
Developmental effect osf some newly induced Ultrabithorax ...Developmental effect osf some newly...
24
/. Embryol. exp. Morph. Vol. 68, pp. 211-234, 1982 211 Printed in Great Britain © Company of Biologists Limited 1982 Developmental effects of some newly induced Ultrabithorax alleles of Drosophila By STEPHEN KERRIDGE 1 AND GINES MORATA 2 From the Centro de Biologia Molecular, Universidad Autonoma de Madrid SUMMARY Nine X-ray-induced mutations of the bithorax complex (BX-C) have been isolated and characterized. They all show the typical features of the Ultrabithorax mutations. They are homozygous lethal, produce a slight enlargement of the haltere in heterozygous condition and fail to complement the mutations at the bx, bxd and pbx loci. Some of them are associated with chromosomal aberrations in the regions 89E 1-4, where the BX-C lies, while others appear normal cytologically. The effect of six of these mutants in the adult cuticle has been studied, producing mutant marked clones in heterozygous individuals. The clones were generated by X-radiation at two points in development: the blastoderm stage and the second larval period. In all cases mutant clones showed the same phenotype: clones appearing in the dorsal structures transform metathorax and first abdominal segment towards mesothorax. That is the additive effect of bx, bxd and pbx mutations. Clones in the legs, if induced during the larval period, show an effect homologous to that seen in the dorsal structures. However, when produced at blasto- derm they show in addition a transformation of the posterior second (mesothoracic) and third (metathoracic) legs into the posterior first (prothoracic) leg. This transformation, named postprothorax (ppx) has been recently described for the alleles Ubx 130 and Vbx 1 (Morata & Kerridge, 1981) and appears to be general for the Ubx mutations. It is concluded that the realm of action of the Ubx gene is defined by part of the rfleso- thoracic segment (posterior second leg compartment) and the entire metathoracic and first abdominal segments. INTRODUCTION The genes of the bithorax complex (BX-C) of Drosophila are necessary for the morphological diversity of thoracic and abdominal segments. Mutations in the complex may transform one or more segments into another but leave the number of segments unaltered (Lewis, 1963, 1978). Segment diversity is achieved by the activation of a specific combination of BX-C genes in each segmental position (Lewis, 1978). In the current view (Lewis, 1978), the lack of BX-C activity results in mesothoracic development; the normal development of the next segment, metathorax, requires the function of the genes bithorax (bx + ) and postbithorax (pbx + ) for the anterior and posterior compartments respectively. The following segment, first abdominal, requires in addition the function of bithoraxoid 1 Author's address: Institut de Genetique Moleculaire, CNRS, Gif-sur-Ivette, France. 2 Author's address: Centro de Biologia Molecular, Universidad Autonoma de Madrid, Canto Blanco, Madrid 34, Spain.
Transcript of Developmental effect osf some newly induced Ultrabithorax ...Developmental effect osf some newly...

/ . Embryol. exp. Morph. Vol. 68, pp. 211-234, 1982 2 1 1Printed in Great Britain © Company of Biologists Limited 1982
Developmental effects of some newly inducedUltrabithorax alleles of Drosophila
By STEPHEN KERRIDGE1 AND GINES MORATA2
From the Centro de Biologia Molecular, Universidad Autonoma de Madrid
SUMMARYNine X-ray-induced mutations of the bithorax complex (BX-C) have been isolated and
characterized. They all show the typical features of the Ultrabithorax mutations. They arehomozygous lethal, produce a slight enlargement of the haltere in heterozygous condition andfail to complement the mutations at the bx, bxd and pbx loci. Some of them are associatedwith chromosomal aberrations in the regions 89E 1-4, where the BX-C lies, while othersappear normal cytologically.
The effect of six of these mutants in the adult cuticle has been studied, producing mutantmarked clones in heterozygous individuals. The clones were generated by X-radiation at twopoints in development: the blastoderm stage and the second larval period. In all cases mutantclones showed the same phenotype: clones appearing in the dorsal structures transformmetathorax and first abdominal segment towards mesothorax. That is the additive effect ofbx, bxd and pbx mutations. Clones in the legs, if induced during the larval period, show aneffect homologous to that seen in the dorsal structures. However, when produced at blasto-derm they show in addition a transformation of the posterior second (mesothoracic) andthird (metathoracic) legs into the posterior first (prothoracic) leg. This transformation,named postprothorax (ppx) has been recently described for the alleles Ubx130 and Vbx1
(Morata & Kerridge, 1981) and appears to be general for the Ubx mutations.It is concluded that the realm of action of the Ubx gene is defined by part of the rfleso-
thoracic segment (posterior second leg compartment) and the entire metathoracic and firstabdominal segments.
INTRODUCTION
The genes of the bithorax complex (BX-C) of Drosophila are necessary for themorphological diversity of thoracic and abdominal segments. Mutations in thecomplex may transform one or more segments into another but leave the numberof segments unaltered (Lewis, 1963, 1978). Segment diversity is achieved by theactivation of a specific combination of BX-C genes in each segmental position(Lewis, 1978). In the current view (Lewis, 1978), the lack of BX-C activity resultsin mesothoracic development; the normal development of the next segment,metathorax, requires the function of the genes bithorax (bx+) and postbithorax(pbx+) for the anterior and posterior compartments respectively. The followingsegment, first abdominal, requires in addition the function of bithoraxoid
1 Author's address: Institut de Genetique Moleculaire, CNRS, Gif-sur-Ivette, France.2 Author's address: Centro de Biologia Molecular, Universidad Autonoma de Madrid,
Canto Blanco, Madrid 34, Spain.

212 S. KERRIDGE AND G. MORATA
(bxd+), for the second abdominal there is the function of infraabdominal-2(iab-2+) (Kuhn, Woods & Cook, 1981), the third abdominal would need thehypothetical iab-3+ etc....In summary, each 'level' of segment development isachieved by the addition of a specific gene function to the pre-existing ones.The last abdominal segment, the eighth, would need the activity of all the BX-Cgenes.
Although Lewis's model explains most of the genetic and developmental dataabout the BX-C, we have found recently (Morata & Kerridge, 1981) that twolethal mutants of the BX-C, Ubx1Z0 and Ubx1 (which behave as deficiencies)produce an unexpected transformation of both posterior meso- and meta-thoracic leg compartments into prothoracic ones. This indicates the existence ofa BX-C function which is necessary for the normal development of the meso-thoracic segment. We named this function postprothorax (ppx) in accordancewith Lewis's nomenclature and suggested the existence of a BX-C gene re-sponsible for this function. Unlike the other BX-C genes, bithorax, post-bithorax and bithoraxoid, which are required throughout larval development(Lewis, 1963: Morata & Garcia-Bellido, 1976), postprothorax is only neededuntil approximately 7 h of development.
The discovery of ppx suggested the existence of a class of early acting BX-Cgenes which would be necessary for the development of mesothorax and perhapsother segments. In particular, as the postprothorax transformation is exclusivelyrestricted to the posterior leg compartment of mesothoracic (second) and meta-thoracic (third) legs, we reasoned that there should exist an 'anterior' prothoracicgene which in mutant form would produce a homologous transformation in theanterior compartments. This argument was based on the existence of two otherBX-C genes, bx and pbx, which perform homologous roles in anterior andposterior compartments respectively (see Morata, 1981 for review).
In an attempt to uncover all the functions of the BX-C, we have begun newexperiments with mutagens and X-rays. Using X-rays we have isolated andcharacterized nine new Ubx alleles. The phenotype of six of them in clonesindicates that they are all deficient for the bx, pbx, bxd and/?/?* activities. Noneof them shows anterior prothoracic transformation or an effect on any segmentbut the mesothorax, metathorax and first abdominal segment.
MATERIALS AND METHODS
(1) Mutagenesis experiment. New BX-C mutants were induced by X-radiationof a multiply marked chromosome mwh jv st red sbd2 e11 ro ca (see Lindsley &Grell, 1968, description of mutants). The dose varied between 3000 and 4000 R.The mutations were detected by a quick method suggested by Gary Struhl. Wecrossed irradiated males to females carrying a weak allele in the bx locus, bx*ie
(the actual genotype was In{3LR)TM3, ri, pv, sep, bx™e, es/Sb62b). The Flgeneration was screened for bx phenotypes. All the possible BX-C mutants,

Developmental effects o/Ultrabithorax alleles o/Drosophila 213even complete deletions for the complex, will survive over foc34e - although somewill be sterile because of the haplo-insufficiency of the Intersex (Ix) gene, whichis located to the right and very close to BX-C (Lewis, 1978). Mutants detected inthis way were then crossed to Df(3R)bxdloo/TMl flies to test their viability overDf(3R)bxd100 (a deletion of part of BX-C) and to isolate stocks over the balancerTML This method, while quick and effective, has the obvious limitation that itcan only detect those BX-C mutants which do not complement &x34e, such as bxor Ubx pseudoalleles or deficiencies. Point mutations, at the bxd or the pbx locifor example, would not be noticed.
(2) Cytological characterization. Mutant chromosomes were analysed cyto-logically to detect possible chromosomal aberrations. Males of each stock werecrossed to females homozygous for the larval marker red. Fl red larvae wereselected and their salivary gland chromosomes examined using standard pro-cedures (Lefevre, 1976).
(3) Genetic characterization. The newly induced mutants were tested overrecessive mutants of the BX-C. For the locus bithorax (bx) two different alleleswere used; bxz (genotype of tester chromosome: sbd2bx3e), and bxsie (a purehomozygous stock was used). For the loci postbithorax (pbx) and bithoraxoid(bxd), one allele was used in each case (tester chromosomes: pbxxexl and bxd511).All the crosses were performed at 25 °C and the flies were mounted on slides formicroscopical examination.
The expressivity of the homeotic transformations in the different combinationswas evaluated and quantified. For transformations of the bithorax type, thenumber of notum and medial triple row bristles in the metathorax was countedand expressed as a percentage of the number of the same type of bristles in thenormal mesothorax (Morata & Kerridge, 1980). combinations showing pbxphenotype were evaluated by scoring the number of bristles in the posteriordouble row which appear in the posterior haltere. This value is expressed as apercentage of the normal number of double row bristles in the wing. Combina-tions showing bxd phenotype were evaluated as the percentage of flies showingabdominal cuticle bearing leg bristles.
(4) Clonal analysis. The method of generating mutant clones is very similar tothat described for Ubx1™ and Ubx1 (Morata & Kerridge, 1981). The followingcross was made:
<? M(l)os*> Dp(3; 1)PU5; Ubxx/Dp(l;3)A59,f+M(l)o+ x; Df(3R)P115/Dp(3;3)P5
In the fathers, the lethal M(l)osp in the first chromosome is covered by Dp(l;3)-A59 which carries the wild-type M(l)o+ gene. The Dp(3; 1)P115 is a duplication ofthe entire BX-C in the proximal part of the first chromosome, while Df(3R)P115is the corresponding deletion of the same genes. Dp(3;3)P5 is a tandem duplica-tion of BX-C. The mutations y, w and /3 6 a are cell-marker mutants used to

214 S. KERRIDGE AND G. MORATA
Table 1. Cytological analysis of new Ubx alleles
Ubx allele
Ubx*-™Ubx6-26
f/fo.5-2326Ubx6-6
Ubx*-12
Ubx*-30
Ubx6-26
Ubx12-6
Ubx1-22
UbxY3
Cytology
NormalNormalNormalNormalNormalT(2;3)34;89E1,2T(2; 3)75C, 54E plus In(3LR) 75C, 89E1,2In(3R)88B, 89E1, 2In(3R)88F, 89E1, 2T(Y; 3) inferred from genetic tests and lost
distinguish mutant clones from surrounding cuticle. Ubx* represents any one ofthe new Ubx mutants.
The Fl of this cross was irradiated and two types of females were collected andscreened for clones:
(1) ywP«a/M(l)osvDp(3; 1)PU5; Ubxx/Df(3R)Pl 15 and(2) ywf™a/M(l)osvDp(3;l)P115; Ubxx/Dp{3;3)P5
in females (1) a mitotic recombination event in the first chromosome (see Fig. 1in Morata & Kerridge, 1981) produces a clone marked with;; wf36a which at thesame time loses the Minute mutation and the Dp{3;l)P115 thus uncovering themutant combination Ubx^/DftffyPllS (these marked Minute+ clones growfaster than surrounding cells, Morata & Ripoll, 1975). There is always a fractionof ywfs6a clones arising from mitotic recombination between Dp{3;l)P115,which is in the heterochromatin, and/3 6 a which is located in the 15 F band.These clones, which represent about 20% of the total, remain wild type for BX-C.In flies of genotype (2) ywfUa clones also grow excessively, but they are notmutant for BX-C because Dp(3;3)P5 carries two doses of all BX-C genes. Thesewere control clones.
Mitotic recombination was induced by X-irradiation at 4 ± 2 h after egglaying (A.E.L.) using a dose of 500-700 R (blastoderm clones) and at 55 ± 12 AELusing a dose of 1000 R (larval clones).
RESULTS
(1) Isolation of new BX-C mutants. A total of 28223 chromosomes werescreened and 28 mutant phenotypes were detected. Of these, 10 were fertile, bredtrue and were isolated as stocks. One of them, that by genetic tests was inferred tobe a translocation involving the third and the Y chromosomes, had low fertilityand was subsequently lost.
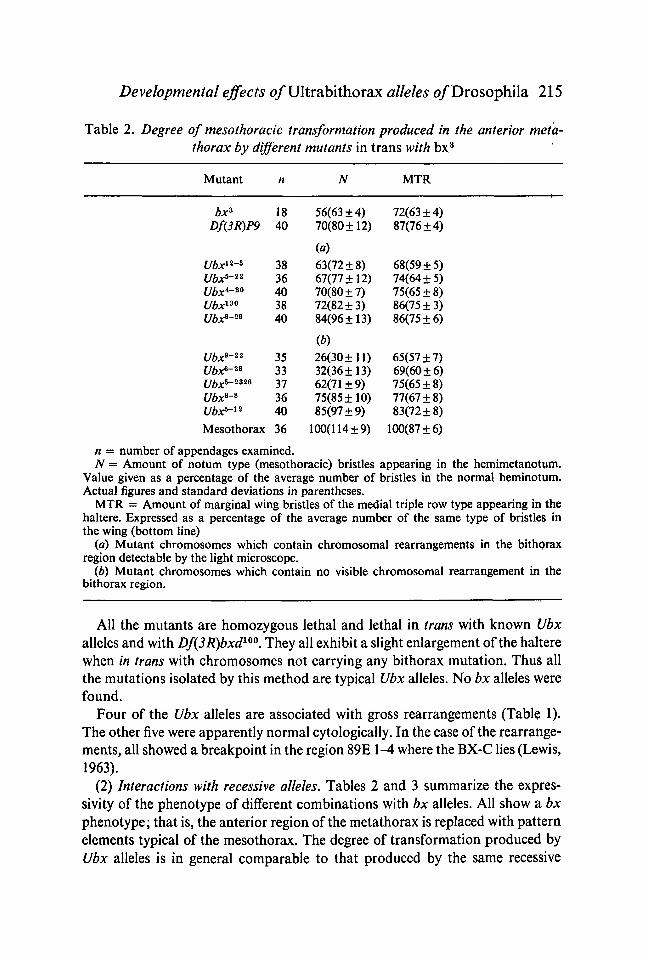
Developmental effects o/Ultrabithorax alleles o/Drosophila 215
Table 2. Degree of mesothoracic transformation produced in the anterior meta-thorax by different mutants in trans with bx3
Mutant MTR
bx*Df(3R)P9
Ubx12~*Ubx*-22
Ubx*-*0
Ubx1*0
Ubx6-26
Ubx9-22
Ubx*-2*Ubx*-2*26
Ubx6-*Ubx*-12
Mesothorax
1840
3836403840
353337364036
56(63 ±4)70(80 ±12)
63(72 ±8)67(77 ±12)70(80 ±7)72(82 ±3)84(96 ±13)
(b)26(30 ±11)32(36 ±13)62(71 ±9)75(85 ±10)85(97 ±9)
100(114 ±9)
72(63 ±4)87(76 ±4)
68(59 + 5)74(64 + 5)75(65 ±8)86(75 + 3)86(75 ±6)
65(57 ±7)69(60 ±6)75(65 ±8)77(67 + 8)83(72 ±8)
100(87 ±6)
n = number of appendages examined.N = Amount of notum type (mesothoracic) bristles appearing in the hemimetanotum.
Value given as a percentage of the average number of bristles in the normal heminotum.Actual figures and standard deviations in parentheses.
MTR = Amount of marginal wing bristles of the medial triple row type appearing in thehaltere. Expressed as a percentage of the average number of the same type of bristle$ inthe wing (bottom line)
(a) Mutant chromosomes which contain chromosomal rearrangements in the bithoraxregion detectable by the light microscope.
(b) Mutant chromosomes which contain no visible chromosomal rearrangement in thebithorax region.
All the mutants are homozygous lethal and lethal in trans with known Ubxalleles and with Df(3R)bxd100. They all exhibit a slight enlargement of the halterewhen in trans with chromosomes not carrying any bithorax mutation. Thu$ allthe mutations isolated by this method are typical Ubx alleles. No bx alleles Werefound.
Four of the Ubx alleles are associated with gross rearrangements (Table 1).The other five were apparently normal cytologically. In the case of the rearrange-ments, all showed a breakpoint in the region 89E 1-4 where the BX-C lies (Lewis,1963).
(2) Interactions with recessive alleles. Tables 2 and 3 summarize the expres-sivity of the phenotype of different combinations with bx alleles. All show a bxphenotype; that is, the anterior region of the metathorax is replaced with patternelements typical of the mesothorax. The degree of transformation produced byUbx alleles is in general comparable to that produced by the same recessive

216 S. KERRIDGE AND G. MORATA
Table 3. Degree of mesothoracic transformation produced in the anterior metathorax by different mutants in trans with bx34e
Mutant
bx3ie
Df(3R)P9
Ubx*-30
Ubx130
Ubx6-26
Ubx9-22
Ubx9-28
Ubx*-2326
Mesothorax
n
3640
4040383638
3835373136
N
26(30 ±5)60(68 ±8)
46(52 + 7)60(68 ±7)47(54 + 8)59(67 ±4)69(79 ±8)
(b)2( 2±1)2( 2 + 2)
24(27 ±7)53(61 ±7)
100(114 ±9)
For explanation see Table 2.
MTR
29(25 ±5)63(55 ±6)
61(53 ±5)57(50 ±7)64(56 ±5)77(67 ±4)68(59 ±6)
53(46 ±4)44(38 ±5)57(50 ±4)60(52 ±6)
100(87 ±6)
mutants over Df(3R)P9 and stronger than that shown by the homozygous reces-sive mutants. Thus most of the new Ubx alleles behave as the complete lack ofbx+ function. Indeed the transformation produced by two of them, Ubx130 andUbx%~2* over bxUe is stronger than that shown by the Df(3R)P9. This is not clearfrom Table 3, where the transformations are evaluated exclusively in terms ofbristle number, but a close examination of haltere appendages of genotypebx3ie/Df(3R)P9 and bxs*e/Ubx130 clearly shows a more complete wing trans-formation of the haltere trichomes and appearance of wing veins (Fig. 1) in thelatter genotype. The difference if probably due to a * transvection' effect (Lewis,1955) of rearrangements with breakpoints in the bithorax complex on the func-tion of the bx function. That is, the hypomorphic function of bxMe may bereduced still further by gross Ubx rearrangements; an effect which may be dueto abnormal somatic pairing of homologous chromosomes in the bithoraxcomplex region.
Two of the point alleles Ubx9~22 and Ubxe~28 showed a transformation weakerthan the recessive alleles. Another point mutant, Ubx1, has also been found tobehave similarly (Morata & Kerridge, 1980). It is of interest to note that in thesethree cases the amount of notum structures is greatly reduced compared to theother alleles, while the amount of wing (MTR) is affected to a lesser degree (Tables2 and 3).
All Ubx mutations, in trans with pbx, transform the posterior metathoracicsegment into a posterior mesothoracic one. Estimations of the expressivity of
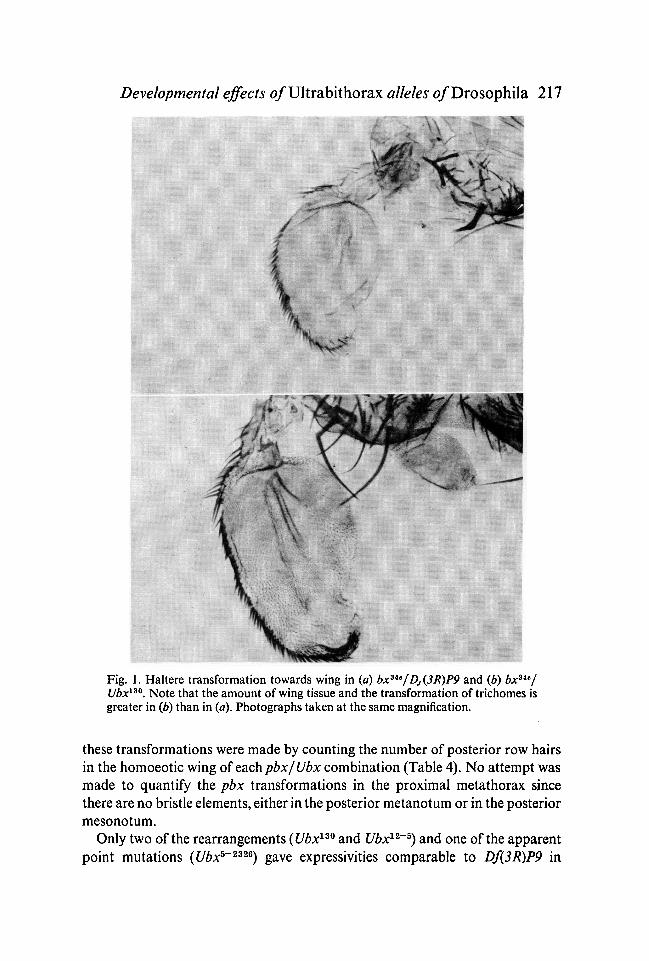
Developmental effects o/Ultrabithorax alleles o/Drosophila 217
I
Fig. 1. Haltere transformation towards wing in (a) bx3le/Dj(3R)P9 and (b) bxsie/Ubx130. Note that the amount of wing tissue and the transformation of trichomes isgreater in (b) than in (a). Photographs taken at the same magnification.
these transformations were made by counting the number of posterior row hairsin the homoeotic wing of each pbx/Ubx combination (Table 4). No attempt wasmade to quantify the pbx transformations in the proximal metathorax sincethere are no bristle elements, either in the posterior metanotum or in the posteriormesonotum.
Only two of the rearrangements (Ubx130 and Ubx12'5) and one of the apparentpoint mutations (Ubx5~2326) gave expressivities comparable to Df(3R)P9 in

218 S. KERRIDGE AND G. MORATA
Table 4. Degree of mesothoracic transformation produced in the posterior meta-thorax by different mutants in trans with pbx
n Transformation (%)
pbxDf(3R)P9
Ubx12-5
Ubx*-22
Ubx*-30
Ubxlz0
Ubx6-™
Ubx9-™Ubx«-2S
C/&C5-2 3 2 6
Ubxs~s
Ubx*-12
Mesothorax
2020
(a)1817181718
(b)172018201823
51(156±14)68(208 ± 7)
63(194 ±13)65(173 ±12)57(176 ±16)62(190 ±14)57(176 ±13)
54(166 ±20)51(156±19)62(191 ±11)52(160 ±23)56(171 ±18)
100(306 ± 5)
n = number of appendages examined.Transformation measured as the amount of double-row (mesothoracic) bristles appearing
in the posterior haltere. Values given as percentages of the average number of double-rowbristles in the normal wing. Actual figures and standard deviations in parenthesis.
(a) and (b) as in Table 2.
combination with pbx. The remaining Ubx/pbx trans combinations are signifi-cantly different from the expressivity of Df(3R)P9/pbx flies. The mutationsUbxs~s, Ubxe~28 and Ubx*~22 are not significantly different from pbx homozygoteswith respect to the numbers of posterior row bristles; indicating that these Ubxalleles have comparable amounts of pbx+ function with the pbx chromosomeitself.
The expressivity of the different Ubx mutations in trans with bxd51i is shown inTable 5. In all cases the main transformation is the removal of the first abdominaltergite and occasionally the replacement with metathoracic structures. In addi-tion, in both the metathoracic and homeotic first abdominal segments, elementsof the posterior metathorax are replaced by posterior mesothoracic ones (Lewis,1963; Kerridge& Sang, 1981).
In no case were Ubx/bxd combinations as extreme as in flies of the genotypeDf{3R)P9/bxdbli, (Table 5). Furthermore, the leg structures seen in the firstabdominal position in Ubx/bxd flies are always rudimentary, with fused legsegments and generally less than 40 leg bristles. On the other hand Df(3R)P9/bxd511 flies can have one or two superficially normal-looking ectopic legs. In nocase were ectopic halteres found.
(3) Clonal analysis. Since all the Ubx alleles used here are lethal when homo-
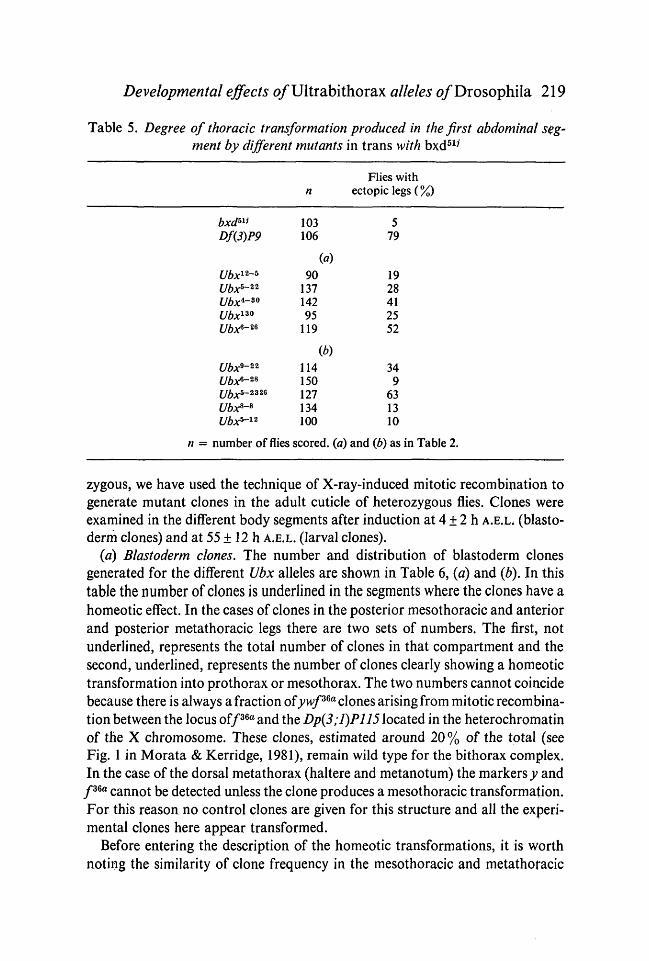
Developmental effects o/Ultrabithorax alleles o/Drosophila 219
Table 5. Degree of thoracic transformation produced in the first abdominal seg-ment by different mutants in trans with bxd517
Flies withn ectopiclegs(%)
bxd™Df(3)P9
Ubx12-*Ubx5-22
Ubx*-*0
Ubx130
Ubx*-26
Ubx9-22
Ubx6-28
Ubx*-2*26
Ubx8-8
103106
(a)90
13714295
119
(b)114150127134
579
1928412552
349
6313
2 100 10
n = number of flies scored, (a) and (b) as in Table 2.
zygous, we have used the technique of X-ray-induced mitotic recombination togenerate mutant clones in the adult cuticle of heterozygous flies. Clones wereexamined in the different body segments after induction at 4 ± 2 h A.E.L, (blasto-derm clones) and at 55 ± 12 h A.E.L. (larval clones).
(a) Blastoderm clones. The number and distribution of blastoderm clonesgenerated for the different Ubx alleles are shown in Table 6, (a) and (b). In thistable the number of clones is underlined in the segments where the clones have ahomeotic effect. In the cases of clones in the posterior mesothoracic and anteriorand posterior metathoracic legs there are two sets of numbers. The first, notunderlined, represents the total number of clones in that compartment and thesecond, underlined, represents the number of clones clearly showing a homeotictransformation into prothorax or mesothorax. The two numbers cannot coincidebecause there is always a fraction ofywf36a clones arising from mitotic recombina-tion between the locus of/36a and the Dp(3;l)P115 located in the heterochromatinof the X chromosome. These clones, estimated around 20 % of the total (seeFig. 1 in Morata & Kerridge, 1981), remain wild type for the bithorax complex.In the case of the dorsal metathorax (haltere and metanotum) the markers y and/3 6 a cannot be detected unless the clone produces a mesothoracic transformation.For this reason no control clones are given for this structure and all the experi-mental clones here appear transformed.
Before entering the description of the homeotic transformations, it is worthnoting the similarity of clone frequency in the mesothoracic and metathoracic

to to O
Tab
le 6
. Num
ber
and
hom
eoti
c ph
enot
ype
ofy
wf3
60
clon
es g
ener
ated
at
blas
tode
rm a
fter
irr
adia
tion
of
the
Fl
of c
ross
es
cJ
M(l
)oSp
Dp(
3;;
Dp
A59
/Ubx
x x ?
y w
f3Sa /F
M6;
D
p{3;
3)P
5/D
f(3)
P11
5c/a
Gen
otyp
e
Ubx
9 -22/D
f(3R
)P11
5U
bx5 -23
™/ D
f(3R
)P11
5U
bxs -8 /D
f(3R
)P11
5U
bx*-
12/D
f(3R
)P11
5U
bx«-
™/D
f(3R
)P11
5U
bx12
-5 /Df(
3R)P
115
*Ubx
130 /D
f(3R
)P9
Ubx
130 /D
p(3;
3)
P5
Con
trol
n 510
474
1098
966
594
330
684
672
Hea
d 6 10 15 10 12 7 14 21
(a) C
lone
s fo
und
in d
orsa
l st
ruct
ures
Mes
otho
rax
14 15 30 19 6 7 17 29
Met
atho
rax
10 12 26 22 11 6 19 —
I 3 — 10 8 4 2 0 22
II 18 — 18 16 12 10 14 19
Abd
omin
al s
egm
ents
III 19 — 27 23 16 14 8 23
IV 25 — 28 25 33 12 18 23
V 23 — 35 30 28 16 19 18
VI 18 — 34 26 25 16 15 17
VII 13 — 19 15 14 7 13 10
7* 3 o a Z O p >

(Tab
le 6
con
t.)
Clo
nes
foun
d in
leg
s
Pro
thor
acic
leg
Mes
otho
raci
c le
g M
etat
hora
cic
leg
Gen
oty
pe
n A
P
A
P
Ubx
9 -22/D
f(3R
)P11
5 51
0 5
2 5
2, 7
Ubx
5 -i32G
/D/(
3R)P
115
474
9 6
7 9,
6U
bx8 -8 /D
f(3R
)P11
5 10
98
14
13
15
8, 0
Ubx
^-i yD
f(3R
)P11
5 96
6 15
10
10
11
, 7
Ubx
G-26
/Df(
3R)P
115
594
10
7 8
5, 2
Ubx
l2-5 /D
f(3R
)P11
5 33
0 6
4 9
5, 4
*Ubx
130 /D
f(3R
)P9
1332
31
22
35
15
, 75
80
600
7 5
8 5,
5
A 7, 13,
19,
19,
16,
10,
35,
10,
19,
6 13 17 16 15 10 27 9 0
P
4, 6, 9, 9, 7, 2, 17, 3, 6,
5 6 8 1 2 14 2 0U
bx13
0 /Dp(
3;
1)P
5 67
2 20
10
18
7,
0
19
,0
6,0
8
Con
trol
g
n =
nu
mbe
r of
sid
es.
<^»
A =
cl
ones
fou
nd i
n th
e an
teri
or
com
par
tmen
t.
S,
P =
cl
ones
fou
nd i
n th
e p
ost
erio
r co
mp
artm
ent.
£
Ital
ic f
igur
es i
ndic
ate
clon
es s
how
ing
ho
meo
tic
tran
sfor
mat
ion.
See
tex
t.
<O*
Par
t of
the
se d
ata
have
bee
n us
ed i
n a
pre
vio
us
publ
icat
ion
(Mor
ata
& K
erri
dge,
198
1).
Df(
3R)P
9 is
a d
elet
ion
of t
he e
ntir
e 72
}bi
thor
ax c
ompl
ex (
Lew
is,
1978
) eq
uiva
lent
to
Df(
3R)P
115.
5
t In
thi
s ca
se t
he f
emal
es u
sed
in t
he c
ross
wer
e yw
P6a
/FM
6;
Dp(
3; 3
)P5/
Ubx
130 .
O bs)
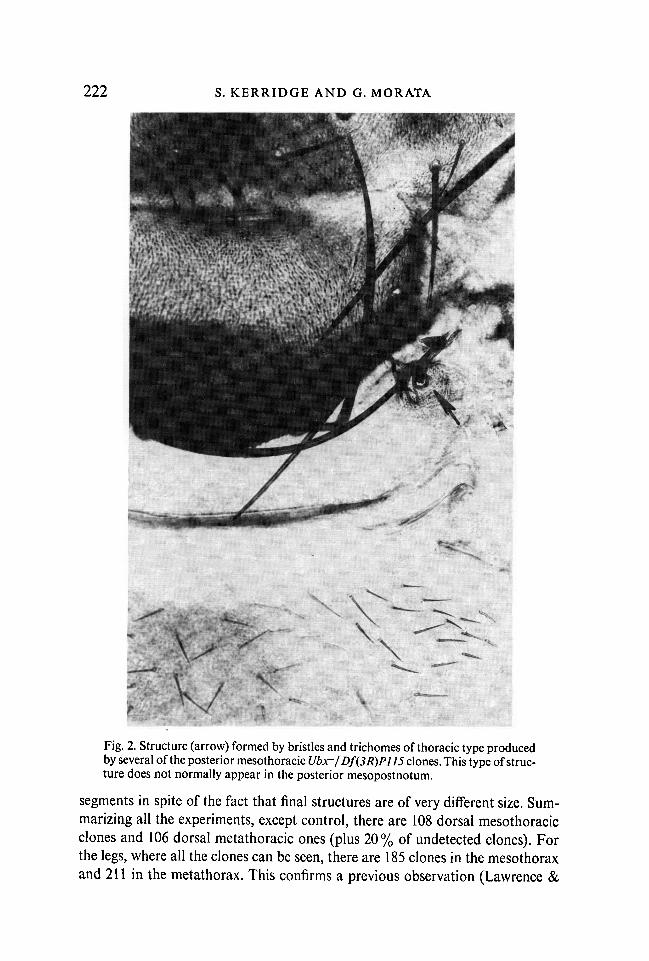
222 S. KERRIDGE AND G. MORATA
Fig. 2. Structure (arrow) formed by bristles and trichomes of thoracic type producedby several of the posterior mesothoracic Ubx~/Df(3R)Pl]5 clones. This type of struc-ture does not normally appear in the posterior mesopostnotum.
segments in spite of the fact that final structures are of very different size. Sum-marizing all the experiments, except control, there are 108 dorsal mesothoracicclones and 106 dorsal metathoracic ones (plus 20% of undetected clones). Forthe legs, where all the clones can be seen, there are 185 clones in the mesothoraxand 211 in the metathorax. This confirms a previous observation (Lawrence &
Developmental effects 0/Ultrabithorax alleles o/Drosophila 223
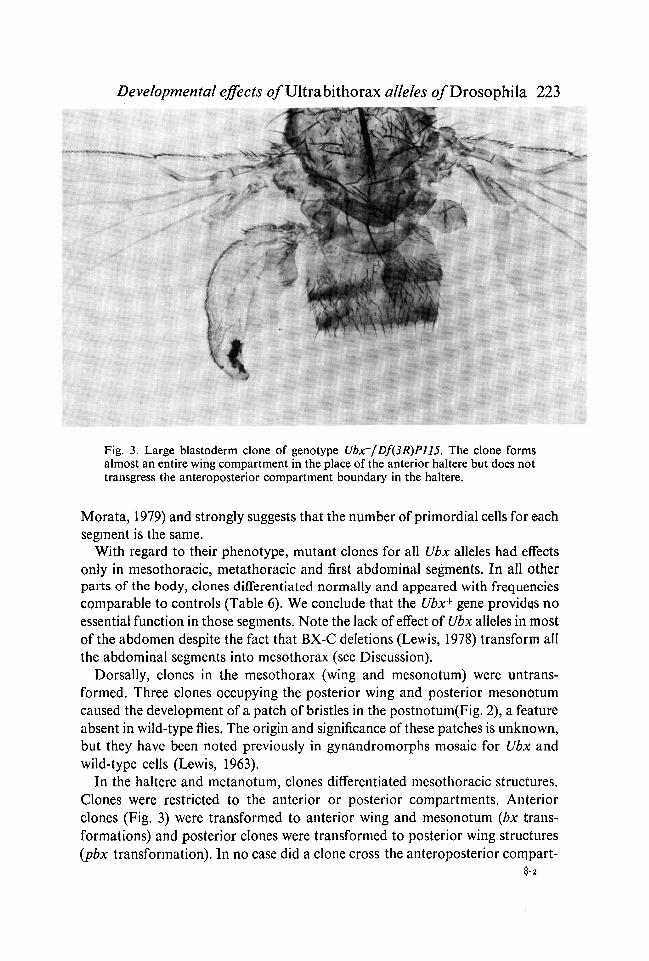
Fig. 3. Large blastoderm clone of genotype Ubx~/Df(3R)P115. The clone formsalmost an entire wing compartment in the place of the anterior haltere but does nottransgress the anteroposterior compartment boundary in the haltere.
Morata, 1979) and strongly suggests that the number of primordial cells for eachsegment is the same.
With regard to their phenotype, mutant clones for all Ubx alleles had effectsonly in mesothoracic, metathoracic and first abdominal segments. In all otherparts of the body, clones differentiated normally and appeared with frequenciescomparable to controls (Table 6). We conclude that the Ubx+ gene provides noessential function in those segments. Note the lack of effect of Ubx alleles in mostof the abdomen despite the fact that BX-C deletions (Lewis, 1978) transform allthe abdominal segments into mesothorax (see Discussion).
Dorsally, clones in the mesothorax (wing and mesonotum) were untrans-formed. Three clones occupying the posterior wing and posterior mesonotumcaused the development of a patch of bristles in the postnotum(Fig. 2), a featureabsent in wild-type flies. The origin and significance of these patches is unknown,but they have been noted previously in gynandromorphs mosaic for Ubx andwild-type cells (Lewis, 1963).
In the haltere and metanotum, clones differentiated mesothoracic structures.Clones were restricted to the anterior or posterior compartments. Anteriorclones (Fig. 3) were transformed to anterior wing and mesonotum (bx trans-formations) and posterior clones were transformed to posterior wing structures(pbx transformation). In no case did a clone cross the anteroposterior cornpart-
$-2
224 S. KERRIDGE AND G. MORATA
bTR
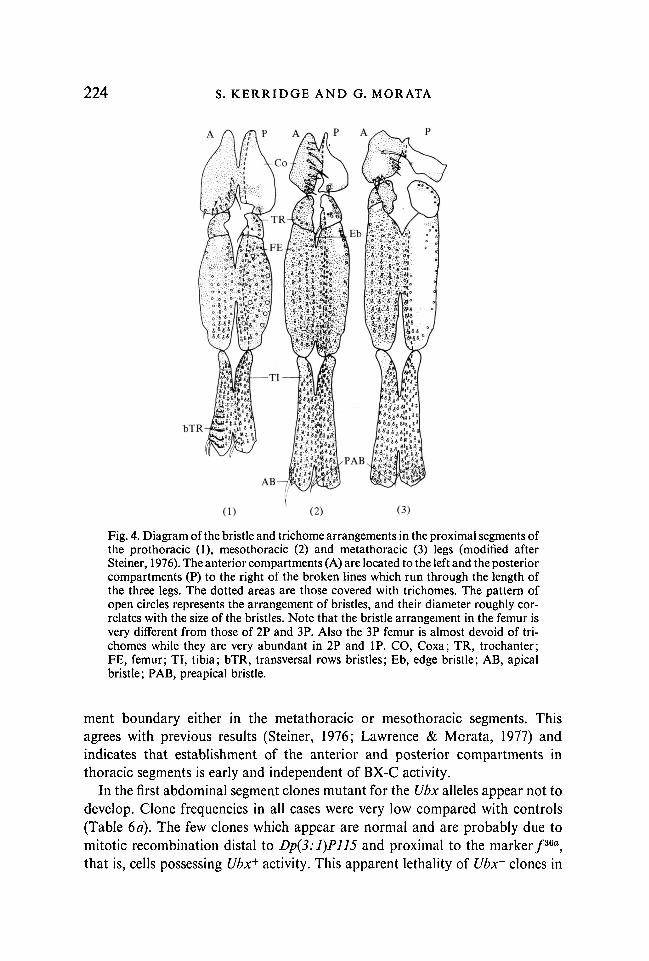
Fig. 4. Diagram of the bristle and trichome arrangements in the proximal segments ofthe prothoracic (1), mesothoracic (2) and metathoracic (3) legs (modified afterSteiner, 1976). The anterior compartments (A) are located to the left and the posteriorcompartments (P) to the right of the broken lines which run through the length ofthe three legs. The dotted areas are those covered with trichomes. The pattern ofopen circles represents the arrangement of bristles, and their diameter roughly cor-relates with the size of the bristles. Note that the bristle arrangement in the femur isvery different from those of 2P and 3P. Also the 3P femur is almost devoid of tri-chomes while they are very abundant in 2P and IP. CO, Coxa; TR, trochanter;FE, femur; TI, tibia; bTR, transversal rows bristles; Eb, edge bristle; AB, apicalbristle; PAB, preapical bristle.
ment boundary either in the metathoracic or mesothoracic segments. Thisagrees with previous results (Steiner, 1976; Lawrence & Morata, 1977) andindicates that establishment of the anterior and posterior compartments inthoracic segments is early and independent of BX-C activity.
In the first abdominal segment clones mutant for the Ubx alleles appear not todevelop. Clone frequencies in all cases were very low compared with controls(Table 6 a). The few clones which appear are normal and are probably due tomitotic recombination distal to Dp(3:l)P115 and proximal to the marker/36",that is, cells possessing Ubx+ activity. This apparent lethality of Ubx~ clones in

Developmental effects o/Ultrabithorax alleles o/Drosophila 225
a b
Fig. 5. Prothoracic transformation shown by clones Ubx-[Dj(3R)P115 (below thedotted line) generated at blastoderm in the posterior compartment of mesothoracic(a) and metathoracic (Z>) legs. Note in (a) that the large marginal bristles, typical of th$posterior prothorax are differentiated by the clone.
the first abdominal segment is probably due to the inability of thoracic cells todevelop in an abdominal segment. A similar observation was made for cloneshomozygous for the viable mutation bxd1, which transform first abdominalsegment into metathorax (Morata & Garcia-Bellido, 1976). We concluded thatthe lack of clones is due to a transformation of first abdominal histoblasts tothoracic cells which fail to differentiate in the abdomen.
In summary, the phenotype of dorsal clones of the genotype Ubx~/Df(3R)P115is that expected for the triple deficiency bx, pbx and bxd, with, as well, indicationof a new phenotype in the posterior mesonotum.
Special mention is necessary of the effect of the clones on the ventral thoracicappendages; that is the legs and pleurae associated with them. Since the bristlepatterns of the three pairs of legs are clearly different (Fig. 4) we can detecthomeotic transformations present in clones. The transverse rows of bristles aretypical of the anterior compartment of the first leg and the sternopleural bristles,edge bristle in the trochanter and apical bristle in the tibia are typical of theanterior compartment of the second leg. For the posterior compartments the

226 S. KERRIDGE AND O. MORATA
I
Fig. 6. Blastoderm clone of genotype Ubx8~s/DJ(3R)PJI5 showing a partial trans-formation into posterior prothoracic leg. The two large bristles (large arrows)located mediodistally are exclusive of the prothorax, but those located more proxi-mally (small arrows) remain largely of mesothoracic type.
differences are very clear in the femur, where the first leg contains more andlarger bristles than the second and third legs (Fig. 4), and the second leg containstrichomes in the ventral side which are missing in the third leg. Also the bristlesin the femur are located more posteriorly in the second than in the third leg.
The leg clones of all the Ubx alleles were scored and their effects noted (Table6b). Clones in the anterior compartment of first and second legs were normal inall the cases and found with normal frequency compared to controls.
As expected most of the Ubx clones in the anterior third leg showed a trans-formation into anterior second, in correspondence with the phenotype of thedorsal clones. This is the phenotype of the bx mutants. The few clones that didnot show a transformation are probably the result of mitotic recombinationproximal to / 3 6 a and distal to Dp{3:l)P115.
In the posterior leg compartments the clones in the second and third legs weretransformed into first (Fig. 5) while those in the first remained normal. Thus, allthe Ubx mutants showed the ppx transformation which we have describedrecently (Morata & Kerridge, 1981).
However, not all the Ubx alleles showed prothoracic transformation with thesame strength, Ubx5~™2\ £/foc9-22and Ubx12'5 in addition to Ubx1™, produceda strong transformation where the posterior prothoracic pattern was reproduced
Tab
le 7
. N
umbe
r of
hom
eoti
c ph
enot
ype
of y
wf36
a clo
nes
gene
rate
d du
ring
the
lar
val p
erio
d in
the
Fl
of t
hecr
osse
s de
scri
bed
in T
able
6
Gen
otyp
en 12
812
015
011
8 96 120
210
Pro
thor
acic
leg
A 12 13 11 11 5 8 14
P 6 5 9 12 3 10 12
Mes
otho
raci
c le
g
A 16 11 13 12 6 12 14
P 4 7 10 2 2 6 9
Met
atho
raci
c le
g
P a o o
Ubx
9 -22/D
f(3R
)PJJ
5U
bx^M
/Dft
fl^P
llU
bxa -s /D
j(3R
)P11
5
Ubx
«-™
/Df(
3R)P
U5
Ubx
12-5 /D
f(3R
)PU
5U
bx13
0 /Df(
3R)P
J15
n =
num
ber
of s
ides
.A
= c
lone
s fo
und
in t
he a
nter
ior
com
part
men
t.P
= c
lone
s fo
und
in t
he p
oste
rior
com
part
men
t.It
alic
fig
ures
indi
cate
clo
nes
show
ing
hom
eotic
tra
nsfo
rmat
ion
into
mes
otho
rax.
15 11 17 8 4 10 10
7 7 7 8 2 9 12
NJ
228 S. KERRIDGE AND G. MORATA
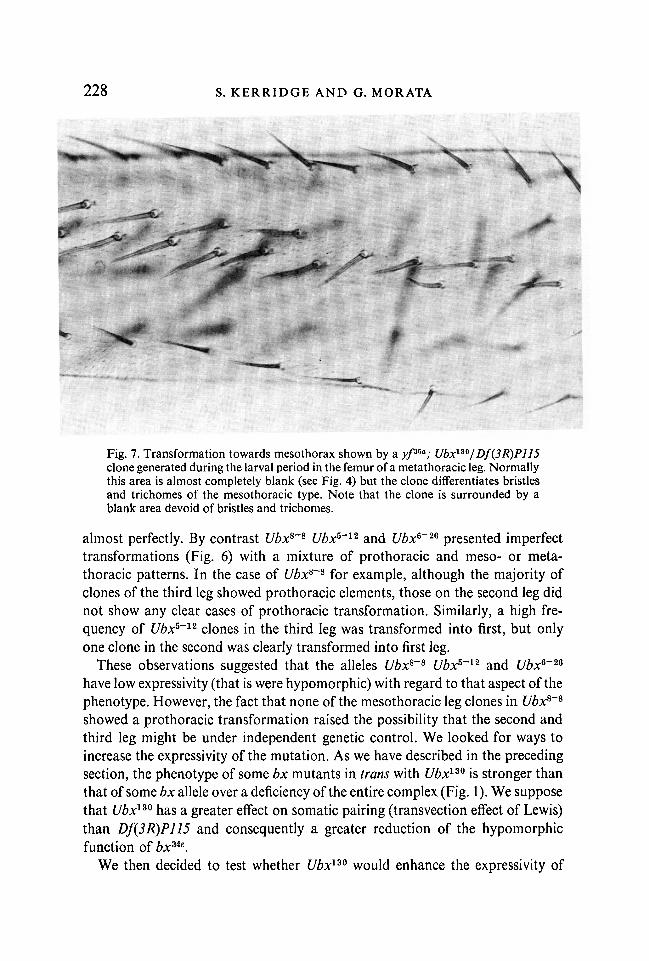
Fig. 7. Transformation towards mesothorax shown by a yp*a; Ubx130/Df(3R)P115clone generated during the larval period in the femur of a metathoracic leg. Normallythis area is almost completely blank (see Fig. 4) but the clone differentiates bristlesand trichomes of the mesothoracic type. Note that the clone is surrounded by ablank area devoid of bristles and trichomes.
almost perfectly. By contrast Ubx8~8 Ubx5~12 and Ubx*~2(i presented imperfecttransformations (Fig. 6) with a mixture of prothoracic and meso- or meta-thoracic patterns. In the case of Ubx8~8 for example, although the majority ofclones of the third leg showed prothoracic elements, those on the second leg didnot show any clear cases of prothoracic transformation. Similarly, a high fre-quency of Ubx5~12 clones in the third leg was transformed into first, but onlyone clone in the second was clearly transformed into first leg.
These observations suggested that the alleles Ubxs~s Ubx5~12 and Ubx*~26
have low expressivity (that is were hypomorphic) with regard to that aspect of thephenotype. However, the fact that none of the mesothoracic leg clones in Ubx8~8
showed a prothoracic transformation raised the possibility that the second andthird leg might be under independent genetic control. We looked for ways toincrease the expressivity of the mutation. As we have described in the precedingsection, the phenotype of some bx mutants in trans with Ubx130 is stronger thanthat of some bx allele over a deficiency of the entire complex (Fig. 1). We supposethat Ubx130 has a greater effect on somatic pairing (transvection effect of Lewis)than Df(3R)P115 and consequently a greater reduction of the hypomorphicfunction of bxUe.
We then decided to test whether Ubx1*0 would enhance the expressivity of

Developmental effects o/Ultrabithorax alleles o/Drosophila 229Ubx8~s and produce prothoracic transformation in the mesothorax. We generatedblastoderm clones of genotype ywf3ea; Ubx8~s / Ubx130 using the same systemdescribed in the Material and Methods, but substituting Ubx1™ for Df(3R)PlJ5.We now find clones in both meso- and metathoracic legs which are transformedinto prothorax (Table 1b). We conclude therefore that Ubx8~8 produces pro-thoracic transformation in both second and third legs, but with differing expres-sivity. We believe that this is also the case for Ubxb~12 and Ubx&~26 although thesealleles have not been tested over Ubx130.
(b) Larval clones. We generated ywf3&a clones mutant for the different Ubxalleles by irradiation during the second larval period. The phenotype of theseclones in the different segments was like that of blastoderm clones but therewere two differences; the first was that there were no clones with patches ofbristles in the postnotum and the second was the absence of the prothoracictransformation of the meso- and metathoracic posterior leg compartments. Theeffect of the clones was restricted to the metathoracic and first abdominal seg-ments. Clones in the haltere showed a transformation into wing, and those inthe metathoracic leg differentiated mesothoracic pattern both in the anteriorand in the posterior compartments. Data from the results obtained in the legsare shown in Table 7. In this table we have considered only those clones whichappear in regions of the legs where the type of bristle pattern whether pro-,meso- or metathoracic could be ascertained. Of the metathoracic clones, we haveconsidered only those which unequivocally presented a homeotic transformation.We took special care in studying the phenotype of the clones in the posteriorcompartments of meso- and metathoracic legs because of the prothoracic trans-formation shown by the blastoderm clones in these compartments. As shown inTable 7, all the 40 mutant clones (for the different Ubx alleles) in the meso-thoracic posterior leg differentiated a typical mesothoracic pattern with no signof prothoracic transformation. In the metathoracic posterior leg 52 clones werefound to produce a homeotic transformation and in every case it was into meso-thorax. One such case is shown in Fig. 7.
As is the case for the blastoderm clones, the larval ones fail to appear in thefirst abdominal segment, while they appear and differentiate normally in the restof the abdominal segments and analia and genitalia.
In conclusion, Ubx~ clones induced during the larval period show bx, pbx andbxd but not ppx transformation. The six new Ubx alleles analysed show thesame temporal sequence of phenotypes described for Ubx130 and Ubx1 (Morata& Kerridge, 1981).
DISCUSSION
Characteristics of the new BX-C mutants
The newly isolated mutants of the BX-C reported here are all Ubx alleles. Allof them in trans with bx, pbx and bxd pseudoalleles give the phenotype of therecessive allele in question, have a dominant phenotype in that the haltere is

230 S. KERRIDGE AND G. MORATA
slightly enlarged and are lethal with all other Ubx alleles and a deficiency for thebithorax complex. In addition, all of them show in clones a ppx phenotype,indicating that the prothoracic transformation is a general feature of the Ubxmutations.
Since a weak bx allele, bx™e, was used to detect some of the new mutants itwas expected that some new bx pseudoalleles would have been isolated. Nonewas found. Current literature also suggests that there are far more Ubx allelesthan bx pseudoalleles (Lewis, 1963; Lindsley & Grell, 1968). However, none ofthe known bx pseudoalleles is associated with gross rearrangements, whereasfrom this analysis approximately half of the Ubx mutants are gross rearrange-ments (Table 2). Of the Ubx mutations isolated over bx3ie, five are putativepoint mutations. The isolation of Ubx mutants only suggests that the DNAresponsible for Ubx alleles is either larger or more sensitive to X-ray mutagenesisthan that giving rise to bx alleles.
Although all the Ubx alleles, old and new, fail to complement bx, pbx and bxdmutants and also show/?/?* phenotype, the data on the expressivity of the pheno-types indicate that some of them are not completely amorphic for the function ofthese genes. This is the case of Ubx9'22 and Ubx*~23 which are both pointmutants. Another point allele, Ubx1 (Morata & Kerridge, 1980), behaves in thesame way. However, other point mutants show an expressivity comparable tothat of the deficiency Df(3R)P9. All the alleles associated with detectable break-points are as strong as the deficiency. This suggests that some of the point mu-tants might have small chromosomal rearrangements undetectable by cytologicalexamination.
The strong effect of chromosomal rearrangements on the bithorax function isprobably related to the phenomenon called 'transvection' (Lewis, 1955), wheresome bithorax mutant phenotypes are enhanced if somatic pairing in the BX-Cregion is disrupted. In fact the combination of bx34e over Ubx120 (Fig. 2) isstronger than over the Df(3R)P9 at least with regard to the transformation of thehaltere trichomes. The same effect is seen for the prothoracic transformation ofUbx8'9, which is stronger over Ubxlso than over Df{3R)P115. Our interpretationis that in Ubx130 the somatic pairing is more disrupted than in the deficiency andconsequently some of the remaining wild-type activity in Ubx8~s is eliminated.It is worth noting that in the case of Ubxs~s it produced bx and pbx transforma-tions as strong as those of Ubx130 or Df(3R)P9, whereas the ppx transformationis weaker. This indicates that the effect of a given Ubx on each of the basic BX-Cfunctions such as bx, pbx, bxd or ppx can be independent.
The interactions of the Ubx mutants with bxd and pbx (Table 4 & 5) show asimpler pattern. In these cases the degree of the transformations is less extremethan with Df(3R)P9, although sometimes the difference is small. Both pointmutants and rearrangements behave similarly, which may suggest that pbx andbxd transformations do not show transvection effects.

Developmental effects o/Ultrabithorax alleles o/Drosophila 231
Early and late functions of the bit ho rax complex
The results presented in this paper confirm and extend a previous result(Morata & Kerridge, 1981) that some of the BX-C genes act on a temporalsequence or that their products act differently at different times of development.Because all the Ubx alleles tested show the early ppx transformation, we con-clude that this is a general feature of the Ubx phenotype. A point which is notyet completely established is the separate existence of ppx as another BX-C geneof the same category but different from bx, pbx and bxd. Indeed to date no inde-pendent mutation has been found for ppx. However, we think its existence isstrongly suggested by the following observations. (1) There are independentisolates of bx, bxd and pbx mutations. They all show a specific transformationand none of them shows, even in their more extreme combinations, any sign ofppx transformation. (2) ppx affects a segment, the mesothorax, which is notaffected by bx, pbx or bxd. (3) The requirement for the normal function of ppx+
is early, until only approximately 7 h of development (Morata & Kerridge, 1981).By contrast bx+, pbx+ and bxd+ are needed until the third larval period (Morata& Garcia-Bellido, 1976). (4) Partial duplications of BX-C like Dp(3;3)bxd100
cover ppx but not pbx or bxd, thus indicating that their wild-type functions are indifferent positions within the complex (Morata & Kerridge, 1981).
It still could be argued that ppx might be the result of the additive effects ofthe triple mutant combination bx, bxd and pbx. Not having analysed this com-bination in clones this possibility cannot be completely ruled out. However, webelieve this is unlikely because (a) extreme double combinations such as bx3
pbx/Ubx1™ (for bx andpbx) or bxdblb/Df{3R)P9 (for bxdaxi&pbx) do not showany sign of ppx transformation; (b) it would be difficult to explain why, if ppxis the summation of bx, pbx and bxd defects, it is shown only by blastodermclones, while bx+, pbx+ and bxd+ are required until much later.
We have proposed (Morata & Kerridge, 1981) that the wild-type function ofpostprothorax (ppx) is necessary for the normal development of posteriormeso- and metathoracic legs. If this function is eliminated, then both compart-ments develop into posterior prothorax. The period of function of ppx is earlyand probably preceeds that of bx or pbx.
In theory this difference in the period of function could be explained by: (a)differential gene activity at BX-C, (b) differential response of other genes due tolack of same/or different BX-C activity. Also, unlike the other genes that affectonly one segment or compartment, ppx+ is required in two non-adjacent com-partments separated by an intervening one (the anterior metathorax) which doesnot require ppx+ activity.
We believe that the phenotype of ppx may suggest a temporal mechanism ofleg determination. The type of leg development, whether prothoracic, meso-thoracic or metathoracic, depends on the switching state of BX-C genes in eachleg primordium. For the posterior compartments, the formula.ppx~,pbx~ results

232 S. KERRIDGE AND G. MORATA
in prothorax, ppx+, pbx~ would be mesothorax and ppx+, pbx+ metathorax.Assuming that anterior and posterior compartments develop in the samemanner, we expected the existence of an 'anterior' gene as we state in theIntroduction. However, we find that neither the Ubx mutants in clones of theadult cuticle nor BC-X deletions in larvae (Lewis, 1978; Struhl, personal com-munication) show the anterior prothoracic transformation. Therefore we areforced to conclude that if the 'anterior' gene exists, it is not in the bithoraxcomplex.
The realm of action of the Ubx gene
All the Ubx mutations tested show a similar phenotype equivalent to theadditive effects of bx, bxd, pbx and ppx transformations. On the idea of theexistence of a separate ppx gene in addition to bx, bxd and pbx, the Ubx mutantsproduce the inactivation of these four genes. The mutant clones have an effecton the adult cuticle of three segments, mesothorax (in part), metathorax andfirst abdominal segment. The clones differentiate normally in the rest of thecephalic or abdominal segments. Since all the Ubx alleles behave in the same waywith respect to the extent of the transformation this result strongly suggests that,for the adult cuticle, the realm of action of the Ubx gene is defined by these threesegments. The fact that Ubx mutations always inactivate bx, bxd, pbx and ppxgenes while there are independent mutations of the latter suggests that these fourgenes have a common overall control for which the normal function of Ubx isindispensable.
As we have not yet studied the larval phenotype of our Ubx mutants, it is notpossible to compare directly the larval and adult phenotypes. However, we canuse to this effect the published larval phenotypes of BX-C deficiencies whichlacking all the genes defective in the Ubx mutations, for example Df(3R)bxd100
(see Lewis, 1978 for description of the different deficiencies and larval pheno-types). What is observed in these larvae is that the metathoracic and firstabdominal segments are transformed towards mesothorax, but there is no reportof a prothoracic transformation homologous to that seen in early clones.Indeed even the biggest deletions where the entire complex is missing, as inDf{3R)P9 or Df(3R)P115, fail to show prothoracic transformations of meso-and metathorax (Lewis, 1978; G. Struhl, personal communication). A likelyexplanation for this apparent discrepancy is that all landmarks used to definethe segment type in the larvae belong to the anterior compartment, which is nottransformed into prothorax. In the larval segments there are not clear differencesbetween posterior pro- and mesothorax, and such a transformation would passunnoticed. In the adult cuticle, on the contrary, the differences between posteriorprothorax and mesothorax are very clear. Thus we think that the larval andadult cuticular structures show the same transformation. According to ourresults, in the larvae homozygous for Df(3R)bxd100, for example, where thegenetic and cytogenetic analysis (Lewis, 1978) indicates that the functions bx,bxd, pbx and ppx are deleted, the mesothorax, metathorax and first abdominal

Developmental effects 0/Ultrabithorax alleles o/Drosophila 233segments would develop as mosaics made of anterior mesothorax and posteriorprothorax.
The realm of action of the bithorax complex
One important difference emerges when we compare the effect of the Ubxalleles and physical deletions such as Df{3K)P9 on the abdominal segmentsposterior to the first. While all the Ubx alleles, even the very strong ones likeUbx130, have their effect limited by the first abdominal segment, Df(3R)P9 showsin addition all abdominal segments transformed towards mesothorax (Lewis,1978, in our view half mesothorax and half prothorax). Df(3R)P9 defines therealm of action of the entire complex; all the segments posterior to mesothoraxrequire BX-C activity. Assuming that the effect of the Ubx mutants on the larvaland adult segments is the same, as discussed above, this indicates that the pheno-type of Ubx mutants accounts for only part of Df(3R)P9 phenotype. This Sug-gests that only some of the BX-C genes are inactivated by the Ubx mutations.Since the inactivation always affects the same genes bx, pbx, bxd and ppx, itsuggests that these form a separate integrated functional subunit of BX-C. Thiswould require a normal activity of the Ubx gene for its normal expression. Therest of BX-C, the 'abdominal gene' region, would be under a different control.It is then possible that the BX-C contains at least two functional subunits, onecontaining the genes controlling the more anterior segment - mesothorax, meta-thorax and first abdominal - and the other containing the genes controlling theremaining abdominal segments.
It is of interest to note that the most anterior limit of BX-C activity seems notto be a segment boundary but the anteroposterior compartment boundary inthe mesothoracic segment. This result, although unexpected, is not unreason-able, as the thoracic anteroposterior boundaries are laid down very early indevelopment possibly at the same time as the segment boundaries (Steiner,1976; Lawrence & Morata, 1977).
REFERENCES
BRYANT, P. J. (1975). Pattern formation in the imaginal wing disc of Drosophila melanogaster:fate map, regeneration and duplication. / . exp. Zool. 193, 49-78.
KERRIDGE, S. & SANG, J. H. (1981). Developmental analysis of the homoeotic mutationbithoraxoid of Drosophila melanogaster. J. Embryol. exp. Morph. 61, 69-86.
KUHN, D. T., WOODS, D. E. & COOK, J. C. (1981). Analysis of a new homeotic mutation(iab-2) within the Bithorax complex in Drosophila melanogaster. Mol. gen. Genet. 181,82-86.
LAWRENCE, P. A. & MORATA, G. (1977). The early development of mesothoracic compart-ments in Drosophila. Devi Biol. 56, 40-51.
LAWRENCE, P. A. & MORATA, G. (1979). Early development of wing and haltere discs inDrosophila. Wilhelm Roux' Archiv. devl Biol. 187, 375-379.
LEFEVRE, G. JR (1976). A photographic representation and interpretation of the polytenechromosomes of Drosophila melanogaster salivary glands. In The Genetics and Biology ofDrosophila, vol. \a (ed. M. Ashburner & E. Novitski), pp. 32-66. London, New York:Academic Press.
LEWIS, E. B. (1955). Some aspects of position pseudoallelism. Am. Nat. 88, 225-239.

234 S. KERRIDGE AND G. MORATA
LEWIS, E. B. (1963). Genes and developmental pathways. Am. Zool. 3, 33-56.LEWIS, E. B. (1978). A gene complex controlling segmentation in Drosophila. Nature 276,
565-570.LINDSLEY, D. L. & GRELL, E. H. (1968). Genetic variations of Drosophila melanogaster.
Carnegie Inst. Wash. Publ. no. 627.MORATA, G. (1981). The mode of action of the bithorax genes of Drosophila melanogaster.
Am. Zool. (in the press).MORATA, G. & GARCIA-BELLIDO, A. (1976). Developmental analysis of some mutants of the
bithorax system of Drosophila. Wilhelm Roux' Arch, devl Biol. 179, 125-143.MORATA, G. & KERRIDGE, S. (1980). An analysis of the expressivity of some bithorax
transformations. In Development and Neurobiology o/Drosophila (ed. O. Sidiqqi, P. Babu,C. Hall, J. Hall), pp. 141-154. London: Plenum Press.
MORATA, G. & KERRIDGE, S. (1981). Sequential functions of the bithorax complex. Nature290,778-781.
MORATA, G. & RIPOLL, P. (1975). Minutes: mutants of Drosophila autonomously affectingcell division rate. Devi Biol. 42, 211-221.
STEINER, E. (1976). Establishment of compartment in the developing leg imaginal discs ofDrosophila. Wilhelm Roux' Archiv devl Biol. 180, 31-46.
{Received 5 October 1981, revised 25 November 1981)









![[OSF] Seed Forum Global EBAN](https://static.fdocuments.us/doc/165x107/587187a21a28ab2c198b5537/osf-seed-forum-global-eban.jpg)








