
Development of Subtype-Selective GABAA Receptor Compounds ...

Developmental Expression of GABAA
Receptor Subunit and GAD Genes inMouse Somatosensory Barrel Cortex
P. GOLSHANI, H. TRUONG, AND E.G. JONES*Department of Anatomy and Neurobiology, University of California, Irvine, California 92717
ABSTRACTIn situ hybridization histochemistry with radioactive cRNA probes was used to study
patterns of gene expression for a1, a2, a4, a5, b1, b2, and g2 subunit mRNAs of typeAgammaaminobutyric acid (GABAA) receptors and for 67-kDa glutamic acid decarboxylase (GAD67)mRNA in mouse barrel cortex during the period (postnatal days 1–12; P1–P12) whenthalamocortical innervation of layer IV barrels is occurring.
The a1, b2, and g2 subunit mRNAs increased substantially with age, especially in layersV and VI, and throughout the period studied, invariably had the same laminar-specificpatterns of expression. All three mRNAs were highly expressed in the dense cortical plate atP1. In layer IV after differentiation of barrels, they were expressed in cells of both barrel wallsand hollows but especially in the walls.
The a2, a4, a5, and b1 subunit mRNAs were expressed at lower levels and had differentlaminar patterns of distribution; a2 and a4 showed switches between layers over time; a5 wasinvariably associated with the subplate or its derivative, b1 with layer IV. Levels of a2 mRNAdid not change over time; a4 and b1 mRNAs increased and a5 decreased. GAD67 mRNA washighest in layer I at P1 and progressively increased in other layers.
These results suggest that postnatal development of GABAA receptors is mainly directedat the production of receptors assembled from a1, b2, and g2 subunits, with b1 contributing inlayer IV. Other subunits may be associated with receptors involved in trophic actions of GABAduring development and may give GABAA receptor-mediated responses in the developing cortextheir particular physiological profile. J. Comp. Neurol. 383:199–219, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: in situ hybridization; inhibition; cortical development; plasticity
The inhibitory neurotransmitter g-amino butyric acid(GABA) is produced by large numbers of neurons in thecerebral cortex (Hendry et al., 1987; Meinecke and Peters,1987; Schwartz et al., 1988; Beaulieu et al., 1994; Prieto etal., 1994). In rodent cortex in which GABA cells formapproximately 15% of the population (Beaulieu, 1993),GABAA receptor-mediated inhibition strongly depressesintrinsic excitability of pyramidal cells, increasing spikethresholds and abolishing repetitive firing (Connors et al.,1988). Small increases in GABAA receptor-based inhibi-tion can increase horizontal spread of excitation, leading toinitiation and propagation of epileptiform activity (Chag-nac-Amitai and Connors, 1989a,b).Cells immunoreactive for GABA or its synthesizing
enzyme, glutamic acid decarboxylase (GAD), and cellsexpressing GABAA receptor subunit mRNAs or immunore-activity for receptor subunit polypeptides are present fromthe earliest stages of cortical development (Wolff et al.,1984; Van Eden et al., 1989; Huntley et al., 1990; Cobas etal., 1991; Shaw and Scarth, 1991; Zhang et al., 1991; Del
Rio et al., 1992; Meinecke and Rakic, 1992), even beforeonset of synaptogenesis. Until recently, however, it wasdifficult to demonstrate inhibitory synapses (Blue andParnavelas, 1983a,b; Miller, 1986; Kageyama and Robert-son, 1993) or inhibitory synaptic activity in rodent cortexbefore the second postnatal week (Luhman and Prince,1991; Agmon and O’Dowd, 1992). Recent evidence in thesomatosensory cortex suggests that functional GABAergicsynapses are present from the beginning of innervation ofthe cortex by thalamocortical fibers. Inhibitory responsesto afferent stimulation can be demonstrated in layer VI ofmice at postnatal day (P) 0 and in layer IV at P2, the times
Grant sponsor: National Institutes of Health, United States PublicHealth Service; Grant number: NS 30109.*Correspondence to: Edward G. Jones, Department of Anatomy and
Neurobiology, University of California at Irvine, Irvine, CA 92717. E-mail:[email protected] 10 October 1996; Revised 7 January 1997; Accepted 13 January
1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 383:199–219 (1997)
r 1997 WILEY-LISS, INC.

3at which thalamocortical fibers first reach these layers(Agmon et al., 1993), but prior to P8, responses are weakand occur at long latency (Agmon et al., 1996). This findingmay reflect a weak excitatory drive to cortical interneu-rons, immaturity or small numbers of GABAergic syn-apses, and/or expression of unusual groupings of GABAAreceptor subunits.The functional attributes of GABAA receptors are deter-
mined by heteromeric combinations of subunit polypep-tides, encoded by 15 known genes classified in families(a1–6, b1–4, g1–3, d, r1,2) by sequence homology (MacDon-ald and Olsen, 1994; Mohler et al., 1992; Herb et al., 1992;Wisden et al., 1992). During fetal and early postnatal life,there is a switch in the subunit genes that is most highlyexpressed in the cortex. The a2, a3, a5, and b2 mRNAs areinitially expressed at the highest levels, but decline andare replaced by high levels of a1, a4, and b2 mRNAs; g2mRNA remains constant (Laurie et al., 1992). In the firstfew postnatal days, many barrel cortex neurons thatinitially display a2 receptor subunit immunoreactivitybegin to display a1 receptor subunit immunoreactivityinstead (Fritschy et al., 1994). The extent of changes of thistype is unknown. The present study examined patterns ofreceptor subunit gene expression in the barrel field duringthe stages when inhibitory neuronal function is estab-lished, when innervation by thalamocortical fibers is occur-ring and when barrel cortex is susceptible to activity-dependent plasticity (Schlaggar et al., 1993; Fox, 1994;Micheva and Beaulieu, 1995a,b).
MATERIALS AND METHODS
In situ hybridization histochemistry
P1 or P4 ICR mice were anesthetized by hypothermia,and the brains were removed and fixed by immersion in 4%paraformaldehyde in 0.1 M phosphate buffer. P7, P12, andadult (.P32) mice were given an overdose of Nembutaland perfused transcardially with 0.1 M phosphate (pH 7.4)buffer followed by 4% paraformaldehyde in 0.1 M phos-phate buffer (pH 7.4). Brains were postfixed overnight in4% paraformaldehyde in 0.1 M phosphate buffer and thencryoprotected in 30% sucrose in 0.1 M phosphate buffer.Brains were frozen directly on dry ice (for parasagittalsections), or the cerebral cortex was dissected out andflattened as it was frozen on dry ice (for tangentialsections). Serial sections were cut at 25 µm on a slidingmicrotome in parasagittal or tangential planes and col-lected in three groups: two groups were collected into 4%paraformaldehyde in 0.1 M phosphate buffer for in situhybridization histochemistry, and the other group wascollected into 0.1 M phosphate buffer (pH 7.4) for cyto-chrome oxidase histochemistry (CO).Sections reserved for in situ hybridization histochemis-
try were mounted on Vectabond-coated slides, dried over-night, lipid extracted with a 1:1 mixture of 100% chloro-form and 100% ethanol, rehydrated through descendingconcentrations of ethanol, and allowed to dry overnight.The slide-mounted sectionswere then pretreated by succes-sive incubations in 0.1 M glycine in 0.1 M phosphate buffer(pH 7.4), 0.5 mg/ml of proteinase K (pH 8.0) for 30 minutesat 37°C, 0.25% acetic anhydride in 0.1 M triethanolamine(pH 8.0), and 23 saline sodium citrate (SSC). Sectionswere then incubated in hybridization solution containing50% formamide, 10% dextran sulfate, 0.7% Ficoll, 0.7%polyvinyl pyrolidone, 0.5 mg/ml yeast tRNA, 0.33 mg/ml
denatured herring sperm DNA, 20 mM dithiothreitol(DTT), and 1 3 106 cpm/ml of a[35S] or [33P] antisensecRNA probe to a1, a2, a4, a5, b1, b2, and g2 GABAAreceptor subunit variants and to 67-kDa GAD (GAD67)mRNA for at least 40 hours at 60°C. The GABAA receptorsubunits chosen represent those most highly expressed inthe adult cortex (a1, a2, and g2), those that show thestrongest developmental regulation (a2, a4, and a5), andone that is reported to be weakly expressed at all ages (b1).Sections from the same brain were always treated simulta-neously and exposed on the same sheet of film. The cloningof cDNAs and preparation of cRNA probes have beendescribed previously (Huntsman et al., 1994, 1995). InNorthern blots, each GABAA receptor subunit cDNArecog-nized transcripts with distinct molecular weights corre-sponding to the a1, a2, a4, a5, b1, b2, and g2 subunits. Thespecificity is reflected in the results of hybridization withmany of the probes, e.g., the b1 and b2 probes (see Results,Figs. 1, 3). Although they share a high degree of sequenceidentity (Ymer et al., 1989), the relative distributions ofsignal with these two probes were quite different in boththe adult and developing mouse cortex. In cases wheredistribution patterns are similar, cross hybridization can-not be automatically ruled out, although the results ofNorthern blotting are against this. All probes were ofsimilar length and had similar G/C ratios and specificactivities. Sense cRNA probes applied to control sectionsresulted in light background labeling only.Following hybridization, slide-mounted sections were
washed in 43 SSC at 60°C, digested with 20 mg/mlribonuclease A (pH 8.0) for 30 minutes at 45°C, andwashed through descending concentrations of SSC with 5mM DTT to a final stringency of 0.13 SSC at 60°C for 1hour. Slides were allowed to dry and then exposed toAmersham b Max film for 3–12 days. After development ofthe film, sections were dipped in Kodak NTB2 emulsiondiluted 1:1 with water and exposed for 4 weeks at 4°C. Theautoradiographs were then developed in Kodak D19, fixedin Kodak Rapid Fixer, washed in water, stained throughthe emulsion with cresyl violet, and coverslipped in DPX.Film autoradiograms of sections containing somatosen-
sory barrel cortex were selected by reference to adjacentCO-stained sections. These autoradiograms were quanti-fied by densitometry by using a microcomputer imagingdevice (MCID/M4; Imaging Research, Inc., St. Catherine’s,Ontario). Optical density readings were taken in scans ofdefined width through the layers from the pia mater to thewhite matter. All readings illustrated were taken fromadjacent sections of the same brain at each time point andexposed on the same sheet of film. Variations betweenanimals at each age did not differ by more than 5–10%.Absolute values (nCI/g) of radioactivity were obtainedfrom the integrated optical density readings by referenceto [14C] plastic standards (Amersham, Arlington Heights,IL) exposed on the same sheet of film.
Immunocytochemistry
P1, P4, P7, P12, and adult ICR mice were eitheranesthetized by hypothermia (P1 and P4), decapitated, thebrains removed and fixed by immersion in 2% paraformal-dehyde and 0.1% glutaraldehyde in 0.1M phosphate buffer(pH 7.4) or given an overdose of Nembutal and perfusedtranscardially with 0.1 M phosphate (pH 7.4) buffer fol-lowed by the same fixative. All brains were removed andpostfixed overnight in 2% paraformaldehyde in 0.1 M
200 P. GOLSHANI ET AL.

phosphate buffer and then cryoprotected in 30% sucrose in0.1 M phosphate buffer. Brains were frozen to facilitateresectioning in parasagittal or tangential planes as de-scribed above, and serial sections were cut at 25 µm on asliding microtome and collected in three groups: twogroups were set aside for immunocytochemistry and onegroup for cytochrome oxidase histochemistry. Sectionsreserved for immunocytochemistry were mounted on gela-tin-coated slides, allowed to dry overnight, and thenpreincubated in a solution of 3% normal horse serum and1% Triton X-100 in 0.1 M phosphate buffer for 1 hour. Thesections were then incubated in 3% normal horse serum,1% Triton X-100, and a 1:500 dilution of a previouslycharacterizedmousemonoclonal antibody directed againstthe b2/b3 subunits of the GABAA receptor (de Blas et al.,1988) in 0.1 M phosphate buffer at 4°C for 72 hours.Section-mounted slides were then washed three times for20 minutes in 0.1 M phosphate buffer and then incubatedin a solution of 3% normal horse serum, 1% Triton X-100,and 1% horse anti-mouse secondary IgM antibody for 1hour. Fifteen minutes before sections were removed fromthe secondary antibody solution, a solution of acetyl-avidinbiotinylated peroxidase complex (ABC solution; Elite, Vec-tor ABC) was prepared at 2 drops/5 ml 0.1 M phosphatebuffer and allowed to react for 30 minutes prior to incuba-tion. Slides were then washed in 0.1 M phosphate buffer,three times for 10 minutes each, and then incubated in theABC solution for 1 hour. Slides were washed subsequentlythree times for 20 minutes each with 0.1 M phosphatebuffer and stained with the Vector VIP peroxidase sub-strate kit (2 drops/5 ml). Slides were then washed twice for10 minutes each with 0.1 M phosphate buffer, dehydratedthrough ascending concentrations of ethanol and xylene,and coverslipped with DPX. Control sections stained byomitting the primary antibody revealed no specific stain-ing.
RESULTS
Laminar distribution
a1 subunit. At all postnatal ages studied, autoradio-graphic labeling with the a1, b2, and g2 subunit-specificprobes showed the highest density compared with autora-diographic labeling by the other subunit-specific probes,with a1 usually showing the highest density of labeling,followed by b2 and then g2 (Figs. 1–5).At P1, the strongest labeling was seen in the dense
cortical plate (DCP; Figs. 1, 2, 4). The marginal zoneshowed weak labeling and was clearly demarcated fromthe DCP, while deep to the DCP, both layers V and VIshowed weak, homogeneous labeling slightly more densethan the labeling seen in the marginal zone but far weakerthan labeling of the DCP. Examination of emulsion autora-diographs showed enrichment of labeling in superficialand middle subdivisions of the DCP, compared with asomewhat lower level in the deep DCP. Very denselypacked small grain clusters 10–12 µm in diameter wereobserved over neuronal somata in the DCP and in layer VI,and loosely packed larger grain clusters of 15–20 µm indiameter were observed over somata in layer V, reflectingthe decreased packing density and larger size of layer Vneuronal somata.There was a marked increase in density of hybridization
with the a1 probe in all the layers of the barrel cortexbetween P1 and P4 (Figs. 3, 4), when barrels emerge in
layer IV and the cortical layers are all formed. At P4,layers III and IV showed the highest levels of hybridiza-tion; labeling of the two layers was homogeneous and wasdistinct from a band of lower intensity labeling in layer II.Hybridization signal was faintest in layer I and intermedi-ate in layers V and VI. In emulsion autoradiographs,labeling of layers III, IV, and VI was in the form of grainclusters 10–12 µm in size, but labeling of layer Vwas in theform of larger grain clusters of 16–18 µm in diameter,again reflecting labeling of larger cell bodies in layer V.Between P4 and P7 (Fig. 4), there was little change in
the distribution and intensity of hybridization for the a1subunit-specific probe. The contrast between the intensesignal in layer IV and the intermediate signal in layer Vbecame even more apparent. Barrel morphology was moredistinct in layer IV, and there was a slightly lower densityof label in the barrel hollows.Hybridization intensity for thea1 subunitmRNAsteadily
increased in all layers of the somatosensory cortex be-tween P7 and P12 (Figs. 4, 5). The most dramatic increasein intensity was observed in layer VI, with an almostdoubling of labeling intensity between P7 and P12. The a1mRNA levels were still highest in layers II, III, and IV,with layer III having the most prominent staining. Theborder between layers IV and V was as distinct as at allyounger ages, and the border between layers V and VIbecame more prominent because of the increase in thehybridization intensity in layer VI. In emulsion autoradio-graphs, tightly packed, small grain clusters 10–12 µm indiameter were found in layers II, III, and IV, and looselypacked large grain clusters 17–20 µm in diameter werefound in layer V. In layer VI, both large and small grainclusters 9–22 µm in diameter were found.
a2 subunit. Autoradiographic labeling for the a2 sub-unit mRNA was extremely faint at all postnatal ages,showing the weakest levels of hybridization among all theGABAA receptor subunit riboprobes (Figs. 1–5). Close-to-background levels of hybridization were observed in alllayers of the presumptive barrel cortex at P1 and P4. AtP7, a faint laminated appearance was seen, with a homoge-neous band of somewhat more intense label in layers II,III, and superficial layer IV. At P12, labeling intensityslightly increased in all cortical layers, with layer IIhaving the most prominent labeling; labeling of all otherlayers was homogeneous.
a4 subunit. Overall, autoradiographic labeling for thea4 subunit was faint but somewhat stronger than labelingfor the a2 subunit.At P1, autoradiographic signal was onlyslightly above background level, and labeling was fairlyhomogeneous in all layers of cortex (Figs. 1, 3). Between P1and P4, there was a marked increase in labeling intensity,with layers II, III, and superficial layer IV appearing as adistinct band between the faintly labeled layers I and V(Fig. 4). There were background levels of labeling in layerVI. At P7, there was a steady increase in hybridizationintensity in all cortical layers, with layers III and IVshowing the greatest enhancement of signal. There wasfaint labeling of all the other layers, although layers V andVI showed a slight increase in labeling intensity. BetweenP7 and P12, there was little change in hybridizationintensity, although the distribution of autoradiographiclabeling was distinct from that seen at P7 (Fig. 5): arelatively strong signal in layer IV, well demarcated byfaint staining of layers III and V. Hybridization in layer Iwas very faint, followed by a slightly more intense label in
DEVELOPMENT OF GABAA RECEPTOR SUBUNITS 201

Fig.1.
Pairedphotom
icrographsfrom
theposterodorsalpartofthepresumptivesomatosen-
sory
cortex
atpostnatal
day1(P1),show
ingNisslstaining(leftmem
berofeach
pair)ofthe
perpendicularsectionsfrom
whichtheautoradiograms(rightm
emberofeach
pair)w
eretaken.
TypeA
g-aminobutyricacid
(GABAA)receptor
subunit
mRNA
or67-kDaglutamic
acid
decarboxylase(GAD67)mRNA
isindicatedat
topleft
ofeach
autoradiograph.I,
V,VI,
differentiated
layers
ofcerebral
cortex;cp,undifferentiated
cortical
plate;sp,subplate:St,
striatum;W
M,w
hitematter.Scalebar
5250µm.

layers II and III. Deep to layer IV, hybridization inten-sity was very weak in layer V and was slightly stronger inlayer VI.
a5 subunit. Hybridization patterns with the a5 sub-unit-specific riboprobe were dramatically different fromthe hybridization patterns seen with all other subunit-specific probes; areas of greatest enrichment of signalcoincided with areas of relatively low cell density (Figs. 1,3, 5). Furthermore, there was a marked and steadydecrease in hybridization intensity in these particularlayers throughout early postnatal development. At P1,autoradiographic labeling was most intense in the newlydifferentiated layer V, with staining most prominent insuperficial and middle zones of layer V (Figs. 1, 2).Hybridization intensity in layer V at this age was amongthe highest seen for all the subunit-specific probes; onlyriboprobes for the a1 and g2 subunit mRNAs showedslightly higher levels of hybridization. The high level of a5signal in layer V was striking because the cell packingdensity in layer V was among the lowest of all corticallayers in the P1 mouse, reflecting intense specific labelingof layer V neurons. In emulsion autoradiographs, interme-diate and large grain clusters 15–20 µm in diameter wereseen over the large somata of the layer V cells. Superficialto layer V, the DCPwas labeled half as intensely as layer Vwas. Superficial DCP was labeled most weakly, and therewas increasing density of labeling in the intermediate anddeep layers of the DCP. Densely packed small grainclusters 8–10 µm in diameter were observed over smallneurons in this layer. The border between the DCP andlayer V was distinct, and there was little labeling of the
marginal zone. Layer VI was labeled about as intensely asthe superficial DCP, although labeling was slightly moreintense than the intermediate level of hybridization indeep layer V. Small and intermediate grain clusters 10–15µm in diameter were observed over somata in layer VI.Beneath layer VI, the cortical subplate (‘‘layer VIb’’) washeavily labeled, differentiating it from the moderatelylabeled layer VI (Fig. 2B,C). Grain clusters 14–18 µm indiameter were observed, reflecting labeling of intermedi-ate and large neuronal somata approaching the size oflarge layer V cells.Between P1 and P4, there was a moderate reduction in
labeling of layer V (Figs. 1, 3, 4, 7); labeling of other corticallayers remained unchanged. At P4, the laminar pattern ofhybridization was the same as that at P1, with oneimportant exception: labeling of the subplate was moredistinct and could be readily distinguished from theweakerlabeling of layer VI. Intermediate grain clusters 14–16 µmin diameter were found overlying neuronal somata in layerVIb. Labeling of layer V was the most intense of all corticallayers, showing loosely packed large grain clusters 18–25µm in diameter over the large neurons of the layer.Between P4 and P12, there was a considerable decrease
in hybridization levels for the a5 subunit-specific probe inall cortical layers (Figs. 4, 5, 7). Labeling in layers I–IV andVI approached background levels, and labeling in layer V,although still twice that of other cortical layers, wasthreefold less intense at P12 than at P4. Deeper regions oflayer V were considerably more enriched with signal thanthe superficial and middle regions of the layer. Hybridiza-tion intensity was greatest in layer VIb in P4–P7 animals
Fig. 2. A,B: Darkfield photomicrographs through the presumptive somatosensory cortex at P1,showing different patterns of expression of a1 and a5 subunit mRNAs. C: Higher magnificationbrightfield photomicrograph from B, showing cells expressing a5 subunit mRNA in the subplate (sp).Scale bar 5 100 µm inA,B, 50 µm in C.
DEVELOPMENT OF GABAA RECEPTOR SUBUNITS 203

and in a layer VIb/subcortical white matter stratum of theolder animals. At all ages, it was slightly more intensethan the labeling of layer V. Layer VI proper was alwaysweakly labeled. In emulsion autoradiographs, no concen-
trated grain clusters associated with cell bodies werefound in layers I–IV or VI. In layer V and in the subplate/superficial white matter stratum, grain clusters lay overlarge cells and were mostly 18–20 µm in diameter.
Fig. 3. Left and right: Paired photomicrographs from the postero-medial barrel subfield at P4. Barrels are just differentiating in layerIV. Left member of each pair shows Nissl staining of the perpendicularsections from which the antoradiographs (right member of each pair)
were taken. Subunit mRNAs or GAD mRNA indicated at top left ofeach autoradiograph. I–VI, layers of cerebral cortex; VIb, derivative ofsubplate; St, striatum; WM, white matter. Scale bars 5 250 µm.
204 P. GOLSHANI ET AL.

a1 subunit. Themain distinguishing feature of hybrid-ization for b1 subunit mRNA was weak to moderatelabeling overall, with intensity of hybridization at levelsintermediate between that of b2 and a4 subunit probes(Figs. 1, 3, 4, 7). There was fairly selective labeling of layerIV at ages when this layer became identifiable. Through-out postnatal development, there was a notable increase in
hybridization intensity for b1 subunit mRNA in all corticallayers. Between P7 and P12, there was a dramatic in-crease in labeling of layers V and VI relative to the stronglabeling of layer IV, and hybridization levels in layer VIapproached those in layer IV.At P1, the most intense labeling for the b1 subunit
mRNA was found in the DCP (Figs. 1, 3). There was a
Figure 3 (Continued.)
DEVELOPMENT OF GABAA RECEPTOR SUBUNITS 205

slight increase in labeling of the deeper part of the DCPrelative to the superficial and middle portions. The DCPwas well delineated in film autoradiographs from theoverlying marginal zone and underlying layer V. Layer Vwas labeled with half the intensity of the DCP but withgreater intensity than layer VI. Labeling in layer VI wasnot much greater than the faint labeling of the marginalzone. Examination of emulsion autoradiographs revealedsmall grain clusters 10–12 µm in diameter over neurons inthe DCP and large grain clusters 18–20 µm in diameterover neurons in layer V.Between P1 and P4, there was little change in the
hybridization patterns for b1 mRNA. At P4, the mostintense autoradiographic label was found in layers II–IV,with layer IV being slightly more enriched than the othertwo layers (Figs. 3, 4). Superficial to layer II, there wasonly faint labeling of layer I. Layer V was labeled abouthalf as intensely as layer IV, with the superficial portionsof layer V being labeled at slightly higher levels than thedeep aspects. Labeling of layer VI was similar in intensityto that of layer V. Small grain clusters 10–12 µm indiameter were observed over layer IV cells in emulsionautoradiographs.At P7, there was little change in hybridization patterns
for the b1 riboprobe. The most intense band of label was in
layer IV; the hybridization band in layer IV was moredistinct at P7 than at P4 because of the decrease in therelative labeling of layers II and III. Labeling intensity inlayers V and VI was about half as intense as the labeling inlayer IV, with some superficial to deep increase. There wasonly faint labeling in layer I. In emulsion autoradiographs,intermediate grain clusters 14–16 µm in diameter werefound in layer IV.Between P7 and P12, there was a considerable change in
both the intensity and pattern of hybridization for the b1riboprobe (Figs. 4, 5). Signal strength virtually doubled inall cortical layers, with the greatest increase being in deeplayer VI, where the intensity of hybridization almostequaled that of layer IV. Layer IV still showed the highestlevel of hybridization, although signal intensities in layersII and III had risen to levels that were close to those inlayer IV. Examination of emulsion autoradiographs re-vealed intermediate-sized grain clusters 14–16 µm indiameter over neuronal somata in layer IV. Labeling oflayers II and III merged into the hybridization band oflayer IV, but layer IV showed a clear border with layer V.Large grain clusters 18–22 µm in diameter were found inlayer V overlying large neuronal somata. Hybridizationintensity in layer V was almost equal to that in layer VI.There were peaks of enhanced signal in deep layer V and
Fig. 4. Relative levels of hybridization for the seven GABAA receptor subunit mRNAs and GAD67mRNA across the thickness of the barrel cortex at P1, P4, P7, and P12, as obtained from optical densitymeasurements in transepts of constant width from pia mater to white matter. Integrated optical densityreadings (I0D) were converted to levels of radioactivity (nCi/g) by reference to [14C] standards.
206 P. GOLSHANI ET AL.

deep layer VI. Small grain clusters 10–12 µm in sizeoverlying small neurons were found in layer VI. There wasonly faint labeling in layer I.
b2 subunit. Hybridization of b2 subunit-specific probesled to a pattern of autoradiographic labeling that re-sembled that seen with the a1- and g2-specific riboprobes(Figs. 3–5, 7). Hybridization with the b2 probe was usuallyless intense than hybridization with the a1 subunit-specific probe, except at P7. Hybridization of the b2 probe,however, was more intense or equal in intensity comparedwith hybridization of the g2 subunit-specific probe. Therewas a substantial increase in b2 hybridization intensitiesfrom P1 to P7 and a moderate decrease between P7 andP12 (Figs. 3–5). The most dense hybridization signal wasfound in the DCP at P1 and in layers II–III in all laterages. The developmental changes in b2 hybridizationpatterns resembled the developmental changes in a1 andg2 hybridization patterns: between P7 and P12, there wasa relative increase in labeling of layers V and VI comparedwith labeling of layers II–IV.At P1, the most dense band of hybridization of the b2
riboprobe was over the DCP, and the intensity of labelingwas similar to that of the g2 riboprobe (Figs. 1, 4, 7). Inemulsion autoradiographs, mainly small grain clusters8–12 µm in diameter were found in the DCP. There was amoderate decrease of signal strength in layer V (Figs. 3, 4).Grain clusters in layer V were variable in size and were
12–20 µm in diameter. Deep layer V showed less labelingthan superficial and middle layer V. Hybridization levelsin layer VI were similar to those in the DCP. Small grainclusters 10–12 µm in diameter were associated withneuronal somata in layer VI.Between P1 and P4, there was a considerable increase in
the intensity of hybridization in all cortical layers. At P4,the most intense band of labeling was a homogeneousstripe coextensive with layers II–IV. Hybridization inten-sity of these layers was almost twice that in the DCP at P1.Labeling of layer V was about half the intensity of that inlayers II–IV. There was homogeneous labeling of layers Vand VI, with label slightly decreasing deeper in the cortex.Labeling of layer I was faint.Between P4 and P7, there was a moderate increase in
hybridization intensity in all cortical layers for the b2subunit-specific riboprobe (Fig. 4). Levels were highest atP7 and declined thereafter to levels similar to those foundat P4. There was little change in the patterns of hybridiza-tion except for a relative increase in labeling of layers Vand VI vs. that of layers II–IV. The most dense band oflabeling was still found in layers II–IV, with small tointermediate-sized grain clusters 10–15 µm in diameter inemulsion autoradiographs. The border between layers IVand V remained distinct, although layers II and III wereindistinguishable from layer IV on film autoradiographs.Labeling of layers V and VI remained homogeneous. In
Figure 4 (Continued.)
DEVELOPMENT OF GABAA RECEPTOR SUBUNITS 207

layer V, large grain clusters 20–25 µm in diameter re-flected labeling of large pyramidal cells. Mostly intermedi-ate to large grain clusters 15–20 µm in diameter werefound in layer VI. There was only faint labeling of layer I.
Between P7 and P12, b2 hybridization levels declined tolevels found at P4 (Figs. 3–5). However, hybridizationpatterns with the b2 riboprobe at P12 were considerablydifferent from those found at P4. The most significant
Fig. 5. Left and right: Paired photomicrographs from the posteromedial barrel subfield at P12.Cortical layers have fully differentiated. Left member of each pair shows Nissl staining of theperpendicular sections from which the autoradiograms (right member of each pair) were taken. VIb,derivative of subplate; St, striatum; WM, white matter. Scale bars 5 250 µm.
208 P. GOLSHANI ET AL.
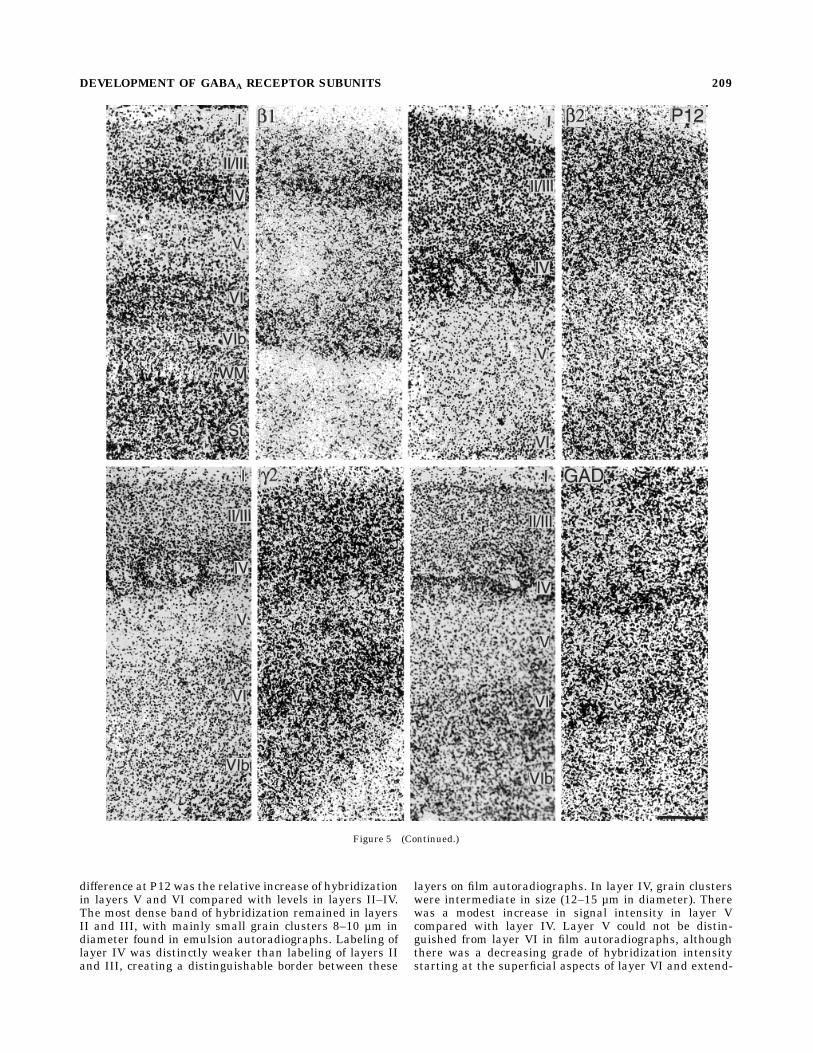
difference at P12 was the relative increase of hybridizationin layers V and VI compared with levels in layers II–IV.The most dense band of hybridization remained in layersII and III, with mainly small grain clusters 8–10 µm indiameter found in emulsion autoradiographs. Labeling oflayer IV was distinctly weaker than labeling of layers IIand III, creating a distinguishable border between these
layers on film autoradiographs. In layer IV, grain clusterswere intermediate in size (12–15 µm in diameter). Therewas a modest increase in signal intensity in layer Vcompared with layer IV. Layer V could not be distin-guished from layer VI in film autoradiographs, althoughthere was a decreasing grade of hybridization intensitystarting at the superficial aspects of layer VI and extend-
Figure 5 (Continued.)
DEVELOPMENT OF GABAA RECEPTOR SUBUNITS 209

ing into the deep layer VIb stratum and the white matter.In layer V, large grain clusters 18–20 µm in diameter werefound overlying large pyramidal neurons; small, interme-diate, and large grain clusters 10–20 µm in diameter werefound in layer VI.
g2 subunit. Hybridization patterns for the g2 subunit-specific probe resembled hybridization patterns for the a1-and b2-specific probes, with the exception that at P7 andP12 the most intense labeling for g2mRNAwas in layer VIin contrast to the hybridization patterns for a1 and b2mRNAs, which labeled layers II–IV the most intensely(Figs. 1, 3–5, 7). The g-2 subunit hybridization levels weregenerally less than a1 subunit hybridization levels andabout equal to b2 subunit hybridization levels. The maindevelopmental change in the pattern of hybridization forg2 subunit mRNA occured between P4 and P7, with aconsiderable relative decrease in labeling of layers II–IVand a comparable increase in labeling of layers V and VI.At P1, the most dense band of g2 subunit-specific
hybridization was over the DCP (Figs. 1, 4, 7). Intensity ofg2 mRNA hybridization in the DCP was somewhat lessintense than for the a1 subunit-specific riboprobe butslightly more intense than for the b2 subunit-specificriboprobe. A homogeneous band of label extended from thesuperficial border of the DCP to the middle of layer V.There was a slight drop of hybridization intensity in themiddle of layer V, but hybridization levels increased againin deep layer V. Overall, labeling of layer V was lessintense and less homogeneous than labeling of the DCP.No boundary between layer V and layer VI could bedistinguished on film autoradiographs. There was a de-creasing grade of hybridization signal that began in super-ficial layer VI and extended to deep layer VIb and thewhite matter. There was a moderate level of hybridizationoverlying the marginal zone.Between P1 and P4, there was little change in the
intensity of hybridization for g2 subunit mRNA, althoughthere were moderate changes in the patterns of hybridiza-tion. The most intense levels of hybridization were foundin layers II and III. Labeling of layer IV was somewhatweaker than labeling of layers II and III (Figs. 3, 4, 7).Labeling of layer IV was continuous with similar labelingof superficial and middle layer V. There was a moderatedecrease in signal strength in deep layer V. Labeling ofdeep layer V was homogeneous with labeling of layer VI.Hybridization levels were low in layer I.There was considerable change in hybridization pat-
terns for the g2 subunit mRNAbetween P4 and P7 (Fig. 4).At P7, the most intense band of hybridization was found inlayer V and superficial layer VI, although labeling oflayers II–IV was almost equally intense. There was aslight increase of signal in the deep aspects of layer Vcompared with its superficial aspects. The boundary be-tween layer V and layer VI could not be discerned on filmautoradiographs, and the peak level of hybridization ap-peared to fall on the border of layers V and VI. Labeling oflayer VI was not homogeneous; there was a steep decreasein hybridization intensity beginning superficially in layerVI and extending into layer VIb and white matter.The dramatic changes in hybridization pattern for g2
subunit mRNA that occurred between P4 and P7 contin-ued through P12 (Fig. 5). There was a relative decrease oflabeling of layers V and VI compared with labeling oflayers II–IV. The most dense band of hybridization was ahomogeneous stripe of hybridization overlying deep layer
V and superficial layer VI. There was a sharp drop inhybridization levels in middle and deep layer VI, andlabeling fell to background levels in the deepest aspects oflayer VI. In emulsion autoradiographs, mainly intermedi-ate and large grain clusters 15–20 µm in diameter werefound overlying layer VI neuronal somata. The boundarybetween layer IV and layer V was distinct on film autora-diographs, with an initial decrease of hybridization inten-sity in superficial layer V compared with layer IV, followedby a steep increase in hybridization signal. Large grainclusters 20–25 µm in diameter were found overlying largepyramidal neurons of layer V. Layer IV was prominentbecause of weaker hybridization in comparison with theadjoining layers III and V. Layers II and III were labeledwith equal intensity, although there was a sharp peak ofincreased hybridization in the middle of layer III. Small,intermediate, and large grain clusters 10–20 µm in diam-eter were found in layers II–IV. Layer I was only faintlylabeled.GAD. Hybridization patterns with the GAD67-specific
riboprobe changed dramatically between P1 and P7 in thebarrel cortex (Figs. 1, 3–5, 7). GAD gene expression washighest in the marginal zone at P1 but substantiallyincreased in all the layers of the cortex in the subsequent 7days. At P7, the most dense band of hybridization coin-cided with layers II and III, with slightly less intensehybridization in the other cortical layers.At P1, the densest band of hybridization was a thin
stripe of intense signal coinciding with the marginal zone(Figs. 1, 4, 7). The peak of hybridization was very sharpand ended abruptly at the border with the DCP. Examina-tion of emulsion autoradiographs revealed intermediate-sized grain clusters 14–16 µm in diameter overlying cellsin themarginal zone. Hybridization levels in the DCPwereless than half that of the marginal zone, and layers V andVI could not be readily distinguished from the DCP on filmautoradiograms. There was a mild increase in hybridiza-tion levels in layer V compared with the DCP and a moremarked decrease of labeling in layer VI, but occasionallabeled cells were still detectable in the subplate.Between P1 and P7, there was a dramatic increase in
hybridization levels for GAD in all cortical layers (Figs. 3,4). The most intense band of hybridization was over layersII and III; hybridization signal increased more than two-fold in these layers. There was an abrupt drop of hybridiza-tion level in superficial and middle layer IV and an equallyabrupt increase of signal levels in deep layer IV. Hybridiza-tion intensity remained high in layer V and in the mostsuperficial aspects of layer VI but precipitously dropped tonear background levels in middle and deep aspects of layerVI. Labeling of layer I remained intense.Between P7 and P12 (Figs. 4, 5), there was a moderate
change in hybridization patterns with the GAD-specificriboprobe. The highest peak of hybridization was in layerII and superficial layer III. There was a significant reduc-tion of hybridization signal in the middle of layer III. Theborder between layers III and IV was not distinct. Inemulsion autoradiographs, mainly small grain clusters10–12 µm in diameter were found over cells in layers II–IV.Hybridization levels increased considerably in deep layerIV and fell again in layer V before rising slightly in layerVI. Intermediate-sized grain clusters 14–16 µm in diam-eter were found in layer VI.
210 P. GOLSHANI ET AL.

Tangential distribution in layer IV
The somatotopic arrangement of the barrels was distinctin film and emulsion autoradiographs in sections hybrid-ized with a1, b1, b2, and g2 subunit-specific riboprobesfrom P4, the age when barrels first become evident in CO-and Nissl-stained preparations. The a2, a4, and a5 sub-unit mRNAs, although present, did not have barrel-specific patterns of label (Figs. 8, 9). The a1, b1, b2, and g2subunit mRNAs were expressed at highest levels in thecell-dense and CO-weak barrel wall, and at significantlylower levels in the cell-poor CO-dense barrel hollow (Fig.6). In general, expression levels were higher in the walls ofbarrels corresponding to the more rostral vibrissae, wherecell density was highest. Overlaying camera lucida draw-ings of GABAA receptor subunit distributions and ofcytochrome oxidase positive patches in adjacent sections
revealed the complementary pattern: subunit-specific ribo-probes most densely labeled areas around CO patches,overlapping only slightly into the periphery of each COpatch.Examination of emulsion autoradiograms of tangential
sections through layer IV hybridized with a1- and g2-specific riboprobes revealed small and intermediate-sizedgrain clusters 10–16 µm in diameter over neuronal somata(Fig. 6A,F). Although there were fewer cells labeled in thebarrel hollows, some of the cells in the hollows werelabeled very densely. In contrast, labeling of cells in thebarrel walls was fairly constant, with an intermediatenumber of grains overlying most cells.In emulsion autoradiograms of tangential sections
through layer IV hybridized with the b2 subunit-specificriboprobe, small to intermediate-sized grain clusters 10–16
Fig. 6. In situ hybridization for a1, b1, b2, and g2 subunit mRNAsat P12. A,B: Brightfield and darkfield photomicrographs of the samebarrel showing high expression of a1 mRNA, especially in cells ofbarrel walls.C,D:Brightfield photomicrograph of a barrel wall (C) andlower magnification darkfield photomicrograph of the whole barrel,
showing b2 subunit mRNA localization. E: Low level of b1 subunitmRNAhybridization in cells of a barrel wall. F: The g2 subunit mRNAlocalization in cells of a barrel wall. Scale bar 5 75 µm inA,B,D, 15 µmin C, 10 µm in E, 5µm in F.
DEVELOPMENT OF GABAA RECEPTOR SUBUNITS 211

Fig.7.
Graphsshow
ingrelative
densitiesofhybridization
forthesevenGABAAreceptorsubunitmRNAsandGAD67mRNAinthelayersofthecerebralcortex
during
theperiod
ofinnervation
ofthebarrelcortex
bythalam
ocorticalfibers.LevelsareexpressedinnCi/g,asobtained
from
integrated
opticaldensity(IOD)m
easurements.

Fig.8.
Tangentialsectionsthrough
layerIV
ofthebarrelfieldatP12.Top
rowshow
sNisslstainingof
thesectionsfrom
whichantoradiogramsofthemiddle
rowweretaken.B
ottom
rowshow
scytochrome
oxidase(CO)stainingof
animmediately
adjacentsection.Expressionof
a1,
b1,
b2,
and
g2subunit
mRNAsisdensestinthebarrelwalls.Scalebar
5500µm.

µm in diameter were found overlying cells in both barrelwalls and hollows (Fig. 6C,D). Most cells in the barrelwalls were labeled, whereas only a few labeled cells couldbe found inside barrel hollows.In emulsion autoradiograms of tangential sections
through layer IV hybridized with the b1-specific riboprobe,small to intermediate-sized grain clusters 10–16 µm indiameter were found overlying the barrel walls (Fig. 6E).These cells were only faintly labeled. There were nolocalized grain clusters over cells in the barrel hollows.Examination of emulsion autoradiograms of tangential
sections through layer IV hybridized with a2-, a4-, anda5-specific riboprobes revealed no concentrated grain clus-ters in relation to the barrels.
Immunocytochemical distribution of theb2/b3 subunit protein in layer IV
Between P0 and P5, immunocytochemical staining forthe b2/b3GABAA receptor subunits revealed light homoge-neous staining of the neuropil, with no clustering ofreceptor immunostaining. After P5, immunostaining forGABAA receptors in tangential sections of layer IV re-vealed staining of the neuropil of the barrel hollows butlittle staining of the barrel wall; the pattern matched theCO staining pattern of the barrels (Fig. 10). High magnifi-cation examination of the immunostaining in the barrelhollow revealed outlines of neuronal somata surroundedby dense immunoreactivity, which was often punctate.Comparison with adjacent CO-stained sections by overlay-ing camera lucida drawings revealed that areas of in-
creased GABAA receptor immunoreactivity coincided al-most perfectly with the CO patches (Fig. 10). This patternof immunostaining emerged at about P5, which is coinci-dent with the emergence of CO-stained barrels. At P5, inaddition to the staining of the neuropil in the barrelhollows, very light staining of the cytoplasm of many cellbodies could be seen in both the barrel hollows and barrelwalls.
DISCUSSION
Genes encoding the a1, b2, and g2 subunits of theGABAA receptor are expressed at highest levels in thebarrel cortex during postnatal development. These sub-units show lamina-specific, developmentally regulated pat-terns of expression, and a1 expression levels tend to begreater than of b2 or g2. The other four subunit mRNAsexamined (a2, a4, a5, and b1) are expressed at much lowerlevels than a1, b2, and g2, but they also display laminar-specific and developmentally regulated expression pat-terns. The a5 mRNAs display the most unique pattern ofexpression, with hybridization densities highest in thecortical subplate or its derivative.
Differential laminar distributionof a1, b2, and g2 subunit mRNAs
Previous studies in the rat have found that the expres-sion of a1, b2, and g2 subunits begins postnatally and thatexpression levels increase markedly with age (Gambarana
Fig. 9. Maps made by superimposing camera lucida drawings ofCO-stained barrel hollows (dark filled outlines) and densities of in situhybridization for a1, b1, b2, and g2 mRNAs in an adjacent section(wave filled outlines) at P12, for a1 mRNA in an adult, and immuno-
staining for b2/b3 subunit polypeptides in an adult. Areas shownwithout hybridization in P12 and the adult reflect the fact that theplane of section of the autoradiogram misses layer IV at these points.Scale bar 5 1 mm.
214 P. GOLSHANI ET AL.

et al., 1991; Zhang et al., 1991; Laurie et al., 1992). In thisstudy, we also found a significant developmental increasein hybridization levels of a1, b2, and g2, with an almostdoubling of signal intensity in all layers of the barrel cortexbetween P1 and P12. The b2 and g2 hybridization levelswere consistently lower than a1 hybridization levels, butthe laminar distribution was the same.At P1, a1, b2, and g2 mRNAs are expressed at signifi-
cant levels in the undifferentiated DCP and at moderatelevels in the newly differentiated layers V and VI. Expres-sion of a1, b2, and g2 mRNA in the marginal zone at lateembryonic ages and in layers V and VI by P1 suggests thepresence of GABAA receptors in the synaptic neuropil ofthese layers. Expression in the DCP is unlikely to betokenthe presence of GABA synapses and raises the question asto whether the GABAA receptors formed from these sub-units in the undifferentiated cortical plate are associatedwith trophic actions of GABA during neuronal differentia-tion.There is enrichment of a1 immunoreactivity in layers III
and IV of the somatosensory cortex in the first postnatalweek, followed by a relative increase in staining of deeperlayers of cortex in the second postnatal week (Fritschy etal., 1994). The changes in laminar distribution of the a1subunit mRNA shown in the present study reflect thisobservation. Although there are no immunocytochemical
studies of the developmental expression of b2 and g2polypeptides, the distribution patterns of these polypep-tides in the adult (Fritschy and Mohler, 1995) are inagreement with the in situ hybridization results in adults.Therefore, the levels of the translated polypeptides arelikely to follow the levels shown for the mRNAs throught-out postnatal development.
Differences in a2, a4, a5,and b1 subunit expression
Hybridization levels for a2, a4, a5, and b1 subunits weremuch lower than hybridization levels for a1, b2, and g2.Unlike a1, b2, and g2 subunit-specific riboprobes thatgenerally labeled layers of highest cell density and showedgreat overlap in their hybridization patterns, a2, a4, a5,and b1 subunit-specific probes often preferentially labeledlayers of lower cell density and did not show much overlapin the layers they labeled.Alpha 2 mRNA expression is highest at late embryonic
ages, when it is expressed in ventricular and intermediatezones (Laurie et al., 1992). Alpha 2 is expressed at veryweak levels and changes very little throughout postnataldevelopment. However, hybridization peaks switch fromsuperficial layer V at the end of the first postnatal week tolayer II in the second postnatal week. These results
Fig. 10. Immunocytochemical staining of barrel centers for b2/b3 subunit immunoreactivity at P5 (A)and P12 (B). C shows CO staining of barrel centers, and D shows high levels of expression of a1 subunitmRNA in barrel walls at P12 in sections adjacent to B. Scale bar 5 250 µm.
DEVELOPMENT OF GABAA RECEPTOR SUBUNITS 215

correspond with the developmental expression of a2 sub-unit immunoreactivity (Fritschy et al., 1994).Alpha 4 mRNA transcripts have been localized to the
lower intermediate and ventricular zones in late embry-onic rats and found in the outer cortical layers in postnatalrats (Laurie et al., 1992). Alpha 4 subunit hybridizationlevels increase during the first postnatal week but de-crease into the second week. In the present study, laminardistribution patterns of a4 mRNA also change consider-ably; a4 subunit mRNA was equally enriched in layersII–IV at early postnatal ages but became localized to layerIV by the second postnatal week.Levels of a5 subunit-specific hybridization were highest
in layers with the lowest cell-packing densities, indicatinghigh expression in individual cells; among these layerswas the cortical subplate (layer VIb). Alpha 5 subunitmRNA levels showed a dramatic decrease between earlyand late postnatal ages. Association with the subplatesuggests possible involvement of GABAA receptor contain-ing a5 subunits in trophic functions of the subplate. Alpha5 levels have been shown to decrease during postnataldevelopment of the rat cerebral cortex (Laurie et al., 1992).The present study shows that this result is associated witha shift in laminar distribution of the a5 subunit from peaklevels of expression in layer V and the subplate duringearly postnatal ages to a solitary peak of hybridizationover the superficial white matter at later postnatal ages.Other workers have described a5 subunit mRNA in layerVI (Wisden et al., 1992) or in ‘‘a group of medium to largeneurons located at the border of layer VI and the corpuscallosum’’ in the rat cerebral cortex (MacLennan et al.,1991). Decreases in a5 mRNA between P4 and P12 coin-cides with marked changes in the reversal potential ofGABAA receptor-mediated inhibitory postsynaptic poten-tials (IPSPs) in developing barrel cortex (Agmon et al.,1996) and may play a role in developmental changes in thepermeability of GABAA receptors to anions. The distribu-tion of a5 mRNA found in this study closely follows thedistribution of a5 polypeptide (Fritschy andMohler, 1995).The b1 subunit-specific hybridization increased steadily
throughout postnatal development in all layers, but hybrid-ization levels were highest in layer IV. The b1 subunitmRNAhas been described in the intermediate and ventricu-lar zones at late embryonic ages and in the superficial anddeep cortical layers at early postnatal and late postnatalages in rats (Laurie et al., 1992), with steady increasespostnatally (Zhang et al., 1991). The present resultsidentify layer IV as the layer most enriched by b1 through-out development and in the adult.
Distribution of GAD67 mRNA
GAD67 gene expression was highest in themarginal zoneat P1 and rose dramatically in the other layers during thefirst postnatal week, especially after layers II–IV haddifferentiated from the DCP. There was no evidence of adrop in GAD gene expression at any time during the periodstudied. Micheva and Beaulieu (1995a) reported a signifi-cant reduction in GABA-immunoreactive cells in the ratbarrel cortex between P5 and P10. Any comparable reduc-tion in the mouse is not accompanied by a commensuratedrop in mRNA levels.GADmRNAlevels were relatively low in layers V and VI
and the subplate and lowest of all in the DCP in the firstpostnatal days. GABA and GAD (including GAD67) immu-noreactive cells have been reported in all these layers in
the prenatal and early postnatal rat and mouse and evenin cells migrating through the intermediate zone (Wolff etal., 1984; Van Eden et al., 1989; Cobas et al., 1991; Del Rioet al., 1992; DeDiego et al., 1994; Vincent et al., 1995). Thepresent results suggest that this finding is based on arelatively low level of gene transcription, although lowlevels of GAD67 mRNA might be compensated for byenhanced transcription of GAD65 (see Behar et al., 1994;Esclapez et al., 1994). GABA levels in the cerebral cortex ofthe newborn rat are only about half those in the adult(Coyle, 1982), a fact that is in keeping with an early lowlevel of GAD gene expression, as found in the presentstudy and consonant with the relative weakness of GABA-based inhibition in the first postnatal week (Agmon et al.,1996).
Tangential distribution of GABAA receptors
Previous autoradiographic studies have demonstratedthat GABAA receptor binding is concentrated in the cen-ters of the barrels of rat somatosensory cortex(Chmielowska et al., 1987; Skangiel-Kramska et al., 1994)and immunocytochemical staining with a1-(Paysan et al.,1994) or b2/b3-specific antibodies (Land et al., 1995;present study) presents a similar picture. The hollows ofthe barrels, the site to which most cells in the barrel wallssend their dendrites (Woolsey et al., 1975), contain the vastmajority of excitatory, thalamocortical synapses (Bensha-lom andWhite, 1986) and also contain significant numbersof GABAcells and inhibitory synapses made by GABAcellsin the walls, hollows, and possibly other layers (Keller andWhite, 1987; Solberg et al., 1988; Bernardo et al., 1990;Micheva and Beaulieu, 1995a).High expression levels for a1, b2, and g2 subunits in the
barrel walls provide further evidence that these are thesubunits most highly expressed in the mammalian cere-bral cortex and suggest that most native GABAA receptorsin the adult cortex are composed of a combination of thesethree subunits. Expression of b1 in the barrel walls, albeitat lower levels, suggests that the b1 subunit may also beincluded in some native GABAA receptors in layer IV of therodent barrel cortex. This possibility could affect thepharmacological profile of the receptors. Subunits a2, a4,and a5 were expressed at low levels in the barrel layer.
Functional consequencesof differential subunit expression
GABAA-mediated inhibitory polysynaptic responses canbe elicited in layers V and VI as early as P0 and in layer IVas early as P2, indicating that intracortical inhibition isfunctional in the earliest postnatal days and is presentduring the period at which thalamic innervation of thebarrel cortex occurs (Agmon et al., 1996). GABAA-medi-ated polysynaptic inhibitory responses to thalamic stimu-lation differ in latency and reversal potential from thosefound in more mature animals. By P8, the inhibitoryresponses have become disynaptic instead of polysynapticand have a more negative reversal potential than thosefound in early postnatal animals (Agmon and O’Dowd,1992). Although changes in the latency of inhibitory re-sponses probably result from increases in the excitatorydrive to inhibitory neurons and increases in GABAergicsynapses between P6 and P8, changes in the reversalpotential of inhibitory responses may result from changesintrinsic to the GABAA receptor. GABAA receptors of early
216 P. GOLSHANI ET AL.

postnatal animals may be more permeable to bicarbonateions, and this increase in permeability may result fromdevelopmentaly regulated changes in subunit compositionof the GABAA receptor (Kaila et al., 1993; Staley et al.,1995). The present study found major changes in theintensity and hybridization patterns for a5 and b1 subunitmRNAs between P4 and P12, the period of time whenchanges in GABAergic reversal potentials have been ob-served. During this period, there is a notable decrease ina5 subunit mRNA levels accompanied by a dramaticincrease of b1 subunit mRNA levels in all cortical layers.These changesmay reflect changes in the subunit composi-tion of GABAA receptors, leading to the observed changesin GABAA-mediated responses in developing barrel cortexneurons.
GABAA receptors in cortical development
We observed high expression levels for GAD and mostGABAA receptor subunits in the first postnatal days, thetime during which inhibitory activity is weak and beforethe peak of synaptogenesis in the rodent somatosensorycortex, which occurs from P4 to P32 (De Felipe et al., 1997),suggesting that GABA may play roles other than neuro-transmission during this period. There is considerableevidence to suggest that GABA may be neurotrophic;addition of GABA to immature cultures of the centralnervous system can induce neurite outgrowth, receptorsynthesis, and synaptogenesis (Wolff, 1981; Hanson et al.,1987; Meier et al., 1987; Wolff et al., 1987; Kater andGuthrie, 1990). Treatment of developing animals withGABA agonists has been shown to accelerate and enhancethe expression of developmental markers during the earlypostnatal period (Meier et al., 1987). There is evidence tosuggest that GABAergic neurons can synthesize GABAduring neurite outgrowth and that GABA is present in andcan be released from growth cones of these neurons in anonvesicular, Ca21-independent fashion (Taylor et al.,1990), so GABAA receptors may also be important inprenatal development. Although GABAA receptor subunit-specific knock-out mice have not clearly elucidated a rolefor GABAA receptors in prenatal neural development, theyhave clearly demonstrated an unexpected and crucial rolefor GABAA receptors in craniofacial development, provid-ing the most solid evidence that GABAhas functions otherthan neural transmission in the immature animal (Culiatet al., 1994, 1995). GABA receptor activation has beenshown to regulate brain-derived neurotrophic factor(BDNF) expression levels, further indicating that there issignificant interaction between the GABAergic system andneurotrophic systems in the developing animal (Metsis etal., 1993). The neurotrophic properties of GABAmight dependon selective expression of unique GABAA receptor subunitcombinations during development, leading to GABA-induced depolarization followed by calcium entry via volt-age-gated calcium channels and Ca1-mediated trophiceffects. Populations of native GABAA receptors containinga5 subunits, immunoprecipitated from brain extracts, arecharacterized by increased binding affinities that distin-guish them from all other known GABAA receptor popula-tions (Mertens et al., 1993). We observed highest expres-sion levels for the a5 subunit throughout early postnataldevelopment in the early generated and developmentallyregulated cortical subplate/layer VIb. Subplate neuronsplay key roles in the normal innervation of the cerebralcortex by afferent fibers (Ghosh et al., 1990) and the direction
of cortical efferent projections toward their subcortical targets(McConnell et al., 1989, 1994; DeCarlos and O’Leary, 1992).Ultrastructural (Herrmann et al., 1994) and electrophysiologi-cal (Friauf et al., 1990) analyses have shown that growingthalamocortical axons form functional synapses on subplatecells during this early period; these synapses may function assubstrates for the later recognition of target cells and thereorganization of thalamic afferents in the cortex proper.The presence of a5 subunits in recombinant GABAA
receptors significantly increases sensitivity of the recep-tors to GABA and is also required for cooperativity ofchannel binding, suggesting that the presence of a5 sub-units can endow GABAA receptors with unique functionalproperties that may be essential for any neurotrophicactions of GABA (Meier et al., 1987; Cherubini et al., 1991;Laurie et al., 1992). Thus, selective expression of thesedevelopmentally specific subunit combinations may beessential in the development of the cerebral cortex andmay be a key factor in promoting death of large numbers ofsubplate cells when their developmental role is completed.
LITERATURE CITED
Agmon, A., and D.K. O’Dowd (1992) NMDA receptor-mediated currents areprominent in the thalamocortical synaptic response before maturationof inhibition. J. Neurophysiol. 68:345–349.
Agmon, A., L.T. Yang, D.K. O’Dowd, and E.G. Jones (1993) Organizedgrowth of thalamocortical axons from the deep tier of terminations intolayer IV of developing mouse barrel cortex. J. Neurosci. 13:5365–5382.
Agmon, A., G. Hollrigel, and D.K. O’Dowd (1996) Functional GABAergicsynaptic connection in neonatal mouse barrel cortex. J. Neurosci.16:4684–4695.
Beaulieu, C. (1993) Numerical data on neocortical neurons in adult rat,with special reference to the GABApopulation. Brain Res. 609:284–292.
Beaulieu, C., G. Campistron, and C. Crevier (1994) Quantitative aspects ofthe GABA circuitry in the primary visual cortex of the adult rat. J.Comp. Neurol. 339:559–572.
Behar, T.N., A.E., Schaffner, C.A. Colton, R. Somogyi, Z. Olah, C. Lehel, andJ.L. Barker (1994) GABA-induced chemokinesis and NGF-inducedchemotaxis of embryonic spinal cord neurons. J. Neurosci. 14:29–38.
Benshalom, G., and E.L. White (1986) Quantification of thalamocorticalsynapses with spiny stellate neurons in layer IV of mouse somatosen-sory cortex. J. Comp. Neurol. 253:303–314.
Bernardo, K.L., J.S. McCasland, T.A. Woolsey, and R.N. Strominger (1990)Local intra- and interlaminar connections in mouse barrel cortex. J.Comp. Neurol. 291:231–255.
Blue, M.E., and J.G. Parnavelas (1983a) The formation and maturation ofsynapses in the visual cortex of the rat. I. Qualitative analysis. J.Neurocytol. 12:599–616.
Blue, M.E., and J.G. Parnavelas (1983b) The formation and maturation ofsynapses in the visual cortex of the rat. II. Quantitative analysis. J.Neurocytol. 12:697–712.
Chagnac-Amitai, Y., and B.W. Connors (1989a) Horizontal spread ofsynchronized activity in neocortex and its control by GABA-mediatedinhibition. J. Neurophysiol. 61:747–758.
Chagnac-Amitai, Y., and B.W. Connors (1989b) Synchronized excitation andinhibition driven by intrinsically bursting neurons in neocortex. J.Neurophysiol. 62:1149–1162.
Cherubini, E., J.-L. Gaiarsa, and Y. Ben-Ari (1991) GABA: An excitatorytransmitter in early postnatal life. Trends Neurosci. 14:515–519.
Chmielowska, J., M.G. Stewart, and R.C. Bourne (1987) Autoradiographiclocalization of g-aminobutyric acid receptors in mouse barrel field.Brain Res. 425:283–289.
Cobas, A., A. Fairen, G. Alvarez-Bolado, and M.P. Sanchez (1991) Prenataldevelopment of the intrinsic neurons of the rat neocortex: a comparativestudy of the distribution of GABA-immunoreactive cells and the GABAAreceptor. Neuroscience 40:357–397.
Connors, B.W., R.C. Malenka, and L.R. Silva (1988) Two inhibitorypostsynaptic potentials, and GABAA and GABAB receptor-mediatedresponses in neocortex of rat and cat. J. Physiol. Lond. 406:443–468.
Coyle, J.T. (1982) Development of neurotransmitters in the neocortex.Neurosci. Res. Prog. Bull. 20:479–491.
DEVELOPMENT OF GABAA RECEPTOR SUBUNITS 217

Culiat, C.T., L.J. Stubbs, C.S. Montgomery, B. Russell, and E.M. Rinchik(1994) Phenotypic consequences of deletion of the g3, a5, or b3 subunitof the typeA g-aminobutyric acid receptor in mice. Proc. Natl. Acad. Sci.USA 91:2815–2818
Culiat, C.T, L.J. Stubbs, R.P. Woychick, L.B. Russell, D.K. Johnson, andE.M. Rinchick (1995) Deficiency of the b3 subunit of the type Ag-aminobutyric acid receptor causes cleft palate in mice. Nature Genet.11:344–346
de Blas, A.L, J. Vitorica, and P. Friedrich (1988) Localization of the GABAA
receptor in rat brain with a monoclonal antibody to the 57,000 Mrpeptide of the GABAA receptor/benzodiazepine receptor/C1-channelcomplex. J. Neurosci. 8:602–614.
DeCarlos, J.A., and D.D.M. O’Leary (1992) Growth and targeting ofsubplate axons and establishment of major cortical pathways. J.Neurosci. 12:1194–1211.
DeDiego, I., A. Smith-Fernandez, and A. Fairen (1994) Cortical cells thatmigrate beyond area boundaries: Characterization of an early neuronalpopulation in the lower intermediate zone of prenatal rats. Eur. J.Neurosci. 6:983–997.
De Felipe, J., P. Marco, A. Fairen, and E.G. Jones (1996) Inhibitorysynaptogenesis in mouse somatosensory cortex. Cereb. Cortex, (inpress).
Del Rio, J.A., E. Soriano, and I. Ferrer (1992) Development of GABA-immunoreactivity in the neocortex of the mouse. J. Comp. Neurol.326:501–526.
Esclapez, M., N.J. Tillakaratne, D.L. Kaufman,A.J. Tobin, and C.R. Houser(1994) Comparative localization of two forms of glutamic acid decarbox-ylase and their mRNAs in rat brain supports the concept of functionaldifferences between the forms. J. Neurosci. 14:1834–1855.
Fox, K. (1994) The cortical component of experience-dependent synapticplasticity in the rat barrel cortex. J. Neurosci. 14:7665–7679.
Friauf, E., S.K. McConnell, and C.J. Shatz (1990) Functional synapticcircuits in the subplate during fetal and early postnatal development ofcat visual cortex. J. Neurosci. 10:2601–2613.
Fritschy, J.M., and H. Mohler (1995) GABAA-receptor heterogeneity in theadult rat brain: Differential regional and cellular distribution of sevenmajor subunits. J. Comp. Neurol. 359:154–194.
Fritschy, J.M., J. Paysan, A. Enna, and H. Mohler (1994) Switch in theexpression of rat GABAA receptor subtypes during postnatal develop-ment: An immunohistochemical study. J. Neurosci. 14:5302–5324
Gambarana, C., C.E. Beattie, Z.R. Rodriguez, and R.E Siegel (1991)Region-specific expression of messenger RNAs encoding GABAA recep-tor subunits in the developing rat brain. Neuroscience 45:423–432.
Ghosh,A., A.Antonini, S.K. McConnell, and C.J, Shatz (1990) Requirementfor subplate neurons in the formation of thalamocortical connections.Nature 347:179–181.
Hanson, G.H., E. Meier, J. Abraham, and A. Schousboe (1987) Trophiceffects of GABA on cerebellar granule cells in culture. In D.A. Redburnand A. Schousboe (eds): Neurology and Neurobiology, vol. 32: Neuro-trophic Activity of GABADuring Development. New York: Alan R. Liss,pp. 109–138.
Hendry, S.H.C., H.D. Schwark, E.G. Jones, and J. Yan (1987) Numbers andproportions of GABA-immunoreactive neurons in different areas ofmonkey cerebral cortex. J. Neurosci. 7:1503–19.
Herb, A., W. Wisden, H. Luddens, G. Puia, S. Vicini, and P.H. Seeburg(1992) The third g subunit of the GABAA receptor family. Proc. Natl.Acad. Sci. USA 89:1433–1437.
Herrmann, K., A. Antonini, and C.J. Shatz (1994) Ultrastructural evidencefor synaptic interactions between thalamocortical axons and subplateneurons. Eur. J. Neurosci. 16:1729–1742.
Huntley, G.W., A.L. de Blas, and E.G. Jones (1990) GABAA receptorimmunoreactivity in adult and developing monkey sensory-motor cor-tex. Exp. Brain Res. 82:519–535.
Huntsman, M.-M., P.J. Isackson, and E.G. Jones (1994) Lamina-specificexpression and activity dependent regulation of seven GABAA receptorsubunit mRNAs in monkey visual cortex. J. Neurosci. 14:2236–2259.
Huntsman, M.-M., T.M. Woods, and E.G. Jones (1995) Laminar patterns ofexpression of GABAA receptor subunit mRNAs in monkey sensorymotor cortex. J. Comp. Neurol. 362:565–582.
Kageyama, G., and R.T. Robertson (1993) Development of geniculocorticalprojections to visual cortex in rat: Evidence of early ingrowth andsynaptogenesis. J. Comp. Neurol. 335:123–148.
Kaila, K., J. Voipio, P. Paalasmaa, M. Pasternack, and R.A. Deisz (1993)The role of bicarbonate in GABAA receptor-mediated IPSPs of ratneocortical neurones. J. Physiol. Lond. 464:273–289.
Kater, S.B., and P.B. Guthrie (1990) Neuronal growth cone as an integratorof complex environmental information. Cold Spring Harbor Symp.Quant. Biol. 55:359–370.
Keller, A., and E.L. White (1987) Synaptic organization of GABAergicneurons in the mouse SmI cortex. J. Comp. Neurol. 262:1–12.
Land, P.W., A.L. de Blas, and N. Reddy (1995) Immunocytochemicallocalization of GABAA receptors in rat somatosensory cortex and effectsof tactile deprivation. Somatosens. Mot. Res. 12:127–141.
Laurie, D.J., P.H. Seeburg, and W. Wisden (1992) The distribution of 13GABAA receptor subunit mRNAs in the rat brain. III. Embryonic andpostnatal development. J. Neurosci. 12:4151–4172.
Luhmann, H.J., and D.A. Prince (1991) Postnatal maturation of theGABAergic system in the rat neocortex. J. Neurophysiol. 6:247–263.
MacDonald, R.L., and R.W. Olsen (1994) GABAA receptor channels. Annu.Rev. Neurosci. 17:569–602.
MacLennan, A.J., N. Brecha, M. Krestchatisky, C. Sternini, N.J.K. Till-akaratne, M.-Y. Chang, K. Anderson, M. Lai, and A.J. Tobin (1991)Independent cellular and ontogenetic expression of mRNAs encodingthree polypeptides of the rat GABAA receptor. Neuroscience 43:369–380.
McConnell, S.K., A. Ghosh, and C.J. Shatz (1989) Subplate neurons pioneerthe first axon pathway from the cerebral cortex. Science 245:978–982.
McConnell, S.K.,A. Ghosh, and C.J. Shatz (1994) Subplate pioneers and theformation of descending connections from cerebral cortex. J. Neurosci.14:892–907.
Meier, E., O.S. Jorgensen, and A. Schousboe (1987) Effect of repeatedtreatment with a g-aminobutryic acid receptor agonist on postnatalneuronal development in rats. J. Neurochem. 49:1462–1470.
Meinecke, D.L., andA. Peters (1987) GABA immunoreactive neurons in ratvisual cortex. J. Comp. Neurol. 261:388–404.
Meinecke, D.L., and P. Rakic (1992) Expression of GABA and GABAA
receptors by neurons of the subplate zone in developing primateoccipital cortex: Evidence for transient local circuits. J. Comp. Neurol.317:91–101.
Mertens, S., D. Benke, and H. Mshler (1993) GABAA receptor populationswith novel subunit combinations and drug binding profiles identified inbrain by a5- and g-subunit-specific immunopurification. J. Biol. Chem.268:5965–5973.
Metsis, M., T. Timmusk, A. Enest, and H. Persson (1993) Differential usageof multiple brain-derived neurotrophic factor promoters in the rat brainfollowing neuronal activation. Proc Natl. Acad. Sci. USA 90:8802–8806.
Micheva, K.D., and C. Beaulieu (1995a) Postnatal development of GABAneurons in the rat somatosensory barrel cortex: A quantitative study.Eur. J. Neurosci. 7:419–430.
Micheva, K.D., and C. Beaulieu (1995b) An anatomical substrate forexperience-dependent plasticity of the rat barrel field cortex. Proc. Natl.Acad. Sci. USA 92:11834–11838.
Miller, M.W. (1986) Maturation of rat visual cortex. III. Postnatal morpho-genesis and synaptogenesis of local circuit neurons. Dev. Brain Res.25:271–285.
Mohler, H., D. Benke, S. Mertens, and M. Fritschy (1992) GABAA-receptorsubtypes differing in alpha-subunit composition display unique pharma-cological properties. Adv. Biochem. Psychopharm. 47:41–52.
Paysan, J., J. Bolz, H. Mohler, and J.M. Fritschy, (1994) The GABAA
receptor a1 subunit: An early marker of cortical parcellation. J. Comp.Neurol. 350:133–149.
Prieto, J.J., B.A. Peterson, and J.A. Winer (1994) Morphology and spatialdistribution of GABAergic neurons in cat primary auditory cortex (AI).J. Comp. Neurol. 344:349–382.
Schlaggar, B.L., K. Fox, and D.D.M. O’Leary, (1993) Postsynaptic control ofplasticity in developing somatosensory cortex. Nature 364:623–625.
Schwartz, M.L., D.-S. Zheng, and P.S. Goldman-Rakic (1988) Periodicity ofGABA-containing cells in primate prefrontal cortex. J. Neurosci. 8:1962–1970.
Shaw, C., and B.A. Scarth (1991) Characterization and differential regula-tion of GABAA and benzodiazepine receptors in rat neocortex. Mol.Brain Res. 11:273–282.
Skangiel-Kramska, J., S. Glazewski, B. Jablonska, E. Siucinska, and M.Kossut (1994) Reduction of GABAA receptor binding of [3H] muscimol inthe barrel field of mice after peripheral denervation: Transient andlong-lasting effects. Exp. Brain Res. 100:39–46.
Solberg, Y., E.L. White, and A. Keller (1988) Types and distribution ofglutamic acid decarboxylase (GAD)-immunoreactive neurons in mousemotor cortex. Brain Res. 459:168–172.
218 P. GOLSHANI ET AL.

Staley, K.J., B.L. Soldo, and W.R. Proctor (1995) Ionic mechanisms ofneuronal excitation by inhibitory GABAA receptors. Science 269:977–981.
Taylor, J., M. Docherty, and P.R. Gordon-Weeks (1990) GABAergic growthcones: Release of endogenous g-aminobutyric acid precedes the expres-sion of synaptic vesicle antigens. J. Neurochem. 54:1689–1699.
Van Eden, C.G., L. Mrzljak, P. Voorn, and H.B. Uylings (1989) Prenataldevelopment of GABA-ergic neurons in the neocortex of the rat. J.Comp. Neurol. 289:213–227.
Vincent, S.L., L. Pabreza, and F.M. Benes (1995) Postnatal maturation ofGABA-immunoreactive neurons of rat medial prefrontal cortex. J.Comp. Neurol. 355:81–92.
Wisden, W., D.J. Laurie, H. Monyer, and P.H. Seeburg (1992) The distribu-tion of 13 GABAA receptor subunit mRNAs in the rat brain. I.Telencephalon, diencephalon, mesencephalon. J. Neurosci. 12:1040–1062.
Wolff, J.R. (1981) Evidence for a dual role of GABA as a synaptictransmitter and a promotor of synaptogenesis. In F.V. DeFeudis andP.M. Mandel (eds): Amino Acid Neurotransmitters. New York: Raven,pp. 459–465.
Wolff, J.R., H. Bottcher, T. Zetzsche, W.H. Oertel, and B.M. Chronwall(1984) Development of GABAergic neurons in rat visual cortex asidentified by glutamate decarboxylase-like immunoreactivity. Neurosci.Lett. 47:207–212.
Wolff, J.R., F. Joo, and P. Kasa (1987) Synaptic, metabolic and morphoge-netic effects of GABA in the superior cervical ganglion of rats. In D.A.Redburn andA. Schousboe (eds): NeurotrophicActivity of GABADuringDevelopment. NewYork: Alan R. Liss, pp. 221–252.
Woolsey, T.A., M.L. Dierker, and D.F. Wann (1975) Mouse SmI cortex:Qualitative and quantitative classification of Golgi-impregnated barrelneurons. Proc. Nat. Acad. Sci. USA 72:2165–2169.
Ymer, S., P.R. Schofield,A. Draguhn, P.Werner, M. Kohler and P.H. Seeburg(1989) GABAA receptor b subunit heterogeneity: Functional expressionof cloned cDNAs. EMBO J. 8:1165–1670.
Zhang, J.-H., M. Sato, and M. Tohyama (1991) Different postnatal develop-ment profiles of neurons containing distinct GABAA receptor b subunitmessenger RNAs (b1, b2 and b3) in the rat forebrain. J. Comp. Neurol.308:586–613.
DEVELOPMENT OF GABAA RECEPTOR SUBUNITS 219
![A - Benzodiazepine-Chloride Receptor-Targeted Therapy for ......nisms through GABAA and GABAB receptors [12]. GABA is classified into two main categories: GABAA and GABAB. GABAA and](https://static.fdocuments.us/doc/165x107/60f82a0e0bab2d34196b5ccd/a-benzodiazepine-chloride-receptor-targeted-therapy-for-nisms-through.jpg)

















