
Derived KMP-11 Antigen via Activation of...
13
Research Article Immunomodulation in Human Dendritic Cells Leads to Induction of Interferon-Gamma Production by Leishmania donovani Derived KMP-11 Antigen via Activation of NF-B in Indian Kala-Azar Patients Rajesh Chaudhary, 1 Ajay Amit, 1 Anupam Yadav, 1 Anurag Singh, 1 Vikash Kumar, 1 S. K. Singh, 1 Shyam Narayan, 1 Vidyanand Rabidas, 1 K. Pandey, 1 Anil Kumar, 1 Pradeep Das, 1 and Sanjiva Bimal 1,2 1 Rajendra Memorial Research Institute of Medical Sciences, Agamkuan, Patna, Bihar 800007, India 2 Division of Immunology, Rajendra Memorial Research Institute of Medical Sciences, Agamkuan, Patna, Bihar 800007, India Correspondence should be addressed to Sanjiva Bimal; [email protected] Received 16 September 2013; Revised 12 December 2013; Accepted 17 December 2013; Published 22 January 2014 Academic Editor: Mehdi Chenik Copyright © 2014 Rajesh Chaudhary et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Dendritic cells (DCs) and macrophages (MΦs) are well-known antigen presenting cells with an ability to produce IL-12 which indicates that they have potential of directing acquired immunity toward a 1-biased response. e aim of this study was to examine the effect of Leishmania specific KMP-11 antigen through comparison of immune responses aſter presentation by DCs and MΦs to T cells in Indian patients with VL. Patients with DCS and MΦs were directed against a purified Leishmania donovani antigen (KMP-11) and phytohaemagglutinin (PHA). e cytokines (IL-12, IL-10, and TGF-) producing abilities of the DCs and MΦs against these antigens were determined by flow cytometry. e transcription factor (NF-B) and T-cell cytokine support (IFN-, IL-10), which could be significant in effector immune function, were also determined. Severe hindrance in the immune protection due to Leishmania parasites, as revealed by decreased expression of IL-12 and upregulation of IL-10 and TGF- expression in the MΦs compared to DCs, occurred in VL patients. e production of IL-12 in response to L. donovani KMP-11 antigen was increased in DCs which was reduced in MΦs of VL patients. In contrast, the presentation of KMP-11 antigen by DCs to T-lymphocytes in VL patients significantly increased the IFN- produced by these immune cells, whereas the levels of IL-10 were significantly elevated aſter presentation of KMP-11antigen by MΦs. e VL patients were observed with severely dysfunctional MΦs in terms of NF-B activity that could be recovered only aſter stimulation of DCs with L. donovani KMP-11 antigen. Immunologically the better competitiveness of KMP-11 antigen through a dendritic cell delivery system may be used to revert T-cell anergy, and control strategy can be designed accordingly against kala-azar. 1. Introduction Among the various forms of leishmaniasis, visceral leishma- niasis (VL) caused by Leishmania donovani (L. donovani) in the Indian subcontinent is the most severe and fatal if untreated. InVL, the causative agent, is transmitted by the phlebotomine sandfly to mammalian hosts, where they reside primarily within macrophages (MΦs) and dendritic cells (DCs). MΦs are well known as effectors of the innate immune system and are able to direct acquired immunity toward a 1/2 biased response by their abilities to produce IL-12 and IL-10. DCs are the most potent type of the antigen- presenting cells (APC) and play a critical role in the initiation of immunity by producing soluble factors such as chemokines and cytokines [1]. InVL MΦs ingest promastigotes but these cells seem to synergies with parasite to facilitate infection [2–4]. Different studies have shown that, in addition to MΦs, infection of Leishmania parasite in DC has also been Hindawi Publishing Corporation BioMed Research International Volume 2014, Article ID 947606, 12 pages http://dx.doi.org/10.1155/2014/947606
Transcript of Derived KMP-11 Antigen via Activation of...

Research ArticleImmunomodulation in Human Dendritic CellsLeads to Induction of Interferon-Gamma Production byLeishmania donovani Derived KMP-11 Antigen viaActivation of NF-𝜅B in Indian Kala-Azar Patients
Rajesh Chaudhary,1 Ajay Amit,1 Anupam Yadav,1 Anurag Singh,1
Vikash Kumar,1 S. K. Singh,1 Shyam Narayan,1 Vidyanand Rabidas,1
K. Pandey,1 Anil Kumar,1 Pradeep Das,1 and Sanjiva Bimal1,2
1 Rajendra Memorial Research Institute of Medical Sciences, Agamkuan, Patna, Bihar 800007, India2Division of Immunology, Rajendra Memorial Research Institute of Medical Sciences, Agamkuan, Patna, Bihar 800007, India
Correspondence should be addressed to Sanjiva Bimal; [email protected]
Received 16 September 2013; Revised 12 December 2013; Accepted 17 December 2013; Published 22 January 2014
Academic Editor: Mehdi Chenik
Copyright © 2014 Rajesh Chaudhary et al. This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
Dendritic cells (DCs) and macrophages (MΦs) are well-known antigen presenting cells with an ability to produce IL-12 whichindicates that they have potential of directing acquired immunity toward aTh1-biased response.The aimof this studywas to examinethe effect of Leishmania specific KMP-11 antigen through comparison of immune responses after presentation by DCs and MΦsto T cells in Indian patients with VL. Patients with DCS and MΦs were directed against a purified Leishmania donovani antigen(KMP-11) and phytohaemagglutinin (PHA). The cytokines (IL-12, IL-10, and TGF-𝛽) producing abilities of the DCs and MΦsagainst these antigens were determined by flow cytometry. The transcription factor (NF-𝜅B) and T-cell cytokine support (IFN-𝛾,IL-10), which could be significant in effector immune function, were also determined. Severe hindrance in the immune protectiondue to Leishmania parasites, as revealed by decreased expression of IL-12 and upregulation of IL-10 and TGF-𝛽 expression in theMΦs compared to DCs, occurred in VL patients.The production of IL-12 in response to L. donovaniKMP-11 antigen was increasedin DCs which was reduced in MΦs of VL patients. In contrast, the presentation of KMP-11 antigen by DCs to T-lymphocytesin VL patients significantly increased the IFN-𝛾 produced by these immune cells, whereas the levels of IL-10 were significantlyelevated after presentation of KMP-11antigen by MΦs. The VL patients were observed with severely dysfunctional MΦs in termsof NF-𝜅B activity that could be recovered only after stimulation of DCs with L. donovani KMP-11 antigen. Immunologically thebetter competitiveness of KMP-11 antigen through a dendritic cell delivery system may be used to revert T-cell anergy, and controlstrategy can be designed accordingly against kala-azar.
1. Introduction
Among the various forms of leishmaniasis, visceral leishma-niasis (VL) caused by Leishmania donovani (L. donovani)in the Indian subcontinent is the most severe and fatal ifuntreated. InVL, the causative agent, is transmitted by thephlebotomine sandfly tomammalian hosts, where they resideprimarily within macrophages (MΦs) and dendritic cells(DCs).MΦs are well known as effectors of the innate immune
system and are able to direct acquired immunity toward aTh1/Th2 biased response by their abilities to produce IL-12and IL-10. DCs are the most potent type of the antigen-presenting cells (APC) and play a critical role in the initiationof immunity by producing soluble factors such as chemokinesand cytokines [1]. InVL MΦs ingest promastigotes but thesecells seem to synergies with parasite to facilitate infection[2–4]. Different studies have shown that, in addition toMΦs, infection of Leishmania parasite in DC has also been
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014, Article ID 947606, 12 pageshttp://dx.doi.org/10.1155/2014/947606

2 BioMed Research International
reported earlier [5, 6]. These cells as antigen-presenting cells(APCs) are able to induce a primary immune responseand establishment of immunological memory [7]. In vivoinfectivity of DC by L. donovani was also shown in mousemodels [8]. Furthermore, different studies have shown thatDC and MΦ are thought to distinguish different pathogensthrough the recognition of pathogen-associated molecularpatterns (PAMP) via the expression of pattern recognitionreceptors (PRR) such as the Toll-like receptor family thatdecode the microbial surface proteins [9–11]. The outermembrane of Leishmania is covered by a dense glycocalyxconsisting predominantly of lipophosphoglycan (LPG) [12].Kinetoplastid Membrane Protein 11 (KMP-11) is a majorsurface protein which remains noncovalently associated withLPG complex especially in L. donovani species [13]. In vitroand in vivo studies using purified parasite protein haveidentified KMP-11 as a multifunctional immunogenic factorrequired for the protection from infection and there iswidespread T-cell epitope conservation of this protein as well[14]. The roles ascribed to KMP-11 include reversal of T-cell anergy and upregulation of the expression of induciblenitric oxide synthase and synthesis of interleukin-12 (IL-12)in animal model [15, 16]. However, although such reports areencouraging, the efficacy may be variable between animalsand human patients. Macrophages are preferred host forLeishmania, but these cells seem to synergies with parasiteto facilitate infection [2–4] and because of this, there is astrong possibility for diminution of such effects of KMP-11in infected MΦs in human VL cases. But its involvementin antigen presentation in the clinical scenario, especiallyin case of VL however, remains to be investigated. In thepresent study, we investigated whether monocyte-deriveddendritic cells (moDCs) have different potential than MΦsto generate cytokines and drive CD4Th1 cytokine expressionafter stimulation with a purified L. donovani protein (KMP-11) in the Indian VL patients.
2. Material and Methods
2.1. Expression, Isolation, and Purification of rKMP-11 Pro-tein. To obtain the recombinant protein preparation fromLeishmania, the plasmid (pQE-30) containing the 273 bpL. donovani specific KMP-11 gene which was 3.4 kb wascultured in Luria Broth suspension medium containingampicillin (25 𝜇g/mL), kanamycin (100 𝜇g/mL) overnight at37∘C under 200 RPM in shaker incubator for 4-5 h. Therecombinant protein was subsequently induced by Isopropyl𝛽-D-1-thiogalactopyranoside (IPTG) (1mM/mL) overnightin shaker incubator at 22∘C and 200 RPM.The bacterial cellswere harvested, pelleted (5000 RPM), and lysed in cell lysingsolution (150mM NaCl, 10mM Tris HCl, 2% SDS; pH 8.0)in 1 : 4 ratio for 30min at 4∘C, and further ultrasonicated at85% amplitude and 0.5-second pulses for 5min. Followingcentrifugation (14000 RPM at 4∘C), recombinant proteinwas purified on a Ni+2-NTA superflow column (Qiagen)according to the manufacturer’s instructions. The protein inits denatured form was eluted in 8M urea salt buffer with250mM imidazole and dialysed in PBS at pH 7.4.The protein
expression was analysed by immunoblotting using mouseanti-KMP-11 Mab, clone L157 (GenWay, USA). Bacteriallipopolysaccharide (LPS) contamination in the recombinantprotein (KMP-11) preparation was determined using theLimulus amoebocyte lysate (LAL) test (Thermo-Scientific,USA). A LPS contamination (0.11𝜇g/mg of protein) wasdetected in the KMP-11 preparation. LPS in recombinantprotein KMP-11 was completely removed by passing throughpolymyxin B-agarose column (Sigma, USA) according to themanufacturer’s instructions and confirmed by LAL test. Therecombinant KMP-11 protein was then stored at −80∘C forfurther use [17].
2.2. Samples from VL Patients and Control. Peripheral-bloodmononuclear cells (PBMCs) were obtained from buffy coatsfrom age and gendermatchedVL patients and healthy donorsusing Ficoll-Hypaque (Pharmacia Biotech, Uppsala, Sweden)density gradient centrifugation at 800×g, 15min at 20∘C [18].APCs (MΦs and moDCs) were taken from these healthyand L. donovani infected individuals and were stimulatedwith recombinant protein (KMP-11) prepared to look atthe immunological response of the cells. These APCs werederived from 10 human patients with acute kala-azar (KA) intheir pretreatment stage and 10 normal controls. Blood sam-ples from KA patients were collected from the Departmentof the Clinical Medicine Unit, Rajendra Memorial ResearchInstitute of Medical Sciences, Patna, Bihar, India. These KApatients were all male and of mixed age (ranging from 15to 45 years) and were rK-39 and direct agglutination testpositive but human immunodeficiency virus (HIV) negative.The parasite burden in splenic aspirates was determined asdescribed previously by Chulay and Bryceson [19]. Each ofthe control subjects had no apparent history of VL and theydid not reside in the VL endemic areas.
2.3. Generation of Macrophage (MΦs) and Monocyte-DerivedDendritic Cells (moDCs). Heparinised samples of venousblood from patients and controls were used to procure APCsand PBMCs were isolated from Buffy coats using Ficoll-Hypaque density gradient centrifugation. Monocytes fromPBMCs of KA patients were isolated by adherence of thecells in a plastic petri-dish and its purity was determinedby FACSCalibur (Becton-Dickinson FACS-Calibur equippedwithCellQuestPro Software,USA) ofCD14, CD19, CD45, andCD3 expression [20], whichwere further cultured for another72 h.Cell preparationswere>92%MΦs and<0.5%T lympho-cytes as assessed by fluorescence (anti-CD14 and anti-CD19monoclonal antibodies, BD, USA) and size (FACS Calibur,BD, USA) [21]. The unfixed cells were stained with trypanblue which demonstrated that >95% cells were viable duringincubation. DCs were obtained from monocytes after treat-ment with IL-4 (1000U/mL) and granulocyte-macrophagecolony stimulating factor (GM-CSF) (800U/mL) on days 0,3, and 6 in complete RPMI medium for 7 days. ImmatureDCs harvested on day 7 were further cultured for 48 h todifferentiate in the presence of recombinant-Human TNF-𝛼 purified (20 ng/mL; Clone: MAb1, Isotype: Mouse IgG1,𝜅; eBioscience, Inc.). Viability of these DCs were found to

BioMed Research International 3
be >90% during incubation. The phenotypes of DC weredetermined in FACS by expression of CD1a, CD14, HLADR, CD80, CD86, and CD83 expression. Antibody stainedcells were evaluated using four-color FACS Calibur (BecktonDickinson, USA) [22, 23]. The absence of endotoxins in DCsand MΦs was demonstrated by LAL test as discussed in thepreceding section.
2.4. Intracellular Cytokine Produced from APCs. We initiallyexamined the capacity of moDCs and MΦs to generatecytokines (IL-12, IL-10, and TGF-𝛽) after stimulation withpurified KMP-11 protein. In brief, to give an initial anti-gen pulsing, cells (APCs) were stimulated with fixed (2%formaldehyde) Ld promastigote antigen in responder (APCs)to stimulator (fixed Ld) ratio of 1 : 50 for 2 h at 37∘C [24] andof them 1 × 106 cells were cultured in 96-well round bottomedplates in the presence of purified KMP-11 protein (10𝜇g/mL).Control cultures were set up in medium alone or mediumcontaining phytohaemagglutinin (PHA) 10 𝜇g/mL. Intracy-toplasmic cytokine level was detected on FACS-Calibur aspreviously described. Cells were cultured for 14 h followed by4 h incubation with a protein transport inhibitor, brefeldin-A (1 𝜇g/mL). The harvested cells were consecutively coincu-bated with PE-conjugated anti-CD83 antibodies/anti-CD14-PE antibodies, cytofix/cytoperm solution, and FITC conju-gated IL-10/IL-12/TGF-𝛽 (BDPharmingen, USA) before eachsample was resuspended in 500 𝜇L stain buffer. Samples wereanalysed on FACS-Calibur.
2.5. Cytokine Enzyme-Linked Immunosorbent Assay. To exa-mine the relationship between the cytokine production pro-files by the FACS based intracellular cytokine assay, afterpolyclonal stimulation with KMP-11, SLA, and PHA stimu-lation, we cultured representative DCs-T-cells and MΦs-T-cells cultured samples (1 × 106 cells/mL) from patients andcontrols for 72 h. Cytokine levels (IL-12, IFN-𝛾, and IL-10)were measured in culture supernatants by sandwich ELISAkit (BD Pharmingen, San Diego, CA, USA).
2.6. Effect of KMP-11 Antigen on NF-𝜅B Production in moDCsand MΦs. moDCs or MΦs (1 × 106) were suspended in 1mLof RPMI-1640 medium and pulsed with formalin fixed L.donovani promastigotes in 24-well tissue culture plates for14 h. The cells were then stimulated with KMP-11 antigenfor 16 h at 37∘C. Nucleic preparation was then obtained byincubating the washed (400 g, 5min at room temperature)cells with 250 𝜇L of a solution containing trypsin, a sperminetetrahydrochloride detergent buffer (BD Pharmingen, SanDiego, CA, USA) for 10min at RT to digest cell membraneand to stabilize the nuclear chromosome. The cells werelater treated with 200𝜇L of a solution containing trypsininhibitor and RNase buffer (BD Pharmingen, San Diego,CA, USA). The nuclei were incubated with PE-labelled anti-nuclear factor-𝜅B (anti-NF-𝜅B p65) antibody (Santa CruzBiotechnology, Inc.). Washed cells were resuspended in500𝜇L of stain buffer and production of NF-𝜅B in moDCsand MΦs of patients and control samples was determined byFACS-Calibur.
In parallel experiments, 20–30 𝜇g of nuclear extractof moDCs and MΦs, either stimulated with KMP-11 or
unstimulated, was separated by 12.5% polyacrylamide gelelectrophoresis (PAGE) with sodiumdodecyl sulphate (SDS).Separated proteins in the gel were transferred onto 0.22𝜇mnitrocellulose membranes (Sigma Aldrich, USA) by an elec-troblot apparatus (Bio-Rad). The blot paper was then leftfor 2 h at RT before being washed thrice with PBS-T (PBScontaining 0.1% Tween-20) and then soaked in 2% (IgG-free)BSA for 2 h at 37∘C, to block any nonspecific binding sites.The blot paper was washed once again with PBS-T beforebeing incubated for 2 h at 37∘C with a 1 : 1000 dilution of theNF-𝜅B (Santa Cruz Biotechnology, Inc.). UnboundMAb wasthen removed by three washes with PBS-T before any boundantibodywas labelled for 2 h at 37∘Cwith avidin-horseradish-peroxidase conjugate used in 1 : 5000 (BD Biosciences, USA).After 5 washes, color was developed with diaminobenzidine(0.06% in 0.5M citrate-saline, with 0.1% H
2O2) for 15min in
the dark.
2.7. Cytokine Released from CD4+ T-Lymphocytes. Subse-quent experiments were performed to look at the capacityof these two different APC subsets to drive CD4 T-cellcytokine response after stimulation with purified L. donovaniprotein (KMP-11) in VL patients and controls. In brief, 5 ×106 autologous PBMCs were cocultured with stimulated ornonstimulated moDCs and MΦs from patients and controls.For this, the PBMCs were added to the wells with alreadyprimed MΦs or moDCs and cultured for 6 days witheither medium alone or antigens of WHO reference strainLeishmania (20 : 1). The levels of expression of IFN-𝛾 andIL-10 in CD4+ T cells of the patients were determined byusing anti-CD4 antibodies and FITC labelled IFN-𝛾/IL-10antibodies by FACS-Calibur. Remaining steps were the sameas described in the preceding sections.
2.8. Statistical Analysis. All data were expressed asmean± SE(standard error of the mean). One way analysis of variancewith Tukey post hoc test was carried out using GraphPadPrism5, USA, software. A value of significance 𝑃 < 0.05 wasconsidered statistically significant.
3. Results
3.1. Characterization of KMP-11. Results on SDS polyacry-lamide gel electrophoresis of KMP-11 purified recombi-nant protein and the localization of KMP-11 protein afterimmunoblotting with anti-KMP-11 antibody are shown inFigure 1. The extraction of KMP-11 protein was achieved byNi2+-NTA immobilized column chromatography.The boundprotein was eluted in the presence of imidazole and latersubjected to 12.5% SDS-PAGE (Figure 1(a)). The anti-KMP-11 mAbs were later used in immunoblots on SDS-PAGEseparated KMP-11 proteins from Leishmania spp. which werepreviously purified by Ni2+-NTA affinity column. The mAbsdetected bands of approximately 11 KDa (Figure 1(b)).
3.2. Phenotypic Characterization of DCs. Human PBMCswere cultured in complete RPMI-1640 medium containingGM-CSF and IL-4 for 7 days followed by 2-day matura-tion in the presence of recombinant-human-TNF-𝛼. Phe-notypic characterization of mature DCs was demonstrated

4 BioMed Research International
WBM CP E1 E2 E3 F1 F2 F3
KMP-11(11kDa)
97.4
66.2
45.0
31.0
21.5
14.4
∗
∗
(a) (b)
Figure 1: (a) SDS polyacrylamide gel electrophoresis of L. donovani of KMP-11 purified recombinant protein and (b) western blot analysis.(a) SDS polyacrylamide gel electrophoresis of L. donovani of KMP-11 purified recombinant protein. Coomassie brilliant blue staining Lane 1Marker, Lane 2 CP- Crude Protein, Lane 3 E1 (eluted protein after first washing), Lane 4 E2 (eluted protein after the second washing), Lane5 E3 (eluted protein after the third washing), Lane 6 F1, Lane 7 F2, Lane 8 F3 (Lane 6-7 protein fraction of KMP-11), and F3 fraction in Lane8 show the KMP-11 recombinant protein. (b) KMP-11 recombinant protein purified by affinity chromatography, transferred on nitrocellulosepaper, and probed with anti-KMP-11 antibody. The arrow indicates the localization of KMP-11 protein. The asterisks show the dimeric andtrimeric forms of the KMP-11 protein.
for expression of costimulatory molecules (CD80, CD83,and CD86) in VL cases. Figure 2(a) shows that large cellswere gated and Figure 2(b) (B1-B2) showing differentiallyexpressed molecule CD83 (B1) and HLA-DR (B2). Theexpression was done on at least 3 separate patients. Thedifferentially expressedmolecule is CD1a positive. Figure 2(c)shows mature DCs showing a lesser percentage of CD1apositivity in CD14 and CD1a double stained cells. Figure 2(d)(D1-D2) shows inverted microscopic picture of immatureand mature DCs. Figure 2(e) shows mature MΦs showing agreater percentage of CD14+ cells than CD19+ cells after 72 hculture.
3.3. Cytokine Production by KMP-11 Stimulated MΦs andmoDCs. To understand APCs related differences, cytokineimmune cell response was examined in moDCs thatexpressed CD83, CD86, CD80, and HLA-DR and MΦs thatexpressed CD14 in response to purified L. donovani antigen(KMP-11) with flow cytometry (Figures 3 and 4). The initialparameters examined were IL-12, which is required for hostprotection, and IL-10 and TGF-𝛽, known to induce strongimmunosuppression following L. donovani infection.
The results revealed higher percentage of IL-10 and TGF-𝛽 in patients with VL compared with the healthy controls(𝑃 < 0.001). However, after activation of DCs with Leishma-niaKMP-11 antigen expression of IL-10 and TGF-𝛽 remainedlow whereas expression level of IL-10 and TGF-𝛽 by MΦremained higher than in DCs (𝑃 < 0.001). These resultsindicate that Leishmania did not influence the triggering ofimmune-suppression factors, especially during presentationof Leishmania KMP-11 antigen to DCs in VL (𝑃 < 0.05)compared to unstimulated. Indeed,VLparticipants presentedwith greater upliftment of IL-12 expression in DCs after
activation with KMP-11 antigen compared with MΦ resultsand the control group (𝑃 < 0.001).
3.4. Leishmania Contributes to Inhibiting NF-𝜅B Signalling inMΦs Which Are However Upregulated by KMP-11 Antigen inmoDCs in VL Patients. The differences in the productionof IL-12 by moDCs and MΦs elicited by KMP-11 antigenprompted us to examine the activation of theNF-𝜅B byKMP-11 antigen.As illustrated in Figure 5,many striking differenceswere observed in moDCs andMΦs with regard to the NF-𝜅Bactivation in the VL patients. In general, it was observed thatLeishmania contributed to inhibiting NF-𝜅B signalling, theevidence of whichwasmore predominant inMΦs rather thanmoDCs of the VL patients. A significant increase in NF-𝜅Bpattern was observed inMΦs and DCs when stimulated withKMP-11 and PHA in comparison with their respective un-stimulated (VL and ctrl.) populations (𝑃 < 0.001). However,we observed a high expression of NF-𝜅B in KMP-11 or PHAstimulated DCs in comparison to KMP-11 or PHA stimulatedMΦs. NF-𝜅B production in DCs increased after stimulationwith KMP-11 antigen about 11.22-fold (𝑃 < 0.001) higherfrom base value (ex vivo) in VL patients. Comparatively,there was observed significantly lower (3.2-fold, 𝑃 < 0.05)ability of MΦs to produce NF-𝜅B after the KMP-11 antigenstimulation frombase value.When compared betweenAPCs,ability of MΦs to produce NF-𝜅B was lower (2.1-fold, 𝑃 <0.001) in KMP-11 stimulated group than the DCs in patients.A significant difference was observed among VL patients(𝑃 < 0.001) and control groups (𝑃 < 0.01) of MΦ and DCpopulations, respectively. There was no significant differenceobserved between VL and Ctrl. groups of MΦs and DCsin different groups, respectively. However VL patients (un-stimulated) have shown 4.92-fold and 1.30-fold increase in

BioMed Research International 5
FSC-
HSSC-H
(a)
CD83
-PE
CD86-FITC CD80-FITC
HLA
-DR-
PE(B1) (B2)
(b)
CD1a-FITC
CD14
-PE
(c)
(D1) (D2)
(d)
CD14
-PE
CD19-FITC
(e)
Figure 2: Phenotypic characterization of mature dendritic cells for expression of costimulatory molecules (CD80, CD83, and CD86) in VLcases. Maturation was induced by 7-days stimulation of DC with TNF-𝛼 for 48 hours. (a) Large cells were gated. (b) (B1-B2)The differentiallyexpressed molecule is CD83 (B1) and HLA-DR (B2). The expression was done on at least 3 separate patients. The differentially expressedmolecule is CD1a. (c)Mature DCs showing a lesser percentage of CD1a positivity in CD14 and CD1a double stained cells. (d) (D1-D2) Invertedmicroscopic picture of immature andmatureDCs. (e)Maturemacrophage showing a greater percentage of CD14+ cells and a lesser percentageof CD19+ cells after 72 h culture.
NF-𝜅B pattern in MΦs and DCs population, respectively(Figure 5).
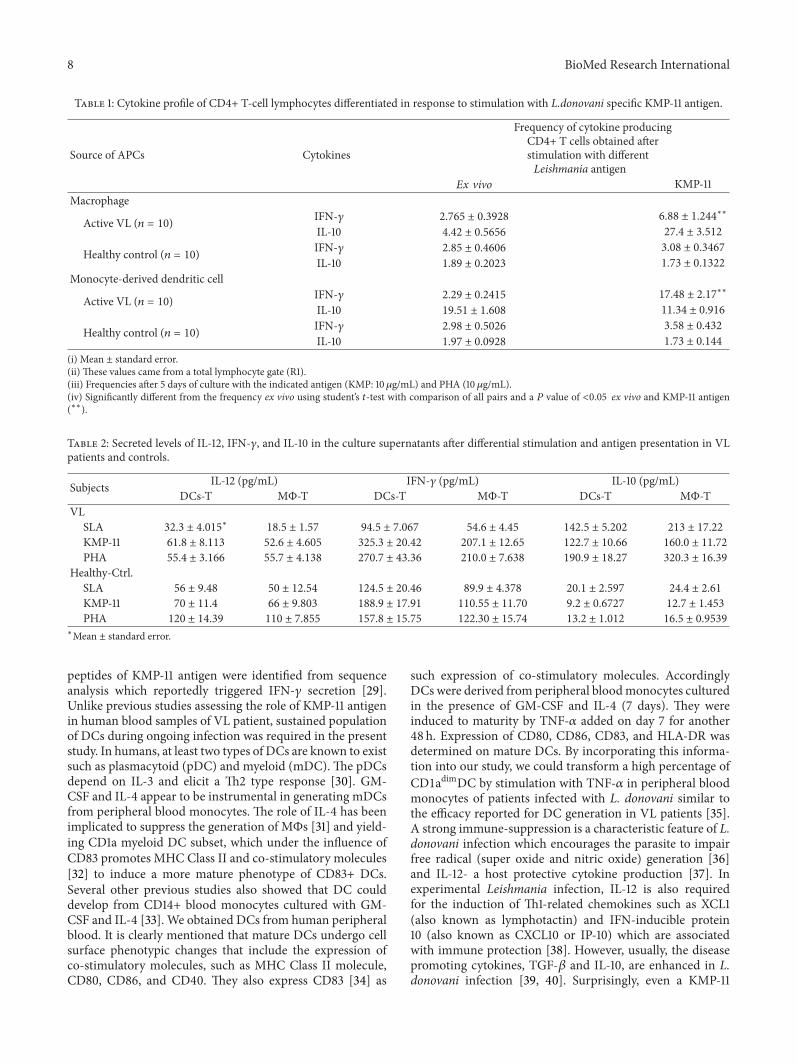
3.5. Interaction of DCs with CD4+ T Cells Contribute toIntense IFN-𝛾 Production in Response to KMP-11 Antigen.We finally validated the potency of DCs in priming anddirecting CD4 T cell cytokine immune response to KMP-11 in VL cases (Table 1). There was absence of Leishmaniaspecific T cell response in healthy control T cells. The IFN-𝛾 and IL-10 ranges for the healthy control T cells in theabsence or presence of costimulation were lower. In fact, nodifference was seen comparing IFN-𝛾 and IL-10 range beforeand after costimulation for moDCs and MΦs in the control.
On the other hand, considerable differences were observedin these APCs at the level of costimulation through KMP-11antigen compared to the corresponding values obtained inthe absence of costimulation in VL patients. In general theIFN-𝛾 produced during the interaction of DCs with CD4+T cells was 7.63-fold higher in response to KMP-11 than exvivo value (𝑃 < 0.001). When theMΦs presented the KMP-11antigen, IFN-𝛾 production in the sameCD4+T cells was only2.49-fold greater in response to KMP-11 than ex vivo value(𝑃 > 0.05). On the other hand, the presentation of KMP-11 antigen by MΦs induced 2.41-fold IL-10 in the T cells ofVL patients compared to when DCs presented the KMP-11antigen (𝑃 < 0.001). The difference between moDCs and

6 BioMed Research International
0
10
20
30
Ex vivo KMP-11 PHA
######
^^^@
$$$
$$$$$$
$$$
IL-1
2 po
sitiv
e cel
ls (%
)
VL-
MΦ
VL-
DC
VL-
DC
Ctrl-
MΦ
Ctrl-
MΦ
Ctrl-
DC
VL-
MΦ
VL-
DC
Ctrl-
MΦ
Ctrl-
DC
Ctrl-
DC
VL-
MΦ
^^^∗∗∗
+
(a)
Ex vivo
Ex vivo
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
CD4
-PE
CD83
-PE
IL-12-FITC IL-12-FITC IL-12-FITC
IL-12-FITC IL-12-FITC IL-12-FITC
Macrophages (IL-12)
Dendritic cells (IL-12)
KMP-11 sti
KMP-11 sti
(1.27%) (7.1%)PHA-sti
PHA-sti
(15.6%)
(6.45%) (19.65%) (14.3%)
FL2
-HFL2
-H
FL2
-H
FL2
-HFL2
-H
FL2
-H
100 101 102 103 104 100 101 102 103 104 100 101 102 103 104
100 101 102 103 104100 101 102 103 104100 101 102 103 104
FL1-H
FL1-H FL1-H FL1-H
FL1-H FL1-H
Ex vivo(1.27%)
KMP-11 sti(19.65%)
PHA-sti(14.3%)
(b)
Figure 3: (a) IL-12 release by MΦ and moDC after KMP-11 and PHA stimulation. (a) Comparison of cytokine production by APCs aftercostimulation by KMP-11. 1 × 106/mL of either CD14+ MΦ or CD83+ moDCs was stimulated with KMP-11 (10𝜇g/mL) and PHA (10𝜇g) for16 h. Harvested cells were consecutively coincubated with Bref-A (1𝜇g/mL), surface CD-4 (PE)/CD-83 (PE) antibodies, and cytofix/Permsolution before cytoplasmic staining with FITC for IL-12 and acquired and analyzed on FACS-calibur. Values were expressed in mean ± SEM.Each sample was run in duplicate. ∗∗∗𝑃 < 0.001 versus VL-MΦ; ###
𝑃 < 0.001 versus VL-DC; ∧∧∧𝑃 < 0.001 versus Ctrl-DC; @𝑃 < 0.05versus VL-MΦ-KMP-11 and $$$
𝑃 < 0.001 versus respective MΦ and DC groups; and +𝑃 < 0.05 between VL-DC-KMP-11 and Ctrl-DC-KMP-11 groups. (b) Representative dot plot for the intracellular staining of IL-12 in MΦs and DCs of VL patients.
MΦs became obvious with the finding that IL-10 producedduring the interaction of MΦs with T-cells were 6.19-foldhigher than control (𝑃 < 0.001) whereas CD4 cells during aninteraction with moDCs yielded about 1.72-fold lower IL-10response to KMP-11 antigen of L. donovani.
The cytokine polarization index (Loge IFN-𝛾: Loge IL-10) (Figure 6) was observed to be more polarized towardsIFN-𝛾 after stimulation of MΦs and DCs with KMP-11 in VLpatients compared to controls. The comparison revealed that
this polarizationwasmore towards IFN-𝛾 after stimulation ofDCs with KMP-11 protein.
3.6. Secreted Levels of IL-12, IFN-𝛾, and IL-12 in CultureSupernatants. Since the supernatants of cultures of all thesamples were not stored, in the representative supernatants ofthe cultures of 3VL patients and 3 healthy controls, secretedlevels of IL-12, IFN-𝛾, and IL-12 were estimated by ELISA.Levels of IL-12 and IFN-𝛾 were high after presentation of L.

BioMed Research International 7
010203040
#
###
aaa
bbb
ccc
$$$
$
Ex vivo KMP-11 PHA
IL-1
0 po
sitiv
e cel
ls (%
)
VL-
MΦ
VL-
DC
VL-
DC
Ctrl-
MΦ
Ctrl-
MΦ
Ctrl-
DC
VL-
MΦ
VL-
DC
Ctrl-
MΦ
Ctrl-
DC
Ctrl-
DC
VL-
MΦ
∗∗∗∗∗∗
(a)
010203040
###aaa
@@@bbb
cc$$$
$$$
$$$
Ex vivo KMP-11 PHA
TGF-
B po
sitiv
e cel
ls (%
)
VL-
MΦ
VL-
DC
VL-
DC
Ctrl-
MΦ
Ctrl-
MΦ
Ctrl-
DC
VL-
MΦ
VL-
DC
Ctrl-
MΦ
Ctrl-
DC
Ctrl-
DC
VL-
MΦ
∗∗∗
∗∗∗
+ + +
(b)
Ex vivo
Ex vivo
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
CD4
-PE
CD83
-PE
IL-10-FITC IL-10-FITC IL-10-FITC
IL-10-FITC IL-10-FITC IL-10-FITC
Macrophages (IL-10)
Dendritic cells (IL-10)
KMP-11 sti
KMP-11 sti
PHA-sti
PHA-sti(3.15%)
FL2
-H
FL2
-H
FL2
-H
FL2
-H
FL2
-H
FL2
-H
100 101 102 103 104100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100 101 102 103 104
(2.56%) (19.17%) (10.67%)
(17.5%) (17.73%)
FL1-H FL1-H FL1-H
FL1-HFL1-HFL1-H
Ex vivo(2.56%)
(c)
Figure 4: Flow cytometry analysis on reversal of immune-suppression inmoDCs in response to L. donovani antigen after stimulation ofKMP-11. (a)-(b) Comparison of cytokine production byAPCs after costimulation byKMP-11. 1× 106/mL of either CD14+MΦ or CD83+moDCswasstimulated with KMP-11 (10𝜇g/mL) and PHA (10𝜇g) for 16 h. Harvested cells were consecutively coincubated with Bref-A (1𝜇g/mL), surfaceCD-4 (PE)/CD-83 (PE) antibodies, and cytofix/Perm solution before cytoplasmic staining with FITC for IL-10 and TGF-𝛽 and acquired andanalyzed on FACS-calibur. Values were expressed inmean± SEM. Each sample was run in duplicate. ∗∗∗𝑃 < 0.001 versus VL-MΦ; #
𝑃 < 0.05,###𝑃 < 0.001 versus VL-DC; aaa
𝑃 < 0.001 versus Ctrl-MΦ; @@@𝑃 < 0.001 versus VL-MΦ-KMP-11; bbb𝑃 < 0.001 versus VL-DC-KMP-11;
cc𝑃 < 0.01, ccc
𝑃 < 0.001 versus Ctrl-KMP-11; $𝑃 < 0.05, $$$
𝑃 < 0.001 versus respective MΦ and DC groups; and +++𝑃 < 0.001 between
respective VL-DC and Ctrl-DC groups. (c) Representative dot plot for the intracellular staining of IL-10 in MΦs and DCs of VL patients.
donovani specific antigen KMP-11 by DCs to T cells in VLpatients. On the contrary, levels of IL-10 were high whenKMP-11 antigen of L. donovani was delivered by MΦs to Tcells in the VL patients compared with DCs-T counterpartsand healthy controls (Table 2).
4. Discussion
The present study successfully demonstrates that L. donovanispecific KMP-11 antigen can act to promote host protectiveimmune response during chronic phase of infection with L.donovani. Furthermore, by comparing the role of DCs and
MΦs in the presence of this L. donovani specific antigenduring the chronic phage of VL, we could show that whereasboth DCs and MΦs contribute to suppress host protectiveimmune response ex vivo, it was this KMP-11 antigen whichwas capable of promoting the activation ofDCs inVLpatientsthat produced IL-12.
There were two reasons why KMP-11 was chosen as thetesting antigen. Firstly, lipophosphoglycan linked KMP-11 isthe major protein on the promastigote surface which playsa key role in immunity against this parasite [14, 25, 26].The induction of IFN-𝛾 is critical for resistance and curein all forms of leishmaniasis [27, 28]. In this context, 30
8 BioMed Research International
Table 1: Cytokine profile of CD4+ T-cell lymphocytes differentiated in response to stimulation with L.donovani specific KMP-11 antigen.
Source of APCs Cytokines
Frequency of cytokine producingCD4+ T cells obtained afterstimulation with differentLeishmania antigen
Ex vivo KMP-11Macrophage
Active VL (𝑛 = 10) IFN-𝛾 2.765 ± 0.3928 6.88 ± 1.244∗∗
IL-10 4.42 ± 0.5656 27.4 ± 3.512
Healthy control (𝑛 = 10) IFN-𝛾 2.85 ± 0.4606 3.08 ± 0.3467
IL-10 1.89 ± 0.2023 1.73 ± 0.1322
Monocyte-derived dendritic cell
Active VL (𝑛 = 10) IFN-𝛾 2.29 ± 0.2415 17.48 ± 2.17∗∗
IL-10 19.51 ± 1.608 11.34 ± 0.916
Healthy control (𝑛 = 10) IFN-𝛾 2.98 ± 0.5026 3.58 ± 0.432
IL-10 1.97 ± 0.0928 1.73 ± 0.144
(i) Mean ± standard error.(ii) These values came from a total lymphocyte gate (R1).(iii) Frequencies after 5 days of culture with the indicated antigen (KMP: 10𝜇g/mL) and PHA (10 𝜇g/mL).(iv) Significantly different from the frequency ex vivo using student’s 𝑡-test with comparison of all pairs and a 𝑃 value of <0.05 ex vivo and KMP-11 antigen(∗∗).
Table 2: Secreted levels of IL-12, IFN-𝛾, and IL-10 in the culture supernatants after differential stimulation and antigen presentation in VLpatients and controls.
Subjects IL-12 (pg/mL) IFN-𝛾 (pg/mL) IL-10 (pg/mL)DCs-T MΦ-T DCs-T MΦ-T DCs-T MΦ-T
VLSLA 32.3 ± 4.015
∗
18.5 ± 1.57 94.5 ± 7.067 54.6 ± 4.45 142.5 ± 5.202 213 ± 17.22
KMP-11 61.8 ± 8.113 52.6 ± 4.605 325.3 ± 20.42 207.1 ± 12.65 122.7 ± 10.66 160.0 ± 11.72
PHA 55.4 ± 3.166 55.7 ± 4.138 270.7 ± 43.36 210.0 ± 7.638 190.9 ± 18.27 320.3 ± 16.39
Healthy-Ctrl.SLA 56 ± 9.48 50 ± 12.54 124.5 ± 20.46 89.9 ± 4.378 20.1 ± 2.597 24.4 ± 2.61
KMP-11 70 ± 11.4 66 ± 9.803 188.9 ± 17.91 110.55 ± 11.70 9.2 ± 0.6727 12.7 ± 1.453
PHA 120 ± 14.39 110 ± 7.855 157.8 ± 15.75 122.30 ± 15.74 13.2 ± 1.012 16.5 ± 0.9539
∗Mean ± standard error.
peptides of KMP-11 antigen were identified from sequenceanalysis which reportedly triggered IFN-𝛾 secretion [29].Unlike previous studies assessing the role of KMP-11 antigenin human blood samples of VL patient, sustained populationof DCs during ongoing infection was required in the presentstudy. In humans, at least two types of DCs are known to existsuch as plasmacytoid (pDC) and myeloid (mDC). The pDCsdepend on IL-3 and elicit a Th2 type response [30]. GM-CSF and IL-4 appear to be instrumental in generating mDCsfrom peripheral blood monocytes. The role of IL-4 has beenimplicated to suppress the generation of MΦs [31] and yield-ing CD1a myeloid DC subset, which under the influence ofCD83 promotes MHC Class II and co-stimulatory molecules[32] to induce a more mature phenotype of CD83+ DCs.Several other previous studies also showed that DC coulddevelop from CD14+ blood monocytes cultured with GM-CSF and IL-4 [33]. We obtained DCs from human peripheralblood. It is clearly mentioned that mature DCs undergo cellsurface phenotypic changes that include the expression ofco-stimulatory molecules, such as MHC Class II molecule,CD80, CD86, and CD40. They also express CD83 [34] as
such expression of co-stimulatory molecules. AccordinglyDCs were derived from peripheral bloodmonocytes culturedin the presence of GM-CSF and IL-4 (7 days). They wereinduced to maturity by TNF-𝛼 added on day 7 for another48 h. Expression of CD80, CD86, CD83, and HLA-DR wasdetermined on mature DCs. By incorporating this informa-tion into our study, we could transform a high percentage ofCD1adimDC by stimulation with TNF-𝛼 in peripheral bloodmonocytes of patients infected with L. donovani similar tothe efficacy reported for DC generation in VL patients [35].A strong immune-suppression is a characteristic feature of L.donovani infection which encourages the parasite to impairfree radical (super oxide and nitric oxide) generation [36]and IL-12- a host protective cytokine production [37]. Inexperimental Leishmania infection, IL-12 is also requiredfor the induction of Th1-related chemokines such as XCL1(also known as lymphotactin) and IFN-inducible protein10 (also known as CXCL10 or IP-10) which are associatedwith immune protection [38]. However, usually, the diseasepromoting cytokines, TGF-𝛽 and IL-10, are enhanced in L.donovani infection [39, 40]. Surprisingly, even a KMP-11

BioMed Research International 9
0102030
Ex vivo KMP-11 PHA
######$$$
$$$V
L-MΦ
VL-MΦ
VL-
DC
VL-
DC
VL-DC
Ctrl-
MΦ
Ctrl-
MΦ
Ctrl-
DC
VL-
MΦ
VL-
DC
Ctrl-
MΦ
Ctrl-
DC
Ctrl-
DC
VL-
MΦ
^^^^^^
∗∗∗
4050
aaaa
$$$$
∗
65 kDa65 kDa
UNS KMP-11 UNS KMP-11
NF-𝜅
B po
sitiv
e cel
ls (%
)
Figure 5: Differential NF-𝜅B pattern after stimulation with KMP-11 in APCs of VL patients and healthy controls. The capacityof moDCs and MΦs for the activation of NF-𝜅B was comparedthrough intracellular staining using fluorescent conjugated anti-NF-𝜅B antibodies on flow cytometry. Following stimulations (KMP-11and PHA), cells were harvested and then incubated with PE-anti-p65 NF-𝜅B antibodies, acquired and analyzed on FACS Calibur.Total amount of NF-𝜅Bwas produced byMΦ andmoDC stimulatedand unstimulated with rKMP-11 evaluated through FACS calibur.Immunoblotting of VL MΦ and VL DCs (insert, Figure 5) withNF-𝜅B antibody shows that KMP-11 triggered significant phospho-rylation of a protein migrating at 65 kDa of NF-𝜅B. There was nosignificant difference observed between DC-Ctrl and VL-DC ofrespective groups. ∗𝑃 < 0.05, ∗∗∗𝑃 < 0.001 versus VL-MΦ; ###𝑃 <0.001 versus VL-DC; a
𝑃 < 0.05, aaa𝑃 < 0.001 versus Ctrl-MΦ;
∧∧∧
𝑃 < 0.001 versus Ctrl-DC; @𝑃 < 0.05 versus VL-MΦ-KMP-11and $$𝑃 < 0.01, $$$
𝑃 < 0.001 versus respective MΦ and DC groups.
Log eI
FN-𝛾
-pro
duci
ng ce
lls/L
oge
IL-10
prod
ucin
g ce
lls
VL Ctrl VL Ctrl VL Ctrl VL Ctrl
MΦ MΦDC DC
KMP-11 stimulated8
6
4
2
0
Ex vivo
Figure 6: Cytokine polarization index after exposure ofmacrophages andmoDCs to KMP-11 and subsequent coculture withT cells in VL patients and healthy controls. Cytokine polarizationindex after exposure ofMΦs andmoDCs to KMP-11 and subsequentco-culturing with T cells in VL patients. The index was obtainedafter calculating the ratio of loge IFN-𝛾: loge IL-10, based on FACSCalibur using CellQuest Pro software.
stimulation given to MΦs could not augment adequate IL-12production in VL patients. Earlier studies also suggest thatLeishmania has the ability to suppress IL-12 production inMΦs [2, 4, 41]. Of note, in spite of stimulation, TGF-𝛽 andIL-10 were observed to increase in MΦs which suggestedthat all parameters of host resistance that we measured were
suppressed in MΦs even after stimulation with KMP-11 anti-gen. Importantly we could not distinguish much between theeffects ex vivo and after KMP-11 stimulation. An importantcharacteristic feature of VL is production of the immuno-regulatory cytokine IL-10 and targeting of IL-10 signalling hasbeen identified as a potential therapeutic strategy [24, 42–44].IL-10 suppresses IFN-𝛾 induced NO production [45]; hencethe increase of IL-10 expressing MΦs may itself underlie thesuppression inMΦs due to Leishmania infection. In contrast,the cytokine response profile triggered by KMP-11 antigen inDCs had differential characteristics such as an increase in IL-12 production concomitantly with amarked decrease in TGF-𝛽 and IL-10 production.
During the study on differential effects of APCs on T cellsagainst the KMP-11 antigen, we provided a long incubation toT cells exposed Leishmania KMP-11 antigen anticipating thatthey had previously been sensitized Leishmania from MΦsand it would be difficult to observe the effect of LeishmaniaKMP-11 antigen presented via DCs to the T cells in a shorterperiod. Reiner et al. (1990) [46] showed that incubationtime did not affect the usual pattern of cytokine response.However, in this earlier study, parasite delivery was done byMΦs only while we attempted to sensitize the T cells withLeishmania KMP-11 antigen from two different sources.
Unlike previous data showing an impact of KMP-11antigen in the stimulation of effector IFN-𝛾 producing CD4cells, the stimulation of T cells after presentation of KMP-11antigen by MΦs during infection did not significantly affectIFN-𝛾 production by CD4+ T cells, suggesting that testingthe efficiencies of an immunogenic candidate via deliverythrough MΦs may suffer as the MΦs could not be essentialfor the maintenance of effector T-cell response. However, ourdata were in contrast when we delivered the KMP-11 antigento T cells via DCs showing that the presentation of the sameantigen by DCs during established infection with L. donovaniresulted in a significant increase in IFN-𝛾 production byCD4+ T cells.The chronic L. donovani infection is associatedwith expansion of CD4+ T cells that express IL-10. Thefurther analysis made here indicates that, during L. donovaniinfection, KMP-11 antigen decreases IL-10-producing CD4+T cells after its delivery via DCs which was more pronouncedthrough DCs compared to the MΦs. Thus, though KMP-11antigen was an effective immunogen, it required providingoptimum immune-stimulatory effects only when delivered toT cells via DCs. Hence, the requirement for KMP-11 antigentomaintainT-cell IFN-𝛾productionwould appear to bemoreprotection-specific, when presented byDCs rather thanMΦs.
MΦs have multiple functions; they serve as host cellsfor parasite replication, as antigen-presenting cells, and as asource of cytokines modulating the T-cell mediated immuneresponse. Moreover, after appropriate activation by Th1 cells,they serve as effector cells for Leishmania killing. Inferentialresults suggest that Leishmania abrogates the abilities of MΦsto abruptly disturb emergence of a Th1-like cell responseduring L. donovani infection.The data generated in this studyalso have hinted toward the lesser potential of MΦs as APCsduring VL in humans. This might be due to inadequateexpression of MHC Class II molecules on MΦs required forthe induction of effective immunity. This speculation is well

10 BioMed Research International
in accordancewith the findings of a previous study inL.majorsuggesting MΦ antigen presentation may not be required foreffector T-cell function [47].
Nevertheless, KMP-11 primed moDCs generated ade-quate IL-12 which reflected that it can have a specific effecton T-cell response that was severely impeded in KMP-11stimulatedMΦs of the samepatients. Reports on infectivity ofL. donovani are available [6] andDCs have also been observedto produce IL-12 after infection with L. major [48] and L.amazonensis [49].
The differential production of IL-12 between MΦs andDCs was observed mainly due to differences in NF-𝜅Bproduction after the presentation of L. donovani KMP-11antigen. This difference may offer distinct signals to immunecells that can affect differentiation of CD4+ T cells to Th1or Th2. In support of this possibility, it is known that thesecells require NF-𝜅B support for maturation [50]. Moreover,the signalling initiated by TNF receptor associated factor2, 3 (TRAF 2-3) at cholesterol rich membrane rafts onthe plasma membrane of DCs leads to p38 MAP kinasephosphorylation for CD40 mediated IL-12 production ofdifferent types of Leishmania [51, 52]. The src family proteinkinase, Lyn, is reported to simultaneously activate the ERKpathway during the process [51]. Further, the NF-𝜅B hasbeen shown to be required for the production of IL-12 andIFN-𝛾 as well as inducible nitric oxide synthase (iNOs)[53–56].
This extent of formidable immunological changes inmoDCs was hitherto not observed to occur in MΦs, sugges-tive of the importance of APCs in judging the immunogenicpotential of KMP-11 as amean to achieve a protective immuneresponse in VL patients. Thus, we found that the KMP-11 antigen of L. donovani had the ability to produce IL-10and TGF-𝛽 in MΦs and IL-12 in DCs in VL patients. Suchalterations in cytokine production in response to KMP-11antigen which we have observed in this study can contributeto bias the immune response which may be important for theoutcome of the disease.
Ethical Approval
Ethical approval was taken from Ethical Committee of Insti-tute.
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper.
Acknowledgments
This work was supported by the Indian Council of MedicalResearch, Ministry of Health and Family Welfare, Govern-ment of India. The authors are indebted to ICMR for thefellowship awarded to Rajesh Chaudhary to pursue this study.The authors are also thankful to Dr. Prakash Chandra andDr. Kumar Vaibhav (GRU, Augusta, GA) for their support toprepare this paper.
References
[1] J. Argueta-Donohue, N. Carrillo, L. Valdes-Reyes et al., “Leish-mania mexicana: participation of NF-𝜅B in the differentialproduction of IL-12 in dendritic cells andmonocytes induced bylipophosphoglycan (LPG),” Experimental Parasitology, vol. 120,no. 1, pp. 1–9, 2008.
[2] L. Carrera, R. T. Gazzinelli, R. Badolato et al., “Leishmania pro-mastigotes selectively inhibit interleukin 12 induction in bonemarrow-derived macrophages from susceptible and resistantmice,” Journal of Experimental Medicine, vol. 183, no. 2, pp. 515–526, 1996.
[3] A. Sartori,M.A. P.Oliveira, P. Scott, andG. Trinchieri, “Metacy-clogenesis modulates the ability of Leishmania promastigotes toinduce IL-12 production in human mononuclear cells,” Journalof Immunology, vol. 159, no. 6, pp. 2849–2857, 1997.
[4] Y. Belkaid, B. Butcher, andD. L. Sacks, “Analysis of cytokine pro-duction by inflammatory mouse macrophages at the single-celllevel: selective impairment of IL-12 induction in Leishmania-infected cells,” European Journal of Immunology, vol. 28, no. 4,pp. 1389–1400, 1998.
[5] P. Scott and C. A. Hunter, “Dendritic cells and immunity toleishmaniasis and toxoplasmosis,”Current Opinion in Immunol-ogy, vol. 14, no. 4, pp. 466–470, 2002.
[6] M. Ghosh and S. Bandyopadhyay, “Interaction of Leishmaniaparasites with dendritic cells and its functional consequences,”Immunobiology, vol. 209, no. 1-2, pp. 173–177, 2004.
[7] D. N. J. Hart, “Dendritic cells: unique leukocyte populationswhich control the primary immune response,” Blood, vol. 90,no. 9, pp. 3245–3287, 1997.
[8] P. M. A. Gorak, C. R. Engwerda, and P. M. Kaye, “Dendriticcells, but nomacrophages, produce IL-12 immediately followingLeishmania donovani infection,” European Journal of Immunol-ogy, vol. 28, pp. 687–695, 1998.
[9] S. Gordon, “Pattern recognition receptors: doubling up for theinnate immune response,” Cell, vol. 111, no. 7, pp. 927–930, 2002.
[10] C. A. Janeway Jr. and R. Medzhitov, “Innate immune recogni-tion,” Annual Review of Immunology, vol. 20, pp. 197–216, 2002.
[11] S. Akira, “Toll-like receptor signaling,” Journal of BiologicalChemistry, vol. 278, no. 40, pp. 38105–38108, 2003.
[12] S. M. Beverley and S. J. Turco, “Lipophosphoglycan (LPG) andthe identification of virulence genes in the protozoan parasiteLeishmania,” Trends in Microbiology, vol. 6, no. 1, pp. 35–40,1998.
[13] A. Jardim, V. Funk, R.M. Caprioli, and R.W.Olafson, “Isolationand structural characterization of the Leishmania donovanikinetoplastid membrane protein-11, a major immunoreactivemembrane glycoprotein,”Biochemical Journal, vol. 305, no. 1, pp.307–313, 1995.
[14] D. L. Tolson, A. Jardim, L. F. Schnur et al., “The kinetoplastidmembrane protein 11 of Leishmania donovani and Africantrypanosomes is a potent stimulator of T-lymphocyte prolifer-ation,” Infection and Immunity, vol. 62, no. 11, pp. 4893–4899,1994.
[15] J. A. L. Kurtzhals, A. S. Hey, A. Jardim et al., “Dichotomyof the human T cell response to Leishmania antigens. II.Absent of Th2-like response to gp63 and Th1-like responseto lipophosphoglycan- associated protein in cells from curedvisceral leishmaniasis patients,” Clinical and ExperimentalImmunology, vol. 96, no. 3, pp. 416–421, 1994.
[16] R. Basu, S. Bhaumik, J. M. Basu, K. Naskar, T. De, and S. Roy,“Kinetoplastid membrane protein-11 DNA vaccination induces

BioMed Research International 11
complete protection against both pentavalent antimonial-sensitive and -resistant strains of Leishmania donovani thatcorrelates with inducible nitric oxide synthase activity and IL-4generation: evidence for mixed Th1- and Th2-like responses invisceral Leishmaniasis,” Journal of Immunology, vol. 174, no. 11,pp. 7160–7171, 2005.
[17] D. I. Lacerda, L. Cysne-Finkelstein, M. P. Nunes et al., “Kineto-plastid membrane protein-11 exacerbates infection with Leish-mania amazonensis in murine macrophages,” Memorias doInstituto Oswaldo Cruz, vol. 107, no. 2, pp. 238–245, 2012.
[18] M. Olivier, K. G. Baimbridge, and N. E. Reiner, “Stimulus-response coupling in monocytes infected with Leishmania:attenuation of calcium transients is related to defective agonist-induced accumulation of inositol phosphates,” Journal ofImmunology, vol. 148, no. 4, pp. 1188–1196, 1992.
[19] J. D. Chulay and A. D. M. Bryceson, “Quantitation of amastig-otes of Leishmania donovani in smears of splenic aspiratesfrom patients with visceral leishmaniasis,” American Journal ofTropical Medicine and Hygiene, vol. 32, no. 3, pp. 475–479, 1983.
[20] A. Jayakumar,M. J. Donovan, V. Tripathi,M. Ramalho-Ortigao,and M. A. McDowell, “Leishmania major infection activatesNF-𝜅B and interferon regulatory factors 1 and 8 in humandendritic cells,” Infection and Immunity, vol. 76, no. 5, pp. 2138–2148, 2008.
[21] M. S. Bernstein, S. E. Tong-Starksen, and R. M. Locksley,“Activation of human monocyte-derived macrophages withlipopolysaccharide decreases human immunodeficiency virusreplication in vitro at the level of gene expression,” Journal ofClinical Investigation, vol. 88, no. 2, pp. 540–545, 1991.
[22] C. Prussin and D. D. Metcalfe, “Detection of intracytoplasmiccytokine using flow cytometry and directly conjugated anti-cytokine antibodies,” Journal of Immunological Methods, vol.188, no. 1, pp. 117–128, 1995.
[23] M. Litton, J. Anderson, L. Bjork, T. Fehniger, A. K. Ulfgren,and U. Anderson, “Cytoplasmic cytokine staining in individualcells,” in Human Cytokine Protocols, Debets and Savelkakoul,Eds., Human Press, 1996.
[24] S. Bimal, S. K. Singh, S. Sinha et al., “Leishmania donovani: roleof CD2 on CD4+ T-cell function in Visceral Leishmaniasis,”Experimental Parasitology, vol. 118, no. 2, pp. 238–246, 2008.
[25] S. Mukhopadhyay, P. Sen, H. K. Majumder, and S. Roy,“Reduced expression of lipophosphoglycan (LPG) and kineto-plastid membrane protein (KMP)-11 in Leishmania donovanipromastigotes in axenic culture,” Journal of Parasitology, vol. 84,no. 3, pp. 644–647, 1998.
[26] T. Naderer, J. E. Vince, and M. J. McConville, “Surface determi-nants of Leishmania parasites and their role in infectivity in themammalian host,” Current Molecular Medicine, vol. 4, no. 6, pp.649–665, 2004.
[27] H. W. Murray, “Tissue granuloma structure-function in exper-imental visceral leishmaniasis,” International Journal of Experi-mental Pathology, vol. 82, no. 5, pp. 249–267, 2001.
[28] H. W. Murray, K. C. Flanders, D. D. Donaldson et al., “Antag-onizing deactivating cytokines to enhance host defense andchemotherapy in experimental visceral leishmaniasis,” Infectionand Immunity, vol. 73, no. 7, pp. 3903–3911, 2005.
[29] R. Basu, S. Roy, and P. Walden, “HLA class I-restricted T cellepitopes of the kinetoplastid membrane protein-11 presented byLeishmania donovani-infected humanmacrophages,” Journal ofInfectious Diseases, vol. 195, no. 9, pp. 1373–1380, 2007.
[30] J. Chehimi, D. E. Campbell, L. Azzoni et al., “Persistentdecreases in blood plasmacytoid dendritic cell number and
function despite effective highly active antiretroviral therapyand increased blood myeloid dendritic cells in HIV-infectedindividuals,” Journal of Immunology, vol. 168, no. 9, pp. 4796–4801, 2002.
[31] F. Sallusto and A. Lanzavecchia, “Efficient presentation ofsoluble antigen by cultured human dendritic cells is main-tained by granulocyte/macrophage colony-stimulating factorplus interleukin 4 and downregulated by tumor necrosis factor𝛼,” Journal of Experimental Medicine, vol. 179, no. 4, pp. 1109–1118, 1994.
[32] M. Ulanova, A. Tarkowski, M. Hahn-Zoric, and L. A. Hanson,“The common vaccine adjuvant aluminum hydroxide up-regulates accessory properties of human monocytes via aninterleukin-4-dependent mechanism,” Infection and Immunity,vol. 69, no. 2, pp. 1151–1159, 2001.
[33] J. I. Mayordomo, T. Zorina, W. J. Storkus et al., “Bone marrow-derived dendritic cells pulsed with synthetic tumour peptideselicit protective and therapeutic antitumour immunity,” NatureMedicine, vol. 1, no. 12, pp. 1297–1302, 1995.
[34] S. Grabbe, E. Kampgen, and G. Schuler, “Dendritic cells: multi-lineal and multi-functional,” Immunology Today, vol. 21, no. 9,pp. 431–433, 2000.
[35] K. C. Roy, G. Bandyopadhyay, S. Rakshit, M. Ray, and S. Bandy-opadhyay, “IL-4 alone without the involvement of GM-CSFtransforms human peripheral blood monocytes to a CD1adim,CD83+ myeloid dendritic cell subset,” Journal of Cell Science,vol. 117, no. 16, pp. 3435–3445, 2004.
[36] S. Stenger, K. R. Niazi, and R. L. Modlin, “Down-regulationof CD1 on antigen-presenting cells by infection with Mycobac-terium tuberculosis,” Journal of Immunology, vol. 161, no. 7, pp.3582–3588, 1998.
[37] A. Adhikari, G. Gupta, S. Majumdar et al., “Mycobacteriumindicus pranii (Mw) Re-establishes host protective immuneresponse in Leishmania donovani infectedmacrophages: criticalrole of IL-12,” PLoS One, vol. 7, no. 7, article e40265, 2012.
[38] C. Zaph and P. Scott, “Interleukin-12 regulates chemokine geneexpression during the early immune response to Leishmaniamajor,” Infection and Immunity, vol. 71, no. 3, pp. 1587–1589,2003.
[39] J. RodriguesV., J. S. Da Silva, andA.Campos-Neto, “Transform-ing growth factor 𝛽 and immunosuppression in experimentalvisceral leishmaniasis,” Infection and Immunity, vol. 66, no. 3,pp. 1233–1236, 1998.
[40] C. Bogdan, Y. Vodovotz, and C. Nathan, “Macrophage deacti-vation by interleukin 10,” Journal of Experimental Medicine, vol.174, no. 6, pp. 1549–1555, 1991.
[41] S. L. Reiner and R. M. Locksley, “The regulation of immunity toLeishmania major,” Annual Review of Immunology, vol. 13, pp.151–177, 1995.
[42] S. Nylen and D. Sacks, “Interleukin-10 and the pathogenesis ofhuman visceral leishmaniasis,” Trends in Immunology, vol. 28,no. 9, pp. 378–384, 2007.
[43] B. M. J. Owens, L. Beattie, J. W. J. Moore et al., “IL-10-producingTh1 cells and disease progression are regulated by distinctCD11c+ cell populations during visceral Leishmaniasis,” PLoSPathogen, vol. 8, no. 7, article e1002827, 2012.
[44] E. M. Carvalho, O. Bacellar, C. Brownell, T. Regis, R. L.Coffman, and S. G. Reed, “Restoration of IFN-𝛾 production andlymphocyte proliferation in visceral leishmaniasis,” Journal ofImmunology, vol. 152, no. 12, pp. 5949–5956, 1994.

12 BioMed Research International
[45] R. T. Gazzinelli, I. P. Oswald, S. L. James, and A. Sher, “IL-10inhibits parasite killing and nitrogen oxide production by IFN-𝛾- activated macrophages,” Journal of Immunology, vol. 148, no.6, pp. 1792–1796, 1992.
[46] N. E. Reiner, W. Ng, C. B. Wilson, W. R. McMaster, andS. K. Burchett, “Modulation of in vitro monocyte cytokineresponses to Leishmania donovani. Interferon-𝛾 preventsparasite-induced inhibition of interleukin 1 production andprimesmonocytes to respond to Leishmania by producing bothtumor necrosis factor-𝛼 and interleukin 1,” Journal of ClinicalInvestigation, vol. 85, no. 6, pp. 1914–1924, 1990.
[47] M. P. Lemos, F. Esquivel, P. Scott, and T. M. Laufer, “MHCclass II expression restricted to CD8𝛼+ and CD11b + dendriticcells is sufficient for control of Leishmania major,” Journal ofExperimental Medicine, vol. 199, no. 5, pp. 725–730, 2004.
[48] M. A. McDowell, M. Marovich, R. Lira, M. Braun, and D.Sacks, “Leishmania priming of human dendritic cells for CD40ligand-induced interleukin-12p70 secretion is strain and speciesdependent,” Infection and Immunity, vol. 70, no. 8, pp. 3994–4001, 2002.
[49] H. Qi, V. Popov, and L. Soong, “Leishmania amazonensis-dendritic cell interactions in vitro and the priming of parasite-specific CD4+ T cells in vivo,” Journal of Immunology, vol. 167,no. 8, pp. 4534–4542, 2001.
[50] M. Rescigno, M. Martino, C. L. Sutherland, M. R. Gold, andP. Ricciardi-Castagnoli, “Dendritic cell survival andmaturationare regulated by different signaling pathways,” Journal of Exper-imental Medicine, vol. 188, no. 11, pp. 2175–2180, 1998.
[51] P.-O. Vidalain, O. Azocar, C. Servet-Delprat, C. Rabourdin-Combe, D. Gerlier, and S. Manie, “CD40 signaling in humandendritic cells is initiated within membrane rafts,” EMBOJournal, vol. 19, no. 13, pp. 3304–3313, 2000.
[52] B. Lu, H. Yu, C.-W. Chow et al., “GADD45𝛾 mediates theactivation of the p38 and JNK MAP kinase pathways andcytokine production in effector TH1 cells,” Immunity, vol. 14, no.5, pp. 583–590, 2001.
[53] Q.-W. Xie, Y. Kashiwabara, and C. Nathan, “Role of transcrip-tion factor NF-𝜅B/Rel in induction of nitric oxide synthase,”Journal of Biological Chemistry, vol. 269, no. 7, pp. 4705–4708,1994.
[54] G. Grigoriadis, Y. Zhan, R. J. Grumont et al., “The Rel subunitof NF-𝜅B-like transcription factors is a positive and negativeregulator of macrophage gene expression: distinct roles for Relin different macrophage populations,” EMBO Journal, vol. 15,no. 24, pp. 7099–7107, 1996.
[55] A. Sica, L. Dorman, V. Viggiano et al., “Interaction of NF-𝜅BandNFATwith the interferon-𝛾 promoter,” Journal of BiologicalChemistry, vol. 272, no. 48, pp. 30412–30420, 1997.
[56] S. Sanjabi, A. Hoffmann, H.-C. Liou, D. Baltimore, and S. T.Smale, “Selective requirement for c-Rel during IL-12 P40 geneinduction macrophages,” Proceedings of the National Academyof Sciences of the United States of America, vol. 97, no. 23, pp.12705–12710, 2000.

Submit your manuscripts athttp://www.hindawi.com
Stem CellsInternational
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Disease Markers
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation http://www.hindawi.com Volume 2014
Immunology ResearchHindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Parkinson’s Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttp://www.hindawi.com


















