder of electroencepalography, Hans Berger. His numerous publications during the decade from 1929 to 1938 strong- ly attest this fact. Although his psychological interpretations have not always proved completely acceptable, his empirical dis- coveries constituted the opening wedge that led others to confirm and extend his ob- servations, especially with respect to the characteristics of the EEG. In addition to the rulin 9 out of artifacts and the establish~ ment of the EEG as a characteristic electrical property of the brain, the early studies of Berger and others clearly delineated the al~ pha and beta components of the EEG tracing as we know it today.
It is also well to recall the efforts in the laboratories of Adrian, Bishop, Jasper, Davis, Gibbs, Kornm/iller. Loomis, Kreezer. Bremer, Fessard, Travis, Hoagland, and others during the early days of EEG study. when attempts were made not only to un- derstand the basic aspects of the EEG, but to relate them to physiological and psycho- logical processes. Throughout the 1930's one and two channel recording units in a handful of laboratories in this country and abroad often worked overtime attempting to establish relationships between various char- acteristics of the EEG and the psychological aspects of sensation, perception, attention, emotion, learning, intelligence, personality and the like (see Jasper 1937: Kreezer 1938; Lindsley 194't). Some of the studies were superficial, with little control of fundamental variables: others were carefully done in a detailed and painstaking manner.
For the most part these initial attempts at correlation of neurophysiological and psy- chological events were not very successful. The correlations themselves were not re- markable and no comprehensive theory arose from them. Yet a careful re-reading of some of the early studies reveals that many of the original EEG observations have stood the test of time.
W h y have not EEG and psychological correlations been pursued more systematically and persistently ? W h y has there been so little success in relating neurophysiological data to subjective states and behavioral manifestations ? There are probably many reasons. Almost from the start, the lure and excitement of new clinical discoveries was a distraction to serious effort in this direction. Secondly, the premature, if not actually grandiose, hope that the mysterious and ubi- quitous alpha rhythm might be related to intelligence, personality and other similarly broad and in themselves undefined para- meters, soon exhausted interests in that direction. There were other matters such as constantly improved equipment which led to the hope that the next new development, whether it was more channels, an automatic analyzer, or what not, might facilitate the process of correlation.
Despite a certain amount of preoccupation with empirical results and the startlin 9 and significant clinical correlations revealed by the EEG, effort persisted in trying to un- cover the mechanism of the autonomous rhythms of the brain. Although a basic un- derstandin 9 of the origin and nature of the alpha rhythm, as well as other aspects of the EEG, still eludes us today, tremendous strides have been made in recent years in gaining factual information about the mech-
[ 443 1

444 DONALD B. LINDSLEY
anism of control. Al though far from com- plete in detail, the f ramework seems to be emer9in 9, and it is in the light of some of these new developments coupled with some reinterpretat ion of older observations that I would like to review the problem of psy- chological and E E G correlates.
This will inevitably verge on the spec- ulative, and in some instances 9 ° beyond the facts, but this I believe we must do if we are to propose new hypotheses for ex- perimental test. Adr ian (19't7) in his little book "Physical Background of Perception", outlines how much we have learned about receptor processes, nerve conduction, and interaction, and the point by point followin 9 of sensory messages as they traverse their pa thways to the brain. H e states: "So far we have thought of the cerebral cortex as a screen on which pat terns are thrown by the different sense organs. T h e pattern cor- responds more or less with the pattern of sensory stimulation because the pa thways from the receptor endings preserve the same anatomical a r rangement on the way up to the cortical receivin 9 area." But he goes on to indicate that mappin 9 of the sensory messages on the cortex is not enou9 h, and despite all of our accumulated neurophys- iological data we come face to face with the problems of psychology, namely, recognition, memory, habit formation and others.
Five years have elapsed since this little book was published and some of our most illuminating information on the central in- tegrat ing mechanism has come to the fore. This information seems to be constantly 9etting closer to the mechanisms needed to explain psychological phenomena. But I wonder if it is not a t wo-way street, and that psychological data are needed to help us explain some of our neurophysiological ob- servations. It seems to me the time is fast approaching when new hypotheses and new experiments are going to require psycho- logical and behavioral data, both in their formulation and execution. W e need a closer liaison between established observation and fact with regard to psychological and neuro-
physiological processes. T h e problem of the transition between nerve impulses arrivin 9 at the cortex and the percepts, symbols and ideas which result will require more than neurophysiological data for its resolution.
M a n y facets of reliable psychological data are open to us, especially in the areas of sensation, perception, action and learnin 9. One of the outstandin 9 parameters of these data is time. Stimulus propert ies of kind, in- tensity and spatiali ty may also be important parameters, but our most common and per- haps most useful parameter in t ransformin 9 and integratin 9 two sets of data is that of time. Furthermore , time and its derivatives, rate, rhythm, sequence and so forth seem to be fundamental properties or conditions of nervous system integration.
I [ I ¢ I T I [ D
I E L A X £ D
DROWSY
A S L [ [ P
OI[EP S L E E P
I ! S rC . i IOt~V*
Fig. 1 General excitatory states showing range of variation in normal electroencephalogram. (From Jasper, 1941, by permission of author and publisher,)
Let us return now to the E E G and examine some of its characteristics, espe- cially with regard to the time parameter . W e may observe in figure 1 that the pat tern of activity (but especially frequency, a time characterist ic) , varies widely during the range of behavioral states common to a normal adult. T h e pat tern varies from a fast desynchronized activity durin 9 emotional ex- citement or alert at tentiveness to a degree

PSYCHOLOGICAL PHENOMENA AND EEG 445
of rhythmic synchrony known as the alpha rhythm during quiet, relaxed wakefulness with eyes closed, and to a progressively slower rate of synchrony in the successive stages of sleep.
In our attempts to relate psychological phenomena to the EEG we have been con-
correlations with things psychological ? The very special conditions of relaxation, dark- ness and quiet requisite to the alpha rhythm, are hardly representative of much of our normal waking life. Instead such conditions are really precursors of sleep, and the slightest effort, noise, or other stimulation
TABLE I
PSYCHOLOGICAL STATES AND THEIR EEG, CONSCIOUS AND BEHAVIORAL CORRELATES
BEHAVIORAL CONTINHHM
Strong, Excited Emo- tion (Fear) (Rage) (Anxiety)
Alert Attentiveness
Relaxed Wakefulness
I
:Drowsiness
Light Sleep
Deep Sleep
Coma
I Death
ELECTRO- ENCEPHALOGRAM
Desynchronized: Low to moderate amplitude; fast mixed frequencies.
Partially synchronized : Mainly fast, low ampli- tude waves.
Synchronized : Optimal alpha rhythm.
Reduced alpha "5 occa- sional low amplitude slow waves.
Spindle bursts '5 slow waves (larger) Loss of alphas.
Large and very slow waves (synchrony but on slow time base) Ran- dom, irregular pattern.
Isoelectric to irregular large slow waves.
Isoelectric: Gradual and permanent disappearance of all electrical activity.
STATE OF AWARENESS
Restricted awareness; divided attention; diffuse, hazy; "Confusion"
Selective attention, bu,t may vary or shift. "Con- cerrtration" an;ticipation, "set"
BEHAVIORAL i EFFICIENCY
Poor: (lack o f - - - - ~ o n t r o l , freezing-up, disorgan- ized), j
Good: (efficient, selec- t;v e~ 2eUdi c k ~ j ; a c~ ie;njl ) Or-
responses.
f A t t e n ! i o n w a n d e r s - - notlGood: (routine reaction: orced, t 'avors tree I'5 creative thought).
association.
Borderline. partial Poor: (uncoordinated awareness. Imagery ,5 [sporadic, lacking reverie. "Dream-like sequential timing). states".
Markedly reduced con- Absent sciousness (loss of con- sciousness ) Dream state.
Complete loss of Absent awareness (no memory for stimulation or for dreams).
Complete loss of con- Absent sciousness little or no re- sponse to stimulation; amnesia.
Complete loss of aware-Absent ness as death ensues.
cerned mainly with only one of these stages of normal brain activity, namely that of the alpha rhythm. Is it possible that our pre- occupation with the alpha rhythm, because of its dominance, regularity and measurability has led us astray and away from significant
will quickly break up this apparently optimal state of rhythmic oscillation during waking.
Table I presents what appears to be a behavioral continuum, based upon both EEG and psychological correlates as determined empirically. There are undoubtedly in-

4zt6 DONALD B. LINDSLEY
adequacies and also inaccuracies in such a categorization, but in the present state of our empirical knowledge this table is a reasonably accurate presentation of ac- cumulated data from many sources. The fact that as wide a range of patterns and fre- quencies as are observed in the EEG during the stages from drowsiness to excited emotion in a normal person can occur, to- gether with the variations of awareness and behavioral efficiency indicated, suggests im- mediately why there is difficulty in assessing personality or in differentiating between psy- chiatric conditions by means of the EEG. But all the more so because we have con- cerned ourselves for the most part with the measurable aspects of the alpha rhythm. In a sense the alpha rhythm is an abstraction both electroencephalographically and psy- chologically; at best it represents an unusual and limited state of affairs in the life of the organism. Yet it has a remarkable reg- ularity and constancy of rhythm and pattern which is virtually a hallmark for the in- dividual.
Let us turn now to some more specific relationships between the alpha rhythm and psychological and behavioral processes. Be- fore doing so it is necessary to discuss two points upon which these relationships de- pend. One of these has to do with a distinc- tion between alpha rhgthrn and alpha activitt¢. This distinction was first emphasized by Bartley (1940), although the assumption un- derlying it was brought out in the writings of Adrian, Jasper and others who held to the view that the alpha rhythm represented an optimal synchronization of electrical ac- tivity of cortical elements, and that breaking up or disappearance of the alpha rhythm re- presented a desynchronization of neuronal activity. In essence the distinction between alpha rhtlthm and alpha activittt might be put quite succinctly in this form: Alpha activity may exist without a recordable alpha rhythm.
I should like to add to this view the as- sumption that alpha activity is a basic me- tabolic or respiratory rhythm of the in-
dividual brain cell, and its electrical variation alone or in small aggregates of cells is nor- mally too small to be recorded from the surface of the scalp. Only when literally thousands of cells are responding in syn- chrony is sufficient summation attained to produce a recordable alpha rhythm. A low amplitude fast or desynchronized EEG as in alert attentiveness might then represent fractional synchronization in many smaller aggregates of cells but with random phase relations, or actually slightly different fre- quencies, as has been shown by Walter (1950a, 1950b). This would mean that an alpha activity rhythm could exist in many small, independent aggregates of brain cells, in the absence of an over-all alpha rhythm in the usual sense.
5 vvw
Fig. 2 Desynchronization of alpha rhythm in normal adults, spontaneously in A, C and D; with visual stimula- tion in B. Time in 1/5 sec. (From Lindsley, 1938, by permission of publisher.)
In one of my old oscillograph records (fi 9. 2), where the sensitivity and magni- fication was much 9reater than in modern inkwriters, the necessary detail is provided to see the nature of spontaneous (A, C, D) and stimulated (B) desynchronization of the occipital alpha rhythm. In A a brief lapse in the alpha rhythm, leaves a definite rhythm of lower voltage and double the frequency of the original and subsequent alpha rhythm. In B the blocking of the occipital alpha rhythm by a light stimulus leaves in its wake

PSYCHOLOGICAL PHENOMENA AND EEG 4't7
a rhythm of approximately double the fre- quency and perhaps one quarter the am- plitude of the original alpha rhythm. Record C shows the occasional emergence of alpha rhythm from an underlyin 9 activity of ap- proximately double the frequency and one- half the amplitude.
The next assumption is that the alpha ac- tivity cycle represents for the individual cell or the a99re9ates of cells with which it is associated, an alternatin 9 excitability cycle. This conception was first proposed by Bishop (1933) and was later amplified by Bishop (1936) and by Jasper (1936), and was sup- ported in part by inferential evidence. For example, Bartley and Bishop (1933) and Bishop (1933) demonstrated that impulses initiated in the optic nerve of the rabbit found access to the cortex in the form of
BRi,k:KE
,~,~ ~.
4 El i~ i i i
l ight-dark ratio of 1 to 1, and shows the Talbot effect of one-half the brightness of s teady illumination at frequencies of ap- proximately 28 and above for this particular intensity level of illumination. However when the frequency of the flickerin 9 light was decreased to the neighborhood of 9 or 10 c/sec., there was marked brightness en- hancement, the so-called Briicke effect. 1 The fact that an increase in apparent bright- ness of 100 per cent over steady illumination occurred at a flicker frequency of the alpha rhythm, with decreasin 9 brightness on either side of that frequency, stron91y sug- gests that the light flashes were synchroniz- ed with the alpha activity and its excitability cycle. Here we see that a psycholo9ical datum of experience affords support for a neurophysiological theory, but at the same
B
EFFECT
B~.C, HTNE$S OF $1"F.N)y ILLUUINATI~/~-.~
~ _ _ _ _ ~ F F . C.FF.
TA~.BOT EFFECT
CIrF.
FLASHES PER SECONO | 2 16 20 2 4 2 G 3 2 36
Fig. 3 Curves showing change in apparent briqhtness at flash frequencies above and below critical flicker frequency (C.F.F.) for varying light-dark ratios, A, L.D.R. 1 : 1, B, 7 : 2, C. 8 : 1. In curve A brightness enhancement (Bart!ey effect rather than Briicke effect) occurs at a flash frequency corresponding to an alpha rhythm frequency of about 9 to 10 per sec. From Bartley (1939, 1941), by permission of author and publishers.
evoked responses only at certain intervals corresponding to the spontaneous rhythmic or alpha activity cycle at about 5 per sec. Accordingly Bishop (1936) interpreted this as evidence of a cortical excitability cycle.
Psychological data supporting this con- cept have been supplied by Bartley (1939). Figure 3 summarizes his results on apparent subjective brightness of a flickering light at different frequencies. Curve A illustrates a
time is made more understandable as a psy- chological experience, since it has been shown that brightness discrimination is modified, presumably by an intrinsic ex- citability cycle associated with alpha ac-
z The brightness enhancement effect of flickering light at 9 to 10 c/sec., although originally labelled by Bartley as the Briicke effect has since been de- signated by him as "brightness enhancement effect" (see Bartley, 1941, p. 137). It might more properly be called the "Bartley effect".

448 DONALD B. LINDSLEY
tivity. T h a t a recordable alpha rhythm does not have to be present for this to occur is further indication of the distinction between alpha rhythm and alpha activity, the latter being assumed to be present in those ag- gregates of cells participating in the re- sponse.
Recent experimental work by Chang (1950, 1951) strongly reinforces the con- cept of an excitability cycle in the cortex and perhaps also in the thalamus. If this is true, two levels of screening of incoming
coding system are necessary in order that our perceptual world and our reactions to it are not distorted or smeared by the more or less continuous influx of sensory stimuli.
A brief example from an unpublished paper by Meister (1951) will serve to illustrate this point. W h e n one moves the eyeball by finger pressure, or when it is moved involuntarily by vestibular or brain- stem reflex the visual field moves with it; but when it moves voluntarily in reading or tracking movements discrete fixations oc-
Fig. 4 Schematic representation of concept of "neuronic shutter mechanism" after Meister (1951)'. Movement of eyes voluntarily as in reading or tracking initiated from motor eye-fields (area 8) accompanied by momentary inhibition in visual cortex, thus preventing smearing of visual images. See text for fuller explanation.
sensory impulses are possible. W h e t h e r these work in harmony and synchrony or at times are mutually exclusive of one another, may well be a determinant of the varied states of awareness which are possible under normal conditions, but especially under con- ditions of selective and differential aware- ness, during heightened degrees of alertness versus drowsiness, during emotional states, under partial anesthesia, hypnosis and so forth. Apar t from these considerations it appears from the nature of psychological experience and behavior that a pulsing and
cur without smearing. Th e eye fixations in reading a line of type are a case in point. Meister has proposed, and I have illustrated diagrammatically in figure 4, the manner in which voluntary eye movements initiated from the motor eye fields of area 8, may simultaneously and momentarily inhibit or block incoming impulses through connections with the occipital cortex. Th e exact manner in which this may occur remains to be de- monstrated, but the psychological effect im- plies that some type of neuronic shutter mechanism is operating. Conceivably this
PSYCHOLOGICAL PHENOMENA AND EEG "t't9
could be through simultaneous discharges from area 8 to the extraocular muscles and the occipital cortex, the latter in effect in- hibitin 9 reception momentarily by resetting the alpha activity cycle in the given ag- gregate of cortical neurons involved in the perception. On the other hand it could arise through a timing mechanism which would permit activation of the eye muscles only in synchrony with the inexcitable phase of the alpha activity of the occipital cortex. In either case a kind of neuronic shutter effect would take place during the movement of the eye, in much the same fashion that a shutter in a movie projector permits two
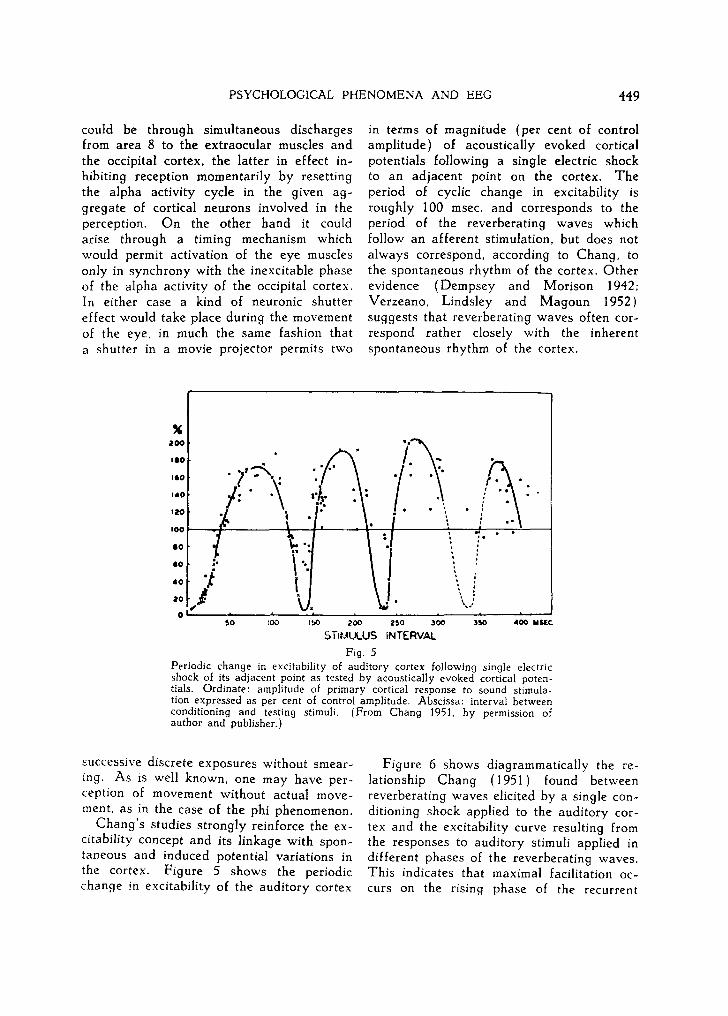
in terms of magnitude (per cent of control amplitude) of acoustically evoked cortical potentials following a single electric shock to an adjacent point on the cortex. The period of cyclic change in excitability is roughly 100 msec. and corresponds to the period of the reverberating waves which follow an afferent stimulation, but does not always correspond, according to Chang, to the spontaneous rhythm of the cortex. Other evidence (Dempsey and Morison 1942; Verzeano, Lindsley and Magoun 1952) suggests that reverberating waves often cor- respond rather closely with the inherent spontaneous rhythm of the cortex.
zoo
leo
l lo I&O
IZo
I00
8 0
I 0
4 0
IO'
0
• l
" i '
L L
f r o • " •
t
I
,. :
~0 ~00 I ~ 20¢ 2SO 300 3 ~ 400 MS(r.
ST#~t.N..US INTERVAL Fig. 5
Periodic change in e×citabi]ity of auditory cortex following single electric shock of its adjacent point as tested by acoustica]|y evoked cortical poten- tials. Ordinate: amp|Rude of primary cortica| response to sound stimula- tion expressed as per cent of control amplitude. Abscissa: interval between conditioning and testing stimuli. (From Chang 195], by permission of author and publisher.)
successive discrete exposures without smear- ing. As is well known, one may have per- ception of movement without actual move- ment, as in the case of the phi phenomenon.
Chang's studies strongly reinforce the ex- citability concept and its linkage with spon- taneous and induced potential variations in the cortex. Figure 5 shows the periodic change in excitability of the auditory cortex
Figure 6 shows diagrammatically the re- lationship Chang (1951) found between reverberating waves elicited by a single con- ditioning shock applied to the auditory cor- tex and the excitability curve resulting from the responses to auditory stimuli applied in different phases of the reverberating waves. This indicates that maximal facilitation oc- curs on the rising phase of the recurrent

450 DONALD B. LINDSLEY
wave and maximal inhibition on the de- scending portion, with excitability unchang- ed at the time of the peaks and troughs.
In another s tudy Chang (1950) has shown the effect of interaction of two au- ditory stimuli (clicks), with the second delivered at varying intervals of time after the first stimulus (fig. 7). T h e single click
is at least 50 msec. does a sizeable response .to the second of the two clicks occur. The maximal evoked response to the second click will be seen to center about the time of the first recurrent wave following the first stimulus. Th is again emphasizes the cortical excitability cycle associated with recurrent cortical af ter-effects .
I1 ] n-
C~
TIME " - * Fig. 6
Diagrammatic illustration of phase relations between reverberat- ing waves and cortical excitability. Contour of shaded areas represents potentials of two consecutive reverberating waves. Sinusoidal curve above represents two cycles of periodic varia- tion of cortical excitability. Two curves drawn on same time scale. (From Chang 1941, by permission of author and publisher. )
stimulus (A) produces an evoked response with recurrent waves following at a rate approximating that of the inherent spon- taneous rhythm. In the dual presentat ions (B-I ) it will be seen that only when the
interval between the first and second clicks
Figure 8 from Chang ' s work (1952) on the visual system summarizes four types of excitability processes which exist simultan- eously. These curves were derived from the recording of cortically evoked responses in the visual cortex of the cat to series of

PSYCHOLOGICAL PHENOMENA AND EEG 451
stimuli delivered to the lateral geniculate body. Curve one shows a general process of facilitation induced by steady illumination on the eye during the course of stimulation. Curve 2 (heavy solid line) post-excitatory depression following immediately upon the evoked response. Curve 3 (light solid line) shows the periodic variation of excitability believed by Chang to accompany cortico- thalamic reverberation. Finally, curve 4 shows a waxing and waning excitability be-
respond with the more prolonged spontan- eous periodic variation of excitability rep- resented by curve 4.
Bartley (1942) has shown that a single short photic stimulus of certain durations and intensities gives rise to two flash ex- periences. It is interesting that at lower levels of illumination (see fig. 9) the duration centers about the 100 msec. point. This sug- gests that the subjective experience of the second flash may depend upon the recurrent
A
I v e , , ~ V c ,' ,~v V
° ' ~ ' T V '
' ' T ' " 'T I'
' t ' , ¢"~ \ ~ " - ~
V V m
V V m
V V v V
V X V V
V V V
V
V V - -
X, ! I I I I I I 0 50 lO0 IbO ZOO 250 300
TiME {MSEC.) Fig. 7
Compilation of nine diagrams of evoked cortical response illustrating the action of the primary response on the occurrence of the repetitive dis- charges. The vertical line represents the stimulus; the solid pyramid, the primary response; and the open triangles, the repetitive discharges. Note that the second of two sharp clicks (auditory stimuli) reaches optimal magnitude (G) at lO0 msec. corresponding to the time of appearance of the first reverberating wave following a single click (A). (From Chang 1950, by permission of author and publisher.)
lieved by Chang to be a generalized process nonspecific to the sensory cortex. From a psychological point of view there is gross evidence at least that visual experience shows fluctuations which coincide roughly with the neurophysiological data represent- ed by curves 2 and 3 during the first second following visual stimulation. These are an initial flash experience, a very brief ab- sence, and recurrence and fluctuation of the positive after-image. Fluctuations of the negative after-image would appear to c o r -
reverberation described by Chang in curve 3, above.
One additional bit of evidence is worth considering, namely, the response to visual stimulation in the case of a one year old child with an alpha rhythm of about 5 per sec. and that of an adult with an alpha rhythm of about 10 per sec. (see fig. 10). Bernard and Skoglund (1943) have sys- tematically and extensively studied the dif- ferential blocking times as a function of age, which Lindsley (1938) had described ear-

452 DONALD B. LINDSLEY
lier. It will be observed that the blockin 9 time in the one year old child was of the order of .4 sec. whereas that for the adult was about .2 sec. Tab le II shows progressive shortenin 9 of the blockin 9 time in inverse relation to the f requency of the alpha rhythm. This relationship suggests that tim- in 9 relations of the excitability of the cortex,
range is limited to 2 to 't per sec., as is the alpha rhythm in a three month old child (see table I I ) . This fact taken together with some of the precedin 9 results su99ests that the inherent or spontaneous rhythm of a given aggregate of cells has somethin 9 in common with recurrent activity initiated by sensory stimulation or by electrical stimula-
. . . . |
m 11 . ° . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
ii j
I ' " . . . . . . . . . . . . . . . . . . . .~ .
, , _ L , '~ ' '" " \ .,:" ".., . ; '
.... ."d;';; ' . . Y ".. . . /
k I o ~ z 3 4 s ~o ~s ao zs
T~ME w SEC0~0S a r T r . ONSet ~ ~ U ~ A t ~ 0 ~
Fig. 8 Diagrammatic representation of four different processes of excitability change of visual system immediately after and during illumination of retina. 1, Progressive facilitation; 2, post-excitatory depression; 3, periodic variation of excitability accompanying cortico-thalamic reverberation; and 't, slow spontaneous periodic variation of excitability. (From Chang 1952, by permission of author and publisher.)
and possibly also in either relay nuclei of the thalamus or in those nuclei of diffuse pro- jection, are such that an excitability cycle which al ternates only half as fast in a young child as in an adult will permit access to the cortex only with a longer latency of twice that in the adult. It may be possible to throw further light on this relationship from a psychological point of view, but it will be difficult in a child so youn 9 to find a suit- able mechanism of response which can be measured adequately, and which will serve in lieu of the subjective report of an adult.
Eichorn and Lindsley (1951) have shown in a youn 9 infant of about two months of age and prior t o the establishment of an occipital a lpha rhythm that it is possible by photic drivin 9 to induce a rhythm, of con- siderably lower ampli tude than the alpha rhythm which normally appears at three months of a9¢. H o w e v e r its f requency
DURATION OF FLASH Fig. 9
Conditions under which a single short photic stim- ulation gives rise to the sensation of two flashes (stippled area). Intensity of light in terms of candles per square foot. Solid lines mark boundaries of flash too weak or too short, and too intense or too long, to produce dual impression. (From Bartley 1942, by permission of author and publisher.)

PSYCHOLOGICAL PHENOMENA AND EEG 453
tion of sensory pa thways . Likewise it might seem also to bear a common timing relation- ship to the so-called recruiting phenomena first described by Morison and Dempsey (1942) and more recently studied by Starzl and Magoun (1951) .
B , ; '
i
Fig. 10 Blocking of occipital alpha rhythm by light stimulus in A, a one year old child with 5 per sec. alpha rhythm (blockin9 time, .4 sec,), and in B, an adult with a 9 to 10 per sec. alpha rhythm (blockin 9 time, .2 sec.) From Bernhard and Skoglund, 1943. Time in 1/100 sec.
Chang has diagrammatical ly illustrated some possible mechanisms of reverberat ing activity, as shown in figure 11. Because he had shown that the pr imary response of the
TABLE II ALPHA FREQUENCY. DURATION AND BLOCKING TIME
FOR VARIOUS AGE LEVELS. (From Bernhard and Skofllund 1943.)
Average Age freq. years alpha/sec.
xA 5.3 2 6.5 4 7.4 6 8.3 8 8.5
l0 8.8 12 9.3 14 9.3 16~ 9.5 25 10.2
Range alpha/see.
3.5-- 6.6 5.5--- 7.4 6.2-- 8.9 7.5-- 9.2 7.5---10.3 7.5--10.2 8.4--10.,t 8.2--11.1 8.2--I 1.1 9.0~11.0
Alpha- durat. in see.
0.189 0.154 0.135 0.121 0.118 0.11't 0.108 0.I08 0.105 0.098
Blocking time
calculated in see.
0.43 0.35 0.31 0,28 0.27 0.26 0.25 0.25 0.24 0.23
second of two click stimuli was capable of abolishing the repetitive discharges to the first stimulus (see fig. 7) he argues that the blockage of the reverberat ing waves occurs in the cortex. There fore he finds figure l lB with its separate thalamo-cort ical reverberat ing pa thways more acceptable. Jarcho (1949) on the other hands submits similar evidence which he interprets in favor of pr imary and repetitive responses sharing the same pathways , as is illustrated in figure l l A .
A B
° "Y
f - . . . _ . _ . .
I i
t l
t
THe.AMOS
I 4
6
" T " q
Fi 9. 11 Diagrams showing two possible pathways of the corticothalamic circuit. In (A) the corticipetal path- way of the reverberating circuit shares the same neurons which mediate the great afferent volleys. In (B) they do not use the same neurons. (From Chang 1950, by permission of author and publisher).
The data of Moruzzi and M a g o u n (19't9) and Starzl and Magoun (1951) dealing, respectively, with the reticular activating system and its widespread desynchronizin9 effects and the recruiting system and its confinement of responses to the associational and motor cortex to the exclusion of pr imary sensory areas (see fig. 12), raises still further questions about the nature of the i n t e r a c t i o n b e t w e e n cort ica l and subcortical centers. Assuming as D e m p s e y a n d Mori-

454 DONALD B. LINDSLEY
son (19 ' t2 ) d id tha t the recru i t in 9 r e s p o n s e is to be iden t i f i ed wi th the mechan i sm of in t r ins ic r h y t h m s of the cor tex , a n d a lso because of its op t ima l f r e q u e n c y c o r r e s p o n d - ence wi th s p o n t a n e o u s r h y t h m s , it w o u l d a p p e a r tha t some form of C h a n g ' s d i a g r a m (fi 9. l l B ) wi th mod i f i ca t ions w o u l d be n e e d e d to exp la in the i n t eg ra t i on and t iming re la t ions invo lved . T h e p ic ture is far f rom comple t e a n d is b y no means s imple, but it a p p e a r s tha t a g r e a t dea l of p r o g r e s s is bein 9 m a d e t o w a r d an u n d e r s t a n d i n 9 of the bas ic m e c h a n i s m s of con t ro l of the r h y t h m s of the cor tex .
Fig. 12
Schematic representation of direct, or classical sensory relay pathways to primary receiving areas of cortex, and of the diffuse projection system via the brain- stem reticular formation. Note the widespread cor- tical influence of the latter (Moruzzi and Magoun 1949). According to Starzl and Magoun (1951) the recruiting mechanism influences only association areas and the motor cortex, but not primary receiving areas. The brainstem activating system (reticular formation) is believed to receive collaterals from ascending sensory paths, and it is believed to involve a series of teqmental relays as illustrated.
In the l i f h t of some of these n e w e r con- cep t ions , it a p p e a r s tha t a 9ood m a n y new h y p o t h e s e s a r e n e e d e d for fu r the r e x p e r - imenta l s tudy . A s each new f indin 9 is
i n t e f r a t e d wi th the o lde r ones, and wi th n e u r o p h y s i o l o g i c a l a n d p sycho log i ca l d a t a a d v a n c i n 9 t o f e t h e r , it is to be h o p e d tha t a c loser co r re l a t ion of p sycho log i ca l phe - n o m e n a a n d the E E G will be possible .
SUMMARY
A n a t t e m p t has been m a d e to show how psyc ho log i c a l a n d n e u r o p h y s i o l o g i c a l d a t a m a y be r e l a t e d t h r o u g h t ime re la t ionsh ips . A d i s t inc t ion has been m a d e b e t w e e n r e c o r d - ab le alpha rhgthm a n d alpha actiuitq. T h e l a t t e r is a s s u m e d to be a bas ic ce l lu la r r hy thm. E v i d e n c e is s u b m i t t e d wh ich su 9- 9es ts tha t the a lpha ac t iv i ty cycle is a s so - c ia ted wi th an exc i t ab i l i t y cycle in pa r t i c u l a r a g g r e g a t e s of cells. T h e exc i t ab i l i t y cyc le is p r o p o s e d as a means of puls in 9 a n d codin 9 s e n s o r y impulses , dnd e x a m p l e s a re p r o v i d e d f rom b e h a v i o r a n d sub jec t ive expe - r ience in suppo r t of this concept .
DISCUSSION
CHARLES E. HENRY
The lateness of the hour must emphasize my brief discussion. A summary and reintegration of facts such as that Dr. Lindsley has just given us needs little annotation. The reciprocity between matters psychological and matters physiological must surely now be obvious ~ and particularly so to us who cultivate the common ground of neurophysiology with our electrical equipment.
For those who may have attempted to answer the put question as to why there have not been more vigorous and more successful attempts to elucidate hard correlations between electroenceph- alographic and psychological phenomena may I sug- gest an aspect often neglected by EEG workers. This is the difficulty in getting meaningful, useful and discriminating measures of the elusive parameters of behavior which may compare with the order of precision possible to obtain (though not without effort) in electroencephalography. We tended, there- fore, to evaluate our records against gross and too- inclusive clinical diagnostic categories.
Scansion of the literature would suggest that this early eager pursuit of the electrical will o' the wisp that keeps the lid on the id was unsuccessful. But does this follow from the fact that we have identified no patterns pathognomonic of schizophrenia or of exhibitionism ? This morning's Symposium is evidence

P S Y C H O L O G I C A L P H E N O M E N A A N D E E G 4 5 5
that this is not entirely so and that some degree of order is evolving slowly out of apparent contra- diction.
I should like to suggest an addition to Dr, Lind- sley's discussion of the significance and the control of the alpha rhythm. I am thinking of the work of Hallowell Davis and colleagues first reported (and largely neglected) some 15 years ago, and more re- cently given a fuller documentation. Their elucidation of the relationship between a strong, stable alpha rhythm and passivity, and the relationship between faster frequencies and hostile, aggressive tendencies is both clearly phrased and clearly supported by the data they presented. W e all remember the more general physiological reformulation of this thinking into the Homeostasis of Cortical Excitability pre- sented as the presidential address before this Society two years ago. These studies are highly pertinent to this particular Symposium: I wonder if we are not being unduly wary about further ventures into this complex but probably rewarding field.
Nevertheless, it would appear that as the orienta- tion of workers has become more specific their efforts have become more productive. As Dr. Lindsley has indicated, the field of perception, in particular, show- ed sudden signs of flowering when cross fertilized by cybernetics. Loop, reveratin9, circus activity has been hypothesized in such singularly attractive ex- planatory form that I am almost convinced myself. I am sure that we both agree in recommending the stimulating papers and discussions in the recently ap- pearing Hixon Symposium to your attention.
Dr. Lindsley has done an able job of pulling together a number of observations and relating them to visual phenomena, thus demonstrating the proof of his observation that the molar psychological data are helpful in elucidating the micro neurophysiological observations. It is to be hoped that his reiterated distinction between the alpha rhythm and alpha activity will lay permanently that paradoxical ghost that has so long confused us. His insight into the important implications of Chan0's work shows that it would have been as appropriately published in a psychological as a physiological journal.
Reference to the visual smearing associated with involuntary eye movements and the lack of such smearing when the neuronic shutter is clicking in good synchrony immediately suggests an experiment waiting to be carried out. While electrical stimulation of the frontal eye fields may certainly elicit eye movements I do not think a reciprocal relationship has been shown to exist between area 8 and the visual cortex. The fact that eye movements are ob- tainable on electrical stimulation of visual cortex
itself suggests that the more probable trigger for the shutter is somewhere in the geniculo-striate system. A possible complication is the additional fact that pa- tients with congenital nystagmus suffer very little visual incapacity, indicating that their alpha activity may not be critically implicated in visual perception.
In conclusion, Dr. Lindsley is to be congratulated on the photic activation he has given to some older ideas. He has sharpened our wits with new insight and suggested new experiments. His paper is thus doubly successful and it is a pleasure to thank him for it.
REFERENCES
ADRIAN, E. D. The Physical Background of Per- ception. Oxford, Clarendon Press, 194'/, 95 pp.
BARTLEY, S. H. Subjective brightness in relation to flash rate and the light dark ratio. ]. exp. Psychol., 1938, 23: 313-319.
BARTLEY, S. H. Some factors in brightness discrimina- tion. Psychol. Rev., 1939, 46: 337-358.
BARTLEY, S. H. The relation between cortical re- sponse to visual stimulation and changes in the alpha rhythm. ]. exp. Psychol., 1940, 27: 624- 639.
BARTLEY, S. H. Vision: A Study of its Basis. New York. Van Nostrand. 1941.
BARTLEY, S. H. The features of the optic-nerve discharge underlying recurrent vision. ]. exp. Psvchol., 1949., 30: 125-135.
BARTLEY, S. H. and BISHOp, G. H. The corticaI response to stimulation of the optic nerve in the rabbit. Amcr. ]. Physiol., 1933, 103: 159-172.
BERNHARD, C. G. and SKOGLLIND, C. R. On the blocking of the cortical alpha rhythm in children. Acta Ps.uchiat. Neurol., 1943, 18: 159-170.
BISHOP, G. H. Cyclic changes in excitability of the optic pathway of the rabbit. Amer. ]. Physiol., 1933, 103: 213-224.
BISHOp, G. H. The interpretation of cortical poten- tials. Cold Spr. Harb. Symp. quant. Biol., 1936, 4: 305-319.
CHANG, H.-T. The repetitive discharges of cortico- thalamic reverberating circuit. ]. Neurophysiol., 1950, 13: 235-258,
CHANG, H.-T. Changes in excitability of cerebral cortex followin~ sinqle electric shock applied to cortical surface. ]. Neurophysiol., 1951, 14: 95-112.
CHANG, H.-T. Cortical response to stimulation of lateral geniculate body and the potentiation thereof by continuous illumination of retina. ]. Neurophysiol., 1952, 15: 5-26.
DEMPSEY, E. W. and MOalSON, R. S. The produc- tion of rhythmically recurrent cortical potentials after localized thalamic stimulation. Amer. ]. Ph.vsiol., 1949., 135: 293-300.
DEMPSEY, E. W. and MORISON, R. S. The interaction of certain spontaneous and induced cortical potentials. Amer. ]. Physiol., 1942, 135: 301-308.
EICHORN, D. and LINDSLEY, D. B. Electrocortical and autonomic response in infants to visual and auditory stimuli. (Unpublished, 19§1).

456 DONALD B. LINDSLEY
JARCHO, L. W. Excitability of cortical afferent sys- tems during barbiturate anesthesia. ]. Neuro- physiol., 1949, 12: 447,457.
JASPER, H. H. Cortical excitatory state and syn- chronism in the control of bioelectric autonomous rhythms. Cold Spr. Harb. Syrup. quant. Biol., 1936, 4: 320-338.
JASPER, H. H. Electrical signs of cortical activity. Psychol. Bull., 1937, 3'/: 411-'t81.
JASPER, H. H. Electroencephalography, Chap. 14, in W. Penfield and T. C. Erickson. Epilepsy and Cerebral Localization. Springfield. Ill.. Thomas. 1941.
KREEZER, G. The electro-encephalogram and its uses in psychology. Amer. J. PsychoI., 5I: 737-759.
LINDSLEY, D. B. Electrical potentials of the brain in children and adults. ]. Gen. Psychol., 1938, 18: 285-306.
LINDSLEY, D. B. Electroencephalography. Chap. 33 in Hunt (ed.): Personality and the Behavior Disorders. New York. Ronald, 1944. Vol. II. Pp. 1033-1103.
MEISTER, R. K. A hypothesis concerning the function of the occipital alpha rhythm in vision with special reference to the perception of movement. (Unpublished doctoral dissertation, Univ. of Chicago, 1951).
MORISON, a. S. and DEMPSEY, E. W. A study of thalamo-cortical relations. Amer. ]. Physiol., 1942, 135: 281-292.
MoRuzzl, G. and MAGOUN, H. W. Brain stem reticular formation and activation of the EEG. LEG Clin. Neurophysiol., 1949, I: 455-473.
STARZL, T. E. and MAGOUN, H. W. Organization of the diffuse thalamic projection system. ]. Neurophysiol., 1951, 14: 133-146.
VERZE~,NO, M., LINDSLEY, D. B. and MAC, OUN, H. W. On the nature of the recruiting response. ]. Neurophysiol. (Submitted for publication).
WALTER, W. G. Normal rhythms -- their develop- ment. distribution and significance. Chap. VII in D. Hill and G. Parr: Electroencephalography. New York, Macmillan, 1959a.
WALTER, W. G. The twenty-fourth Maudsley lecture: The functions of electrical rhythms in the brain. J. ment. Sei., 1950b, 96: 1-31.
Reference: L1NDSLEY, D. B. Psychological phenomena and the electroencephalogram. physiol., 1952, 4: '143-456.
LEG Clin. Neuro-


















