
Diversity of stonefly hexamerins and implication for … · Diversity of stonefly hexamerins and...
11
Insect Biochemistry and Molecular Biology Insect Biochemistry and Molecular Biology 37 (2007) 1064–1074 Diversity of stonefly hexamerins and implication for the evolution of insect storage proteins Silke Hagner-Holler a , Christian Pick b , Stefan Girgenrath c , James H. Marden c , Thorsten Burmester b, a Institute of Zoology, Johannes Gutenberg University of Mainz, D-55099 Mainz, Germany b Institute of Zoology and Zoological Museum, University of Hamburg, D-20146 Hamburg, Germany c Department of Biology, Pennsylvania State University, USA Received 4 April 2007; received in revised form 30 May 2007; accepted 1 June 2007 Abstract Hexamerins are large storage proteins of insects in the 500 kDa range that evolved from the copper-containing hemocyanins. Hexamerins have been found at high concentration in the hemolymph of many insect taxa, but have remained unstudied in relatively basal taxa. To obtain more detailed insight about early hexamerin evolution, we have studied hexamerins in stoneflies (Plecoptera). Stoneflies are also the only insects for which a functional hemocyanin is known to co-occur with hexamerins in the hemolymph. Here, we identified hexamerins in five plecopteran species and obtained partial cDNA sequences from Perla marginata (Perlidae), Nemoura sp. (Nemouridae), Taeniopteryx burksi (Taeniopterygidae), Allocapnia vivipara (Capniidae), and Diamphipnopsis samali (Diamphipnoidae). At least four distinct hexamerins are present in P. marginata. The full-length cDNA of one hexamerin subunit was obtained (PmaHex1) that measures 2475 bp and translates into a native polypeptide of 702 amino acids. Phylogenetic analyses showed that the plecopteran hexamerins are monophyletic and positioned at the base of the insect hexamerin tree, probably diverging about 360 million years ago. Within the Plecoptera, distinct hexamerin types evolved before the divergence of the families. Mapping amino acid compositions onto the phylogenetic tree shows that the accumulation of aromatic amino acids (and thus the evolution of ‘‘arylphorins’’) commenced soon after the hexamerins diverged from hemocyanins, but also indicates that hexamerins with distinct amino acid compositions reflect secondary losses of aromatic amino acids. r 2007 Elsevier Ltd. All rights reserved. Keywords: Hexamerin; Molecular clock; Plecoptera; Phylogeny; Stoneflies; Storage protein 1. Introduction All insect species investigated so far harbor large proteins in their hemolymph that are referred to as hexamerins. Sequence comparisons have shown that these proteins belong to the superfamily of arthropod hemocya- nins, phenoloxidases, crustacean pseudohemocyanins and dipteran hexamerin receptors (Beintema et al., 1994; Burmester and Scheller, 1996; Burmester, 2001, 2002). Like arthropod hemocyanins, hexamerins usually consist of six identical or similar subunits in the range of 80 kDa, giving rise to native molecules of about 500 kDa (Telfer and Kunkel, 1991; Burmester, 1999). However, in contrast to the respiratory hemocyanins, hexamerins do not bind oxygen because the copper-binding histidines residues have been replaced by other amino acids (Beintema et al., 1994; Burmester, 2001, 2002). In many insect species, hexamerins accumulate in the hemolymph to extraordinarily high concentrations, reach- ing up to 50% of the total salt-extractable proteins of the animal in premolt developmental stages (Scheller et al., 1990; Telfer and Kunkel, 1991). Therefore, hexamerins are thought to act mainly as storage proteins, which are used as a source of energy and amino acids during non-feeding periods, such as pupal or nymphal stages (Telfer and Kunkel, 1991; Burmester, 1999). Hexamerins have also ARTICLE IN PRESS www.elsevier.com/locate/ibmb 0965-1748/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.ibmb.2007.06.001 Corresponding author. Tel.: +49 40 42838 3913; fax: +49 40 42838 3937. E-mail address: [email protected] (T. Burmester).
Transcript of Diversity of stonefly hexamerins and implication for … · Diversity of stonefly hexamerins and...

ARTICLE IN PRESS
InsectBiochemistry
andMolecularBiology
0965-1748/$ - se
doi:10.1016/j.ib
�Correspondfax: +4940 428
E-mail addr
Insect Biochemistry and Molecular Biology 37 (2007) 1064–1074
www.elsevier.com/locate/ibmb
Diversity of stonefly hexamerins and implication for the evolutionof insect storage proteins
Silke Hagner-Hollera, Christian Pickb, Stefan Girgenrathc,James H. Mardenc, Thorsten Burmesterb,�
aInstitute of Zoology, Johannes Gutenberg University of Mainz, D-55099 Mainz, GermanybInstitute of Zoology and Zoological Museum, University of Hamburg, D-20146 Hamburg, Germany
cDepartment of Biology, Pennsylvania State University, USA
Received 4 April 2007; received in revised form 30 May 2007; accepted 1 June 2007
Abstract
Hexamerins are large storage proteins of insects in the 500 kDa range that evolved from the copper-containing hemocyanins.
Hexamerins have been found at high concentration in the hemolymph of many insect taxa, but have remained unstudied in relatively
basal taxa. To obtain more detailed insight about early hexamerin evolution, we have studied hexamerins in stoneflies (Plecoptera).
Stoneflies are also the only insects for which a functional hemocyanin is known to co-occur with hexamerins in the hemolymph. Here, we
identified hexamerins in five plecopteran species and obtained partial cDNA sequences from Perla marginata (Perlidae), Nemoura sp.
(Nemouridae), Taeniopteryx burksi (Taeniopterygidae), Allocapnia vivipara (Capniidae), and Diamphipnopsis samali (Diamphipnoidae).
At least four distinct hexamerins are present in P. marginata. The full-length cDNA of one hexamerin subunit was obtained (PmaHex1)
that measures 2475 bp and translates into a native polypeptide of 702 amino acids. Phylogenetic analyses showed that the plecopteran
hexamerins are monophyletic and positioned at the base of the insect hexamerin tree, probably diverging about 360 million years ago.
Within the Plecoptera, distinct hexamerin types evolved before the divergence of the families. Mapping amino acid compositions onto the
phylogenetic tree shows that the accumulation of aromatic amino acids (and thus the evolution of ‘‘arylphorins’’) commenced soon after
the hexamerins diverged from hemocyanins, but also indicates that hexamerins with distinct amino acid compositions reflect secondary
losses of aromatic amino acids.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Hexamerin; Molecular clock; Plecoptera; Phylogeny; Stoneflies; Storage protein
1. Introduction
All insect species investigated so far harbor largeproteins in their hemolymph that are referred to ashexamerins. Sequence comparisons have shown that theseproteins belong to the superfamily of arthropod hemocya-nins, phenoloxidases, crustacean pseudohemocyanins anddipteran hexamerin receptors (Beintema et al., 1994;Burmester and Scheller, 1996; Burmester, 2001, 2002).Like arthropod hemocyanins, hexamerins usually consistof six identical or similar subunits in the range of 80 kDa,
e front matter r 2007 Elsevier Ltd. All rights reserved.
mb.2007.06.001
ing author. Tel.: +4940 42838 3913;
38 3937.
ess: [email protected] (T. Burmester).
giving rise to native molecules of about 500 kDa (Telferand Kunkel, 1991; Burmester, 1999). However, in contrastto the respiratory hemocyanins, hexamerins do not bindoxygen because the copper-binding histidines residues havebeen replaced by other amino acids (Beintema et al., 1994;Burmester, 2001, 2002).In many insect species, hexamerins accumulate in the
hemolymph to extraordinarily high concentrations, reach-ing up to 50% of the total salt-extractable proteins of theanimal in premolt developmental stages (Scheller et al.,1990; Telfer and Kunkel, 1991). Therefore, hexamerins arethought to act mainly as storage proteins, which are usedas a source of energy and amino acids during non-feedingperiods, such as pupal or nymphal stages (Telfer andKunkel, 1991; Burmester, 1999). Hexamerins have also

ARTICLE IN PRESSS. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–1074 1065
been identified as constituents of the sclerotizing system ofthe insect’s cuticle (Peter and Scheller, 1991). They mayalso serve as carriers for ecdysteroids (Enderle et al., 1983),juvenile hormone (Braun and Wyatt, 1996) and otherorganic compounds such as riboflavin (Magee et al., 1994).More recent studies suggest that hexamerins function incaste differentiation of termites via regulation of thejuvenile hormone levels (Zhou et al., 2006, 2007). Thereis also evidence that some hexamerins play a role in theinsect’s humoral immune response (Phipps et al., 1994;Beresford et al., 1997). Molecular phylogenetic analyseshave shown a complex pattern of hexamerin evolution insome insect orders, with several hexamerin subtypes, suchas the highly aromatic arylphorins or methionine-richhexamerins occurring in parallel in a single species(Burmester et al., 1998; Burmester, 1999, 2001). Hexamerinsequences have been successfully used as markers for theinference of insect phylogeny (Burmester et al., 1998;Burmester, 1999).
Although hexamerins are probably ubiquitously presentin insects, most information derives from investigations ofdipteran and lepidopteran species, whereas data from othertaxa are scarce (for a review see Telfer and Kunkel, 1991;Burmester, 1999). Here, we investigate for the first time thehexamerins from Plecoptera (stoneflies). Stoneflies arethought to have arisen near the base of the winged insectsand have retained several ancient morphological andbehavioral features (Hennig, 1969, 1981). Recently, wehave demonstrated that the stonefly Perla marginata
harbors a functional hemocyanin in its hemolymph(Hagner-Holler et al., 2004). Thus, the Plecoptera are theonly known insect order which possesses both types ofhexameric proteins in their hemolymph. This observationprompted us to investigate the occurrence and evolution ofhexamerins in selected stonefly species.
2. Material and methods
2.1. Analyses of hemolymph proteins
Hemolymph was withdrawn from the dorsal abdomenof living P. marginata nymphs (1–3 years) and adults bythe aid of a syringe, and immediately diluted with anapproximately equal volume of 100mM Tris–HCl, pH 7.5,10mM Ca2+, 10mM Mg2+. Hemocytes and cell debriswere removed by 10min centrifugation at 10,000� g at4 1C. The total protein concentration in the hemolymphwas determined according to the method of Bradford(1976). Denaturing SDS-PAGE was performed on 7.5%gels according to standard procedures. After electrophor-esis, the gel was fixed in ethanol/acetic acid and stainedwith 0.1% Coomassie Brilliant Blue R-250.
2.2. RT-PCR cloning of stonefly hexamerins
Total RNA was extracted from adult and larvalP. marginata, Nemoura sp., Taeniopteryx burksi, Allocapnia
vivipara and Diamphipnopsis samali employing the guani-dine-thiocyanate method (Chirgwin et al., 1979) or the ureaprocedure according to Holmes and Bonner (1973). Wedesigned a set of degenerated oligonucleotide primersaccording to conserved amino acid sequences of insecthexamerins and hemocyanins (Burmester, 2001). The for-ward primers were: 50-GAGGGNSAGTTCGTNTACGC-30,50-CCNCCNCCNTAYGARRTCTACCC-30; reverse pri-mers: 50-GAANGGYTTGTGGTTNAGRCG-30, 50-TCG-TACTTGGGTCCNAGGAAGAC-30. Reverse-transcription(RT)-PCR experiments were carried out applying theOneStep-kit according to the manufacturer’s instructions(Qiagen, Hilden, Germany). PCR-fragments of theexpected size were cloned into the pCR4-TOPOs-TAor pGem-T Easy cloning vectors, and sequenced bi-directionally by a commercial service (GENTERPRISE,Mainz, Germany) or on-site using an ABI Prism 377 DNAsequencer. Additional 30 sequence of T. burksi andD. samali hexamerin was obtained by the 30 rapidamplification of cDNA ends (RACE) method using RACEprimers supplied with a kit (Roche, Indianapolis, IN, USA)in combination with gene specific PCR primers.
2.3. Molecular cloning of P. marginata hexamerin cDNA
A cDNA expression library from 3 years old P. marginata
nymphs was constructed as previously described employingthe STRATAGENE l-ZAP system (Hagner-Holler et al.,2004). PCR-fragments of various P. marginata hexamerinswere labeled with digoxigenin (Roche PCR labeling kit) andused as probes to screen the cDNA library. Positive phageclones were converted to pBK-CMV plasmid vectors usingthe material provided by STRATAGENE, and sequencedon both strands as described (GENTERPRISE, Mainz).
2.4. Sequence analyses
Sequences were assembled with the Vector NTI 10program (Invitrogen) or with the software packageLasergeneTM (DNA STAR, Madison, WI, USA). Thetools provided by the ExPASy Molecular Biology Serverof the Swiss Institute of Bioinformatics (http://www.expasy.org) were used for the analyses of DNA and aminoacid sequences. Signal peptides were predicted using theonline version of SignalP V1.1 (Nielsen et al., 1997). Analignment of the stonefly hexamerins was constructed byhand using GeneDoc 2.6 (Nicholas and Nicholas, 1997).Statistical analysis was carried out with ANOVA, asimplemented in the Microsoft EXCEL 2003 spreadsheetprogram. The Plecoptera sequences were added to analignment of insect hexamerins and hemocyanins, as it hasbeen used in previous studies (Burmester et al., 1998;Burmester, 1999). More recently available sequences havebeen added using ClustalX (Thompson et al., 1997) andmanually adjusted with GeneDoc. Except for the plecop-teran hexamerins, incomplete sequences were excludedfrom the analyses. A list of sequences used in this study is

ARTICLE IN PRESSS. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–10741066
provided in Supplemental Table 1. Amino acid composi-tions were deduced from the translated cDNA sequences.
2.5. Molecular phylogeny
The program packages PHYLIP 3.6b (Felsenstein,2004), TREE-PUZZLE 5.2 (Strimmer and von Haeseler,1997) and MrBayes 3.1 (Huelsenbeck and Ronquist, 2001)were used for the phylogenetic tree reconstructions.Distance matrices were calculated with TREE-PUZZLEusing the WAG (Whelan and Goldman, 2001) model ofamino acid evolution, assuming gamma distributions ofsubstitution rates with eight categories. Neighbor-joiningtrees were inferred with the program NEIGHBOR fromthe PHYLIP package. The reliability of the branchingpattern was tested by bootstrap analysis (Felsenstein, 1985)with 100 replications, employing the PUZZLEBOOT shellscript (M. Holder and A. Roger). Bayesian phylogeneticanalyses were performed by MrBayes, using the WAGmodel and assuming a gamma distribution of substitutionrates. Prior probabilities for all trees were equal. Metro-polis-coupled Markov chain Monte Carlo (MCMCMC)sampling was performed with one cold and three heatedchains that were run for 1,000,000 generations. Startingtrees were random, trees were sampled every 100thgeneration and posterior probabilities were estimated onthe final 2000 trees (burnin ¼ 8000).
2.6. Molecular clock estimates
Linearized trees were calculated under the assumption ofa relaxed molecular clock employing the program R8S 1.71(Sanderson, 1997, 2002). We applied the Langley–Fitchmaximum likelihood method (Langley and Fitch, 1974)with the Powell algorithm to estimate divergence times.The tree was calibrated with fossils constraints (Kukalova-Peck, 1991; Ross and Jarzembowski, 1993). Stratigraphicinformation was obtained from http://www.fossilrecord.net (Benton and Donoghue, 2007); numerical ages derivedfrom the ‘‘International Stratigraphic Chart’’ (Gradstein
Fig. 1. Coomassie-stained SDS-gel with hemolymph from Perla marginata. A
(1–3) and adult stoneflies were applied. The molecular mass standard is given
et al., 2004). We assumed that brachyceran and nemato-ceran Diptera diverged between 238.5 and 295.4 millionyears ago (MYA), Diptera and Lepidoptera between 281.5and 307.2MYA, Hymenoptera and Mecopteria (i.e.,Diptera+Lepidoptera) more than 299.0MYA, Isopteraand Blattaria more than 145.5MYA, and that the firstHexapoda appeared at least 407.0MYA.
3. Results
3.1. Plecopteran hexamerins
Hemolymph samples from 1- to 3-year-old larval andadult specimens of P. marginata were withdrawn andanalyzed by SDS–PAGE. In all four stages, we observedseveral prominent bands in the range of 75–80 kDa, whichis the typical mass of arthropod hemocyanins and insecthexamerins (Telfer and Kunkel, 1991; Burmester, 1999,2001) (Fig. 1). The two lower bands at about 75 kDa werepreviously identified as hemocyanins by using specificantibodies in Western blotting (Hagner-Holler et al., 2004).In 3-years-old larvae and in adult stoneflies, multiple(about four) additional bands with higher molecularmasses (about 80 kDa) appear, which we suspect to behexamerins. Due to the lack of a specific antibody, wecannot conclusively evaluate this assumption. The mole-cular evidence presented below, however, demonstrates thepresence of multiple hexamerin chains.
3.2. Molecular cloning and analyses of P. marginata
hexamerins
We used an alignment of insect hexamerins and hemo-cyanins to deduce degenerate oligonucleotide primers.These primers were applied in reverse transcription-PCRon total RNA from 3-years-old nymphs and adults ofP. marginata, which resulted in several cDNA products ofabout 1000bp lengths. Cloning and sequencing of thecDNAs revealed that these fragments represent four diffe-rent hexamerin sequences, which we named PmaHex1–4.
bout 10 mg of total hemolymph proteins from 1, 2- and 3-year-old larvae
on the left side; the arrow denotes the position of the hexamerins.

ARTICLE IN PRESSS. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–1074 1067
A cDNA library had been constructed from poly(A)+
RNA isolated from 3 years old P. marginata nymphs(Hagner-Holler et al., 2004). By screening about600,000 pfu with four different probes that are derivedfrom these sequences, we obtained one full-length clone(PmaHex1) and two 50 incomplete clones (PmaHex3and PmaHex4). PmaHex2 could not be identified in thelibrary.
The cDNA sequence of PmaHex1 comprises a total of2475 bp and covers an open reading frame of 2109 bp,beginning with a ATG at bp 13 (Supplemental Fig 1).The translation stop codon (TAA) is present at positionbp 2119 and is followed by a polyadenylation signal(AATAAA) at bp 2441 and poly(A) tail of 18 bp. Theincomplete sequences of PmaHex3 and PmaHex4 comprise1899 and 1847 bp, respectively. By comparison withPmaHex1, we inferred that 408 and 308 bp of the 50 codingends are missing in PmaHex3 and Hex4 (SupplementalFig 2). The partial PmaHex2 sequence covers 1086 bp ofthe middle region, and misses �450 bp of the 50 and�600 bp of the 30 coding regions. Within the overlappingregions, the P. marginata sequences are 65.8–87.6%identical on the DNA level and 54.0–83.1% identical onthe amino acid level (Table 1).
The deduced primary structure of PmaHex1 yields apolypeptide of 702 amino acids with a calculated molecularmass of 82.52 kDa. Computer analysis of the sequencerevealed the presence of a typical 16 amino acid signalpeptide necessary for transmembrane transport and exportinto the hemolymph (von Heijne, 1986). Therefore, thenative secreted PmaHex1 subunit comprises 686 aminoacids with a mass of 80.95 kDa (pI 5.68), which isin excellent agreement with the data obtained fromSDS-PAGE (Fig. 1). The six copper-binding sites essentialfor the function in O2 binding are not conserved inPmaHex1, although two of the six histidines have beenmaintained (Fig. 2). A potential N-glycosylation site(NXS/T) is present at Asn 207. The PmaHex1 amino acidsequence was used to search the databases employing theBLAST algorithm. Hexamerin 2 (RflHex2; accessionno. AY572859) from the termite Reticulitermes flavipes
(Scharf et al., 2005) was found to be the closest relative ofPmaHex1 (Fig. 2). Both hexamerins were clearly distinctfrom crustacean or insect hemocyanins, as exemplified bythe absence of the copper-binding histidines.
Table 1
Comparison of P. marginata hexamerins
PmaHex1 PmaHex2 PmaHex3 PmaHex4
PmaHex1 87.6 65.8 73.9
PmaHex2 83.1 69.1 75.2
PmaHex3 60.4 58.6 71.9
PmaHex4 67.4 67.7 54.0
Percent identities between hexamerins were calculated from nucleotide
(above diagonal) and amino acid sequences (below).
3.3. Identification of hexamerins in other plecopteran species
Using various combinations of degenerate primers,we obtained partial sequences from four additionalplecopteran species: Nemoura sp. (Nemouridae), T. burksi
(Taeniopterygidae), A. vivipara (Capniidae), and D. samali
(Diamphipnoidae). They represent two other major super-families of the Plecoptera. While P. marginata belongs tothe Perloidea, Nemoura sp., T. burksi and A. vivipara aremembers of the Nemouroidea, and D. samali belongs to theEusthenioidea. Three distinct hexamerins were obtainedfrom Nemoura sp., and one for each T. burksi, A. vivipara,and D. samali (Supplemental Fig 2). The cDNA sequencesmeasured 841–1931 bp and translate into polypeptidefragments from 279 to 567 amino acids. Comparisons ofthe amino acid sequences showed a high degree of identity.The closest relatives among hexamerins from differentspecies were the proteins from T. burksi (TbuHex1) andA. vivipara (AviHex1), which display 96.2% identity on thenucleotide and 95.3% on the amino acid level. Withinspecies, hexamerins 1 and 3 from Nemoura sp. (NspHex1and 3) were found closely related, with 97.4% identity onthe nucleotide and 97.5% on the amino acid level.
3.4. Phylogeny of insect hexamerins
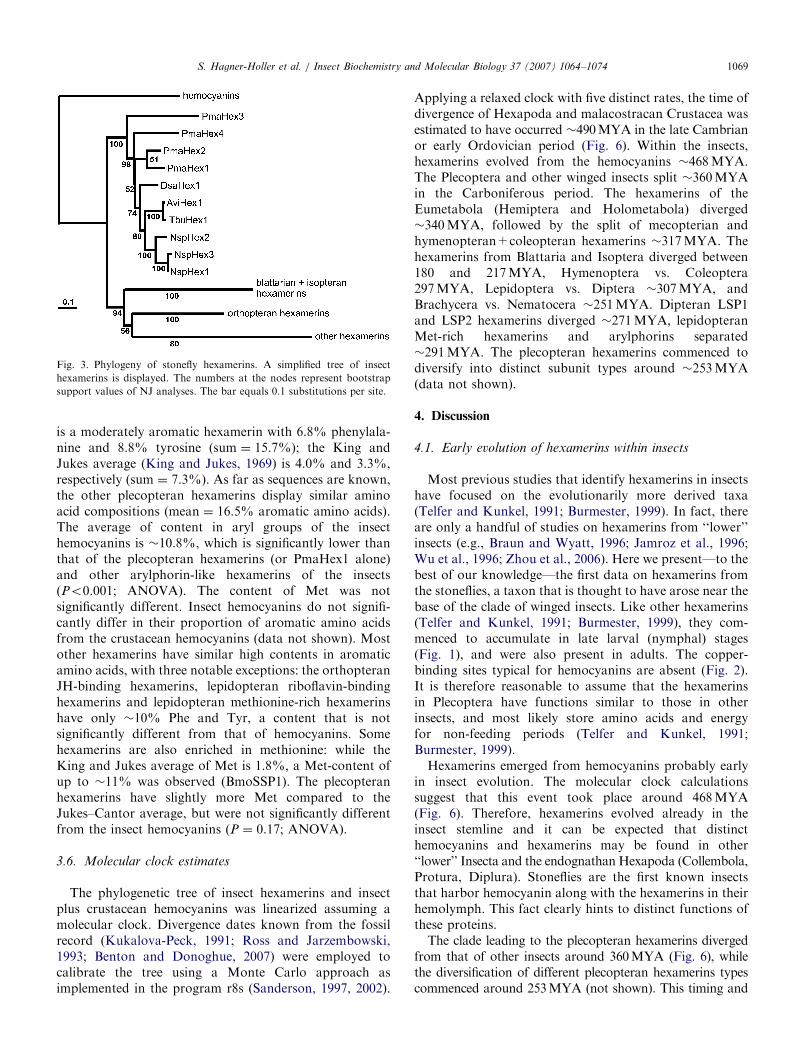
The hexamerin sequences of the Plecoptera wereincluded in an alignment of insect hemocyanins andselected other hexamerins (Supplemental Table 1). Weomitted hexamerin sequences carrying long insertions orfor which only partial database entries were available. Wefurther excluded several sequences of the dipteran LSP-1and LSP-2 types, for which multiple similar entries fromclosely related species were available. The final amino acidalignment includes four crustacean hemocyanins, fiveinsect hemocyanins, 10 plecopteran and 67 other hexamer-ins. Phylogenetic trees were constructed employing Baye-sian inference and the neighbor-joining (NJ) method. Bothanalyses show that the plecopteran hexamerins form a well-supported monophyletic clade (100% bootstrap value; 1.0Bayesian posterior probability) (Figs. 3 and 4). Within thePlecoptera, PmaHex3 was found at the basal mostposition; however, the hexamerins of P. marginata wereparaphyletic, with the clade leading to PmaHex1 and 2being closer to the other plecopteran hexamerins thanPmaHex3 or 4 (Fig. 3). The three hexamerins fromNemoura sp. were monophyletic, and also the hexamerinsfrom the Nemouroidea (Nemoura sp., T. burksi andA. vivipara) formed a monophylum.In neighbor-joining analysis, the plecopteran hexamerins
were found to be in sistergroup position to all other insecthexamerins. This relationship is well supported (94%bootstrap value in NJ analysis; Fig. 3). In Bayesiananalysis, the plecopteran hexamerins were found assistergroup of the cockroach and termite hexamerins (datanot shown). The analyses were rerun after exclusion ofthe nine incomplete plecopteran hexamerin sequences.

ARTICLE IN PRESS
Fig. 2. Multiple alignment of selected hexamerin and hemocyanin sequences. P. marginata hexamerin 1 (PmaHex1) was compared with hemocyanin
subunit a from the spiny lobster Panulirus interruptus (PinHcA, accession no. P04254), hemocyanin subunit 1 from P. marginata (PmaHc1; AJ555403) and
the hexamerin from the termite R. flavipes (RflHex2; AY572858). The copper-binding histidines are shaded in black, other strictly conserved residues are
shaded in gray. The signal peptides are underlined, the secondary structure of PinHcA is given in the lower row.
S. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–10741068
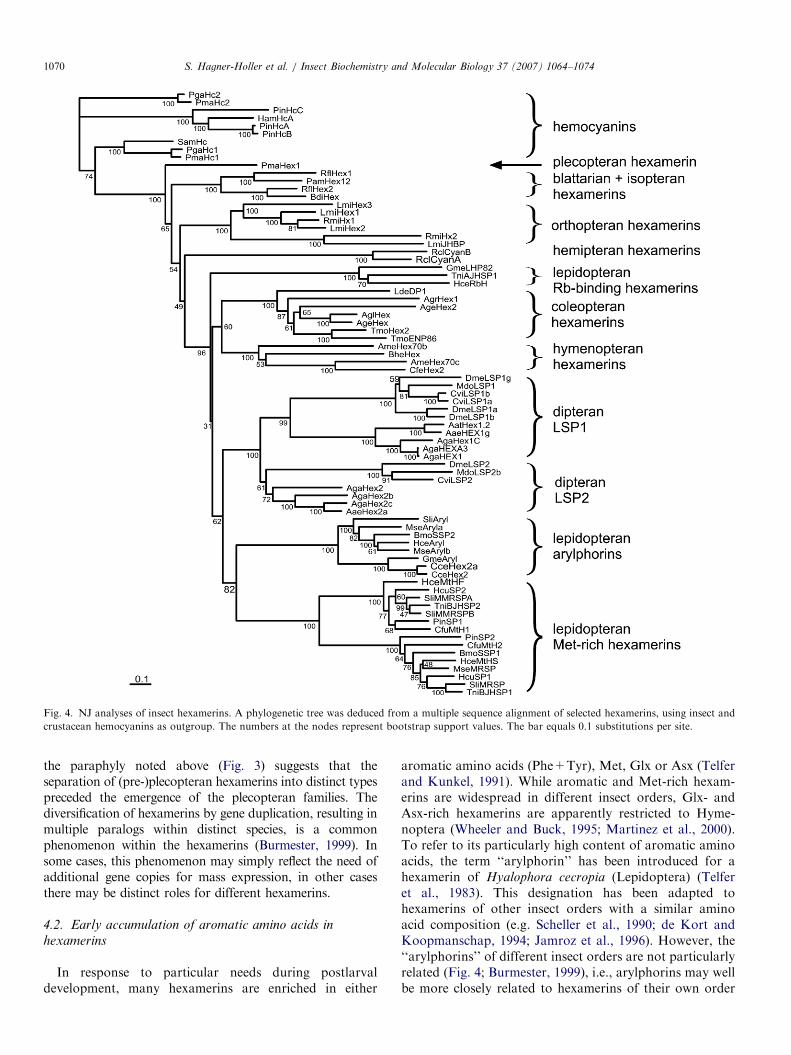
The basal position of the PmaHex1 was recovered, butthis topology does not reach the significance levels (65%bootstrap support in NJ analysis; Fig. 4). The otherhexamerins display an essentially identical tree topology asthose published before (Burmester, 1999, 2001): thehexamerins from Diptera (LSP1 and LSP2) were mono-phyletic, as well as the methionine-rich and aromatichexamerins (arylphorins) from the Lepidoptera. Theposition of the lepidopteran riboflavin (Rb)-bindinghexamerins was uncertain: they were found either insistergroup position to the other hexamerins from Holo-metabola (NJ; Fig. 4) or form a common clade with theother lepidopteran and the dipteran hexamerins (Bayesiananalyses; data not shown). Coleopteran and hymenopteranhexamerins form a reasonably well-supported monophy-
lum, which is basal to the mecopterian hexamerins. Thehemipteran hexamerins were sister to all hexamerins fromHolometabola. The recently identified hexamerins of thetermites (Isoptera) (Scharf et al., 2005) join the cockroach(Blattaria) hexamerins, but were not monophyletic. R. flavipes
hexamerin 1 (RflHex1) groups with the Periplaneta
americana hexamerin (PamHex12), whereas R. flavipes
hexamerin 2 (RflHex2) joins the Blaberus discoidalis
hexamerin (BdiHex).
3.5. Analyses of amino acid compositions
The proportions of the aryl-groups (Phe and Tyr) and ofMet were deduced from the cDNA sequences for each ofthe insect hemocyanins and hexamerins (Fig. 5). PmaHex1
ARTICLE IN PRESS
Fig. 3. Phylogeny of stonefly hexamerins. A simplified tree of insect
hexamerins is displayed. The numbers at the nodes represent bootstrap
support values of NJ analyses. The bar equals 0.1 substitutions per site.
S. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–1074 1069
is a moderately aromatic hexamerin with 6.8% phenylala-nine and 8.8% tyrosine (sum ¼ 15.7%); the King andJukes average (King and Jukes, 1969) is 4.0% and 3.3%,respectively (sum ¼ 7.3%). As far as sequences are known,the other plecopteran hexamerins display similar aminoacid compositions (mean ¼ 16.5% aromatic amino acids).The average of content in aryl groups of the insecthemocyanins is �10.8%, which is significantly lower thanthat of the plecopteran hexamerins (or PmaHex1 alone)and other arylphorin-like hexamerins of the insects(Po0.001; ANOVA). The content of Met was notsignificantly different. Insect hemocyanins do not signifi-cantly differ in their proportion of aromatic amino acidsfrom the crustacean hemocyanins (data not shown). Mostother hexamerins have similar high contents in aromaticamino acids, with three notable exceptions: the orthopteranJH-binding hexamerins, lepidopteran riboflavin-bindinghexamerins and lepidopteran methionine-rich hexamerinshave only �10% Phe and Tyr, a content that is notsignificantly different from that of hemocyanins. Somehexamerins are also enriched in methionine: while theKing and Jukes average of Met is 1.8%, a Met-content ofup to �11% was observed (BmoSSP1). The plecopteranhexamerins have slightly more Met compared to theJukes–Cantor average, but were not significantly differentfrom the insect hemocyanins (P ¼ 0.17; ANOVA).
3.6. Molecular clock estimates
The phylogenetic tree of insect hexamerins and insectplus crustacean hemocyanins was linearized assuming amolecular clock. Divergence dates known from the fossilrecord (Kukalova-Peck, 1991; Ross and Jarzembowski,1993; Benton and Donoghue, 2007) were employed tocalibrate the tree using a Monte Carlo approach asimplemented in the program r8s (Sanderson, 1997, 2002).
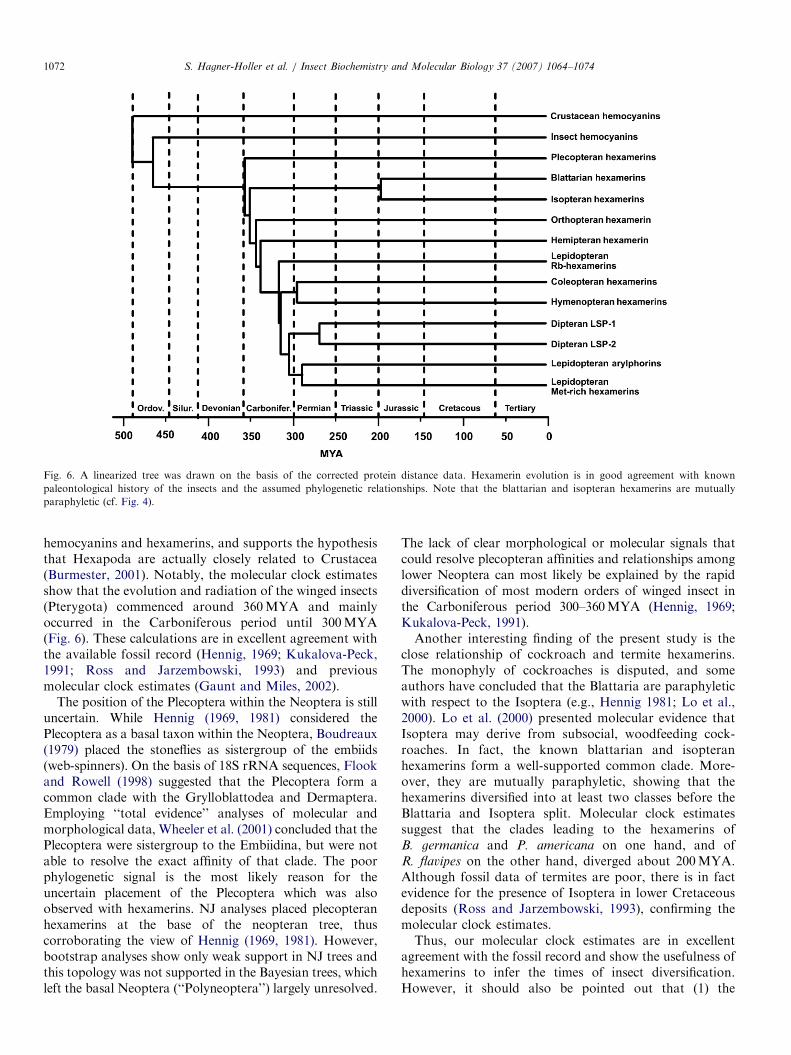
Applying a relaxed clock with five distinct rates, the time ofdivergence of Hexapoda and malacostracan Crustacea wasestimated to have occurred �490MYA in the late Cambrianor early Ordovician period (Fig. 6). Within the insects,hexamerins evolved from the hemocyanins �468MYA.The Plecoptera and other winged insects split �360MYAin the Carboniferous period. The hexamerins of theEumetabola (Hemiptera and Holometabola) diverged�340MYA, followed by the split of mecopterian andhymenopteran+coleopteran hexamerins �317MYA. Thehexamerins from Blattaria and Isoptera diverged between180 and 217MYA, Hymenoptera vs. Coleoptera297MYA, Lepidoptera vs. Diptera �307MYA, andBrachycera vs. Nematocera �251MYA. Dipteran LSP1and LSP2 hexamerins diverged �271MYA, lepidopteranMet-rich hexamerins and arylphorins separated�291MYA. The plecopteran hexamerins commenced todiversify into distinct subunit types around �253MYA(data not shown).
4. Discussion
4.1. Early evolution of hexamerins within insects
Most previous studies that identify hexamerins in insectshave focused on the evolutionarily more derived taxa(Telfer and Kunkel, 1991; Burmester, 1999). In fact, thereare only a handful of studies on hexamerins from ‘‘lower’’insects (e.g., Braun and Wyatt, 1996; Jamroz et al., 1996;Wu et al., 1996; Zhou et al., 2006). Here we present—to thebest of our knowledge—the first data on hexamerins fromthe stoneflies, a taxon that is thought to have arose near thebase of the clade of winged insects. Like other hexamerins(Telfer and Kunkel, 1991; Burmester, 1999), they com-menced to accumulate in late larval (nymphal) stages(Fig. 1), and were also present in adults. The copper-binding sites typical for hemocyanins are absent (Fig. 2).It is therefore reasonable to assume that the hexamerinsin Plecoptera have functions similar to those in otherinsects, and most likely store amino acids and energyfor non-feeding periods (Telfer and Kunkel, 1991;Burmester, 1999).Hexamerins emerged from hemocyanins probably early
in insect evolution. The molecular clock calculationssuggest that this event took place around 468MYA(Fig. 6). Therefore, hexamerins evolved already in theinsect stemline and it can be expected that distincthemocyanins and hexamerins may be found in other‘‘lower’’ Insecta and the endognathan Hexapoda (Collembola,Protura, Diplura). Stoneflies are the first known insectsthat harbor hemocyanin along with the hexamerins in theirhemolymph. This fact clearly hints to distinct functions ofthese proteins.The clade leading to the plecopteran hexamerins diverged
from that of other insects around 360MYA (Fig. 6), whilethe diversification of different plecopteran hexamerins typescommenced around 253MYA (not shown). This timing and
ARTICLE IN PRESS
Fig. 4. NJ analyses of insect hexamerins. A phylogenetic tree was deduced from a multiple sequence alignment of selected hexamerins, using insect and
crustacean hemocyanins as outgroup. The numbers at the nodes represent bootstrap support values. The bar equals 0.1 substitutions per site.
S. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–10741070
the paraphyly noted above (Fig. 3) suggests that theseparation of (pre-)plecopteran hexamerins into distinct typespreceded the emergence of the plecopteran families. Thediversification of hexamerins by gene duplication, resulting inmultiple paralogs within distinct species, is a commonphenomenon within the hexamerins (Burmester, 1999). Insome cases, this phenomenon may simply reflect the need ofadditional gene copies for mass expression, in other casesthere may be distinct roles for different hexamerins.
4.2. Early accumulation of aromatic amino acids in
hexamerins
In response to particular needs during postlarvaldevelopment, many hexamerins are enriched in either
aromatic amino acids (Phe+Tyr), Met, Glx or Asx (Telferand Kunkel, 1991). While aromatic and Met-rich hexam-erins are widespread in different insect orders, Glx- andAsx-rich hexamerins are apparently restricted to Hyme-noptera (Wheeler and Buck, 1995; Martinez et al., 2000).To refer to its particularly high content of aromatic aminoacids, the term ‘‘arylphorin’’ has been introduced for ahexamerin of Hyalophora cecropia (Lepidoptera) (Telferet al., 1983). This designation has been adapted tohexamerins of other insect orders with a similar aminoacid composition (e.g. Scheller et al., 1990; de Kort andKoopmanschap, 1994; Jamroz et al., 1996). However, the‘‘arylphorins’’ of different insect orders are not particularlyrelated (Fig. 4; Burmester, 1999), i.e., arylphorins may wellbe more closely related to hexamerins of their own order

ARTICLE IN PRESS
Fig. 5. Evolution of amino acid compositions of insect hexamerins. A simplified phylogenetic tree of insect hexamerins is displayed. The mean relative
contents (in percent) of aromatic amino acids (tyrosine and phenylalanine; first number at the branch and first column of chart) and of methionine (second
number at branch and second column) are mapped to the tree. The bar equals 0.1 substitutions per site.
S. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–1074 1071
with lower content in aromatic amino acids than toarylphorins of other orders.
To address the question whether aromatic hexamerinsevolved multiple times independently or whether this is aplesiomorphic character of the hexamerins, we mapped therelative content in Phe+Tyr and Met on the phylogenetictree (Fig. 5). The plecopteran hexamerins were found tohave high content in aromatic amino acids. Thus, theaccumulation of aromatic amino acids, probably reflectingthe need of building blocks for the formation of the cuticle(Scheller et al., 1990; Burmester, 1999), appears to haveoccurred in a very early stage of the evolution ofhexamerins. Therefore, the enrichment in aromatic aminoacids should be considered as plesiomorphic character ofthese proteins.
Hexamerins with lower content in Phe+Tyr most likelyreflect other requirements, e.g., in the higher extantLepidoptera, two distinct types of Met-rich hexamerins(averages 4.2% and 7.5% Met) have been identified (Ryanet al., 1985; Tojo and Yoshiga, 1994). Met-rich hexamerinsare more abundant in the female than in the male, and it
has been assumed that they support female reproductionand egg development by enhancing the pool of sulphur-containing amino acids at the time of vitellogenesis (Panand Telfer, 1996). Likewise, the juvenile-hormone bindinghexamerins of Orthoptera (cf. Braun and Wyatt, 1996)have a comparatively low content of Phe+Tyr (10.2%),but are rich in Met (4.5%). The phylogenetic tree impliesthat these proteins have evolved independently fromhighly aromatic hexamerins. Thus, the losses of aromaticamino acids most likely reflect secondary evolutionaryevents (Fig. 5).
4.3. Implications for insect phylogeny
Hexamerins have been successfully applied to estimateevolutionary patterns and divergence times of insects(Burmester et al., 1998; Burmester, 2001). Assuming arelaxed molecular clock with multiple substitution ratesand fossil constraints, we calculated that Hexapoda andmalacostracan Crustacea diverged around 490MYA. Thisis in good agreement with previous estimates using
ARTICLE IN PRESS
Fig. 6. A linearized tree was drawn on the basis of the corrected protein distance data. Hexamerin evolution is in good agreement with known
paleontological history of the insects and the assumed phylogenetic relationships. Note that the blattarian and isopteran hexamerins are mutually
paraphyletic (cf. Fig. 4).
S. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–10741072
hemocyanins and hexamerins, and supports the hypothesisthat Hexapoda are actually closely related to Crustacea(Burmester, 2001). Notably, the molecular clock estimatesshow that the evolution and radiation of the winged insects(Pterygota) commenced around 360MYA and mainlyoccurred in the Carboniferous period until 300MYA(Fig. 6). These calculations are in excellent agreement withthe available fossil record (Hennig, 1969; Kukalova-Peck,1991; Ross and Jarzembowski, 1993) and previousmolecular clock estimates (Gaunt and Miles, 2002).
The position of the Plecoptera within the Neoptera is stilluncertain. While Hennig (1969, 1981) considered thePlecoptera as a basal taxon within the Neoptera, Boudreaux(1979) placed the stoneflies as sistergroup of the embiids(web-spinners). On the basis of 18S rRNA sequences, Flookand Rowell (1998) suggested that the Plecoptera form acommon clade with the Grylloblattodea and Dermaptera.Employing ‘‘total evidence’’ analyses of molecular andmorphological data, Wheeler et al. (2001) concluded that thePlecoptera were sistergroup to the Embiidina, but were notable to resolve the exact affinity of that clade. The poorphylogenetic signal is the most likely reason for theuncertain placement of the Plecoptera which was alsoobserved with hexamerins. NJ analyses placed plecopteranhexamerins at the base of the neopteran tree, thuscorroborating the view of Hennig (1969, 1981). However,bootstrap analyses show only weak support in NJ trees andthis topology was not supported in the Bayesian trees, whichleft the basal Neoptera (‘‘Polyneoptera’’) largely unresolved.
The lack of clear morphological or molecular signals thatcould resolve plecopteran affinities and relationships amonglower Neoptera can most likely be explained by the rapiddiversification of most modern orders of winged insect inthe Carboniferous period 300–360MYA (Hennig, 1969;Kukalova-Peck, 1991).Another interesting finding of the present study is the
close relationship of cockroach and termite hexamerins.The monophyly of cockroaches is disputed, and someauthors have concluded that the Blattaria are paraphyleticwith respect to the Isoptera (e.g., Hennig 1981; Lo et al.,2000). Lo et al. (2000) presented molecular evidence thatIsoptera may derive from subsocial, woodfeeding cock-roaches. In fact, the known blattarian and isopteranhexamerins form a well-supported common clade. More-over, they are mutually paraphyletic, showing that thehexamerins diversified into at least two classes before theBlattaria and Isoptera split. Molecular clock estimatessuggest that the clades leading to the hexamerins ofB. germanica and P. americana on one hand, and ofR. flavipes on the other hand, diverged about 200MYA.Although fossil data of termites are poor, there is in factevidence for the presence of Isoptera in lower Cretaceousdeposits (Ross and Jarzembowski, 1993), confirming themolecular clock estimates.Thus, our molecular clock estimates are in excellent
agreement with the fossil record and show the usefulness ofhexamerins to infer the times of insect diversification.However, it should also be pointed out that (1) the

ARTICLE IN PRESSS. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–1074 1073
orthology of various hexamerins is not always thoroughlyestablished and that (2) molecular clock estimates usuallyhave large standard errors. Therefore, all our calculationsshould only be considered as rough estimates rather than asexact dates.
Acknowledgments
We thank Klaus Enting and Rainer Rupprecht (Mainz)for P. marginata specimens, and Falko Roeding andRobert Schopflin for the help with the implementationof the r8s program. This work has been supported bygrants of the Deutsche Forschungsgemeinschaft (Bu956/5;Bu956/9) and by NSF grant IBN-9722196. The nucleotidesequences reported in this paper have been submitted tothe EMBL/GenBankTM Databases under the accessionnos. AM690365 to AM690371, EF620538, EF617597 andEF617598.
Appendix A. Supplementary data
Supplementary data associated with this article can befound in the online version at doi:10.1016/j.ibmb.2007.06.001.
References
Beintema, J.J., Stam, W.T., Hazes, B., Smidt, M.P., 1994. Evolution of
arthropod hemocyanins and insect storage proteins (hexamerins). Mol.
Biol. Evol. 11, 493–503.
Benton, M.J., Donoghue, P.C.J., 2007. Paleontological evidence to date
the tree of life. Mol. Biol. Evol. 24, 26–53.
Beresford, P.J., Basinski-Gray, J.M., Chiu, J.K., Chadwick, J.S.,
Aston, W.P., 1997. Characterization of hemolytic and cytotoxic
Gallysins: a relationship with arylphorins. Dev. Comp. Immunol. 21,
253–266.
Boudreaux, H.B., 1979. Arthropod Phylogeny with Special Reference
Process to Insects. Wiley, New York.
Bradford, M.M., 1976. A rapid and sensitive method for quantification of
microgram quantities of proteins using the principle of protein dye
binding. Anal. Biochem. 72, 248–254.
Braun, R.P., Wyatt, G.R., 1996. Sequence of the juvenile hormone
binding protein from the hemolymph of Locusta migratoria. J. Biol.
Chem. 271, 31756–31762.
Burmester, T., 1999. Evolution and function of the insect hexamerins. Eur.
J. Entomol. 96, 213–225.
Burmester, T., 2001. Molecular evolution of the arthropod hemocyanin
superfamily. Mol. Biol. Evol. 18, 184–195.
Burmester, T., 2002. Origin and evolution of arthropod hemocyanins and
related proteins. J. Comp. Physiol. B 172, 95–117.
Burmester, T., Scheller, K., 1996. Common origin of arthropod
tyrosinase, arthropod hemocyanin, insect hexamerin, and dipteran
arylphorin receptor. J. Mol. Evol. 42, 713–728.
Burmester, T., Massey Jr., H.C., Zakharkin, S.O., Benes, H., 1998. The
evolution of hexamerins and the phylogeny of insects. J. Mol. Evol. 47,
93–108.
Chirgwin, J.M., Przbyla, A.E., MacDonald, R.J., Rutter, W.J., 1979.
Isolation of biologically active ribonucleic acid from sources enriched
in ribonuclease. Biochemistry 18, 5294–5299.
de Kort, C.A.D., Koopmanschap, A.B., 1994. Nucleotide and deduced
amino acid sequence of a cDNA clone encoding diapause protein 1, an
arylphorin-type storage hexamer of the Colorado potato beetle.
J. Insect Physiol. 40, 527–535.
Enderle, U., Kauser, G., Renn, L., Scheller, K., Koolman, J., 1983.
Ecdysteroids in the hemolymph of blowfly are bound to calliphorin.
In: Scheller, K. (Ed.), The Larval Serum Proteins of Insects: Function,
Biosynthesis, Genetic. Thieme, Stuttgart, New York, pp. 40–49.
Felsenstein, J., 1985. Confidence limits on phylogenies: an approach using
the bootstrap. Evolution 39, 783–791.
Felsenstein, J., 2004. PHYLIP (Phylogeny Inference Package), version 3.6.
Distributed by the author. Department of Genetics, University of
Washington, Seattle.
Flook, P.K., Rowell, C.H.F., 1998. Inferences about orthopteroid
phylogeny and molecular evolution from small subunit nuclear
ribosomal DNA sequences. Insect Mol. Biol. 7, 163–178.
Gaunt, M.W., Miles, M.A., 2002. An insect molecular clock dates the
origin of the insects and accords with palaeontological and biogeo-
graphic landmarks. Mol. Biol. Evol. 19, 748–761.
Gradstein, F.M., Ogg, J.G., Smith, A.G., et al., 2004. A Geologic
Timescale 2004. Cambridge University Press, Cambridge.
Hagner-Holler, S., Schoen, A., Erker, W., Marden, J.H., Rupprecht, R.,
Decker, H., Burmester, T., 2004. A respiratory hemocyanin from an
insect. Proc. Natl. Acad. Sci. USA 101, 871–874.
Hennig, W., 1969. Stammesgeschichte der Insekten. W. Kramer,
Frankfurt/M.
Hennig, W., 1981. Insect Phylogeny. Academic Press, New York.
Holmes, D.S., Bonner, J., 1973. Preparation, molecular weight, base
composition, and secondary structure of giant nuclear ribonucleic acid.
Biochemistry 12, 2330–2338.
Huelsenbeck, J.P., Ronquist, F., 2001. MRBAYES: Bayesian inference of
phylogenetic trees. Bioinformatics 17, 754–755.
Jamroz, R.C., Beintema, J.J., Stam, W.T., Bradfield, J.Y., 1996.
Aromatic hexamerin subunit from adult female cockroaches
(Blaberus discoidalis): molecular cloning, suppression by juvenile
hormone, and evolutionary perspectives. J. Insect Physiol. 42,
115–124.
King, J.L., Jukes, T.H., 1969. Non-Darwinian evolution. Science 164,
788–798.
Kukalova-Peck, J., 1991. Fossil history and evolution of hexapodstruc-
tures. In: Naumann, I.D. (Ed.), The Insects of Australia. Melbourne
University Press, Melbourne, Australia, pp. 141–179.
Langley, C.H., Fitch, W., 1974. An estimation of the constancy of the rate
of molecular evolution. J. Mol. Evol. 3, 161–177.
Lo, N., Tokuda, G., Watanabe, H., Rose, H., Slaytor, M., Maekawa, K.,
Bandi, C., Noda, H., 2000. Evidence from multiple gene sequences
indicates that termites evolved from wood-feeding cockroaches. Curr.
Biol. 10, 801–804.
Magee, J., Kraynack, N., Massey Jr., H.C., Telfer, W.H., 1994. Properties
and significance of a riboflavin-binding hexamerin in the hemo-
lymph of Hyalophora cecropia. Arch. Insect Biochem. Physiol. 25,
137–157.
Martinez, T., Burmester, T., Veenstra, J., Wheeler, D., 2000. Sequence and
evolution of a hexamerin from the ant, Camponotus festinatus. Insect
Mol. Biol. 9, 427–431.
Nicholas, K.B., Nicholas Jr., H.B., 1997. GeneDoc: Analysis and
Visualization of Genetic Variation. /www.psc.edu/biomed/genedoc/S.Nielsen, H., Engelbrecht, J., Brunak, S., von Heijne, G., 1997.
Identification of prokaryotic and eukaryotic signal peptides and
prediction of their cleavage sites. Protein Eng. 10, 1–6.
Pan, M.L., Telfer, W.H., 1996. Methionine-rich hexamerin and
arylphorin as presursor reservoirs for reproduction and metamor-
phosis in female luna moths. Arch. Insect Biochem. Physiol.
33, 149.
Peter, M.G., Scheller, K., 1991. Arylphorin and the integument. In:
Retnakaran, A., Binnington, K. (Eds.), The Physiology of Insect
Epidermis. CSIRO, Australia, pp. 113–122.
Phipps, D.J., Chadwick, J.S., Aston, W.P., 1994. Gallysin-1, an
antibacterial protein isolated from hemolymph of Galleria mellonella.
Dev. Comp. Immunol. 18, 13–23.
Ross, A.J., Jarzembowski, E.A., 1993. Hexapoda. In: Benton, M.J. (Ed.),
The Fossil Record 2. Chapman & Hall, London, pp. 363–426.

ARTICLE IN PRESSS. Hagner-Holler et al. / Insect Biochemistry and Molecular Biology 37 (2007) 1064–10741074
Ryan, R.O., Anderson, D.R., Grimes, W.J., Law, J.H., 1985. Arylphorin
from Manduca sexta: carbohydrate structure and immunological
studies. Arch. Biochem. Biophys. 243, 115–124.
Sanderson, M.J., 1997. A nonparametric approach to estimating
divergence times in the absence of rate constancy. Mol. Biol. Evol.
14, 1218–1231.
Sanderson, M.J., 2002. Estimating absolute rates of molecular evolution
and divergence times: a penalized likelihood approach. Mol. Biol.
Evol. 19, 101–109.
Scharf, M.E., Wu-Scharf, D., Zhou, X., Pittendrigh, B.R., Bennett, G.W.,
2005. Gene expression profiles among immature and adult reproduc-
tive castes of the termite Reticulitermes flavipes. Insect Mol. Biol. 14,
31–44.
Scheller, K., Fischer, B., Schenkel, H., 1990. Molecular properties,
functions and developmentally regulated biosynthesis of arylphorin in
Calliphora vicina. In: Hagedorn, H.H., Hildebrand, J.G., Kidwell,
M.G., Law, J.H. (Eds.), Molecular Insect Science. Plenum Press, New
York, pp. 155–162.
Strimmer, K., von Haeseler, A., 1997. Likelihood-mapping: a simple
method to visualize phylogenetic content of a sequence alignment.
Proc. Natl. Acad. Sci. USA 94, 6815–6819.
Telfer, W.H., Kunkel, J.G., 1991. The function and evolution of insect
storage hexamers. Annu. Rev. Entomol. 36, 205–228.
Telfer, W.H., Keim, P.S., Law, J.H., 1983. Arylphorin, a new protein from
Hyalophora cecropia: comparison with calliphorin and manducin.
Insect Biochem. 13, 601–613.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins,
D.G., 1997. The ClustaIX windows interface: flexible strategies for
multiple sequence alignment aided by quality analysis tools. Nucleic
Acids Res. 25, 4876–4882.
Tojo, S., Yoshiga, T., 1994. Purification and characterization of three
storage proteins in the common cutworm, Spodoptera litura. Insect
Biochem. Mol. Biol. 24, 729–738.
Von Heijne, G., 1986. A simple method for predicting signal peptide
cleavage sequences. Nucleic Acids Res. 14, 4683–4690.
Wheeler, D.E., Buck, N.A., 1995. Storage proteins in ants during
development and colony founding. J. Insect. Physiol. 41, 885–894.
Wheeler, W.C., Whiting, M., Wheeler, Q.D., Carpenter, J.M., 2001. The
phylogeny of the extant hexapod orders. Cladistics 17, 113–169.
Whelan, S., Goldman, N., 2001. A general empirical model of protein
evolution derived from multiple protein families using a maximum-
likelihood approach. Mol. Biol. Evol. 18, 691–699.
Wu, C.H., Lee, M.F., Liao, S.C., Luo, S.F., 1996. Sequencing analysis of
cDNA clones encoding the American cockroach Cr-PI allergens.
Homology with insect hemolymph proteins. J. Biol. Chem. 271,
17937–17943.
Zhou, X., Oi, F.M., Scharf, M.E., 2006. Social exploitation of hexamerin:
RNAi reveals a major caste-regulatory factor in termites. Proc. Natl.
Acad. Sci. USA 103, 4499–4504.
Zhou, X., Tarver, M.R., Scharf, M.E., 2007. Hexamerin-based regulation
of juvenile hormone-dependent gene expression underlies phenotypic
plasticity in a social insect. Development 134, 601–610.


![Basel 3 & Implication[1]](https://static.fdocuments.us/doc/165x107/577d1e721a28ab4e1e8e9042/basel-3-implication1.jpg)













