
Detection of Enterotoxins Genes in … of Iraq Ministry of Higher Education & Scientific Research...
162
Republic of Iraq Ministry of Higher Education & Scientific Research University of Baghdad College of Science Detection of Enterotoxins Genes in Staphylococci Isolated from Milk and Cheese A Thesis Submitted to the College of Science-University of Baghdad in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Ph.D) in Microbiology/ Food Poisoning. By Marwa Hameed Mtashar Al-Khafaji B. Sc. Biology/College of Science/University of Baghdad 2002 M. Sc. Microbiology/College of Science/University of Baghdad 2008 Supervised by Professor Dr. May Talib Flayyih May 2013
Transcript of Detection of Enterotoxins Genes in … of Iraq Ministry of Higher Education & Scientific Research...

Republic of Iraq Ministry of Higher Education & Scientific Research University of Baghdad College of Science
Detection of Enterotoxins Genes in Staphylococci Isolated from Milk and
Cheese
A Thesis Submitted to the College of Science-University of Baghdad in
partial fulfillment of the requirements for the degree of Doctor of Philosophy (Ph.D) in Microbiology/ Food Poisoning.
By Marwa Hameed Mtashar Al-Khafaji
B. Sc. Biology/College of Science/University of Baghdad 2002
M. Sc. Microbiology/College of Science/University of Baghdad 2008
Supervised by
Professor Dr. May Talib Flayyih
May 2013

حيم حمن الر بسم هللا الر
))سبح اسم ربك األعلى((
صدق هللا العلي العظيم
سورةاألعلى
) ۱اآلية (

Certification
I certify that this thesis was prepared under my supervision at
College of Science, University of Baghdad, as a partial requirement
for the degree of Doctor of Philosophy (Ph.D) in
Microbiology/Food poisoning.
Signature:
Dr. May Talib Flayyih
Professor
Date: / /2013
In view of the available recommendation, I forward this thesis for debate
the examining committee.
Signature:
Prof. Dr. Sabah N. Alwachi Head of Department of Biology
College of Science
University of Baghdad
Date: / /2013

Committee’s Certification
We, the examining committee, certify that we have read this dissertation and have examined the student Marwa Hameed Mtasher Al-Khafaji in its contents and that in our opinion it is adequate with good standing as a dissertation for the degree of Doctor of philosophy (PhD) in
Microbiology/Food Poisoning.
Dr. Rashid M. Musleh
Professor Chairman
/ 6 / 2013
Dr. Hayfa H. Hassani Dr. Intisar M. Juma Professor Assistant Professor
Member Member / 6 / 2013 / 6 / 2013
Dr. Ayad M. A. Fadhil Dr. Shrooq R. Kadhum Assistant Professor Assistant Professor
Member Member / 6/ 2013 / 6 / 2013
Dr. May T. Flayyih Professor Advisor
/ 6 / 2013
Approved for the College of committee of graduate studies. Prof. Dr. Saleh Mahdi Ali
Dean College of Science Baghdad University
/ 6 / 2013

I dedicate this little effort to: To the persons who guided me to the right path…
My leaders and teachers… The prophet of God "Mohammed'' and his relatives
"mercy and peace are up on them" To the great home, where I fell safe, although bombs surrounding me from every direction…
My injured country …Iraq To those who loved me more than themselves gave me their lives … To those who always are surprising me with a very beautiful words and works... To those who lives and settles inside my heart…
My lovely family My dear friends
Marwa

ACKNOWLEDGEMENT
First of all, I thank God Allah for all his endless blessings and giving me the power and the intention to accomplish this work in this final shape. Mercy and peace are up on the prophet of God "Mohammed'' and his relatives. This research project would not have been possible without the support of many people. I wish to express my grateful thanks to my deer supervisor Professor Dr. May T. Flayyih for her scientific guidance, recommendation, advice, encouragement, support, patience and kindness during my scientific and practical life. Special thanks go to my consultant supervisor Dr. Majeed A. Sabah Biotechnology Research Center, for his good observations and valuable efforts in molecular working through this work. My gratitude also extends to the head and all members: my colleagues and friends at the Department of Biology, College of Science, University of Baghdad who gave me their hands and support to finish my work. I would like to thank the head and all the staff of biotechnology research center, with special thanks to Mrs. Noor Hashim, Miss. Reghad, Mr. Ali Talib and Mr. Anes Noory for their cooperation and offering a friendly work environment. The words are not enough express my great love, thankful and heartfelt gratitude to my lovely family; my dearest and lovely mother and father, my uncles (Ali and Saad), my kind brothers (Ahmed, Laith, Ali and Yusef) and my sweetest beautiful sisters (Safa, Muna, Shahad, Nada, Aula, and Farah). Finally, I would like to express my deepest and warmest gratitude to all who had contributed to the completion and success of this project whom I forgot to mention I dedicate my wishes, thanks and gratitude.
Marwa

Summary
During 2011; 300 milk and cheese samples were collected from Baghdad markets.
Two hundred staphylococcal isolates were isolated from milk and cheese samples, the
predominant species was Staphylococcus aureus, 97 isolates (48%), followed by
S.chromogenes 82 isolates (41%) and 21 (11%) S.epidermidis isolates.
The pattern of antibiotic susceptibility of Coagulase Positive Staphylococci
(COPS) and Coagulase Negative Staphylococci (CONS) isolates to 3 antibiotics
(Methicillin, Tetracycline and Vancomycin) was determined using disc diffusion
method; the results revealed that 80 S. aureus isolates (82.47%) found to be
methicillin resistant (MRSA) and 18 S. aureus isolates (19%) resist tetracycline while
8 isolates (8.24%) were vancomycin resistant (VRSA). Sixty four CONS isolates
(62.13%) were methicillin resistant, 28 CONS isolates (27.18%) resist tetracycline,
and 5 CONS isolates (4.85%) were vancomycin resistant.
Deoxyribo nucleic acid (DNA) extraction from staphylococcal isolates and directly
from milk and cheese samples was done manually.
The isolates were subjected to Polymerase chain reaction (PCR) technique in a
monoplex pattern to amplify coagulase encoding gene: the coa gene; results by this
study showed that 76 (78.35%) S. aureus isolates gave the amplicon size 730 base pair
of the coa gene.
The genetic determinants of methicillin resistance femA and mecA genes were
amplified using monoplex PCR technique in order to identify methicillin resistant
(mecA+) and susceptible (lacking mecA) staphylococci and to differentiate S. aureus
(femA+) from coagulase negative staphylococci (lacking femA). Ninety six S. aureus
isolates (98.96%) were found to harbour femA gene, it is species specific marker for S.
aureus. The mecA gene was detected in 91 (93.81%) MRSA isolates, while it was
detected in 70 (67%) CONS isolates.
The detection of staphylococcal enterotoxigenicity according to four classical
enterotoxins genes which are sea, seb, sec, and sed was performed simultaneously
using multiplex PCR assay. One hundred fifty three staphylococcal isolates (76.5%)
found to be enterotoxigenic, 95 S.aureus isolate (62.09%) found to harbor one or more

enterotoxin gene. CONS isolates showed to be enterotoxigenic in this research, they
accomplished 37.9% of the total enterotoxigenic strains (58 strains).
The sea gene was the most frequent enterotoxin coding gene among the others
tested; sea accomplished 51.85% of the detected enterotoxin genes followed by the sed
gene, which constituted 34.73%, and then the seb coding gene by 12.5% while the sec
gene was very rare as 0.92%.
The results obtained by this study showed that 102 (66.66%) staphylococcal
isolates harboured only one enterotoxin coding gene, while 39 (25.5%) of the isolates
possessed two toxin genes, which were mostly sea and sed genes and 12 staphylococcal
isolates (7.8%) harboured three toxin coding gene: sea, seb and sed genes.
Suckling mouse bioassay was tested to investigate the staphylococcal enterotoxin
biological activity. Results showed that 131 isolates which constitutes 65.5% of the
examined isolates, gave a positive result. Both COPS and CONS isolates were shown
to be enterotoxigenic, COPS represented by S. aureus species occupied the higher
ratio of the enterotoxigenic staphylococci, 86 S.aureus isolates (65.64% of the
enterotoxigenic staphylococci) gave the positive ratio of the intestine weight to the
body weight which was ≥ 0.083 while 45 CONS isolates (34.35%) were enterotoxin
producers. These toxins were thermostable staphylococcal enterotoxins which gave
the same toxic effect after heating to 100°C for 30 minutes.
Comparing the results obtained by multiplex PCR assay detecting four classical
enterotoxin coding genes, with those obtained by suckling mouse bioassay concerning
the phenotypic expression of enterotoxin coding genes, 153 (76.5%) staphylococcal
isolates harboured one or more enterotoxin coding genes, while the suckling mouse
bioassay revealed that 131 (65.5%) of those isolates produced detectable amounts of
enterotoxins.

List of Contents Page No. Title
I Summery IV List of Contents
VIII List of Figures X List of Tables
XII List of Abbreviations 1 Introduction
Page
No.
Chapter One : Literature Review Series
٤ Staphylococcus 1-1
5 Staphylococcus aureus 1-2
6 Coagulase-Negative Staphylococci (CNS) 1-3
7 virulence factors 1-4
8 Staphylococcal Enterotoxins 1-5
11 Staphylococcal Enterotoxin Properties 1-5-1
11 Staphylococcal Enterotoxin Nomenclature 1-5-2
12 Staphylococcal Enterotoxin Structure 1-5-3
14 Mechanisms of Action 1-5-4
16 Emetic effect of SEs 1-5-4-1
16 SE superantigenic property in immunopathogenesis associated
with staphylococcal food poisoning
1-5-4-2
19 Enterotoxin Gene Location 1-5-5
19 Plasmids 1-5-5-1
21 Prophages 1-5-5-2
21 Staphylococcus aureus Pathogenicity Islands 1-5-5-3
23 vSa Genomic Islands 1-5-5-4
24 Enterotoxin Genes in the Proximity of the Staphylococcal
Cassette Chromosome
1-5-5-6
25 Regulation of Enterotoxin Formation 1-5-6
25 The classical enterotoxins (SEA-SEE) 1-5-6-1
27 The non-classical enterotoxins (SElG–SElV) 1-5-6-2
29 Agents that Target the Superantigen Effect of SE 1-6
31 Staphylococcal Enterotoxins and Food Poisoning Outbreaks 1-7
33 Gastro-Intestinal Inflammatory Injury Associated with SFP 1-8
35 Antibiotic Resistance in S. aureus Correlation to Enterotoxigenic
Strains
1-9
37 Impact of Environmental Factors on SE Production 1-10
Page No.
Chapter Two : Materials and Methods Series
39 Materials 2-1 39 Laboratory Equipments and Apparatus 2-1-1 40 Chemicals and Biological Materials 2-1-2 41 Antibiotic discs 2-1-3 41 Culture Media 2-1-4 41 Ready to use culture media 2-1-4-1 42 Laboratory Prepared Media 2-1-4-2 45 Stains, Reagents, Solutions and Emulsions used in the
identification of bacterial isolates 2-1-5
47 Materials used in Agarose Gel Electrophoresis and in PCR amplification
2-1-6
48 Methods 2-2 48 Samples’Collection 2-2-1 49 Staphylococcal Isolation 2-2-2 49 Staphylococcal Isolation from Milk Samples 2-2-2-1 50 Staphylococcal Isolation from Cheese Samples 2-2-2-2 50 Staphylococcal Identification 2-2-3 50 Gram Stain 2-2-3-1 50 Growth on Mannitol Salt Agar 2-2-3-2

51 Growth on Baird-Parker Egg Yolk Tellurite Medium 2-2-3-3 52 Endopigmentation on Milk agar (Staphyloxanthin) 2-2-3-4 52 Detection of Haemolysis on Human Blood agar 2-2-3-5 52 Catalase test 2-2-3-6
5۳ Oxidase test 2-2-3-7 ۳5 Clumping Factor and Coagulase test 2-2-3-8
54 DNase production test 2-2-3-9 54 Lipase Production 2-2-3-10 54 Protease activity 2-2-3-11 54 Urease Production 2-2-3-12 55 Gelatin Liquefaction 2-2-3-13 55 Tolerance to different Concentrations of Salt 2-2-3-14 55 Acetoin production test 2-2-3-15 56 Nitrate Reduction Test 2-2-3-16 56 Motility test 2-2-3-17 56 Detection of the bacterial ability for the slime layer production 2-2-3-18 57 API-STAPH System 2-2-3-19 57 HiStaph Latex Test 2-2-4 58 Antibiotic Susceptibility Test 2-2-5 59 Preservation of Bacterial Strains 2-2-6 60 DNA Extraction 2-2-7 60 DNA Extraction from Bacterial Isolates 2-2-7-1 62 DNA Extraction from Milk Samples Directly 2-2-7-2 63 DNA Extraction from Cheese Samples 2-2-7-3 64 Estimation of DNA Concentration and Purity 2-2-8 65 Agarose Gel Preparation and Electrophoresis 2-2-9 65 Polymerase Chain Reaction (PCR) Technique 2-2-10 66 Genes selection 2-2-11 66 Primers selection 2-2-11-1 67 PCR Amplification 2-2-11-2 69 Determination of PCR Specificity 2-2-11-3 69 Detection of Thermostable Enterotoxins by Suckling Mouse
Bioassay 2-2-12
70 Statistical Analysis 2-2-13 Page No.
Chapter Three : Results and discussion Series
71 Staphylococcal Isolation and Identification 3-1 82 Detection of Virulence Factors of Staphylococci 3-2 86 Antibiotic Susceptibility 3-3

89 Molecular Studies 3-4 89 DNA extraction 3-4-1 94 Polymerase Chain Reaction (PCR) Techniques 3-4-2 94 coa gene amplification by monoplex PCR technique 3-4-2-1 98 femA and mecA genes amplification by monoplex PCR technique 3-4-2-2 107 Enterotoxigenicity detection using multiplex PCR technique 3-4-2-3 122 Detection of Staphylococcal Thermostable Enterotoxins 3-5 131 Conclusions 132 Recommendations
134 References
List of Figures Series Figure Title Page
No.
1.1 Model of SE interaction with T cell Receptors and class II MHC Molecules 18
1.2 Structure and functioning of locus agr 29
1.3 Model of the role of mucosal lamina professional and non professional APCs in SE associated Gastro-Intestinal (GI)
inflammatory injury. 34
3.1 The percentage of Staphylococcus spp. isolated from milk and cheese samples 71
3.2 Mannitol salt agar cultured with Staphylococcus spp. mannitol non
fermentor colonies of S.epidermidis and mannitol fermentor colonies of S.aureus
73
3.3 Baird-Parker Egg Yolk Tellurite agar cultured with S.aureus which appeared as black shiny convex colonies with lipase activity 74
3.4 Skim milk agar cultured with S.aureus which appeared as glistening orange convex colonies (due to Staphyloxanthin
production) with protease activity 75
3.5 The percentage of each Coagulase-Positive and Cogulase-Negative among isolated Staphylococcus spp. 76
3.6 The results of HiStaph Latex Test 77
3.7 The prevalence of Staphylococcus aureus according to all the Staphylococcal isolates isolated from milk and cheese samples 78
3.8 Slime layer producing S. aureus colonies on the congo red agar 84
3.9 S. chromogenes colonies on the congo red agar (non slime layer producers) 84
3.10 Antibiotic Susceptibility of COPS (S. aureus) and CONS isolates 86 3.11 DNA and RNA samples extracted from 11 staphylococcal isolates 89
3.12 DNA samples extracted from 2 staphylococcal isolates using hot shock with the Genomic DNA extraction kit 91
3.13 DNA bands extracted from 5 staphylococcal isolates and Staphylococcal DNA extracted from 5 cheese samples directly 92
3.14 Staphylococcal DNA extracted from 7 milk samples directly 92
3.15 Gel electrophoresis of amplified PCR products of coa gene (730 bp) of S. aureus isolates in monoplex PCR technique 94
3.16 Gel electrophoresis of amplified PCR products of femA gene (318bp) of S. aureus isolates in monoplex PCR technique 98
3.17 Gel electrophoresis of amplified PCR products of mecA gene (533bp) from staphylococci isolates by monoplex PCR technique 101
3.18 Gel electrophoresis of amplified PCR products of staphylococcal
enterotoxin genes (sea, seb, sec, and sed) in multiplex PCR technique
107
3.19 Enterotoxigenicity of both Coagulase-positive
(COPS) and Coagulase-negative Staphylococci(CONS)
113
3.20 The percentage of each staphylococcal enterotoxin gene of the four SEs genes tested (sea, seb, sec, and sed) by multiplex PCR assay 114
3.21 Staphylococcal enterotoxin genes availability among the Staphylococcal isolates 117
3.22 Distribution of the SE encoding genes (sea, seb, sec, and sed) among COPS and CONS strains 117
3.23 Enterotoxigenicity of the staphylococcal isolates isolated from milk and cheese samples, as assessed by the Suckling Mouse Bioassay 126
3.24 Diversity of Results in the biological test between coagulase-positive and coagulase-negative staphylococcal isolates 127
List of Tables Series Table Title Page
No. 1.1 Unique features of some common SEs 10 1.2 Grouping of SEs and SEls based on amino acid sequence
comparisons 12
1.3 General properties of SEs and SEls and genomic location of the 20

encoding genes 1.4 Factors affecting Staphylococcus aureus growth and enterotoxin
formation 38
2.1 Culture media 41 2.2 Milk and cheese samples 49 2.3 Colony characteristics of typical organisms on Baird-Parker Egg
Yolk Tellurite Medium 52
2.4 Diameter interpretive standards of inhibition zones according to NCCLs
59
2.5 The primers and their sequences used in conventional PCR for detection of Staphylococcus aureus
67
2.6 Program used to amplify the femA and mecA genes 68 2.7 Program used to amplify the coa gene 68 2.8 Program used to amplify enterotoxins’ genes sea, seb, sec and sed 68 3.1 The biochemical tests and their results for the staphylococcal
isolates 72
3.2 Isolated staphylococcal species from each milk and cheese samples 81 3.3 Relationship between the samples’ type and the isolated
staphylococcal species 82
3.4 Virulence factors produced by Staphylococcal isolates 85 3.5 The coa gene amplification of the S. aureus isolates 95 3.6 Comparison between coa gene harbouring and Coagulase enzyme
production by S. aureus isolates 96
3.7 The femA gene amplification results of the COPS strains obtained by monoplex PCR
99
3.8 The mecA gene amplification results of the COPS strains obtained by monoplex PCR
102
3.9 The mecA gene amplification of the CONS strains 103 3.10 Relationship between Methicillin resistance and the presence of
femA and mecA genes in both COPS and CONS 105
3.11 Results of the Amplification of enterotoxins genes (sea, seb, sec, and sed) of COPS strains by multiplex PCR
108
3.12 Results of the Amplification of enterotoxins genes (sea, seb, sec, and sed) of CONS strains by multiplex PCR
110
3.13 The biological activity of the COPS enterotoxins in Suckling Mouse Bioassay represented by the intestine weight /body weight
ratio
122
3.14 The biological activity of the CONS enterotoxins in Suckling Mouse Bioassay represented by the intestine weight /body weight
ratio
124
3.15 Effect of heat on staphylococcal enterotoxin activity 128 3.16 Comparison of the genotypic content of enterotoxigenic
staphylococcal isolates of sea, seb, sec, and sed and the phenotypic expression of enterotoxicity (detected by Suckling Mouse
Bioassay)
129

List of Abbreviations
Abbreviate Details
µg/ml Microgram per milliliter µl micro liter 5-HT 5-hydroxytryptamine AAD Antibiotic associated diarrhea Ab Antibody agr Accessory gene regulator AIP Auto Inducing Peptide APCs Antigen Presenting Cells API-STAPH Analytical Profile Index for Staphylococci identification attS Attachment site S BPA Baird Parker Agar bp base pair bsa antibiotic biosynthesis C.O.S.Q.C Central Organization for Standardization and Quality Control
CB Cannabinoid CFU/ml Colony forming unit per milliliter CHIP Chemotaxis inhibitory protein CONS Coagulase negative staphylococci COPS Coagulase positive staphylococci DNA Deoxyribonucleic acid EDTA Ethylene diamine tetra acetic acid EFSA European Food Safety Authority egc Enterotoxin gene cluster ELISA Enzyme-linked immunosorbent assay GI Gastrointestinal HLA Human leukocyte antigen IFN-gamma Interferon gamma
IL-6 Interleukin-6 kDa Kilo Dalton lpl lipoprotein-encoding genes cluster LSD Least significant difference LukED leukocidin MHC Major histocompatibility complex MRSA methicillin resistant S. aureus MSSA Methicillin susceptible S. aureus NCCLs National Committee for Clinical Laboratory Standards O.D. optical density

OB oligosaccharide/oligonucleotide binding PBP2a Penicillin-binding protein PCR Polymerase chain reaction pIB485 penicillinase plasmid, pmol picomole pvl Panton-Valentine leukocidin RAPD Random Amplification of Polymorphic DNA RNA Ribonucleic acid Rot Repressor of toxins rpm Round per minute SAgs Super antigens SaPIs S. aureus pathogenicity islands sar Staphylococcal accessory regulator SCC Staphylococcal cassette chromosome SCCmecs Staphylococcal chromosome cassette methicillin-resistance islands SCIN Staphylococcal complement inhibitor SEA Staphylococcal Enterotoxin A SEB Staphylococcal Enterotoxin B SEC Staphylococcal Enterotoxin C SED Staphylococcal Enterotoxin D SEE Staphylococcal Enterotoxin E SEG Staphylococcal Enterotoxin G SEH Staphylococcal Enterotoxin H SEI Staphylococcal Enterotoxin I SEls Staphylococcal Enterotoxin-like SEs Staphylococcal Enterotoxins SFP Staphylococcal Food Poisoning SFPOs Staphylococcal Food Poisoning Outbreaks spl serine protease gene cluster SPSS Statistical Package for Social Sience ssl Staphylococcal superantigen-like cluster SSLs streptococcal superantigens TBE Tris - Borate - EDTA TCR T-cell receptors TE Tris HCl- EDTA Th T helper cell TNF-α Tumor Necrosis Factor-alpha TSST-1 Toxic shock syndrome toxin-1 VRSA vancomycin resistant S. aureus vSa Genomic island Vβ Variable region Beta WHO World Health Organization σB sigma-B φent1 and φent2
pseudogenes

Introduction

Introduction Staphylococcal Food Poisoning (SFP) is an intoxication that results from the
consumption of improperly prepared or stored foods containing sufficient amounts of one
(or more) preformed enterotoxin (Schelin et al., 2011; Niveditha et al., 2012).
The first well-documented report, which clearly identified a Staphylococcus aureus
toxin as the cause of food poisoning outbreaks, was done by Dack et al.(1930). They
isolated a pigment-forming Staphylococcus present in large numbers in a Christmas cake
responsible for a food poisoning incident, and sterile filtrate from a broth, in which the
organism was grown, produced illness when ingested by human volunteers; initially, five
antigenic variants of S. aureus enterotoxins designated SEA through SEE were identified,
since then, new variants have been identified and designated SEH to SElR, and SElU and V
in the order that they were discovered (Ortega et al., 2010).
According to the International Committee for Staphylococcal Superantigens
Nomenclature (INCSSN), only staphylococcal superantigens that induce emesis after oral
administration in a primate model should be designated as SEs. Other related toxins that
either lack the emetic properties in this model or have not been tested should be designated
as staphylococcal enterotoxin-like (SEI) superantigens (Demir et al., 2011).
Foods that have been frequently incriminated in staphylococcal intoxication include
meat and meat products, poultry and egg products, milk and dairy products, salads, bakery
products, particularly cream-filled pastries and cakes, and sandwich fillings, salted food
products, have also been implicated according to the capacity of S. aureus to grow at
relatively low water activity (aw = 0.86; Scott, 1953) (Tasci et al., 2011). Milk is a good
substrate for S. aureus growth and enterotoxin production and dairy products are a known
source of intoxication, although pasteurization kills S. aureus cells, thermostable SEs
generally retain their biological activity; in addition, enterotoxins retain their biological
activity even after pasteurization (Janstova et al., 2012).
The European Food Safety Authority (EFSA) reported in 2009 that cheese followed by
mixed or buffet meals were the two main food vehicles in verified outbreaks of food
poisoning caused by staphylococcal toxins (EFSA, 2011).
The large numbers of carriers (more than 30-50% of the population), the contamination
of food or one of its gradients during handling, storage at unsuitable temperatures and the

capacity of the microorganism to develop in a wide range of pH, free water concentrations,
and sodium chloride concentrations- and therefore a wide range of food products- are the
main epidemiological features that create the ideal conditions for an outbreak of SFP; work
surfaces and equipments used to prepare foods are an important source for indirect
contamination (Giannatale et al., 2011).
Five conditions are required to induce Staphylococcal Food Poisoning Outbreaks
(SFPOs): (1) a source containing enterotoxin-producing staphylococci: raw materials,
healthy or infected carrier; (2) transfer of staphylococci from source to food: dirty food
preparation tools due to poor hygiene practices; (3) food composition with favorable
physicochemical characteristics for S. aureus growth and toxinogenesis; (4) favorable
temperature and sufficient time for bacterial growth and toxin production; and (5) ingestion
of food containing sufficient amounts of toxin to provoke symptoms; most SFPOs arise due
to poor hygiene practices during processing, cooking or distributing the food product
(Hennekinne et al., 2010).
Considering the facts above and that at present little is known about the occurrence of
virulence genes among the staphylococci isolated from foods in our country, so this study
aimed to:
1- Isolation and identification of the most prevalent staphylococcal species from milk and
cheese samples using bacteriological and immunological methods, and confirming this
identification by PCR technique through the amplification of the coa and femA genes.
2- Assess the prevalence of antimicrobial resistance of COPS and CONS in food (milk and
cheese) samples and investigate the incidence of methicillin resistance genes (mecA and
femA) among the staphylococcal strains isolated from these samples, and thus (from the 2nd
and 3rd aims) developing a system that differentiate methicillin resistant S. aureus (MRSA)
from mecA-positive CONS by detection both of the two genes.
3- Detection of staphylococcal enterotoxin production by an immunological method or a
bioassay.
4- Detection the presence of four classical staphylococcal enterotoxin encoding genes (sea,
seb, sec, and sed) among the isolated staphylococcal strains by multiplex PCR technique.

Chapter One
Literature Review

1- Literature review
1-1 Staphylococcus The genus Staphylococcus is a member of the Staphylococcaceae family; they are
gram-positive, cocci arranged in a grape like cluster, facultative anaerobic,
chemoorganotrophic cocci with a respiratory and fermentative metabolism at an optimal
temperature of 37°C, also, they are non-movable, nonsporulated, catalase positive and
found as pathogens or commensal organisms in both humans and animals (Argudin et al . ,
2010).
According to Euzéby (2010), there are 47 known species and 24 subspecies in the
Staphylococcus genus. Approximately half of the species are endogenous to human beings,
including S. aureus (a coagulase positive species) and coagulase-negative species: S.
epidermidis, S. haemolyticus, S. saprophyticus, S. cohnii, S. xylosus, S. lugdunensis, and S.
schleiferi (Schleifer and Kloos, 1975), S. capitis, S. warneri, S. hominis, S. simulans
(Kloos and Schleifer, 1975), S. saccharolyticus, S. auricularis (Kilpper-Balz and Schleifer,
1981), S. caprae (Devriese et al., 1983) . There are also some subspecies that are
endogenous to humans and other primates, such as S. capitis subsp. ureolyticus
(Bannerman and Kloos, 1991) and S. cohnii subsp. urealyticum (Kloos and Wolfshohl,
1991).
The rest Staphylococcal species are: S. agnetis (Taponen et al., 2012), S.
stepanovicii (Hauschild et al., 2012), S.pettenkoferi (Trülzsch et al., 2007), S.
pseudintermedius (Devriese et al., 2005) , S. simiae (Pantucek et al., 2005), S. carnosus,
S.caseolyticus, S. chromogenes, S.devriesei, S.equorum, S. felis, S. fleurettii, S. gallinarum,
S. intermedius, S. kloosii, S. lentus, S.lugdunensis, S. lutrae, S. muscae, S. massiliensis, S.
nepalensis, S. pasteuri, S. piscifermentans, S. pulvereri, S. rostri, S. sciuri, S. arlettae, S.
microti, S.auricularis, S. condimenti, S. succinus, and S. vitulinus (Euzéby, 2010).
These organisms are resistant to adverse environmental conditions and can be
recovered from non-physiological environments even months after inoculation, a peculiar
characteristic of staphylococci is their capacity to grow in high salt concentrations, and
most of them grow in media with 10% of NaCl (Vos et al., 2009; Hennekinne et al., 2010).
The species in the genus are classified based on the production of enzyme coagulase,

coagulase production capacity divides staphylococci into two major groups: coagulase
positive, including species S.aureus, S. intermedius, S. schleiferi subsp. coagulans and S.
delphini; and coagulase negative, including more than 30 different species, species S.
hyicus is variably coagulase positive and frequently included among coagulase-negative
microorganisms (Cunha, 2009a).
1-2 Staphylococcus aureus S. aureus is an extraordinarily versatile pathogen, and it can cause a large spectrum
of infections, from mild to severe and fatal. It is important in humans and also
economically important when infecting animals, able to cause superficial lesions and
systemic infections, S. aureus is responsible for toxin-mediated diseases, such as the Toxic
Shock Syndrome (TSS), Kawasaki’s Syndrome and staphylococcal food poisoning (Leung
et al., 1993;Vasconcelos and Cunha, 2010). S. aureus is known as one of the most frequent
pathogens in both community and nosocomial infections, and it can cause septicemia,
endocarditis, osteomyelitis, abscesses, pneumonia, wound infections, impetigo, cutaneous
rash, in addition to various toxin-mediated diseases, the variety of such spectrum of clinical
manifestations is mostly dependent on the numerous virulence factors produced by each
strain (Le Loir et al., 2003).
Approximately 30 - 50% of the human population carries S. aureus, and its main habitat is
the nasopharynx, a site where strains can persist as transitory or persistent members of the
normal microbiota without causing any symptomatology (Partida et al., 2010).
1-3 Coagulase-Negative Staphylococci (CONS) Originally, of all staphylococcal species, only S. aureus was considered to be
pathogenic, and it was distinguished from other species by the production of coagulase
enzyme, manitol fermentation and the presence of protein A on the cell surface (Vos et al.,
2009). Then, a great interest in Staphylococcus species, namely the Coagulase-Negative
Staphylococci (CONS), has increased due to their increasing importance in hospital
infection, particularly in nosocomial bacteremias (Brooks et al., 2007).
The CONS group contains the bacteria most frequently isolated in clinical
microbiology laboratories; however, distinguishing clinically significant pathogenic strains
from those that are only sample contaminants is one of the greatest problems faced by
clinical laboratories (Cunha et al., 2006). CONS are the major components of the normal

bacterial flora in the cutaneous system of the human body, which includes the skin and
mucosal membranes, because they are present on the skin, clinical samples are many times
contaminated during collection despite antisepsis procedures, since staphylococci present a
relatively strong degree of adherence to the epithelial cells of the dermis in addition to their
capacity of colonizing catheters and other devices, thus having access to bloodstream and
possibly causing sepsis, an infection of great clinical importance (Cunha et al., 2009b).
The increasing importance of CONS is also partly due to the acknowledgement of
this group of bacteria as essentially opportunistic as well as to the increased use of
transitory or permanent medical devices, such as intravascular catheters and prostheses in
severely impaired or immunocompromised patients, for instance those in intensive care
units, pre-term newborns, cancer and transplanted patients, in these patients, CONS
infections may be severe enough to constitute a death risk. These microorganisms exhibit
various virulence factors which are responsible for the successful invasion and infection of
their hosts (Vasconcelos and Cunha, 2010).
1-4 virulence factors The virulence factors of microorganisms in the Staphylococcus genus include surface
components, such the capsule, peptidoglycans, teichoic acid, protein A, collagen cell
attachment protein, enzymes such as lipases, esterases, fatty-acid modifying enzymes,
various proteases, hialuronidase, hydrolytic enzymes, desoxyribonucleases, catalase,
betalactamase, staphylokinase, and various toxins, such as exfoliative toxin A and B,
leukocidins, enterotoxins, TSST-1 and alpha, beta, gamma and delta hemolysins
(Vasconcelos and Cunha 2010).
Plasma coagulase is an enzyme that functions like thrombin to convert fibrinogen
into fibrin tissue, microcolonies surrounded by fibrin walls are difficult to phagocytes,
coagulase production is the principal criterion used by the clinical microbiology laboratory
for the identification of Staphylococcus aureus isolates numerous allelic forms of S. aureus
coagulase exist, with each isolate producing one or more of these enzyme variants (Kayser
et al., 2005).
The coa gene is one of the most important virulence factors for S. aureus, expression of
this gene is thought to enhance bacterial growth and promote infection in the face of host
defense mechanisms, such as phagocytosis (Karahan, and Cetinkaya 2007).

At least thirty four (34) different extracellular proteins are produced by pathogenic
Staphylococcus strains, and several of them already play a definite role in the pathogenesis
of recognized staphylococcal disease (Lisa, 2004). Some genes responsible for such factors
are frequently transported by genetic elements, such as phages and pathogenicity islets,
these are differently sized and potentially movable DNA segments which encode virulence-
related genes, and are horizontally transferred among the strains (Yamaguchi et al., 2000;
Yoshizawa et al., 2000).
Most severe infections caused by Staphylococcus sp. cannot be explained by the
action of a determined virulence factor, the action of several of such factors during the
infectious process is imperative. S. aureus strains that are capable of causing diseases
express different virulence factors, such as exotoxins, which are molecules on the cell
surface associated with adherence and with resistance to various antimicrobials, in addition
to enterotoxins, which are extracellular proteins with superantigenic activity (Omoe et al.,
2005).
1-5 Staphylococcal Enterotoxins Staphylococcal enterotoxins are members of a family of more than 20 different
staphylococcal and streptococcal exotoxins that are functionally related and share sequence
homology. These bacterial proteins are known to be pyrogenic and are connected to
significant human diseases that include food poisoning and toxic shock syndrome.
These toxins are for the most part produced by S. aureus although other species have
also been shown to be enterotoxigenic, S. aureus is persistent in 20% of the general
population, while another 60% are intermittent carriers, most frequently, the anterior nares
is the site of colonization in humans, and this colonization increases the risk of infections
when host defenses are compromised. This is supported by multiple observations, for
instance, the frequency of infections is higher in carriers than in non-carriers, non-carriers
commonly acquire infections through contaminated food or when food handlers who are
carriers contaminate food during preparation (von Eiff et al., 2001; Havelaar et al., 2010).
Staphylococcus secrets various enzymes, cytotoxins, exotoxins, and exfoliative toxins, the
chief function of these enzymes is to turn host components into nutrients that the bacteria
may use for growth. Among the other secreted factors are exotoxins that include

staphylococcal enterotoxins (SEs), and toxic shock syndrome toxin (TSST)-1, these factors
subvert the host immune system and illicit major responses (Morandi et al., 2009).
The S. aureus enterotoxins (SEs) are potent gastrointestinal exotoxins synthesized by
S. aureus throughout the logarithmic phase of growth or during the transition from the
exponential to the stationary phase (Derzelle et al., 2009). They are active in high
nanogram to low microgram quantities (Larkin et al., 2009), and are resistant to conditions
(heat treatment, low pH) that easily destroy the bacteria that produce them, and to
proteolytic enzymes, hence retaining their activity in the digestive tract after ingestion
(Argudin et al . , 2010).
Most genes coding for SEs are located on mobile elements such as plasmids,
bacteriophages or pathogenicity islands (Lindsay et al., 1998). Thus, horizontal transfer
between strains is not rare , according to Varshney et al.(2009) study most S. aureus
isolates obtained from three separate hospitals had more than one enterotoxin gene,
although there are more than 20 distinct staphylococcal enterotoxins, only a few of them
have been studied in depth.
The most common staphylococcal enterotoxins are SEA and SEB as shown in table
1-1, SEA is the most common toxin in staphylococcus-related food poisoning. While SEB
is associated with food poisoning, it has been studied for potential use as an inhaled
bioweapon (Ler et al., 2006). SED is suggested to be the second most common
staphylococcal toxin associated with food poisoning worldwide, and one study showed that
only very small amounts of this toxin were needed to induce food poisoning, SEE has also
been documented in some cases of food poisoning, while SEF has been implicated in toxic
shock syndrome (Pinchuk et al., 2010) .
SEG, SEH, and SEI are not as well studied as the others, but were associated with
one of the food poisoning outbreaks in Taiwan (Chen et al., 2004). SEH has been also
identified as one of the causes of massive food poisoning associated with the reconstituted
milk consumption in Osaka, Japan in 2000 (Ikeda et al., 2005).
Table 1-1: Unique features of some common SEs (Pinchuk et al., 2010)
Binding to Class II MHC Feature Staphylococcal Enterotoxin

Alpha and beta chains Most common toxin associated with staphylococcal food poisoning
SEA
Alpha chain Studied as a biological weapon SEB Outside the binding groove on the flanking helix from the α chain
Commonly isolated from animals SEC
Alpha and Beta chains Food poisoning SED
Beta chain Food poisoning SEE
Alpha and beta chains Associated with toxic shock syndrome
SEF
SEB-like interaction with a chain
Minor role in food poisoning SEG
Alpha chain Food poisoning SEH
Beta chain Minor role in food poisoning SEI
1-5-1 Staphylococcal Enterotoxin Properties
These SE proteins have a remarkable ability to resist heat and acid, therefore, they
may not be completely denatured by mild cooking of contaminated food. They are
pyrogenic and share some other important properties that include the ability to induce
emesis and gastroenteritis as well as their noted superantigenicity, they are resistant to
inactivation by gastrointestinal proteases including pepsin, trypsin, rennin and papain, thus
they can easily outlast the bacteria that produce them (Le Loir et al., 2003; Pinchuk et al.,
2010) .
1-5-2 Staphylococcal Enterotoxin Nomenclature Staphylococcal enterotoxins belong to the broad family of pyrogenic toxin
superantigens (SAgs); SAgs bypass conventional antigen recognition by interaction with
major histocompatibility complex (MHC) class II molecules on the surface of antigen
presenting cells, and with T-cell receptors (TCR) on specific T-cell subsets, interaction
typically occurs to the variable region of the TCR β chain (Vβ) but binding to the TCR Vα
domain has been reported, this leads to activation of a large number of T-cells followed by

proliferation and massive release of chemokines and proinflammatory cytokines that may
led to potentially lethal toxic shock syndrome (Larkin et al., 2009).
However, SEs have been proposed to be named according to their emetic activities, only
SAgs that induce vomiting after oral administration in a primate model will be designated
as SEs, related toxins that lack emetic activity or have not been tested for it should be
designated as staphylococcal enterotoxin-like (SEls) SAgs, also, newly discovered toxins
with more than 90% amino acid sequence identity with existing SEs or SEls should be
designated as a numbered subtype, however, despite this consensus nomenclature some
subtypes are still just called variants (Lina et al ., 2004; Schlievert and Case, 2007).
The repertoire of S.aureus SEs/SEls comprised 22 members, excluding molecular
variants: (i) the classical SEA, SEB, SEC (with the SEC1, SEC2 and SEC3, SEC ovine and
SEC bovine variants), SED and SEE, which were discovered in studies of S. aureus strains
involved in SFP outbreaks, and classified in distinct serological types ; and (ii) the new
types of SEs (SEG, SEH, SEI, SER, SES, SET) and SEls (SElJ, SElK, SElL, SElM, SElN,
SElO, SElP, SElQ, SElU, SElU2, and SElV) (Thomas et al., 2006). The toxic shock
staphylococcal toxin (TSST-1), initially designated as SEF, lacks emetic activity (Ono et
al., 2008; Argudin et al., 2010).
1-5-3 Staphylococcal Enterotoxin Structure
SEs and SEls constitute a family of structurally related exoproteins that range in size
from ~22 to 28 kDa, based on amino acid sequence comparisons, they have been
distributed into four or five groups (Table1- 2), depending on the inclusion or not of SEH
within group 1(Uchiyama et al., 2006; Thomas et al., 2007; Ono et al., 2008; Larkin et al.,
2009).
Table 1-2: Grouping of SEs and SEls based on amino acid sequence comparisons (Larkin et al., 2009)
Group
SEs and SEls
Group 1 SEA, SED, SEE, (SEH), SElJ, SElN, SElO, SElP, SES Group 2 SEB, SEC, SEG, SER, SElU, SElU2 Group 3 SEI, SElK, SElL, SElM, SElQ, SElV

Group 4 SET Group 5 (SEH)
Enterotoxins encoded by the egc cluster are shown in bold. SEH (in parenthesis) has been placed within Group 1 or Group 5, depending on the author (Uchiyama et al., 2006; Thomas et al., 2007).
The three-dimensional structures of TSST-1 and several SEs and SEls have been
solved by crystallography (Table 1-3). The structures are remarkably conserved, although
they interact differently with MHC class II molecules, and show different TCR specificity
(Fernandez et al., 2007). They are compact ellipsoidal proteins with two unequal domains
separated by a shallow grove. The larger C-terminal domain is a β-grasp fold consisting of
four- to five-strand β-sheet that packs against a highly conserved α-helix , the smaller N-
terminal domain consists of a mixed β-barrel with Greek-key topology, similar to the OB
(oligosaccharide/oligonucleotide binding)-fold also found in many other bacterial toxins
(SSLs, streptococcal superantigens, nucleases and toxins of the AB5 family, including
cholera and pertussis toxins, and verotoxin) (Fraser and Proft, 2008) .
The two domains are stabilized by close packing and by a section of the N-terminus
that extends over the top of the C-terminal domain. The N-terminal extension contributes
substantially to the TCR-binding site, located in the cleft between the two protein domains,
while the MHC class II binding site is in the OB-fold , the top of the N-terminal domain
usually contains a highly flexible disulfide loop, which has been implicated with emetic
activity (Thomas et al., 2007; Fraser and Proft, 2008).
Enterotoxin molecules are rich in lysine, aspartic acid, glutamic acid and tyrosine
residues. Many enterotoxins have a cystein arch that is probably involved in the molecule’s
emetic activity (Le Loir et al., 2003; Orwin et al., 2003).
The toxins SEI, SElK, SElL and SElQ, have been identified with the cysteine fold,
they were characterized as superantigens, but emetic activity is significantly reduced in
magnitude in SEI and lacks in SEK and SEQ (Fitzgerald et al., 2001; Orwin et al., 2003).
TSST-1, which does not have cysteine residues, is considered to be non-emetic (Schlievert
et al., 2000).
Toxins SEA, SEB, SEC, SED, SEE and SEH have clearly been shown to present a
greater or smaller emetic potential, depending on the molecule. Superantigenicity and

emetic activity of SEs are two separate functions located in different domains of the protein
(Hovde et al., 1994; Dinges et al., 2000).
Important efforts have been made to identify specific amino acids and domains
within SEs which may be important for emesis, but results are still limited and
controversial, like TSST-1, SElL, and SElQ are nonemetic, while SEI displays weak emetic
activity (Orwin et al., 2002).These toxins lack the disulfide loop characteristically found at
the top of the N-terminal domain of other SEs. Nonetheless, the loop itself does not appear
to be an absolute requirement for emesis, although it may stabilize a crucial conformation
important for this activity (Hovde et al., 1994).
Hoffman et al. (1996) demonstrated Carboxymethylation of histidines on SEA or
SEB generates proteins devoid of enterotoxicity, which still retain superantigenicity,
analysis of the effects of carboxymethylation of each of the SEA histidines revealed that
His61 is important for emesis, but not for T-cell proliferation . Conversely, Leu48Gly and
Phe44Ser mutant forms of SEA and SEB, respectively, do not bind MHC class II
molecules or cause T-cell activation, but still provoke vomiting, hence separating emesis
and superantigenicity as different functions of the proteins. Despite this, a high correlation
exists between the two activities since, in most cases, genetic mutations resulting in a loss
of superantigen activity also results in loss of emetic activity (Argudin et al . , 2010).
1-5-4 Mechanisms of Action In contrast to the case of many other bacterial enterotoxins, specific cells and
receptors in the digestive system have not been unequivocally linked to oral intoxication by
a SE. It has been suggested that SEs stimulate the vagus nerve in the abdominal viscera,
which transmits the signal to the vomiting center in the brain, supporting this idea,
receptors on vagal afferent neurons are essential for SEA-triggered emesis (Hu et al.,
2007), and capsaicin, a small molecular weight compound from chilli peppers that depletes
peptidergic sensory nerve fibers, also diminishes SE effects in mammals. In addition, SEs
are able to penetrate the gut lining and activate local and systemic immune responses,
release of inflammatory mediators (including histamine, leukotrienes, and neuroenteric
peptide substance P) causes vomiting and the emetic response can be eliminated by H2-
and calcium channel-blockers, which also block the release of histamine, local immune

system activation could also be responsible for the gastrointestinal damage associated with
SE ingestion .Inflammatory changes are observed in several regions of the gastrointestinal
tract, but the most severe lesions appear in the stomach and the upper part of the small
intestine (Argudin et al., 2010).
The diarrhea associated with SEs intoxication may be due to the inhibition of water
and electrolyte reabsorption in the small intestine (Bergdoll and Wong, 2006; Larkin et al.,
2009). In an attempt to link the two distinct activities of SEs, i.e., superantigenicity and
enterotoxicity, it has been postulated that enterotoxin activity could facilitate transcitosis,
enabling the toxin to enter the bloodstream and circulate through the body, thus allowing
the interaction with antigen presenting- and T-cells that leads to superantigen activity
(Kappler et al., 1997). In this way, circulation of SEs following ingestion of SEs as well as
their spread from a S. aureus infection site, could have more profound effects upon the host
versus if the toxin remains localized (Larkin et al. , 2009).
1-5-4-1 Emetic effect of SEs Although the superantigenic activity of SEs has been well characterized, the
mechanisms behind the emetic activity are poorly understood. In large part, this is due to
the dearth of adequate animal models, one animal model that seems well-suited to study the
emetic response of SEs is the house musk shrew, this small mammal that resembles a
mouse responds with vomiting two hours after peroral or intraperitoneal administration of
SEs (Hu et al ., 2003).
A study by Hu et al. (2007) who used the house musk shrew, showed that the small
intestine is a site of emetic action by SEA and appears to involve the 5-hydroxytryptamine
(5-HT) or serotonin pathway, their study showed that SEA-induced emesis was inhibited
by cannabinoid (CB) receptor agonists and the action was reversed by a CB1 antagonist.
Hu et al. (2009) showed that aspartic acid at position 227 of SEA was important in
the emetic activity, since substitution of that amino acid with alanine resulted in a molecule
devoid of emetic activity, histamine and Ca++ channel blockers have also been found to
prevent the emetic response to SEs suggesting an involvement of mast cells in enterotoxin-
induced emesis.

1-5-4-2 SE superantigenic property in immunopathogenesis associated with
staphylococcal food poisoning
Staphylococcal enterotoxins bind to class II MHC molecules on APCs outside of
the antigenic peptide binding groove (Figure 1-1), (Thibodeau et al., 1994). SEA has two
distinct binding sites on both sides of the peptide binding groove of class II MHC. SEA
molecules must be bound to both sites for optimal activity, which allows for class II MHC
crosslinking, and stable interactions with T cells (Hu et al., 2007). SED was shown to have
multiple sites of interaction with class II MHC (Al- Daccak et al., 1998; Marta et al.,
2011). SEB and TSST-1 bind to the same region of HLA-DR1, but TSST-1 is the only
staphylococcal toxin that extends part way over the peptide binding groove when bound to
class II MHC, SEE is similar structurally to SEA and binds to the same region as SEA on
the beta-chain (Karp et al . , 1992).
Fernandez et al. (2006) revealed that SEH binding site on class II MHC overlaps
with one of the SEA binding sites, and SEI binds to the HLA-DR1 beta-chain. Once bound
to class II MHC, SEs may then bind to T cells via the T cell receptor (TCR). T cells
normally require presentation of a specific antigenic peptide to the TCR by APCs in order
to become activated, SEs interact with T cells in a ‟nonspecific” manner, only requiring a
common variable region on the TCR (Figure 1-1). This MHC class II: SEs: TCR tri-
molecular interaction leads to an uncontrolled release of various proinflammatory
cytokines including IFN-gamma, TNF-α, IL-1β, IL-6 and IL-8, the key
cytokines/chemokines causing superantigen-mediated acute inflammation and shock
(Assenmacker et al., 1998; Pinchuk et al., 2010).
Whereas T cells are normally only activated in an antigenic specific way, their
interaction with SEs leads to a massive proliferation and differentiation of T cells
predominantly toward Th1 and Th17 phenotypes both of which are associated with acute
inflammatory responses (Grumann et al ., 2008).
The extensive inflammation induced by the immune response to SEs leads to an
increase in intestinal epithelial permeability and a decrease in expression of tight junction
proteins. Disruption of barrier function leads to an influx of antigens through the mucosal
layer, further activating immune responses to these antigens as they interact with immune

cells. SEs are able to cross the epithelial barrier intact and, by traversing this barrier, gain
access to T cells (Pinchuk et al., 2010).
Figure 1-1: Model of SE interaction with T cell Receptors and class II
MHC Molecules (Pinchuk et al., 2010)
Kappler et al. (1997) demonstrated that SEB was more efficient at traversing the
epithelial barrier than SEA, and thus, is more likely to reach the blood, in addition to
inducing T cell responses, SEs also induce proinflammatory responses from professional
and non professional APCs when binding to MHC class II on these cells. In a mouse
model, SEA, SEB, and TSST-1 were able to induce dendritic cell migration and maturation
dependant on T cell activation, macrophages are also activated by SEs and upon binding
release neutrophil chemotactic factors that induce neutrophil migration and increased
release of proinflammatory cytokines (Desouza et al., 2002; Muraille et al., 2002).
A lot of studies suggested that SEs may bind to a variety of cell types via MHC class
II molecules and these interactions leads to their activation resulting in proinflammatory
cytokines and chemokines production and uncontrolled activation of T cells (Fischer et al.,

1989; Krakauer et al., 1994; Fromont et al., 1995; Fujisawa et al., 1998; Byrne et al., 2002;
Pinchuk et al., 2007).
1-5-5 Enterotoxin Gene Location
All se and sel genes are located on accessory genetic elements, including plasmids,
prophages, S. aureus pathogenicity islands (SaPIs), genomic island vSa, or next to the
staphylococcal cassette chromosome (SCC) elements (Table 1-3). Most of these are mobile
genetic elements, and their spread among S. aureus isolates can modify their ability to
cause disease and contribute to the evolution of this important pathogen (Argudin et al.,
2010).
1-5-5-1 Plasmids
Plasmids have been long recognized as efficient vehicles for the spread of resistance
and virulence determinants through horizontal gene transfer. In S. aureus, two kinds of
plasmids carrying se/sel genes have been characterized (Table 1-3). Both contain selj and
ser associated with either sed (pIB485-like) or with ses and set (pF5) (Ono et al., 2008).
The first plasmid described to carry an enterotoxin gene was pIB485, a 27.6 kilobase (kb)
plasmid, in which first sed and latter selj were identified (Bayles and Iandolo, 1989).
Enterotoxin SER was discovered by Omoe et al. (2003) in S. aureus strains associated with
a food poisoning outbreak that occurred in Fukuoka City, Japan, in 1997, and the ser gene
was shown to be located on a family of closely related plasmids, termed pF5 and pF5-like,
these plasmids have similar restriction profiles and carry selj along with ser. Later two
novel SE genes (ses and set) have also been detected on the Fukuoka plasmids (Ono et al.,
2008). Interestingly, the ser gene, together with sed and selj, has also been found in
pIB485-like plasmids from laboratory strains, food poisoning outbreak isolates and healthy
human isolates in Japan (Omoe et al., 2003) and pIB485-like plasmids, varying in size
and/or restriction profile were present in S. aureus isolates recovered in Spain from human
nasal carriers and manually handled foods, two of them; named pUO-Sa-SED1 (~33 kb)
and pUO-Sa-SED2 (~36 kb), carried sed, selj and ser, and have restriction patterns
identical or similar to that of pIB485, while pUO-Sa-SED3 (53.5 kb; containing sed, selj
and ser-like) has a different profile (Fueyo et al., 2005). A blast search of the sed, selj, ser,
ses and set genes revealed additional pIB485-like and pF5-like plasmids obtained from
human clinical isolates, the evolutionary relationship between the two types of plasmids is
unknown (Argudin et al ., 2010) .
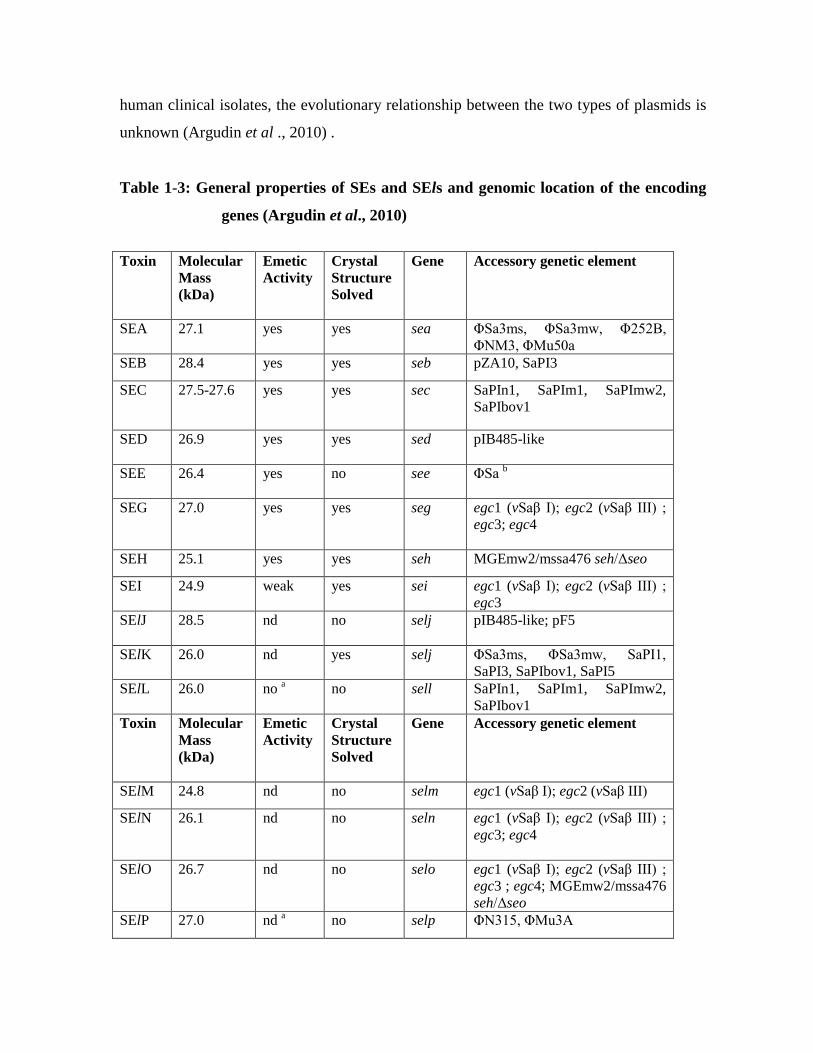
Table 1-3: General properties of SEs and SEls and genomic location of the encoding
genes (Argudin et al., 2010)
Toxin Molecular
Mass (kDa)
Emetic Activity
Crystal Structure Solved
Gene
Accessory genetic element
SEA
27.1 yes yes sea ΦSa3ms, ΦSa3mw, Φ252B, ΦNM3, ΦMu50a
SEB 28.4 yes yes seb pZA10, SaPI3
SEC 27.5-27.6 yes yes sec SaPIn1, SaPIm1, SaPImw2, SaPIbov1
SED 26.9 yes yes sed pIB485-like
SEE 26.4 yes no see ΦSa b
SEG
27.0 yes yes seg egc1 (vSaβ I); egc2 (vSaβ III) ; egc3; egc4
SEH 25.1 yes yes seh MGEmw2/mssa476 seh/Δseo
SEI 24.9 weak yes sei egc1 (vSaβ I); egc2 (vSaβ III) ; egc3
SElJ
28.5 nd no selj pIB485-like; pF5
SElK 26.0 nd yes selj ΦSa3ms, ΦSa3mw, SaPI1, SaPI3, SaPIbov1, SaPI5
SElL 26.0 no a no sell SaPIn1, SaPIm1, SaPImw2, SaPIbov1
Toxin Molecular Mass (kDa)
Emetic Activity
Crystal Structure Solved
Gene
Accessory genetic element
SElM 24.8 nd no selm egc1 (vSaβ I); egc2 (vSaβ III)
SElN 26.1 nd no seln egc1 (vSaβ I); egc2 (vSaβ III) ; egc3; egc4
SElO 26.7 nd no selo egc1 (vSaβ I); egc2 (vSaβ III) ; egc3 ; egc4; MGEmw2/mssa476 seh/Δseo
SElP 27.0 nd a no selp ΦN315, ΦMu3A
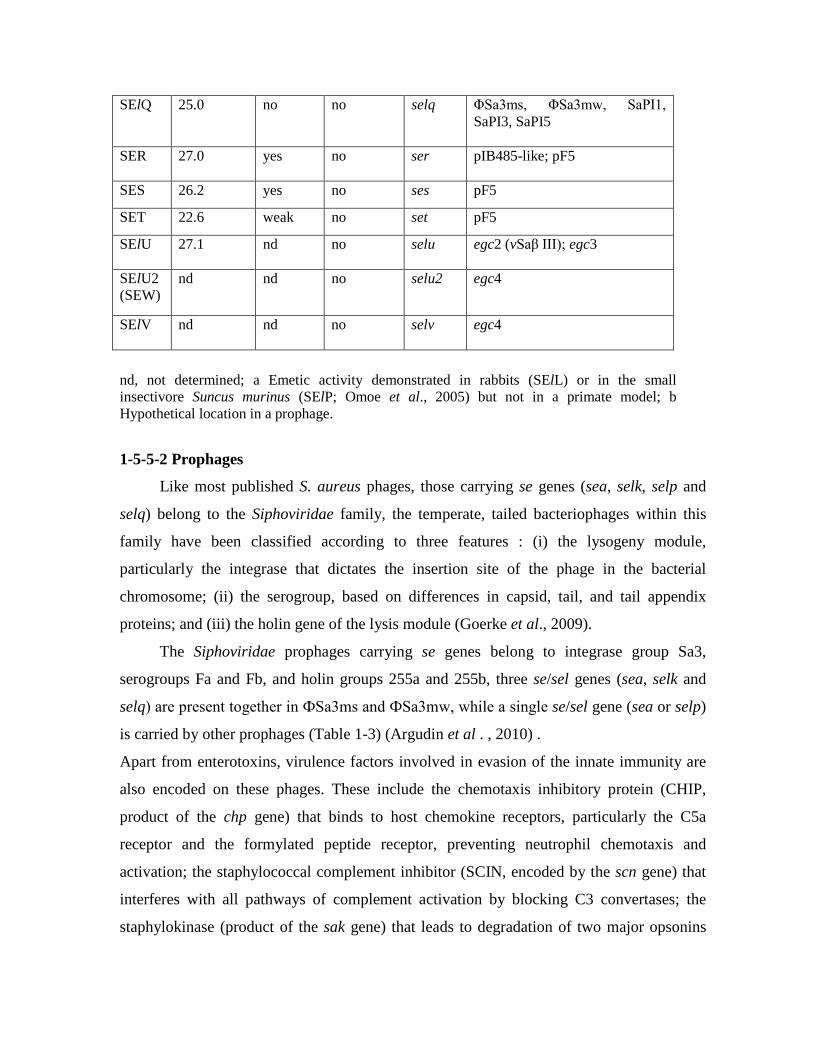
SElQ 25.0 no no selq ΦSa3ms, ΦSa3mw, SaPI1, SaPI3, SaPI5
SER 27.0 yes no ser pIB485-like; pF5
SES 26.2 yes no ses pF5
SET 22.6 weak no set pF5
SElU
27.1 nd no selu egc2 (vSaβ III); egc3
SElU2 (SEW)
nd nd no selu2 egc4
SElV
nd nd no selv egc4
nd, not determined; a Emetic activity demonstrated in rabbits (SElL) or in the small insectivore Suncus murinus (SElP; Omoe et al., 2005) but not in a primate model; b Hypothetical location in a prophage.
1-5-5-2 Prophages
Like most published S. aureus phages, those carrying se genes (sea, selk, selp and
selq) belong to the Siphoviridae family, the temperate, tailed bacteriophages within this
family have been classified according to three features : (i) the lysogeny module,
particularly the integrase that dictates the insertion site of the phage in the bacterial
chromosome; (ii) the serogroup, based on differences in capsid, tail, and tail appendix
proteins; and (iii) the holin gene of the lysis module (Goerke et al., 2009).
The Siphoviridae prophages carrying se genes belong to integrase group Sa3,
serogroups Fa and Fb, and holin groups 255a and 255b, three se/sel genes (sea, selk and
selq) are present together in ФSa3ms and ФSa3mw, while a single se/sel gene (sea or selp)
is carried by other prophages (Table 1-3) (Argudin et al . , 2010) .
Apart from enterotoxins, virulence factors involved in evasion of the innate immunity are
also encoded on these phages. These include the chemotaxis inhibitory protein (CHIP,
product of the chp gene) that binds to host chemokine receptors, particularly the C5a
receptor and the formylated peptide receptor, preventing neutrophil chemotaxis and
activation; the staphylococcal complement inhibitor (SCIN, encoded by the scn gene) that
interferes with all pathways of complement activation by blocking C3 convertases; the
staphylokinase (product of the sak gene) that leads to degradation of two major opsonins

(IgG and C3b) through activation of surface-bound plasminogen into plasmin, and also
inhibits the bactericidal effect of α-defensins, the region encoding these virulence factors is
known as the "innate inmune evasion cluster" and is located at one or both ends of the
phages (van Wamel et al., 2006 ; Goerke et al., 2009).
1-5-5-3 Staphylococcus aureus Pathogenicity Islands The Staphylococcus aureus Pathogenicity Islands (SaPIs) are mobile pathogenicity
islands, which are widely distributed in S. aureus and have also been found in other species
of Staphylococcus. SaPIs have a highly conserved overall organization, parallel to that of
typical temperate bateriophages. Each one occupies a specific chromosomal site (attS), and
always appears in the same orientation. From its integration site, the island can be induced
to excise and replicate by one or more specific staphylococcal helper phages (Tallent et al.,
2007). Following replication the SaPI DNA is efficiently encapsidated into infectious
small-headed phage-like particles resulting in extremely high transfer frequencies (Argudin
et al., 2010).
SaPIs are very common in S. aureus (Table 1-3). They range in size from 15–17 kb,
with the exceptions of SaPIbov2 (27 kb) and a highly degenerated SaPI (3.14 kb) present in
some sequenced genomes. The complete nucleotide sequence is known for 20 SaPIs, and
some of them carry genes encoding TSST-1 and/or one or more SEs , for instance, tst is
found together with selk and selq in SaPI1, with sec3 and sell in SaPIm1 and SaPIn1, and
with sell and sec in SaPIbov1; seb, selq and selk have been reported in SaPI3; selk and selq
in SaPI5; and sec4 and sell2 in SaPImw2 , the induction of a SaPI is likely to originate an
increase in the copy number of the toxin genes, and therefore to an increase in toxin
production, as described for lysogenic phages (Novick and Subedi, 2007; Baba et al.,
2008).
1-5-5-4 vSa Genomic Islands
The term vSa refers to non-phage and non-SCC genomic islands that are exclusively
present in S. aureus, often (but not always) encode virulence determinants, are inserted at
specific loci in the chromosome (Baba et al., 2008).
According to Argudin et al. (2010), two major vSa genomic islands, namely vSaα
and vSaβ, each of about 20–30 kb, are present in all S. aureus genomes sequenced so far,

but absent in other Staphylococcus species, including S. epidermidis. Though vSaα and
vSaβ could have been acquired by horizontal gene transfer, actually there is not evidence
that they can move. Both vSaα and vSaβ contain clusters of genes encoding known or
putative virulence factors, vSaα carries a cluster of lipoprotein-encoding genes (lpl cluster),
and the sel (staphylococcal exotoxin-like) cluster which then re-named as the ssl
(staphylococcal superantigen-like) cluster (Chung et al., 2007). vSaβ carries a serine
protease gene (spl) cluster, genes for the components of the LukED leukocidin (lukD and
lukE), genes for antibiotic biosynthesis (bsa) and/or the enterotoxin gene cluster (egc),
which includes a variable number of se/sel genes forming an operon (Fraser and Proft,
2008). Two representative types of vSaβ, the genomic island carrying se genes have been
described vSaβ and vSaβ III (Baba et al., 2008).
Jarraud et al. (2001) had discovered the first egc (egc1) which consists of two SE genes
(seg and sei), three SEl genes (selm, seln and selo), and two pseudogenes (φent1 and φent2)
afterward, a second egc variant (egc2) containing an additional SEl gene (selu) was
described (Letertre et al., 2003). In addition, allelic variants of each of the egc2 genes
compose the egc3 cluster (Thomas et al., 2006; Collery et al., 2009). Moreover, the fact
that each of the three major homology groups of SEs/SEls (Table 1-2) contains
enterotoxins encoded by genes of the egc operon led to the proposal that all se/sels
originated from the egc cluster (Thomas et al., 2007).
1-5-5-6 Enterotoxin genes in the Proximity of the Staphylococcal
Cassette Chromosome
There are some enterotoxin genes occurs in the Proximity of the Staphylococcal
Cassette Chromosome (SCC), the seh gene, flanked by a truncated selo gene and a putative
transposase gene, have been found in close proximity of the non-mecA containing SCC
element harbored by MSSA (methicillin susceptible S. aureus) strain 476; the SCCmec
type IV of S. aureus MW2; and the SCCmec type IV of a collection of highly related
community-associated S. aureus , in the latter strains, acquisition of the seh element could
have stabilized the integration of SCCmec type IV, which is unable to excise (Noto and
Archer, 2006) .

1-5-6 Regulation of Enterotoxin Formation
The best-known staphylococcal regulatory systems are agr (accessory gene
regulator) (Peng et al., 1988; Vasconcelos and Cunha, 2010), sar (staphylococcal accessory
regulator, divided into sarA, sarS, sarT and sarR) (Cheung et al., 1992; McCulloch, 2006)
and rot (repressor of toxins) (McNamara et al., 2000), which can directly affect
staphylococcal enterotoxin production. There are also regulatory systems saeRS, σB
(sigma-B), arlRS (McCulloch, 2006) and srrAB (Throup et al., 2001; Yarwood et al.,
2001).
1-5-6-1 The classical enterotoxins (SEA-SEE)
I-Prophage-encoded enterotoxins (sea and see)
The sea gene is carried by a polymorphic family of temperate bacteriophages, the
bacteriophage is inserted into the bacterial chromosome as a prophage and behaves like
part of the bacterial genome, however, under environmental stress conditions, such as mild
food preservation conditions, the prophage can be induced to replicate the phage genome
and release new bacteriophages (Wallin-Carlquist et al., 2010).
There are at least six completely sequenced S. aureus strains containing different sea-
carrying prophages, Φ252B, ΦMu3, ΦMu50A, ΦNM3, ΦSa3ms and ΦSa3mw, have been
found, all of which frequently carry the genes for enterotoxin A, staphylokinase and the
complement inhibitor, it was demonstrated that the transcription of sea is linked to some
extent to the lifecycle of the SEA-encoding prophage, in contrast to many other non-phage
encoded enterotoxin genes such as seb, sec and sed (Goerke et al.,2009).
The see gene is situated on a defective prophage, in contrast to the prophage encoding sea
and see expression appears to be unaffected by bacterial growth (Derazelle et al., 2009).
II-agr-regulated enterotoxins (seb, sec and sed)
The seb gene is carried on the S. aureus pathogenicity island, SaPI3, while enterotoxin C
(SEC) exists in multiple variants, C1, C3, Cbov, which are situated on SaPI4, SaPIn1/m1
and SaPIbov, respectively (Novick et al.,2010). The sed gene is situated on a 27.6 kb
penicillinase plasmid, pIB485, in S. aureus (Marta et al., 2011). Despite being encoded by

different mobile genetic elements, the expression of seb, sec and sed genes is induced
during the transition from the exponential to the stationary phase, an expression pattern
characteristic of proteins encoded by genes regulated by the Agr regulatory system, the two
se genes encoded by SaPIs, seb and sec, undergo a much more drastic induction than the
plasmid-encoded sed (Derazelle et al.,2009).
The agr is a group of genes with quorum sensing activity which regulate the expression of
various virulence factors; Quorum sensing is the name given to the mechanism of
“communication” among bacteria by means of which a bacterium can “perceive” the
population density in the medium, this mechanism is important in Staphylococcus since
some accessory proteins (such as virulence factors) are only expressed in certain growth
phases (Thoendel et al., 2011).
The agr locus generates two different transcripts, RNAII and RNAIII, driven by the
promoters P2 and P3, respectively (Figure 1-2). RNAII encodes the structural genes for the
quorum sensing system agrB, agrD, agrC and agrA. AgrD and AgrB act to generate the
quorum sensing molecule [autoinducing peptide (AIP)], which after reaching a threshold
level stimulates activation of AgrC and AgrA, a two component regulatory system.
Activated AgrA then upregulates the promoters P2 and P3, generating more RNAII and
RNAIII transcripts. The P3 transcript, RNAIII, encodes delta-hemolysin but, more
importantly, the RNAIII itself is the intracellular effector of gene regulation in the cell
(Novick and Geisinger, 2008). As the cell grows the intracellular level of RNAIII increases
due to the autoregulatory circuit of the Agr system, leading to increased transcription of
secreted virulence factors such as enterotoxins, and reduced transcription of a subset of
genes encoding cell wall proteins, loss of the Agr signal transduction system is reported to
result in substantial loss in the transcript level of seb, sec and sed and thus the
corresponding SEB, SEC and SED production (Schelin et al., 2011).
The RNAIII-mediated impact on the transcription of seb and sed is indirect and is
dependent on the presence of a functional Rot (repressor of toxins), which is a member of
the Sar family of transcriptional factors of S. aureus. Rot binds to promoter regions, as
shown for the seb promoter, thereby repressing the transcription of genes , when the Agr
system is induced during post-exponential growth RNAIII base pairs with rot mRNA, this

mediates translational repression of rot mRNA, and subsequently lowers the amount of
cellular Rot (Biosset et al., 2007).
The enterotoxins B, C and D are, however, only partially upregulated by the Agr system
and can be produced independently of agr. Although SarA is required for full agr loci
transcription, SarA has also been shown to regulate seb transcription independently of
RNAIII, and the alternative sigma factor, sigmaB, has been reported to reduce seb
expression, possibly by repressing both the agr system and a second unidentified inducer,
notably, many of the environmental conditions known to repress seb transcription, such as
high salt content and alkaline conditions, are also known activators of sigma B (Fujimoto et
al., 2009).
1-5-6-2 The non-classical enterotoxins (SElG–SElV)
Regarding regulation of the non-classical enterotoxins, results from a kinetic study indicate
that the expression of the majority of the newly described se genes is not controlled by the
agr system (Derazelle et al., 2009).
Only the transcript level of seh, ser and sel increases in the post-exponential phase, which
implies possible regulation by the Agr regulatory system. seh mRNA was found to undergo
a much more drastic induction than ser and sel, and activation of seh took place earlier in
the growth cycle than the classical agr-controlled seb and sed genes, this expression pattern
is consistent with results reported showing that maximal SEH production takes place in the
late exponential phase, while SEB is mainly produced in the stationary phase (Sakai et al.,
2008; Derazelle et al., 2009). The transcript level of other investigated se genes either
remained unchanged during growth (sej, sek, seq, sep), or decreased slightly (seg, sei, sem,
sen, seo, seu) after exponential growth. Most of the se genes with unchanged transcription
are phage-encoded, and may therefore be regulated by the processes that govern lysogeny,
in contrast, the se genes that showed a slight decrease in transcript level during growth are
encoded by the egc operon and, notably, these enterotoxins could not be detected using
two-dimensional gel electrophoresis (Pocsfalvi et al., 2008). It is still unclear whether the
non-classical enterotoxins are responsible for food poisoning, and so far SEH is the only
non-classical enterotoxin detected in foods responsible for food poisoning (Ikeda et al.,
2005).

The interaction of all regulatory systems shows the complexity of regulation of
accessory genes in Staphylococcus. Positive- and negative-feedback events occur among
them, amplifying or inhibiting signs, and a competition takes place between such systems
to regulate a gene. For instance, the agr system increases the expression of alpha-
hemolysin, whereas, the arlRS system decreases the expression of the same gene
(McCulloch, 2006).
Figure 1-2: Structure and functioning of locus agr
(Vasconcelos and Cunha, 2010)
1-6 Agents that Target the Superantigen Effect of SE Despite all the advances in the understanding of the SE mechanism of action, the SE-
associated diarrheal disease due to food poisoning or nosocomial S. aureus infection is of
major concern in health programs worldwide. World Health Organization (WHO) pointed
out in 2003 that the best approach to reduce the number of food poisoning-related disease
outbreaks are preventative measures and treatments against SEs, the preventive measures
include stricter food control, hand and environmental hygiene, identification and isolation
of carriers, and proper S. aureus antibiotic therapy (Much et al ., 2009; Lin et al ., 2010) .
SE-associated diarrheal disease symptoms are abrupt, and may be severe enough to
warrant hospitalization, antimicrobial agents with activity against S. aureus should be given
to all patients with suspected toxic shock syndrome and MRSA infections (Murray, 2005).
However, the increase in MRSA strains poses a challenge to efficient therapy, therefore,

novel ways targeting the prevention of SE production by S. aureus or
blocking/neutralization of SE interaction with the host are required to ameliorate the
disease outcome (Cooper et al ., 2004; Gbaguidi et al ., 2009).
SE immunopathological effects are strongly associated with their capacity to act as
superantigens. Thus, the SE superantigenic properties represent a very attractive
therapeutic target, and many potential targets to prevent the toxic effects of bacterial
superantigens have been well reviewed by Krakauer in 2005, Fraeser and Proft in 2008,
and Larkin et al. in 2009.
Since the discovery of SE structures and immune receptors, multiple
immunotherapeutic strategies have been proposed. Those strategies include neutralization
of SEs by intravenous Ig therapy that consists of anti-SE polyclonal Abs from multiple
donors (Yanagisawa et al., 2007), blocking the interaction of SEs with MHC class II or
TCR (Buonpane et al., 2007; Yang et al., 2008).
The inhibition of SE-induced proinflammatory cytokine/chemokine cascade by using
neutralizing Abs, anti-inflammatory cytokine (e.g., IL-10), or potent immunosuppressants,
a setrategy proposed by Pender et al. (1998), Stiles et al.(1999), and Pinchuk et al. (2007),
while Lui et al.(2009), Krakauer et al. (2010), and Tilahun et al. (2010) proposed another
strategy, included the inhibition of signal transduction pathways activated by these
superantigens, particularly NF- k B.
More than one study proposed original approach was to use of the innate immunity
modulators (Hayworth et al., 2009; Perez-Bosque et al., 2010; Tilahun et al., 2010). For
instance, Hayworth et al.(2009) demonstrated that bovine lactoferrin was able to attenuate
SEB-induced proliferation, IL-2 production, and CD25 expression in HLA-DR4 transgenic
mouse T cells , this inhibition was due to the lactoferrin iron-binding capacity. Dietary
plasma protein supplements have been shown to prevent release of SEB-induced mucosal
proinflammatory mediators (IFN-γ, TNF- α, IL-6 and LTB4) in rats (Perez-Bosque et al.,
2010).
All the available data has demonstrated that the early blockade of the mechanisms
involved in the SE induced hyperactivation of immune responses may represent attractive
strategy for the development of new specific anti-SE therapeutic approaches.

1-7 Staphylococcal Enterotoxins and Food Poisoning Outbreaks Independently of their origin, enterotoxigenic S. aureus often differ in the number of
mobile genetic elements and se/sel genes therein, as well as in the enterotoxins they
produce. SEA, either alone or together with other SEs/SEls, is the enterotoxin most
commonly reported in foods, and is also considered as the main cause of SFP, probably due
to its extraordinarily high resistance to proteolytic enzymes (Balaban and Rasoolly 2000;
Argudin et al., 2010) .
Several studies have investigated the distribution of SEs and se/sel genes in S. aureus
from foods and SFP outbreaks in Asian countries. Among strains recovered from patients
associated with SFP outbreaks during 2001-2003 in Taiwan, sea was the most common
gene, followed by seb and sec (Chiang et al., 2008).
In Korea, about 90% of food poisoning isolates were reported to contain the sea gene
(Cha et al., 2006). SEA also was the most common SE associated to SFP in Japan (Shimizu
et al., 2000). In this country, an extensive outbreak that occurred in 2000 was attributed to
low-fat milk containing SEA (Asao et al., 2003), while another outbreak (2009) was due to
crepes containing SEA and SEC (Kitamoto et al., 2009).
SEB, SEC or SED alone have been also implicated in SFP outbreaks through the
world (Kerouanton et al., 2007; Veras et al., 2008). Interestingly, an outbreak, which
affected three members of the same family in USA, was caused by coleslaw-containing
SEC produced by a community-acquired methicillin resistant S. aureus from an
asymptomatic food handler (Jones et al., 2002).
The fifth classical enterotoxin, SEE, has been infrequently reported in foods and
food-producing animals, and its involvement in SFP outbreaks has only been demonstrated
in rare occasions. However, six SFP outbreaks, which occurred in France at the end of
2009, were caused by SEE present in soft cheese made from unpasteurized milk. This
enterotoxin has also been associated with outbreaks in USA and UK (Mc Lauchlin et al.,
2000; Ostyn et al., 2010).
In contrast to classical SEs, the relationship between the novel SEs/SEls and SFP is
not fully understood. Among them, SEG, SEH and SEI, SER, SES, and SET have shown to
be emetic after oral administration in a primate model, while the emetic activity of SElL

and SElP has only been demonstrated in rabbits and the small insectivore Suncus murinus,
respectively (Orwin et al., 2003; Omoe et al., 2005).
The remaining SEls either lack emetic properties (SElQ), or have not been tested
(SElJ, SElK, SElM, SElN, SElO, SElU, SElU2 and SElV). Moreover, commercial kits are
not available for immunological detection of these SEs and SEls, although ELISA
(enzyme-linked immunosorbent assay) has been described for SEH (Su and Wong 1996)
and for SEG and SEI (Omoe et al., 2002). Of the new enterotoxins, only SEH-producing
strains have clearly been involved in SFP outbreaks (Ikeda et al., 2005; Jorgensen et al.,
2005a), but results from different researchers have shown the high incidence of genes
encoding new SEs and SEls among food-borne S. aureus (Blaiotta et al., 2004 ; Bania et
al., 2006 ) .
McLauchlin et al. (2000) revealed that 23 staphylococcal strains implicated in SFP
outbreaks in UK, in which classical se genes were not detected, harbored one or more of
the new se/sel genes, i.e., seg, seh, sei or selj. It is possible that the corresponding SEs
might have been the cause of these outbreaks. The presence of egc genes was also shown in
food-associated S. aureus from other countries (Martin et al., 2004; Jorgensen et al.,
2005b; Bania et al., 2006).
In 2007 Hwang et al. revealed that the newly described SE or SEl genes, particularly
those belonging to the egc cluster, were more frequently detected in S. aureus strains
isolated from raw pork and chicken meat in Korea than genes encoding classical SEs.
Despite this, egc-encoded SEs or SEls have not yet been directly implied in typical cases of
SFP, although SEG and SEI have been reported as the cause of chronic diarrhea associated
with severe but reversible enteropathy in two malnourished neonates (Naik et al., 2008).
1-8 Gastro-Intestinal Inflammatory Injury Associated with SFP
SFP leads to inflammatory changes throughout the gastrointestinal tract with severe
lesions in the jejunum and ileum. The direct inhibitory effect of purified SEs on intestinal
tone, contractility and colonic transit has been noted in the laboratory animals, ingestion of
SEs within food cause food poisoning, which is characterized by severe vomiting and
diarrhea, those symptoms occur within hours after eating of SE-contaminated food
(Hennekinne et al., 2010; Schelin et al., 2011).

Despite the significant progress in the understanding of SE associated inflammation of GI
tract, it is still unclear how this inflammation is initiated in vivo and what is the exact role
of each of the immune and non immune cells that contribute to the progression of the
disease. Many recent ex vivo and in vitro studies suggest that GI inflammatory injury
associated with staphylococcal enterotoxigenic disease is mediated mostly through the SE
superantigenic effect on MHC class II expressing APCs and CD4+ T cells, cells expressing
major receptors for SEs (Pinchuk et al., 2007). Those interactions may result in
hyperactivation of professional as well as non-professional APCs and T cells leading to the
excessive proliferation of CD4+ T cells and the release of proinflammatory cytokines and
chemokines that contribute to the SE inflammatory effect on GI tract (Figure 1-3) (Pinchuk
et al., 2010).
Figure 1-3: Model of the role of mucosal lamina professional and non
professional APCs in SE associated Gastro-Intestinal (GI) inflammatory injury (Pinchuk et al., 2010).
GI inflammatory injury during SFP is mediated mostly through the SE superantigenic
effect on MHC class II expressing mucosal professional (macrophages and dendritic cells,
DC) and non professional (such as myofibroblasts) APCs and TCR expressing CD4+ T
cells.
SE can cross the intestinal epithelial barrier in intact form and bind to class II MHC
molecules that expressed on subepithelial myofibroblast. These processes will lead to a

strong production of the proinflammatory cytokines and chemokines, including IL-6, IL-8
and MCP-1. The last one may leads to the increased chemotaxis of professional immune
cells (CD4+ T cells, Macrophages, DC) from gut associated lymphoid tissue (GALT) to the
site of SE associated inflammation in GI mucosa. Those MHC class II: SEs:TCR
interactions may in turn result in hyperactivation of the APCs and the T cells leading to the
excessive proliferation of T cells and the uncontrolled burst of various proinflammatory
cytokines and chemokines causing the superantigen-mediated acute inflammation and
shock (Xu and McCormick 2012).
1-9 Antibiotic Resistance in S. aureus Correlation to Enterotoxigenic Strains
Food is an important factor for the transfer of antibiotic resistance. Such transfer can occur
by means of antibiotic residues in food, through the transfer of resistant food-borne
pathogens or through the ingestion of resistant strains of the original food microflora and
resistance transfer to pathogenic microorganisms, S. aureus strains are known to be
frequently resistant to antibiotic therapy due to their capacity to produce an
exopolysaccharide barrier and because of their location within microabscesses, which limit
the action of drugs (Gündogan et al., 2006; Pesavento et al., 2007).
As previously related, it is well known that determinants of resistance to antibiotics and
other toxic substances in staphylococci, as in other pathogens, are largely carried by
accessory genetic elements, especially plasmids, transposons and their relatives, a
particularly important resistance determinant in staphylococci is that for methicillin and
other β -lactam compounds, giving rise to the notorious MRSA acronym. Although it has
been clear for some time that a novel penicillin-binding protein, PBP2a, is responsible for
this resistance, and that the PBP2a gene is not native to S. aureus, there has been
considerable uncertainty regarding the nature of the accessory genetic elements that carry
it. These were thought at one time to be plasmids or transposons, but then have been shown
definitively to be a family of large chromosomal insertions that belong to the general
category of chromosomal islands and are appropriately regarded as resistance islands. The
staphylococcal pathogenicity islands (SaPIs) and staphylococcal chromosome cassette
methicillin-resistance islands (SCCmecs) represent two well-defined classes of novel
mobile elements that are presumably responsible for the horizontal transmission of SAg

and certain resistance determinants. It seems remarkable that the SaPIs carry only SAg
determinants and the SCCmecs carry only resistance determinants (Ortega et al., 2010).
Several reports had analyzed the prevalence of enterotoxin genes in methicillin-resistant
and methicillin-susceptible (MSSA) isolates of S. aureus, Sila et al. (2009) found 7 genes
more frequently detected in MRSA isolates: sea, seb, sed, seg, sei, sej and eta, coding for
the production of enterotoxins A, B, D, G, I, J and the exfoliative toxin A. On the other
hand, the pvl, tst and sec genes for Panton-Valentine leukocidin, TSST-1 and enterotoxin C
were most frequent in MSSA. However, the role and the source of food contamination are
still unclear since only few reports on the presence and possible origin of MRSA in foods
are available.
The correlation of antibiotic resistance and enterotoxigenicity of S. aureus has not been
clearly supported by published results showing that food poisoning caused by MRSA is
different of that caused by MSSA, except for the difference in prevalence of SE genes in
these two populations. In turn the role of antibiotic resistance in the pathogenicity of SE-
producing S. aureus has been revealed under specific conditions, e.g., such as those leading
to AAD (antibiotic associated diarrhea) development. Alteration of gut flora by antibiotic
therapy seems to be important in regard to the expression of pathogenic properties of
intestinal MRSA, frequently the only bacteria able to survive the presence of
antimicrobials. Clinical parameters are higher in AAD patients colonized with
enterotoxigenic MRSA than in those suffering from diarrhea not associated with MRSA or
colonized with non-enterotoxigenic MRSA (Boyce and Havill 2005).
1-10 Impact of Environmental Factors on SE Production SFP is often associated with growth in protein-rich food such as meat and dairy
products, these products are highly complex matrices compared with broth, with respect to,
e.g., microbial content, salt, pH, nutrient availability, oxygen availability and temperature
(Valero et al., 2009). Generally, growth of S. aureus is necessary for enterotoxin
production, although enterotoxin production does not always accompany growth, and in a
few cases toxin production has been observed in non-replicating cell cultures, as
documented by Wallin-Carlquist et al. (2010). Studies have been performed to identify key
parameters that prevent or stimulate enterotoxin production in laboratory media and in
diverse food products, and a multifaceted network of environmental and genetic factors

seems to regulate enterotoxin production and some of the identified effects of
environmental conditions on enterotoxin production are listed in table 1-4 (Schelin et al.,
2011).
Table 1-4: Factors affecting Staphylococcus aureus growth and enterotoxin formation (Schelin et al., 2011)
Factor Optimal
growth Growth limits
Optimal SE production
SE production limit
Enterotoxin (s) reported affected
Notes to effect (s) on enterotoxin production
Examples of analysis of the specific factor in food products
Temperature 35–41°C 6–48°C 34–40°C 10–46°C SEA, SEB, SEC, SED
Temperature seems to affect enterotoxin synthesis more that growth.
Milk Ham Egg products
pH 6–7 4–10 7–8 5–9.6 SEA, SEB, SEC, SED, SEE
Higher tolerance under aerobic compared with anaerobic growth conditions. Lactic acid particularly inhibits toxin formation. agr dependent regulation (SEC).
Ham Sausage
aw 0.99 0.83 – 0.99
0.99 0.86– 0.99 SEA, SEB, SEC, SEH
SEB and SEC may be more sensitive than SEA and SEH. SEH enterotoxin production at aw: 0.97 > 1 > 0.95.
Cured beef slurry Cured pork slurry Bacon Scrimp slurry Sausage
NaCl 0% 0-20% 0% <12% SEA, SEB, SEC
Raises temperature limit for SEA production. Low osmolality increases enterotoxin production. SEB production seems more strongly inhibited than growth.
Ham Sausage
Oxygen
Aerobic Anaerobic-aerobic
Aerobic Anaerobic-aerobic
SEA, SEB, SEC, SEH
Increases yield of SEB up to 10-fold. 10% dissolved oxygen is optimal for SEB production.
Ham Prawn Sausage
Redox potential (Eh)
>+200 mV
≥200 to > +200 mv
>+200 mV ≥100 to > +200 mv
- - -

Lactococcus lactis
- - - - sec, sel (sek, seg, seh) sea
Strongly reduces transcription of sec and sel and sightly sek, seg, she May favor the maintenance of sea in stationary phase.
Cheese
Chapter Two
Materials & Methods

2-1 Materials 2-1-1 Laboratory Equipments and Apparatus Apparatus and Equipments Used in This Study
Manufacturing Company/ Origin Equipment
Hirayama /Japan Autoclave Hettich /Germany Centrifuge Sanyo/Japan Deep-freezer Canon /Japan Digital camera Techne /UK Dry bath
Precisa /Switzland Electrical balance Sterellin Ltd /UK Eppendorf tubes Cleaver /UK Gel electrophoresis apparatus IKA /Germany Hot plate with magnetic stirrer Memmert /Germany Incubator Techne /UK Laminar air flow hood Olympus /Japan Light microscope Multigene (Gradient) /USA Master cycler gradient PCR Hettich /Germany Microcentrifuge Brand /Germany Micropipette Sigma / USA Microwave oven SherWood /USA Oven Jenwey /UK pH meter Consort E815 /Belgium Power supply Beko / UK Refrigerator BBL /USA Screw capped test tubes Gallenkamp /England Shaker Incubator Shimadzu /Japan Spectrophotometer Ultra Violet products institute /USA UV Transilluminator documentation
system Labcoo /Germany Vortex Memmert /Germany Water bath G.L.F. / Germany Water distillator
2-1-2 Chemicals and Biological Materials Chemicals and Biological Materials Used in This Study
Manufacturing Company/ Origin Chemical and biological material

BIONEER / Koria 100bp DNA Ladder Promega / USA 1kb DNA Ladder Merk / England Absolute ethanol (99%) Hi-Media/ India Agar-Agar Bio Basic INC/ Canada Agarose BDH / England Alpha-naphthol Bio-Merieux / France API – Staph system BDH / England Barium chloride (BaCl2) Oxoid / England Congo red BIONEER / Koria Deionized sterile distilled water BDH / England Dimethyl-alpha-naphthylamine Geneaid/ Thailand DNA Loading dye Geneaid/ Thailand DNA rehydration solution (TE) Bio Basic INC/ Canada Ethidium bromide dye BDH / England EDTA GCC / England Gelatin powder Geneaid/ Thailand Genomic DNA mini kit Fluka / Switzerland Glacial acetic acid BDH / England Glucose BDH / England Glycerol Syrbio /S.A.R. Gram stain reagents HiMedia / India HiStaph Latex Test Kit BDH / England Hydrochloric acid (HCl) BDH / England Hydrogen peroxide (H2O2) 30% BDH / England Meat extract BDH / England Isopropanol BDH / England Methyl green dye BDH / England N,N,N,N-tetramethyl-p-phenylene diamine
dihydrochloride
S.D.I / Iraq Normal Saline (Sterile) Hi-Media/ India Peptone water BIONEER / Koria PCR premix Fisons / UK Potassium hydroxide (KOH)
Manufacturing Company/ Origin Chemical and biological material BDH / England Potassium Tellurite
Hi-Media/ India Skim milk powder BDH / England Sodium chloride (NaCl) BDH / England Sodium citrate BDH / England Sodium hydroxide (NaOH) Redial-dehaeny / Germany Sulfanilic acid
BDH / England Sulfuric acid (H2SO4)

BDH / England Sucrose Bio Basic INC/ Canada Tris – Borate EDTA Buffer (TBE) BDH / England Triton X-100 BDH / England Trypton Fluka / Switzerland Urea Hi-Media/ India Yeast extract BDH / England Zinc powder
2-1-3 Antibiotic discs Antibiotic discs used in this study Antibiotic disc Concentration (µg) Company /Origin Methicillin (ME) 5 Hi-Media/India Tetracyclin (TE) 30 Hi-Media/India Vancomycin (VA) 30 Hi-Media/India 2-1-4 Culture Media 2-1-4-1 Ready to use culture media Table 2-1: Ready to use culture media Medium Company /Origin Baird-Parker agar Hi-media / India Blood agar base Hi-media / India Brain-Heart-Infusion broth Hi-media / India DNase agar Oxoid / England Mannitol Salt agar Hi-media / India Medium Company /Origin Muller-Hinton agar Hi-media / India Nitrate Broth Merk / Germany Nutrient broth Biolife / Italy Nutrient agar Hi-media / India Staph 110 agar Hi-media / India Tripticase Soy Broth Hi-media / India Urea agar base Hi-media / India
All media listed in table 2-1 were prepared according to the manufacturing company
instructions, pH was adjusted with 0.1N NaOH or 0.1N HCl, then sterilized with
autoclaving at 121oC/1 pound for 15min. The prepared media were then distributed into
sterile tubes or Petri dishes.
2-1-4-2 Laboratory Prepared Media 2-1-4-2-1 Baird-Parker Egg Yolk Tellurite Medium
(Baird-Parker, 1962)

Sixty three grams of Baird-Parker agar was suspended in 1 liter of distilled water
and autoclaved, then cooled to 50°C, afterward, 50ml of freshly prepared Egg Yolk
Emulsion (It was prepared by mixing 30 ml of egg yolk "the egg was carefully broken to
avoid contamination of the contents followed by separation the yolk from the white
{albumin} and the latter was discarded" with 70 ml of normal saline -under aseptic
conditions- with shaking vigorously) and three ml of potassium tellurite solution (which
was prepared by dissolving 3.5 gm of potassium tellurite in 100 ml of distilled water) were
added, after mixing well, the medium was poured into sterile Petri dishes.
2-1-4-2-2 Skim – Milk agar (Collee et al., 1996; Benson, 2001) The medium was prepared by dissolving 2.45 gm of nutrient agar in 87.5 ml of distilled
water, and autoclaved, cooled to 45-50°C and then 12.5 ml of previously sterilized (by
autoclaving at 121oC and 1 pound/square for 5 min) skim milk was added, thereafter the
medium poured in sterile Petri dishes.
This medium was used to detect the ability of bacteria to produce protease enzyme and
bacterial pigments.
2-1-4-2-3 Blood agar medium (Atlas, 2005) This medium was prepared according to the manufacturer’s instructions, and
autoclaved. The medium was cooled to 45-50oC, then 5%, as a final concentration, of
defibrinated horse blood was added. The medium was mixed gently and poured into sterile
Petri dishes .This medium is suitable for the isolation and culturing of bacteria and for the
detection of haemolytic activity and the type of haemolysis.
2-1-4-2-4 DNase medium
Thirty gram of DNase agar powder and 0.05 gm of methyl green dye were dissolved
completely in 1liter of distilled water. pH was adjusted to 7.4, then it was sterilized with
autoclave and poured in sterilized Petri dishes. This medium was used for detecting the
ability of bacteria to produce DNase enzyme.
2-1-4-2-5 Acetoin production medium (Collee et al., 1996)
The medium was composed of the following materials: Material Quantity Tryptone 10 gm Meat extract 3 gm Yeast extract 1 gm Glucose 20 gm

All these materials were dissolved in 900 ml of distilled water, pH was adjusted to
7.2, then the volume was completed to 1 liter with distilled water. Afterward, aliquots of
5ml were dispensed into tubes and autoclaved. The medium was used to detect the ability
of bacteria to ferment glucose with acetoin production.
2-1-4-2-6 Urea agar medium (Collee et al., 1996)
This medium consists of :
Material Quantity Urea base agar (Christensen’s medium)
95 ml
Urea solution 5 ml The Christensen’s medium was prepared according to the manufacturer’s instructions; pH
was adjusted to 6.8-6.9 and autoclaved. The medium was cooled to 45-50oC, then 5ml of
sterile filtrated 40% urea solution was aseptically added to each 95ml of Christensen’s
medium, and mixed well then dispended into sterile tubes (poured as a slant). This medium
was used to detect the ability of bacterial isolates to produce urease enzyme.
2-1-4-2-7 Nutrient Gelatin Agar (Harley and Prescott, 2002) This medium consists of:
Material Quantity Peptone 5 gm Meat extract 3 gm Gelatin 120 gm
All these materials were dissolved in 900ml of distilled water, pH was adjusted to 6.8,
thereafter the volume was completed to 1 liter with D.W., and autoclaved. This medium
was used to demonstrate the gelatin liquefaction by bacteria.
2-1-4-2-8 Semisolid medium (Collee et al., 1996)
This medium was prepared by dissolving (1.3gm of nutrient broth and 0.7gm of agar-agar)
in 100 ml of distilled water and dispensed aseptically in test tubes, autoclaved and left to
set in a vertical position. This medium was used to determine bacterial motility.
2-1-4-2-9 Congo red agar (Freeman et al., 1989)

This medium was prepared by dissolving 37gm of brain heart infusion broth with 50gm of
sucrose and 15gm of agar-agar in 900 ml of distilled water, autoclaved and then it was
cooled to 55-60oC, and mixed with congo red solution (which was prepared by dissolving
0.8gm of congo red dye in 100 ml of distilled water, and autoclaved), thereafter the
medium was poured in a sterile Petri dishes. This medium was used to determine bacterial
ability to produce the slime layer.
2-1-5 Stains, Reagents, Solutions and Emulsions used in the
identification of bacterial isolates
2-1-5-1 Gram Stain Kit (Benson, 2001)
It consists of:
· Crystal violet solution
· Lugol Iodine
· Alcohol Acetone solution
· Basic Fuchsine solution (This solution was diluted 1/5 before used).
2-1-5-2 Catalase reagent (Benson, 2001)
Hydrogen peroxide (H2O2) 3% was prepared for detecting catalase production.
2-1-5-3 Oxidase reagent (Vandepitte et al., 2003)
By dissolving 1gm of N,N,N,N-tetramethyl p- phenylene diamine hydrochloride in 100 ml
of distilled water (1% solution of N,N,N,N-tetramethyl p- phenylene diamine
hydrochloride) was prepared, stored in dark bottle and used immediately to detect the
ability of bacteria to produce oxidase enzyme.
2-1-5-4 McFarland standard solution (Benson, 2001)
McFarland standard solution is the turbidity standard solution, which is the most widely
used method for inocula preparation or standardization, especially the McFarland No.0.5
standard solution which was prepared by adding 0.5 ml of 1% solution of barium chloride
(BaCl2) to 9.5 ml of 1% solution of sulphuric acid (H2SO4) (these amounts must be added
to clean dry ampoules which should have the same diameters as the test tube to be used in
subsequent density determinations) to obtain barium sulfate solution with the specific
optical density to provide a turbidity comparable to that of bacterial suspension containing
1.5×108 CFU/ml.

2-1-5-5 Acetoin production test reagent (Barritt’s reagent) (Collee et al., 1996;
Benson, 2001)
This reagent was consisted of two solutions:
Solution (A): Prepared by dissolving 5 gm of alpha-naphthol in 100
ml of absolute ethanol.
Solution (B): Prepared by dissolving 40 gm of potassium hydroxide
(KOH) in 100 ml of distilled water.
2-1-5-6 Nitrite Test Reagents (Benson, 2001)
• Solution A: Prepared by dissolving 8 gm of sulfanilic acid in 1000 ml 5N acetic acid (1
part glacial acetic acid to 2.5 parts water).
• Solution B: Prepared by dissolving 5gm of dimethyl-alpha-naphthylamine in 1000 ml
5N acetic acid.
• Zinc powder. 2-1-6 Materials used in Agarose Gel Electrophoresis and in PCR amplification
2-1-6-1 Tris - Borate - EDTA (TBE) buffer
TBE buffer (10X) was supplied by (Bio Basic INC/Canada). Consisted of Tris base, boric
acid and EDTA. The pH was adjusted to 8.3; the volume of buffer was completed to one
liter with distilled water then 100 ml of TBE (10X) was added to 900 ml of D.W. to obtain
1X TBE solution used for agarose gel electrophoresis .
2-1-6-2 DNA Ladder (100 base pairs)
100 bp DNA ladder was specially obtained for determining the size of double stranded
DNA from 100 to 2000 bp provided as a liquid by the supplier (BIONEER), Lambda DNA
prepared in a concentration of 135 ng/µL in a storage buffer consists of: 10mM Tris-HCl
(pH 8.0), 1mM EDTA, 2.5% Ficoll, 0.005% Bromophenol Blue and 0.005% Xylene
Cyanol.
2-1-6-3 DNA Ladder (1000bp)
The 1 kb DNA ladder is ideal for determining the size of double stranded DNA from 250 to
10000 bp. Provided by the supplier (Promega) as a liquid prepared in a concentration of
100 µg/ml in a storage buffer consists of : 10mM Tris-HCl (pH 8.0) and 1 mM EDTA.

This ladder was provided with a Blue /Orange 6X loading dye which composed of: 10mM
Tris-HCl (pH 7.5), 50 mM EDTA, 15% Ficoll, 0.03% Bromophenol Blue, 0.03% Xylene
Cyanol and 0.4% Orange G.
The 1 kb DNA ladder had to be mixed with its loading dye as 5/1 immediately before
usage.
2-1-6-4 PCR Premix
It was provided by the supplier (BIONEER), the PCR premix consists of: 1U Top DNA
polymerase, 250 µM Each: dNTP (dATP, dCTP, dGTP, dTTP), 10 mM Tris-HCl (pH 9.0),
30 mM KCl, 1.5 mM MgCl2, stabilizer and tracking dye.
2-1-7 Standard strain The standard strain Staphylococcus aureus ATCC 25923 supplemented from the
teaching laboratories of Madienet Al-Teb.
2-2 Methods 2-2-1 Samples’Collection
Three hundred samples of milk and cheese were obtained from local markets. These
samples included raw, pasteurized, flavored, condensed, evaporated milk, raw salty,
processed, soft, hard and semi-hard cheese as shown in table 2-2.
Table 2-2: Milk and cheese samples
Sample types No. of samples
Raw milk 40
Pasteurized milk 40
Flavored milk 20
Evaporated milk 40
Condensed milk 10
Raw soft cheese 50
Raw salty cheese 40
Processed hard cheese 25
Processed semi-hard cheese 35
Total No. 300

The samples were collected randomly according to the instructions of the Iraqi Standard
Criterion No.2/2270 in Sampling, (2006); afferent by the Iraqi Central Organization for
Standardization and Quality Control (C.O.S.Q.C.).
2-2-2 Staphylococcal Isolation
The Iraqi Standard Criterion No.3/2270 in Isolation, Enumeration and Identification of
Microbiological Groups in Foods, (2006) was employed in staphylococcal isolation.
2-2-2-1 Staphylococcal Isolation from Milk Samples
One milliliter of each milk sample was placed (in triplets) in a sterilized Petri dish, then 15-
18 ml of the agar medium Staph No.110 (2-1-4-1) were poured (the medium should be
maintained melted at 44-46oC), the plates left to solidify at room temperature, and
thereafter they were incubated at 37oC for 24-48 h. Because of its constituent of 7.5%
NaCl; Staph No.110 is a selective medium so the isolates which can tolerate this salt
concentration strictly could be isolated. Then the grown colonies were further investigated.
2-2-2-2 Staphylococcal Isolation from Cheese Samples
Ten grams of each cheese sample was placed in 90 ml of peptone water, stomached for 1
min, three (ten serial) dilutions were made and 0.1ml of each dilution was inoculated on the
agar medium Staph No.110 (2-1-4-1), the plates left to solidify at room temperature, and
thereafter they were incubated at 37oC for 24-48 h. The grown colonies were further
investigated.
2-2-3 Staphylococcal Identification
Staphylococcal identification was performed according to: MacFaddin, (2000); Benson,
(2001); Harley and Prescott, (2002); Kayser, (2005); Atlas and Snyder, (2006); Brooks et
al., (2007), Vos et al., (2009) in addition to the Iraqi Standard Criterion No.3/2270 in
Isolation, Enumeration and Identification of Microbiological Groups in Foods, (2006).
The standard strain S. aureus ATCC 25923 (2-1-7) was used for confimation of the results
obtained by all the tests performed for the staphylococcal identification.
2-2-3-1 Gram Stain
For all bacterial isolates, smears were prepared from young culture suspensions, and
stained by Gram stain (2-1-5-1). Gram stainability, shape, size, and arrangement of cells
were determined under light microscope.
2-2-3-2 Growth on Mannitol Salt Agar

Mannitol salt agar (2-1-4-1) plates were streaked with a pure colony of tested bacteria, and
then incubated at 37oC for 24 h. This medium was used for selective isolation (contains
7.5% NaCl) and culturing of staphylococci and to differentiate medically important;
coagulase positive staphylococci which produce yellow colonies with yellow zone on
mannitol salt agar as a result of utilizing mannitol (as a positive result for mannitol
fermentation; yellow colonies); whereas other species of staphylococci produce small pink
or large deep yellow to deep orange colonies with no color change to medium (because of
its component: the monnitol sugar and the phenol red as an indicator).
2-2-3-3 Growth on Baird-Parker Egg Yolk Tellurite Medium
Baird-Parker agar (2-1-4-2-1) is a selective and differential medium used for the isolation
and identification of Staphylococcus species from foods. Its components gave it its
selectivity, these selective agents (glycine, lithium, and tellurite) have been carefully
balanced to suppress the growth of most bacteria present in foods, without inhibiting
Staphylococcus species, sodium pyruvate protects damaged cells and aid their recovery,
while egg yolk emulsion as a diagnostic agent.
Prior to use the surface of agar plate was dried with a sterile cotton swab, then 0.1ml of
food dilutions were spread with a glass spatula on the agar surface until it is dry, incubated
at 37oC for 24 h. The plates were examined for the typical colonies according to the table
2-3. The negative cultures were re-incubated for further 24h.
Table 2-3: Colony characteristics of typical organisms on Baird-Parker Egg Yolk
Tellurite Medium (O’Brien et al., 2009)
Organism Growth Colonial morphology
Staphylococcus aureus Good Dark grey-black shiny convex 1-1.5 mm diameter (18hrs) up to 3 mm (48hrs) narrow white entire margin surrounded by zone of clearing 2-5mm
Staphylococcus epidermidis Variable Not shiny black and seldom produces clearing
Staphylococcus saprophyticus Variable Irregular and may produce clearing. Wide opaque zones may be produced in 24 hrs
Staphylococcus chromogenes Good Not shiny orange-brown average size colonies, may produce clearing

Micrococcus species Variable Very small in shades of brown and black No clearing
Bacillus species Variable Dark brown matt with occasional clearing after 48hrs
Escherichia coli Variable Large brown-black
Proteus species Variable Brown-black with no clearing
Yeasts Variable White, no clearing
2-2-3-4 Endopigmentation on Milk agar (Staphyloxanthin) Skim milk agar (2-1-4-2-2) was inoculated with tested bacteria by streaking and incubated
at 37oC for 24 h. After incubation period, the colony and pigment of colonies were
observed.
2-2-3-5 Detection of Haemolysis on Blood agar Blood agar (2-1-4-2-3) was inoculated with an overnight bacterial growth culture and
incubated at 37oC for 24 h. A clear zone around the colonies was considered as beta
haemolysis behavior.
2-2-3-6 Catalase test One or two drops of catalase test reagent (3% H2O2) (which was prepared according to the
item 2-1-5-2) were placed on a slide; a growth from the center of a fresh pure colony from
nutrient agar plate was mixed with the reagent on the slide. The formation of bubbles
indicates a positive result.
2-2-3-7 Oxidase test A piece of filter paper placed in a clean Petri dish and 2-3 drops of freshly prepared oxidase
reagent (2-1-5-3) were added to the filter paper. A colony from tested organism was
transferred to the filter paper and rubbed on to the reagent with a sterile wooden stick. The
positive result was indicated by a violet or purple color appearance within (10-15) sec.
2-2-3-8 Clumping Factor and Coagulase test This test was used to detect the ability of isolates to produce clumping factor which is a
surface compound that is responsible for adherence of the microorganisms to fibrinogen
and fibrin when mixed with plasma, form clumps. The coagulase test is used to detect
coagulase, which is an enzyme-like protein that clots oxalated or citrated plasma, it binds to
prothrombin, together they become enzymatically active and initiate fibrin polymerization.
Clumping Factor:

An isolated pure colony was emulsified in a drop of saline on a microscopic slide with a
minimum of spreading. A similar suspensions of control positive and negative strains to
confirm the activity of plasma, were made, then one drop of undiluted human (or rabbit)
plasma was added to each suspension and mixed gently. The result was observed within 10
sec, the formation of visible coarse clumping, indicated the positive result, the negative
result or any slow-reacting strain was re-examined by a tube test.
Coagulase test (tube test):
Citrated human (or rabbit) plasma diluted 1:5 was mixed with an equal volume of
overnight bacterial broth culture or growth from colonies on agar, and incubated at 37oC.
The formation of clots in (1-4) h indicates a positive result. Negative results re-examined
for 24 h. A tube of plasma mixed with sterile broth is included as a control.
2-2-3-9 DNase production test
The fact that coagulase-positive bacteria are also able to hydrolyze DNA; makes the
DNase test a reliable means of confirming S. aureus identification. DNase agar (2-1-4-2-4)
was heavily streaked by the activated bacteria, and incubated at 35-37oC for 18-24 h.
Conversion of medium blue color to yellow color indicates the positive result.
2-2-3-10 Lipase Production Plates of Baird-Parker Egg Yolk Tellurite Medium agar (which was prepared as in the
item 2-1-4-2-1) were inoculated with the staphylococcal isolates (heavy streaking) and the
plates were incubated at 37oC for 24 h. Lipase production observed as opaque haloes
around the colonies and streaking line.
2-2-3-11 Protease activity Skim milk agar (2-1-4-2-2) was inoculated with a pure isolated colony, and incubated
at 37oC for 24 h. A positive result was observed as clear zone (for complete proteolysis) or
white precipitate (for partial proteolysis) around the colonies.
2-2-3-12 Urease Production This test shows the bacterial ability to produce urease enzyme. The bacterial isolate was
streaked on urea agar slant (2-1-4-2-6), and then incubated at 37oC for 18-24 h. The
positive result was evidenced by the change in the medium color from deep orange to pink.
2-2-3-13 Gelatin Liquefaction The nutrient gelatin deep (which was prepared according to the item 2-1-4-2-7) was
inoculated with the tested bacterial isolate by stabbing the medium 3/4 of the way down to

the bottom of the tube, and incubated at 37oC. The incubation time depends on the species
of bacteria, some organisms produce gelatinase at a very slow rate. Tubes that are negative
should be incubated for extra 4 or 5 days to see if gelatinase is produced slowly. After the
incubation period, the nutrient gelatin deep tubes were removed from incubator and placed
them in a refrigerator at 4oC for 30 min. or in an ice bath for 3-5 min until the bottom
resolidifies. The tubes were removed and tilt them from side to side to check if a portion of
the medium is still liquid which indicates gelatin liquefaction (positive result). If no
hydrolysis occurred; the medium will retain a gel.
2-2-3-14 Tolerance to different Concentrations of Salt The tested isolates were grown on the surface of nutrient agar containing 7.5%, 10%
and 15% concentrations of sodium chloride salt (NaCl). Appearance of growth indicates a
positive result, which means that the bacteria can tolerate the different concentrations of
salt.
2-2-3-15 Acetoin production test
An overnight grown (2-3) bacterial colonies were inoculated with a sterile loop into tubes
containing 5ml of acetoin production broth (2-1-4-2-5), then incubated at 30oC for 14 days,
after that 1ml of 40% potassium hydroxide reagent and 3ml of α-naphthol reagent (2-1-5-5)
were added to bacterial growth, then reagents were mixed and left at room temperature,
appearance of pink color after (1-2) h is an evidence of positive result.
2-2-3-16 Nitrate Reduction Test
Nitrate medium (2-1-4-1) was inoculated with a pure isolated colony and incubated at
37oC for 24 h, at the end of incubation, 1ml of both solution A and solution B (2-1-5-6)
were added to the broth culture, the development of a red color within 30 sec after adding
the test reagents indicates the presence of nitrate and represents a positive result for nitrate
reduction.
If no color developed after adding the test reagents, a small quantity of zinc powder was
added, the formation of a red color confirms a true negative result; the nitrate were not
reduced to nitrites by the bacteria. If a red color does not appear, the nitrate in the medium
were reduced past the nitrite stage to either ammonia or nitrogen gas.
2-2-3-17 Motility test

Semisolid medium (prepared according to the item 2-1-4-2-8) was inoculated by
stabbing down the center of the tube to about half of the depth of the medium and
incubated at 37oC for 24 h. Spreading turbidity from the stab line or turbidity throughout
the medium was considered as positive result.
2-2-3-18 Detection of the bacterial ability for the slime layer production
(Freeman et al., 1989)
The congo red agar method was used to detect the bacterial ability to produce the slim
layer; this was performed by culturing the bacterial colony on the congo red agar plate (2-
1-4-2-9), which then incubated at 37oC for 24h. Black colonies indicate highly production
of a slim layer; red colonies indicate moderately production of a slim layer, while light
colonies is a sign of negative result.
2-2-3-19 API-STAPH System: This system consists of strips containing dehydrated substrates in individual micro
tubes. These tests are reconstituted by adding to each micro tube an aliquot of API-STAPH
medium that has been inoculated with the strain of concern. The strip is then incubated for
8-24 h at 35-37oC, after incubation, metabolism produced color change that are either
spontaneous or revealed by the reagents addition.
Identification of the staphylococcal species was made by using indicators, and differential
charts supplied by the manufacturer. This test system is considered as a confirmatory test
for the identification of Staphylococcus spp. (Appendix 1).
2-2-4 HiStaph Latex Test (Rapid slide agglutination test) Before using the HiStaph latex reagent, it was allowed to equilibrate to room
temperature, then it was shaked well, and one drop of the latex reagent was added into a
circle of the test card, 2-3 average sized isolated colonies from a fresh overnight culture
plate was picked up by a clean mixing stick and emulsified in the latex drop, the card was
then rotated slowly and read within 1 minute. A positive result was indicated by visible
aggregation of the latex particles with a clear background.
In latex agglutination test (slide test), latex particles covered with fibrinogen and IgG were
used. IgG antibodies bind with coagulase on S.aureus, which results in latex particles
clumping in about 20 sec. This slide test may be negative in a low percent; in this case a

tube test, which detects both free and bound coagulase, has to be performed. About 97% of
human S. aureus isolates possess both forms of coagulase.
2-2-5 Antibiotic Susceptibility Test (Benson, 2001; Morello et al., 2006) The modified Kirby-Bauer method was used as the following: 2-2-5-1- Preparation of Mueller-Hinton Plates Mueller-Hinton agar was prepared according to the manufacturer’s instructions (2-1-4-1),
then the medium was cooled to 45-50oC and poured into the plates, allowed to set on a level
surface to a depth of approximately 4mm. When the agar was solidified, the plates were
stored at 4oC until use.
2-2-5-2 Inocula Preparation (Turbidity Standard)
To prepare the inocula, colonies from overnight culture of staphylococcal isolates were
transferred to 5 ml tube of normal saline to obtain culture with 1.5×108 CFU/ml by
adjusting to 0.5 McFarland standard.
2-2-5-3 Inoculation of the Test Plate
A- The plates were inoculated by dipping a sterile swab into the inocula; care must be
taken to express excess broth from the swab prior to inoculation, by pressing and rotating
the swab firmly against the side of the tube above the level of the fluid.
B- The swab was rubbed over the surface of the medium three times rotating the plate
through at an angle of 60 after each application. Finally the swab was passed around the
edge of agar surface.
C- The inocula were left for a few minutes to dry at room temperature with the lid is
closed.
D- By using a sterile forceps, antibiotic discs were placed on the inoculated plate.
Discs should be warmed to room temperature, and then dispensed on the agar surface; they
should gently pressed down with sterile forceps.
E- The plates were incubated within 30 min for 18-24 h at 37oC.
2-2-5-4 Reading the Results
After incubation, the diameters of the complete zone of inhibition were noted and measured
in millimeters. The diameter of inhibition zone for individual antimicrobial agent was

translated in terms of sensitive, intermediate and resistant categories by comparison with
the standard inhibition zone (Table 2-4).
Table 2-4: Diameter interpretive standards of inhibition zones according to NCCLs
(Benson, 2001; Morello et al., 2006)
ID Antimicrobial Agent
Disc Potency
Diameter of inhibition zone (mm)
Resistant Intermediate Sensitive
1 Methicillin 5 µg ≤ 9 10-13 ≥ 14
2 Tetracyclin 30 µg ≤ 14 15-18 ≥ 19
3 Vancomycin 30 µg ≤ 12 14-16 ≥ 17
2-2-6 Preservation of Bacterial Strains
2-2-6-1 Short Time Preservation (Harely and Prescott, 2002)
Single pure colony of bacterial isolate was streaked on the nutrient agar culture plate
and on the nutrient agar slants, incubated at 37oC for 24 h and stored in the refrigerator at
4oC, for one and three months for the plate and slants respectively.
2-2-6-2 Long Time Preservation (Vandepitte et al., 2003)
A- Stab culture method: An optimal (18-24 h) pure isolated colony was stabbed into
nutrient agar, then incubated at 37oC for 24 h and stored in dark place at room temperature.
B- Glycerol method : A nutrient broth was inoculated by a loopful of overnight pure
bacterial culture, and incubated at 37oC , after 18h glycerol was added to the inocula in a
final concentration of 15-30% and stored at (-20oC) for 12-18 months.
2-2-7 DNA Extraction
2-2-7-1 DNA Extraction from Bacterial Isolates
DNA extraction method described by De Baere et al., (2002) was modified in this study to
be suitable for the DNA extraction from staphylococcal isolates as described below:
De Baere et al., (2002) method was: bacterial colonies suspended in 500µL of TE buffer,
heated for 95oC for 30 min, then frozen (-20oC) for 30 min. While the modified method
was: five average sized pure isolated colonies from a fresh overnight culture plate (Brain
heart infusion agar) were picked up, placed in an eppendorf tube containing *200µL of

distilled water, the tube was vortexed, then *incubated in a dry bath at *85oC for *20 min,
then immediately frozen (-20oC) for *10 min. Thereafter it had been centrifuged at 10000
rpm for 5 min. Supernatant was placed in a new sterilized eppendorf tube, and examined
for quantity, purity and quality via the estimation of DNA concentration and purity (2-2-8),
and agarose gel electrophoresis (2-2-8). The standard strain S. aureus ATCC 25923 (2-1-7)
DNA was extracted by this method to be subjected to PCR for confimation of the results
obtained by the primers used for the staphylococcal identification.
In an attempt to optimize the DNA extraction from staphylococcal isolates, the classical
method using Genomic DNA mini kit, (Geneaid, Tiland) protocol for gram positive
bacteria was used with a modification: by depending the physical cell wall disruption
instead of enzymatic degradation and then the method was completed as the following: the
supernatant was subjected to the extraction protocol
1. GT buffer (200 µL) was added to the supernatant by vigorous shaking then
incubated at room temperature for 5min.
2. GB buffer (200 µL) was added by shaking vigorously for 5sec then incubated at
70˚C for 10min. during incubation the tubes were inverted every 3min.
3. Absolute ethanol (200 µL) was added and mixed by shaking vigorously, then
transferred to a mini column and centrifuged at 13000 rpm for 2min.
4. The collection tube was discarded and placed the GD column in a new one.
5. W1 buffer (400µL) was added then centrifuged at 13000 rpm for 30 sec and the
flow-through was discarded.
6. Washing buffer (600µL) were added, centrifuged at 13000 rpm for 30 sec and the
flow-through were discarded.
7. Washing buffer (400µL) were added, centrifuged at 13000 rpm for 30 sec and the
flow-through were discarded.
8. To dry the column matrix, GD column centrifuged at 13000 rpm for 3min.
9. Elution buffer (50µL), previously incubated at 70˚C for 10min, was added to GD
column then transferred to a clean microfuge tube and let stand for 3-5 min. then
centrifuged at 13000 rpm for 30 sec.
10. The DNA was stored in a deep freezer until PCR analysis is carried out.

2-2-7-2 DNA Extraction from Milk Samples Directly
Fusco et al. (2011) described a method for direct DNA extraction from milk sample; but
here in this study a modified simple method was depended as the following:
One milliliter of each milk sample mixed with one volume of (2%) sodium citrate, then the
lysate was centrifuged at *8000 rpm for *7min, pellets were washed with *distilled water
and resuspended in 100µL of *TE (Lysostaphin in a final concentration 1U/100 µL was
used instead of TE with incubation for 30 min at 37 oC in the initial method by Fusco et al.
and continued direct DNA extraction using Wizard Genomic DNA purification kit),
vortexed, then they were incubated in a dry bath at 90oC for 20min, the heated samples
were immediately frozen (-20oC) for 10min. Thereafter they were centrifuged at 10000 rpm
for 5min, pellets resuspended in 100µL of TE, and examined for quantity, purity and
quality.
For further purification, an addition of isopropanol and ethanol is required as the following:
One hundred microliter (an equal volume DNA extract) of isopropanol was added to the
extracted DNA, and mixed gently by inversion until white thread-like strand of DNA form
a visible mass, then centrifuged at 13000 rpm for 2min. The supernatant was discarded and
pellets washed with 100 µL of (70%) ethanol, with rotation of the tubes, then they were
centrifuged at 13000 rpm for 3 min. Ethanol was aspirated carefully then tubes were
inverted on clean gauze. The pellets were air-dried for 10-15 min.
The DNA pellets were rehydrated by adding 100 µL of DNA rehydration solution (TE) and
incubated overnight at 4oC then kept in the deep freezer. DNA must be thawed before
subjecting to PCR.
DNA extraction from milk samples directly impedes the problems of costing, time
consuming, routine work using genomic DNA extraction kits which need the addition of
cost effective lysostaphin with lysozyme.
2-2-7-3 DNA Extraction from Cheese Samples Directly
The method by Baruzzi et al.(2005) was modified to extract DNA directly from cheese
samples. The modified method used in this research included:
One gram of each cheese sample washed with one volume of (2%) sodium citrate,
homogenized for 2 min by vortexing, then the homogenate was centrifuged at * 10000 rpm
for 6 min, pellets were washed twice with distilled water, then pellet dissolved in 500 µL of

1:39 Triton X-100 solution with shaking vigorously, and heating at 70oC for 10 min., then
the lysate was centrifuged at *10000 rpm for *5min, pellets were rinsed with distilled water
and (initial method continued direct DNA extraction using Wizard Genomic DNA
purification kit) while the modified method described in this study continued as:
*resuspended in 100 µL of distilled water then they were incubated in a dry bath at 90oC
for 20 min, the heated samples were frozen immediately (-20oC) for 10 min. Thereafter
they were centrifuged at 10000 rpm for 5 min, pellets resuspended in 100 µL of TE, and
examined for quantity, purity and quality.
For further purification, an addition of isopropanol and ethanol is required as the following:
One hundred microliter (an equal volume DNA extract) of isopropanol was added, and
mixed gently by inversion, thus genomic DNA precipitated out forming clumps of
intertwined threads, then centrifuged at 13000 rpm for 2min. The supernatant was
discarded and 100 µL of 70% ethanol was added, gently, inverted several times to wash the
DNA pellets.
Pellets washed with 100 µL of (70%) ethanol, by rotating the tubes, they were centrifuged
at 13000 rpm for 3 min. Ethanol was aspirated carefully then tubes were inverted on clean
gauze. The pellets were air-dried for 10-15 min.
The DNA pellets were rehydrated by adding 100 µL of DNA rehydration solution (TE) and
incubated overnight at 4oC then kept in the deep freezer. DNA must be thawed before
subjecting to PCR.
2-2-8 Estimation of DNA Concentration and Purity (Stephenson, 2003)
The DNA concentration was determined by using spectrophotometer; 5 µL of each
DNA sample were added to 995 µL of distilled water and mixed well. Spectrophotometer
was used for measuring the optical density (O.D.) at wave length of 260 nm and 280 nm.
An O.D of one corresponds to approximately 50µg/ml for double stranded DNA. The
concentration of DNA was calculated according to the formula:
DNA concentration (µg/ml) = O.D 260nm × 50 × dilution factor
Spectrophotometer was used also to estimate the purity ratio of DNA according to the
following formula:

DNA purity = O.D 260nm / O.D 280nm
The ratio used for detecting DNA contamination with protein preparation. DNA
quality could be assessed by 0.8% agarose gel electrophoresis (Maniatis et al., 1982).
2-2-9 Agarose Gel Preparation and Electrophoresis Agarose gel was prepared in 0.8 % concentration for quality of the extracted DNA, by
dissolving 0.8 gm of agarose powder in 100 ml of 1X TBE buffer, and melted, then the
agarose gel was cooled to 50-60oC, 5 µL of ethidium bromide dye was added with mixing,
agarose was poured out into the gel jar to prevent bubble formation, then cooled to 20oC.
When agarose gel was poured, several wells were carefully made with a comb at one side
of the gel about 5-10 mm away from the end of gel; after final solidification. The comb was
carefully removed; the jar was put in the electrophoresis tank.
Six microliters of the 1 kb DNA ladder were placed in the first left well of the agarose
electrophoresis gel. Before loading to the gel wells; DNA samples were first mixed with a
loading dye, so that 7 µL of each DNA sample were mixed with 3 µL loading dye, then this
10 µL of loaded DNA were carefully transferred to a well of the agarose electrophoresis
gel, then the electrophoresis tank closed with its special lid, and electric current was
matched (70 volt for 1 h).
2-2-10 Polymerase Chain Reaction (PCR) Technique
The polymerase chain reaction (PCR) is an in vitro amplification of target DNA with a pair
of primers and a DNA polymerase, resulting in several million fold amplification of the
target sequence within few hours (Dale and Park, 2003).
PCR assay was performed in a monoplex and multiplex patterns in order to amplify
different fragments of genes under study in a single tube for detecting Staphylococcus
aureus (femA) and virulence associated genes (virulence factors: coagulase, enterotoxins,
methicillin resistance).
2-2-10-1 Genes selection
Seven genes were selected to be amplified either separately in monoplex (coa, femA
and mecA) or in multiplex PCR technique (sea, seb, sec and sed).
PCR was performed to amplify coa gene that encodes coagulase enzyme which
divides Staphylococcus genus to coagulase positive and coagulase negative staphylococci,

femA gene as a specific genomic marker for S. aureus and it is factor essential for
methicillin resistance, while mecA gene was amplified because it encodes methicillin
resistance.
Multiplex PCR was used to amplify four genes including:
sea, seb, sec and sed were selected because they are the genes that encodes the four
classically described staphylococcal enterotoxins SEA, SEB, SEC, and SED respectively.
2-2-10-2 Primers selection
The primers listed in table 2-5 were selected for this study; these primers were
provided in a lyophilized form, dissolved in sterile distilled water to give a final
concentration of 100 pmol/ µL as recommended by provider and stored in deep freezer
until used in PCR amplification.
Table 2-5: The primers and their sequences used in conventional PCR for detection of Staphylococcus aureus
Gene Primer
name Sequence
5´ 3´ Len--gth
Predica--tive PCR product
References
fem A Fem A F CATGATGGCGAGATTACAGGT 21 318 bp Manikandan et al. (2011) fem A Fem A R GTCATCACGATCAGCGAAAGC 21
mec A Mec A F AAAATCGATGGTAAAGGTTGGC 19 533 bp Merlino et al. (2002) mec A Mec A R AGTTCTGCAGTACCGGATTTGC 22
coa Coa F CGAGACCAAGATTCAACAAG 20 730 bp Aslantas et al. (2007) coa Coa R AAAGAAAACCACTCACATCA 20
sea Sea F CCTTTGGAAACGGTTAAAACG 21 127 bp El-Seedy et al. (2010) sea Sea R TCTGAACCTTCCCATCAAAAC 21
seb Seb F TCGCATCAAACTGACAAACG 20 477 bp El-Seedy et al. (2010) seb Seb R GCAGGTACTCTATATAGTGCC 21
sec Sec F AGATGAAGTAGTTGATGTGTATGG
24 451 bp El-Seedy et al. (2010)
sec Sec R CACACTTTTAGAATCAACCG 20 sed Sed F CCAATAATAGGAGAAAATAAAAG 23 278 bp El-Seedy et al.
(2010) sed Sed R ATTGGTATTTTTTTTCGTTC 20
2-2-10-3 PCR Amplification
The extracted DNA, primers and PCR premix (Accupower, Bioneer), were thawed at
4oC, vortex and centrifuged briefly to bring the contents to the bottom of the tubes. PCR
mixture was set up in a total volume of 25 µL included 5µL of PCR premix, 2µL of each

primer and 4µL of template DNA have been used. The rest volume was completed with
sterile de-ionized distilled water, then vortexed and finally 4µL of template DNA was
added. Negative control contained all material except template DNA, so instead that
distilled water was added.
PCR reaction tubes were centrifuged briefly to mix and bring the contents to the bottom of
the tubes, and placed into thermocycler PCR instrument where DNA was amplified as
indicating in the tables 2-6, 2-7, and 2-8.
Table 2-6: Program used to amplify the femA and mecA genes
Stage Temperature (time)
Initial denaturation 95˚C (5min) Denaturation 95˚C (1min)
30 cycles Annealing 63˚C (1min) Extension 72˚C (1min) Final extension 72˚C (5min)
Table 2-7: Program used to amplify the coa gene
Stage Temperature (time)
Initial denaturation 95˚C (5min) Denaturation 95˚C (1min)
40 cycles Annealing 55˚C (1min) Extension 72˚C (1min) Final extension 72˚C (10min)
Table 2-8: Program used to amplify enterotoxins’ genes sea, seb, sec and sed
Stage Temperature (time)
Initial denaturation 95˚C (5min)
Denaturation 95˚C (1min)
40 cycles Annealing 52˚C (1min)
Extension 72˚C (1min)
Final extension 72˚C (10min)
2-2-11 Determination of PCR Specificity

The extracted DNA from staphylococcal isolates (which were isolated from milk and
cheese samples), and the directly extracted DNA from milk and cheese samples, were
checked for their concentration and purity, and thereafter they were analyzed by PCR and
the results confirmed by using 1.5% agarose gel electrophoresis. Agarose gel was prepared
in 1.5 % concentration for the determination of the PCR specificity by dissolving 1.5 gm of
agarose powder in 100 ml of 1X TBE buffer, and boiling, then the agarose was cooled to
50-60oC, 5 µL of bromide dye was added with mixing, agarose was poured out into the gel
jar to prevent bubble formation, then cooled to 20oC. When agarose gel was poured, several
wells were carefully made with a comb at one side of the gel about 5-10 mm away from the
end of gel; after final solidification, the comb was carefully removed; the jar was put in the
electrophoresis tank. Six microliters of the 100 bp DNA ladder were placed in the first left
well or the middle one of the agarose electrophoresis gel, then 10 µL of each PCR product
were put in the rest wells of the agarose gel. Thereafter the electrophoresis tank closed with
its special lid, and electric current was matched (70 volt for 3 h).
2-2-12 Detection of Thermostable Enterotoxins by Suckling Mouse Bioassay
(Guarino et al., 1989; Kisiel et al., 2011)
Suckling Mouse Bioassay was used in order to detect the ability of staphylococcal
isolates to secrete extracellular heat stable enterotoxins in their broth culture supernates, by
testing the enteotoxins’ biological activity;
1 mouse unit of SE activity was defined arbitrarily as the amount of toxin producing a ratio
of intestinal weight to carcass weight of 0.083
This bioassay was illustrated by:
Newborn Swiss albino suckling mice (2-4 days old) were separated from their mothers
immediately before use. A 0.1ml of broth culture supernatant had been administered orally
to each mouse, and then the mouse was left at 25°C. Two hours after administration, the
mice were killed by decapitation, the abdomen was opened, and the entire intestines (not
including the stomach), was removed with forceps. The intestines from each mouse were
pooled and weighed, and the ratio of gut weight to remaining carcass weight was
calculated.
v Preparation of crude culture filtrates

Staphylococcal isolates were inoculated in a Trypticase Soy Broth (TSB) containing 0.6% yeast extract for 48 h at 37oC in a shaker incubator. Then the culture was centrifuged for 10 min in an eppendorf centrifuge with a speed of 3000rpm, the obtained supernatant was transported in the amount ranging from 1 to 1.5 ml to a test tube, and prepared to be administered orally to the suckling mice directly or after heated at different temperatures (65 and 100oC) for various periods of time (15, 30 min). 2.2.13 Statistical Analysis
Experimental data were presented in terms of observed numbers and percentage
frequencies, SPSS (Statistical Package for Social Sience) program version 20 was used
and some of these data analysed by SAS program (2004)-LSD (Least significant
difference) was used to the significant compare difference. Regarding other data were
analyzed by the followings:
1. Chi-square test: P values ≤ 0.05 or ≤ 0.01 were considered statistically significant.
2. Paired samples T-test: P value ≤ 0.05 was considered statistically significant.
3. Pearson correlation.

Chapter Three
Results & Discussion

3- Results and Discussion 3-1 Staphylococcal Isolation and Identification
The collected milk and cheese samples were cultured on the agar medium Staph No.110;
because of its constituent of sodium chloride (7.5%) Staph No.110 is a selective medium,
so that only staphylococci and any bacteria which can tolerate this salt concentration will
grow.
Out of 234 bacterial isolates grown on Staph No.110 (Figure 3-1) were more purified by
ABC streaking method, and then all of them were examined microscopically for gram
stainability, shape and cluster arrangement.
Figure 3-1: The percentage of Staphylococcus spp. isolated from milk and cheese samples
Two hundred colonies were identified morphologically as gram positive cocci arranged in
grape-like irregular clusters this clusters occur because the bacterial cells divide in three
planes in an irregular pattern producing branches of cocci which are characteristics to
Staphylococcus spp. (Brooks et al., 2007; Dubey and Maheshwari, 2009).
Table 3-1 illustrates the results of the biochemical tests applied to identify the
staphylococcal isolates.
0102030405060708090
100
Staphylococci Other bacteria
15%
85%
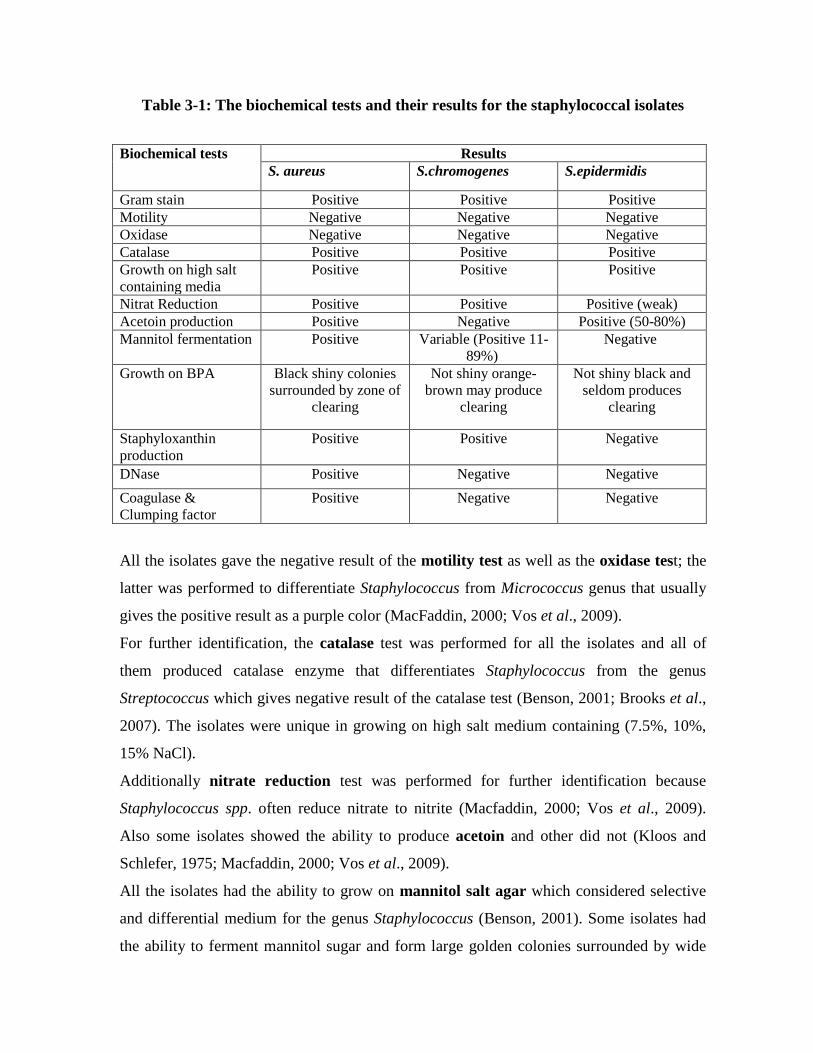
Table 3-1: The biochemical tests and their results for the staphylococcal isolates
Biochemical tests Results S. aureus S.chromogenes S.epidermidis
Gram stain Positive Positive Positive Motility Negative Negative Negative Oxidase Negative Negative Negative Catalase Positive Positive Positive Growth on high salt containing media
Positive Positive Positive
Nitrat Reduction Positive Positive Positive (weak) Acetoin production Positive Negative Positive (50-80%) Mannitol fermentation Positive Variable (Positive 11-
89%) Negative
Growth on BPA Black shiny colonies surrounded by zone of
clearing
Not shiny orange-brown may produce
clearing
Not shiny black and seldom produces
clearing
Staphyloxanthin production
Positive Positive Negative
DNase Positive Negative Negative Coagulase & Clumping factor
Positive Negative Negative
All the isolates gave the negative result of the motility test as well as the oxidase test; the
latter was performed to differentiate Staphylococcus from Micrococcus genus that usually
gives the positive result as a purple color (MacFaddin, 2000; Vos et al., 2009).
For further identification, the catalase test was performed for all the isolates and all of
them produced catalase enzyme that differentiates Staphylococcus from the genus
Streptococcus which gives negative result of the catalase test (Benson, 2001; Brooks et al.,
2007). The isolates were unique in growing on high salt medium containing (7.5%, 10%,
15% NaCl).
Additionally nitrate reduction test was performed for further identification because
Staphylococcus spp. often reduce nitrate to nitrite (Macfaddin, 2000; Vos et al., 2009).
Also some isolates showed the ability to produce acetoin and other did not (Kloos and
Schlefer, 1975; Macfaddin, 2000; Vos et al., 2009).
All the isolates had the ability to grow on mannitol salt agar which considered selective
and differential medium for the genus Staphylococcus (Benson, 2001). Some isolates had
the ability to ferment mannitol sugar and form large golden colonies surrounded by wide

yellow zones and turned the color of the medium from pink to yellow, others mannitol non
fermentor which appeared as small pink colonies as S.epidermidis or large deep yellow to
deep orange colonies as S. chromogenes and in both cases no color change was observed in
the medium (Brooks et al., 2007) (Figure 3-2).
Figure 3-2: Mannitol salt agar cultured with Staphylococcus spp. mannitol non fermentor colonies of S.epidermidis (to the left) and mannitol fermentor colonies of S.aureus (to the right)
All the isolates grown on Baird-Parker Egg Yolk Tellurite Medium which is selective
and differential medium used for the isolation and identification of Staphylococcus species
from foods, three types of colonies appeared: dark grey-black shiny convex 1-1.5 mm
diameter (18hrs) up to 3 mm (48hrs) narrow white entire margin surrounded by zone of
clearing 2-5mm colonies identified as S. aureus (Figure 3-3), black not shiny colonies and
seldom produces clearing as S. epidermidis and orange-brown not shiny average size
colonies, may produce clearing as S. chromogenes (O’Brien et al., 2009).

Figure 3-3: Baird-Parker Egg Yolk Tellurite agar cultured with S.aureus which appeared as black shiny convex colonies with lipase activity (shown as hydrolyzed clear zone surrounding the colonies)
On milk agar plates a number of pigmented colonies appeared from white to deep orange,
S.epidermidis appeared as white colonies, golden-yellow colonies expected to be S.aureus,
while deep yellow and deep orange colonies expected to be S. chromogenes (Figure 3-4).
Among the best-recognized bacterial pigments are the carotenoids that impart the
eponymous golden color to the major human pathogen, S. aureus. This organism produces
multiple carotenoid pigments via a well described biosynthetic pathway that culminates
with golden staphyloxanthin as the major product and yellow 4040-diaponeurosporene as a
minor product. Staphyloxanthin also acts as a virulence factor. It has an antioxidant action
that helps the microbe evade death by reactive oxygen species produced by the host
immune system.

Figure 3-4: Skim milk agar cultured with S.aureus which appeared as glistening orange convex colonies (due to Staphyloxanthin production) with protease activity (shown as hydrolyzed clear zone surrounding the colonies)
DNase production was detected by culturing the isolates on DNase agar, the appearance of
yellow zone surrounding the colonies considered as a positive result. The results showed
that 97 S. aureus isolates (100%) were DNase producers. Most of pathogenic isolates of
S.aureus produce DNase enzyme. DNase degrades the host DNA and that increases the
invasiveness and pathogenecity of staphylococci that possess it (Brooks et al., 2007).
After isolates identification at the generic level, the coagulase test was performed to
identify the bacterial isolates at the species level, ninety seven isolates (48.5%) showed the
ability to produce coagulase enzyme (coagulase positive) and 103 isolates (51.5%) were
coagulase negative as shown in figure 3-5, this result belong to the fact that coagulase
enzyme is the most important identification agent that recognizes coagulase positive from
coagulase negative staphylococci. Coagulase is an important virulence factor; it’s acting by
deposit fibrin on the surface of Staphylococci, thereby altering their destruction inside the
phagocytes.
Clumping factor is a surface S. aureus compound that is responsible for adherence of the
organisms to fibrin and fibrinogen (Brooks et al., 2007).

Figure 3-5: The percentage of each Coagulase-Positive and Cogulase-Negative among isolated Staphylococcus spp.
API Staph system was used to confirm the identification of the most prevalent
Staphylococcus spp. isolated from milk and cheese samples S.aureus which previously
identified by conventional biochemical tests. The test was applied only for five isolates,
and the results obtained from API Staph system were in agreement with those obtained
from biochemical identifications. (Appendix 2 and 3).
HiStaph Latex Test is a rapid slide agglutination test was performed to all the coagulase
positive Staphylococci which were previously identified by biochemical tests, 90 (92.8%)
COPS isolates gave the positive result of the test which was indicated by visible
aggregation of the latex particles with a clear background as shown in figure 3-6.
0102030405060708090
100
COPS CONS
51.50% 48.50%

Figure 3-6: The results of HiStaph Latex Test (2 S.aureus; 4 S.chromogenes; 6 Control positive)
The results showed that 97 (48.5%) of the staphylococcal isolates isolated from milk and
cheese samples were S.aureus, followed by 82 (41%) S.chromogenes and 21 (10.5%)
S.epidermidis (Figure 3-7).
Figure 3-7: The prevalence of Staphylococcus aureus according to all the Staphylococcal isolates isolated from milk and cheese samples
Staphylococci are ubiquitous bacteria found on the skin and mucous membranes of humans
and worm-blooded animals; they also can be recovered from different environmental
48.5 41
10.5
0
10
20
30
40
50
60
%
S.aureus S.chromogenes S.epidermidis

sources such as soil and water and from wide range of foodstuffs (Irlinger, 2008). S. aureus
is the causative agent of mastitis in lactating animals (Bartolomeoli et al., 2009). Other
possible sources that contribute to high levels of S. aureus in milk are the improper hygiene
and poor farm management (Ateba et al., 2010).
Milk is a good substrate for S. aureus growth, and dairy products are common sources of
intoxication (Morandi et al., 2007). S. aureus can gain access to milk either by direct
excretion from udders with clinical or subclinical staphylococcal mastitis or by
contamination from the environment during handling and processing raw milk (Peles et al.,
2007).
Cheese is a complex and dynamic microbial ecosystem characterized by the presence of a
large variety of bacteria, yeasts, and molds, some microorganisms including species of
lactobacilli or lactococci, are known to contribute to the organoleptic quality of cheese,
whereas the presence of other microorganisms may lead to spoilage or constitute a health
risk, S. aureus is recognized worldwide as an important food-borne pathogen, owing to the
production of enterotoxins in food matrices (Duquenne et al., 2010).
Fooladi et al. (2010) found that 32% of the dairy products were contaminated by S. aureus.
Contamination of food products with S. aureus pathogens may result from their presence in
the basic raw material - milk, this is of great importance especially in countries with large
production of dairy products such as cheeses (Holeckova et al., 2002).
Algero et al. (2007), who analyzed 172 food samples including milk, soft cheese, hard
cheese, ice cream, yoghurt, and fast food like sandwiches in a market in Brazil, reported
that 26 samples (15.1%) of the food were contaminated with S. aureus.
S. aureus was found in 70.4% in raw and pasteurized bovine milk samples as demonstrated
by Rall et al.(2008).
Dastmalchi Saee et al. (2009) evaluated 370 cow milk samples, using culture and PCR
methods. They reported that 58 samples (15.67%) were contaminated with coagulase-
positive S. aureus.
The presence of this microorganism (S. aureus) after pasteurization can be attributed to
inefficacy of the thermal process (Rall et al., 2008).
The CONS species most commonly isolated from mastitic milk were S.chromogenes and S.
simulans. S. chromogenes was the major CONS species in subclinical mastitis, the majority

of CONS isolates in milk samples in a research in USA, were S. chromogenes (Matthews et
al., 1991).
The predominant CONS species of 298 isolates in milk samples from subclinical mastitis in
Germany were S. chromogenes (33%) and S. simulans (23%) (Luthje and Schwarz, 2006).
In addition to the predominant species, various other CONS species are less frequently
isolated from milk samples. S. hyicus, S. epidermidis and S. haemolyticus were detected in
some samples (Devriese and De Keyser, 1980; Jarp, 1991; Aarestrup et al., 1995; Chaffer
et al., 1999; Luthje and Schwarz, 2006), but S. warneri, S. sciuri and S. xylosus, and
several other CONS species usually only occur seldom (Rather et al., 1986; Aarestrup et
al., 1995; Waage et al., 1999; Luthje and Schwarz, 2006).
A wide variety of CONS species have been isolated from cows’ skin, the predominant
CONS species in cows’ beddings and environment were reported to be S. xylosus, S. sciuri,
and S. saprophyticus (Matos et al., 1991). The same species are frequently isolated from
cows’ haircoat. The predominant species in milk and extramammary samples are mainly
different, but S.chromogenes is an exception. It seems to be adapted both to conditions on
the skin and in the udder. The only CONS species isolated both from mastitis and from
milkers’ hands were S.chromogenes and S.succinus subsp. succinus. The isolates from the
hands were, however, of different pulsotypes than the mastitis isolates. Although we cannot
exclude the possibility of elimination and re-infection with the same species, we assume
that the CONS infections remained persistent in the udder. S. simulans is a more specific
mastitis pathogen and adapted to udder conditions. In contrast, S. chromogenes is able to
live on bovine skin, but can also infect the mammary gland and cause mastitis.
In the study of Soares et al.(2011) S.epidermidis species occupied 14.3% of the
staphylococcal spp. isolated from dairy products in Portugal.
S.epidermidis species was detected in food samples in a percentage of 40% in the research
of Cunha et al. (2006), this species is a common inhabitant is a common inhabitant of
human skin and mucous membranes of individuals manipulating food and animals, and is
able to contaminate raw products and processed foods (Bergdoll, 1995; Cunha et al., 2006).
Fusco et al.(2011) found that S. aureus as 54% in raw milk samples, while Jorgensen et
al.(2005) isolated S. aureus in 75% from bovine milk samples.

The numerous examples of S. aurues causing bacteremia were reported in human with
predisposing conditions of dairy farms (Normanno et al., 2007; Sharma et al., 2011). The
presence of S. aurues in the milk sample is a new and appealing as well as an important
finding of this study.
Regarding samples’ type and the isolated staphylococcal species, there was a relationship
between them as demonstrated in tables 3-2 and 3-3.
Table 3-2: Isolated staphylococcal species from each milk and cheese samples
Milk and Cheese Samples No. of the staphylococcal species in each sample S. aureus S. chromogenes S. epidermidis
Raw milk 11 29 6 Pasteurized milk 5 24 2 Flavored milk 2 1 1 Evaporated milk 3 0 0 Condensed milk 2 3 0 Raw soft cheese 32 13 8 Raw salty cheese 27 9 3 Processed hard cheese 3 1 0 Processed semi-hard cheese
12 2 1
Table 3-3: Relationship between the samples’ type and the isolated staphylococcal
species
Samples’ type Total No. of the isolated species of staphylococci
COPS CONS
S. aureus S. chromogenes S. epidermidis
Milk 23 57 9
Cheese 74 25 12
X2-Test value 39.62* 17.18* 0.4608**
*: significant (P<0.05), **: non significant (P>0.05).

Statically there were significant differences between milk and cheese samples in the
number of staphylococcal species isolated from them. From milk samples S. chromogenes
was the most prevalent isolated staphylococcal species; while cheese samples contaminated
mainly by S. aureus.
Inadequate pasteurization of initially contaminated milk samples from animal skin or hair
contributed in such high number of S. chromogenes in these samples, while food handling
during cheese processing may be the source of S. aureus in these samples.
3-2 Detection of Virulence Factors of Staphylococci
The production of haemolysin, coagulase and clumping factor, DNase, lipase, protease,
urease and gelatinase enzymes and slime layer production were detected in this study.
Haemolysin: Haemolysin production was detected by culturing the isolates on blood agar;
positive results appeared as a haemolytic zone around the colonies. Seventy eight (80%) S.
aureus isolates showed haemolytic activity and the lysis was β type.
Protease production was detected on skim milk agar plates, casein is the predominant
protein in milk, and its presence causes milk to have its characteristic white appearance,
protease which hydrolyzes casein to produce more soluble, transparent derivatives. The
hydrolyzed clear zone around the colonies considered positive result, the results obtained
by this study showed that most of the isolates were protease producers.
Lipase production was detected by culturing isolates on Baird Parker Egg Yolk Tellurite
medium, the hydrolyzed clear zone surrounding the colonies considered as positive result
(Figure 3-3).
All the isolates were lipase producers, these enzymes acting by helping bacteria to invade
host tissues throughout hydrolyzing cells membranes and spoiling foods such as cheese and
milk.
Urease production was done on urea agar slants, changing the color of phenol red indicator
from yellow to pink represents the positive result. Results showed that all S.aureus isolates
were urease producers.
Gelatinase production test was done by stabbing the isolates in gelatinase medium,
liquefaction of the medium is the positive result, results showed that all S.aureus isolates
were gelatinase producers.

These results evidence on the ability of the staphylococcal isolates to produce a variety of
enzymes that helps in invading the host’s tissues as well as the foods texture (proteases,
lipases and gelatinases).
Slime layer production detection for both COPS and CONS isolates, was investigated by
using Congo Red Agar method revealed slime production in 74.22% of COPS (Figure 3-
8), while CONS isolates did not produce it (Figure 3-9).
Figure 3-8: Slime layer producing S. aureus colonies on the congo red agar
Figure 3-9: S. chromogenes colonies on the congo red agar (non slime layer producers)
All these results of the detection of some virulence factors produced by Staphylococcal
isolates are illustrated in table 3-4.
Table 3-4: Virulence factors produced by Staphylococcal isolates Virulence factor S. aureus S. chromogenes S. epidermidis
Protease Positive (100%) Positive (95%) Positive (78%)
Urease Positive (100%) Variable Positive (78%)
Lipase Positive (100%) Positive (68%) Positive (100%)
Gelatinase Positive (100%) Positive (62%) Negative
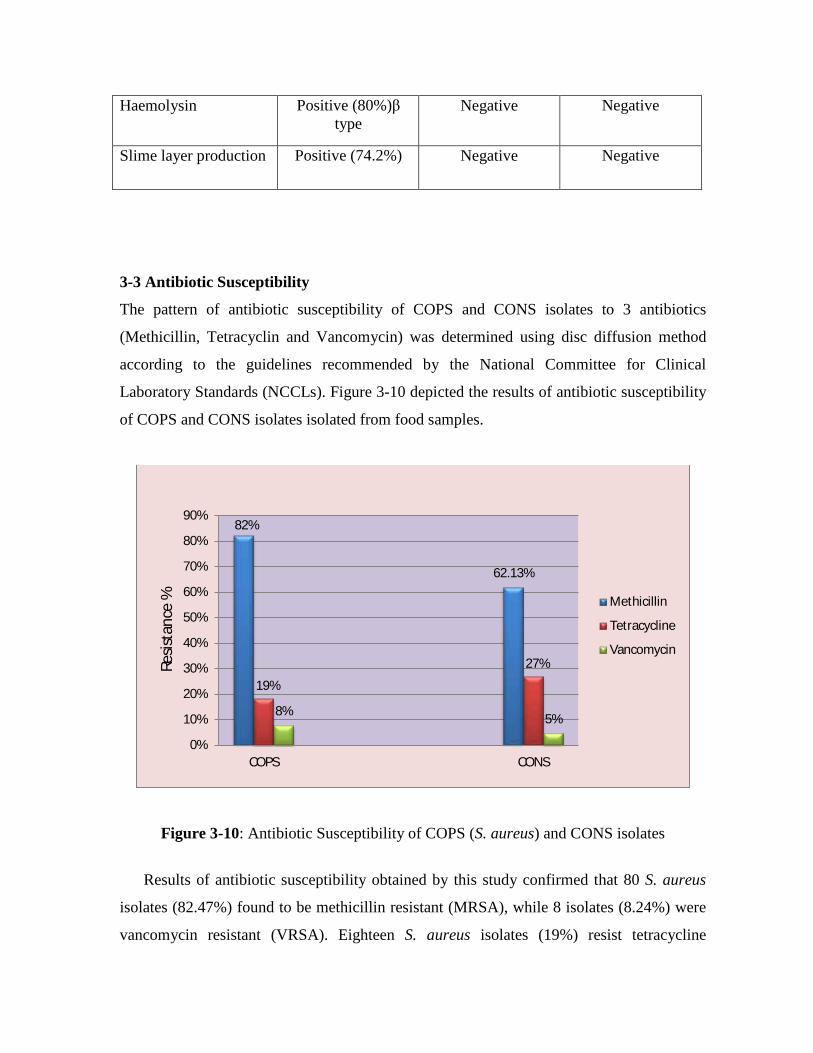
Haemolysin Positive (80%)β type
Negative
Negative
Slime layer production Positive (74.2%) Negative
Negative
3-3 Antibiotic Susceptibility
The pattern of antibiotic susceptibility of COPS and CONS isolates to 3 antibiotics
(Methicillin, Tetracyclin and Vancomycin) was determined using disc diffusion method
according to the guidelines recommended by the National Committee for Clinical
Laboratory Standards (NCCLs). Figure 3-10 depicted the results of antibiotic susceptibility
of COPS and CONS isolates isolated from food samples.
Figure 3-10: Antibiotic Susceptibility of COPS (S. aureus) and CONS isolates
Results of antibiotic susceptibility obtained by this study confirmed that 80 S. aureus
isolates (82.47%) found to be methicillin resistant (MRSA), while 8 isolates (8.24%) were
vancomycin resistant (VRSA). Eighteen S. aureus isolates (19%) resist tetracycline
82%
62.13%
19%
27%
8% 5%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
COPS CONS
Resis
tanc
e %
Methicillin
Tetracycline
Vancomycin

antibiotic. Resistance to tetracycline occurs by three mechanisms efflux, ribosomal
protection, and chemical modification (Brooks, 2007).
In an investigation done by El-Seedy et al.(2012), a high sensitivity (98%) was recorded to
methicillin in the examined S. aureus isolates isolated from buffaloes.
Al-Shamary (2012) found that S. aureus isolates were highly sensitive to tetracycline and
vancomycin as 100%, vancomycin sensitive S. aureus were also reported by Jayavanth and
Napori (2011). Fourty one percent of S. aureus isolates showed to be tetracycline resistant
in a study done by Duran et al.(2012), regarding vancomycin, high sensitivity was reported
as 100%.
Results obtained from a study done in a neighbouring country by Mirzaie et al. (2012)
showed that 54% of S. aureus isolates isolated from raw and pasteurized milk and ice
cream were MRSA, while only 7% of them were VRSA, and 23% were tetracycline
resistant.
Antibiotic resistance is a major public health concern since resistant bacteria can persist
and circulate in the environment with possible transmission to humans via contaminated
food and water, consequently MRSA is currently the most commonly identified antibiotic-
resistant pathogen in many countries worldwide (Bukharie, 2010; Ippolito et al., 2010).
COPS isolates derived from raw milk and cheese exhibited resistance to vancomycin, this
probably because MRSA isolates that are resistant to beta lactam drugs may develop
induced resistance to vancomycin (Deresinski, 2007; Ateba et al., 2010).
Regarding CONS isolates, they were methicillin resistant in 62.13 percent (64 isolates), 28
CONS isolates (27.18%) resist tetracycline, and only 5 CONS isolates (4.85%) were
vancomycin resistant (Figure 3-10). Results by local investigators showed higher
methicillin resistance rates among CONS isolates of 84% and 86.6% respectively in the
studies of Abd-Lateef (2011) and Al-Maliki (2009).
Duran et al.(2012) reported that 30.8% of CONS were tetracycline resistant, but none of
them resist vancomycin. Vancomycin has been the major drug used for treatment of
methicillin resistant staphylococci, but recovery of isolates with intermediate resistance and
the reports of several cases of high level resistance to vancomycin have spurred the search
for newer agents (Brooks, 2007).

CONS isolatesin the study of Soares et al.(2011) were multi-resistant in a percent 57.1%,
these results provided evidence that carrying antimicrobial resistance genes has became
reasonably widespread in milk and cheese samples.
The resistance to penicillin G is explained by the production of β-lactamase (Taponen and
Pyorala, 2009), while resistance to methicillin is encoded by mec A gene which is located
on Staphylococcus cassette chromosome (SCC), the SCC can be transferred horizontally
between various staphylococcal species which suggests that CONS acts as reservoir for the
dissemination of resistance genes to COPS (Irlinger, 2008; Piette and Verschraegen, 2009).
Resistance to methicillin in CONS of dairy products was recorded worldwide (Faria et al.,
2009; Sawant et al., 2009; Even et al., 2010).
The consumption of dairy products containing CONS is considered to be safe, no available
reports of CONS associated health hazards following the ingestion of dairy products
(Coton et al., 2010; Even et al ., 2010). Nevertheless, the occurrence of multidrug-resistant
CONS in dairy products and the possible transfer of drug resistance between
microorganisms in food matrices, suggests that only specifically selected antibiotic
susceptible isolates of CONS should be used in starter cultures (Resch et al., 2008; Even et
al., 2010).
3-4 Molecular Studies 3-4-1 DNA Extraction
3-4-1-1 DNA Extraction from Staphylococcal isolates
Staphylococcus is a gram positive bacteria, its thick peptidoglycan layer makes DNA
extraction an expensive and difficult work because of the requirement of lysostaphin with
lysozyme in addition to the cost of the extraction kit, for all this, here in this study a simple
method modified from that described by De Baere et al. (2002) was depended. Thus, using
boiling method, both DNA and RNA extracted simultaneously, as shown in figure 3-11,
DNA samples then subjected to agarose gel electrophoresis of 0.8% agarose for 1 hour at
70 volts.
Figure 3-11: DNA and RNA bands extracted from 11 staphylococcal isolates
Gel electrophoresis :agarose (0.8%), TBE buffer (1X), 5V/Cm for 1 h stained with ethidium bromide. M: The DNA molecular weight marker (1kb ladder); Lane 1: DNA extracted

from the isolate S 14; Lane 2: DNA extracted from the isolate S 17; Lane 3: DNA extracted from the isolate S 144; Lane 4: DNA extracted from the isolate S 33; Lane 5: DNA extracted from the isolate S 219; Lane 6: DNA extracted from the isolate S 211; Lane 7: DNA extracted from the isolate S 25; Lane 8: DNA extracted from the isolate S 117; Lane 9: DNA extracted from the isolate S 53; Lane 10: DNA extracted from the isolate S 95; Lane 11: DNA extracted from the isolate S 65. Physical cell wall disruption substituted the enzymatic cell wall digestion, no incubation
periods needed, only heating at 85oC for 20 min, and immediate freezing for 10 min, so,
after 30 min only, the DNA was extracted. While according Montanaro et al.(1999),
lysostaphin was added to the cell suspension with an incubation at 37oC for 30min, then
proteinase K with Tris-HCl (pH 7.5) were added, a further incubation period at 37oC for
20min, finally; heating at 95oC for 10 min (to inactivate proteinase K).
The protocol described in this study is suitable for DNA extraction from bacterial growth
from solid and liquid media. It was preferred to use growths from solid media as this
enhanced biosafety by reducing the chance of aerosol generation and laboratory spillover.
The classical method for extracting DNA from staphylococcal isolates is performed always
via initial digestion of the staphylococcal cells with lysostaphin or lysozyme and proteinase
K, and by subjecting the cell lysate to a wizard genomic DNA purification kit, and the
extraction continued according to the manufacturers’ protocol, which includes: an
extraction solution, column, washing solution, and an elution buffer.
Instead of all these long, expensive and difficult steps, here in this study; a physical cell
wall lysis substituted the initial enzymatic cell wall digestion and the using of commercial
kits. DNA samples extracted from staphylococcal isolates using heat shock method and
completed the extraction with the Genomic DNA extraction kit demonstrated in figure 3-
12.

Figure 3-12: DNA bands extracted from 2 staphylococcal isolates using heat shock with the Genomic DNA extraction kit
Gel electrophoresis: agarose (0.8%), TBE buffer (1X), 5V/Cm for 1 h stained with ethidium
bromide. Lane 1: DNA extracted from the isolate S 55; Lane 2: DNA extracted from the isolate S
117.
3-4-1-2 DNA Extraction from milk and cheese samples directly
DNA extraction from bacterial isolates which isolated from milk and cheese samples
after being collected and cultured on at least three types of media to confirm the isolation
of the intent bacteria is an expensive and time consuming routine work, at this study, an
alternative method to extract the bacterial DNA was depended, this method modified from
the method described by De Baere et al. (2002). Now simply with low coast materials,
with no need to isolate the bacteria, otherwise, directly from the food sample, DNA could
be extracted, as shown in figures 3-13 and 3-14.
Figure 3-13: DNA bands extracted from 5 staphylococcal isolates (to the left) and
Staphylococcal DNA extracted from 5 cheese samples directly (to the right) Gel electrophoresis :agarose (0.8%), TBE buffer (1X), 5V/Cm for 1 h stained with ethidium
bromide. M: The DNA molecular weight marker (1kb ladder); Lane 1: DNA extracted from the
isolate S 230; Lane 2: DNA extracted from the isolate S 83; Lane 3: DNA extracted from the isolate
S 23; Lane 4: DNA extracted from the isolate S 91; Lane 5: DNA extracted from the isolate S 211;
Lanes 6-10: DNA extracted from five cheese samples.

Figure 3-14: Staphylococcal DNA extracted from 7 milk samples directly Gel electrophoresis: agarose (0.8%), TBE buffer (1X), 5V/Cm for 1 h stained with ethidium bromide. DNA extraction from milk samples modified from Fusco et al. (2011); who depended a
pre-treatment of the milk sample by 1 volume of 2% sodium citrate, vortexing,
centrifugation at 20000 rpm for 15 min, then washing with TE buffer and lysostaphin
addition with incubation at 37oC for 30 min and then processing the lysate with an
extraction kit.
In this study, a modified way was followed; instead of enzymatic lysis of the rigid
staphylococcal cell wall by lysostaphin, a heat shock technique was used to achieve
physical cell wall disruption through applying heating the sample and the sudden freeze
after the washing steps. For further purification, alcohols added to obtain purified DNA.
DNA obtained using the methods described in this study consistently gave A260/A280 ratio
of 1.7–2.0 i.e. the extracted DNA was pure (Glasel, 1997) and was amenable to further
molecular techniques, including agarose gel electrophoresis, and PCR. PCR-based
techniques do not necessarily require highly pure DNA. However, pure DNA is
recommended for PCR-based assays such as the rep-PCR and RAPD, which are susceptible
to polymorphism, with the quality of DNA often affecting the banding pattern.
The method described by Baruzzi et al.(2005) for the DNA extraction from cheese sample
was modified in this study; Baruzzi et al.(2005) method illustrated by: a homogenized 1 ml
of 1:10 cheese suspension in 2% sodium citrate, centrifuged 14000 rpm, the pellet
dissolved in 500 µl of 1:39 Triton X-100 solution with shaking, a heating step was needed
at 70oC for 10 min, after centrifugation; the lysate was rinsed twice with distilled water and
thereafter a wizard genomic DNA purification kit was used and the DNA extraction
continued according to the manufacturers’ protocol (as described previously).
Here in this study a modified method was depended by employing physical cell wall
disruption instead of enzymatic usage (as a kit appendage for gram positive bacteria; which
was lysozyme buffer), and the kit purification was substituted by alcohols with several
washing steps.

3-4-2 Polymerase Chain Reaction (PCR) Techniques
3-4-2-1 coa gene amplification by monoplex PCR technique
To detect S. aureus isolates with coa gene, it was subjected to PCR technique in a
monoplex pattern.
coa positivity was confirmed by agarose gel electrophoresis in a 1.5% agarose stained with
ethidium bromide, electrophoresed in 75 volt for 2 hours and photographed under
ultraviolet (UV) transilluminator (Figure 3-15).
Figure 3-15: Gel electrophoresis of amplified PCR products of coa gene (730 bp) of S.
aureus isolates in monoplex PCR technique Agarose (1.5%), TBE buffer (1X), 5V/Cm for 3hrs stained with ethidium bromide. M: The DNA molecular weight marker (100bp ladder); Lane 1, Lane 2 and Lane 3: Positive amplification of 730 bp for coa gene of the isolates S 17, S 30 and S 94 respectively; Lane 4, and Lane 8: CONS isolates S 48 and S 105; Lane 5, Lane 6 and Lane 7: Positive amplification of 730 bp for coa gene of the isolates S 117, S 132 and S 195 respectively Lane 9, Lane 10 and Lane 11: Positive amplification of 730 bp for coa gene of the isolates S 198, S 204 and S 232 respectively. Table 3-5 showed the results of the coa gene amplification for S. aureus isolates from milk
and cheese samples.
Table 3-5: The coa gene amplification of the S. aureus isolates
Strain coa gene amplification
Strain coa gene amplification
Strain coa gene amplification
1 S 14 + 34 S 97 - 67 S 173 - 2 S 15 - 35 S 98 + 68 S 174 - 3 S 16 + 36 S 100 - 69 S 177 + 4 S 17 + 37 S 102 + 70 S 180 + 5 S 21 + 38 S 103 + 71 S 185 + 6 S 27 + 39 S 108 + 72 S 191 + 7 S 30 + 40 S 109 + 73 S 194 + 8 S 31 + 41 S 111 - 74 S 195 + 9 S 40 + 42 S 112 + 75 S 196 + 10 S 41 + 43 S 113 - 76 S 198 + 11 S 50 - 44 S 117 + 77 S 199 + 12 S 51 - 45 S 119 + 78 S 200 -

13 S 55 + 46 S 120 - 79 S 203 - 14 S 56 + 47 S 121 + 80 S 204 + 15 S 57 + 48 S 125 + 81 S 205 + 16 S 58 + 49 S 130 + 82 S 206 + 17 S 59 + 50 S 132 + 83 S 207 + 18 S 60 - 51 S 133 + 84 S 208 + 19 S 63 + 52 S 136 - 85 S 214 + 20 S 64 + 53 S 140 + 86 S 215 + 21 S 65 + 54 S 144 - 87 S 217 - 22 S 68 + 55 S 145 + 88 S 219 + 23 S 70 + 56 S 147 + 89 S 220 + 24 S 73 + 57 S 150 + 90 S 223 - 25 S 77 - 58 S 155 + 91 S 227 + 26 S 80 + 59 S 156 + 92 S 228 - 27 S 81 + 60 S 157 - 93 S 230 + 28 S 83 + 61 S 161 + 94 S 232 + 29 S 85 + 62 S 163 + 95 S 233 + 30 S 91 + 63 S 164 + 96 S 234 + 31 S 93 - 64 S 165 + 97 S 235 + 32 S 94 + 65 S 168 + 33 S 95 + 66 S 171 + (+) the isolate amplified coa gene (-) the isolate did not amplify coa gene Results were showed that 76 S. aureus isolates (78.35%) carried the coa gene. There
was no amplification product of the DNA from CONS (S.chromogenes and S. epidermidis).
Momtaz et al., (2011) demonstrated that 48.83% of S. aureus isolates harbored the coa
gene, because the 3' end of the coa gene contains a series of 81-bp tandem repeats, which
differ among S. aureus isolates.
Karahan and Cetinkaya (2007) in Turkey, reported that 80.6% of S. aureus isolated from
milk samples contained coa gene, while Veras et al. (2008) found that all S. aureus isolates
(100%) possessed the coa gene.
Aslantas et al. (2007) showed in their study that 98.7% of S. aureus isolates possessed coa
gene.
The coa and femA genes have been determined to be characteristic of S.aureus; coa is
100% specific for the species S. aureus while femA encodes a protein precursor essential
for peptidoglycan biosynthesis in S. aureus; therefore, primers for fem A amplification were
included as a positive control for coagulase-positive staphylococci (Veras et al., 2008).
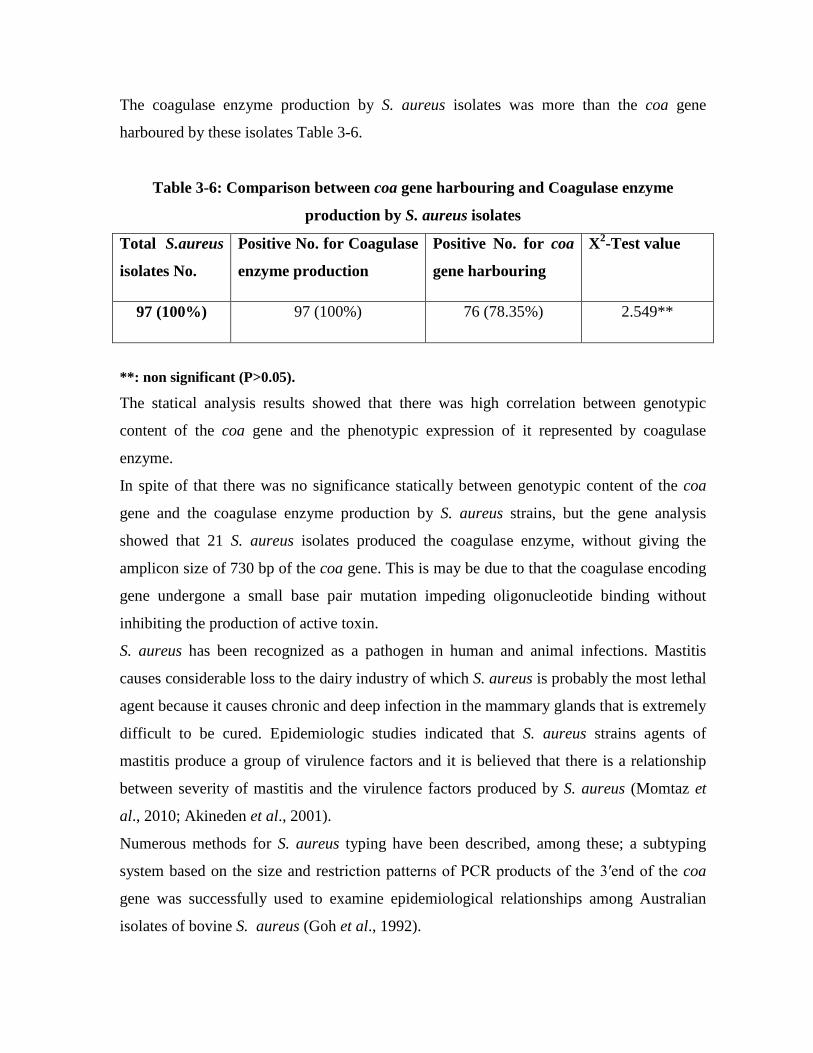
The coagulase enzyme production by S. aureus isolates was more than the coa gene
harboured by these isolates Table 3-6.
Table 3-6: Comparison between coa gene harbouring and Coagulase enzyme
production by S. aureus isolates
Total S.aureus
isolates No.
Positive No. for Coagulase
enzyme production
Positive No. for coa
gene harbouring
X2-Test value
97 (100%) 97 (100%) 76 (78.35%) 2.549**
**: non significant (P>0.05).
The statical analysis results showed that there was high correlation between genotypic
content of the coa gene and the phenotypic expression of it represented by coagulase
enzyme.
In spite of that there was no significance statically between genotypic content of the coa
gene and the coagulase enzyme production by S. aureus strains, but the gene analysis
showed that 21 S. aureus isolates produced the coagulase enzyme, without giving the
amplicon size of 730 bp of the coa gene. This is may be due to that the coagulase encoding
gene undergone a small base pair mutation impeding oligonucleotide binding without
inhibiting the production of active toxin.
S. aureus has been recognized as a pathogen in human and animal infections. Mastitis
causes considerable loss to the dairy industry of which S. aureus is probably the most lethal
agent because it causes chronic and deep infection in the mammary glands that is extremely
difficult to be cured. Epidemiologic studies indicated that S. aureus strains agents of
mastitis produce a group of virulence factors and it is believed that there is a relationship
between severity of mastitis and the virulence factors produced by S. aureus (Momtaz et
al., 2010; Akineden et al., 2001).
Numerous methods for S. aureus typing have been described, among these; a subtyping
system based on the size and restriction patterns of PCR products of the 3′end of the coa
gene was successfully used to examine epidemiological relationships among Australian
isolates of bovine S. aureus (Goh et al., 1992).

Additionally, molecular typing of staphylococcal isolates can provide useful information
about their epidemiological relatedness for the confirmation of staphylococcal food-borne
outbreaks. Several virulence factors including coagulase gene are produced by S.aureus
and may contribute to its pathogenicity and its detection is an important feature used
worldwide for the identification of S. aureus, expression of this gene is thought to enhance
bacterial growth and promote infection in the face of host defense mechanisms, such as
phagocytosis (Aarestrup et al., 1995).
3-4-2-2 femA and mecA genes amplification by monoplex PCR technique
The accurate and rapid diagnosis of antibiotic resistance genes in the treatment of
staphylococcal infections is extremely important in preventing the spread of infections.
PCR-based molecular methods are often preferred for determination of antibiotic resistance
genes. Using PCR technique, the genetic determinants of methicillin resistance femA and
mecA were co-amplified to identify susceptible (lacking mecA) and resistant (mecA+)
staphylococci and to differentiate S. aureus (femA+) from coagulase- negative
staphylococci (lacking femA). The results of the present study showed that, femA gene
band detected at 318bp region, all MRSA isolated strains were produced 318 bp band (Fig
3-16).
Figure 3-16: Gel electrophoresis of amplified PCR products of femA gene (318bp) of
S. aureus isolates in monoplex PCR technique Agarose (1.5%), TBE buffer (1X), 5V/Cm for 3hrs stained with ethidium bromide. M: The DNA molecular weight marker (100bp ladder "BIONEER''); Lane 1, Lane 2 and Lane 3: Positive amplification of 318 bp for femA gene of the isolates S 21, S 30 and S 102 respectively; Lane 4: femA negative CONS isolate S213; Lane 5, Lane 6 and Lane 7: Positive amplification of 318 bp for femA gene of the isolates S 117, S 140 and S 205 respectively. Ninety six S. aureus isolates (98.96%) were found to harbour femA gene, and all the MRSA
isolates do so (Table 3-7).
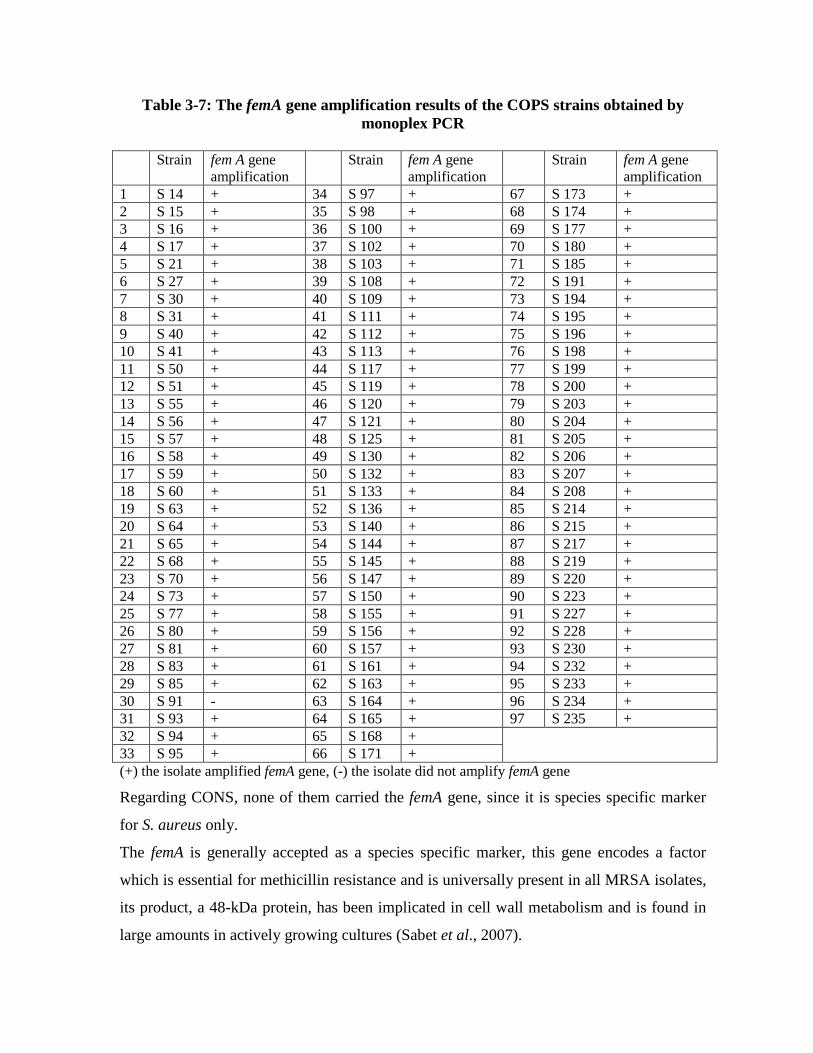
Table 3-7: The femA gene amplification results of the COPS strains obtained by monoplex PCR
Strain fem A gene
amplification Strain fem A gene
amplification Strain fem A gene
amplification 1 S 14 + 34 S 97 + 67 S 173 + 2 S 15 + 35 S 98 + 68 S 174 + 3 S 16 + 36 S 100 + 69 S 177 + 4 S 17 + 37 S 102 + 70 S 180 + 5 S 21 + 38 S 103 + 71 S 185 + 6 S 27 + 39 S 108 + 72 S 191 + 7 S 30 + 40 S 109 + 73 S 194 + 8 S 31 + 41 S 111 + 74 S 195 + 9 S 40 + 42 S 112 + 75 S 196 + 10 S 41 + 43 S 113 + 76 S 198 + 11 S 50 + 44 S 117 + 77 S 199 + 12 S 51 + 45 S 119 + 78 S 200 + 13 S 55 + 46 S 120 + 79 S 203 + 14 S 56 + 47 S 121 + 80 S 204 + 15 S 57 + 48 S 125 + 81 S 205 + 16 S 58 + 49 S 130 + 82 S 206 + 17 S 59 + 50 S 132 + 83 S 207 + 18 S 60 + 51 S 133 + 84 S 208 + 19 S 63 + 52 S 136 + 85 S 214 + 20 S 64 + 53 S 140 + 86 S 215 + 21 S 65 + 54 S 144 + 87 S 217 + 22 S 68 + 55 S 145 + 88 S 219 + 23 S 70 + 56 S 147 + 89 S 220 + 24 S 73 + 57 S 150 + 90 S 223 + 25 S 77 + 58 S 155 + 91 S 227 + 26 S 80 + 59 S 156 + 92 S 228 + 27 S 81 + 60 S 157 + 93 S 230 + 28 S 83 + 61 S 161 + 94 S 232 + 29 S 85 + 62 S 163 + 95 S 233 + 30 S 91 - 63 S 164 + 96 S 234 + 31 S 93 + 64 S 165 + 97 S 235 + 32 S 94 + 65 S 168 + 33 S 95 + 66 S 171 + (+) the isolate amplified femA gene, (-) the isolate did not amplify femA gene
Regarding CONS, none of them carried the femA gene, since it is species specific marker
for S. aureus only.
The femA is generally accepted as a species specific marker, this gene encodes a factor
which is essential for methicillin resistance and is universally present in all MRSA isolates,
its product, a 48-kDa protein, has been implicated in cell wall metabolism and is found in
large amounts in actively growing cultures (Sabet et al., 2007).

Analysis of the femA product indicated that this protein is associated with the expression of
high levels of methicillin resistance without affecting PBP-2` production, the significance
of the femA genes in the mechanism of methicillin resistance was supported by the
demonstration that S.aureus strain with femA gene inactivated lost; the methicillin
resistance trait, but with the transduction of femA genes restored the resistance (Giannouli
et al., 2010). Another biochemical analysis suggested that femA product may be involved in
the metabolism of cell wall synthesis (Maidhof et al., 1991).
mecA gene, which encodes production of an altered penicillin binding protein (PBP)
(PBP2a or PBP2’) a modified transpeptidase that has a low affinity for all beta-lactam
antimicrobials (penicillins, cephalosporins, carbapenems) (Scott et al., 2010).
The 533 bp mecA was detected in 91 (93.81%) MRSA isolates, while it was detected in 70
(67%) CONS isolates (Figure 3-17).
Figure 3-17: Gel electrophoresis of amplified PCR products of mecA gene (533bp) from staphylococci isolates by monoplex PCR technique

Agarose (1.5%), TBE buffer (1X), 5V/Cm for 3hrs stained with ethidium bromide: M: The DNA molecular weight marker (100bp ladder "BIONEER''); Lane 1: mecA negative S 112; Lane 2, Lane 3 and Lane 4: Positive amplification of 533 bp for mecA gene of the COPS isolates S 14, S 55 and S 117 respectively; Lane 5: Negative control (Standard strain S. aureus ATCC 25923 free of antibiotic resistance genes); Lane 6, Lane 7 and Lane 8: Positive amplification of 533 bp for mecA gene of the CONS isolates S 33, S 39 and S 174 respectively. Results of mecA gene amplification by COPS and CONS isolates were demonstrated in the
tables 3-8, and 3-9 respectively.
Table 3-8: mecA gene amplification results of the COPS strains obtained by monoplex
PCR
Strain mecA gene amplification
Strain mecA gene amplification
Strain mecA gene amplification
1 S 14 + 34 S 97 + 67 S 173 + 2 S 15 - 35 S 98 + 68 S 174 + 3 S 16 + 36 S 100 + 69 S 177 + 4 S 17 + 37 S 102 + 70 S 180 + 5 S 21 + 38 S 103 + 71 S 185 + 6 S 27 + 39 S 108 + 72 S 191 + 7 S 30 + 40 S 109 + 73 S 194 + 8 S 31 + 41 S 111 + 74 S 195 + 9 S 40 + 42 S 112 - 75 S 196 + 10 S 41 + 43 S 113 + 76 S 198 + 11 S 50 + 44 S 117 + 77 S 199 + 12 S 51 + 45 S 119 + 78 S 200 + 13 S 55 + 46 S 120 + 79 S 203 + 14 S 56 + 47 S 121 + 80 S 204 + 15 S 57 + 48 S 125 + 81 S 205 + 16 S 58 + 49 S 130 - 82 S 206 + 17 S 59 + 50 S 132 + 83 S 207 + 18 S 60 + 51 S 133 + 84 S 208 + 19 S 63 + 52 S 136 + 85 S 214 + 20 S 64 - 53 S 140 + 86 S 215 + 21 S 65 + 54 S 144 + 87 S 217 + 22 S 68 + 55 S 145 + 88 S 219 + 23 S 70 + 56 S 147 + 89 S 220 + 24 S 73 + 57 S 150 + 90 S 223 + 25 S 77 + 58 S 155 + 91 S 227 + 26 S 80 + 59 S 156 + 92 S 228 - 27 S 81 + 60 S 157 + 93 S 230 + 28 S 83 + 61 S 161 + 94 S 232 + 29 S 85 + 62 S 163 + 95 S 233 + 30 S 91 + 63 S 164 + 96 S 234 + 31 S 93 + 64 S 165 + 97 S 235 + 32 S 94 - 65 S 168 +

33 S 95 + 66 S 171 + (+) COPS isolate amplified mecA gene (-) COPS isolate did not amplify mecA gene
Table 3-9: mecA gene amplification of the CONS strains
Strain mecA gene amplification
Strain mecA gene amplification
Strain mecA gene amplification
1 S 1 + 36 S 48 + 71 S 124 + 2 S 2 + 37 S 49 - 72 S 126 - 3 S 3 + 38 S 52 + 73 S 127 + 4 S 4 - 39 S 53 + 74 S 128 + 5 S 5 + 40 S 54 + 75 S 129 + 6 S 7 + 41 S 61 - 76 S 131 - 7 S 8 + 42 S 62 + 77 S 134 + 8 S 9 + 43 S 66 + 78 S 135 + 9 S 10 + 44 S 67 - 79 S 160 + 10 S 11 - 45 S 69 + 80 S 162 + 11 S 12 - 46 S 71 - 81 S 175 - 12 S 13 + 47 S 72 + 82 S 178 + 13 S 18 + 48 S 74 + 83 S 179 + 14 S 19 + 49 S 75 - 84 S 183 + 15 S 20 - 50 S 76 - 85 S 190 + 16 S 22 + 51 S 78 - 86 S 192 - 17 S 23 - 52 S 79 + 87 S 193 + 18 S 24 + 53 S 82 + 88 S 197 + 19 S 25 + 54 S 84 - 89 S 201 + 20 S 26 - 55 S 86 + 90 S 202 + 21 S 28 + 56 S 87 - 91 S 210 + 22 S 29 + 57 S 88 + 92 S 211 + 23 S 33 + 58 S 89 + 93 S 212 + 24 S 34 + 59 S 90 + 94 S 213 - 25 S 35 - 60 S 92 - 95 S 216 + 26 S 36 - 61 S 96 + 96 S 218 + 27 S 37 + 62 S 99 + 97 S 221 + 28 S 38 - 63 S 101 - 98 S 222 - 29 S 39 + 64 S 104 + 99 S 224 + 30 S 42 - 65 S 105 + 100 S 225 + 31 S 43 + 66 S 107 + 101 S 226 + 32 S 44 - 67 S 114 - 102 S 229 + 33 S 45 + 68 S 115 - 103 S 231 - 34 S 46 - 69 S 122 + 35 S 47 - 70 S 123 + (+) CONS isolate amplified mecA gene (-) CONS isolate did not amplify mecA gene

The virulence genes mecA and femA were examined because they are believed to be the
major contributors to methicillin resistance. It is known that methicillin resistance could
manifest with or without mecA, auxillary genes like femA influence the extent of resistance
implying that there could be other mechanisms that circumvent absence of gene to confer
methicillin-resistance (Chikkala et al., 2012).
The inclusion of an internal positive control femA in the reaction provides assurance against
false-negative results (Mehrotra et al., 2000).
Food is a main source for the transfer of antibiotic resistance, such transfer may occur by
means of antibiotic residues in food, through the transfer of resistant food-borne pathogens,
or through the ingestion of resistant strains of the original food microflora and resistance
transfer to pathogenic microorganisms (Pereira et al., 2009; Pesavento et al., 2007).
Although the detection of mecA gene remains the gold standard for detecting methicillin-
resistance, its detection alone does not confirm the presence of S. aureus and there is no
consensus on the molecular target that could be used to confirm the S. aureus species,
constitutively expressed genes such as femA, femB and nuc are being used as molecular
targets for the identification of S. aureus polymorphisms within these constitutive genes
have been reported worldwide, from another point of view, mecA alone does not solely
confer the methicillin resistance, studies have shown that fem (factors essential for
methicillin-resistance) or the auxiliary genes like fem A/B/X in addition to mecA are also
important in expression of methicillin resistance, the fem ABX operon encodes factors
which are responsible for the formation of pentaglycine bridges in the cell wall of
Staphylococci (Carroll, 2008;Chikkala et al., 2012).
In the present study, the results of antibiotic susceptibility by disk diffusion method were
compared with gene analysis results in staphylococcal isolates (Table 3-10), because the
phenotypic expression of antimicrobial resistance has been reported to be influenced by
various factors.
Table 3-10: Relationship between Methicillin resistance and the presence of femA and mecA genes in both COPS and CONS
Staphylococcal
isolates Positive No. of Methicillin resistant Staphylococci by
phenotypic method
The distribution of methicillin resistance genes
X2-Test value
Positive No. of Positive No. of

femA mecA 97 COPS isolates (100%)
80 (82.47%) 96(98.96%) 91(93.81%) 1.506**
103CONS isolates (100%)
64 (62.13%) 0 (0%) 70 (67.96%) 0.269**
**: non significant (P>0.05).
The statical analysis results showed that there was high correlation between genotypic
content of the femA and mecA genes and the phenotypic expression of them when tested by
antibiotic disc diffusion method.
There was no significance statically between genotypic content and phenotypic expression
of them represented by methicillin resistance.
Gene analysis results confirmed that none of the CONS strains harboured the femA gene;
all of them harboured the mecA gene, which means that CONS’ methicillin resistance
refers to the mecA gene only, while COPS’ methicillin resistance refers to both of the femA
and mecA genes.
Nafisi et al. (2008) screened on 52 isolates of coagulase positive S. aureus, using agar
screen and duplex PCR methods and reported that phenotypically 23 cases (44%) of the
isolates, and genotypically mecA 27 cases of the isolates (52 %) are resistant to methicillin.
Duran et al., (2012) had reported that methicillin resistance was observed in 17.8 % isolates
when tested by methicillin disk diffusion method, whereas 27.9 % isolates had mecA gene.
Phenotypically methicillin susceptible 30 isolates also carried the mecA gene.
Chikkala et al. (2012) observed a rather high rate (58.3%) detection of mecA among MSSA
isolates.
Standardized methods of susceptibility test have been used for the detection of MRSA
strains: Kirby-Bauer antibiotic testing (disc diffusion antibiotic sensitivity test), oxacillin
agar screen test, agar dilution and the Epsilometer test (E-test) are commonly used to
determine the MRSA phenotype in the clinical microbiology laboratory, these tests have
limitations and frequently show variations, however, phenotypic expression of methicillin-
resistance can be heterogenous, in addition, methicillin resistance is influenced by culture
conditions such as temperature, pH and sodium chloride (NaCl) content in the medium;
these factors complicate the detection of methicillin resistance, especially for strains with

low level resistance, the PCR methods have high sensitivity and specificity and are
independent of the physical and chemical conditions of the culture (Zamani et al., 2007;
Mirzaei et al., 2012).
The results of this study showed a high presence of S. aureus resistant to methicillin in the
milk and cheese, therefore, food handlers should take appropriate measures to prevent the
spread of MRSA by contaminated raw foods and to prevent the occurrence, growth and
survival of MRSA in prepared food.
3-4-3 Detection of Enterotoxigenicity using multiplex PCR technique One of the criteria for confirmation of staphylococcal food poisoning (SFP) is whether the
recovered isolates have enterotoxigenicity, in contrast, non-SE-producing S. aureus strains
are not considered etiologic organisms for food poisoning (Hennekinne et al., 2011). The
detection of staphylococcal enterotoxigenicity according to four classical enterotoxins
genes which are sea, seb, sec, and sed was simultaneously performed using multiplex PCR
assay, the results were shown in figure 3-18.
Figure 3-18: Gel electrophoresis of amplified PCR products of staphylococcal
enterotoxin genes (sea, seb, sec, and sed) in multiplex PCR technique Agarose (1.5%), TBE buffer (1X), 5V/Cm for 3hrs stained with ethidium bromide: M: The DNA molecular weight marker (100bp ladder); Lane 1 and Lane 3: Positive amplification of 477 bp for enterotoxin B of the isolates S 97 and S 36 respectively; Lane 2: Positive amplification of 127 bp
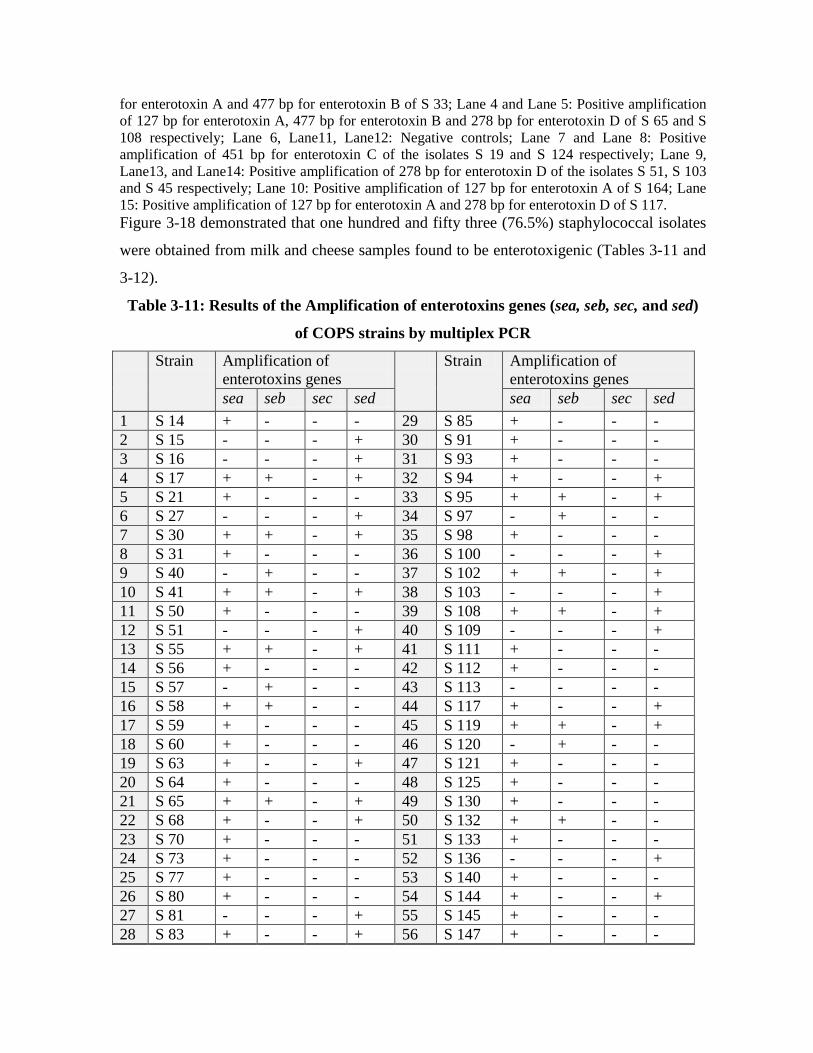
for enterotoxin A and 477 bp for enterotoxin B of S 33; Lane 4 and Lane 5: Positive amplification of 127 bp for enterotoxin A, 477 bp for enterotoxin B and 278 bp for enterotoxin D of S 65 and S 108 respectively; Lane 6, Lane11, Lane12: Negative controls; Lane 7 and Lane 8: Positive amplification of 451 bp for enterotoxin C of the isolates S 19 and S 124 respectively; Lane 9, Lane13, and Lane14: Positive amplification of 278 bp for enterotoxin D of the isolates S 51, S 103 and S 45 respectively; Lane 10: Positive amplification of 127 bp for enterotoxin A of S 164; Lane 15: Positive amplification of 127 bp for enterotoxin A and 278 bp for enterotoxin D of S 117. Figure 3-18 demonstrated that one hundred and fifty three (76.5%) staphylococcal isolates
were obtained from milk and cheese samples found to be enterotoxigenic (Tables 3-11 and
3-12).
Table 3-11: Results of the Amplification of enterotoxins genes (sea, seb, sec, and sed)
of COPS strains by multiplex PCR
Strain Amplification of enterotoxins genes
Strain Amplification of enterotoxins genes
sea seb sec sed sea seb sec sed 1 S 14 + - - - 29 S 85 + - - - 2 S 15 - - - + 30 S 91 + - - - 3 S 16 - - - + 31 S 93 + - - - 4 S 17 + + - + 32 S 94 + - - + 5 S 21 + - - - 33 S 95 + + - + 6 S 27 - - - + 34 S 97 - + - - 7 S 30 + + - + 35 S 98 + - - - 8 S 31 + - - - 36 S 100 - - - + 9 S 40 - + - - 37 S 102 + + - + 10 S 41 + + - + 38 S 103 - - - + 11 S 50 + - - - 39 S 108 + + - + 12 S 51 - - - + 40 S 109 - - - + 13 S 55 + + - + 41 S 111 + - - - 14 S 56 + - - - 42 S 112 + - - - 15 S 57 - + - - 43 S 113 - - - - 16 S 58 + + - - 44 S 117 + - - + 17 S 59 + - - - 45 S 119 + + - + 18 S 60 + - - - 46 S 120 - + - - 19 S 63 + - - + 47 S 121 + - - - 20 S 64 + - - - 48 S 125 + - - - 21 S 65 + + - + 49 S 130 + - - - 22 S 68 + - - + 50 S 132 + + - - 23 S 70 + - - - 51 S 133 + - - - 24 S 73 + - - - 52 S 136 - - - + 25 S 77 + - - - 53 S 140 + - - - 26 S 80 + - - - 54 S 144 + - - + 27 S 81 - - - + 55 S 145 + - - - 28 S 83 + - - + 56 S 147 + - - -

Strain Amplification of enterotoxins genes
Strain Amplification of enterotoxins genes
sea seb sec sed sea seb sec sed 57 S 150 + - - - 78 S 200 + + - - 58 S 155 + + - - 79 S 203 - - - + 59 S 156 + - - - 80 S 204 + - - + 60 S 157 - - - + 81 S 205 + + - + 61 S 161 + - - + 82 S 206 + - - - 62 S 163 + + - - 83 S 207 + - - + 63 S 164 + - - - 84 S 208 + + - - 64 S 165 + - - - 85 S 214 + - - + 65 S 168 + - - + 86 S 215 + - - - 66 S 171 - - - - 87 S 217 + - - + 67 S 173 + - - - 88 S 219 + - - + 68 S 174 - - - + 89 S 220 + - - - 69 S 177 + - - - 90 S 223 - + - - 70 S 180 + - - - 91 S 227 - - - + 71 S 185 + - - - 92 S 228 - - - + 72 S 191 + - - + 93 S 230 + - - - 73 S 194 + + - + 94 S 232 + + - + 74 S 195 + - - + 95 S 233 - - - + 75 S 196 + - - - 96 S 234 + - - + 76 S 198 + - - + 97 S 235 - - - + 77 S 199 - - - + (+) COPS isolate amplified one or more of enterotoxins coding genes sea, esb, sec, sed (-) COPS isolate did not amplify one or more of enterotoxins coding genes sea, esb, sec, sed
Table 3-12: Results of the Amplification of enterotoxins genes (sea, seb, sec, and sed)
of CONS strains by multiplex PCR
Strain Amplification of enterotoxins genes
Strain Amplification of enterotoxins genes
sea seb sec sed sea seb sec sed 1 S 1 - - - - 34 S 46 - - - -

2 S 2 + - - + 35 S 47 + - - - 3 S 3 - - - + 36 S 48 - - - - 4 S 4 - - - - 37 S 49 + - - + 5 S 5 - - - + 38 S 52 - - - - 6 S 7 - - - - 39 S 53 - - - - 7 S 8 + - - - 40 S 54 + - - - 8 S 9 + - - + 41 S 61 - - - - 9 S 10 - - - - 42 S 62 + - - - 10 S 11 - - - - 43 S 66 - - - - 11 S 12 - - - - 44 S 67 - - - - 12 S 13 + - - + 45 S 69 - - - - 13 S 18 - - - + 46 S 71 - - - - 14 S 19 - - + - 47 S 72 - - - - 15 S 20 + - - - 48 S 74 + + - - 16 S 22 - - - - 49 S 75 - - - + 17 S 23 + - - - 50 S 76 - + - - 18 S 24 - - - + 51 S 78 + - - - 19 S 25 + - - - 52 S 79 - - - + 20 S 26 - - - - 53 S 82 - - - - 21 S 28 + - - - 54 S 84 - - - + 22 S 29 - - - + 55 S 86 - - - - 23 S 33 + + - - 56 S 87 + - - - 24 S 34 - - - - 57 S 88 - - - - 25 S 35 - - - - 58 S 89 + - - + 26 S 36 - + - - 59 S 90 - - - + 27 S 37 + - - + 60 S 92 - - - - 28 S 38 - - - - 61 S 96 - - - - 29 S 39 - - - + 62 S 99 - - - - 30 S 42 + - - - 63 S 101 - - - + 31 S 43 - - - - 64 S 104 - - - - 32 S 44 + - - + 65 S 105 + - - + 33 S 45 - - - + 66 S 107 + - - - Strain Amplification of
enterotoxins genes Strain Amplification of
enterotoxins genes sea seb sec sed sea seb sec sed
67 S 114 + - - - 86 S 192 - - - - 68 S 115 + - - - 87 S 193 + - - - 69 S 122 - - - - 88 S 197 - - - + 70 S 123 + - - - 89 S 201 + - - - 71 S 124 - - + - 90 S 202 + - - + 72 S 126 - - - - 91 S 210 - - - - 73 S 127 + - - + 92 S 211 + - - + 74 S 128 + - - - 93 S 212 + - - + 75 S 129 + - - - 94 S 213 - - - -
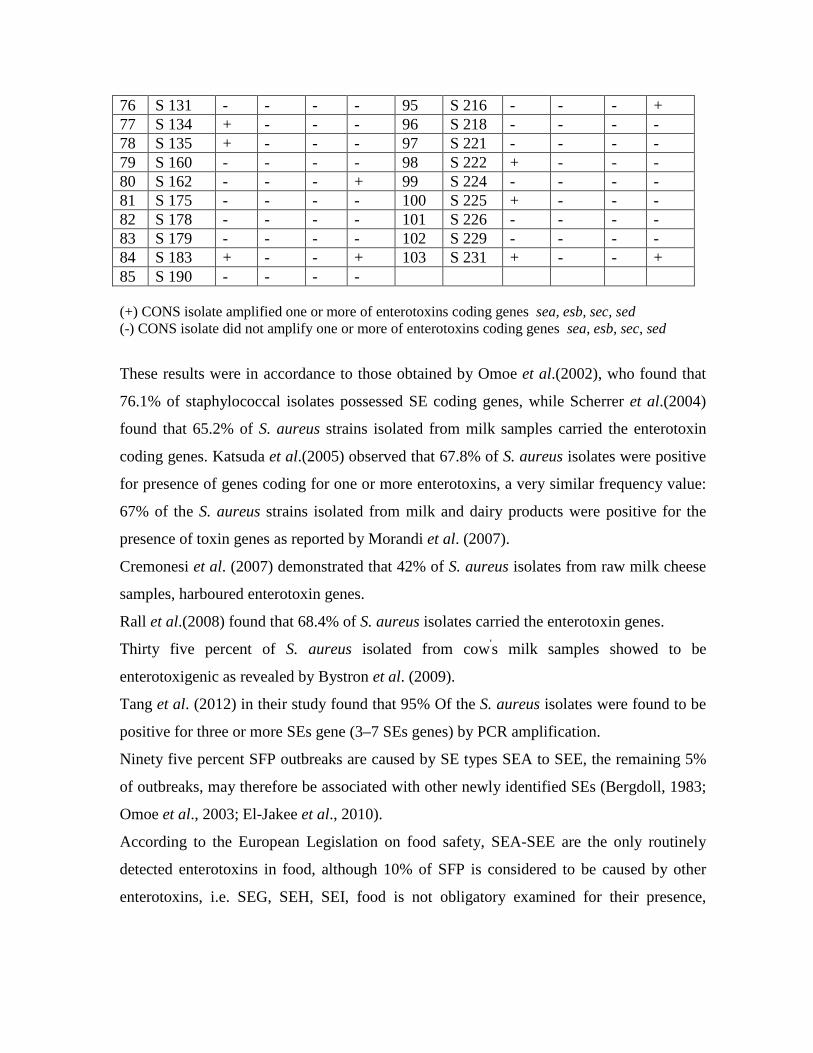
76 S 131 - - - - 95 S 216 - - - + 77 S 134 + - - - 96 S 218 - - - - 78 S 135 + - - - 97 S 221 - - - - 79 S 160 - - - - 98 S 222 + - - - 80 S 162 - - - + 99 S 224 - - - - 81 S 175 - - - - 100 S 225 + - - - 82 S 178 - - - - 101 S 226 - - - - 83 S 179 - - - - 102 S 229 - - - - 84 S 183 + - - + 103 S 231 + - - + 85 S 190 - - - - (+) CONS isolate amplified one or more of enterotoxins coding genes sea, esb, sec, sed (-) CONS isolate did not amplify one or more of enterotoxins coding genes sea, esb, sec, sed
These results were in accordance to those obtained by Omoe et al.(2002), who found that
76.1% of staphylococcal isolates possessed SE coding genes, while Scherrer et al.(2004)
found that 65.2% of S. aureus strains isolated from milk samples carried the enterotoxin
coding genes. Katsuda et al.(2005) observed that 67.8% of S. aureus isolates were positive
for presence of genes coding for one or more enterotoxins, a very similar frequency value:
67% of the S. aureus strains isolated from milk and dairy products were positive for the
presence of toxin genes as reported by Morandi et al. (2007).
Cremonesi et al. (2007) demonstrated that 42% of S. aureus isolates from raw milk cheese
samples, harboured enterotoxin genes.
Rall et al.(2008) found that 68.4% of S. aureus isolates carried the enterotoxin genes.
Thirty five percent of S. aureus isolated from cow's milk samples showed to be
enterotoxigenic as revealed by Bystron et al. (2009).
Tang et al. (2012) in their study found that 95% Of the S. aureus isolates were found to be
positive for three or more SEs gene (3–7 SEs genes) by PCR amplification.
Ninety five percent SFP outbreaks are caused by SE types SEA to SEE, the remaining 5%
of outbreaks, may therefore be associated with other newly identified SEs (Bergdoll, 1983;
Omoe et al., 2003; El-Jakee et al., 2010).
According to the European Legislation on food safety, SEA-SEE are the only routinely
detected enterotoxins in food, although 10% of SFP is considered to be caused by other
enterotoxins, i.e. SEG, SEH, SEI, food is not obligatory examined for their presence,

currently PCR is the only method used to demonstrate the presence of SE genes in S.
aureus strains (Bystron et al., 2009).
The results of this study included both COPS (S. aureus) and CONS (S. chromogenes and
S. epidermidis) isolates, CONS isolates showed to be enterotoxigenic in this research, they
accomplished 37.9% of the total enterotoxigenic strains (58 strains), as previously
demonstrated in table (3-13) while COPS were the predominant enterotoxigenic strains,
since 95 S.aureus isolate (62.09%) found to harbor one or more enterotoxin gene, (Figure
3-19).
Figure 3-19: Enterotoxigenicity of both Coagulase-positive (COPS) and Coagulase-negative Staphylococci (CONS)
Valle et al.(1990) revealed that 22% of CONS strains produced enterotoxins, whereas
Cunha et al.(2006) found that 15% of CONS isolates harboured the sea gene and 5%
carried the sec gene.
Soares et al.(2011) found that CONS isolates had low incidence of enterotoxin genes as
8.6%.
According to Cunha et al.(2006); no enterotoxin production was observed by
immunological method, while PCR assay showed that the toxigenic capacity of CONS
should not be ignored, and thus requiring more investigation of this group of
microorganisms in food. The detection of an even small number of CONS isolates
Enterotoxigenic CONS 38%
Enterotoxigenic COPS 62%

harbouring enterotoxin genes, is important, because confirms that CONS isolated from
foods can produce enterotoxins.
Staphylococcal intoxication occurs after the ingestion of food contaminated with
enterotoxin produced by bacteria of the Staphylococcus, and S. aureus is the main agent,
however, some authors have reported the isolation of other enterotoxigenic coagulase
positive species, such as S.intermedius and S.hyicus, studies have also reported the
enterotoxigenicity of coagulase negative staphylococci, including, S. epidermidis,
S.chromogenes, S. saprophyticus, S.sciuri, S.warneri, S.cohnii and S. lentus (Cunha and
Calsolari, 2007).
The sea gene was the most frequent enterotoxin coding gene among the others tested. It
was found that 112 staphylococcal isolates isolated from milk and cheese carried the sea
gene, sea accomplished 51.85% of the detected enterotoxin genes followed by the sed
gene, which constituted 34.73%, 75 staphylococcal isolates harboured it, then the seb
coding gene by 12.5%, it was carried by 27 staphylococcal isolates, while the sec gene was
very rare as 0.92%; two staphylococcal isolates only found to harbour the sec gene (Figure
3-20).
Figure 3-20: The percentage of each staphylococcal enterotoxin gene of the four SEs genes tested (sea, seb, sec, and sed) by multiplex PCR assay
0.00%
10.00%
20.00%
30.00%
40.00%
50.00%
60.00%
sea seb sec sed
51.85%
12.50%
0.92%
34.73%
Ente
roto
xin
Perc
enta
ge
sea gene
seb gene
sec gene
sed gene

The results of this study were closely related to those obtained by Cremonesi et al. (2007),
that sea and sed genes were prevalent in S. aureus isolates isolated from raw milk cheese
samples. Efuntoye et al.(2011) results showed that SEA was the predominant enterotoxin
produced by the S. aureus isolates, it was detected in 85% isolates and in combination with
SED in 20% isolates and with SEC in 10% isolates. SEA is the most frequent enterotoxin
involved in staphylococcal food poisoning.
It is widely accepted that SE production is characteristic of COPS, and most studies have
dealt with S. aureus. Geniogeorgis (1989) concluded that there is no prevailing type of SEs,
apart from the strains isolated from foods involved in staphylococcal gastroenteritis, where
SEA is the main type of SE recovered. In an epidemiological analysis, it has been reported
that a total of 100 to 200 ng of SEA in food was sufficient to cause food poisoning
(Evenson et al., 1988; Gencay et al., 2010).
It was stated that the toxigenic strains isolated from cheese samples were sea positive
(Giannatale et al., 2011).
Previous data also indicated that SEA is mostly involved in outbreaks of staphylococcal
food poisoning, Lim et al. (2004) determined 22.28 % of S. aureus isolates harboured sea,
seb, and sec genes while sea was the most frequent SE gene as it occupies 86.48%.
In the study of Bautista et al.(1988) the most frequently detected enterotoxins in S. aureus
isolates isolated from sheep milk were SEA and SED produced as 35.48% and 34.67%
respectively, whereas SED and SEC were elaborated by 6.45% and 21.77% respectively.
sea gene was the most frequent enterotoxin coding gene as 41%, followed by sec (20.5%),
sed (12.8%), and seb (7.7%) in the study of Rall et al. (2008).
Manfredi et al.(2010) found that 55.7% of the staphylococcal isolates carried the sea gene,
14.8% carried the seb gene, while only 6.5% harboured the sec gene.
Detection of S. aureus strains which harbour the gene for SEA synthesis is important
because the SEA is toxic in low concentrations (0.6 ng/ml) Holeckova et al.,(2002). In the
study of Naresh, (2000) fifty percent of the staphylococcal isolates carried the sed gene.
Harvey et al.(1982) have previously reported a predominance of sed positive strains in their
study on food isolates also the occurrence of the sed gene was the highest (47.4%) among
the classical enterotoxin coding genes in the study of Tang et al., (2012) followed by sea
(36.8%).
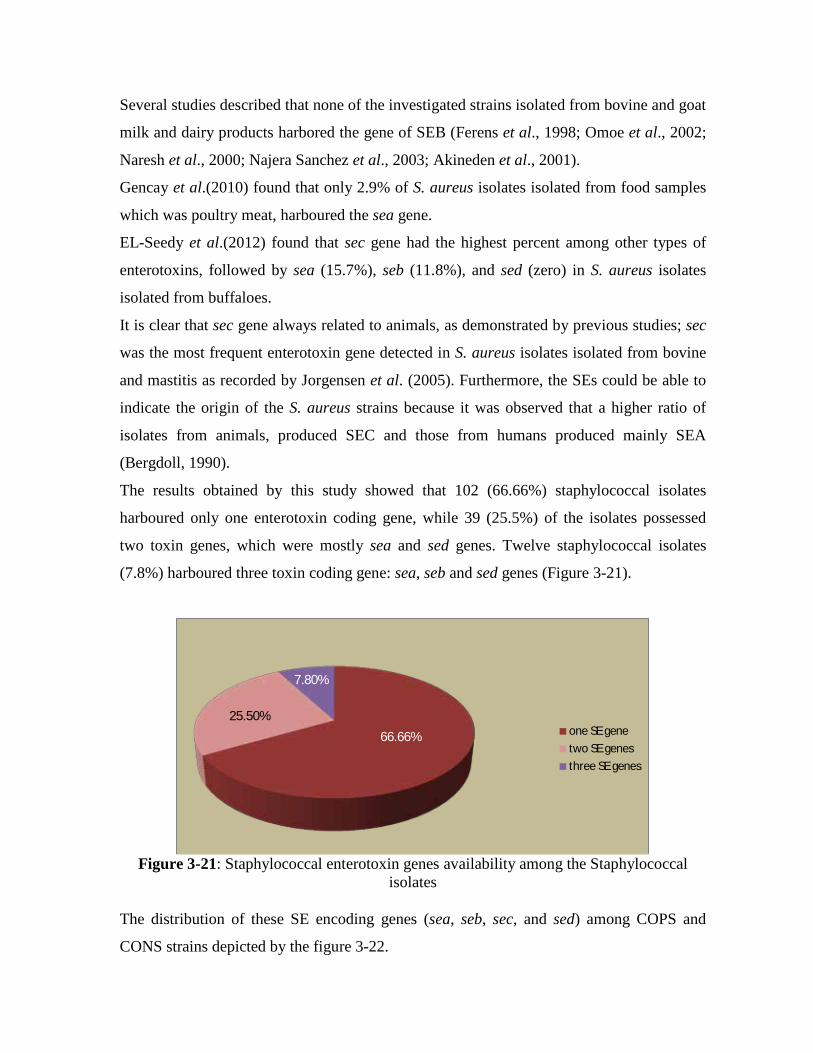
Several studies described that none of the investigated strains isolated from bovine and goat
milk and dairy products harbored the gene of SEB (Ferens et al., 1998; Omoe et al., 2002;
Naresh et al., 2000; Najera Sanchez et al., 2003; Akineden et al., 2001).
Gencay et al.(2010) found that only 2.9% of S. aureus isolates isolated from food samples
which was poultry meat, harboured the sea gene.
EL-Seedy et al.(2012) found that sec gene had the highest percent among other types of
enterotoxins, followed by sea (15.7%), seb (11.8%), and sed (zero) in S. aureus isolates
isolated from buffaloes.
It is clear that sec gene always related to animals, as demonstrated by previous studies; sec
was the most frequent enterotoxin gene detected in S. aureus isolates isolated from bovine
and mastitis as recorded by Jorgensen et al. (2005). Furthermore, the SEs could be able to
indicate the origin of the S. aureus strains because it was observed that a higher ratio of
isolates from animals, produced SEC and those from humans produced mainly SEA
(Bergdoll, 1990).
The results obtained by this study showed that 102 (66.66%) staphylococcal isolates
harboured only one enterotoxin coding gene, while 39 (25.5%) of the isolates possessed
two toxin genes, which were mostly sea and sed genes. Twelve staphylococcal isolates
(7.8%) harboured three toxin coding gene: sea, seb and sed genes (Figure 3-21).
Figure 3-21: Staphylococcal enterotoxin genes availability among the Staphylococcal
isolates
The distribution of these SE encoding genes (sea, seb, sec, and sed) among COPS and
CONS strains depicted by the figure 3-22.
66.66% 25.50%
7.80%
one SE genetwo SE genesthree SE genes

Figure 3-22: Distribution of the SE encoding genes (sea, seb, sec, and sed) among COPS
and CONS strains Fooladi et al.(2010) investigated the enterotoxigenicity of S. aureus isolates and found that
15.6% of these isolates possessed the sea gene, 9.3% had the seb gene, and 6.2% possessed
both genes.
A study done by Manfredi et al.(2010) revealed that 9.9% carried both of the sea and seb
genes, 3.3 % had the sea and sec genes, and only 1.7% of the staphylococcal isolates
carried both of the sea and sed genes.
Peles et al. (2007) found that 27.1% of S. aureus isolates isolated from unprocessed cow's
milk, harboured one or more enterotoxin coding genes, 25.42% carried a single gene
(mainly seb), and 1.69 % carried two genes.
Rall et al. (2008) revealed that 64.1% exhibited only one enterotoxin gene, 23.1% carried
genes coding for two enterotoxins, and 5.1% were positive for three enterotoxin coding
genes.
It was found that 15% of S. aureus isolates isolated from meat samples carried the sed
gene, while only 1% possessed the sea gene; but neither seb nor sec had been harboured by
these isolates (Pu et al., 2011).
A+B+D A+D A+B A B C DCOPS 12 17 6 38 5 0 17CONS 0 14 2 23 2 2 15
0
5
10
15
20
25
30
35
40
No.
of s
taph
yloc
occa
l str
ains

Giannatale et al.(2011) found that 44.4% S. aureus isolates isolated from food samples,
harboured one enterotoxin coding gene (sea or sec), 61.6% carried two genes, and16.1%
carried three toxin coding genes.
Foods of animal origin, especially milk and dairy products, are associated with food-born
disease (Jorgensen et al., 2005a). The fact that S.aureus is the main etiological agent of
bovine mastitis makes milk a great vehicle for its dispersion (Stamford et al., 2006).
It is still common to find non-refrigerated milk containers by the roads waiting for
collection by the cooperative,s truck, in spite of this problem, most of the milk produced is
processed, however, in small or even in large cities, commercialization of raw milk and
cheese, which called "informal marketing" is still usual, this practice increases the risk of
staphylococcal food poisoning due to inadequate storage conditions before pasteurization
(Rall et al., 2008) .
According to Schmitz et al.(1998) in clinical practice staphylococcal strains carrying a
toxin gene should be considered potential producers of this toxin since toxin production in
vivo can not be excluded.
Milk is a good substrate for S.aureus growth and enterotoxin production; in addition
enterotoxins retain their biological activity even after pasteurization (Asao et al., 2003).
According to Anderson et al.(1996), staphylococcal enterotoxins are very resistant to heat;
SEA, for example, retains some biological activity after 28 min at 121oC.
Many factors affect S. aureus growth and enterotoxin production in foodstuffs, such as the
presence of specific amino acids that vary with the strains (Onoue and Mori, 1997),
glucose, pH, acetic acid, lactic acid, sodium chloride and competing microorganisms (Loir
et al., 2003).
Thus considerable research effort is still required for better understanding of the interaction
between CONS and the food matrix, and the mechanisms of enterotoxin production in
foodstuffs.
PCR has been showed to be a rapid and sensitive method for the detection of
enterotoxigenic S. aureus. The detection of specific gene for an enterotoxin doesn’t
necessarily indicate the ability of the microorganism to produce the intact and biologically
active toxin, or enough amounts to cause disease (McLauchlin et al., 2000). However it has
been shown that strains containing genes for certain toxins should still be considered toxin

producers, since their ability to produce toxin in actual food poisoning outbreaks should not
be excluded (Schimitz et al., 1998; Veras et al., 2008).
As underlined by other authors (Ercolini et al., 2004), in case of food matrix, a PCR
detection of toxin encoding genes coupled with the specific detection of producing species,
indicates the potential to find toxins in the food and, depending on the level of
contamination, the possibility to have hazardous food products.
It is important to recognize that this technique only can be used identify strains harboring
the toxin genes and is independent of the expression and secretion of the toxin. To verify
toxin production by any given isolate, time- and labor-intensive immunological methods
may be used to detect the excreted toxins.
The literature shows very variable results concerning the occurrence of enterotoxigenic S.
aureus strains in foods. This is probably due to the differences among the kinds of
examined foods, number of samples, in detection methods used, and in the ecological
origin of strains (Holeckova et al., 2002).
SEA, SEB and SED are the most common enterotoxins recovered from food-poisoning
outbreaks in man, the significance of these novel toxins for public health is unclear
(Zschock et al., 2005). Therefore, an interesting development of multiplex PCR assay in
this study would be its application directly on milk and food samples, saving two days of
analysis. The molecular method developed for the first time in Iraq can be applied for rapid
detection of S. aureus bacteria in dairy milk samples.
This study revealed the wide spread presence of S. aureus strains that are carriers of genes
coding for toxins other than those identifiable with traditional methods. The real
significance of the presence of these toxins in food and their possible impact as a cause of
S. aureus food poisoning in humans is still unknown and further studies are required using
both traditional and molecular methods. The use of innoviate techniques for the
identification of genes coding for the production of the enterotoxins along with traditional
detection methods, would enable the identification of strains that carry genes that might, in
suitable conditions, produce as yet unknown toxins potentially capable of producing
sickness in humans.
The high percentage of positive samples of milk and cheese for staphylococcal enterotoxins
draw attention to the need to improve the management of animals at the farm level, in

particular through the implementation of specific monitoring programs designed to reduce
subclinical mastitis which is often the cause of milk contamination. An effective reduction
of contamination levels could be achieved by improving sanitation and hygiene procedures
(Giannatale et al., 2011).
3-5 Detection of Staphylococcal Enterotoxin Production
After the detection of staphylococcal enterotoxins encoding genes (sea, seb, sec, and
sed) by multiplex PCR technique and confirming the enterotoxigenic potential of
staphylococcal isolates from milk and cheese samples, and depending on the fact that SEs
inducing diarrhea and causes inflammatory changes in the gastrointestinal tract (Veras et
al., 2008; Hennekinne et al., 2010; Schelin et al., 2011; Xu and McCormick 2012), so that
the suckling mouse bioassay was tested to investigate the staphylococcal enterotoxin
biological activity.
Among the 200 staphylococcal isolates from milk and cheese samples, examined with
suckling mouse bioassay, 131 isolates which constitutes 65.5% of the examined isolates,
gave a positive result as depicted by the tables 3-13 and 3-14.
Table 3-13: The biological activity of the COPS enterotoxins in suckling mouse
bioassay represented by the intestine weight /body weight ratio Isolate IW/BW Isolate IW/BW 1 S 14 0.083 19 S 63 0.103 2 S 15 0.084 20 S 64 0.085 3 S 16 0.087 21 S 65 0.113 4 S 17 0.113 22 S 68 0.084 5 S 21 0.083 23 S 70 0.091 6 S 27 0.112 24 S 73 0.116 7 S 30 0.083 25 S 77 0.076 8 S 31 0.083 26 S 80 0.085 9 S 40 0.112 27 S 81 0.037 10 S 41 0.085 28 S 83 0.096 11 S 50 0.087 29 S 85 0.049 12 S 51 0.033 30 S 91 0.084 13 S 55 0.111 31 S 93 0.095 14 S 56 0.089 32 S 94 0.083 15 S 57 0.093 33 S 95 0.117 16 S 58 0.086 34 S 97 0.083

17 S 59 0.114 35 S 98 0.094 18 S 60 0.087 36 S 100 0.082 Isolate IW/BW Isolate IW/BW 37 S 102 0.115 68 S 174 0.084 38 S 103 0.085 69 S 177 0.094 39 S 108 0.116 70 S 180 0.087 40 S 109 0.113 71 S 185 0.096 41 S 111 0.068 72 S 191 0.086 42 S 112 0.044 73 S 194 0.106 43 S 113 0.037 74 S 195 0.113 44 S 117 0.088 75 S 196 0.085 45 S 119 0.110 76 S 198 0.109 46 S 120 0.092 77 S 199 0.083 47 S 121 0.083 78 S 200 0.094 48 S 125 0.086 79 S 203 0.088 49 S 130 0.085 80 S 204 0.091 50 S 132 0.099 81 S 205 0.115 51 S 133 0.090 82 S 206 0.087 52 S 136 0.084 83 S 207 0.109
53 S 140 0.100 84 S 208 0.085 54 S 144 0.116 85 S 214 0.115 55 S 145 0.105 86 S 215 0.091 56 S 147 0.054 87 S 217 0.110 57 S 150 0.088 88 S 219 0.089 58 S 155 0.112 89 S 220 0.084 59 S 156 0.063 90 S 223 0.105 60 S 157 0.086 91 S 227 0.084 61 S 161 0.099 92 S 228 0.083 62 S 163 0.083 93 S 230 0.094 63 S 164 0.096 94 S 232 0.093 64 S 165 0.115 95 S 233 0.086 65 S 168 0.112 96 S 234 0.090 66 S 171 0.024 97 S 235 0.087 67 S 173 0.083

All isolates were grown in TSB-YE medium for 48 h in shaker incubator, mouse incubation period 2 h
at 25oC. Intestine weight/body weight ratios of the control group ranged from 0 to 0.003.
Table 3-14: The biological activity of the CONS enterotoxins in suckling mouse bioassay represented by the intestine weight /body weight ratio
Isolate IW/BW Isolate IW/BW 1 S 1 0.042 33 S 45 0.026 2 S 2 0.087 34 S 46 0.039 3 S 3 0.079 35 S 47 0.098 4 S 4 0.051 36 S 48 0.067 5 S 5 0.092 37 S 49 0.083 6 S 7 0.037 38 S 52 0.013 7 S 8 0.075 39 S 53 0.065 8 S 9 0.088 40 S 54 0.086 9 S 10 0.005 41 S 61 0.038 10 S 11 0.039 42 S 62 0.085 11 S 12 0.046 43 S 66 0.021 12 S 13 0.099 44 S 67 0.034 13 S 18 0.048 45 S 69 0.028 14 S 19 0.089 46 S 71 0.053 15 S 20 0.079 47 S 72 0.047 16 S 22 0.013 48 S 74 0.096 17 S 23 0.097 49 S 75 0.082 18 S 24 0.089 50 S 76 0.091 19 S 25 0.084 51 S 78 0.093 20 S 26 0.057 52 S 79 0.089 21 S 28 0.082 53 S 82 0.017 22 S 29 0.057 54 S 84 0.087 23 S 33 0.088 55 S 86 0.050 24 S 34 0.028 56 S 87 0.083 25 S 35 0.046 57 S 88 0.056 26 S 36 0.085 58 S 89 0.109 27 S 37 0.106 59 S 90 0.080 28 S 38 0.017 60 S 92 0.028 29 S 39 0.083 61 S 96 0.035 30 S 42 0.091 62 S 99 0.043 31 S 43 0.040 63 S 101 0.087

32 S 44 0.100 64 S 104 0.032 Isolate IW/BW Isolate IW/BW 65 S 105 0.108 85 S 190 0.032 66 S 107 0.089 86 S 192 0.026 67 S 114 0.093 87 S 193 0.084 68 S 115 0.089 88 S 197 0.092 69 S 122 0.054 89 S 201 0.085 70 S 123 0.087 90 S 202 0.082 71 S 124 0.095 91 S 210 0.054 72 S 126 0.069 92 S 211 0.092 73 S 127 0.111 93 S 212 0.094 74 S 128 0.085 94 S 213 0.039 75 S 129 0.091 95 S 216 0.085 76 S 131 0.044 96 S 218 0.056 77 S 134 0.088 97 S 221 0.020 78 S 135 0.070 98 S 222 0.084 79 S 160 0.048 99 S 224 0.032 80 S 162 0.083 100 S 225 0.085 81 S 175 0.035 101 S 226 0.036 82 S 178 0.056 102 S 229 0.045 83 S 179 0.032 103 S 231 0.093 84 S 183 0.089 All isolates were grown in TSB-YE medium for 48 h in shaker incubator, mouse incubation period 2 h at 25oC. Intestine weight/body weight ratios of the control group ranged from 0 to 0.003. From the obtained results it was concluded that none of both COPS and CONS isolates that
did not possessed any of the tested enterotoxin genes gave a positive result when tested by
suckling mouse bioassay, the highest positive ratio of IW/BW was 0.116 that was the
biological activity of three COPS isolates: S108, S144 and S205 which possessed the
following SE encoding genes sea+seb+sed, sea+sed, and sea+seb+sed respectively,
regarding CONS the highest positive ratio of IW/BW was 0.111 that was the biological
activity of the isolate S127 followed by 0.109 IW/BW resulted by S89 then 0.108 IW/BW
by S105; all these three isolates possessed the SE encoding genes sea+sed, this may be
related to the synergic effect of the toxin genes together when the tested staphylococcal
isolates possessed two or three of them.
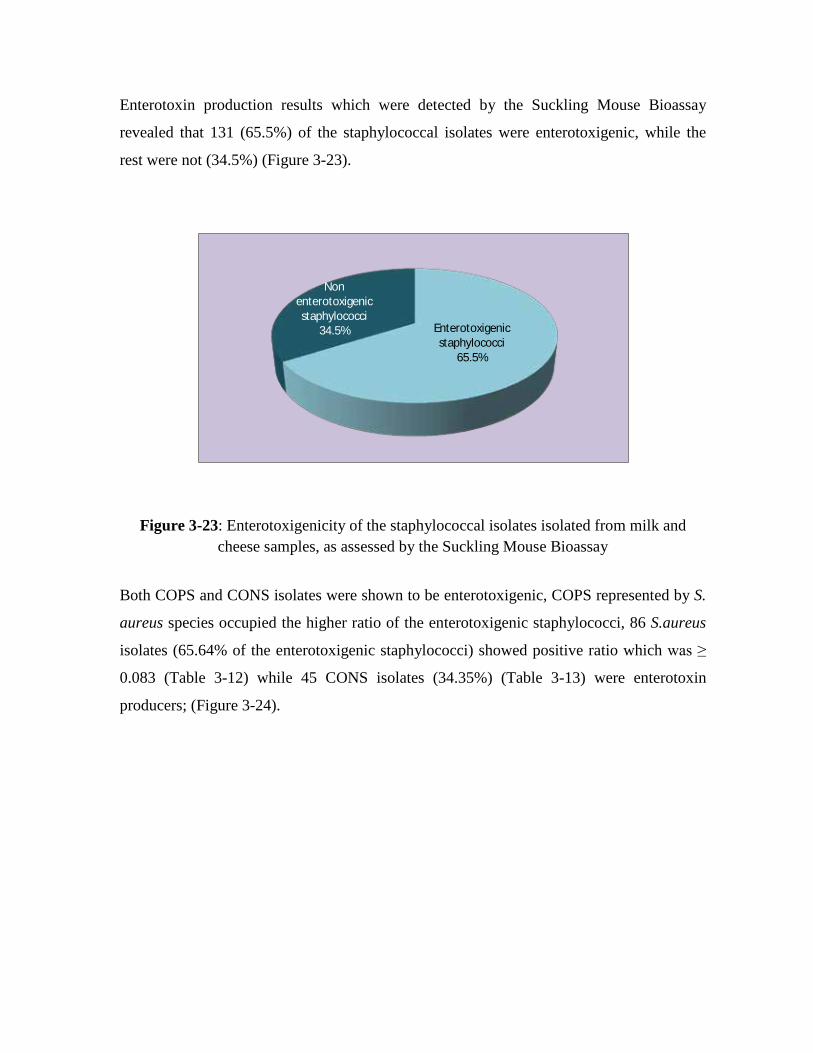
Enterotoxin production results which were detected by the Suckling Mouse Bioassay
revealed that 131 (65.5%) of the staphylococcal isolates were enterotoxigenic, while the
rest were not (34.5%) (Figure 3-23).
Figure 3-23: Enterotoxigenicity of the staphylococcal isolates isolated from milk and cheese samples, as assessed by the Suckling Mouse Bioassay
Both COPS and CONS isolates were shown to be enterotoxigenic, COPS represented by S.
aureus species occupied the higher ratio of the enterotoxigenic staphylococci, 86 S.aureus
isolates (65.64% of the enterotoxigenic staphylococci) showed positive ratio which was ≥
0.083 (Table 3-12) while 45 CONS isolates (34.35%) (Table 3-13) were enterotoxin
producers; (Figure 3-24).
Enterotoxigenic staphylococci
65.5%
Non enterotoxigenic staphylococci
34.5%

Figure 3-24: Diversity of Results in the biological test between coagulase-positive and coagulase-negative staphylococcal isolates
The staphylococcal enterotoxins, not the bacterium, settles in the small intestine and cause
inflammation and swelling, this in turn can cause abdominal pain, cramping, dehydration,
diarrhea and fever, SEs act as superantigens, binding to MHC II molecules and stimulating
T cells to divide and produce lymphokines such as IL-2 and TNF-alpha, which induces
diarrhea (Pinchuk et al., 2010; Rao, 2006).
Assessment of staphylococcal isolates ability to produce enterotoxins was conducted by the
use of suckling mouse bioassay, this test was devised initially by Dean et al.(1972) and
modified by Guarino et al.(1989), it was meant to assess the ability to produce enterotoxins
by E. coli isolates, and after slight modifications it is still used as done by Kisiel et
al.(2011) to assess the ability of Y.enterocolitica isolates to produce enterotoxins.
It had been reported that the orally and intragastrically introduction of SEA and SEB to
laboratory animals induced emetic responces, diarrhea and GI inflammatory changes
(Benjamin et al., 1998).
The effect of heat on staphylococcal enterotoxins activity in suckling mice was detected; it
had been applied on 4 Staphylococcal isolates.
0
20
40
60
80
100
COPS CONS
86
45
2
28
9
30
Positive Doubtful Negative

Among all the examined crude preparations of ST, none were affected by heating at 65 Co
for 15 min and the same result was observed when the preparations of ST heated to 100 Co
for 30 min (Table 3-15).
Table 3-15: Effect of heat on staphylococcal enterotoxin activity
Staphyloco--
ccal isolates
Toxin Activity
IW/BW
No heat
mean ± SE
Toxin Activity
IW/BW
65Co, 15 min
mean ± SE
X2-Test
value
Toxin Activity
IW/BW
100Co, 30 min
mean ± SE
X2-Test
value
S 95 (0.117 ± 0.0003) (0.117 ± 0.0012) 0.229 a (0.116± 0.0012) 0.918 **
S 14 (0.083 ± 0.0015) (0.085 ± 0.0035) 0.423 a (0.084± 0.0010) 0.866 **
S 84 (0.087 ± 0.0058) (0.091 ± 0.0064) 0.326 a (0.091± 0.0081) 0.293 **
S 37 (0.106 ± 0.0025) (0.104 ± 0.0055) 0.569 a (0.104 ± 0.0015) 0.622 **
All isolates were grown in TSB-YE medium for 48 h in shaker incubator, mouse incubation period 2 h
at 25oC all values are mean.
**: non significant statically.
Statically analysis showed that there were no significant differences among the tested
groups; the toxic activity has not been affected by applying heat even at 100oC for 30 min.
these results confirmed that these toxins are thermostable staphylococcal enterotoxins.
Kisiel et al. (2011) revealed that there was no influence of high temperature on the activity
of the examined enterotoxins produced by Yersinia enterocolitica; even after heating to
65oC for 15 min.
The detection of S. aureus and SEs in food is often difficult because food processing may
kill the bacteria without destroying SEs, which are stable to high temperature and to
inactivation by gastrointestinal proteases such as pepsin (Cremonesi et al., 2007).
Comparing the results obtained by multiplex PCR assay detecting four classical enterotoxin
coding genes, with those obtained by suckling mouse bioassay concerning the phenotypic
expression of enterotoxin coding genes, 153 (76.5%) staphylococcal isolates harboured one
or more enterotoxin coding genes, while the suckling mouse bioassay revealed that 131
(65.5%) of these isolates produced detectable amounts of enterotoxins (Table 3-16).

Table 3-16: Comparison of the genotypic content of enterotoxigenic staphylococcal isolates of sea, seb, sec, and sed and the phenotypic expression of enterotoxicity
(detected by Suckling Mouse Bioassay)
Staphylococcal isolates
Enterotoxigenicity detected by Suckling
Mouse Bioassay
Enterotoxigenicity detected by multiplex
PCR
X2-Test value
197 COPS 86 (65.64%) 95 (62.09%) 0.448**
103 CONS 45 (34.35%) 58 (37.90%) 1.641**
**: non significant (P>0.05)
It is obviously from the table (3-15) that the genotypic content was higher than the
phenotypic expression of the tested enterotoxin coding genes (sea, seb, sec, and sed). There
was high correlation between the results of the two methods (PCR and the bioassay).
Although the PCR technique is more specific than the suckling mouse bioassay in detecting
the staphylococcal enterotoxigenicity, but statically no significant differences detected
between the genotypic and phenotypic detection techniques.
Comparing the PCR and the bioassay data, some isolates were shown to be PCR-positive
for enterotoxin gene but did not evidence detectable production of enterotoxin. Similar
findings have been reported by other investigators (Schmitz et al., 1998; Cunha et al.,
2006).
This is possibly because; the toxin gene was expressed and the toxin is produced below the
detection limit of the bioassay used, or the toxin gene had been inactivated by point
mutation with insertion of stop codon, other factors for in vitro toxin production may were
absent.

Conclusions
&
Recommendation

Conclusions From the results of this study, several conclusions can be listed as the following:
1- Presence of high number of COPS and CONS in milk and cheese samples.
2- The results provide evidence that the presence of antimicrobial resistant strains of
Staphylococcus species to the antibiotics: methicillin, tetracycline and vancomycin, had
become remarkably widespread in food samples.
4- Effectiveness of using the genetic determinants of methicillin resistance femA and mecA
genes together in the detection of methicillin resistant (mecA+) and susceptible (lacking
mecA) staphylococci and in differentiating S. aureus (femA+) from (mecA+) coagulase
negative staphylococci (lacking femA).
5- This study demonstrated that both COPS and CONS staphylococcal isolates from milk
and cheese samples, possessed various toxin genes.
6- Prevalence of sea and sed genes among the isolated staphylococci, and the dearth of sec
gene; it was harboured by two isolates of CONS only.
Recommendations
1- Application of this approach on other microorganisms especially other gram positive
bacteria, since the DNA extraction from such cells being no more difficult.
2- Using multiplex PCR assay in detecting the presence of newly described staphylococcal
enterotoxin genes, because there is no available immunoassay to detect them because of the
difficulty to purify them.
3- Depending the direct detection of egc in raw materials can provide rapid results and
highlights the presence of loads of staphylococci potentially representing risk of
intoxication.
4- Study the role of Staphyloxanthin in staphylococci pathogenesis.

5- Further studies for characterization of enterotoxigenic potential of CONS in foods were
need.
6- Using coa gene polymorphism for typing and differentiation of S. aureus isolates that
locally isolated from various foods and food handlers.
7- The molecular procedures described in this study was especially developed to fit into the
daily work requirement of routine food quality control laboratories since detection and
identification of the toxin genes of a pathogen from diverse food sources is becoming an
important component of the diagnostic inventory of such laboratories.
8- The enterotoxin genes from enterotoxigenic staphylococci continue to constitute a
potential risk for consumers’ health, therefore, not only enterotoxin production, but also
detection of the enterotoxin genes must be taken into account for all food-borne
staphylococcal isolates.
9- The presence of such high percentage of methicillin resistant staphylococci among
COPS and CONS isolates from milk and cheese samples confirms the need for strict
control measurments to be applied from farm to fork.
10- Vigilant food safety practice needs to be implemented for people who handle raw milk
and cheese to prevent food-borne infections and intoxications due to staphylococcal
contamination, and the rapid and efficient detection of S. aureus isolates from dairy
products is essential for consumer safety.
11- Sanitary strategies must be designed to control the apparently widespread
contamination of milk and cheese with staphylococci, and thereby reducing the associated
economical losses and public health problems.

References

A § Aarestrup, F.M.; Wegener, H.C.; Rosdahl, V.T.; Jensen, N.E. (1995).
Staphylococcal and other bacterial species associated with intramammary infections in Danish dairy herds. Acta Vet. Scand. 36:475-487.
§ Abd-Elateef, A. N. (2011). Comparative Study between Local Isolates of Methicillin Sensitive and Resistance of Staphylococcus epidermidis. MSc. Thesis. College of Science. University of Baghdad.
§ Akineden, O.; Annemuller, C.; Hassan, A. A.; Lammler, C.; Wolter, W. and Zschock, M. (2001). Toxin genes and other characteristics of Staphylococcus aureus isolates from milk of cows with mastitis. Clin Diagn Lab Immunol 8:959–964.
§ Al-Daccak, R.; Mehindate, K.; Damdoumi, F.; Etongue-Mayer, P.; Nilsson, H.; Antonsson, P.; Sundstrom, M.; Dohlsten, M.; Sekaly, R.P. and Mourad, W. (1998). Staphylococcal enterotoxin D is a promiscuous superantigen offering multiple modes of interactions with the MHC classII receptors. J. Immunol. 160:225–232.
§ Alegro, A. L.C.; Konta, E. M.; Suzuki, K.; Silva, M.G. Junior, A.F. Rall, R. and Mores, R.V.L. (2007). Occurrence of coagulase-positive Staphylococcus in various food products commercialized in Botucatu, SP, Brazil and detection of toxins from food and isolated strains. Food Control. 18:630-634.
§ Al-Maliki, A.A.A. (2009). A Study of Some Methicillin Resistant Staphylococci (MRSA) and (MRSE) Isolated from Baghdad Hospital Patients. MSc. Thesis. College of Science. Al-Mustansiriya University.
§ Al-Shamary, R. (2012). Inhibition of lipase produced from Staphylococcus aureus isolated from acne by Lactobacillus bacteriocins. MSc. Thesis. College of Science. University of Baghdad.
§ Anderson, J. E.; Beelman, R.R. and Doores, S. (1996). Persistance of serological and biological activities of staphylococcal enterotoxin A in canned mushrooms. J. Food Prot. 59:1292-1299.
§ Argudin, M. A.; Mendoza, M. C. and Rodicio, M. R. (2010). Food Poisoning and
Staphylococcus aureus Enterotoxins. Toxins 2: 1751-1773.
§ Asao, T.; Kumeda, Y.; Kawai, T.; Shibata, T.; Oda, H.; Haruki, K.; Nakazawa, H.; Kozaki, S. (2003). An extensive outbreak of staphylococcal food poisoning due to

low-fat milk in Japan: estimation of enterotoxin A in the incriminated milk and powdered skim milk. Epidemiol. Infect. 130: 33–40.
§ Aslantas, O.; Demir, C.; Turutoglu, H.; Cantekin, Z.; Ergun, Y. and Dogruer, G. (2007). Coagulase gene polymorphism of Staphylococcus aureus isolated from subclinical bovine mastitis. Turk. J. Vet. Anim. Sci. 31(7):253-257.
§ Assenmacher, M.; Lohning, M.; Scheffold, A.; Manz, R.A.; Schmitz, J.; and Radbruch, A. (1998). Production of IL-2, IFN-Gamma and IL-10 by individual staphylococcal enterotoxinn B-activated T helper lymphocytes. Eur. J. Immunol. 28: 1534–1543.
§ Ateba, C. N.; Mbew, M.; Moneoang, M. S. and Bezuidenhout, C.C. (2010). Antibiotic resistant Staphylococcus aureus isolated from milk in the Mafikeng area, North West province, South Africa. S. Afr. J. Sci. 106:1-6.
§ Atlas, R. M. (2005). Handbook of media for environmenatal microbiology.2nd ed. Tylor & Francis Group, LIC. USA.
§ Atlas, R. M. and Snyder, J. W. (2006). Handbook of media for clinical microbiology. CRC Press.
B § Baba, T.; Bae, T.; Schneewind, O.; Takeuchi, F.; and Hiramatsu K. (2008).
Genome sequence of Staphylococcus aureus strain Newman and comparative analysis of staphylococcal genomes: polymorphism and evolution of two major pathogenicity islands. J. Bacteriol. 190: 300–310.
§ Baird-Parker, A. C. (1962). An improved diagnostic and selective medium for isolating coagulase positive staphylococci. Journal of Applied Bacteriology. 25:12-19.
§ Balaban, N. and Rasooly, A. (2000). Staphylococcal enterotoxins. Int. J. Food Microbiol. 61(1): 1-10.
§ Bannerman, T. L. and Kloos, W. E. (1991). Staphylococcus capitis subsp.
ureolyticus subsp. nov. from human skin. Int. J. Syst. Bacteriol. 41: 144-147.
§ Bania, J.; Dabrowska, A.; Bystron, J.; Korzekwa, K.; Chrzanowska, J.; and Molenda, J. (2006). Distribution of newly described enterotoxin-like genes in Staphylococcus aureus from food. Int. J. Food Microbiol. 108: 36–41.

§ Bartolomeoli, I.; Maifreni, M.; Frigo, F.; Urli, G. and Marino, M. (2009). Occurrence and characterization of Staphylococcus aureus isolated from raw milk for cheese making. Int. J. Dairy Technol. 62:366-371.
§ Baruzzi, F.; Matarante, A.; Caputo, L. and Morea, M. (2005). Development of a culture-independent PCR-based assay for the detection of Lactobacilliin stretched cheese. J. Rapid Meth Aut Mic. 13:177-192.
§ Bautista, L.; Gaya, P.; Medina, M. and Nunes, M. (1988). A Quantitative study of enterotoxin production by sheep milk staphylococci. Applied and Environmental Microbiology. 45: 566-569.
§ Bayles, K.W. and Iandolo, J.J. (1989). Genetic and molecular analyses of the gene encoding staphylococcal enterotoxin D. J. Bacteriol. 171: 4799-4806.
§ Benjamin, M.A.; Lu, J.; Donnelly, G.; Dureja, P.; McKay, D.M. (1998). Changes in murine jejunal morphology evoked by the bacterial superantigen Staphylococcus aureus enterotoxin B are mediated by CD4+ T cells. Infect. Immun. 66: 2193–2199.
§ Benson, J.M. (2001). Microbiological Applications: Laboratory Manual in General
Microbiology. 8th ed. McGraw Hill.
§ Bergdoll, M.S. (1983). Enterotoxins. In Staphylococci and Staphylococcal Infections; Easman, C.S.F., Adlam, C., Eds.; Academic Press Inc: London, UK. 2:559–598.
§ Bergdoll, M. S. (1990). Analytical methods for Staphylococcus aureus. International Journal of Food Microbiology. 10: 91-100.
§ Bergdoll, M. S. (1995). Importance of staphylococci that produce nanogram quantities of enterotoxin. Zbl Bakt. 282:1-6.
§ Bergdoll, M. S. and Wong, A.C.L. (2006). Foodborne Intoxications. Elsevier Inc.14.
§ Boisset S, Geissmann T, Huntzinger E, Fechter P, Bendridi N, Possedko M. (2007).
Staphylococcus aureus RNAIII coordinately represses the synthesis of virulence factors and the transcription regulator Rot by an antisense mechanism. Genes Dev. 21:1353-66.
§ Blaiotta, G.; Ercolini, D.; Pennacchia, C.; Fusco, V.; Casaburi, A.; Pepe, O. and Villani, F. (2004). PCR detection of staphylococcal enterotoxin genes in

Staphylococcus spp. strains isolated from meat and dairy products. Evidence for new variants of seG and seI in S. aureus AB-8802. J. Appl. Microbiol. 97: 719–730.
§ Boyce, J.M. and Havill, N.L. (2005). Comparison of BD Gene Ohm methicillin-resistant Staphylococcus aureus (MRSA) PCR versus the CHROMagar MRSA assay for screening patients for the presence of MRSA strains. J. Clin. Microbiol. 46 (1): 350-351.
§ Brooks, G. F.; Caroll, K. C. and Morse, S. A. (2007). Staphylococcus (Jawetz). Melnick and Adelberg’s, Medical Microbiology. 24th edition. The McGraw-Hill. New York.
§ Bukharie, H.A. (2010). Increasing threat of community-acquired methicillin-resistant Staphylococcus aureus. Am. J. Med. Sci. 340: 378-381.
§ Buonpane, R.A.; Churchill, H.R.; Moza, B.; Sundberg, E.J.; Peterson, M.L.;
Schlievert, P.M. and Kranz, D.M. (2007). Neutralization of Staphylococcal Enterotoxin B by Soluble, High-Affinity Receptor Antagonists. Nat. Med 13: 725–729.
§ Byrne, B.; Madrigal-Estebas, L.; McEvoy, A.; Carton, J.; Doherty, D.G.; Whelan, A.; Feighery, C.; O'Donoghue, D.P.; and O'Farrelly, C. (2002). Human duodenal epithelial cells constitutively express molecular components of antigen presentation but not costimulatory molecules. Hum. Immunol. 63: 977–986.
§ Bystron, J.; Bania, J.; Lis, E.; Molenda, J. and Bednarski, M. (2009). Characterization of Staphylococcus aureus strains isolated from cows’ milk. Bull Vet Inst Pulawy. 53: 59-63.
C
§ Carroll, K. C. (2008). Rapid Diagnostics for Methicillin-Resistant Staphylococcus aureus: Current Status. Molecular Diagnosis & Therapy. 12 (1):15-24.
§ Cha, J.O.; Lee, J.K.; Jung, Y.H.; Yoo, J.I.; Park, Y.K.; Kim, B.S. and Lee, Y.S. (2006). Molecular analysis of Staphylococcus aureus isolates associated with staphylococcal food poisoning in South Korea. J. Appl. Microbiol. 101: 864–871.
§ Chaffer, M.; Leitner, G.; Winkler, M.; Glickman, A.; Krifucks, O.; Ezra, E. and Saran, A. (1999). Coagulase negative staphylococci and mammary gland infections in cows. J. Vet. Med. B 46:707-712.

§ Chen, T.R.; Chiou, C.S. and Tsen, H.Y. (2004). Use of novel PCR primers specific to the genes of staphylococcal enterotoxin G, H, I for the survey of Staphylococcus aureus strains isolated from food-poisoning cases and food samples in Taiwan. Int. J. Food Microbiol. 92: 189–197.
§ Cheung A.L., Koomey J.M., Butler C.A., Projan S.J., and Fischetti V.A. (1992). Regulation of exoprotein expression in Staphylococcus aureus by alocus (sar) distinct from agr. Proc. Nati. Acad. Sci. USA 89: 6462-6466.
§ Chiang, Y.C.; Liao, W.W.; Fan, C.M.; Pai, W.Y.; Chiou, C.S.; and Tsen, H.Y. (2008). PCR detection of staphylococcal enterotoxins (SEs) N, O, P, Q, R, U, and survey of SE types in Staphylococcus aureus isolates from food-poisoning cases in Taiwan. Int. J. Food Microbiol. 121: 66–73.
§ Chikkala, R.; George, N. O.; Ratnakar, K. S. R.; Iyer, N. and Sritharan, V. (2012). Heterogeneity in femA in the Indian isolates of Staphylococcus aureus limits its usefulness as a species specific marker. Advances in Infectious Diseases 2: 82-88.
§ Collee, J. G.;Fraser, A.G.; Marmison, B.P. and Simpson, S. A. (1996). Mackie and
MaCartneys. Practical medical microbiology. 14th ed. Churchill Livingstone, U.K.
§ Collery M.M.; Smyth, D.S.; Tumilty, J.J.; Twohig, J.M.; and Smyth, C.J. (2009). Associations between enterotoxin gene cluster types egc1, egc2 and egc3, agr types, enterotoxin and enterotoxin-like gene profiles, and molecular typing characteristics of human nasal carriage and animal isolates of Staphylococcus aureus. J. Med. Microbiol. 58: 13–25.
§ Cooper, B.S.; Medley, G.F.; Stone, S.P.; Kibbler, C.C.; Cookson, B.D.; Roberts, J.A.; Duckworth, G.; Lai, R.; and Ebrahim, S. (2004). Methicillin-Resistant Staphylococcus aureus in Hospitals and the Community: Stealth Dynamics and Control Catastrophes. Proc. Natl. Acad. Sci. USA. 101: 10223–10228.
§ Coton, E.; Desmonts, M.H.; Leroy, S.;Coton, M. and Jamet, E. (2010). Biodiversity of coagulase-negative staphylococci in French cheese, dairy fermented sausages, processing environments and clinical samples. Int. J. Food Microbiol. 137:221-229.
§ Cremonesi, P.;Perez, G.;Pisoni, P.;Moroni, P.; Morandi, S.;Luzzana, M.; Brasca, M. and Castiglioni, B. (2007). Detection of enterotoxigenic Staphylococcus aureus isolates in raw milk cheese. Letters in Applied Microbiology.45: 586-591.

§ Cunha, M. L. R. S.; Rugolo, L. M. S. S. and Lopes, C. A. M. (2006). Study of virulence factors in coagulase-negative staphylococci isolated from newborns. Mem. Inst. Oswaldo Cruz. 101: 661-668.
§ Cunha, M. L. R. S. (2009a). Pathogenesis of Staphylococcus aureus and Bovine
Mastitis. Int. J. Med. Biol. Frontiers. 15: 1031-1042.
§ Cunha, M. L. R. S. (2009b). Catheter-Related Infections with Emphasis on Pathogenesis and Diagnosis. Int. J. Med. Biol. Frontiers 15: 1021-1030.
§ Cunha, M. L. R. S. and Calsolari, R. A. O. (2007). Toxigenecity in Staphylococcus with emphasis on coagulase-negative staphylococci. Communicating Current Research and Educational Topics and Trends in Applied Microbiology. 778-782.
§ Chung, M.C.; Wines, B.D.; Baker, H.; Langley, R.J.; Baker, E.N.; and Fraser, J.D. (2007). The crystal structure of staphylococcal superantigen-like protein 11 in complex with sialyl Lewis X reveals the mechanism for cell binding and immune inhibition. Mol. Microbiol. 66: 1342–1355.
D § Dack, G.M.; Cary, W.E.; Woolpert, O. and Wiggins, H.J. (1930). An outbreak of
food poisoning proved to be due to a yellow haemolytic Staphylococcus. J. Prev. Med. 4: 167–175.
§ Dastmalchi, H.; Ahmadi, M.; Mardani, K. and Batavani, R.A. (2009). Molecular typing of Staphylococcus aureus isolated from bovine mastitis based on polymorphism of the coagulase gene in the north west of Iran. Vet. Microbiol. 137:202-206.
§ Dean, A. G.; Ching, Y.C.; Williams, R.G. and Harden, B.L. (1972). Test for Escherichia coli Enterotoxin Using Infant Mice: Application in a Study of Diarrhea in Children in Honolulu. J. Infect. Dis. 125 (4): 407-411.
§ De Baere, T.; de Mendonça, R.;Claeys,G. Verschraegen,G.; Mijs,W.; Verhelst,R.; Rottiers,S. Van Simaey, L.; De Ganck, C. and Vaneechoutte, M.(2002). Evaluation of amplified rDNA restriction analysis (ARDRA) for the identification of cultured mycobacteria in a diagnostic laboratory. BMC Microbiology. 2:4.
§ Demir, C.; Aslantas, O.; Duran, N.; Ocak, S. and Ozer, B.(2011). Investigation of toxin genes in Staphylococcus aureus strains isolated in Mustafa Kemal University Hospital. Turk J Med Sci. 41 (2): 343-352.

§ Deresinski, S., (2007). Counterpoint: Vancomycin and Staphylococcus aureus-An antibiotic enters obsolescence. Clin. Infect. Dis. 44: 1543-1548.
§ Derzelle, S.; Dilasser, F.; Duquenne, M. and Deperrois, V. (2009). Differential temporal expression of the staphylococcal enterotoxins genes during cell growth. Food Microbiol. 26: 896–904.
§ Desouza, I.A.; Hyslop, S.; Franco-Penteado, C.F.; and Ribeiro-DaSilva, G. (2002). Evidence for the Involvement of a Macrophage-Derived Chemotactic Mediator in the Neutrophil Recruitment Induced by Staphylococcal Enterotoxin B in Mice. Toxicon. 40: 1709–1717.
§ Devriese, L.A., De Keyser, H. (1980). Prevalence of different species of coagulase-
negative staphylococci on teats and in milk samples from dairy cows. J. Dairy Res. 47:155-158.
§ Devriese, L. A., Vancanneyt, M., Baele, M., Vaneechoutte, M., De Graef, E., Snauwaert, C., Cleenwerck, I., Dawyndt, P., Swinges, J., Decostere, A. and Haesebrouck, F. (2005). Staphylococcus pseudintermedius sp. nov., a coagulase-positive species from animals. Int. J. Syst. Evol. Microbiol. 55: 1569-1573.
§ Devriese, L. A., Poutrel, B., Kilpper-Balz, R. and Schleifer, K. F. (1983). Staphylococcus gallinarum and Staphylococcus caprae, two new species from animals. Int. J. Syst. Bacteriol. 33: 480-486.
§ Dinges, M.M. Orwin, P.M. and Schlievert, P.M. (2000). Exotoxins of
Staphylococcus aureus. Clin. Microbiol. 13: 16-34.
§ Dubey, R.C. and Maheshwari, D. K. (2009). A textbook of Microbiology. 1st ed. S. Chand and Company LTD. New Delhi.
§ Duquenne M, Fleurot I, Aigle M, Darrigo C, Borezée-Durant E, Derzelle S. (2010). Tool for quantification of staphylococcal enterotoxin gene expression in cheese. Appl Environ Microbiol. 76:1367-74.
§ Duran, N.; Ozer, B.; Duran, G. G.; Onlen, Y. and Demir, C. (2012). Antibiotic
resistance genes & susceptibility patterns in staphylococci. Indian. J Med Res. 135: 389-396.

E § EFSA. (2011). The European Union Summary Report on Trends and Sources of
Zoonoses, Zoonotic Agents and food-borne outbreaks in 2009. EFSA J; 2090.
§ Efuntoye, M. O.; Bakare, A. A. and Sowunmi A. A. (2011). Virulence factors and antibiotic resistance in Staphylococcus aureus and Clostridium perfringens from landfill leachate. African Journal of Microbiology Research. 5(23): 3994-3997.
§ El-Jakee, J. A.; Nagwa, S.; El-Said, G. W.A.; Bakry, M.A; Samy, A.A.; Khairy, E.A.; Elgabry, E.A. (2010). Diversity of Staphylococcus aureus Isolated from Human and Bovine Estimated by PCR - Gene Analysis. Journal of American Science. 6(11):487-498.
§ Ercolini, D. (2004). PCR-based detection of enterotoxigenic Staphylococcus aureus in the early stages of the raw milk cheese making. Journal of Applied Microbiology. 96: 1090-1096.
§ El-Seedy, F.R.; El-Shabrawy, M.; Hakim, A.S.; Syame, S.F. and Osman, N.M. (2012). Advanced Techniques Used for Isolation and Characterization of Staphylococcus aureus Isolated from Mastitic Buffaloes. Global Veterinaria 8 (2): 144-152.
§ Euzéby, J.P. (2010). List of Prokaryotic names with Standing in Nomenclature.Genus Staphylococcus. http://www.bacterio.cict.fr/s/Staphylococcus.html.
§ Even, S.; Leroy, S.; Charlier, C.; Zakour, N.B. and Chacornac, J. P. (2010). Low occurrence of safety hazards in coagulase negative staphylococci isolated from fermented foodstuffs. Int. J. Food Microbiol. 139: 87-95.
§ Evenson, M.L.; Hindes, M. W. and Bernstein, R. S. (1988). Estimation of human dose of staphylococcal enterotoxin A from a large outbreak involving chocolate milk. International Journal of Food Microbiology. 7:311-316.
F § Faria, C.; Vas-Moreira, I.; Serapicos, E.; Nunes, O. C. and Manaia, C.M. (2009).
Antibiotic resistance in coagulase negative staphylococci isolated from waste water and drinking water. Sci. Total Environ. 407:3876-3882.

§ Ferens, W. A.; Davis, W. C.; Hamilton, M. J.; Park, Y. H.; Deobald, C. F.; Fox, L. and Bohach, G. (1998). Activation of bovine lymphocyte subpopulations by staphylococcal enterotoxin C. Infect. Immun. 66:573–580.
§ Fernandez, M.M.; Guan, R.; Swaminathan, C.P.; Malchiodi, E.L.; and Mariuzza, R.A. (2006). Crystal Structure of Staphylococcal Enterotoxin I (SEI) in Complex with a Human Major Histocompatibility Complex Class II Molecule. J. Biol. Chem. 281: 25356–25364.
§ Fernández, M.M.; Bhattacharya, S.; De Marzi, M.C.; Brown, P.H.; Kerzic, M.; Schuck, P.; Mariuzza, R.A. and Malchiodi, E.L. (2007). Superantigen natural affinity maturation revealed by the crystal structure of staphylococcal enterotoxin G and its binding to T-cell receptor Vbeta8.2. Proteins. 68: 389–402.
§ Fischer, H.; Dohlsten, M.; Lindvall, M.; Sjogren, H.O.; and Carlsson, R. (1989). Binding of Staphylococcal Enterotoxin A to HLA-DR on B Cell Lines. J. Immunol. 142: 3151–3157.
§ Fitzgerald, J.R.; Monday, S.R.; Foster, T.J.; Bohach, G.A.; Hartigan, P.J.; Meaney, W.J.; and Smith, C.J. (2001). Characterization of putative pathogenicity island from bovine Staphylococcus aureus encoding multiple superantigens. J. Bacteriol., 183, 63–70.
§ Fooladi, I. A. A.; Tavakoli, H.R.; Nadaeri, A. (2010). Detection of enterotoxigenic Staphylococcus aureus isolates in domestic dairy products. Iranian Journal of Microbiology. 2 (3): 135-140.
§ Fraser, J.D. and Proft, T. (2008). The bacterial superantigen and superantigen-like proteins. Immunol. Rev. 225: 226–243.
§ Freeman, D.J.; Falkiner, F.R. and Keane, C.T. (1989). New Method for detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 42:872-874.
§ Fromont, G.; Cerf-Bensussan, N.; Patey, N.; Canioni, D.; Rambaud, C.; Goulet, O.; Jan, D.; Revillon, Y.; Ricour, C.; and Brousse, N. (1995). Small Bowel Transplantation in Children: an Immunohistochemical Study of Intestinal Grafts. Gut 37: 783–790.
§ Fueyo, J.M.; Mendoza, M.C.; Alvarez, M.A.; and Martín, M.C. (2005). Relationships between toxin gene content and genetic background in nasal carried

isolates of Staphylococcus aureus from Asturias, Spain. FEMS Microbiol. Lett. 243: 447–454.
§ Fujimoto, D.F.; Higginbotham, R.H.; Sterba, K.M.; Maleki, S.J. Segall, A.M. and Smeltzer, M.S. (2009). Staphylococcus aureus SarA is a regulatory protein responsive to redox and pH that can support bacteriophage lambda integrasemediated excision/recombination. Mol Microbiol . 74:1445-58.
§ Fujisawa, N.; Hayashi, S.; Kurdowska, A.; Noble, J.M.; Naitoh, K.; and Miller, E.J. (1998). Staphylococcal Enterotoxin A-Induced Injury of Human Lung Endothelial Cells and IL-8 Accumulation Are Mediated by TNF-Alpha. J. Immunol. 161: 5627–5632.
§ Fusco, V.; Quero, G.M.; Morea, M.; Blaiotta, G. and Visconti, A. (2011). Rapid and reliable identification of Staphylococcus aureus harbouring the enterotoxin gene cluster (egc) and quantitative detection in raw milk by real time PCR. International Journal of Food Microbiology 144:528-537.
G § Gbaguidi-Haore, H.; Thouverez, M.; Couetdic, G.; Cholley, P.; Talon, D.; and
Bertrand, X. (2009). Usefulness of Antimicrobial Resistance Pattern for Detecting PVL- or TSST-1-Producing Meticillin-Resistant Staphylococcus aureus in a French University Hospital. J. Med. Microbiol. 58: 1337–1342.
§ Gencay, Y. E.; Ayaz, N. D. and Dugro, A. K. (2010). Enterotoxin gene profiles of Staphylococcus aureus and other staphylococcal isolates from various foods and food ingredients. J Fac Vet Med Univ Erciyes. 7(2): 75-80.
§ Genigeorgis, C.A. (1989). Present state of knowledge on staphylococcal intoxication. Int J Food Microbiol. 9:327-60.
§ Giannatale, E., D. Prencipe, V.; Tonelli, A.; Marphoglia, C. and Migliorati, G. (2011). Characterization of Staphylococcus aureus strains isolated from food for human consumption. Veterinaria Italiana. 47: 165-173.
§ Glasel, J. A. (1997). Validity of nucleic acid purities monitored by 260 nm/280 nm absorbance ratios. Bio Techniques 18:3-62.
§ Giannouli, S.; Labrou, M. and Kyritsis, A. (2010). Detection of Mutations in the FemXAB Protein Family in Oxacillin-Susceptible mecA-Positive Staphylococcus aureus clinical Isolates. Journal of Antimicrobial Chemotherapy. 65 (4): 626-633.

§ Goerke, C.; Pantucek, R.; Silva Holtfreter, S.; Berit Schulte, B.; Manuel Zink, M.;
Grumann, D.; Bröker, B.M.; Doskar, J.; and Wolz, C. (2009). Diversity of prophages in dominant Staphylococcus aureus clonal lineages. J. Bacteriol. 191: 3462–3468.
§ Goh, S.H.; Byrne, S.K.; Zhang, J.L. (1992). Molecular typing of Staphylococcus aureus on the basis of coagulase gene polymorphisms. J. Clin. Microbiol. 30: 1642-1645.
§ Grumann, D.; Scharf, S.S.; Holtfreter, S.; Kohler, C.; Steil, L.; Engelmann, S.; Hecker, M.; Volker, U.; and Broker, B.M. (2008). Immune Cell Activation by Enterotoxin Gene Cluster (Egc)-Encoded and Non-Egc Superantigens from Staphylococcus aureus. J. Immunol.181: 5054–5061.
§ Guarino, A.; Alessio, M.; Tarallo, L.; Fontana, K.; Lacono, S. and Cosali, G. (1989). Heat stable enterotoxin produced by Escherichia coli in acute diarrhoea. Arch. Dis. Child. (ADC). 64:808-813.
§ Gundogan, N.; Citak, S. and Turan, E. (2006). Slime production, DNAse activity
and antibiotic resistance of Staphylococcus aureus isolated from raw milk, pasteurised milk and ice cream samples. Food Control .17:389-392.
H
§ Harley, J. p. and Prescott, L. M. (2002). Laboratory Exercises in Microbiology. 5th ed. McGraw Hill.
§ Harvey, J.; Paterson, J.T. and Gibs, P.A. (1982). Enterotoxigenicity of Staphylococcus aureus strains isolated from poultry: raw poultry carcasses as a potential food poisoning hazard. J. Appl. Bacteriol. 52: 251-258.
§ Hauschild, T., Stepanovic, S. and Zakrzewska-Czerwinska, J.(2010). Staphylococcus stepanovicii sp. nov., a novel novobiocin-resistant oxidase-positive staphylococcal species isolated from wild small mammals. Syst. Appl. Microbiol. 33:183-187.
§ Havelaar, A.H.; Brul, S.;de Jong, A.; de Jonge, R.; Zwietering, M.H. and Ter Kuile, B.H. (2010). Future challenges to microbial food safety. Int J Food Microbiol. 139:79-94.

§ Hayworth, J.L.; Kasper, K.J.; Leon-Ponte, M.; Herfst, C.A.; Yue, D.; Brintnell, W.C.; Mazzuca, D.M.; Heinrichs, D.E.; Cairns, E.; Madrenas, J.; Hoskin, D.W.; McCormick, J.K.; and Haeryfar, S.M. (2009). Attenuation of Massive Cytokine Response to the Staphylococcal Enterotoxin B Superantigen by the Innate Immunomodulatory Protein Lactoferrin. Clin. Exp. Immunol. 157: 60–70.
§ Hennekinne, J.A.; Ostyn, A.; Guillier, F.; Herbin, S.; Prufer, A. L. and Dragacci, S.
(2010). How Should Staphylococcal Food Poisoning Outbreaks Be Characterized? Toxins 2: 2106-2116.
§ Hennekinne, J. A.; DeBuyser, M. L. and Dragacci, S. (2011). Staphylococcus aureus and its food poisoning toxins: characterization and outbreak investigation. FEMS Microbiol. Rev. 10:1574- 6976.
§ Hoffman, M.; Tremaine, M.; Mansfield, J. and Betley, M. (1996). Biochemical and mutational analysis of the histidine residues of staphylococcal enterotoxin A. Infect. Immun. 64: 885–890.
§ Holeckova, B.; Holoda, E.; Fotta, M.; Kalinacova, V.;Gondol, J.; Grolmus, J. (2002). Occurrence of enterotoxigenic Staphylococcus aureus in food. Ann Agric Environ Med. 9: 179–182.
§ Hovde, C.J.; Marr, J.C.; Hoffmann, M.L.; Hackett, S.P.; Chi, Y.I.; Crum, K.K.; Stevens, D.L.; Stauffacher, C.V.; and Bohach, G.A. (1994). Investigation of the role of the disulphide bond in the activity and structure of staphylococcal enterotoxin C1. Mol. Microbiol. 13: 897-909.
§ Hu, D.L.; Omoe, K.; Shimoda, Y.; Nakane, A.; and Shinagawa, K. (2003). Induction of Emetic Response to Staphylococcal Enterotoxins in the House Musk Shrew (Suncus Murinus). Infect. Immun. 71: 567–570.
§ Hu, D.L.; Zhu, G.; Mori, F.; Omoe, K.; Okada, M.; Wakabayashi, K.; Kaneko, S.;
Shinagawa, K.; and Nakane, A. (2007). Staphylococcal Enterotoxin Induces Emesis Through Increasing Serotonin Release in Intestine and It Is Downregulated by Cannabinoid Receptor 1. Cell. Microbiol. 9: 2267–2277.
§ Hu, D.L.; Omoe, K.; Sashinami, H.; Shinagawa, K.; and Nakane, A. (2009).
Immunization with a Nontoxic Mutant of Staphylococcal Enterotoxin A, SEAD227A, Protects Against Enterotoxin-Induced Emesis in House Musk Shrews. J. Infect. Dis. 199: 302–310.

§ Hwang, S.Y.; Kim, S.H.; Jang, E.J.; Kwon, N.H.; Park, Y.K.; Koo, H.C.; Jung, W.K.; Kim, J.M. and Park, Y.H. (2007). Novel multiplex PCR for the detection of the Staphylococcus aureus superantigen and its application to raw meat isolates in Korea. Int. J. Food Microbiol. 117: 99–105.
I § Ikeda, T.; Tamate, N.; Yamaguchi, K. and Makino, S. (2005). Mass Outbreak of
Food Poisoning Disease Caused by Small Amounts of Staphylococcal Enterotoxins A and H. Appl. Environ. Microbiol. 71: 2793–2795.
§ Ippolito, G.; Leone, S.; Lauria, F. N.; Nicastri, E. and Wenzel, R.P. (2010). Methicillin-Resistant Staphylococcus aureus: The suoerbug. Int. J. Infect. Dis. 14: S7-S11.
§ Iraqi Central Organization for Standardization and Quality Control (C.O.S.Q.C). (2006). Iraqi Standard Criterion No.2/2270 in Sampling.
§ Iraqi Central Organization for Standardization and Quality Control (C.O.S.Q.C). (2006). Iraqi Standard Criterion No.3/2270 in Isolation, Enumeration and Identification of Microbiological Groups in Foods.
§ Irlinger, F. (2008). Safety assessment of Dairy microorganisms: Coagulase negative staphylococci. Int. J. Food Microbiol. 126: 302-310.
J § Janstova, B.; Necidova, L. and Bohumíra, J. (2012). Comparing the growth of
Staphylococcus aureus and production of staphylococcal enterotoxin C in sheep’s and goat’s milk. Journal of Microbiology, Biotechnology and Food Sciences. 758-768.
§ Jarp, J. (1991). Classification of coagulase-negative staphylococci isolated from bovine clinical and subclinical mastitis. Vet. Microbiol. 27:151-158.
§ Jayavanth, P. and Napori, N.M. (2011). Ceftazidime and vancomycin sensitive Beta haemolytic Staphylococcus aureus. Int. J. Pharm. Biomed. Res. 2(2):63-66.
§ Jones, T.F.; Kellum, M.E.; Porter, S.S.; Bell, M.; and Schaffner, W. (2002). An outbreak of community-acquired foodborne illness caused by methicillin-resistant Staphylococcus aureus. Emerg. Infect. Dis. 8: 82–84.

§ Jorgensen, H.J.; Mathisen, T.; Lovseth, A.; Omoe, K.; Qvale, K.S.; and Loncarevic, S. (2005a). An outbreak of staphylococcal food poisoning caused by enterotoxin H in mashed potato made with raw milk. FEMS Microbiol. Lett. 252: 267–272.
§ Jorgensen, H.J.; Mørk, T.; Caugant, D.A.; Kearns, A.; and Rorvik, L.M. (2005b). Genetic variation among Staphylococcus aureus strains from Norwegian bulk milk. Appl. Environ. Microbiol. 71: 8352–8361.
K
§ Kappler, J.W; Hamad, A.R. and Marrack P. (1997).Transcytosis of staphylococcal superantigen toxins. J. Exp. Med. 185: 1447–1454.
§ Karahan, M. and Cetinkaya, B. (2007). Coagulase gene polymorphism detected by PCR in Staphylococcus aureus isolated from subclinical bovine mastitis in Turkey. The Veterinary Journal 174: 428-431.
§ Karp, D.R.; and Long, E.O. (1992). Identification of HLA-DR1 Beta Chain
Residues Critical for Binding Staphylococcal Enterotoxins A and E. J. Exp. Med. 175: 415–424.
§ Kayser, F.H; Bienz,K.A.; Eckert, J. and Zinkernagel, R.M.(2005). Color Atlas of Medical Microbiology.11th ed. Thieme Verlag, NewYork, USA: 300-303.
§ Katsuda, K.; Hata, E.; Eguchi, M. (2005). Molecular typing of S.aureus isolated from bovine mastitic milk on the basic of toxin genes and coagulase gene polymorphisms. Veterinary Microbiology. 105:301-305.
§ Kérouanton, A.; Hennekinne, J.A.; Letertre, C.; Petit, L.; Chesneau, O.; Brisabois, A.; and De Buyser, M.L. (2007). Characterization of Staphylococcus aureus strains associated with food poisoning outbreaks in France. Int. J. Food Microbiol. 115: 369–375.
§ Kilpper-Balz, R. and Schleifer, K. H. (1981). Transfer of Peptococcus saccharolyticus Foubert and Douglas to the genus Staphylococcus: Staphylococcus saccharolyticus (Foubert and Douglas) comb nov. Zentralbl. Bakteriol. Hyg. Abt. 1 Orig. Reihe C 2: 324-331.
§ Kisiel, A.; Turek, A.S. and Szweda, W. (2011). Application of the suckling mouse bioassay to assess the enterotoxigenic properties of Yersinia enterocolitica strains isolated from fattening pigs. Bull Vet Inst Pulawy. 55: 21-25.

§ Kitamoto, M.; Kito, K.; Niimi, Y.; Shoda, S.; Takamura, A.; Hiramatsu, T.; Akashi, T.; Yokoi, Y.; Hirano, H.; Hosokawa, M.; Yamamoto, A.; Agata, N. and Hamajima, N. (2009). Food poisoning by Staphylococcus aureus at a university festival. Jpn. J. Infect. Dis. 62: 242–243.
§ Kloos, W.E. and Schleifer, K. H. (1975). Isolation and characterization of staphylococci from human skin. II. Descriptions of four new species: Staphylococcus warneri, Staphylococcus capitis, Staphylococcus hominis, and Staphylococcus simulans. Int. J. Syst. Bacteriol. 25: 62-79.
§ Kloos,W. E. and Wolfshohl, J. F. (1991). Staphylococcus cohnii subspecies: Staphylococcus cohnii subsp. cohnii subsp. nov. and Staphylococcus cohnii subsp. urealyticum subsp. nov. Int. J. Syst. Bacteriol. 41: 284- 289.
§ Krakauer, T. (1994). Costimulatory Receptors for the Superantigen Staphylococcal Enterotoxin B on Human Vascular Endothelial Cells and T Cells. J. Leukoc. Biol. 56: 458–463.
§ Krakauer, T. (2005). Chemotherapeutics Targeting Immune Activation by Staphylococcal Superantigens. Med. Sci. Monit. 11: 290–295.
§ Krakauer, T.; Buckley, M.; Issaq, H.J.; and Fox, S.D. (2010). Rapamycin Protects Mice From Staphylococcal Enterotoxin B-Induced Toxic Shock and Blocks Cytokine Release in vivo and in vivo. Antimicrob. Agents Chemother. 54: 1125–1131.
L § Larkin, E.A.; Carman, R.J.; Krakauer, T. and Stiles, B.G. (2009). Staphylococcus
aureus: the toxic presence of a pathogen extraordinaire. Curr. Med. Chem. 16: 4003–4019.
§ Le Loir, I.; Baron, F. and Gautier, M. (2003). Staphylococcus aureus and food poisoning. Genet. Mol. Res. 2: 63-76.
§ Ler, S.G.; Lee, F.K. and Gopalakrishnakone, P. (2006). Trends in Detection of Warfare Agents. Detection Methods for Ricin, Staphylococcal Enterotoxin B and T-2 Toxin. J. Chromatogr. A. 1133: 1–12.

§ Letertre C.; Perelle, S.; Dilasser, F.; and Fach, P. (2003). Identification of a new putative enterotoxin SEU encoded by the egc cluster of Staphylococcus aureus. J. Appl. Microbiol. 95: 38–43.
§ Leung, D. Y., Meissner, H. C., Fulton, D. R., Murray, D. L., Kotzin, B. L. and Schlievert, P. M. (1993). Toxic shock syndrome toxin-secreting Staphylococcus aureus in Kawasaki syndrome. Lancet. 342(8884): 1385-1388.
§ Lim, Y.; Jana, M.; Luong, T. T. and Lee, C. Y. (2004). Control of Glucose and NaCl-Inducded Biofilm Formation by rbf in Staphylococcus aureus. J. Bacteriol. 186(3): 722-729.
§ Lin, Z.; Kotler, D.P.; Schlievert, P.M.; and Sordillo, E.M. (2010). Staphylococcal Enterocolitis: Forgotten but Not Gone? Dig. Dis. Sci. 55: 1200–1207.
§ Lina, G.; Bohach, G.A.; Nair, S.P.; Hiramatsu, K.; Jouvin-Marche, E. and
Mariuzza, R. (2004). Standard nomenclature for the superantigens expressed by Staphylococcus. J. Infect. Dis. 189: 2334–2336.
§ Lindsay, J.A.; Ruzin, A.; Ross, H.F.; Kurepina, N. and Novick, R.P. (1998). The Gene for Toxic Shock Toxin Is Carried by a Family of Mobile Pathogenicity Islands in Staphylococcus aureus. Mol. Microbiol. 29: 527–543.
§ Lisa R.W. (2004). Staphylococcus aureus exfoliative toxins: How they cause disease. Derm. Foundat. 122: 1070-1077.
§ Loir, Y.L.; Baron, F. and Gautier, M. (2003). Staphyloccus aureus and food poisoning. Genet. Mol.Res. 2(1):63-76.
§ Liu, D.; Zienkiewicz, J.; DiGiandomenico, A. and Hawiger, J. (2009). Suppression of Acute Lung Inflammation by Intracellular Peptide Delivery of a Nuclear Import Inhibitor. Mol. Ther. 17: 796–802.
§ Luthje, P. and Schwarz, S. (2006). Antimicrobial resistance of coagulase-negative staphylococci from bovine subclinical mastitis with particular reference to macrolide-lincosamide resistance phenotypes and genotypes. J. Antimicrob. Chemother. 57:966-969.
M

§ Maidhof, H.; Reinicke, B.; Blumel, P.; Berger-Bachi, B. and Labischinski, H. (1991). femA, which encodes a factor essential for expression of methicillin resistance, affects glycine content of peptidoglycan in methicillin-resistant and methicillin-susceptible Staphylococcus aureus strains. J. Bacteriol. 173:3507-3513.
§ Manfredi, E. A.; Leotta, G. A. and Rivas, M. (2010). Multiplex PCR for the detection of sea, seb, sec, sed and see genes of Staphylococcus aureus. Characterization of isolates from food. Rev Argent Microbiol. 42(3): 212-215.
§ Maniatis, T.; Fritsch, E. and Sambrook, J. (1982). Molecular Cloning: a Laboratory Manual. Cold Spring Harbor Laboratory. New York.
§ Marta, D.; Wallin-Carlquist, N.; Schelin, J.; Borch, E. and Radstrom, P. (2011). Extended staphylococcal enterotoxin D expression in ham products. Food Microbiol. 28:617-20.
§ Martin, M.C.; Fueyo, J.M.; González-Hevia, M.A. and Mendoza, M.C. (2004).
Genetic procedures for identification of enterotoxigenic strains of Staphylococcus aureus from three food poisoning outbreaks. Int. J. Food Microbiol. 94: 279–286.
§ Matos, J.S.; White, D.G.; Harmon, R.J. and Langlois, B.E. (1991). Isolation of Staphylococcus aureus from sites other than the lactating mammary gland. J. Dairy Sci. 74:1544-1549.
§ Matthews, K.R.; Harmon, R.J.; and Langlois, B.E. (1991). Effect of naturally
occurring coagulase-negative staphylococci infections on new infections by mastitis pathogens in the bovine. J. Dairy Sci. 74:1855-1859.
§ McCulloch, J. A. (2006). Avaliação da funcionalidade do locus accessory gene regulator (agr) em cepas de Staphylococcus aureus brasileirascom suscetibilidade reduzida aos glicopeptídeos. Faculdade de Ciências Farmacêuticas, São Paulo.
§ MacFaddin, J. F. (2000). Biochemical tests for identification of medical bacteria. 3rd ed. Lipicott Williams and Wilkins USA.
§ McLauchlin, J.; Narayanan, G.L.; Mithani, V.; and O’Neil, G. (2000). The detection of enterotoxins and toxic schock síndrome toxin genes in Staphylococcus aureus by polymerase chain reaction. J. Food Prot. 63: 479–488.
§ McNamara, P. J.; Milligan-Monroe, K. C.; Khalili, S. and Proctor, R. A. (2000). Identification, cloning, and initial characterization of rot, a locus encoding a

regulator of virulence factor expression in Staphylococcus aureus. J. Bacteriol. 182 (11): 3197-3203.
§ Mehrotra, M.; Wang, G. and Johnson, W. M. (2000). Multiplex PCR for detection of genes for Staphylococcus aureus enterotoxins, exfoliative toxins, toxic shock syndrome toxin 1, and methicillin resistance. J. Clin. Microbiol. 38: 1032-1035.
§ Merlino, J.; Watson, J.; Rose, B.; Beard-Pegler, M.; Gottlieb, T.; Bradbury, R. and
Harbour. C. (2002). Detection and expression of methicillin/oxacillin resistance in multidrug-resistant and non-multidrug-resistant Staphylococcus aureus in central Sydney, Australia. J. Antimicrob. Chemother. 49793-801.
§ Mirzaei, H.; Farhoudi, H.; Tavassoli, H.; Farajli, M. and Monadi A. (2011). Presence and antimicrobial susceptibility of methicillin resistant Staphylococcus aureus in raw and pasteurized milk and ice cream in Tabriz by culture and PCR techniques. African Journal of Microbiology Research. 6(32): 6224-6229.
§ Mirzaie, H.; Farhoudi, H.; Tavassoli, H.; Farajli, M. and Monadi, A. (2012). Presence and antimicrobial susceptibility of methicillin resistant Staphylococcus aureus in raw and pasteurized milk and ice cream in Tabriz by culture and PCR techniques. African Journal of Microbiology Research. 6(32): 6224-6229.
§ Momtaz, H.; Rahimi, E. and Tajbakhash, E. (2010). Detection of some virulence factors in Staphylococcus aureus isolated from clinical and subclinical bovine mastitis in Iran. African Journal of Biotechnology. 9 (25): 3753-3758.
§ Momtaz, H.; Tajbakhash, E. and Rahimi, E. (2011). Coagulase gene polymorphism of Staphylococcus aureus isolated from clinical and subclinical bovine mastitis in Isfahan and Chaharmahal Va Bakhtiari Provinces of Iran. Comp. Clin. Pathol. 20: 519-522.
§ Montanaro, L. Arciola, C.R.; Baldassarrri, L. and Borsetti, E. (1999). Presence and expression of collagen adhesion gene (cna) and slime production in Staphylococcus aureus strains from orthopaedic prosthesis infections. Biomaterials. 20: 1945-1949.
§ Morandi, S.; Brasca, M.; Lodi, R.; Cremones, P. and Castiglioni, B. (2007). Detection of classical enterotoxins and identification of enterotoxin genes in Staphylococcus aureus from milk and dairy products. Veter. Microbiol. 124: 66-72.

§ Morandi, S.; Brasca, M.; Andrighetto, C.; Lombardi, A. and Lodi, R. (2009). Phenotypic and Genotypic Characterization of Staphylococcus aureus Strains from Italian Dairy Products. Int. J. Microbiol. 501362:1–501362:7.
§ Morello, J. A.; Mizer, H. E.; and Granato, P. A. (2006). Laboratory Manual and Workbook in Microbiology Applications to Patient Care. 18th ed. McGraw Hill.
§ Much, P.; Pichler, J.; Kasper, S.S.; and Allerberger, F. (2009). Foodborne
Outbreaks, Austria 2007. Wien. Klin. Wochenschr. 121: 77–85.
§ Muraille, E.; De Trez, C.; Pajak, B.; Brait, M.; Urbain, J.; and Leo, O. (2002). T Cell-Dependent Maturation of Dendritic Cells in Response to Bacterial Superantigens. J. Immunol. 168: 4352–4360.
§ Murray, R.cJ. (2005). Recognition and Management of Staphylococcus aureus
Toxin-Mediated Disease. Intern. Med. J. 35:106–119.
N § Nafisi, M.R.; Kalhor, H.; Zamanzad, B.; Karimi, A.; Farokhi, A. and Validi, M.
(2008). Comparison of agar screen and duplex-PCR in determination of methicillin resistant Staphylococcus aureus (MRSA) strains isolated from nose of personnel in Hajar hospital of Shahre-kord. A. M.U.J. 11:94-100.
§ Naik, S.; Smith, F.; Ho, J.; Croft, N.M.; Domizio, P.; Price, E.; Sanderson, I.R.; and Meadows, N.J. (2008). Staphylococcal enterotoxins G and I, a cause of severe but reversible neonatal enteropathy. Clin. Gastroenterol. Hepatol. 6: 251–254.
§ Naresh, K.; Sharma, D.; Catherine, E.; Rees, D. and Christine, E. R. (2000). Development of a single-reaction multiplex PCR toxin typing assay for staphylococcus aureus strains. Appl and Environ. Microbiol. 66(4):1347-1353.
§ National Committee for Clinical Laboratory Standards (NCCLS) (2006).
Performance standards for antimicrobial Susceptibility testing. NCCLS approved standard M100 – S16.
§ Niveditha, S.; Shylaja, R.; Murali, H. S. and Batra, H. V. (2012). A novel mPCR
for the detection of prominent toxins in MRSA strains of S. aureus recovered from diverse sources. International Journal of Research in Biological Sciences. 2 (1): 26-32.

§ Normanno, G.; La Salandra, G.; Dambrosio, A.; Quaglia, N.C.; Corrente, M.; Parisi, A.; Santagada, G.; Firinu, A.; Crisetti, E. and Celano, G.V. (2007). Occurrence, characterization and antimicrobial resistance of enterotoxigenic Staphylococcus aureus isolated from meat and dairy products. Int. J. Food Microbiol. 115:290-296.
§ Noto, M. J.; and Archer, G.L. (2006). A subset of Staphylococcus aureus strains harboring staphylococcal cassette chromosome mec (SCCmec) type IV is deficient in CcrAB-mediated SCCmec excision. Antimicrob. Agents Chemother. 50: 2782–2788.
§ Novick, R. P. and Subedi, A. (2007). The SaPIs: mobile pathogenicity islands of Staphylococcus. Chem. Immunol. Allergy. 93: 42–57.
§ Novick, R.P.; Christie, G.E.; and Penades, J.R. (2010). The phage-related chromosomal islands of Gram-positive bacteria. Nat Rev Microbiol . 8:541 51.
§ Novick, R.P. and Geisinger, E. (2008). Quorum sensing in staphylococci. Annu Rev Genet. 42:541-64.
O § O’Brien, M.; Hunt, K.; McSweeney, S. and Jordan, K. (2009). Occurrence of
foodborne pathogens in Irish farmhouse cheese. Food Microbiol. 26: 910-914.
§ Omoe, K.; Ishikawa, M.; Shimoda, Y.; Hu, D.L.; Ueda, S. and Shinagawa, K. (2002). Detection of seg, seh, and sei genes in Staphylococcus aureus isolates and determination of the enterotoxin productivities of S. aureus isolates harboring seg, seh, or sei genes. J. Clin. Microbiol. 40: 857–862.
§ Omoe, K.; Hu, D.L.; Takahashi-Omoe, H.; Nakane, A. and Shinagawa, K. (2003). Identification and characterization of a new staphylococcal enterotoxin related putative toxin encoded by two kinds of plasmids. Infect. Immun. 71: 6088-6094.
§ Omoe, K.; Hu, D.L.; Takahashi-Omoe, H.; Nakane, A. and Shinagawa, K. (2005). Comprehensive analysis of classical and newly described staphylococcal superantigenic toxin genes in Staphylococcus aureus isolates. FEMS Microbiol. Lett. 246:191–198.
§ Ono, H.K.; Omoe, K.; Imanishi, K.; Iwakabe, Y.; Hu, D.L.; Kato, H.; Saito, N.; Nakane, A.; Uchiyama, T. and Shinagawa, K. (2008). Identification and

characterization of two novel staphylococcal enterotoxins, types S and T. Infect. Immun. 76: 4999–5005.
§ Onoue, Y. and Mori, M. (1997). Amino acid requirements for the growth and enterotoxin production by Staphylococcus aureus in chemically defined media. Int J Food Microbiol. 36:77-82.
§ Ortega, E.; Hikmate, A.; Lucas, R.and Gálvez A. (2010). Multiple Roles of Staphylococcus aureus Enterotoxins: Pathogenicity, Superantigenic Activity, and Correlation to Antibiotic Resistance Toxins. 2: 2117-2131.
§ Orwin, P.M.; Leung, D.Y.; Tripp, T.J.; Bohach, G.A.; Earhart, C.A.; Ohlendorf, D.H. and Schlievert, P.M. (2002). Characterization of a novel staphylococcal enterotoxin-like superantigen, a member of the group V subfamily of pyrogenic toxins. Biochemistry. 41: 14033–14040.
§ Orwin, P. M.; Fitzgerald, J. R.; Leung, D. Y. M.; Gutierrez, J. A.; Bohach, G. A.; and Schlievert, PM (2003). Characterization of Staphylococcus aureus Enterotoxin L. Infect. Immun. 71(5): 2916–2919.
§ Ostyn, A.; De Buyser, M. L.; Guillier, F.; Groult, J.; Felix, B.; Salah, S.; Delmas, G. and Hennekinne, J. A. (2010). First evidence of a food poisoning outbreak due to staphylococcal enterotoxin type E, France, 2009. Euro. Surveill. 15: pii. 19528.
P § Pantucek, R., Sedlacek, I., Petras, P., Koukalova, D., Svec, P., Stetina, V.,
Vancanneyt, M., Chrastinova, L., Vokurkova, J., Ruzickova, V., Doskar, J., Swinges, J. and Hajek, V. (2005). Staphylococcus simiae sp. nov., isolated from South American squirrel monkeys. Int. J. Syst. Evol. Microbiol. 55:1953-1958.
§ Partida, A.; Espunes,T.; Marinez, J.B. (2010). Characterization and Persistence of Staphylococcus aureus Strains Isolated from the Anterior Nares and Throats of Healthy Carriers in a Mexican Community. J. of Clin. Microbiol. 48(5): 1701-1705.
§ Peles, F.; Wagner, M.; Varga, L.; Hein, I.; Rieck, P.; Gutser, K.; Keresztúri, P.;
Kardos, G.; Turcsányi, I.; Béri, B. and Szabo, A. (2007). Characterization of Staphylococcus aureus strains isolated from bovine milk in Hungary. Int. J. Food Microbiol. 118:186-193.
§ Pender, S. L.; Breese, E. J.; Gunther, U.; Howie, D.; Wathen, N. C.; Schuppan, D.
and MacDonald, T. T. (1998). Suppression of T Cell-Mediated Injury in Human

Gut by Interleukin 10: Role of Matrix Metalloproteinases. Gastroenterology. 115: 573–583.
§ Peng, H. L.; Novick, R. P.; Kreiswirth, B.; Kornblum, J.; and Schlievert, P. (1988). Cloning, characterization, and sequencing of an accessory gene regulator (agr) in Staphylococcus aureus. J. Bacteriol. 170(9): 4365-4372.
§ Pereira, V.; Lopes, C.; Castro, A.; Silva, J.; Gibbs, P. and Teixeira, P. (2009). characterization for enterotoxin production. Virulence factors, and antibiotic susceptibility of staphylococcus aureus isolates from various foods in Portugal. Food Microbiol. 26:278-282.
§ Perez-Bosque, A.; Miro, L.; Polo, J.; Russell, L.; Campbell, J.; Weaver, E.; Crenshaw, J. and Moreto, M. (2010). Dietary Plasma Protein Supplements Prevent the Release of Mucosal Proinflammatory Mediators in Intestinal Inflammation in Rats. J. Nutr. 140: 25–30.
§ Pesavento, G.; Ducci, B.; Comodo, N. and Nostro, A.L. (2007). Antimicrobial
resistance profile of Staphylococcus aureus isolated from raw meat: a research for methicillin resistant Staphylococcus aureus (MRSA). Food Control 18:196-200.
§ Pfaller, M. A. and Herwaldt, L. A. (1988). Laboratory, clinical, and epidemiological aspects of coagulase-negative staphylococci. Clin. Microbiol. Rev. 1: 281-299.
§ Piette, A. and Verschraegen, G. (2009). Role of coagulase-negative staphylococci in human disease. Vet. Microbiol. 134: 45-54.
§ Pinchuk, I.V.; Beswick, E.J.; Saada, J.I.; Suarez, G.; Winston, J.; Mifflin, R.C.; Di
Mari, J.F.; Powell, D.W. and Reyes, V.E. (2007). Monocyte Chemoattractant Protein-1 Production by Intestinal Myofibroblasts in Response to Staphylococcal Enterotoxin a: Relevance to Staphylococcal Enterotoxigenic Disease. J. Immunol. 178: 8097–8106.
§ Pinchuk, I. V.; Beswick, E. J. and Reyes, V. E. (2010). Staphylococcal Enterotoxins. Toxins. 2: 2177-2197.
§ Pocsfalvi, G.; Cacace, G.; Cuccurullo, M.; Serluca, G.; Sorrentino, A. and
Schlosser, G. (2008). Proteomic analysis of exoproteins expressed by enterotoxigenic Staphylococcus aureus strains. Proteomics. 8: 2462-76.

§ Pu, S.; Wang, F. and Ge, B. (2011). Characterization of toxin genes and antimicrobial susceptibility of Staphylococcus aureus isolates from Louisiana retail meats. Foodborne Pathog Dis. 8(2): 299-306.
R § Rall, V.L.M.; Vieira, F. B., Rall, R.; Vieitis, R.L.; Fernandes, A.; Candeias, J.M.G.;
Cardoso, K.F.G. and Araujo, J. P. (2008). PCR detection of staphylococcal enterotoxin genes in Staphylococcus aureus strains isolated from raw and pasteurized milk. Veterinary Microbiology. 132: 408-413.
§ Rao, S. P.N. (2006). Food Poisoning. www.microrao.com.
§ Rather, P.N.; Davis, A.P. and Wilkinson, B.J. (1986). Slime production by bovine milk Staphylococcus aureus and identification of coagulase-negative staphylococcal isolates. J. Clin. Microbiol. 23: 858-862.
§ Resch, M.; Nagel, V. and Hertel, C. (2008). Antibiotic resistance of coagulase-negative staphylococci associated with food and used in starter cultures. Int. J. Food Microbiol. 127: 99-104.
§ Riffon, R.; Sayasith, K.; Khalil, H.; Dubreuil, P.; Drolet, M. and Lagace, J. (2001). Development of a rapid and sensitive test for identification of major pathogens in bovine mastitis by PCR. J. Clin. Microbiol. 39:2584–2589.
S
§ Sabet, N. S.; Subramaniam, G.; Navaratnam, P. and Sekaran, S.D. (2007). Detection of methicillin-aminoglycoside-resistant genes and simultaneous identification of S. aureus using triplex real-time PCR Taqman assay. Journal of Microbiological methods. 68:157-162.
§ Sakai, F.; Ihara, H.; Aoyama, K.; Igarashi, H.; Yanahira, S. and Ohkubo, T. (2008). Characteristics of enterotoxin H-producing Staphylococcus aureus isolated from clinical cases and properties of the enterotoxin productivity. J. Food Prot. 71:1855-60.
§ Sanchez, N.G.; Rodriguez, R.; Ruiz, O. and Garza, L.M. (2003). Development of two multiplex polymerase chain reactions for the detection of enterotoxigenic strains of Staphylococcus aureus isolated from foods. J. Food Prot. 66: 1055-1062.

§ Sawant, A.A.; Gillespie, B.E. and S.P. Oliver, (2009). Antimicrobial susceptibility of coagulase-negative Staphylococcus species isolated from bovine milk. Vet. Microbiol. 134: 73-81.
§ Schelin, J.; Wallin-Carlquist, N.; Cohn, M. T.; Lindqvist, R.; Barker,G. C. and
Radstorm, P. (2011). The formation of Staphylococcus aureus enterotoxin in food environments and advances in risk assessment. Virulence 2(6): 580-592.
§ Scherrer, D.; Corti, S.; Muehlherr, J. E.; Zweifel, C. and Stephan, R. (2004). Phenotypic and genotypic characteristics of Staphylococcus aureus isolates from raw bulk-tank milk samples of goats and sheep. Vet Microbiol. 101: 101-107.
§ Schleifer, K.H. and Kloos, W. E. (1975). Isolation and characterization of
staphylococci from human skin. I. Amended description of Staphylococcus epidermidis and Staphylococcus saprophyticus and descriptions of three new species: Staphylococcus cohnii, Staphylococcus haemolyticus, and Staphylococcus xylosus. Int. J. Syst. Bacteriol. 25: 50-61.
§ Schlievert, P .M.; Jablonski, L. M.; Roggiani, M.; Sadler, I.; Callantine, S.; Mitchell, D. T.; Ohlendorf, D. H. and Bohach, G. A. (2000). Pyrogenic toxin superantigen site specificity in toxic shock syndrome and food poisoning in animals. Infect. Immun. 68(6): 3630-3634.
§ Schlievert, P. M. and Case, L. C. (2007). Molecular analysis of staphylococcal superantigens. Methods Mol. Biol. 391: 113-126.
§ Schmitz, F.J.; Steiert, M.; Hofmann, B.; Verhoef, J.; Hadding, U.; Heinz, H.P. and Kohrer, K. (1998). Development of a multiplex-PCR for direct detection of the genes for enterotoxin B and C and toxic shock syndrome toxin-1 in Staphylococcus aureus isolates. J. Med. Microbiol. 47: 335-340.
§ Scott, W. J. (1953). Water relations of Staphylococcus aureus at 30 degrees C. Aust. J. Biol. Sci. 6: 549–564.
§ Scott, W.J. and Van Duijkeren, E. (2010). Methicillin-resistant Staphylococcun aureus and Staphylococcus pseudintermedius in veterinary medicine.Vet. Microbiol. 140(3-4):418-429.
§ Sharma, D.; Sharma, P. K. and Malik, A. (2011). Prevalence and Antimicrobial Susceptibility of Drug Resistant Staphylococcus aureus in Raw Milk of Dairy Cattle. International Research Journal of Microbiology (IRJM). 2(11): 466-470.

§ Shimizu, A.; Fujita, M.; Igarashi, H.; Takagi, M.; Nagase, N.; Sasaki, A. and
Kawano, J. (2000). Characterization of Staphylococcus aureus coagulase type VII isolates from staphylococcal food poisoning outbreaks (1980–1995) in Tokyo, Japan, by pulsed field gel electrophoresis. J. Clin. Microbiol. 38: 3746–3749.
§ Sila, J.; Sauer, P.; Kolar, M. (2009). Comaprison of the prevalence of genes coding for enterotoxins, exfoliatins, Panton-Valentine leukocidin and TSST-1 between methicillin-resistant and methicillin-susceptible isolates of Staphylococcus aureus at the University Hospital in Olomouc. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub.153: 215–218.
§ Stamford, T. L. M.; Silva, C.G.M.; Mota, R.A. and Neto, C. A. (2006). Enterotoxigenicidade de staphylococcus sp isolates de leite in natura. Cienc. Tecnol. Aliment. 26: 41-45.
§ Stephenson, F. H. (2003). Calculations for Molecular Biology and Biotechnology:
A guide to mathematics in the laboratory. Elsevier Science. USA.
§ Stewart, G.C. (2005). Staphylococcus aureus. In Foodborne pathogens: Microbiology and Molecular Biology; Fratamico, P.M.; Bhunia, A.K.; Smith, J.L.; Eds.; Caister Academic Press: Norfolk, UK, 273–284.
§ Stiles, B.G.; Campbell, Y.G.; Castle, R.M. and Grove, S.A. (1999). Correlation of Temperature and Toxicity in Murine Studies of Staphylococcal Enterotoxins and Toxic Shock Syndrome Toxin 1. Infect. Immun. 67: 1521–1525.
§ Soares, J. C.; Marques, M. R.; Tavaria, F. K.; Pereira, J. O.; Xavire, F. and Pintado, M.M. (2011). Biodiversity and characterization of Staphylococcus species isolated from a small manufacturing dairy plant in Portugal. International Journal of Food Microbiology. 146: 123-129.
§ Su, Y.C. and Wong, A.C. (1996). Detection of staphylococcal enterotoxin H by an enzyme-linked immunosorbent assay. J. Food Prot. 59: 327–330.
T § Tallent, S.M.; Langston, T.B.; Moran, R.G. and Christie, G.E. (2007). Transducing
particles of Staphylococcus aureus pathogenicity island SaPI1 are comprised of helper phage-encoded proteins. J. Bacteriol. 189: 7520–7524.
§ Tang, J; Tang, C., Wang, Y., Chen, J.; Liu, J.; Liu, L. and Yue, H. (2012). Surveillance study of enterotoxin genes in Staphylococcus aureus isolates from

goats of different slaughterhouses in Sichuan, China. Annals of Microbiology. 62: 1247-1253.
§ Taponen, S., Surre, K., Piessens, V., Van Coillie, E., De Vliegher, S. and Koort, J. M. K. (2012). Staphylococcus agnetis sp. nov., a coagulase-variable species from bovine subclinical and mild clinical mastitis. Int. J. Syst. Evol. Microbiol. 62: 61-65.
§ Taponen, S. and Pyorala, S. (2009). Coagulase-negative staphylococci as cause of bovine mastitis-no so different from Staphylococcus aureus. Vet. Microbiol. 134: 29-36.
§ Tasci, F., Sahindokuyucu, and Ozturk, D. (2011). Detection of Staphylococcus species and staphylococcal enterotoxins by ELISA in ice cream and cheese consumed in Burdur Province. African Journal of Agricultural Research. 6(4): 937-942.
§ Thibodeau, J.; Cloutier, I.; Lavoie, P.M.; Labrecque, N.; Mourad, W.; Jardetzky, T. and Sekaly, R.P. (1994). Subsets of HLA-DR1 Molecules Defined by SEB and TSST-1 Binding. Science. 266: 1874–1878.
§ Thoendel, M.; Kavanaugh, J.S.; Flack, C.E. and Horswill, A.R. (2011). Peptide signaling in the staphylococci. Chem Rev. 111:117-51. PMID: 21174435.
§ Thomas, D.Y.; Jarraud, S.; Lemercier, B.; Cozon, G.; Echasserieau, K.; Etienne, J.; Gougeon, M.L.; Lina, G. and Vandenesch, F. (2006). Staphylocccal enterotoxin-like toxins U2 and V, two new staphylococcal superantigens arising from recombination within the enterotoxin gene cluster. Infect. Immun. 74: 4724–4734.
§ Thomas, D.; Chou, S.; Dauwalder, O. and Lina, G. (2007). Diversity in Staphylococcus aureus enterotoxins. Chem. Immunol. Allergy. 93: 24–41.
§ Throup, J. P.; Zappacosta, F.; Lunsford, R. D.; Annan, R. S.; Carr, S. A.; Lonsdale, J. T.; Bryant, A. P.; McDevitt, D.; Rosenberg, M. and Burnham, M. K. (2001). The srhSR gene pair from Staphylococcus aureus: genomic and proteomic approaches to the identification and characterization of gene function. Biochem. 40: 10392-10401.
§ Tilahun, A.Y.; Theuer, J.E.; Patel, R.; David, C.S. and Rajagopalan, G. (2010). Detrimental Effect of the Proteasome Inhibitor, Bortezomib in Bacterial Superantigen- and Lipopolysaccharide-Induced Systemic Inflammation. Mol. Ther. 18: 1143–1154.

§ Trülzsch, K.; Grabein, B.; Schumann, P.; Mellmann, A.; Antonenka, U.; Heesemann, J. and Becker, K. (2007). Staphylococcus pettenkoferi sp. nov., a novel coagulase-negative staphylococcal species isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 57:1543-1548.
U § Uchiyama, T.; Imanishi, K.; Miyoshi-Akiyama, T. and Kato, H. (2006).
Staphylococcal superantigens and the diseases they cause. In The Comprehensive Sourcebook of Bacterial Protein Toxins, 3rd ed.; Alouf, J.E., Popoff, M.R., Eds.; Academic Press: Burlington, VT, USA. 830–843.
V § Valle, J.; Lucia, G. E.; Piriz, S.; Goyache, J.; Orden, J. A. and Vadillo, S. (1990).
Enterotoxin production by staphylococci isolated from healthy goats. Appl Environ Microbiol. 56: 1323-1326.
§ Valero, A.; Perez-Rodriguez, F.; Carrasco, E.; Alventosa, J.M.; Garcia-Gimeno, R.M. and Zurera, G. (2009). Modelling the growth boundaries of Staphylococcus aureus: Effect of temperature, pH and water activity. Int J Food Microbiol .133:186-94.
§ Vandepitte, J.; Verhaegen, J.; Engbaek, K.; Rohner, P.; Piot, P. and Heuck, C.C. (2003). Basic Laboratory Procedures in Clinical Bacteriology. 2nd. ed. World Health Organization, Geneva.
§ Van Wamel, W.J.; Rooijakkers, S.H.; Ruyken, M.; van Kessel, K.P. and van Strijp, J.A. (2006). The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J. Bacteriol. 188: 1310–1315.
§ Varshney, A. K.; Mediavilla, J. R.; Robiou, N.; Guh, A.; Wang, X.; Gialanella, P.; Levi, M. H.; Kreiswirth, B. N. and Fries, B.C. (2009). Diverse Enterotoxin Gene Profiles Among Clonal Complexes of Staphylococcus aureus Isolates From the Bronx, New York. Appl. Environ. Microbiol. 75: 6839–6849.
§ Vasconcelos, N. G. and Cunha, M. L. R. S. (2010). Staphylococcal enterotoxins:
Molecular aspects and detection methods. Journal of Public Health and Epidemiology. 2(3): 29-42.
§ Veras, J. F.; Carmo, L. S.; Tong, L.C.; Shupp, J. W.; Cummings, C.; Dos Santos, D. A.; Cerqueira, M. M.; Cantini, A.; Nicoli, J.R. and Jett, M. (2008). A study of the enterotoxigenicity of coagulase-negative and coagulase-positive staphylococcal

isolates from food poisoning outbreaks in Minas Gerais, Brazil. Int. J. Infect. Dis. 12: 410–415.
§ von Eiff, E. C.; Becker, K.; Machka, K.; Stammer, H. and Peters, G. (2001). Nasal Carriage as a Source of Staphylococcus aureus Bacteremia. Study Group. N. Engl. J. Med. 344: 11–16.
§ Vos, P.; Garrity, G.; Jones, D.; Krieg, N. R.; Ludwig, W.; Rainey, F.A.; Schleifer, K. H. and Whitman, W. B. (2009). The Firmicutes In: Bergey’s manual of determinative bacteriology. 2nd ed. Volume 3. Springer, New York.
W
§ Waage, S.; Mork, T.; Roros, A.; Aasland, D.; Hunshamar, A. and Odegaard, A. (1999). Bacteria associated with clinical mastitis in dairy heifers. J. Dairy Sci. 82:712-719.
§ Wallin-Carlquist, N.; Cao, R.; Marta, D.; da Silva, A.S.; Schelin, J. and Radstrom, P. (2010). Acetic acid increases the phage encoded enterotoxin A expression in Staphylococcus aureus. BMC Microbiol. 10:147; PMID:20487538.
X
§ Xu, S.X. and McCormick, J. K. (2012). Staphylococcal superantigens in colonization and disease. Cellular and Infection Microbiology. 2:(52) 1-11.
Y § Yamaguchi, T.; Hayachi, T.; Takami, H.; Nakasome, K.; Ohnish, M.; Nakayama,
K.; Yamada, S.; Komatsuzawa, H. and Sugai, M. (2000). Phage conversion of esfoliative toxin A production in Staphylococcus aureus. Mol. Microbiol. 38: 694-705.
§ Yanagisawa, C.; Hanaki, H.; Natae, T. and Sunakawa, K. (2007). Neutralization of Staphylococcal Exotoxins in Vitro by Human-Origin Intravenous Immunoglobulin. J. Infect. Chemother. 13: 368–372.
§ Yang, X.; Buonpane, R. A.; Moza, B.; Rahman, A. K.; Wang, N.; Schlievert, P. M.;
McCormick, J. K.; Sundberg, E. J. and Kranz, D. M. (2008). Neutralization of Multiple Staphylococcal Superantigens by a Single-Chain Protein Consisting of Affinity-Matured, Variable Domain Repeats. J. Infect. Dis. 198: 344–348.

§ Yarwood, J. M.; McCormick, J. K. and Schlievert, P. M. (2001). Identification of a novel two-component regulatory system that acts in global regulation of virulence factors of Staphylococcus aureus. J. Bacteriol. 183:1113-1123.
§ Yoshizawa, Y.; Sakurada, J.; Sakurai, S.; Machida, K.; Kondo, I. and Masuda, S. (2000). An exfoliative toxin A-converting phage isolated from Staphylococcus aureus strain ZM. Microbiol. Immunol. 44: 189-191.
Z § Zamani, A.R.; Sadaghian, S.; Najafiemosleh, M.; Goudarzi, M.T. Yoysefi-
Moshouf, R. and Ghaderkhani, J. (2007). Detection of methicillin-resistance (mec-A) gene in Staphylococcus aureus strains by PCR and determination of antibiotic susceptibility. Ann. Microbiol. 57:273-276.
§ Zschock, M.; Kloppert, B.; Wolter, W.; Hamann, H.P. and Lammler, C. (2005). Pattern of enterotoxin genes seg, seh, sei and sej positive Staphylococcus aureus isolated from bovine mastitis. Vet Microbiol. 108:243–249.

Appendices

Appendix1 Reading table for API Staph. strip results Test Symbol Negative
result Positive result
Negative control 0 Red Yellow Acidification of D-glucose GLU Red Yellow Acidification of D-fructose FRU Red Yellow Acidification of D-mannose MNE Red Yellow Acidification of D-maltose MAL Red Yellow Acidification of D-lactose LAC Red Yellow Acidification of D-trehalose TRE Red Yellow Acidification of D-mannitol MAN Red Yellow Acidification of xylitol XLT Red Yellow Acidification of D-melibiose MEL Red Yellow Reduction of nitrate to nitrite NIT Colorless-
Light pink Red
Alkaline phosphatase PAL Yellow Violet Acetyl-methyl-carbinol production VP colorless Violet-Pink Acidification of raffinose RAF Red Yellow Acidification of xylose XYL Red Yellow Acidification of sucrose SAC Red Yellow Acidification of α-methyl-D-glucoside MGD Red Yellow Acidification of N-acetyl glucoseamine NAG Red Yellow Arginine dihydrolase ADH Yellow Orange-Red Urease URE Yellow Red-Violet

Appendix 2 Results of the biochemical test for identification of Staphylococcus spp. by API Staph System
Biochemical Test S. aureus S.chromogenes S. epidermidis Negative control 0 - - -
Acidification of D-glucose GLU
+ + +
Acidification of D-fructose FRU
+ + +
Acidification of D-mannose MNE
+ + +
Acidification of D-maltose MAL
+ + +
Acidification of D-lactose LAC
+ + +
Acidification of D-trehalose TRE
+ + -
Acidification of D-mannitol MAN
+ - -
Acidification of xylitol XLT
- - -
Acidification of D-melibiose MEL
- - -
Reduction of nitrate to nitrite NIT
+ + +
Alkaline phosphatase PAL + + + Acetyl-methyl-carbinol
production VP - - -
Acidification of raffinose RAF
- - -
Acidification of xylose XYL
- - -
Acidification of sucrose SAC
+ + +
Acidification of α-methyl-D-glucoside MDG
- - -
Acidification of N-acetyl glucoseamine NAG
+ + -
Arginine dihydrolase ADH + + + Urease URE + - -

Appendix 3 Result of S. aureus obtained by API Staph System

الخالصة ۲۰۰، وتم عزل ۲۰۱۱عينة من الحليب والجبن من األسواق المحلية في مدينة بغداد خالل العام ۳۰۰جمعت
Staphylococcusعزلة من المكورات العنقودية من العينات التي تم جمعها، وكانت المكورات العنقودية الذهبية
aureus تالها النوع %48عزلة ( ۹۷هي األكثر شيوعا في العينات إذ بلغ مجموع العزالت ،(S.chromogenes
). ۱۱عزلة (% S.epidermidis ۲۱ثم النوع (%41)عزلة 82
تم تحديد اختبار الحساسية لعزالت المكورات العنقودية الموجبة و المكورات العنقودية السالبة لإلنزيم المخثر
للبالزما لثالث من مضادات الحياة (المثيسيلين، التتراسايكلين، والفانكومايسين) باستخدام طريقة االنتشار باألقراص.
S. aureus) عزلة من ۱۹(% ۱۸) كانت مقاومة للمثيسيلين و ٤۷.S. aureus )%۸۲عزلة ۸۰أظهرت النتائج أن
) من المكورات ٦۲.۱۳عزالت مقاومة للفانكومايسين. أربع وستون عزلة (% ۸مقاومة للتتراسايكلين بينما كانت
٥سايكلين و ) مقاومة للتترا۲۷.۱۸عزلة (% ۲۸العنقودية السالبة لإلنزيم المخثر للبالزما كانت مقاومة للمثيسيلين و
مقاومة للفانكومايسين. (%4.85)عزالت
ومن عينات الحليب المزروعة على األوساط الزرعيةمن عزالت المكورات العنقودية DNA)استخلص الدنا (
) باستخدام الطرائق الفيزيائية دون استخدام عدد االستخالص على األوساط الزرعيةوالجبن مباشرة (غير المزروعة
ة.الجاهز
للتحري عن الجين المشفر لإلنزيم المخثر monoplex PCR)المنفرد ( أستخدمت تقنية تفاعل سلسلة البلمرة
.) bp ۷۳۰امتلكت هذا الجين ( S. aureus) من ۷۸.۳٥عزلة (% ۷٦)، وأظهرت النتائج أن coaللبالزما (
تقنية تفاعل سلسلة باستخدام mecAو femAات تم التحري المحددات الجزيئية لمقاومة المثيسيلين وهي جين
S. aureus) وللتفريق بين -mecA) والحساسة له (mecAالمنفرد للتحري عن العزالت المقاومة للمثيسيلين (+ البلمرة
+)femA) و المكورات العنقودية السالبة لإلنزيم المخثر للبالزما (-femA.(
S. aureus، إذ إن هذا الجين هو مؤشر نوعي للنوع femA) حاملة لجين S. aureus %)۹۸.۹٦عزلة ۹٦كانت
mecAمن المكورات العنقودية المقاومة للمثيسيلين كانت حاملة لجين (%93.81)عزلة ۹۱.كما وأظهرت النتائج أن
للبالزما. من المكورات العنقودية السالبة لإلنزيم المخثر (%67)عزلة ۷۰بينما وجد هذا الجين في
للتحري عن وجود أربعة جينات تقليدية للسموم multiplex PCR)( المتعدد استخدمت تقنية تفاعل سلسلة البلمرة
من المكورات (%76.5)عزلة ۱٥۳بتفاعل واحد وأظهرت النتائج أن sedو secو sebو seaالمعوية العنقودية هي
حاملة لواحد أو أكثر من S.aureus عزلة(%62.09) 95 وم المعوية ، كانتالعنقودية كان لها القابلية إلنتاج السم
ساللة من المكورات العنقودية السالبة لإلنزيم المخثر ٥۸جينات السموم التي تم اختبارها. أظهرت نتائج هذه الدراسة أن
لسالالت الحاملة لجينات السموم. وقد كان من مجموع ا ۳۷.۹للبالزما كان لها القابلية إلنتاج السموم المعوية إذ شكلت %
هو الجين األكثر انتشارا بين الجينات المشفرة للسموم التي تم اختبارها في هذه الدراسة. إذ وجد بنسبة seaالجين
نادر إذ وجد بنسبة secبينما كان تواجد الجين %12.5بنسبة sebثم الجين %34.73بنسبة sed، ثم الجين 51.85%
0.92%.

عزلة من المكورات العنقودية كانت حاملة لنوع واحد من الجينات (%66.66)۱۰۲أظهرت نتائج هذه الدراسة أن
و sea امتلكت اثنين من هذه الجينات ؛ وأغلبها كانت (%25.5)عزلة ۳۹األربعة المشفرة للسموم التي تم اختبارها، و
sed عزلة من المكورات العنقودية امتلكت ثالث جينات مشفرة للسموم المعوية ۱۲وsea وseb وsed.
واظهرت عن الفعالية الحيوية للسموم المعوية للمكورات العنقودية التحري في الرضيعة الفئران تجريع اختبار تم
المكورات العنقودية الموجبة بينت النتائج أن كال من .اعزلة انتجت سموما معوية فاعلة احيائي(%65.5) ۱۳۱النتائج ان
من المجموع S. aureus 65.64% و السالبة لإلنزيم المخثر للبالزما كانت منتجة للسموم المعوية، وقد شكل النوع
لمعوية بنسبة الكلي للعزالت المنتجة بينما كانت المكورات العنقودية السالبة لإلنزيم المخثر للبالزما منتجة للسموم ا
، وتم اختبار ثبات هذه السموم في ظروف الحرارة العالية وأكدت النتائج إن هذه السموم المعوية ثابتة ۳٥.%۳٤
. C°100دقيقة في درجة حرارة ۳۰بالحرارة إذ بقيت فعالة إحيائيا حتى بعد تعرضها للتسخين لمدة
المتعدد للتحري عن وجود الجينات المشفرة اعل سلسلة البلمرةتقنية تفقورنت النتائج التي تم الحصول عليها من إجراء
عن انتاجية التحري في الرضيعة الفئران تجريع اختبار ألربع من السموم المعوية مع تلك التي تم الحصول عليها من
من %65.5من المكورات العنقودية حاملة لجينات السموم المعوية بينما كانت %76.5السموم المعوية وقد وجد أن
هذه العزالت منتجة لهذه السموم.

جمهورية العراق وزارة التعليم العالي والبحث العلمي
جامعة بغداد / كلية العلوم
المكورات التحري عن جينات السموم المعوية في
العنقودية المعزولة من الحليب والجبن
أطروحة مقدمة الى
كلية العلوم / جامعة بغداد وهي جزء من متطلبات نيل درجة الدكتوراه فلسفة علم االحياء المجهرية/ التسمم الغذائي
من
مروى حميد مطشر الخفاجي
۲۰۰۲جامعة بغداد –كلية العلوم -الحياة قسم علوم -بكالوريوس/علوم حياة / احياء مجهرية
۲۰۰۸جامعة بغداد –كلية العلوم -قسم علوم الحياة -ماجستير/علوم حياة احياء مجهرية
بإشراف
أ.د.مي طالب فليح
1434 رجب 2013 أيار


















