
Developmental changes in the synthesis of haemolymph polypeptides and their sequestration by the...
8
Insect Biochem, Vol. 12. No. 4, pp 431-440, 1982 0020-1790/82/040431-10503.00/0 Printed in Great Britain © 1982 Per~lamon Press Ltd DEVELOPMENTAL CHANGES IN THE SYNTHESIS OF HAEMOLYMPH POLYPEPTIDES AND THEIR SEQUESTRATION BY THE PREPUPAL FAT BODY IN CALPODES ETHLIUS STOLL (LEPIDOPTERA: HESPERIIDAE) JOHN LOCKE*, HEATHER MCDERMID, TIM BRAC and BURR G. ATKINSON Cell Science Laboratories, Department of Zoology, University of Western Ontario, London, Ontario, Canada N6A 5B7 (Received 24 March 1981, revised 12 November 1981) Abstract--Developmental changes in haemolymph protein and In its synthesis were examined in Cal- podes ethhus from late in the fourth stadium to early pupation The major haemolymph proteins have mol wts of 470,000, 580,000 and 720,000 Upon dissociation, the 470 K and 580 K proteins each resolve into single polypeptldes with mol. wts of 82,000 and 86,000 respectively. The 720 K protein separates into two distinct polypeptides with mol. wts of 86,000 and 90,000. The 82,000 polypeptide is present in the haemolymph at all times but not synthesized either very early in the fifth stadium or after the second critical period The 86,000 and 90,000 polypeptides are present and synthesized only between the first and second critical periods Disappearance of the 86,000 and 90,000 polypeptides from C ethhus haemo- lymph is correlated with a decrease in haemolymph protein concentration and an increase in fat body protein granules Injection of [~4C]-Iabelled haemolymph proteins into larvae after the second critical period results in the fat body acquiring radioactive polypeptides having Mrs of 86,000 and 90,000 These results suggest that the prepupal fat body of C ethhus sequesters at least one and probably two (the 720 K and 580 K proteins) of the three major fifth stadium haemolymph proteins Key Word Index: Haemolymph proteins, Calpodes ethhus, ecdysteroid tltre, fat body. PAGE, fluoro- graphy INTRODUCTION DURING the metamorphosis of holometabolous insects, the last larval instar must prepare itself for the non-feeding pupal instar. Part of this preparation involves the synthesis of considerable amounts of hae- molymph proteins during the feeding phase of the last larval stadium. In C. ethlius an accumulation or in- crease in haemolymph protein is not apparent until after the first critical period, 66 hr after ecdysis (E66) (LOCKE, 1970). This accumulation continues until the second critical period, 156hr after ¢cdysis (E156), after which there is a decline in the haemolymph pro- tein concentration. LOCKE and COLLINS (1968) initially reported these relationships and suggested that the dechne in haemolymph protein involves the seques- tration, accumulation and storage of these proteins by the prepupal fat body. The uptake of haemolymph proteins by the prepu- pal fat body has been demonstrated in Calliphora sty- gia (MARTIN et al., 1971, KINNEAR and THOMSON, 1975) by using autoradlography to identify [i4C]-labelled haemolymph proteins in fat body extracts from larvae injected with [i*C]-labelled hae- molymph proteins. In Lepidopterans this uptake has * Present address and to whom reprint requests should be sent. Department of Biology, Queen's University. King- ston, OntariO, Canada K7L 3N6 only been suggested (KRAMER et al, 1980; TOJO et al., 1980; ToJo et al., 1978) by the dechne in storage pro- teln titre in the haemolymph which accompanies a concurrent increase of proteins m the fat body. The present study identifies the major haemolymph pro- teins of C. ethhus, defines when each is synthesized and demonstrates the selective sequestration of some of these proteins by the prepupal fat body. MATERIALS AND METHODS Animals Larvae of Calpodes ethhus St~ll (Lepidoptera'Hespern- dae) were raised on Camla leaves in our greenhouse until ecdysls to the fifth stadium, at which time they were re- moved to a 22°C incubator with a 12hr hght:12hr dark cycle. In our text and figures animal age is indicated as E followed by the number of hours after ecdysls into the fifth stadium or P followed by the number of hours after pupa- tlon. Under these conditions the first critical period (the time after which the head is no longer necessary for pupa- tion) occurs at E66, the second critical period (the time after which the prothorax is no longer necessary for pupa- tion) at E156 and the fifth stadium lasts approximately 192 hr (LOCKE, 1970) Labelling with [I 4C].leucm e and ~ample preparation Larvae were injected through the base of the third proleg with 5 #Ci/g body weight of L-[t'*C(U)]-leuclne (0 l #Ci/l #l) (Ne~ England Nuclear, specific activity 431
-
Upload
john-locke -
Category
Documents
-
view
212 -
download
0
Transcript of Developmental changes in the synthesis of haemolymph polypeptides and their sequestration by the...

Insect Biochem, Vol. 12. No. 4, pp 431-440, 1982 0020-1790/82/040431-10503.00/0 Printed in Great Britain © 1982 Per~lamon Press Ltd
DEVELOPMENTAL CHANGES IN THE SYNTHESIS OF HAEMOLYMPH POLYPEPTIDES
AND THEIR SEQUESTRATION BY THE PREPUPAL FAT BODY IN CALPODES ETHLIUS STOLL
(LEPIDOPTERA: HESPERIIDAE)
JOHN LOCKE*, HEATHER MCDERMID, TIM BRAC and BURR G. ATKINSON
Cell Science Laboratories, Department of Zoology, University of Western Ontario, London, Ontario, Canada N6A 5B7
(Received 24 March 1981, revised 12 November 1981)
Abstract--Developmental changes in haemolymph protein and In its synthesis were examined in Cal- podes ethhus from late in the fourth stadium to early pupation The major haemolymph proteins have mol wts of 470,000, 580,000 and 720,000 Upon dissociation, the 470 K and 580 K proteins each resolve into single polypeptldes with mol. wts of 82,000 and 86,000 respectively. The 720 K protein separates into two distinct polypeptides with mol. wts of 86,000 and 90,000. The 82,000 polypeptide is present in the haemolymph at all times but not synthesized either very early in the fifth stadium or after the second critical period The 86,000 and 90,000 polypeptides are present and synthesized only between the first and second critical periods Disappearance of the 86,000 and 90,000 polypeptides from C ethhus haemo- lymph is correlated with a decrease in haemolymph protein concentration and an increase in fat body protein granules Injection of [~4C]-Iabelled haemolymph proteins into larvae after the second critical period results in the fat body acquiring radioactive polypeptides having Mrs of 86,000 and 90,000 These results suggest that the prepupal fat body of C ethhus sequesters at least one and probably two (the 720 K and 580 K proteins) of the three major fifth stadium haemolymph proteins
Key Word Index: Haemolymph proteins, Calpodes ethhus, ecdysteroid tltre, fat body. PAGE, fluoro- graphy
I N T R O D U C T I O N
DURING the metamorphosis of holometabolous insects, the last larval instar must prepare itself for the non-feeding pupal instar. Part of this preparation involves the synthesis of considerable amounts of hae- molymph proteins during the feeding phase of the last larval stadium. In C. ethlius an accumulation or in- crease in haemolymph protein is not apparent until after the first critical period, 66 hr after ecdysis (E66) (LOCKE, 1970). This accumulation continues until the second critical period, 156hr after ¢cdysis (E156), after which there is a decline in the haemolymph pro- tein concentration. LOCKE and COLLINS (1968) initially reported these relationships and suggested that the dechne in haemolymph protein involves the seques- tration, accumulation and storage of these proteins by the prepupal fat body.
The uptake of haemolymph proteins by the prepu- pal fat body has been demonstrated in Calliphora sty- gia (MARTIN et al., 1971, KINNEAR and THOMSON, 1975) by using autoradlography to identify [i4C]-labelled haemolymph proteins in fat body extracts from larvae injected with [i*C]-labelled hae- molymph proteins. In Lepidopterans this uptake has
* Present address and to whom reprint requests should be sent. Department of Biology, Queen's University. King- ston, OntariO, Canada K7L 3N6
only been suggested (KRAMER et al, 1980; TOJO et al., 1980; ToJo et al., 1978) by the dechne in storage pro- teln titre in the haemolymph which accompanies a concurrent increase of proteins m the fat body. The present study identifies the major haemolymph pro- teins of C. ethhus, defines when each is synthesized and demonstrates the selective sequestration of some of these proteins by the prepupal fat body.
MATERIALS A N D M E T H O D S
Animals
Larvae of Calpodes ethhus St~ll (Lepidoptera'Hespern- dae) were raised on Camla leaves in our greenhouse until ecdysls to the fifth stadium, at which time they were re- moved to a 22°C incubator with a 12hr hght:12hr dark cycle. In our text and figures animal age is indicated as E followed by the number of hours after ecdysls into the fifth stadium or P followed by the number of hours after pupa- tlon. Under these conditions the first critical period (the time after which the head is no longer necessary for pupa- tion) occurs at E66, the second critical period (the time after which the prothorax is no longer necessary for pupa- tion) at E156 and the fifth stadium lasts approximately 192 hr (LOCKE, 1970)
Labelling with [I 4C].leucm e and ~ample preparation Larvae were injected through the base of the third proleg
with 5 #Ci/g body weight of L-[t'*C(U)]-leuclne (0 l #Ci/l #l) (Ne~ England Nuclear, specific activity
431

432 JOHN LOCKE, et al
342mCl/m-mole). Pupae were injected with eqmvalent amounts of radioactivity through the mtersegmental cuti- cle. After 4 hr at room temperature (22~C), ammals were bled from a pricked proleg (larvae) or dorsal thorax (pupae) Into an ice cold Petn &sh containing 20 #1 of satu- rated phenylthlourea (PTU) The haemolymph/PTU mix- ture (100-200/ll/larva) was transferred to a test tube and centrifuged to pellet haemocytes. The supernatant was mixed with 6~o w/v Tns-HC1 (pH 6.8) or a sodmm dodecyl sulphate (SDS) buffer 180 mM Tris-HCl, pH 6.8, 1 mM phenyl-methyl-sulphonyl-fluonde (PMSF), 5~ow/v SDS and 10Yo w/v glycerol] Samples prepared m the SDS buffer were heated at 100°C for 2mm prior to use Since the addmon of 5 mM 2-mercaptoethanol did not dlssocmte the major haemolymph proteins run on native gels and &d not affect the mobility of haemolymph polypeptldes separated by SDS-PAGE, 2-mercaptoethanol was not routinely added to the solubdlzed proteins
Preparation of 1"14C].labelle d haemolymph proteins Female fifth mstar larvae were collected from a green-
house stock culture prior to the appearance of wax on the posterior ventral surface (the appearance of wax approxi- mately corresponds to the first crmcal period). Approxi- mately 18-36 hr after the appearance of wax (between El00 and El40), larvae were m3ected through the base of the third proleg with 5pCl/g body wt of L-[xaC(U)]-leucme (specific activity 378 mC~/m-mole). A similar rejection was admimstered 12 hr later. Twelve hours after the second injection (during th~s labelhng period all three major hae- molymph proteins are being synthesized) larvae were bled through a pricked proleg into an ice-cold Petn d~sh con- taming 20#1 of water saturated w~th PTU Th~s haemo- lymph-PTU m~xture was centrifuged to removed haemo- cytes An ahquot was kept so that total and acid-precept- table [14C]-radloactlwty could be determined (Table 1) Eight larvae gave approximately 2 ml of haemolymph which was dmlysed for 12 hr at 4°C against art~ficml hae- molymph (RvERSE, 1978) containing PTU An additional 1.0g/l of non-ra&oactlve leuclne (7.6 mM) was added to ddute the [l~C]-leucme remaining m the labelled haemo- lymph. This was followed by a second 12 hr dialysis at 4°C against art~ficml haemolymph w~thout PTU, to remove the PTU m the sample. An ahquot of this sample was kept for assay of the total and ac~d-prec~pltable I-~4C]-ra&oact~vlty (Table 1). The dmlysed haemolymph was then centrifuged (50000 x 10mln at 4°C) and the supernatant used for mject~on into carefully aged larvae
Injectton of 1-t 4C]-Iabelled haemolymph proteins Protein granules begin to appear m the fat body by the
t~me the rectum empties (RE) (M. Locke, personal com- mumcatlon), therefore we chose to reject larvae when gut purging was half complete (~ e E162, a txme at which hae- molymph proteins are no longer being synthesized) The transparent cuticle m C ethhus permitted easy determi- nation of the amount of gut emptying that had occurred as well as the sex of the larva The autonomous nature of
development from RE results m synchronous timing (RvERSE, 1978) At gut-half-empty, female larvae were rejected through the base of the third proleg into the hae- mocoel with 50#1 (600/tg of protein containing 6 25 x 105 DPM) of dmlyzed [~4C]-labelled haemolymph proteins
Tissue preparation and solubdizatton Larvae were bled at 0, 1, 4, 8, 20 and 48 hr after rejection
with [~4C]-labelled haemolymph proteins. Haemolymph was obtained by bleeding through a pricked proleg onto an ice-cold Petrl dish containing 20 #1 of water saturated with PTU This mixture was centrifuged to remove haemocytes and the supernatant solubdlzed m a sodium dodecyl sul- phate (SDS) buffer (5~o w/v SDS, 80 mM Tns-HCl, pH 6.8, 1 mM PMSF and 10~o v/v glycerol) Bled larvae were cut open and rinsed with cold 0 15 M NaC1 The fat body was removed by &ssectlon, thoroughly rinsed m cold 0 15 M NaC1 and homogenized m SDS buffer Both haemolymph and homogemzed fat body samples were placed m a bod- ing water bath for 2 mm and stored frozen at -20°C until use.
Protein and radtoactwlty determination Protein concentration of SDS-solubdlzed haemolymph
and fat body lysates was determined by turbl&metnc assay (COMINGS and TACK, 1972). Ra&oactlwty of [~4C]-labelled proteins was measured in tnchloroacetlc acid-precipitated macromolecules. An aliquot of the haemolymph sample was precipitated in 05ml of 1 mg/ml bovine serum albumin (carrier) using an equal volume of 20~o (w/v) trl- chloroacetlc acid The pellet was washed with 10Yo tn- chloroacet~c acid, sodmm acetate saturated ethanol, etha- nol and ðyl ether. The washed pellet was solublhzed in 0 25 ml of NCS (Amersbam Corp, Arhngton Heights, Ilh- nols), added to a Tnton-Xl00-toluene-ommfluor (New England Nuclear, Boston, Mass) cocktad (TURNER, 1968) and counted m a Beckman LS-255 hqmd sclntdlatlon counter
Polyacrylamzde gel electrophoresls SDS-polyacrylamlde denaturing gels were prepared
according to the method of LAE~IMLI (1970), except that the separating gel consisted of a 3-15~ polyacrylamide gradient slab gel overlaid w~th a 3Vo polyacrylamlde stack- ing gel (ATKINSON, 1981). Non-denaturing polyacrylamide gels were prepared in the same manner as the SDS- denaturing gels, except that SDS was omitted from the gels and buffer solutions
After electrophoresls, proteins and polypeptldes were visualized by staining with Coomassie blue G (WEBER and OSBORN, 1969) Proteins were also stained w~th peno&c aod Schlff (PAS) reagent (CLARKE, 1964) and Sudan black B (CHIPPENDALE and BECK, 1966) Polypeptlde and protem molecular weights (Mr) were determined by comparison wxth concurrently electrophoresed proteins and peptldes of known mol wt (Pharmacm High and Low Molecular
Table 1. Recovery of acid-insoluble [l"C]-radioactlvlty before and after dlalys~s of haemo- lymph proteins from mid-stadium C ethhus larvae
Acid-insoluble Percent [~C] in Haemolymph Total disintegrations &smtegratlons acid insoluble
sample (mm - 1/gl sample) (mm - 1/~1 sample) fraction
Before dmlysls 22,990 18,370 80~o treatment
After dialysis 14,520 13350 95~/o treatment
These represent the counts obtained from one set of determinations.

433
4 4 0 - I i i
2 3 2 - t ,
1 4 0 -
Std ? 0 x
A
Mr
6 6 9 - ~ , ~
232"-
1 4 0 - - ~
6 7 - ~
Std mix SB PAS
B
Fig. 1 Electrophoretlc separatzon of native haemolymph protems from mld-lnstar C ethhus on a 3-15~, polyacrylamide gra&ent gel (A) A comparison of male (3) and female (~) haemolymph proteins co- electrophoresed with known mol wt standards (Std., thyroglobuhn (669,000), ferrltm (440,000), catalase (232,000), lactate dehydrogenase 040,000) and albumin (67,000). (B) Electrophoretlcally separated pooled haemolymph from both sexes are stained with Coomassne blue (Mix), Sudan black B (SB) and periodic
aod Schlff reagent (PAS)
A
669 t !
4 4 0 ~ B i l l ~ q ~ " 4 7 0
232,- 9 i,
140-- ~
Std WH
PAGE
8
t --,94
QgP q l B e -67
- , 43
- ,30
720 580 470 WH Std
SDS - PAGE
Fig. 2 Polyacrylamlde gel electrophoresis of mld-lnstar native haemolymph proteins (A) and their individual polypeptlde constRuents (B) Indlvndual native haemolymph proteins, separated on non- denaturing gels (A), were exosed from the gels, equilibrated in SDS buffer and apphed to an SDS- polyacrylamlde gel The electrophoretlcally-separated polypeptide subumts from each of the native proteins and from whole haemolymph (WH) are shown in B The mol wt standards (Std) used in A are the same as those used in Fig 1, while those used m the SDS-denaturlng gel (B) were phosphorylase I~
(94,000), albumin (67,000), ovalbumm (43,000) and carbomc anhydrase (20,000)
436
Hemolymph
90 )- 82,-
- -94
--,67
4lID - ,43
--"30
201Jg Protein
H FB
LH
Fat body ~ .
0 4
- - ~ --,20
8 20 Std
90=- 86)- 82 "
I
9 0 " 82 )-
. . . . . i i -',94
--,67
g lD -,43
- ,30
8 hour
- . . 1 1 1 - - 2 0
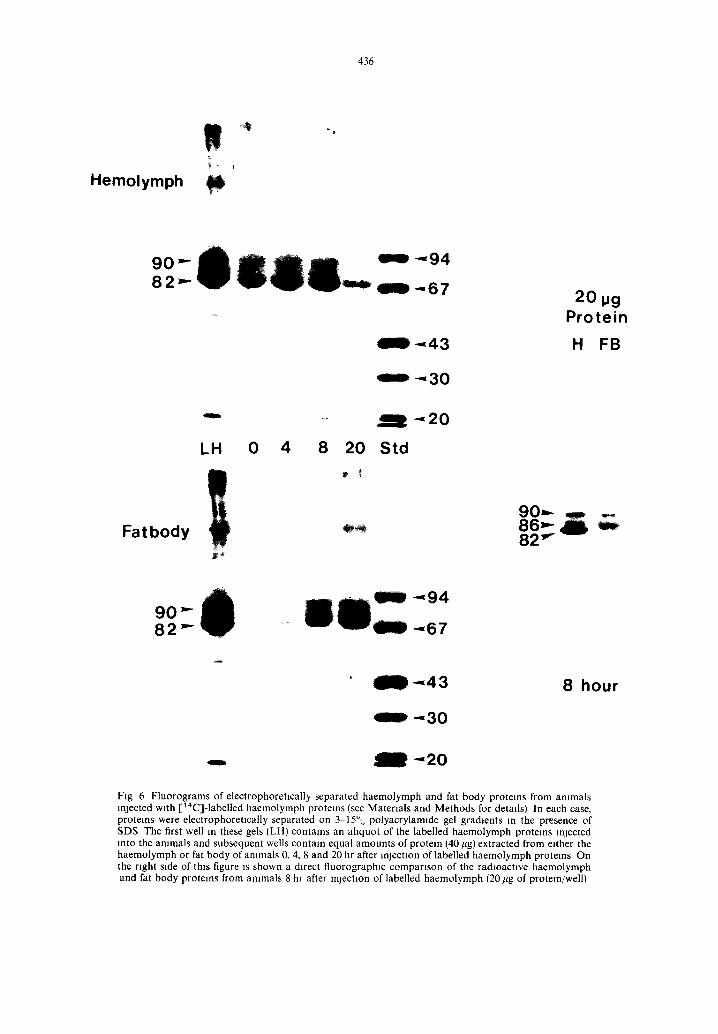
Fig 6 Fluorograms of electrophoretlcally separated haemolymph and fat body proteins from ammals rejected with [14C]-labelled haemolymph proteins (see Materials and Methods for detads) In each case, proteins were electrophoretlcally separated on 3 15°,o polyacrylamlde gel gradients m the presence of SDS The first well in these gels (LH) contains an ahquot of the labelled haemolymph protems rejected into the ammals and subsequent wells contain equal amounts of protein (40/~g) extracted from either the haemolymph or fat body of ammals 0, 4, 8 and 20 hr after m lect]on of labelled haemolymph protems On the right side of this figure is shown a direct fluorographtc compartson of the radioactive haemolymph and fat body proteins from ammals 8 hr after rejection of labelled haemolymph (20/tg of protein/well)

Fate of haemolymph proteins 437
Weight ElectrophoresIs Cahbration Kits; Pharmhcla Fine Chemicals, Div of Pharmacla Inc., Plscataway, N.J.).
Fluoro#raphy Stained slab gels were photographed, impregnated with
2,5-dtphenyloxazole (BONNER and LASKEY, 1974) and dried down onto Whatman 3 MM filter paper with a Bio-Rad 224 Gel Slab Dryer. Fluorographs were prepared by apposing dried gels at -70°C to Kodak RP Royal X-Omat film (XR-1) which was pre-flashed to an O.D. of 0.15 (LASKEY and MILLS, 1975).
Radzoactw~ty measurements by scmtdlat~on counting of poly- pepttde bands exc~sed from gels
Regions from dried gels, corresponding to bands on the fluorograms, were excised, hydrated and incubated with 1 0 ml of 90% NCS solubilizer (Amersham) for 6 hr at 45°C (ATKINSON, 1981, LIEBERMANN et al., 1980) Radioactivity was determined by scintillation counting of the solubdlzed samples m 10 ml of a toluene-based fluor (TuRSER, 1968) on a Beckman LS-255 scintillation counter. Similar-sized cuts from blank areas of the gel were used to determine background and efficiency was determined by the internal standard method of quench correction (H~NDEE, 1972).
RESULTS
Electrophoret~c separation of native haemolymph pro- teins
No major differences are detectable between mid- fifth instar male and female haemolymph proteins separated m 3-159/o polyacrylamide gradient gels (Fig. 1A). In each case there are three major protein bands with apparent mol. wts of 470,000 (470 K), 580,000 (580 K) and 720,000 (720 K). The 470 K pro- tein stains strongly with Sudan black B and PAS, indicating that It IS a lipoglycoprotein (Fig. IB). The other major proteins, 580 K and 720 K, stain weakly with Sudan black B (not visible in photograph) and PAS, suggesting that they too may be lipoglycopro- teins.
Polypeptide constituents of native haemolymph proteins The major protein bands (see Fig. 2A) were excised
from stained native polyacrylam~de gels, equilibrated in SDS buffer (80 mM Tris-HCl, pH 6.8, 2% w/v SDS, 1 mM PMSF, 10% v/v glycerol) for 15 mm and separ- ated into constituent polypeptides on SDS-denaturing 3-15% polyacrylamide gradient gels (Fig. 2B). The 470 K and 580 K proteins each resolved into single polypeptides with tool. wts of 82,000 and 86,000 re- spectively. The 720 K protein separated into two dis- tinct polypeptldes with moi. wts of 86,000 and 90,000. All three proteins, when denatured, also showed very minor constituents of approximately three times the subunit polypeptide mol. wt. These are probably undissociated forms of the polypeptide monomers.
Developmental changes in haemolymph protein and polypeptide synthesis
Haemolymph from animals which were labeled with [14C]-leucme at various ages from late in the fourth stadium to early in the pupal stadium was elec- trophoresed on SDS-denaturing 3-15% polyacryl- am~de gradient gels. The polypeptides separated in these gels were visualized by staining with Coomassie blue G (Fig.. 3A) and newly synthesized polypeptldes
were detected by fluorography (Fig. 3B). The results show that although the polypeptides forming the major haemolymph proteins of mid-fifth instar females have tool. wts ranging from 82,000 to 90,000 (Fig. 2B), only an 82,000 mol. wt polypeptide is detect- able (Fig. 3A) in feeding late fourth mstar larvae (IA). Fluorographic analysis (Fig 3B) indicates that this polypeptide is being synthesized at that time. Six hours after ecdysis to the fifth stadium (E6) an 82,000 mol. wt polypeptide Is present in the haemo- lymph but is no longer being actively synthesized. Synthesis of this polypeptide Is resumed by 45 hr after ecdysis (E45), just prior to the first critical period and continues until approximately the second critical period (El60), after which time synthesis of it stops (Fig. 3B). Beginning at approximately E80, this poly- pepude accumulates and it is maintained at a high level in the haemolymph for at least six days beyond pupation.
Polypeptides with mol. wts of approximately 86,000 and 90,000 are detectable in C ethhus haemolymph only after the first critical period, at E80 (Fig. 3A). While the density of stammg in Fig. 3A masks the separation of the 86,000 and 90,000 polypeptides, other electrophoretically-separated preparations were used to estabhsh the presence of both of these poly- peptides at E80, E145 and El60. Fluorographic analy- sis indicates that these polypeptides are being synthe- sized at E80 and continue to be synthesized up to El60 (Fig. 3B). The presence of these polypeptldes is not electrophoretically detectable in C. ethhus haemo- lymph after El80. The pattern of polypeptide appear- ance and synthesis in males (not shown) is similar to that found m females. The 82,000 mol. wt polypeptide is present in male haemolymph at all times, but is not synthesized just after ecdysis into the fifth stadium or after the second critical period. Polypeptldes with mol. wts of 86-90,000 are present m male haemo- lymph and synthesized only between the first and second critical periods (data not shown).
Specific uptake of rejected [14C]-labelled haemolymph proteins
Twelve of the fourteen female larvae injected at gut- one-half-empty (E162) with 50#1 of [14C]-labelled haemolymph proteins showed no lasting detrimental effects. One larva was permitted to metamorphose completely and moulted into a morphologically nor- mal female adult. The death of the other two larvae was probably due to physical damage during injection since they died soon after injection (possibly due to gut puncture).
These injections did not interfere w~th the appear- ance of protein granules in the fat body, normally seen at this time (LOCKE and COLLINS, 1965, 1968). The cells at 20 h after injection (Fig 4) show no differ- ences from unlnjected larvae. [Compare this micro- graph with those from uninjected larvae of similar age shown m LocKE and COLUNS (1965)]. Protein gran- ules, w~th a denser crystalline core, are present along with the urate granules and glycogen, supporting the proposed storage role for the fat body at this time m development. Most mltochondria and rougl~ enflo- plasmic reticulum have undergone autolysi§-by this time although some are still present, but m reduced amounts

438 JOHN LOCKE, et al
After injection of [14C]-labelled haemolymph pro- teins, the specific radioactivity of fat body proteins increased with time (Fig. 5). Most of this increase occurred within the first eight hours after injection. It then leveled off and possibly showed a slight decline 48hr later; approx 18hr after pupation. Specific radioactivity of haemolymph proteins remained rela- tively constant, but substantially above fat body specific radioactivity, until 20 hr after injection; hae- molymph protein specific radioactivity then declined to relatively constant levels. The temporal increase in fat body protein specific radioactivity along with the concurrent decline in haemolymph protein specific radioactivity suggests that the injected [I4C]-labelled haemolymph proteins are being sequestered from the haemolymph by the fat body.
Sodium dodecyl sulphate (SDS) polyacrylamide gel electrophoresls was used to separate the polypeptides present m haemolymph and fat body homogenates of C. ethhus larvae and pupae that had been injected at gut one-half empty with [14C]-labelled haemolymph proteins. Fluorographlc analysis of these gels con- firmed that the prepupal fat body had sequestered [14C]-labelled proteins from the haemolymph. More- over, fluorographic analysis of constant amounts of electrophoretieally separated haemolymph protein from injected larvae (Fig. 6) shows that the distribu- tion of radioactivity in the haemolymph proteins resemble that of the [~4C]-Iabelled injected proteins. The amount of 14C in electrophoretically separated fat body homogenates, as detected by fluorography, shows an increase in the polypeptldes of the major haemolymph proteins from the time of injection to 20 hr after injection and clearly demonstrates that the fat body sequesters haemolymph proteins.
%
l<
E ¢L 0
20-
15-
10-
5 -
Fatbody
Hours
Fig. 7. An assessment of the amount of radioactivity in individual polypeptides which were electrophoretically separated from proteins extracted from the fat body of animals injected with [14C]-labelled haemolymph proteins Radioactive bands corresponding to the 86,000 (O O) and 90,000 (O----~) molecular weight polypepttdes were cut out of the gels shown in Fig 6 and the amount of radioactivity at each t~me point determined (see Materials
and Methods for details). Counting efficiency was 90%
The sequestration of protein by the prepupal fat body is not non-specific. Fluorographlc analysis of the electrophoretically separated polypeptldes, in fat body that has sequestered [14C]-labelled haemo- lymph proteins, demonstrate that the major [14C]-labelled polypeptldes, present in the haemo- lymph, are not present in the same proportions as in the prepupal fat body (Fig. 6). The 82,000mol. wt polypeptlde is not sequestered while the 86,000 and 90,000 reel. wt polypeptldes are sequestered by the prepupal fat body The density of the 86,000 mol. wt band on the fluorogram appears greater than the 90,000 band Bands, cut from fluorographed gels (Fig. 7) of fat body samples at 4, 8 and 20 hr after injection of [14C]-labelled haemolymph proteins, show considerably more 14C present in the 86,000 reel. wt position than the 90,000 posmon.
DISCUSSION
Polypeptlde constituents of the major haemolymph pro- teins m C. ethllus larvae
In C. ethhus fifth instar larvae there are three major haemolymph proteins with estimated mol. wts of 470,000 (470 K), 580,000 (580 K) and 720.000 (720 K), based on 3-15% linear gradient gel electrophoretlc separations. Reported mol. wts of major larval hae- molymph protein in other Lepldopterans range from 480,000 to 530,000 and appear to consist of one or two proteins rather than three (KRAMER e t al., 1980; ToJo et al., 1980; ToJo et al., 1978) The 470 K protein stains strongly with Sudan black B and periodic acid- Schiff reagent indicating that it contains lipid and carbohydrate This protein could be similar to the lipoproteins In other insects (PATT~AIK et al., 1979; GELLISSEN and EMMERICH, 1980) The 580K and 720 K haemolymph proteins stain weakly with these reagents suggesting that only slight amounts of lipid and carbohydrate may be associated with them. COL- LINS (1974) also found three protein bands which stained with periodic acld-SchIff in C. ethlius haemo- lymph run on 5% polyacrylamide gels.
Upon SDS denaturation, C. ethhus haemolymph proteins dissociate into subunits wlth mol. wts between 82,000 and 90,000. The 470 K protein is a homopolymer with a subunit molecular weight of 82,000, suggesting a hexamenc form. The 580 K pro- tein IS also a homopolymer with a subumt molecular weight of 86,000. The 720 K protein is composed of two different polypeptlde subumts of 86,000 and 90,000 reel. wt in apparent equal amounts and may exist as an octamer composed of equal numbers of each polypeptide.
Developmental changes in the quantity, quality and syn- thesis of haemolymph proteins
Our results demonstrate that there are major changes in the quantity, quality and synthesis of the major haemolymph proteins during the last larval sta- dium of C. ethlius. These changes are similar in females and males and occur at developmental times which correspond to and might reflect events occur° ring at reported critical periods of the last larval sta- dium (LOCKE, 1970). During the last stadium, the con- centratlon of protein in the haemolymph begins to

Fate of haemolymph proteins 439
increase only after the first critical period (E66), reaches a maximum concentration of approximately 120 #g/#l of haemolymph at the second critical period (E156) and thereafter declines. A similar haemolymph protein concentration profile has been reported for larval Lepidoptera (KRAMER et al., 1980; ToJo et al., 1978) and other insects (ROBERTS et al., 1977).
The increase in haemolymph protein concentration in C. ethlius after the first critical period (E66) is due primarily to the synthesis and accumulation of the three major haemolymph protems. Synthesis of the 470 K protein subunit is initiated between 6 and 45 hr after ecdysis to the fifth stadium and continues until El80. By E80, just after the first critical period, sub- untts for the 580 K and 720 K haemolymph proteins are also being synthesized. Synthesis of these proteins continues until approximately El60 Oust after the second critical period) and thereafter stops. The marked synthesis and subsequent accumulation in the haemolymph of subunlts for the 470K, 580K and 720 K proteins occurs between the first and second critical periods which clearly establishes that these proteins must contribute significantly to the overall increase in haemolymph protein occurring at that time.
The decrease m haemolymph protein concentration which occurs after the second critical period appears not to involve the 470 K protein, but rather to result from a depletion of the 580 K and 720 K proteins. By El80 the presence of subunits for the 580K and 720 K proteins are no longer detectable in C. ethlius haemolymph, while the amount of the 470 K protein subunit remains high through pupation. The disap- pearance of some of these major haemolymph pro- teins from C. ethlius haemolymph occurs at the time when the fat body is reported to be sequestering and storing protein in the form of granules (LocKE and COLLINS, 1968). The possibihty that the 580K and 720 K haemolymph proteins are selectively removed from the haemolymph is suggested by these results
Haemolymph protein uptake by the prepupal fat body
C. ethlius prepupal fat body sequesters polypeptldes of at least one and probably two of the three major fifth stadium haemolymph proteins. This sequestra- tion involves the uptake of the 90,000 and 86,000 Mr polypeptides, which are the subunits of the 720 K hae- molymph protein. Fluorographic analysis at the 86,000 Mr band shows that it contains consistently more I4C than the 90,000 Mr polypeptide. If the fat body were sequestering just the 720 K protein the I4C should be equally distributed between these two bands. The 580 K haemolymph protem contains only an 86,000 Mr subunlt, suggesting that the prepupal fat body also sequesters the 580 K protein as well as the 720 K protein. The presence of only the 82,000 Mr subunit in haemolymph and not in the fat body, 20 hr after injection of labelled haemolymph proteins, sug- gests that the 480 K protein is not sequestered by the prepupal fat body. Thus, haemolymph protein seques- tration by the prepupal fat body appears to be a selec- tive process, involving the 720 K and 580 K haemo- lymph proteins.
Since all three of the major proteins are present m the [~4C]-labelled haemolymph protein preparation used for Injection, it is unhkely that the lack of
sequestration of the 82,000 polypeptide is due to a protein dilution effect. It is also unlikely that this selective sequestration is due to the injection pro- cedure since similar patterns of polypeptides are obtained from fat body of uninjected larvae. The selective sequestration of specific haemolymph pro- teins, as reported herein, is probably a normal event in the development of C. ethlius.
Correlation of protein synthetic changes wzth ecdyster- oid t~tre
Before each change In the pattern of synthesis of haemolymph proteins there is an elevation in the hae- molymph ecdysteroid titre (DEAN et al., 1980). Although not yet demonstrated, we assume there is a moulting titre of ecdysterotd between the feeding fourth instar and the newly ecdysed fifth instar which might be responsible for the cessation of synthesis of the 470 K protein as seen in the newly ecdysed fifth. The ecdysteroid peak at approximately El2 (DEAN et al., 1980) occurs prior to or approximately at the time when the synthesis of the 470 K protein is shown to resume. The ecdysteroid peak at approximately the first critical period (E66) is presumed to be the pupal commitment or reprogramming peak and corre- sponds with the initiation of the synthesis of the 580 K and 720 K proteins. Finally, the moulting peak of ecdysteroid at the second crmcal period (E156) cor- responds with the cessation of major haemolymph protein synthesis and the selective disappearance of the 580 K and 720 K proteins from the haemolymph.
The presence of ecdysterolds has been shown to be necessary for aspects of fat body protein metabolism such as the formation of protein granules with the corresponding decline in haemolymph protein con- centrations after the second critical period in C. eth- lius (COLLINS, 1969). Ecdysterone stimulation of pro- tein granule formation has been confirmed m vitro (DEAN, 1978). Although the changes in synthesis of haemolymph proteins correlates with changes in the ecdysteroid titre, the direct control of synthesis by ecdysteroids has yet to be demonstrated, but it seems probable that ecdysteroids play a crucml role m regu- lating the synthesis and sequestrauon of these major haemolymph proteins.
Acknowledgements--We thank Dr M LOCKE and Dr R DEAN for cnt~ozmg the manuscrapt H M and T.B were supported by NSERC postgraduate scholarships J L. held a summer NSERC undergraduate scholarship during this work Support was provided by an Academic Development Fund grant from the Umvers~ty of Western Ontario to Dr B. G. ATKINSON
REFERENCES
ATKINSON B G. (1981) Synthesis of heat-shock proteins by cells undergoing myogenes~s J Cell Bml. 89, 666-673
BONNER W. M. and LASKEY R A (1974) A film detectaon method for trmum-labelled proteins and nucleic aods m polyacrylamlde gels Eur. J Blochem. 46, 83-88.
CHIPPENDALE G M and BECK S D (1966)Hemolymph proteins of Ostrmm nubilahs during dlapause and pre- pupal differentiation. J Insect Phvsiol 12, 1629-1638
CLARK J. T. (1964) SImphfied "disc' (polyacrylamlde gel) electrophoresls Ann. N.Y Acad. Scz 121,428-436.
COLLINS J V. (1969) The hormonal control of fat body development m Calpodes J. Insect Phymol 15, 341-352

440 JOHN LOCKE, et al
COLLINS J. V. (1974) Hormonal control of protein seques- tration m the fat body of Calpodes ethhus (St611). Can. J. Zool. 52, 639-646.
COMINGS D. E. and TACH L. C. (1972) Similarities in the cytoplasmic proteins of different organs and species examined by SDS electrophoresis. Expl Cell Res. 75, 73-78.
DEAN R. L (1978) The induction of autophagy in isolated insect fat body by ~-ecdysone. J Insect Physiol 24, 439-447.
DEAN R. L, BOLLENBACHER W. E, LOCKE M, SMITH S L and GILBERT L I (1980) Haemolymph ecdysterold levels and cellular events in the lntermoult/moult sequence of Calpodes ethhus, d. Insect Phystol. 26, 267-280
GELLISSEN G. and EMMERICH H. (1980) Purification and properties of a dlglycende-bmding hpoprotem (LPI) of the hemolymph of adult male Locusta migratoria. J camp Physml. 136, 1-9.
HENDEE W. R. 0972) Radtoactwe Isotopes m Btological Research. John Wiley, New York
KINNEAR J. F. and THOMSON J. A. (1975) Nature, origin and fate of major hemolymph proteins in Calhphora Insect Btochern. 5, 531-552
KRAMER S. J., MUNDELL E. C. and LAW J H (1980) Purifi- cation and properties of Manducln, an amino acid stor- age protein of the hemolymph of larval and pupal Man- duca sexta Insect Btochem 10, 279-288.
LAEMMLI U K. (1970) Cleavage of structure proteins dur- ing the assembly of the head of bacteriophage T4. Nature 227, 680-685.
LASKEY R. A. and MmLS A. D. (1975) Quantitative film detection of 3H and ~4C m polyacrylamlde gels by fluor- ography. Eur J. Btochem. 56, 335--341
LIEBERMANN D B., HGFFMAN-LIEBERMANN B. and SACHS L (1980) Molecular dissection of differentiation in normal and leukemic myoblasts, separately programmed path- ways of gene expression. Deal Btol 79, 46-63
LOCKE M. (1970) The molt/intermoult cycle in the epi- dermts and other tissues of an insect Calpodes ethlius
(Lepldoptera Hesperndae) Tzssue and Cell 2(2), 197-223
LOCKE M. and COLLINS J. V. (1965) The structure and for- mation of protein granules m the fat body of an insect. J Cell Biol. 26, 857-885.
LOCKE M. and COLLINS J. V. (1968) Protein uptake into mulUvesicular bodies and storage granules m the fat body of an insect. J Cell Biol. 36, 453-483.
MARTIN M D, KINNEAR J F and THOMSON J. A. (1971) Developmental changes m the late larva of Calhphora stygia IV Uptake of plasma protein by the fat body. Aust. I. blol Scl 24, 291-299.
PATTNAIK N M., MUNDELL E C, TRAMBUSTI B G., LAW J H and K~ZDY F J. (1979) Isolation and characteriz- ation of a larval hpoprotem from the hemolymph of Manduca sexta. Camp. Biochem. Physiol. 630, 469-476.
R'~RSE J S (1978) The hormonal control of Malpighian tubule structure and function during larval-pupal-adult development of the skipper butterfly, Calpodes ethhus Ph.D Thesis, University of Western Ontario, London, Ontario
ROnERTS D B, WOLFE J and ADAM M E. (1977) The developmental profiles of two major hemolymph pro- telns from Drosophda melanogaster J. Insect Physiol 23, 871-878.
ToJo S., BETCHAKU T, ZICCARD1 V J. and WYATT G. R (1978) Fat body protein granules and storage proteins m the sllkmoth Hyalophera cecropia. J. Cell Btol. 78, 823-838.
ToJo S, NAGATA M and KOBAYASm M. (1980) Storage proteins m the silkworm, Bombyx mort Insect Biochem 10, 289-303
TURNER J. C. (1968) Triton X-100 scintdlant for carbon-14 labelled materials. Int. J appl. Radiat Isotopes 19, 557-563.
WEBER K. and OSaORN M. (1969) The reliability of mol- ecular weight determinations by dodecyl sulfate-poly- acrylamide gel electrophorests J btol Chern 244, 4406--4412.


















