Distinct Regions of the Mouse Cyclin A1 Gene, Ccna1, Confer Male ...
30
Distinct Regions of the Mouse Cyclin A1 Gene, Ccna1, Confer Male Germ-Cell Specific Expression and Enhancer Function 1 Expression of Ccna1 Reporter Genes in Mice cyclin, spermatogenesis, meiosis, gene regulation, testis Karen M. Lele 2 and Debra J. Wolgemuth 2,3,4,5,6,7 1 This research was supported by NIH grants HD34915 (D.J.W) and T32 DK07647 (K.M.L.) 2 The Institute of Human Nutrition, Departments of 3 Genetics and Development and 4 Obstetrics and Gynecology, 5 The Center for Reproductive Sciences, 6 The Herbert Irving Comprehensive Cancer Center, Columbia University Medical Center, New York, New York 10032 7 Correspondence: Dr. Debra Wolgemuth, Dept. Genetics and Development, Columbia University Medical Center, 630 West 168 th Street, Black 1613, New York, New York 10032; FAX: 212 305 6084; e-mail: [email protected] BOR Papers in Press. Published on June 23, 2004 as DOI:10.1095/biolreprod.104.030387 Copyright 2004 by The Society for the Study of Reproduction.
Transcript of Distinct Regions of the Mouse Cyclin A1 Gene, Ccna1, Confer Male ...

Distinct Regions of the Mouse Cyclin A1 Gene, Ccna1, Confer Male Germ-Cell Specific
Expression and Enhancer Function1
Expression of Ccna1 Reporter Genes in Mice
cyclin, spermatogenesis, meiosis, gene regulation, testis
Karen M. Lele2 and Debra J. Wolgemuth2,3,4,5,6,7
1This research was supported by NIH grants HD34915 (D.J.W) and T32 DK07647
(K.M.L.)
2The Institute of Human Nutrition, Departments of 3Genetics and Development and
4Obstetrics and Gynecology, 5The Center for Reproductive Sciences, 6The Herbert Irving
Comprehensive Cancer Center, Columbia University Medical Center, New York, New
York 10032
7Correspondence: Dr. Debra Wolgemuth, Dept. Genetics and Development, Columbia
University Medical Center, 630 West 168th Street, Black 1613, New York, New York
10032; FAX: 212 305 6084; e-mail: [email protected]
BOR Papers in Press. Published on June 23, 2004 as DOI:10.1095/biolreprod.104.030387
Copyright 2004 by The Society for the Study of Reproduction.

ABSTRACT 1
The gene encoding mouse cyclin A1, Ccna1, is expressed at highest levels in late 2
pachytene-diplotene spermatocytes, where it is required for meiotic cell division. To 3
begin to understand the mechanisms responsible for its highly restricted pattern of 4
expression, transgenic mouse lines carrying constructs consisting of the cyclin A1 5
regulatory region fused with the reporter gene lacZ were generated. Analysis of tissue-6
specific and testicular cell type-specific transgene expression indicated that sequences 7
within -1.3 kb of the cyclin A1 putative transcriptional start site were sufficient to direct 8
transgene expression uniquely to late spermatocytes while maintaining repression in other 9
tissues. However, sequences located between -4.8 kb and -1.3 kb of the putative 10
transcriptional start site were apparently required to transcribe the reporter at levels 11
needed for consistent X-gal staining. Comparison of the mouse, rat and human proximal 12
promoters revealed regions of high sequence conservation and consensus sequences both 13
for known transcripton factors, some of which are co-expressed with Ccna1, such as A-14
myb and Hsf2, and for elements that control expression of genes in somatic cell cycles, 15
such as CDE, CHR, and CCAAT elements. Thus, the promoter region within 1.3 kb 16
upstream of the putative Ccna1 transcriptional start can direct expression of lacZ to 17
spermatocytes, while sequences located between -4.8 kb and -1.3 kb of the putative 18
transcriptional start site may enhance expression of lacZ. 19
20

2
INTRODUCTION 20
Spermatogenesis is an ordered process during which germ cells enter successive 21
mitotic, meiotic, and post-meiotic phases. The program of gene expression required for 22
this process is tightly controlled, involving both germ cell-specific and common 23
transcription factors, whose expression is also stringently regulated (rev. in [1, 2]). This 24
regulation allows precisely ordered events to occur such as genetic recombination and 25
meiotic cell division, followed by the morphological development of spermatids. 26
Genes for several key somatic cell cycle regulators, including cyclins, cyclin-27
dependent kinases (Cdks), Cdk inhibitors, and Cdc25 family members are expressed 28
during spermatogenesis in specific patterns that suggest their function in cell cycle 29
control in the germ line as well [3, 4]. Cyclins are regulatory subunits of the Cdk 30
complexes and are expressed periodically during the cell cycle. There are at least ten 31
classes of cyclins in higher vertebrates, designated cyclins A to I and T, and multiple 32
members of the A-, B-, and D-type cyclin families (rev. in [5-7]). 33
In higher organisms, there are two A-type cyclins, cyclin A1 and cyclin A2. In 34
mice, both are expressed in male germ cells, albeit during quite different stages of 35
differentiation, suggesting that they are tightly regulated and may have distinct functions 36
[8, 9]. The cyclin A1 gene, Ccna1, appears to be testis-specific and is expressed late in 37
the meiotic cell cycle, just prior to chromosomal desynapsis [8, 9]. Disruption of mouse 38
Ccna1 resulted in male infertility and complete spermatogenic arrest prior to the first 39
meiotic division [10]. In contrast, cyclin A2 (Ccna2) is widely expressed in mouse 40
tissues [8]. In the male germ line, Ccna2 is expressed earlier than Ccna1, in 41
spermatogonia and preleptotene spermatocytes, and is no longer detected as germ cells 42

3
enter the leptotene stage [9]. Disruption of the Ccna2 gene resulted in embryonic 43
lethality [11]. Therefore, its role in spermatogenesis could not be discerned from a null 44
model. 45
Human CCNA1 is expressed most highly in testis, and at lower levels in adult 46
brain and in hematopoietic cells [12, 13]. It is also expressed in several human myeloid 47
leukemia cell lines and an osteosarcoma cell line, where its level of expression appears to 48
be cell cycle-regulated [12, 14]. In testis, human cyclin A1 is localized in late meiotic 49
prophase spermatocytes, similarly to mouse cyclin A1 [15]. A short fragment of the 50
human CCNA1 promoter, comprised of 190 bp of sequence upstream and 145 bp of 51
sequence downstream of the transcriptional start site, has been reported to activate 52
expression of a reporter gene in Hela cells [16] and CV-1 cells [17]. In transgenic mice, 53
a 1.3 kb fragment of the human CCNA1 promoter has further been reported to direct 54
expression of EGFP to male germ cells but in a much less restricted expression pattern 55
than observed for the endogenous mouse gene [18]. That is, in addition to stage IX to 56
XII spermatocytes, EGFP fluorescence was detected in spermatogonia, at earlier stages of 57
spermatocyte development and in spermatids [18, 19]. 58
The DNA regulatory elements required for mouse Ccna1 gene expression in male 59
germ cells have not been identified. We hypothesize that there will be regulatory 60
elements unique to Ccna1, not present in the Ccna2 promoter, reflecting their distinct 61
expression patterns, and further that such elements might be evolutionarily conserved. 62
To begin to define the location of these elements and, thus, begin to understand 63
mechanisms that control the stage-specific expression of Ccna1 and repression at other 64
stages of male germ cell development, we have tested expression of Ccna1 65

4
promoter/reporter constructs in mice. This in vivo approach was chosen to define 66
promoter function in male germ cells since procedures for transfection of these cells in 67
culture have not been established. In the present study, the regulatory function of 68
genomic fragments spanning –8.2 kb to +0.8 kb from the putative transcriptional start site 69
of mouse Ccna1 was examined in transgenic mice, using the reporter lacZ. The patterns 70
of transgene expression revealed that the genomic fragment that lies between –1.3 kb to 71
+0.8 kb of the putative Ccna1 transcriptional start site could fully recapitulate the correct 72
developmental expression of the endogenous Ccna1 gene in male germ cells with no 73
ectopic expression observed, unlike the human CCNA1 reporter constructs. However, 74
sequences between –4.8 to –1.3 of the putative transcriptional start appeared to be 75
necessary for fully penetrant expression of the gene in spermatocytes. Within this 76
genomic region, consensus binding sequences for several transcription factors were 77
identified. They include sequences known to bind factors that are co-expressed with 78
Ccna1 and also sequences that control cell cycle regulated gene expression in somatic 79
cell cycles. Alignment of the proximal promoters of the mouse, rat, and human genes for 80
cyclin A1 revealed regions of high homology; however, the human proximal promoter 81
also contained inserted sequences that were not present in the mouse and rat promoters. 82
83
MATERIALS AND METHODS 84
Transgene constructs and transgenic mice 85
A 5.6 kb fragment that extends 5’ from the BamHI site in exon 2 of Ccna1 to an 86
EcoRI site in the 5’ flanking region was inserted into a vector containing a cassette 87
composed of lacZ and intron 1 and the polyadenylation signal of mouse protamine-1 to 88

5
yield construct 4.8cyA1lacZ (Fig. 1). The reporter cassette had been used previously to 89
produce a Hoxa4 promoter-reporter construct that was expressed in male germ cells [20]. 90
8.2cyA1lacZ was generated from 4.8cyA1lacZ by inserting a SalI-EcoRI fragment 5’ of 91
the EcoRI site. 1.3cyA1lacZ was produced from 4.8cyA1lacZ by digestion with XhoI 92
and religation. All constructs were verified by sequencing. The fusion gene product is 93
expected to have the first 33 amino acids of the cyclin A1 protein linked to β-94
galactosidase (β-gal). The production of transgenic mice was carried out as previously 95
described [20, 21]. All procedures were performed in accord with guidelines of the 96
Institutional Animal Care and Use Committee of the Columbia University Medical 97
Center. Transgenic animals were identified by Southern blot analysis of tail DNA 98
digested with EcoRI, using as a probe a 2 kb EcoRV-EcoRI fragment of lacZ (Fig. 1) 99
labeled with [γ-32P]dCTP by random priming, according to our standard procedures [20]. 100
The probe detects a 3 kb band corresponding to full length lacZ cDNA. 101
102 Tissue staining and histology 103
The X-gal staining procedure was similar to that of Behringer et al. [20]. Briefly, 104
mice were sacrificed by cervical dislocation or CO2 asphyxiation and perfused 105
transcardially with fixative (0.2% glutaraldehyde, 2% formaldehyde, 5 mM EGTA, 0.1% 106
deoxycholic acid, 0.2% Nonidet P-40, 2 mM MgCl2, and 0.1 M phosphate buffer pH 107
7.3). The tissues were dissected out immediately (the testes were decapsulated and the 108
tubules were gently teased apart), and soaked in fixative for 1 hr at room temperature. 109
After thorough washing in rinse buffer (0.1 M Phosphate buffer pH 7.3, 0.1% 110
deoxycholic acid), the tissues were stained overnight at 30°C in X-gal solution (5 mM 111
potassium ferricyanide, 5 mM potassium ferrocyanide, 2 mM MgCl2). After staining, the 112

6
tissues were fixed overnight in 4% paraformaldehyde in 0.1 M phosphate buffer pH 7.3. 113
Intact tubules were viewed and photographed with a Wild MPS 51 Dissecting 114
Microscope (Wild Heerbrugg Ltd., Heerbrugg, Switzerland). For histological study, the 115
stained tissues were dehydrated and embedded in paraffin. Five-µm-thick sections were 116
deparaffinized with Histo-Clear (National Diagnostics, Atlanta, GA) and counterstained 117
with neutral red. The sections were photographed using a Nikon Eclipse 800 microscope 118
(Nikon Instrument Group, Melville, NY) with a SPOT digital camera (Diagnostic 119
Instruments, Inc., Sterling Heights, MI). 120
121
Northern blot hybridization analysis 122
Total RNA was isolated from tissues dissected from euthanized adult mice using 123
TriReagent (Molecular Research Center, Inc., Cincinnati, OH) or an RNeasy kit (Qiagen, 124
Chatsworth, CA) following the manufacturer’s instructions. Twenty µg samples of 125
denatured total RNA were resolved on a 0.8% denaturing agarose gel according to our 126
published procedures [10]. Ethidium bromide staining of the 18S and 28S rRNA bands 127
was used to determine equal sample loading. The gels were blotted onto nitrocellulose 128
membranes. The 32P-labeled antisense cRNA probe was 1 kb and corresponded to the 129
5’end of the lacZ cDNA (Fig. 1). The membranes were hybridized at 65°C overnight 130
using 107 cpm/ml of lacZ cRNA in hybridization solution (5x SSC, 20 mM sodium 131
phosphate buffer (pH 7), 60% formamide, 1% SDS, 5x Denhardt’s solution, 100 ug/ml 132
salmon sperm RNA, and 7% dextran sulfate). The membranes were washed with 133
increasing stringency in solutions of 0.2X SSC and 1% SDS. The hybridization signal 134
was detected using XAR 5 film (Eastman Kodak Co., Rochester, NY). 135

7
136 In situ hybridization analysis 137
Testes and selected other tissues were dissected and fixed in 4% 138
paraformaldehyde in PBS overnight at 4°C and then dehydrated prior to paraffin 139
embedding. Six µm sections were cut. Paraffin was removed using Histo-Clear and the 140
sections were rehydrated. Sense and antisense cRNA probes were transcribed from 141
template consisting of the 5’ 1 kb of lacZ cDNA (Fig. 1) using [35S]UTP as radiolabel. 142
Hybridization was carried out overnight at 65°C in 50% formamide, 0.3M NaCl, 20 mM 143
Tris pH 8.0, 5 mM EDTA, 10 mM sodium phosphate pH 8.0, 10% dextran sulfate, 1x 144
Denhardt’s solution, 500 µg/ml yeast tRNA and 1 x 104 cpm/ml probe. After 145
hybridization, the slides were washed in 5x SSC for 10 min. at 50°C and then in 80% 146
formamide and 2x SSC for 20 min. at 65°C. Single-stranded RNA was digested in 0.3 M 147
NaCl, 10 mM Tris pH 8.0, 5 mM EDTA and 50 µg/ml RNase A for 30 min. at 37°C. 148
The RNase-treated slides were washed with 50% formamide and 2x SSC for 20 min. at 149
65°C and further rinsed with 2x SSC and with 0.1x SSC each for 15 min. at room 150
temperature. The slides were dehydrated, coated with NTB-2 emulsion (Eastman Kodak 151
Co., Rochester, NY) and exposed for 2 weeks at 4°C. The slides were developed and 152
stained with hematoxylin and eosin. Photomicrographs were taken with a DIALUX 20 153
microscope (Leica Microsystems, Wetzlar, Germany) using epifluorescence optics and a 154
SPOT digital camera. 155
156
157 Results 158 159 Generation of cyA1lacZ transgenic mice 160

8
To begin to identify sequences responsible for the highly restricted pattern of 161
mouse Ccna1 expression, constructs composed of varying lengths of Ccna1 5’ genomic 162
sequence linked to lacZ cDNA were prepared and used to generate transgenic mice. The 163
transgenes were constructed with mouse genomic fragments consisting of 8.2 kb, 4.8 kb, 164
or 1.3 kb of Ccna1 5’ flanking sequence and the 5’ end of the Ccna1 structural gene to 165
exon 2 fused in-frame to a reporter cassette consisting of lacZ and the 3’ UTR and intron 166
1 of the testis-specific gene protamine1, yielding 8.2cyA1lacZ, 4.8cyA1lacZ, and 167
1.3cyA1lacZ, respectively (Fig. 1). Two permanent mouse lines each carrying 168
8.2cyA1lacZ and 1.3cyA1lacZ and one permanent mouse line carrying 4.8cyA1lacZ 169
were established. Each of the 5 lines was characterized further. 170
171
Distribution of cyA1lacZ transgene expression in adult tissues 172
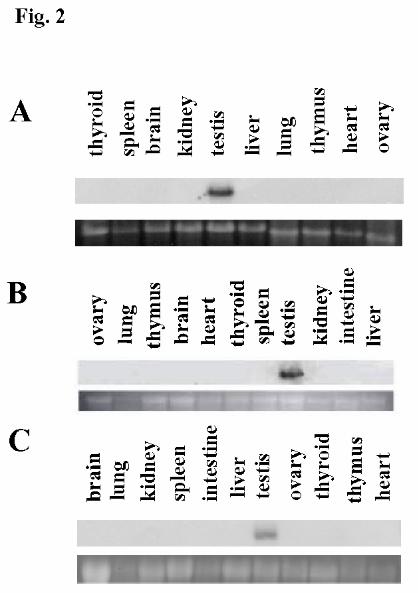
To establish the tissue specificity of expression of the transgenes, RNA was 173
isolated from a variety of tissues from adult transgenic mice and analyzed by Northern 174
blot hybridization analysis. For all lines, a lacZ transcript was detected in RNA from 175
testis only (Fig. 2A – C; and data not shown), similarly to endogenous Ccna1. This result 176
was confirmed by in situ hybridization analysis of selected tissues from each line using a 177
lacZ probe (data not shown). These results suggested that the testis-specificity of 178
expression of Ccna1 is conferred by sequences within 1.3 kb upstream of its putative 179
transcriptional start. 180
181
Pattern of B-galactosidase activity in cyA1lacZ testes 182

9
To determine the cellular specificity of expression of the cyA1lacZ transgenes, 183
testes were stained with X-gal as whole mounts and then sectioned. Visualization of X-184
gal staining in intact tubules of 8.2cyA1lacZ testis (Fig. 3A) and 4.8cyA1lacZ testis (Fig. 185
3B) were identical and revealed dark blue staining except at the edges of the tubules, 186
which appeared unstained. This suggested that the cells closest to the basement 187
membrane of the tubules, the spermatogonia and early meiotic spermatocytes, did not 188
express the transgenes. In contrast, tubules from wild type testis were unstained, 189
although non-specific staining appeared in interstitial cells, which are located between 190
tubules (inset of Fig. 3A). Histological sections of the X-gal stained tubules from 191
8.2cyA1lacZ testis (Figs. 3D, 3G, and 3J) and 4.8cyA1lacZ testis (Figs. 3E, 3H, and 3K) 192
confirmed this observation. No staining was observed in spermatogonia or in 193
spermatocytes through stage VIII (Fig. 3D and 3E). Rather, the onset of expression 194
appeared in stage IX spermatocytes (Fig. 3G and 3H), similar to the appearance of 195
endogenous Ccna1 mRNA and protein [8]. X-gal staining was present in spermatocytes 196
at later stages, including stage XII (Fig. 3J and 3K), and persisted in all round and 197
elongating spermatids (Figs. 3D, 3E, 3G, 3H, 3J, 3K), cells known not to express cyclin 198
A1 (see results below). This staining pattern suggested that the transgenes contained 199
sequences required to direct the onset of expression of lacZ to stage IX spermatocytes. 200
Examination of stained, intact testicular tubules from the 1.3cyA1lacZ mice 201
revealed a less uniform X-gal staining pattern (Fig. 3C) as compared to that of 202
8.2cyA1lacZ (Fig. 3A) or 4.8cyA1lacZ mice (Fig. 3B). Unlike the consistent appearance 203
of X-gal staining in late spermatocytes and spermatids in 8.2cyA1lacZ and 4.8cyA1lacZ 204
testis, the staining of spermatocytes and spermatids within tubules of 1.3cyA1lacZ testis 205

10
was sometimes absent or was of varying intensity (Figs. 3C, 3F, and 3L). However, the 206
cellular specificity of expression exhibited was similar to 8.2cyA1lacZ and 4.8cyA1lacZ 207
lines. First, the onset of expression of the transgenes was similar. Examination of 208
sectioned tubules revealed that staining was absent in spermatogonia and in 209
spermatocytes through stage VIII (Fig. 3F), and β-gal activity first appeared in stage IX 210
spermatocytes (Fig. 3I). Also, β-gal activity persisted in the spermatids in the 211
1.3cyA1lacZ transgenic testis (Figs. 3F, 3I, and 3L). These observations suggest that the 212
Ccna1 genomic fragment within -1.3 kb to +0.8 kb from its transcriptional start contains 213
sequences needed to direct the onset of expression to stage IX spermatocytes, but that 214
additional upstream sequences from -4.8 kb and -1.3 kb of the Ccna1 transcriptional start 215
site were needed to direct detectable expression of 1.3cyA1lacZ in all spermatocytes. In 216
support of these observations, Northern blot analysis of total RNA from adult testis of 217
mice carrying each construct shows that 1.3cyA1lacZ is expressed at lower levels than 218
4.8cyA1lacZ or 8.2cyA1lacZ (Fig. 4). 219
The detection of β-gal activity in spermatids may have been due to expression of 220
the transgenes at stages later than that of endogenous Ccna1. Alternatively, the lacZ 221
reporter may have been expressed at the same developmental stages as Ccna1, but its 222
gene product may have persisted in spermatids. To determine if ectopic expression of the 223
transgene was responsible for the β-gal activity detected in spermatids in cyA1lacZ 224
testes, lacZ expression was assayed by in situ hybridization. LacZ mRNA was detected 225
in spermatocytes as late as stage XII (Fig. 5A), but was not detected in spermatids at 226
stage I (Fig. 5A) or in round or elongating spermatids at later developmental stages (Figs. 227
5A – 5D). Each of the cyA1lacZ constructs exhibited this same pattern of expression 228

11
(Fig. 5 and data not shown). The localization of lacZ mRNA only in spermatocytes 229
suggested that the β-gal activity detected in spermatids was not produced by ectopic 230
expression of the transgenes, but rather to stable β-gal protein. This further suggested 231
that sequences within -1.3 kb to +0.8 kb from the mouse Ccna1 transcriptional start site, 232
unlike its human homologue, were sufficient to control the correct timing of 233
spermatocyte-specific expression of the transgenes in germ cells. 234
A curiosity of the expression of these transgenes in the testis was noted in our in 235
situ hybridization analysis when a sense lacZ probe was used as a control. Signal was 236
detected in stage VI to VIII spermatocytes in transgenic testis sections for all three 237
constructs (Fig. 5E and data not shown). This signal was not present in sections from 238
non-transgenic testis (Fig. 5F). This expression might result from activity of a cryptic 239
promoter driving transcription in the opposite orientation in the protamine1 sequence 240
placed 3’ to lacZ in each transgene. However, promoter elements that support 241
transcriptional initiation, as assessed by TFSearch [22], were not detected in this 242
sequence. 243
244
Consensus elements for transcriptional regulators located between -4.8 kb and +0.8 kb 245
of the Ccna1 putative transcriptional start site 246
Intron 1 and the region between -4.8 kb and the putative transcriptional start site 247
of the Ccna1 gene were sequenced and scanned for putative transcription factor binding 248
sites using the search engine TFSEARCH and the TRANSFAC database [22]. The 249
locations of consensus sequences of DNA binding motifs for known transcription factors 250
and for binding activities are listed in Table 1. Among these motifs are binding sites for 251

12
transcription factors that are co-expressed in spermatocytes with Ccna1, such as A-myb 252
[23], Hsf2 [24], and Sp1 [25]. Consensus sequences for A-myb and Hsf2 binding are 253
located between -4.8 kb and -1.3 kb of the putative Ccna1 transcriptional start site, and 254
A-myb and Sp1 consensus sequences located within -1.3 kb of the putative Ccna1 255
transcriptional start site (Table 1). 256
The proximal promoter of Ccna1 was aligned with the corresponding rat and 257
human promoters. A region of high homology was found from -511 to -448 of the 258
putative Ccna1 start site (Fig. 6A), although this region lacked consensus sequences for 259
transcription factor binding sites listed in the TRANSFAC database [22]. However, 260
highly conserved consensus sequences for Sp1, GATA-1, Ap-1, and delta EF1 binding 261
sites, and a CCAAT box were identified (Table 1 and Fig. 6B) [22]. Downstream of 262
these consensus sequences and 153 bp upstream of the putative transcriptional start site 263
of the Ccna1 gene, the first of two bipartite ‘cell-cycle dependent element’/’cell-cycle 264
genes homology region’ (CDE/CHR) elements were found (Table 1 and Fig. 6B). In the 265
mouse and rat promoters, there are two conserved sets of putative CDE/CHR elements. 266
In the human promoter, the 5’ set and only the CDE sequence of the 3’ set are found; that 267
is, the human promoter lacks the CHR sequence of the 3’ CDE/CHR present in the mouse 268
and rat promoters. In addition, unpaired CDE and CHR elements are present, which are 269
conserved among all three cyclin A1 genes (Fig. 6B). 270
In the human proximal promoter, there are sequences that are poorly conserved or 271
not conserved at all with mouse and rat, for example, several GC boxes (Fig. 6B). These 272
sequences, in the context of a human CCNA1 reporter construct, have been suggested to 273
contribute to transcriptional activation in Hela cells [16]. 274

13
275
Discussion 276
In mice, the Ccna1 gene is expressed specifically in testis in stage IX to XII 277
spermatocytes [8]. Human CCNA1 is reported to be expressed at highest levels in male 278
germ cells [15], and at very low levels in brain and hematopoietic cells of normal tissues 279
[12, 13, 26]. Other sites of expression of mouse Ccna1 are yet to be confirmed. The 280
tissue-specificity of Ccna1 expression contrasts with that of Ccna2, which is expressed in 281
a variety of tissues [8, 27]. Ccna1 and Ccna2 also have non-overlapping expression 282
patterns during male germ cell development, as Ccna2 expression is downregulated early 283
in the meiotic cell cycle before Ccna1 is expressed [8, 9]. We therefore hypothesize that 284
there will be specific regulatory elements unique to each A-type cyclin as well. The 285
essential role of Ccna1 in male germ cell development and the concurrent expression of 286
cyclin A1 message and protein suggested the importance of understanding mechanisms 287
controlling transcription of the Ccna1 gene. The lack of cell lines derived from male 288
germ cells made it necessary to analyze the Ccna1 promoter in these cells in vivo, in 289
transgenic mice. The use of serial deletions of Ccna1 upstream sequence has allowed us 290
to define several functional segments of the promoter. 291
Analysis of transgene expression in the testis, using a combination of X-gal 292
staining and in situ hybridization, revealed their expression specifically in spermatocytes 293
at stages IX to XII of the cycle of the seminiferous epithelium, and not in earlier stages. 294
The endogenous mouse Ccna1 has been shown to be expressed only during this same 295
narrow window of spermatogenesis. This suggests that the genomic fragment of mouse 296
Ccna1 spanning -1.3 kb to +0.8 kb of the putative transcriptional start site contains 297

14
sequences necessary for the stage-specific expression of Ccna1 in the meiotic cell cycle. 298
In contrast, 1.3 kb of human CCNA1 promoter directs expression of EGFP in a much less 299
restrictive pattern in male germ cells [19], reflecting differences in the mouse and human 300
promoters despite their sharing highly conserved regions. 301
Within this fragment are consensus sequences for two sets of paired CDE/CHR 302
elements. These elements were first discovered in the proximal promoter of genes for 303
human cyclin A2, Cdc2, and Cdc25C, which are expressed in S and G2 phases of the 304
mitotic cell cycle [28]. Several lines of evidence suggest that the CDE/CHR elements 305
control the timing of expression of these genes during the cell cycle. In vivo footprinting 306
of the human CCNA2 proximal promoter revealed that the bipartite element was occupied 307
at stages that the gene was not transcribed [28]. Mutation of the CDE/CHR element in 308
the context of CCNA2 or CDK1 promoter/reporter genes caused derepression of the 309
reporters in G1 phase [28]. These elements have now been shown to be present and 310
involved in controlling the timing of expression of other cell cycle-regulated genes, 311
including the genes for cyclin B2 [29], rabkinesin6 [30], polo-like kinase [31], p130 [32], 312
m-survivin [33], and aurora A [34]. Also, the CDE/CHR element appears to 313
downregulate expression of the CDK1 gene in response to TPA-induced differentiation of 314
U937 cells [35] or in response to p53-dependent DNA damage [36]. The promoters of 315
the genes for mouse and rat cyclin A1 are unique in that they contain two sets of 316
CDE/CHR elements, and like the human gene, also have unpaired CDE and CHR 317
consensus sequences. There have been no previous reports of unpaired CDE elements, 318
but the promoters of the genes for human cyclin B2 ([37]) and mouse Cdc25C ([38]) are 319
regulated by CHR elements that are not paired with functional CDE elements. Testing of 320

15
mutated CDE/CHR elements in transgenic mice will determine the role, if any, of these 321
elements in controlling the expression of Ccna1 in the meiotic cell cycle. Factors that 322
bind CDE/CHR [39] or CHR [40, 41] have been detected in various cultured cell lines, 323
but have not been identified. 324
The less efficient X-gal staining in male germ cells from both lines of 325
1.3cyA1lacZ transgenic mice, as compared to germ cells from 8.2cyA1lacZ and 326
4.8cyA1lacZ transgenic mice, suggests that sequences that lie between 4.8 kb and 1.3 kb 327
upstream of the putative Ccna1 transcriptional start contains enhancer elements needed 328
for consistent expression of lacZ. Within this region, consensus elements for several 329
regulatory factors expressed in spermatocytes have been identified (Table 1). Hsf2 [24] 330
and A-myb [23] are reported to be expressed in mid- to late-stage spermatocytes, raising 331
the possibility that they may be involved in the regulation of expression of Ccna1. 332
Interestingly, disruption of A-myb in mice causes spermatogenic arrest during mid-333
prophase and male sterility [42]. Developmentally, arrest in A-myb-/- spermatocytes 334
occurs slightly earlier than the arrest in Ccna1-/- spermatocytes. 335
In summary, our results suggest that the genomic fragment -1.3 kb to +0.8 kb 336
from the putative Ccna1 transcriptional start contains sequences that mediate the correct 337
developmental expression of Ccna1 in male germ cells, as compared to this region in the 338
human CCNA1 gene. Further, sequences located between -4.8 kb and -1.3 kb from the 339
putative Ccna1 transcriptional start site appear to contribute to enhanced expression at 340
theses stages. Finally, sequences that lie between -1.3 kb and +0.8 kb from the Ccna1 341
transcriptional start appear to be sufficient to maintain repression of Ccna1 in the correct 342
adult tissues and stages of spermatogenic differentiation. Additional deletions in the -1.3 343

16
to +0.8 region may identify the location of elements that repress transcription of Ccna1 in 344
other cell types while mutation of known transcriptional regulatory elements and/or the 345
CDE-CHR elements may shed light on enhancer and cell-cycle regulatory roles, 346
respectively. Currently, the Ccna1 promoter we have characterized will provide a useful 347
tool to direct the expression of genes specifically to late meiotic spermatocytes in vivo. 348
349
Acknowledgements 350
We are very grateful to Dr. Xiangyuan Wang for performing microinjection and 351
producing histological sections and to Ms. Stacey Baptiste for staining 4.8cyA1lacZ 352
testes. 353

References
1. Sassone-Corsi P. Unique chromatin remodeling and transcriptional regulation in spermatogenesis. Science 2002; 296: 2176-2178.
2. Eddy EM. Male germ cell gene expression. Recent Prog Horm Res 2002; 57: 103-128.
3. Wolgemuth DJ, Rhee K, Wu S, Ravnik SE. Genetic control of mitosis, meiosis and cellular differentiation during mammalian spermatogenesis. Reproduction, Fertility, and Development 1995; 7: 669-683.
4. Beumer TL, Roepers-Gajadien HL, Gademan LS, Rutgers DH, de Rooij DG. P21(Cip1/WAF1) expression in the mouse testis before and after X irradiation. Mol Reprod Dev 1997; 47: 240-247.
5. Sherr CJ. Mammalian G1 cyclins. Cell 1993; 73: 1059-1065. 6. Pines J. Cyclins and cyclin-dependent kinases: a biochemical view. Biochem J
1995; 308: 697-711. 7. Wolgemuth DJ, Laurion E, Lele KM. Regulation of the mitotic and meiotic cell
cycles in the male germ line. Recent Prog Horm Res 2002; 57: 75-101. 8. Sweeney C, Murphy M, Kubelka M, Ravnik SE, Hawkins CF, Wolgemuth DJ,
Carrington M. A distinct cyclin A is expressed in germ cells in the mouse. Development 1996; 122: 53-64.
9. Ravnik SE, Wolgemuth DJ. Regulation of meiosis during mammalian spermatogenesis: the A-type cyclins and their associated cyclin-dependent kinases are differentially expressed in the germ-cell lineage. Developmental Biology 1999; 207: 408-418.
10. Liu D, Matzuk MM, Sung WK, Guo Q, Wang P, Wolgemuth DJ. Cyclin A1 is required for meiosis in the male mouse. Nature Genetics 1998; 20: 377-380.
11. Murphy M, Stinnakre MG, Senamaud-Beaufort C, Winston NJ, Sweeney C, Kubelka M, Carrington M, Brechot C, Sobczak-Thepot J. Delayed early embryonic lethality following disruption of the murine cyclin A2 gene [published erratum appears in Nat Genet 1999 Dec;23(4):481]. Nature Genetics 1997; 15: 83-86.
12. Yang R, Morosetti R, Koeffler HP. Characterization of a second human cyclin A that is highly expressed in testis and in several leukemic cell lines. Cancer Research 1997; 57: 913-920.
13. Kramer A, Hochhaus A, Saussele S, Reichert A, Willer A, Hehlmann R. Cyclin A1 is predominantly expressed in hematological malignancies with myeloid differentiation. Leukemia 1998; 12: 893-898.
14. Yang R, Muller C, Huynh V, Fung YK, Yee AS, Koeffler HP. Functions of cyclin A1 in the cell cycle and its interactions with transcription factor E2F-1 and the Rb family of proteins. Mol Cell Biol 1999b; 19: 2400-2407.
15. Liao C, Li SQ, Wang X, Muhlrad S, Bjartell A, Wolgemuth DJ. Elevated levels and distinct patterns of expression of A-type cyclins and their associated cyclin-dependent kinases in male germ cell tumors. Int J Cancer 2004; 108: 654-664.

2
16. Muller C, Yang R, Beck-von-Peccoz L, Idos G, Verbeek W, Koeffler HP. Cloning of the cyclin A1 genomic structure and characterization of the promoter region. GC boxes are essential for cell cycle-regulated transcription of the cyclin A1 gene. J Biol Chem 1999; 274: 11220-11228.
17. Muller C, Yang R, Idos G, Tidow N, Diederichs S, Koch OM, Verbeek W, Bender TP, Koeffler HP. c-myb transactivates the human cyclin A1 promoter and induces cyclin A1 gene expression. Blood 1999; 94: 4255-4262.
18. Muller C, Readhead C, Diederichs S, Idos G, Yang R, Tidow N, Serve H, Berdel WE, Koeffler HP. Methylation of the cyclin A1 promoter correlates with gene silencing in somatic cell lines, while tissue-specific expression of cyclin A1 is methylation independent. Mol Cell Biol 2000; 20: 3316-3329.
19. Muller-Tidow C, Readhead C, Cohen AH, Asotra K, Idos G, Diederichs S, Cauvet T, Yang R, Berdel WE, Serve H, Koeffler HP. Successive increases in human cyclin A1 promoter activity during spermatogenesis in transgenic mice. Int J Mol Med 2003; 11: 311-315.
20. Behringer RR, Crotty DA, Tennyson VM, Brinster RL, Palmiter RD, Wolgemuth DJ. Sequences 5' of the homeobox of the Hox-1.4 gene direct tissue-specific expression of lacZ during mouse development. Development 1993; 117: 823-833.
21. Packer AI, Crotty DA, Elwell VA, Wolgemuth DJ. Expression of the murine Hoxa4 gene requires both autoregulation and a conserved retinoic acid response element. Development 1998; 125: 1991-1998.
22. Heinemeyer T, Wingender E, Reuter I, Hermjakob H, Kel AE, Kel OV, Ignatieva EV, Ananko EA, Podkolodnaya OA, Kolpakov FA, Podkolodny NL, Kolchanov NA. Databases on transcriptional regulation: TRANSFAC, TRRD and COMPEL. Nucleic Acids Res 1998; 26: 362-367.
23. Latham KE, Litvin J, Orth JM, Patel B, Mettus R, Reddy EP. Temporal patterns of A-myb and B-myb gene expression during testis development. Oncogene 1996; 13: 1161-1168.
24. Goodson ML, Park-Sarge OK, Sarge KD. Tissue-dependent expression of heat shock factor 2 isoforms with distinct transcriptional activities. Mol Cell Biol 1995; 15: 5288-5293.
25. Persengiev SP, Raval PJ, Rabinovitch S, Millette CF, Kilpatrick DL. Transcription factor Sp1 is expressed by three different developmentally regulated messenger ribonucleic acids in mouse spermatogenic cells. Endocrinology 1996; 137: 638-646.
26. Bladh J, Landberg G, Richter J, Wolgemuth DJ, Persson JL. Regulation of the cyclin A1 protein is associated with its differential subcellular localization in hematopoietic and leukemic cells. Submitted.
27. Ravnik SE, Wolgemuth DJ. The developmentally restricted pattern of expression in the male germ line of a murine cyclin A, cyclin A2, suggests roles in both mitotic and meiotic cell cycles. Developmental Biology 1996; 173: 69-78.
28. Zwicker J, Lucibello FC, Wolfraim LA, Gross C, Truss M, Engeland K, Muller R. Cell cycle regulation of the cyclin A, cdc25C and cdc2 genes is based on a common mechanism of transcriptional repression. Embo J 1995; 14: 4514-4522.

3
29. Lange-zu Dohna C, Brandeis M, Berr F, Mossner J, Engeland K. A CDE/CHR tandem element regulates cell cycle-dependent repression of cyclin B2 transcription. FEBS Lett 2000; 484: 77-81.
30. Fontijn RD, Goud B, Echard A, Jollivet F, van Marle J, Pannekoek H, Horrevoets AJ. The human kinesin-like protein RB6K is under tight cell cycle control and is essential for cytokinesis. Mol Cell Biol 2001; 21: 2944-2955.
31. Uchiumi T, Longo DL, Ferris DK. Cell cycle regulation of the human polo-like kinase (PLK) promoter. J Biol Chem 1997; 272: 9166-9174.
32. Fajas L, Le Cam L, Polanowska J, Fabbrizio E, Servant N, Philips A, Carnac G, Sardet C. A CDE/CHR-like element mediates repression of transcription of the mouse RB2 (p130) gene. FEBS Lett 2000; 471: 29-33.
33. Otaki M, Hatano M, Kobayashi K, Ogasawara T, Kuriyama T, Tokuhisa T. Cell cycle-dependent regulation of TIAP/m-survivin expression. Biochim Biophys Acta 2000; 1493: 188-194.
34. Tanaka M, Ueda A, Kanamori H, Ideguchi H, Yang J, Kitajima S, Ishigatsubo Y. Cell-cycle-dependent regulation of human aurora A transcription is mediated by periodic repression of E4TF1. J Biol Chem 2002; 277: 10719-10726.
35. Sugarman JL, Schonthal AH, Glass CK. Identification of a cell-type-specific and E2F-independent mechanism for repression of cdc2 transcription. Mol Cell Biol 1995; 15: 3282-3290.
36. Badie C, Itzhaki JE, Sullivan MJ, Carpenter AJ, Porter AC. Repression of CDK1 and other genes with CDE and CHR promoter elements during DNA damage-induced G(2)/M arrest in human cells. Mol Cell Biol 2000; 20: 2358-2366.
37. Wasner M, Haugwitz U, Reinhard W, Tschop K, Spiesbach K, Lorenz J, Mossner J, Engeland K. Three CCAAT-boxes and a single cell cycle genes homology region (CHR) are the major regulating sites for transcription from the human cyclin B2 promoter. Gene 2003; 312: 225-237.
38. Haugwitz U, Wasner M, Wiedmann M, Spiesbach K, Rother K, Mossner J, Engeland K. A single cell cycle genes homology region (CHR) controls cell cycle-dependent transcription of the cdc25C phosphatase gene and is able to cooperate with E2F or Sp1/3 sites. Nucleic Acids Res 2002; 30: 1967-1976.
39. Liu N, Lucibello FC, Korner K, Wolfraim LA, Zwicker J, Muller R. CDF-1, a novel E2F-unrelated factor, interacts with cell cycle-regulated repressor elements in multiple promoters. Nucleic Acids Res 1997; 25: 4915-4920.
40. Philips A, Chambeyron S, Lamb N, Vie A, Blanchard JM. CHF: a novel factor binding to cyclin A CHR corepressor element. Oncogene 1999; 18: 6222-6232.
41. Kishore R, Spyridopoulos I, Luedemann C, Losordo DW. Functionally novel tumor necrosis factor-alpha-modulated CHR-binding protein mediates cyclin A transcriptional repression in vascular endothelial cells. Circ Res 2002; 91: 307-314.
42. Toscani A, Mettus RV, Coupland R, Simpkins H, Litvin J, Orth J, Hatton KS, Reddy EP. Arrest of spermatogenesis and defective breast development in mice lacking A-myb. Nature 1997; 386: 713-717.

4
Figure Legends
Fig. 1. Diagram of constructs for generating transgenic mice. The 8.2 kb, 4.8 kb, and 1.3
kb of Ccna1 flanking region and 0.8 kb of Ccna1 structural gene were fused to lacZ and
the polyadenylation signal and intron 1 of protamine1. Solid black lines represent Ccna1
flanking sequence or intron 1. Open boxes represent Ccna1 exon sequence. Exon 2 is
fused with lacZ, which is represented by the boxes with black dots. Boxes with vertical
lines represent the Prm1 polyadenylation signal and intron 1. The boxes with downward
diagonal stripes and with upward diagonal stripes, depicted below the cartoon of the
constructs, represent the regions of lacZ sequence used as probe templates for genotyping
or for Northern and in situ hybridization analysis, respectively. S, SalI; R, EcoRI; X,
XhoI; B, BamHI; V, EcoRV.
Fig. 2. Northern blot hybridization analysis of adult tissues from transgenic mice. Total
RNA from 8.2cyA1lacZ, 4.8cyA1lacZ, or 1.3cyA1lacZ transgenic tissues was hybridized
with a radiolabeled lacZ probe. The top lanes in each panel represent the pattern of the
lacZ hybridization signal, while the bottom lanes show the 18s rRNA ethidium bromide
staining pattern to assess equal sample loading. Results from A, 8.2cyA1lacZ, B,
4.8cyA1lacZ, and C, 1.3cyA1lacZ transgenic lines are shown.
Fig. 3. Expression of the lacZ reporter gene in representative testis tubules and
histological sections from transgenic mice. X-gal stained cells appear blue. A,D,G,J,
8.2cyA1lacZ testis, B,E,H,K, 4.8cyA1lacZ, C,F,I,L, 1.3cyA1lacZ testis and the inset in

5
A, wild type testis. A, B, C, whole mount at 12x magnification. D, E, F, stage VIII
tubule at 40x. G, H, I, stage IX tubule at 40x. J, K, L, stage XII tubule at 40x.
Fig. 4. Northern blot hybridization analysis of adult testes from transgenic mice. Total
RNA from wild type (wt) , 1.3cyA1lacZ, 4.8cyA1lacZ, and 8,2cyA1lacZ testes was
hybridized with a radiolabeled lacZ probe. The top lane represents the pattern of the lacZ
hybridization signal, while the bottom lane shows the 18s rRNA ethidium bromide
staining pattern.
Fig. 5. In situ hybridization analysis in representative histological sections from
transgenic testes. Testes sections were hybridized with antisense lacZ probe (A-D) or
sense lacZ probe (E, F). Green-colored speckled areas represent the positive signal of
reflected fluorescent light off autoradiographic silver grains. A, B, 8.2cyA1lacZ testis
sections at 40x magnification, C, D, E, 1.3cyA1lacZ testis sections at 40x, and F, a non-
transgenic testis section at 40x.
Fig. 6. Alignment of the nucleotide sequence of the 5’ flanking regions of mouse Ccna1
(mCcna1), rat Ccna1 (rCcna1) and human CCNA1 (hCCNA1). The proximal promoter
sequence of mCcna1 located A, from –517 to –438 and B, from –266 to +53 of the
putative transcriptional start site is compared to the corresponding regions in rCcna1 and
hCCNA1. Numbers indicate the position of the nucleotides of mCcna1 sequence relative
to the putative transcription start site, which is indicated by an upward bending arrow.
Nucleotides identical to mouse sequence are dark-shaded. Nucleotide differences relative

6
to mouse sequence are light-shaded, and insertions or deletions are unshaded. The region
between the solid arrowheads is highly conserved sequence. Consensus sequences for
regulatory elements in mCcna1 are labeled above the sequence. GC boxes previously
identified in the human sequence are underlined.

Table 1: Some of the transcription factors whose consensus DNA binding motifs are found within 4.8 kb of the mouse Ccna1 putative transcriptional start site or in intron 1 Transcription factor Consensus sequence
a Location
b
_______________________________________________________________________________________ SRY AAAC(A/T)A(/C) -4292, -4784, -4777, -4371, -3195 -3156, -3125, -1727, +583 GATA-1 (G/C)NNGATNNNN -4597, -1336, -979, -574, -220, +692 Sox-5 NNAACAATNN -3386, -3011, -2598, +435
c-Mybc NNNAAC(G/T)G(G/C)C -2069
YY1 NNNNNCCATNT(A/T)NNN(A/T)N -1460 HSF2 NGAANN(A/T)C(G/T) -1314, -913 USF NCACGTGN -1031, -861 AML-1a TGCGGT -319, -309, -278 CDE (G/C)GCGG -255 Sp1 G(A/G)GGC(G/T)GGG(A/T) -247 delta EF1 NNNCACCTNAN -201 CDE/CHR (G/C)GCGG/TGGAA -153/-142, -89/-79 AP-1 NTGA(C/G)TCAG -102 CHR TGGAA -26 aConsensus sequences are those listed in the TRANSFAC database [22] or, for CDE and CHR elements, were listed in Zwicker et al. [28]. bLocations of motifs are as they appear 5’ to 3’ on the sense strand, regardless of orientation, and fit the consensus sequence exactly or differ by one base. cThe c-Myb consensus sequence that is listed in the database is identical to that for A-myb and B-myb.