Description of joint movements in human and non-human primate locomotion using Fourier analysis
16
ORIGINAL ARTICLE Description of joint movements in human and non-human primate locomotion using Fourier analysis David Webb William Anthony Sparrow Received: 29 May 2006 / Accepted: 31 January 2007 / Published online: 22 May 2007 Ó Japan Monkey Centre and Springer 2007 Abstract To describe and help interpret joint movements in various forms of primate locomotion, we explored the use of Fourier analysis to represent changing joint angles as a series of sine and cosine curves added together to approximate the raw angular data. Results are presented for four joints (shoulder, elbow, hip and knee) with emphasis on the shoulder, and for five types of locomotion (catarhine primate quadrupedal walking, human hands- and-feet creeping and hands-and-knees creeping, and human walking and running). Fourier analysis facilitates functional interpretation of the angles of all four joints, by providing average joint angles and an indication of the number of peaks and troughs in the angular data. The description of limb movements also afforded us the opportunity to compare human and other catarhine joint angles, and we interpret the Fourier results in terms of locomotor posture and type. In addition, the shoulder data are useful for determination of some aspects of interlimb coordination. Non-human primates walking quadrupedally and humans creeping on hands and knees generally evince diagonal couplets interlimb coordination, in which the hand on one side strikes the substrate at about the same time as the contralateral foot or knee. Furthermore, human walking and running seem to follow a similar pattern, as indicated by Fourier analysis. From our data it is concluded that human bipedal gaits are qualitatively similar to diagonal couplets gaits in other primates, but quite different from the lateral couplets gaits used by many non-primate mammals. A number of other benefits of Fourier analysis in primate locomotion studies are also discussed. These include the ability to make statistical comparisons among various types of limb movements in a wide variety of species, a simple archival technique for limb movement data, and a greater understanding of the variability of locomotor movements. Keywords Joint angle Interlimb coordination Locomotion Fourier analysis Introduction Fourier analysis The purpose of this study was to explore the technique of Fourier analysis to describe, in quantifiable terms, the joint movement patterns of primates, including humans. A wide variety of techniques exists for the quantitative description of various aspects of gait. Some examples are: duty factor (from footfall diagrams); speed (both absolute and relative to body size); ground reaction force; cadence; various Froude numbers; and foot angle (from footprints). In the case of joint movements, many authors have used body segment models and joint angle data (see, for example, D’Aou ˆt et al. 2002; Isler 2005). Angular data analysis has suffered from the fact that a series of joint angles is difficult to reduce to a few simple numbers for statistical compar- ison. It is easy enough to describe the angles through which any joint passes, using a graph of angle versus time. Fur- thermore, some standardization of the data can be achieved by replacing time with a relative measure of time, such as percent of stride cycle (see Figs. 1, 3, 5), and data can be D. Webb (&) Department of Anthropology and Sociology, Kutztown University, Kutztown, PA 19530, USA e-mail: [email protected] W. A. Sparrow School of Health Sciences, Faculty of Health and Behavioural Sciences, Deakin University, Burwood, Victoria 3125, Australia 123 Primates (2007) 48:277–292 DOI 10.1007/s10329-007-0043-4
-
Upload
david-webb -
Category
Documents
-
view
213 -
download
0
Transcript of Description of joint movements in human and non-human primate locomotion using Fourier analysis

ORIGINAL ARTICLE
Description of joint movements in human and non-human primatelocomotion using Fourier analysis
David Webb Æ William Anthony Sparrow
Received: 29 May 2006 / Accepted: 31 January 2007 / Published online: 22 May 2007
� Japan Monkey Centre and Springer 2007
Abstract To describe and help interpret joint movements
in various forms of primate locomotion, we explored the
use of Fourier analysis to represent changing joint angles as
a series of sine and cosine curves added together to
approximate the raw angular data. Results are presented
for four joints (shoulder, elbow, hip and knee) with
emphasis on the shoulder, and for five types of locomotion
(catarhine primate quadrupedal walking, human hands-
and-feet creeping and hands-and-knees creeping, and
human walking and running). Fourier analysis facilitates
functional interpretation of the angles of all four joints, by
providing average joint angles and an indication of the
number of peaks and troughs in the angular data. The
description of limb movements also afforded us the
opportunity to compare human and other catarhine joint
angles, and we interpret the Fourier results in terms of
locomotor posture and type. In addition, the shoulder data
are useful for determination of some aspects of interlimb
coordination. Non-human primates walking quadrupedally
and humans creeping on hands and knees generally evince
diagonal couplets interlimb coordination, in which the hand
on one side strikes the substrate at about the same time as
the contralateral foot or knee. Furthermore, human walking
and running seem to follow a similar pattern, as indicated
by Fourier analysis. From our data it is concluded that
human bipedal gaits are qualitatively similar to diagonal
couplets gaits in other primates, but quite different from the
lateral couplets gaits used by many non-primate mammals.
A number of other benefits of Fourier analysis in primate
locomotion studies are also discussed. These include the
ability to make statistical comparisons among various types
of limb movements in a wide variety of species, a simple
archival technique for limb movement data, and a greater
understanding of the variability of locomotor movements.
Keywords Joint angle � Interlimb coordination �Locomotion � Fourier analysis
Introduction
Fourier analysis
The purpose of this study was to explore the technique of
Fourier analysis to describe, in quantifiable terms, the joint
movement patterns of primates, including humans. A wide
variety of techniques exists for the quantitative description
of various aspects of gait. Some examples are: duty factor
(from footfall diagrams); speed (both absolute and relative
to body size); ground reaction force; cadence; various
Froude numbers; and foot angle (from footprints). In the
case of joint movements, many authors have used body
segment models and joint angle data (see, for example,
D’Aout et al. 2002; Isler 2005). Angular data analysis has
suffered from the fact that a series of joint angles is difficult
to reduce to a few simple numbers for statistical compar-
ison. It is easy enough to describe the angles through which
any joint passes, using a graph of angle versus time. Fur-
thermore, some standardization of the data can be achieved
by replacing time with a relative measure of time, such as
percent of stride cycle (see Figs. 1, 3, 5), and data can be
D. Webb (&)
Department of Anthropology and Sociology,
Kutztown University, Kutztown, PA 19530, USA
e-mail: [email protected]
W. A. Sparrow
School of Health Sciences,
Faculty of Health and Behavioural Sciences,
Deakin University, Burwood, Victoria 3125, Australia
123
Primates (2007) 48:277–292
DOI 10.1007/s10329-007-0043-4

further abstracted by dividing the stride cycle into stance
and support phases and plotting joint angle versus percent
of each phase (Fleagle et al. 1981; Hirasaki et al. 2000;
Isler 2005). However, the overall distribution of the data,
the ‘‘shape of the graph’’, cannot be quantified by these
means, leaving scientists to compare joint movement pat-
terns only in qualitative terms or only at specific points in
the stride.
To move beyond qualitative descriptions and compari-
sons, to quantify the overall distribution of gonial data, we
chose to investigate Fourier analysis. In 1822, Jean-Bap-
tiste-Joseph Fourier, a French mathematician and physicist,
published Theorie analytique de la chaleur (Analytical
Theory of Heat). Fourier’s book defines the series of sines
and cosines today known as Fourier series, and describes
their use in studying heat conductance (Bell 1937). While
there were difficulties with Fourier’s proof of his new
theorem, these were subsequently overcome by other
mathematicians and the use of Fourier analysis is wide-
spread today in such fields as electronics and the physics of
sound where transformations of the temporal distribution of
data are desirable (Richmond 1972).
With the methods of Fourier analysis, we hoped to be
able to describe and interpret joint movement patterns in
primates in quantitative terms and more thoroughly than
has been done previously. Quantitative descriptions of ‘‘the
shape of the graph’’ should allow the use of statistical
comparisons among individuals and among species, and
should lead to a greater understanding of the variability of
limb movements. In this way, Fourier analysis should help
elucidate several areas of joint analysis, including intra-
and inter-limb coordination and substrate accommodation.
We also hoped to develop a simple method of archiving
primate movement patterns, using many fewer descriptors
than would be necessary with traditional joint angle data-
points.
In order to describe joint angles and test for joint angle
similarity between humans and other primates, Fourier
analysis was used in this study to analyze the joint angle
data obtained from videotape records. Fourier analysis has
previously been used in physical anthropology to describe
the cross-sectional shapes of hominoid femora and maca-
que skulls and human skulls (Lestrel et al. 1977, 1993,
2005), to analyze sagittal movements of the head and trunk
of walking humans (Cappozzo 1981), and to describe the
extent and type of ‘‘meandering’’ while trying to walk in a
straight line (Uetake 1992). Fourier analysis allows the data
to be represented, not as measurements over time, but as
the sum of sine and cosine curves of various frequencies
and magnitudes. We can therefore approximate any cur-
vilinear data plot by means of a Fast Fourier Transform
(FFT). The analysis technique, as applied to the joint angle
data presented below, begins with a series of angles mea-
sured at regular intervals during the stride cycle. The FFT
program then calculates the magnitudes of a number of sine
and cosine curves which, when summed, approximate the
distribution of the data (Fig. 2). The magnitude of each
wave is expressed as a coefficient by which the height of
the wave is multiplied. These Fourier coefficients, properly
applied, will produce a curve of the correct shape. Then,
the average angle must be added to the resultant curve in
order to match the scale of the original data.
For the purposes of the present study, one advantage of
the technique is that FFTs allow comparison of the upper
limb movements in walking humans with the forelimbs of
quadrupedal primates. This is not possible using footfall
analysis such as revealed in ground contact diagrams
(Burnside 1927; Hildebrand 1967; Sparrow 1989), since
human upper limbs never contact the ground during normal
walking. However, Fourier analysis can be applied to bi-
pedal locomotion, if a standard for the beginning and end
of each stride is used. Thus, the most commonly applied
temporal standard, contact of the right hind limb/lower
limb, can be used in either bipedal or quadrupedal forms
of primate terrestrial locomotion. Fourier transforms also
describe each joint’s contribution to limb movement, rather
than just the results of those joints’ movements on ground
contact, and they allow the production of datasets of
average joint movements for many strides and many
individuals. In addition, quantitative ontogenetic data can
be analyzed, allowing greater elucidation of locomotor
development in various primate species (e.g., Lasko-
Macarthey et al. 1990; Kimura et al. 2005; Shapiro and
Raichlen 2006). Finally, and most importantly, FFTs
quantify angular data in a standardized form, allowing the
use of statistical procedures to compare joint movements
Fig. 1a, b Footfall diagrams illustrating coupling and sequence. LHLeft hind limb, LF left forelimb, RH right hind limb, RF right
forelimb. Thick lines indicate the time during which a given limb is in
contact with the substrate. a Diagonal couplets/diagonal sequence
gait. b Lateral couplets/lateral sequence gait
278 Primates (2007) 48:277–292
123

among different strides of the same individual, among
different individuals, and even among different species.
Fourier analysis is therefore an appropriate method of
assessing the similarities and differences among human
and non-human primate modes of locomotion.
Interlimb coordination
Both within and between species, the timing of the char-
acteristic regular movements of the limbs in walking can be
categorized and compared using two footfall parameters,
sequence and coupling. Sequence refers to the order in
which the forefeet follow the hindfeet in contacting the
ground. A lateral sequence pattern is when the right or left
forefoot strikes the ground immediately following the
hindfoot on the same side. Diagonal sequence gait occurs
when forefoot contact follows contralateral hindfoot con-
tact. Coupling refers to the footfalls of a forefoot and a hind
foot being related in time as a pair. Hildebrand (1967)
defined two types of coupling: lateral couplets and diagonal
couplets, as follows: ‘‘Lateral-couplets gaits have the
footfalls of fore and hindfeet on the same side of the body
related in time as a pair. Diagonal-couplets gaits have the
footfalls of fore and hindfeet on the opposite sides of the
body related in time as a pair’’ (Hildebrand 1967, p 119).
Sequence and coupling are illustrated in Fig. 1 using
footfall diagrams.
In a review of non-human primate gaits, it becomes
clear that the general pattern among great apes (Hildebrand
1967; Vilensky and Larson 1989), Old World monkeys
Fig. 2a–d Process of approximating angular data with a Fast Fourier
Transform (FFT). In each part of the figure, the original data are
represented by the open circles connected by the thickest line. As the
approximation progresses, the current approximation in each part of
the figure is represented by open squares connected by a medium line;
the previous approximation is shown with open triangles; and the
newly added curve is shown with open diamonds. a The original data,
plotted as joint angle over time, with the first approximation made
using a simple sine curve with one peak and one trough within the
cycle (0–350 on the Time scale). The height of the peak and the depth
of the trough are determined by the Fourier coefficient of the first sine
(5.13 in this case). b The approximation is refined by adding a simple
cosine curve (open diamonds), multiplied by the appropriate
coefficient (0.63). This cosine curve is added to the sine curve
in part a (now shown as open triangles), resulting in a new
approximation as indicated by the open squares. c In the interests of
saving time and space, two new curves are simultaneously added to
the previous approximation. Thus the current approximation is the
sum of the first sine and cosine curves of part b and the second sine
and cosine curves combined (open diamonds). Note that the combined
second sine and cosine curve has twice the frequency of the first order
curves, thereby producing a curve with two peaks and two troughs.
This permits the FFT to add finer details to the approximation. d The
final approximation, using fourth order sine and cosine curves,
produces a very good match to the original angular data. Subse-
quently, even higher order sine and cosine curves can be added to
match the original data very precisely. Note that this figure was made
using imaginary, simplified data that are not from primate locomo-
tion. Therefore, ‘‘time’’ and ‘‘angle’’ are not real units
Primates (2007) 48:277–292 279
123
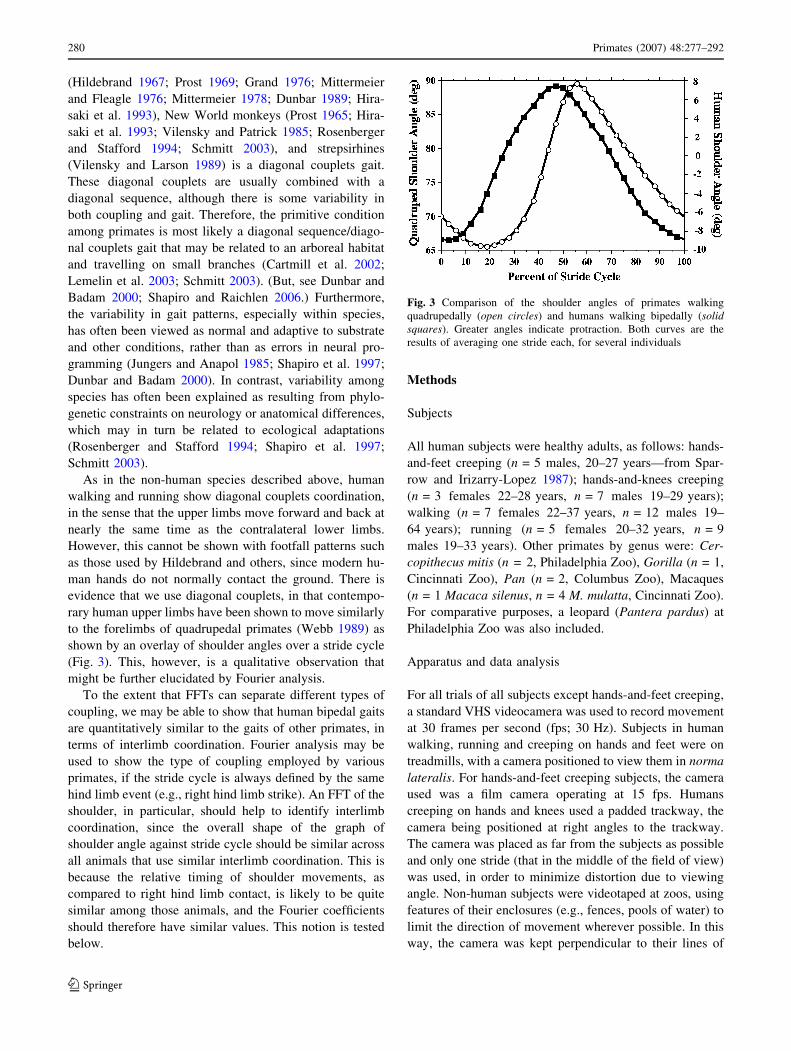
(Hildebrand 1967; Prost 1969; Grand 1976; Mittermeier
and Fleagle 1976; Mittermeier 1978; Dunbar 1989; Hira-
saki et al. 1993), New World monkeys (Prost 1965; Hira-
saki et al. 1993; Vilensky and Patrick 1985; Rosenberger
and Stafford 1994; Schmitt 2003), and strepsirhines
(Vilensky and Larson 1989) is a diagonal couplets gait.
These diagonal couplets are usually combined with a
diagonal sequence, although there is some variability in
both coupling and gait. Therefore, the primitive condition
among primates is most likely a diagonal sequence/diago-
nal couplets gait that may be related to an arboreal habitat
and travelling on small branches (Cartmill et al. 2002;
Lemelin et al. 2003; Schmitt 2003). (But, see Dunbar and
Badam 2000; Shapiro and Raichlen 2006.) Furthermore,
the variability in gait patterns, especially within species,
has often been viewed as normal and adaptive to substrate
and other conditions, rather than as errors in neural pro-
gramming (Jungers and Anapol 1985; Shapiro et al. 1997;
Dunbar and Badam 2000). In contrast, variability among
species has often been explained as resulting from phylo-
genetic constraints on neurology or anatomical differences,
which may in turn be related to ecological adaptations
(Rosenberger and Stafford 1994; Shapiro et al. 1997;
Schmitt 2003).
As in the non-human species described above, human
walking and running show diagonal couplets coordination,
in the sense that the upper limbs move forward and back at
nearly the same time as the contralateral lower limbs.
However, this cannot be shown with footfall patterns such
as those used by Hildebrand and others, since modern hu-
man hands do not normally contact the ground. There is
evidence that we use diagonal couplets, in that contempo-
rary human upper limbs have been shown to move similarly
to the forelimbs of quadrupedal primates (Webb 1989) as
shown by an overlay of shoulder angles over a stride cycle
(Fig. 3). This, however, is a qualitative observation that
might be further elucidated by Fourier analysis.
To the extent that FFTs can separate different types of
coupling, we may be able to show that human bipedal gaits
are quantitatively similar to the gaits of other primates, in
terms of interlimb coordination. Fourier analysis may be
used to show the type of coupling employed by various
primates, if the stride cycle is always defined by the same
hind limb event (e.g., right hind limb strike). An FFT of the
shoulder, in particular, should help to identify interlimb
coordination, since the overall shape of the graph of
shoulder angle against stride cycle should be similar across
all animals that use similar interlimb coordination. This is
because the relative timing of shoulder movements, as
compared to right hind limb contact, is likely to be quite
similar among those animals, and the Fourier coefficients
should therefore have similar values. This notion is tested
below.
Methods
Subjects
All human subjects were healthy adults, as follows: hands-
and-feet creeping (n = 5 males, 20–27 years—from Spar-
row and Irizarry-Lopez 1987); hands-and-knees creeping
(n = 3 females 22–28 years, n = 7 males 19–29 years);
walking (n = 7 females 22–37 years, n = 12 males 19–
64 years); running (n = 5 females 20–32 years, n = 9
males 19–33 years). Other primates by genus were: Cer-
copithecus mitis (n = 2, Philadelphia Zoo), Gorilla (n = 1,
Cincinnati Zoo), Pan (n = 2, Columbus Zoo), Macaques
(n = 1 Macaca silenus, n = 4 M. mulatta, Cincinnati Zoo).
For comparative purposes, a leopard (Pantera pardus) at
Philadelphia Zoo was also included.
Apparatus and data analysis
For all trials of all subjects except hands-and-feet creeping,
a standard VHS videocamera was used to record movement
at 30 frames per second (fps; 30 Hz). Subjects in human
walking, running and creeping on hands and feet were on
treadmills, with a camera positioned to view them in norma
lateralis. For hands-and-feet creeping subjects, the camera
used was a film camera operating at 15 fps. Humans
creeping on hands and knees used a padded trackway, the
camera being positioned at right angles to the trackway.
The camera was placed as far from the subjects as possible
and only one stride (that in the middle of the field of view)
was used, in order to minimize distortion due to viewing
angle. Non-human subjects were videotaped at zoos, using
features of their enclosures (e.g., fences, pools of water) to
limit the direction of movement wherever possible. In this
way, the camera was kept perpendicular to their lines of
Fig. 3 Comparison of the shoulder angles of primates walking
quadrupedally (open circles) and humans walking bipedally (solidsquares). Greater angles indicate protraction. Both curves are the
results of averaging one stride each, for several individuals
280 Primates (2007) 48:277–292
123

progression. From the videos, it was determined that hands-
and-feet creeping subjects used lateral couplets gaits, while
hands-and-knees subjects and non-human primates used
diagonal couplets.
Using a computer with a video ‘‘frame-grabber’’
(ComputerEyes/RT, Digital Vision, Dedham, MA) and a
VHS video player, each frame of each stride was opened to
the computer screen and the x-y coordinates of the elbow,
shoulder, hip, knee, wrist and ankle were digitized using a
program (MouseSpot) developed in-house. Using additional
customized software the x-y coordinate data were smoothed
according to an algorithm wherein the coordinates for any
given frame were averaged with those of the two preceding
and two following frames, weighted as follows: 1-2-10-2-1.
The hands-and-feet creeping data were obtained by manual
digitizing of films taken at 15 fps as described by Sparrow
and Irizarry-Lopez (1987). Subsequently, the angles at the
hip, knee, shoulder and elbow were calculated for each
frame and written to a file for Fourier analysis. The methods
for measuring the angles are shown in Fig. 4. FFTs were
then performed by Mathcad� 3.1, and the resulting Fourier
coefficients analyzed using the statistical package, Stat-
view� 4.02 (www.statview.com).
The cosine and sine coefficients from the Fourier anal-
ysis are the magnitudes by which each cosine or sine curve
must be multiplied, so that the sum of all the curves
approximates the original distribution of angle versus
percent of stride cycle. The coefficients presented in the
Results are named according to the frequency of the sine or
cosine by which they are multiplied. Hence, the coefficient,
cos2, is multiplied by a cosine curve with two peaks and
two troughs during the stride cycle. This second order
cosine curve is therefore one with twice the frequency of a
normal cosine curve. The coefficient, sin3, is multiplied by
a sine curve of three times the normal frequency, and hence
three peaks and three troughs in one cycle. Higher order
coefficients, e.g., sin4 and cos5, describe more detailed
fluctuations in the original angular data, whereas lower
order coefficients (e.g., sin1, cos2) describe only the gen-
eral shape of the data. Fourier coefficients for the various
modes of locomotion were compared using Student’s t-test,
after it was confirmed that the data were normally dis-
tributed. Specifically, the resulting Fourier coefficients
were not skewed, being evenly distributed about the mean.
This was tested not only within groups of subjects, but also
among 20 strides of the same subject.
Results
Tables 1, 2, 3 and 4 present the coefficients of the FFTs for
each joint. The number of subjects in each group (n) is
followed by a0, the joint angle averaged over the entire
stride cycle for each individual and then for all individuals
in the group. From the data in Tables 1–4 an average curve
for the shoulder angles of the five groups was reconstructed
(Fig. 5). The shoulder angle is the best indicator of overall
forelimb movement, and therefore provides a qualitative
comparison of interlimb coordination among the five
groups. The angles in Fig. 5 were obtained by adding all
the sines and cosines indicated, and adding a0, the average
measured joint angle. For each point along the x-axis
(percentage of stride cycle) the reconstructed joint angle (y-
value) was obtained using the formula:
X½cosðX � 1Þ � cos 1þ cosðX � 2Þ � cos 2
þ cosðX � 3Þ � cos 3þ cosðX � 4Þ � cos 4
þ cosðX � 5Þ � cos 5þ sinðX � 1Þ � sin 1
þ sinðX � 2Þ � sin 2þ sinðX � 3Þ � sin 3
þ sinðX � 4Þ � sin 4þ sinðX � 5Þ � sin 5þ a0�
where ‘‘X’’ is the value on the x-axis, cos2 is the second
cosine coefficient (from Table 3), sin3 is the third sine
coefficient, etc. Thus, the second term in the summation,
cos(X*2)*cos2, would read: ‘‘the cosine of twice the x-
value, multiplied by the second cosine coefficient’’.
Each graph in Fig. 5 has only one peak and one trough,
indicating the relatively simple protraction and retraction
of the limb that occurs in each stride cycle. In human
walking and running, hands-and-feet creeping, and cata-
rhine primate walking, the peak that denotes maximum
protraction occurs about 40–50% after the trough denoting
maximum retraction. Maximum protraction and retraction
Fig. 4 Stick figures generated from videotaped quadrupedal and
bipedal subjects. Digitized joints: 1 wrist, 2 elbow, 3 shoulder, 4 hip,
5 knee, 6 ankle. Only the side of the body facing the camera was
digitized. Angles calculated were as shown: E elbow, S shoulder, Hhip and K knee. Note that, by the definition used here, the shoulder
angle of the bipedal figure would be negative, positive values being
ventral to the torso line, 34
Primates (2007) 48:277–292 281
123

Ta
ble
1A
ver
age
Fo
uri
erco
effi
cien
tsfo
rth
esh
ou
lder
ang
les
of
no
n-h
um
anp
rim
ates
and
hu
man
s.F
rom
thes
ed
ata,
the
aver
age
curv
eso
fsh
ou
lder
ang
lev
ersu
sp
erce
nt
stri
de
can
be
dra
wn
as
sho
wn
inF
ig.
4
Gro
up
na0
cos
1co
s2
cos
3co
s4
cos
5si
n1
sin
2si
n3
sin
4si
n5
Pri
mat
ew
alkin
g10
75.4
20
(9.3
43)a
–7.6
31
(2.6
56)
2.5
30
(1.6
69)
–0.8
81
(0.8
26)
0.4
44
(0.4
63)
0.0
13
(0.3
64)
–7.7
04
(3.7
83)
1.6
75
(2.4
20)
–0.5
50
(1.2
12)
0.2
83
(0.3
95)
0.0
31
(0.2
81)
Hum
ancr
eepin
g
(han
ds-
and-k
nee
s)
10
69.9
88
(6.9
70)
–9.8
89
(2.3
62)
0.2
17
(2.7
11)
1.0
10
(0.6
39)
–0.1
75
(0.4
77)
–0.1
13
(0.1
44)
–2.2
26
(3.2
36)
–4.3
82
(1.3
13)
0.2
99
(1.3
07)
0.3
22
(0.5
58)
–0.0
14
(0.2
32)
Hum
ancr
eepin
g
(han
ds-
and-f
eet)
599.6
61
(9.5
04)
0.8
32
(3.6
74)
–2.0
41
(1.9
92)
–1.5
64
(1.4
87)
–0.1
41
(0.5
99)
0.0
74
(0.2
64)
11.0
86
(1.8
64)
1.9
50
(1.4
34)
0.4
02
(1.0
55)
–0.3
78
(0.3
86)
–0.2
28
(0.3
44)
Hum
anw
alkin
g19
–1.2
40
(4.8
31)
–8.0
47
(2.7
34)
0.2
41
(0.5
06)
–0.0
07
(0.2
03)
0.1
53
(0.2
09)
–0.0
08
(0.1
81)
0.8
42
(1.9
82)
–0.6
00
(0.6
59)
–0.1
53
(0.3
38)
–0.1
07
(0.1
80)
0.0
53
(0.1
53)
Hum
anru
nnin
g14
–10.2
14
(6.9
64)
–8.4
89
(2.4
18)
–1.0
38
(0.3
69)
0.0
31
(0.1
92)
0.0
15
(0.2
23)
0.1
22
(0.1
57)
6.7
95
(3.2
13)
0.4
14
(0.6
98)
–0.1
57
(0.2
22)
–0.1
17
(0.2
99)
0.1
28
(0.2
02)
aS
tan
dar
dd
evia
tio
ns
are
giv
enin
par
enth
eses
Ta
ble
2A
ver
age
Fo
uri
erco
effi
cien
tsfo
rth
eel
bo
wan
gle
so
fn
on
-hu
man
pri
mat
esan
dh
um
ans
Gro
up
na0
cos
1co
s2
cos
3co
s4
cos
5si
n1
sin
2si
n3
sin
4si
n5
Pri
mat
ew
alkin
g10
154.5
94
(8.9
17)a
4.9
78
(2.1
19)
–1.6
37
(2.7
68)
–0.0
95
(1.9
53)
0.7
96
(0.9
52)
1.0
32
(2.9
97)
0.5
65
(4.3
32)
4.4
89
(2.9
93)
–0.9
77
(3.8
39)
0.5
10
(0.7
42)
0.1
17
(0.4
36)
Hum
ancr
eepin
g
(han
ds-
and-k
nee
s)
10
160.3
75
(4.9
18)
–2.2
75
(1.6
60)
3.2
32
(1.5
82)
0.6
65
(0.9
48)
–0.2
32
(0.8
71)
–0.0
65
(0.5
20)
–5.2
64
(2.1
46)
–1.5
57
(2.0
88)
1.4
41
(1.6
67)
0.5
08
(0.7
13)
0.0
99
(0.4
00)
Hum
ancr
eepin
g
(han
ds-
and-f
eet)
5162.5
92
(6.5
04)
–1.0
49
(2.0
38)
–0.5
10
(1.6
82)
–0.9
91
(1.0
75)
–0.6
91
(0.5
97)
–0.0
78
(0.4
49)
0.8
66
(1.3
24)
–0.8
63
(1.1
21)
–0.2
61
(0.9
33)
0.0
85
(0.5
96)
0.0
44
(0.1
68)
Hum
anw
alkin
g19
159.7
36
(6.0
31)
6.8
12
(3.6
81)
–1.2
64
(0.9
07)
0.0
48
(0.4
28)
–0.0
69
(0.2
93)
0.0
15
(0.2
25)
0.7
19
(2.2
07)
0.1
04
(1.2
50)
0.1
43
(0.5
88)
–0.0
81
(0.1
84)
0.0
70
(0.2
25)
Hum
anru
nnin
g14
121.1
55
(23.9
93)
1.8
40
(40.8
02)
–1.3
87
(10.6
56)
0.0
87
(0.5
52)
–0.1
26
(0.3
29)
0.1
04
(0.1
70)
–2.1
52
(3.4
25)
2.0
93
(1.2
25)
–0.0
66
(0.4
85)
–0.2
87
(0.4
87)
0.1
57
(0.3
36)
aS
tan
dar
dd
evia
tio
ns
are
giv
enin
par
enth
eses
282 Primates (2007) 48:277–292
123

Ta
ble
3A
ver
age
Fo
uri
erco
effi
cien
tsfo
rth
eh
ipan
gle
so
fn
on
-hu
man
pri
mat
esan
dh
um
ans
Gro
up
na0
cos
1co
s2
cos
3co
s4
cos
5si
n1
sin
2si
n3
sin
4si
n5
Pri
mat
ew
alkin
g9
83.0
01
(8.0
51)a
–11.9
10
(2.5
98)
0.2
69
(1.6
67)
0.8
89
(0.7
05)
0.0
72
(0.4
93)
0.1
20
(0.3
37)
–2.3
35
(1.8
13)
3.4
00
(0.8
74)
0.2
05
(0.3
80)
–0.3
82
(0.4
98)
0.0
93
(0.3
99)
Hum
ancr
eepin
g
(han
ds-
and-k
nee
s)
10
103.5
52
(7.6
76)
–11.1
34
(1.9
98)
–0.3
06
(1.3
70)
–0.0
89
(0.6
04)
0.1
05
(0.2
30)
0.0
43
(0.1
95)
–0.4
04
(1.7
68)
3.0
66
(0.8
98)
0.1
75
(0.5
14)
0.0
84
(0.4
21)
0.0
08
(0.3
26)
Hum
ancr
eepin
g
(han
ds-
and-f
eet)
577.5
59
(11.2
72)
–13.7
90
(2.3
43)
–1.2
51
(1.3
47)
0.6
29
(0.9
24)
–0.2
13
(0.3
03)
–0.0
66
(0.4
36)
–5.4
11
(2.7
97)
1.8
58
(1.5
85)
0.9
35
(0.7
14)
0.4
06
(0.6
04)
0.2
48
(0.2
72)
Hum
anw
alkin
g19
170.7
56
(5.2
19)
–7.5
81
(1.5
02)
1.5
03
(0.7
09)
–0.3
98
(0.3
55)
–0.1
39
(0.3
07)
0.0
72
(0.2
65)
2.7
14
(1.2
03)
0.6
27
(0.9
40)
–0.3
53
(0.4
44)
–0.0
10
(0.2
67)
–0.2
48
(0.2
99)
Hum
anru
nnin
g14
164.6
73
(4.2
65)
–5.7
39
(1.3
63)
1.8
17
(0.7
90)
0.1
81
(0.5
17)
–0.0
02
(0.3
09)
–0.0
76
(0.1
44)
4.0
08
(1.1
74)
–1.1
83
(1.2
56)
–0.2
82
(0.3
98)
0.1
13
(0.1
76)
–0.1
94
(0.2
98)
aS
tan
dar
dd
evia
tio
ns
are
giv
enin
par
enth
eses
Ta
ble
4A
ver
age
Fo
uri
erco
effi
cien
tsfo
rth
ek
nee
ang
les
of
no
n-h
um
anp
rim
ates
and
hu
man
s
Gro
up
na0
cos
1co
s2
cos
3co
s4
cos
5si
n1
sin
2si
n3
sin
4si
n5
Pri
mat
ew
alkin
g10
136.6
13
(10.7
12)a
1.0
73
(3.6
12)
6.5
50
(1.7
97)
2.3
69
(1.4
09)
0.1
56
(0.8
87)
–0.0
03
(0.3
56)
5.9
93
(2.1
28)
1.2
26
(1.7
80)
–2.1
82
(1.0
98)
–1.2
27
(0.9
90)
–0.3
29
(0.5
35)
Hum
ancr
eepin
g
(han
ds-
and-k
nee
s)
10
82.8
66
(6.7
21)
–12.3
08
(2.6
13)
0.2
71
(1.0
32)
0.2
15
(0.8
56)
–0.1
12
(0.5
10)
0.3
15
(0.2
91)
0.5
37
(1.4
59)
3.3
09
(1.4
99)
–0.1
42
(0.6
53)
0.1
37
(0.4
52)
0.2
38
(0.2
25)
Hum
ancr
eepin
g
(han
ds-
and-f
eet)
5120.9
32
(5.1
87)
–8.0
60
(3.5
10)
2.8
18
(1.4
44)
1.9
21
(1.7
84)
0.2
07
(0.6
05)
0.1
71
(0.5
32)
–3.8
80
(3.0
85)
0.5
92
(1.9
22)
–0.2
96
(0.7
10)
–1.0
08
(1.0
38)
–0.1
35
(0.2
04)
Hum
anw
alkin
g19
152.9
88
(3.6
36)
2.3
63
(1.5
40)
6.6
48
(1.5
86)
–0.2
19
(0.7
06)
0.2
26
(0.3
08)
0.0
74
(0.3
55)
10.2
32
(1.5
61)
–4.5
26
(1.8
47)
–1.7
21
(0.4
81)
–0.5
24
(0.3
90)
–0.4
52
(0.3
33)
Hum
anru
nnin
g14
136.6
88
(3.4
43)
5.5
25
(2.2
72)
3.4
33
(2.2
75)
0.0
39
(0.5
79)
–0.2
42
(0.3
30)
–0.2
20
(0.2
27)
7.2
05
(1.5
52)
–7.4
05
(1.5
72)
–1.0
32
(0.4
38)
–0.2
64
(0.3
36)
–0.3
08
(0.3
05)
aS
tan
dar
dd
evia
tio
ns
are
giv
enin
par
enth
eses
Primates (2007) 48:277–292 283
123

are therefore fairly evenly spaced in time, and the forelimb/
upper limb oscillates smoothly. Human creeping on hands
and feet, in addition to being out of phase with the other
modes, differs slightly from this pattern, in that the
shoulder is held at maximum retraction for a moment be-
fore the upper limb swings forward. Human creeping on
hands and knees is quite different from the other modes of
locomotion, since maximum protraction follows maximum
retraction by only about 30%. Hence, we see a steep slope
between those two points, followed by a gentler slope after
maximum protraction is reached. This is an indication of
the relatively short swing phase and long support phase of
the upper limb.
In Tables 1, 2, 3 and 4, there is a general decrease in
magnitude (independent of sign) as we move to higher order
coefficients. Table 5 shows where significant (P £ 0.01)
differences in shoulder angle exist among the various
modes of locomotion. Among the 11 coefficients, average
joint angle and the first and second order coefficients are
frequently different, but the fourth and fifth are usually not;
the third order components are intermediate. With respect to
the other joints, the smallest number of significant differ-
ences is found among elbow angle comparisons, while the
greatest number is found among knee comparisons.
For the shoulder, there are a large number of significant
differences, especially in the average shoulder angle (a0),
the third cosine coefficient, and the first and second sine
coefficients. The fourth and fifth order components, both
sine and cosine, show either few or no differences among
the various types of locomotion. Average shoulder angles
(a0) are significantly different among nearly all forms of
locomotion, except between non-human primate walking
and hands-and-knees creeping, a reflection of the similar
torso positions in these gaits.
Since the values of the fourth and fifth order components
are small (Table 1), and their differences are generally not
significant (Table 5), the Fourier coefficients were reana-
lyzed to determine each coefficient’s contribution to the
Fig. 5a–e Average curves for
shoulder angle versus percent of
stride cycle for the five types of
locomotion studied. Each graph
begins with right hind limb
strike (RHS) at 0%, and greater
angles indicate protraction.
a Primate quadrupedal walking.
b Human creeping on hands
and knees. c Human creeping
on hands and feet. d Human
bipedal walking. e Human
bipedal running
284 Primates (2007) 48:277–292
123

overall estimate of the shoulder data. Table 6 shows the
contribution of each of the first, second and third order
coefficients, expressed as a percentage of the total estimate
of the data, for each mode of locomotion. In this analysis,
only the first five sine and cosine coefficients were con-
sidered. From the last row of Table 6, it is clear that almost
all (approximately 96%) of the information in each FFT is
accounted for by the first three sine and cosine coefficients.
These first, second and third order coefficients are therefore
sufficient to describe all but the fine details of the data, and
the higher order coefficients (i.e., fourth and fifth) are of
relatively little importance to our estimate. Hands-and-feet
creeping is somewhat unusual in this set, in having the
lowest cos1 coefficient (by a factor of ten in most cases),
and the highest sin1 (by at least 20%).
Similar results were obtained in comparisons among
Fourier coefficients of the elbow, hip and knee. For all
joints, the magnitudes of first and second order coefficients
were generally the greatest, the fourth and fifth were
smallest, and the third order coefficients were intermediate.
Accordingly, significant differences appeared most often
among first, second and third sine and cosine components.
Comparisons among knee movements were notable, in that
they show relatively more differences involving hands-and-
knees creeping than other joints and modes of locomotion.
Hands-and-knees creeping is the only mode in which the
knee is placed on the substrate, nearly eliminating the leg,
ankle and foot contributions to locomotion and changing
the function of the knee joint itself. For this reason, it is not
surprising that knee coefficients for hands-and-knees
creeping differ greatly from the others.
In Tables 1, 2, 3 and 4, standard deviations of Fourier
coefficients follow a pattern similar to that seen among the
absolute values of the coefficients; namely, standard devia-
tions decrease as the order of the coefficient increases. Thus,
higher order coefficients have smaller standard deviations.
Discussion
Average joint angles
Differences in the average angles of the four joints (a0) can
be interpreted with reference to body proportions and trunk
positions in various types of locomotion (Tables 1–4;
Fig. 4). During normal walking, the limbs of quadrupedal
primates oscillate in the sagittal plane under a nearly hor-
izontal torso and both hip and shoulder angles will average
approximately 90�. Thus, the quadrupedal forms of loco-
motion show average shoulder angles of 70�–100� and hip
angles of 75–105� (Tables 1, 3). However, increased or-
thogrady will tend to decrease shoulder angle and increase
hip angle. The extreme case is seen among bipedal humans
(Fig. 4), with slightly negative shoulder angles and hip
angles near 170�. Human hands-and-feet creeping, wherein
the shoulder is lower than the hip, is unusual in that it
evinces shoulder angles higher than 90� and hip angles
lower than 90�.
The fully extended elbow or knee has a maximum angle
of approximately 180�, and the average joint angle must
therefore be less than 180� (Tables 2, 4). Hence, average
elbow angles are generally between 155� and 165�, the
only exception being found in human running when the
elbow is usually held in a flexed position. Average knee
angles are generally between 120� and 155�, except for
human hands-and-knees creeping, which is somewhat be-
low 90� since the knee is flexed 90� to allow it to be placed
on the substrate.
Joint movement and Fourier coefficients
A major focus of this study is the shoulder, since Fourier
analysis of shoulder movements can elucidate similarities
in interlimb coordination when footfall diagrams are
irrelevant, as they are when comparing bipeds with quad-
rupeds. As noted above, this is possible when the stride
cycle is defined by hind limb actions such as the time when
the right hind limb strikes the substrate (right hind limb
strike; RHS). If all the datasets begin with the same hind
limb action, similar forelimb joint movements will produce
similar Fourier analysis results.
From Table 6, we see that the greatest percent contri-
bution of any component to the overall estimate of the
shoulder data ranges from 35%, for both first sine and first
cosine in primate quadrupedal walking, to 79% for first
cosine in human walking. Furthermore, in combination, the
sums of the first sine and cosine components range from
64 to 88% of the overall estimate, with higher order
Table 5 Significant differences (P £ 0.01) among shoulder angle coefficients
Hands-and-feet Human walking Human running Primate walking
Hands-and-knees a0, cos1, cos3, sin1, sin2 a0, cos3, sin1, sin2, sin4 a0, cos3, cos5, sin1, sin2 cos3, sin1, sin2
Hands-and-feet a0, cos1, cos3, sin1, sin2 a0, cos1, cos3, sin2 a0, cos1, cos2, sin1, sin4
Human walking a0, cos2, sin1, sin2 a0, cos2, cos3, sin1, sin2, sin4
Human running a0, cos2, cos3, sin1
Primates (2007) 48:277–292 285
123

coefficients adding relatively little to the estimate. These
results highlight the fact that there is only one peak and one
trough in the graphed data of shoulder angle versus percent
stride (Fig. 3), corresponding to the relatively simple
oscillation of the forelimb, as the limb is protracted during
swing phase and retracted during stance phase. Further-
more, the functional differences between human upper
limbs and non-human forelimbs are seen in the combined
contributions of the first sine and cosine coefficients in
Table 6. Specifically, the two modes of human bipedalism
evince the highest contributions of first sine and cosine
components (87–88%), well above those of the other
modes (64–70%). This tells us that the upper limbs in
human bipedalism oscillate fairly smoothly, with a single
peak and trough evenly spaced in time. In contrast, the
somewhat lower combined first sine and cosine contribu-
tions of the quadrupedal modes indicate a skewed distri-
bution of the data, in which the peak and trough are not
evenly spaced. The functional significance of this dis-
crepancy lies in the fact that, in bipedalism, the upper limbs
are not weight-bearing and hence are free to swing much
like pendulums. However, the support phase of the fore-
limb in quadrupeds (including creeping humans) is longer
than the swing phase, since the torso must vault over the
supporting limb. The swing phase is then relatively short,
as the limb is swung forward quickly to prevent the torso
from pitching forward during progression. While compar-
isons among forelimb duty factors could have shown the
disparity between stance and swing phase durations, com-
parison of Fourier coefficients is necessary when bipeds are
included in the analysis, since there is no duty factor for
limbs that do not contact the ground.
Higher order coefficients (second sine, third cosine, etc.)
are better suited to describing data with two or three or
more peaks and troughs and, for shoulder and hip angle,
will generally be most useful only in refining the overall
shape of the estimate. Tables 2, 3 and 4 support this con-
clusion. Table 3 for the hip shows clearly that the most
prominent coefficients are first cosine and first sine com-
ponents, results that agree well with those of Grasso et al.
(2000),who used Fourier analysis to study the hip, knee and
ankle in human walking. Table 2 (elbow) and Table 4
(knee), however, show very strong contributions from the
second sine and cosine components, reflecting the fact that
the elbow and knee undergo two flexion and extension
phases in each stride cycle. The first flexion phase of the
elbow or knee occurs when the limb flexes as the body
vaults over it, while the second flexion phase helps the
cheiridium clear the ground during the forward swing.
The third, fourth and fifth sine and cosine components
are quite small in comparison to the first three, as indicated
in Tables 1–6. In Table 6, for the shoulder only, the first
three sine and cosine components together account for at
least 95% of the shape of the shoulder graph, leaving only
5% or less to be covered by the fourth and fifth order
components. Even the third sine and cosine components
account for relatively little (from less than 0.1% to about
8%). In Tables 1–4, clearly the largest coefficients are
those for the first and second components, not only for the
shoulder, but also for the other joints studied. This is be-
cause none of the limb segments in any mode of locomo-
tion oscillates back and forth four or five times and, again,
these higher order components merely refine the overall
patterns of joint movements. Also, since the fourth and fifth
coefficients account not for the general shapes of the curves
but for the details in the data, they are more likely to show
individual variation. Hence, in Table 5 the variance of
fourth and fifth order components for the shoulder is such
that there are often no significant differences among the
five modes of locomotion. This is evidence that there are
no hidden, higher frequency oscillations in the function
of the limbs. Therefore, the use of high-speed film (e.g.,
120 fps) will generally be unnecessary to understand the
functions of these joints in medium- and large-bodied
primates. Perhaps in studying small primates researchers
will need to use high-speed data collection techniques for
the major limb joints, since their movements will generally
be faster. However, Fourier analysis shows that, in most
cases, the locomotor movements of larger primates can be
studied with standard (30 fps) video.
Table 6 Percent contribution
of the first three sine and cosine
coefficients to the overall
estimate of the shoulder data
Coefficient Primate
walking
Hands-and-knees
creeping
Hands-and-feet
creeping
Human
walking
Human
running
cos1 35.098 53.033 4.450 78.807 49.052
cos2 11.636 1.164 10.917 2.360 5.998
cos3 4.052 5.416 8.365 0.069 0.179
sin1 35.434 11.934 59.296 8.246 39.264
sin2 7.704 23.500 10.430 5.876 2.392
sin3 2.560 1.603 2.150 1.498 0.907
Total % 96.484 96.650 95.608 96.865 97.792
286 Primates (2007) 48:277–292
123

Variance and variability
Standard deviations of the Fourier coefficients can tell us
about the experimental procedures used and the composi-
tions of the groups studied. Hence, different study groups
can show mardedly different standard deviations. From
Tables 1–4, it is clear that in human walking and running
there is somewhat less variation in higher order coefficients
than there is among the other types of locomotion. In
Tables 1 and 3, for the shoulder and hip, respectively, the
standard deviations of first order coefficients (cos1 and
sin1) in human walking and running are similar to those
found among other types of locomotion. However, second
order coefficients are generally much lower for both forms
of human bipedalism than for any of the other modes of
locomotion. This trend continues as we look at progres-
sively higher order coefficients. Figure 6 shows the rela-
tively low standard deviations of shoulder coefficients for
human walking and running, as compared to the other
modes of locomotion studied. Elbow, hip and knee stan-
dard deviations evince a similar pattern, such that the
second and higher order coefficients for human walking
and running are generally smaller than those for primate
walking and human quadrupedal movement.
Lower standard deviations for human bipedalism tell us
about the conditions of the experiment. Since both types of
human bipedal locomotion (walking and running) were
performed on a treadmill, speed and direction were held
nearly constant. Therefore, minor fluctuations in speed
and direction that might normally occur, due to slight
unevenness of the substrate or minor asymmetries in
muscle contractions, are reduced by the experimental
technique. The non-human primates walked on level, but
sometimes slightly irregular, substrates in zoos, adding
variability to their limb movements. Furthermore, although
humans creeping on hands and knees moved across a
smooth, padded floor, small variations in muscle use, from
one stride to the next or between the left and right sides of
the body, would also cause increased variation in joint
movements. The treadmill used by bipedal subjects con-
strains such movements. Indeed, similar results have been
observed in other vertebrates. For example, Webb (1993)
found that fish swimming in artificial flumes (the fish
equivalent of a treadmill) showed less variance in their
movements than when they swam in perfectly still ponds.
The steadily moving water, even with its turbulence, con-
strained the movements of the fish. Fourier analysis thus
demonstrates the effect of constraining (laboratory) con-
ditions on movement variability and serves as a caveat to
those comparing results from laboratory settings with re-
sults from more naturalistic ones.
One might reasonably ask then, why the hands-and-feet
creeping subjects did not show standard deviations as low as
those for bipedal subjects, since they too were on a treadmill.
The answer certainly lies in the fact that creeping on hands
and feet is an unusual mode of locomotion for an adult hu-
man. Indeed, the original study from which those data were
drawn was intended to investigate the effects of learning a
novel task (Sparrow and Irizarry-Lopez 1987). With con-
siderable practice, one would expect subjects creeping on
hands and feet to show less variation in their movements and,
therefore, less variance in their Fourier coefficients.
It is unlikely that the different film speed (15 vs 30 fps)
is a significant factor in the hands-and-feet creeping study,
because the Fourier transform is useful for a variety of
sampling rates. The limiting factor is the total number
of datapoints (frames in this case), expressed in powers of
two. Thus, two datapoints limit research to the first sine and
cosine components, four datapoints limit us to the first- and
second-order components, eight points limit us to third-
order or less, and so forth. Fifteen and 30 fps for a duration
of stride of a little more than 1 s produce more than 16
datapoints, limiting us to the fourth-order components
(16 = 24), or the fifth order when stride period is well over
1 s (‡32 frames). Most of the strides in this study provided
more than 32 frames of data but fewer than 64 frames, so
we do not report any Fourier coefficients beyond the fifth
order. Specifically, those taken from Sparrow and Irizarry-
Lopez’s (1987) hands-and-feet creeping data generally
covered about 40 frames. Also, the vast majority of the
information about joint movement is covered by the first
three components (as discussed above). Hence, Fourier
analysis of primate locomotion is fairly tolerant of differ-
ences in camera speed.
Fig. 6 Standard deviations of the first five coefficients (both cosine
and sine). The two thick lines represent human bipedal locomotion
(triangles indicate running, diamonds indicate walking). Open circlesrepresent hands-and-feet creeping; open squares represent hands-and-
knees creeping; crosses represent primate walking. Standard devia-
tions of second through fourth order coefficients are markedly lower
for human bipedalism than for the other modes of locomotion
Primates (2007) 48:277–292 287
123

Fourier coefficients and mode of locomotion
As indicated in Table 6, the first and second sine and co-
sine coefficients account for most of the descriptive data in
the FFTs of shoulder angle. Hence these components were
further analyzed in Fig. 7. The box plots in Fig. 7 also
show that the first cosine component (Fig. 7a) can clearly
distinguish between lateral couplets gaits (e.g., hands-and-
feet creeping) and diagonal couplets gaits, but the other
coefficients generally did not show such clear separations.
Hence, although various modes of locomotion can be dis-
tinguished on statistical grounds (Table 5), there are few
clear discontinuities and considerable overlap among the
other coefficients, when considered individually. Thus,
human walking, running, and hands-and-knees creeping
appear in many respects to lie on a continuum with primate
quadrupedalism, a continuum that shows behavioral and
neurological similarity among diagonal couplets gaits.
These modes are, however, quite distinct from human
hands-and-feet creeping, which more closely resembles the
quadrupedalism of some other mammals. An example of a
non-primate mammal with a lateral couplets gait is the
leopard (Panthera pardus), indicated by P in Fig. 7. For all
four coefficients (first and second sine and cosine), the
leopard is indistinguishable from the lateral-couplets,
hands-and-feet creeping of humans, most notably for the
first cosine coefficient, which clearly separates the two
types of interlimb coordination. Hence, despite the obvious
mechanical and morphological differences, human biped-
alism and creeping on hands and knees resemble primate
quadrupedalism more than they do the lateral couplets gaits
of other mammals and human hands-and-feet creeping.
Furthermore, the informal observation that human walking
seems to evince diagonal-couplets interlimb coordination is
validated in Fig. 7, without the need for support diagrams.
Bivariate plots and ‘‘Kinesiological Space’’
Although it is impossible to separate all modes of loco-
motion on the basis of a single Fourier coefficient, pairs
of coefficients can distinguish among them fairly clearly.
The major components of an FFT can be plotted against
one another, thereby potentially separating different groups
along two axes; Fig. 8 shows bivariate graphs of pairs of
coefficients. Where different modes of locomotion are
grouped separately on these graphs, a kind of ‘‘kinesio-
logical space’’ is defined. Thus, kinesiological spaces are
defined by consistent association of individuals, species or
types in particular areas of the graph. In this case, FFT
coefficients describe the movement characteristics of a
particular joint in a particular mode of locomotion.
In Fig. 8a, the separation of diagonal couplets from
lateral couplets gaits is made even more obvious than in
Fig. 7a by the combination of the first sine component and
the first cosine component. But Fig. 8a also shows a clear
separation of quadrupedal walking (open diamonds) from
bipedal running (crosses), whereas there is still much over-
lap among human walking, running and hands-and-knees
creeping. Second and third cosine coefficients, by their
nature, can help to refine these distinctions (Fig. 8b). In this
Fig. 7a–c Box plots showing the distribution of first and second
cosine and first sine coefficients for human hands-and-feet creeping
(white), hands-and-knees creeping (diagonal lines), bipedal walking
(vertical lines), primate walking (stipling), and bipedal running (solidblack). In each graph, Pantera pardus is indicated by the letter
P. a cos1, b cos2, c sin1
288 Primates (2007) 48:277–292
123

case, the kinesiological spaces occupied by creeping on the
hands and knees and by human bipedalism are more clearly
separated from one another, indicating that the details of
shoulder movement in these modes are where the differ-
ences lie. Shoulder movements in human walking and
running are very similar and so are not easily distinguished
with most Fourier coefficients. However, a graph of the
first and second sine components helps to separate them,
even though they appear to lie on a continuum of the same
type of motion (Fig. 8c). (Human walking and running are
quite clearly separated by the combination of second cosine
and first sine, and both are very different from quadrupedal
primates, but in the interests of space, not all possible
graphs are shown.) Figure 8c also quite clearly separates
hands-and-knees creeping from all other modes of loco-
motion.
Conclusions and prospectus
In summary, the results of Fourier analysis indicate con-
siderable similarity among primate quadrupedalism, human
bipedalism and human creeping on hands and knees.
Without the use of Fourier analysis, some of this similarity
would be unquantifiable, since footfall analysis cannot be
applied to bipedal humans as it can to quadrupeds. A
number of benefits accrue to the use of FFTs in the study of
primate locomotion. Some of these have been realized
here, while others will be the subject of further study.
Realized benefits of Fourier analysis
In particular, we have used Fourier analysis in the fol-
lowing ways:
1. FFTs are designed to describe cyclical phenomena,
including stride cycles, and are therefore well-suited
to descriptions of joint angles in locomotion research.
Also, they are specifically designed to describe data
in terms of their shape on curvilinear graphs—exactly
what is needed in joint movement studies.
2. FFTs allow us to compare joint movements of the
upper limbs in walking humans with those of the
forelimbs of quadrupedal primates, something that is
not possible with footfall analysis (ground contact
diagrams), since the upper limbs never contact the
ground. Thus, the casual observation, that walking
and running humans seem to use diagonal couplets
gaits like those of other primates, can be shown with
some degree of precision.
3. FFTs are an improvement over the analysis of footfall
patterns to describe interlimb coordination, since they
describe each joint’s contribution to limb movement,
rather than just the results of those joints’ movements
on ground contact. We have focused on the shoulder
joint for the most part—and the shoulder and hip in
quadrupeds make fairly simple movements whose
Fig. 8a–c Bivariate plots of pairs of Fourier coefficients. Solid circlesHands-and-feet creeping, open squares hands-and-knees creeping,
open triangles human walking, crosses human running, opendiamonds non-human primate walking, P Pantera pardus. Note that
hands-and-feet creeping and the leopard generally occupy the same
kinesiological space. a cos1 vs sin1, b cos2 vs cos3, c sin1 vs sin2
Primates (2007) 48:277–292 289
123

relationship to footfall patterns is relatively clear—
but other joints can as easily be studied. Indeed, the
knee and elbow, with two peaks and troughs in their
goniometric data, are not readily understood with
footfall diagrams. Their function is best elucidated by
goniological techniques like those described here.
4. Strides of different duration can be compared as
easily as those of identical duration, since it is the
overall shape of the (graphed) data, not the stride
period, which determines the Fourier coefficients. In
that sense, FFTs act similarly to data transformations
such as expression of the raw data in percent of stride
cycle—both techniques standardize the data with re-
spect to time. In the case of Fourier analysis, how-
ever, the data are removed from the ‘‘time domain’’
entirely, and placed in the ‘‘frequency domain.’’ This
means that sine and cosine waves of various fre-
quencies are added together to produce the same
distribution of data as if a measurement were plotted
against time.
5. FFTs can produce angular data graphs with many
fewer data points than traditional ‘‘time domain’’
graphs (i.e., angle vs time). In this study, datasets of
approximately 50 angles were very well-character-
ized using only 11 Fourier descriptors (five sines and
cosines, and the average angle). Indeed, as noted
above (and in Table 6), 95% of the information can
be described with only seven numbers (three sines
and cosines, and the average angle).
6. Data collected with cameras of different speeds will
usually be compatible, since the number of frames for
each stride is irrelevant, because Fourier analysis will
give very similar results if the same stride is de-
scribed with 24 data points or 48 data points. In this
study, we used frame rates of 15 and 30 fps and the
data were very well-described in both cases.
7. FFTs quantify angular data in a standardized form,
allowing us to use statistical analyses to compare
joint movements among different strides of the same
individual, among different individuals, and even
among different species. Here, we averaged single
strides of several different individuals to arrive at a
general pattern for a particular type of locomotion in
a particular species (or group of species).
8. Fourier coefficients seem to be normally distributed,
so simple statistical tests (like Student’s t-test) can be
used in comparisons. The comparisons in Table 5, for
example, were based on t-tests.
9. The standardization of data by FFT does not distort
the original angular data. This is an advantage over
the technique of dividing the stride cycle into stance
and support phases and plotting joint angle versus
percent of each phase.
10. FFTs provide a simple method of archiving data,
using many fewer datapoints than the original raw
data. It is our hope that other researchers will benefit
from our presentation of the Fourier coefficients in
Tables 1–4. By reversing the Fourier transform, as
described above, others will have joint angle data for
use in comparative studies.
11. The number of flexions/extensions shows up fairly
clearly among the Fourier coefficients, since simple
protraction/retraction movements will have high first
order coefficients, while more complex movements
with two flexions/extensions will have high second
order coefficients.
12. FFTs have allowed us to produce graphs of average
joint movements for many strides and many indi-
viduals.
13. Analysis of the statistical variation of the coefficients
tells us about the variance in movements among
different strides of the same individual and among
different individuals, different species and different
experimental conditions. Variance helps elucidate
mechanical differences in locomotion on a treadmill
versus that on the ground where minute accommo-
dations to irregular substrates are expected.
14. Comparison of coefficients helps us understand the
differing functions of the various limb segments and
joints (in shock absorption, directional control, etc.).
For example, Table 4, for the knee joint, shows the
very large contribution of the first cosine component
to the description of hands-and-knees creeping, as
distinct from all other modes studied.
15. Since FFTs generate average angle, we can easily
investigate differences in average angle due to dif-
ferences in limb proportions and posture.
16. Fourier analysis describes the distribution of the
dataset as a whole, rather than focusing on a few
points in the stride cycle (e.g., a limb striking the
substrate or the timing of maximum extension of a
joint). Therefore, differences between these points are
also quantified for further study. The data for shoulder
angle highlight the smooth, symmetrical oscillation of
human upper limbs during bipedal locomotion, as
opposed to the uneven motion evinced when the upper
limb/forelimb is used for support or propulsion.
Future research
Not all of the benefits of Fourier analysis have been real-
ized in this project. Additional data collection would be
necessary to prove the value of FFTs for other types of
analyses and to answer other questions about primate
locomotion. Future research will undoubtedly begin
with an expansion of the small database provided here.
290 Primates (2007) 48:277–292
123

Thereafter, it may include the study of the effects of sub-
strates on joint movement, and variance may be analyzed
with respect to the level of expertise or experience that
subjects have with a particular mode of locomotion. Some
of these effects may be found most easily using Fourier
analysis. Therefore, several as yet unrealized benefits
should come from the use of Fourier analysis:
1. Because of the nature of Fourier transforms, large
datasets of angular measurement against time can be
reduced to very few datapoints. In theory, we can de-
scribe the shape of a curve which is based on 2n da-
tapoints, using n sines and n cosines. Thus, very high-
speed (120 fps) videorecordings of primate movements
can be fully described with approximately seven sines
and cosines and the average angle. [This is based on a
stride of a little more than 1 s, producing about 128
(=27) angular measurements.] Furthermore, even with
very high frame rates, the vast majority of the distri-
bution of the data can be described with only three
sines and cosines and the average angle.
2. FFTs provide a simple method of archiving data, using
many fewer datapoints than the original raw data. In
the future, even high-speed video of primate locomo-
tion can be reduced to only a few numbers and a large
database can be developed. The database can be used
in biomechanical research, even when the original
primates themselves are unavailable. Unfortunately, it
is quite possible that the greatest value of such a
database will be in archiving the movements of extinct
primates in the decades to come.
3. Since FFTs generate average angle, we can easily
investigate changes in average angle that may arise
from changes in speed or inclination of the substrate,
and we can investigate differences in average angle
due to differences in limb proportions and posture.
Average angles can be produced without Fourier
analysis but are a by-product of FFTs so no additional
work must be performed to obtain that information
while also deriving the other benefits of Fourier anal-
ysis. In addition, differently angled branches or terrain,
or more irregular substrates, may cause kinesiological
changes that do not show up in EMG studies, since
similar muscle contractions may result in different
movements under different conditions. These changes
would be found most easily with Fourier analysis.
4. Study of variance should help us understand the
learning of a new task or the difference between an
expert and a novice. Here, we noted differences in the
variance of Fourier coefficients for a novel task
(hands-and-feet creeping) and walking and running.
Further research will reveal the rate of improvement in
the performance of new locomotor tasks.
5. Analysis of variance will likely have clinical uses. The
degree of deviance from average human FFTs and
therefore average curves of joint angle will help define
rates of post-operative and post-traumatic recovery.
6. Studies of variance can be used to investigate learning
of all types of locomotor behaviors in all primates. For
this reason, analysis of variation in Fourier coefficients
should help us understand the acquisition and refine-
ment of locomotor patterns in young primates.
7. Biomechanical modelling will benefit from Fourier
analysis, since average curves of joint movements can
be used to constrain a model’s output to fit the
parameters of an FFT. Better still, several FFTs of
several joint angles can be combined with body seg-
ment lengths to produce stick figures of primates in
motion. The paths of body points in space (e.g., center
of mass, base of spine, back of head) can also be
described with Fourier analysis, providing additional
constraints on biomechanical models.
8. Biomechanical models based on FFTs can benefit the
entertainment industry in the area of animated film and
special effects. Thus, the representations of primates,
including humans, in motion pictures can be made
more realistic.
Acknowledgments The authors would like to thank Andy Baker,
Philadelphia Zoo, for organizational assistance and permission to
videotape several primates; Stephanie Silfies Donner and Sheela
Davis of the Department of Anthropology and Sociology, Kutztown
University, for their assistance in data entry; Dave Abel and Barry
Peterson, for video equipment loan and technical assistance; and
Dr. Robert Root, Department of Mathematics, Lafayette College, for
mathematical advice. The authors also wish to thank the Kutztown
University Research Committee for their financial support.
References
Bell ET (1937) Men of mathematics. Simon and Schuster, New York
Burnside LH (1927) Coordination in the locomotion of infants. Genet
Psychol Monogr 2:280–372
Cappozzo A (1981) Analysis of the linear displacement of the head
and trunk during walking at different speeds. J Biomech 14:411–
425
Cartmill M, Lemelin P, Schmitt D (2002) A new theory concerning
the adaptive value and evolution of diagonal-sequence gaits in
primates and marsupials. Am J Phys Anthropol 34(Suppl):52
D’Aout K, Aerts P, De Clercq D, De Meester K, Van Elsacker L
(2002) Segment and joint angles of hind limb during bipedal and
quadrupedal walking of the Bonobo (Pan paniscus). Am J Phys
Anthropol 119:37–51
Dunbar DC (1989) Locomotor behavior of Rhesus Macaques
(Macaca mulatta) on Cayo Santiago. P R Health Sci J 8:79–85
Dunbar DC, Badam GL (2000) Locomotion and posture duringter-
minal branch feeding. Int J Primatol 21:649–669
Fleagle JG, Stern JT, Jungers WL, Susman RL, Vangor AK, Wells JP
(1981) Climbing: a biomechanical link with brachiation and with
bipedalism. In: Day MH (ed). Vertebrate locomotion, Symposia
Primates (2007) 48:277–292 291
123

of the Zoological Society of London, No. 48. Academic Press,
New York, pp 359–375
Grand TI (1976) Differences in terrestrial velocity in Macaca and
Presbytis. Am J Phys Anthropol 45:101–108
Grasso R, Zago M, Lacquaniti F (2000) Interactions between posture
and locomotion: motor patterns in humans walking with bent
posture versus erect posture. J Neurophysiol 83:288–300
Hildebrand M (1967) Symmetrical gaits of primates. Am J Phys
Anthropol 26:119–130
Hirasaki E, Kumakura H, Matano S (1993) Kinesiological character-
istics of vertical climbing in Ateles geoffroyi and Macacafuscata. Folia Primatol 61:148–156
Hirasaki E, Kumakura H, Matano S (2000) Biomechanical analysis of
vertical climbing in the spider monkey and the Japanese
macaque. Am J Phys Anthropol 113:455–472
Isler K (2005) 3D-Kinematics of vertical climbing in hominoids. Am
J Phys Anthropol 126:66–81
Jungers WL, Anapol FC (1985) Interlimb coordination and gait in the
brown Lemur (Lemur fulvus) and the talapoin monkey (Miop-ithecus talapoin). Am J Phys Anthropol 67:89–97
Kimura T, Yaguramaki N, Fujita M, Ogiue-Ikeda M, Nishizawa S,
Ueda Y (2005) Development of energy and time parameters in
the walking of healthy human infants. Gait Posture 22:225–232
Lasko-Macarthey P, Beuter A, Biden E (1990) Kinematic variability
and relationships characterizing the development of walking.
Dev Psychobiol 23:809–837
Lemelin P, Schmitt D, Cartmill M (2003) Footfall patterns and
interlimb co-ordination in opossums (family Didelphidae):
evidence for the evolution of diagonal-sequence walking gaits
in primates. J Zool (Lond) 260:423–429
Lestrel PE, Kimbel WH, Prior FW, Fleischmann ML (1977) Size and
shape of the hominoid distal femur: Fourier analysis. Am J Phys
Anthropol 46:281–290
Lestrel PE, Bodt A, Swindler DR (1993) Longitudinal study of cranial
base shape changes in Macaca nemestrina. Am J Phys Anthropol
91:117–129
Lestrel PE, Cesar RM Jr, Takahashi O, Kanazawa E (2005) Sexual
dimorphism in the Japanese cranial base: a Fourier-wavelet
representation. Am J Phys Anthropol 128:608–622
Mittermeier RA (1978) Locomotion and posture in Ateles. Folia
Primatol 30:161–193
Mittermeier RA, Fleagle JG (1976) The locomotor and postural
repertoires of Ateles geoffroyi and Colobus guereza, and a
reevaluation of the locomotor category semibrachiation. Am J
Phys Anthropol 45:235–256
Prost J (1965) The methodology of gait analysis and gaits of
monkeys. Am J Phys Anthropol 23:215–240
Prost J (1969) A replication study on monkey gaits. Am J Phys
Anthropol 30:203–208
Richmond AE (1972) Calculus for electronics. McGraw-Hill, New
York
Rosenberger AL, Stafford BJ (1994) Locomotion in captive Leon-topithecus and Callimico: a multimedia study. Am J Phys
Anthropol 94:379–394
Schmitt D (2003) Evolutionary implications of the unusual walking
mechanics of the common marmoset (C. jacchus). Am J Phys
Anthropol 122:28–37
Shapiro LJ, Raichlen D (2006) Limb proportions and the ontogeny of
quadrupedal walking in infant baboons (Papio cynocephalus).
J Zool 269:191–203
Shapiro LJ, Anapol FC, Jungers WL (1997) Interlimb coordination,
gait, and neural control of quadrupedalism in chimpanzees. Am J
Phys Anthropol 102:177–186
Sparrow WA (1989) Creeping patterns of human adults and infants.
Am J Phys Anthropol 78:387–401
Sparrow WA, Irizarry-Lopez VM (1987) Mechanical efficiency and
metabolic cost as measures of learning a novel gross motor task.
J Mot Behav 19:240–264
Uetake T (1992) Can we really walk straight? Am J Phys Anthropol
89:19–28
Vilensky JA, Larson SG (1989) Primate locomotion:utilization and
control of symmetrical gaits. Annu Rev Anthropol 18:17–35
Vilensky JA, Patrick MC (1985) Gait characteristics of two squirrel
monkeys, with emphasis on relationships with speed and neural
control. Am J Phys Anthropol 68:429–444
Webb D (1989) The function of the upper limbs in human walking.
PhD dissertation. University of Chicago, Chicago
Webb PW (1993) The effect of solid and porous channel walls on
steady swimmining of steel-head trout Oncorhynchus mykiss.
J Exp Biol 178:97–108
292 Primates (2007) 48:277–292
123